university microfilms, a xerox company, ann arbor,...
TRANSCRIPT

EVALUATION OF DEXTROAMPHETAMINE-INDUCED BODY TEMPERATURE CHANGES IN RATS
Item Type text; Dissertation-Reproduction (electronic)
Authors Broadie, Larry Lewis, 1940-
Publisher The University of Arizona.
Rights Copyright © is held by the author. Digital access to this materialis made possible by the University Libraries, University of Arizona.Further transmission, reproduction or presentation (such aspublic display or performance) of protected items is prohibitedexcept with permission of the author.
Download date 18/05/2018 12:40:55
Link to Item http://hdl.handle.net/10150/287563

71-2489
BROADIE, Larry Lewis, 1940-EVALUATION OF DEXTROAMPHETAMINE-INDUCED BODY TEMPERATURE CHANGES IN RATS.
University of Arizona, Ph.D., 1970 Pharmacology
University Microfilms, A XEROX Company, Ann Arbor, Michigan
THIS DISSERTATION HAS BEEN MICROFILMED EXACTLY AS RECEIVED

EVALUATION OF DEXTROAMPHETAMINE-INDUCED
BODY TEMPERATURE CHANGES IN RATS
by
Larry Lewis Broadie
A Dissertation submitted to the Faculty of the
COLLEGE OF PHARMACY
In Partial Fulfillment of the Requirements For the Degree of
DOCTOR OF PHILOSOPHY WITH A MAJOR IN PHARMACOLOGY
In the Graduate College
THE UNIVERSITY OF ARIZONA
19 7 0

THE UNIVERSITY OF ARIZONA
GRADUATE COLLEGE
I hereby recommend that this dissertation prepared under my
direction by Larry Lewis Broadie
entitled Evaluation of Dextroamphetamine-induced Body
Temperature Changes in Rats
be accepted as fulfilling the dissertation requirement of the
degree of Doctor of Philosophy
7̂ $ ' P O
Dissertation Director Date
After Inspection of the dissertation, the following members
of the Final Examination Committee concur in its approval and
recommend its acceptance:*
<OL£&*U-c>fc
• C?.\ vujimA
S~z,st- 7O
s-y- 7^
•y-ig - ?<?
jfcTfiielapprovel aiid acceptance is contingent on the candidate's adequate performance and defense of this dissertation at the final oral examination. The inclusion of this sheet bound Into the library copy of the dissertation is evidence of satisfactory performance at the final examination.

STATEMENT BY AUTHOR
This dissertation has been submitted in partial fulfillment of requirements for an advanced degree at The University of Arizona and is deposited in the University Library to be made available to borrowers under rules of the Library.
Brief quotations from this dissertation are allowable without special permission, provided that accurate acknowledgment of source is made. Requests for permission for extended quotation from or reproduction of this manuscript in whole or in part may be granted by the head of the major department or the Dean of the Graduate College when in his judgment the proposed use of the material is in the interests of scholarship. In all other instances, however, permission must be obtained from the author.
S1GNE

ACKNOWLEDGEMENTS
The author wishes to express sincere gratitude to Dr. Lincoln
Chin for his counsel and guidance, especially during the preparation of
this manuscript.
The author also wishes to express his appreciation to Dr. Albert
L. Picchioni and Dean Willis R. Brewer for their efforts and concern.
Finally, the author is indebted to his wife, Barbara, for her
continual encouragement and understanding.
Ill

TABLE OF CONTENTS
Page
LIST OF ILLUSTRATIONS vii
LIST OF TABLES x
ABSTRACT ' . . xi
1. INTRODUCTION 1
2. THE EFFECT OF INTRAVENTRICULARLY AND INTRAPERITONEALLY ADMINISTERED DEXTROAMPHETAMINE ON BODY TEMPERATURE OF RATS . . 4
Materials and Methods . . . . . 5 Animals and Housing . . 5 Administration of Drugs 5 Measurement of Body Temperature 6 Assay of Tritiated Dextroamphetamine 6 Surgical Techniques 7 Statistical Procedures 7
Results 7 Body Temperature Changes after Intraventricular
Dextroamphetamine in Restrained Rats 7 Body Temperature Changes after Intraventricular
Dextroamphetamine in Unrestrained Rats 8 Body Temperature Changes after Intraperitoneal
Dextroamphetamine in Unrestrained Rats 8 Brain Concentrations of Dextroamphetamine Sulfate ... 8 Comparison of Body Temperature Changes after
Equivalent Intraperitoneal and Intraventricular Doses of Dextroamphetamine at Various Ambient Temperatures ..... 9
Effects of Adrenergic Blocking Drugs on Body Temperature Responses to Dextroamphetamine .... 10
Effect of Pithing on Body Temperature Response to Dextroamphetamine 10
Effect of Adrenalectomy on Body Temperature Response to Dextroamphetamine ..... 11
Discussion 11
3. BEHAVIORAL AND PHYSIOLOGICAL FACTORS ASSOCIATED WITH DEXTROAMPHETAMINE-INDUCED BODY TEMPERATURE CHANGES 28
Materials and Methods 30 Measurement of Oxygen Consumption 30
lv

V
TABLE OF CONTENTS--Continued
Page
Measurement of Spontaneous Activity 31 Quantification of Sniffing Behavior 31
Results 31 Effects of Dextroamphetamine on Oxygen
Consumption 32 Effects of Dextroamphetamine on Spontaneous
Motor Activity ......... 32 Evaluation of Sniffing Behavior ....... 33
Discussion 34
4. THE ROLE OF CENTRAL MONOAMINES IN DEXTROAMPHETAMINE-1NDUCED HYPOTHERMIA 43
Materials and Methods 47 Effects of Monoamine-depleting Agents on
Dextroamphetamine-induced Body Temperat u r e C h a n g e s . . . . . 4 7
Brain Monoamine Levels after Single Intraventricular or Intraperitoneal Administration of Dextroamphetamine . ..... 48
Effects of Repeated Intraventricular Injections of Dextroamphetamine on Hypothermic Response ... 49
Brain Monoamine Levels at Peak Hypothermic Effect Induced by the Last of Three Repeated Intraventricular Injections of Dextroamphetamine .... 49
Brain Monoamine Levels at Time of Recovery of Body Temperature after Repeated Intraventricular Injections of Dextroamphetamine 49
Spectrophotofluorometric Assay of Catecholamines and 5-Hydroxytryptamine 50
Hlstochemical Fluorescence Technique 50 Results 51
Effects of Monoamine-depleting Agents on Dextroamphetamine 'Induced Body Temperature Changes ... 51
Brain Monoamine Levels after Single Intraventricular or Intraperitoneal Administration of Dextroamphetamine 51
Effects of Repeated Intraventricular Injections of Dextroamphetamine on Hypothermic Response ... 52
Brain Monoamine Levels at Peak Hypothermic Effect Induced by the Last of Three Repeated Intraventricular Injections of Dextroamphetamine 52
Brain Monoamine Levels at Time of Recovery of Body Temperature Following Repeated Intraventricular Injections of Dextroamphetamine 53
Discussion 53

vi
TABLE OF CONTEtTTS-- Continued
Page
5. GENERAL DISCUSSION 69
APPENDIX A: INTRACEREBROVENTRICULAR TECHNIQUE 80
APPENDIX B: PREPARATION OF DRUGS 85
APPENDIX C: ASSAY FOR BRAIN DEXTROAMPHETAMINE 88
APPENDIX D: STATISTICAL PROCEDURES 93
APPENDIX E: SPECTROPHOTOFLUOROMETRIC ASSAY OF 5-HYDROXY-TRYPTAMINE, NOREPINEPHRINE AND DOPAMINE 95
APPENDIX F: HISTOCHEMICAL FLUORESCENCE TECHNIQUE FOR DIRECT OBSERVATION. OF NOREPINEPHRINE, DOPAMINE AND 5-HYDROXYTRYPTAMINE 109
LIST OF REFERENCES 118

LIST OF ILLUSTRATIONS
Figure Page
1. Effect of Intraventricular Amphetamine on Body Temperature of Restrained Rats at an Ambient Temperature of 25°C ... 12
2. Effect of Intraventricular Amphetamine on Body Temperature of Unrestrained Rats at an Ambient Temperature of 25°C . . 13
3. Concentration of Amphetamine in Rat Brain Following Intraperitoneal and Intraventricular Administration at an Ambient Temperature of 25°C 15
A. Effect of Intraperitoneal Amphetamine on Body Temperature of Rats at an Ambient Temperature of 25°C 16
5. Effect of Intraperitoneal Amphetamine on Body Temperature of Rats at Various Ambient Temperatures 17
6. Effect of Intraventricular Amphetamine on Body Temperature of Rats at Various Ambient Temperatures 18
7. Effect of Phenoxybenzamine and Propranolol on Hypothermia Produced by Intraventricular Amphetamine in Rats at an Ambient Temperature of 25°C 19
8. Effect of Propranolol on Hyperthermia Produced by Intraperitoneal Amphetamine in Rats at Ambient Temperatures of 25° and 20°C 20
9. Effect of Phenoxybenzamine on Hyperthermia Produced by Intraperitoneal Amphetamine in Rats at Ambient Temperatures of 25° and 20°C 21
10. Effect of Intraperitoneal Amphetamine on Body Temperature Change in Pithed Rats at an Ambient Temperature of 26°C 22
11. Effect of Adrenalectomy on Hyperthermia Produced by Intraperitoneal Amphetamine at an Ambient Temperature of 2S°C 23
12. Effect of Intraperitoneal and Intraventricular Amphetamine on Oxygen Consumption Rate of Rats at an Ambient Temperature of 25°C 35
vii

viii
LIST OF ILLUSTRATIONS--Continued
Figure Page
13. Effect of Intraperitoneal Amphetamine on Spontaneous Activity of Rats at an Ambient Temperature of 25°C .... 36
14. Effect of Intraventricular Amphetamine on Spontaneous Activity of Rats at an Ambient Temperature of 25°C .... 37
15. Effect of Intraperitoneal and Intraventricular Amphetamine on Sniffing Behavior of Rats at an Ambient Temperature of 25°C 38
16. Body Temperature Response to Intraventricular Amphetamine at an Ambient Temperature of 25°C in Rats Pretreated with Ro 4-1284 or a Combination of Alpha-methyl-m-tyrosine (aMMT) and Alpha-methyl-p-tyrosine (aMPT) .... 56
17. Body Temperature Response to Intraperitoneal Amphetamine at an Ambient Temperature of 25°C in Rats pretreated with Ro 4-1284 or a combination of Alpha-methyl-m-tyrosine (aMMT) and Alpha-methyl-p-tyrosine (aMPT) .... 57
18. Effect of Intraventricular and Intraperitoneal Amphetamine on Fluorescence of Anterior Hypothalamic Nucleus Peri-vent ricularis in the Rat at an Ambient Temperature of 25°C 59
19. Effect of Intraventricular and Intraperitoneal Amphetamine on Fluorescence of Nucleus Caudatus Putamen (NCP) and Globus Pallidus (GP) in the Rat at an Ambient Temperature of 25°C 60
20. Peak Hypothermic Responses to Repeated Intraventricular Injections of Amphetamine at an Ambient Temperature of 25°C 61
21. Fluorescence in Anterior Hypothalamic Nucleus Periven-tricularis and Nucleus Caudatus Putamen at Time of Peak Hypothermia after Multiple Intraventricular Injections of A-nphetamine at an Ambient Temperature of 25°C 62
22. Fluorescence in Anterior Hypothalamic Nucleus Periven-tricularis at Time of Body Temperature Recovery after Multiple Intraventricular Injections of Amphetamine at an Ambient Temperature of 25°C 63

lx
LIST OF ILLUSTRATIONS--Continued
Figure Page
23. Fluorescence in Nucleus Caudatus Futamen at Time of Body Temperature Recovery after Multiple Intraventricular Injections of Amphetamine at an Ambient Temperature of 25°C 64

LIST OF TABLES
Table Page
1. Effect of Intraperitoneal Amphetamine on Body Temperature of Rats at an Ambient Temperature of 25°C 14
2. Effect of Ambient Temperature of 6°C, Propranolol and Adrenalectomy on Sniffing Behavior Following Intraperitoneal Amphetamine in Rats 39
3. Effect of Ro 4-1284 on Whole Brain Norepinephrine, Dopamine and 5-Hydroxytryptamine Concentrations in Rats at an Ambient Temperature of 25°C 54
4. Effect of p-Chlorophenylalanine and ?vO /:-1284 on Hypothermia Produced by Intraventricular Amphetamine at an Ambient Temperature of 25°C 55
5. Effect of Intraventricular and Intraperitoneal Amphetamine on Whole Brain Norepinephrine, Dopamine and 5-Hydroxy-tryptamine Concentrations in Rats at an Ambient Temperature of 25°C 58
x

ABSTRACT
There Is general agreement that amphetamine characteristically
produces hyperthermia in rats. However, there is disagreement whether
this effect is due to the central or peripheral action of the drug.
Dextroamphetamine was administered to rats by intraventricular or intra
peritoneal injection in an effort to resolve this controversy. Various
manipulative and pretreatment procedures were employed to modify dextro
amphetamine -induced body temperature responses to obtain evidence of the
relative roles of central and peripheral action of dextroamphetamine in
this regard. Additional studies were conducted to examine possible cor
relation between physiological or behavioral responses and temperature
changes and to examine certain endogenous neurochemicals as probable
mediators of the central hypothermic action of dextroamphetamine.
Intraventricular injection of dextroamphetamine in doses of 4,
100, and 400 meg at 25°C ambient temperature produced hyperthermia, hy
pothermia, or initial hypothermia followed by secondary hyperthermia,
respectively, whereas intraperitoneal injection of 2 mg/kg of the drug
produced hyperthermia. Intraperitoneal injection of 2 mg/kg of dextro
amphetamine and intraventricular Injection of 100 meg produced the same
brain concentration of the drug 30 minutes postadmlnistration. These
two doses are considered as "equivalent doses." Hypothermia appeared to
be the predominant central effect of dextroamphetamine, whereas hyper
thermia appeared to be mainly caused by the peripheral effect of dextro
amphetamine. It was suggested that systemic dextroamphetamine
xi

xll
simultaneously activates peripheral hyperthermic and central hypothermic
mechanisms. Support for this hypothesis was provided by observations
which suggest that high ambient temperature reduces body heat dissipa
tion and allows the hyperthermic action of the drug to become manifested,
whereas low ambient temperature enhances heat dissipation and allows the
hypothermic action of the drug to become manifested. Additionally, ad
renergic blocking agents appeared to antagonize the peripheral hyper
thermic effect of systemic dextroamphetamine and unmask the central
hypothermic effect of the drug. The predominant hyperthermic effect of
dextroamphetamine is apparently of peripheral origin because the drug
retards the rate of progressive hypothermia caused by ablation of the
central nervous system and because the absence of adrenal catecholamines
prevents the hyperthermic effect of dextroamphetamine.
Attempts to correlate physiological or behavioral responses and
dextroamphetamine-induced temperature changes yielded the initial im
pression that changes in spontaneous motor activity and oxygen consump
tion correlated with changes in body temperature, whereas sniffing
behavior did not. Subsequently, overt observations seemed to contradict
the initial findings and Implied a lack of correlation. It was ration
alized that various physiological or behavioral responses may be more
appropriately correlated with body heat generation or dissipation rather
than with changes in body temperature per se.
The probable role of norepinephrine, dopamine, and/or 5-hydroxy-
tryptamlne In the central hypothermic action of dextroamphetamine was
studied by observing the effect of depletion of these amines on the body
temperature effect of dextroamphetamine. All three of the biogenic

xiii
amines were simultaneously depleted with Ro 4-1284, 5-hydroxytryptamine
was depleted with p-chlorophenylalanine, and the two catecholamines were
simultaneously depleted with the combined use of alpha-methyl-m-tyrosine
and alpha-methyl-p-tyrosine. These depletion studies suggested that the
central hypothermic action of dextroamphetamine is mediated, at least in
part, by one or both of the catecholamines.
The Importance of one or both of the catecholamines in the cen
tral hypothermic effect of dextroamphetamine was further investigated
through the use of hlstochemlcal fluorescence analyses of specific areas
of the brain following single or repeated Injections of the drug. Of
the two catecholamines considered, norepinephrine appeared to be impor
tant in mediation of the central hypothermic action of dextroampheta
mine. This concept is particularly appealing because norepinephrine is
the predominant monoamine In the anterior hypothalamus and because this
region is considered to be important in thermoregulation.

CHAPTER 1
INTRODUCTION
Amphetamine has been repeatedly shown to elevate body tempera
ture (Haffner, 1938; Haas, 1939; Simonyi and Szentgyorgyi, 1949; Harrison,
Ambrus and Ambrus, 1952; Cheymol and Levassort, 1957; Lessin and Parkes,
1957; Klissiunis and Dosi, 1959; Tedeschi et al., 1959; Askew, 1962;
Belenky and Vltolina, 1962; Morpurgo and Theobald, 1965; Mantegazza,
Naimzada and Riva, 1968a; Gessa, Clay and Brodie, 1969; Slater and
Turnbull, 1969). However, the role of the central nervous system in
this response is not clearly defined. Amphetamine has been described as
having predominantly ergotropic activity (Brodie, Spector and Shore,
1959), thus leading to the conclusion that the hyperthermia may be pro
duced by stimulation of adrenergic receptors in the brain (Euler, 1961).
Indeed, elevation of brain amphetamine levels is reported to be associ
ated with a prolonged hyperthermic response in rats (Valzelli, Consolo
and Morpurgo, 1966; Valzelli et al., 1968). In an attempt to describe
the central site of action of amphetamine, Belenky and Vltolina (1962)
administered amphetamine to cats and rabbits in which lesions had been
produced at various levels in the brain. They concluded from their re
sults that the site of initiation of the hyperthermic response was in
the region of the diencephalon.
A different conclusion was reached by" Gessa, Clay and Brodie
(1969) who related amphetamine-induced hyperthermia to the mobilization
1

2
of free fatty acids through the release of norepinephrine from neurons
in adipose tissue. Since hyperthermia was not altered by a ganglionic
blocking agent these investigators concluded that amphetamine produces
hyperthermia by a peripheral action. Hence, a controversy exists re
garding the relative Importance of central and peripheral mechanisms in
amphetamine-induced hyperthermia.
The effect of other sympathomimetic amines on body temperature,
particularly the catecholamines, has also been studied. The nature of
body temperature changes induced by catecholamines appears to vary with
factors such as animal species and route of administration. For exam
ple, peripherally administered norepinephrine or epinephrine is reported
to produce hyperthermia in rats (Feldberg and Lotti, 1967; Jori ,
Paglialunga and Garattini, 1967) and rabbits (Masek and Raskova, 1964);
hypothermia in chicks (Allen and Marley, 1967) and monkeys (Essex,
1952); and both hyperthermia and hypothermia in dogs (Essex, 1952). In-
tracerebroventricular administration of these catecholamines is reported
to produce hyperthermia in rabbits (Euler, Linder and Myrin, 1943;
Cooper, Cranston and Honour, 1965) and sheep (Bllgh, 1966a); hypothermia
in mice (Brittain, 1966; Brittain and Handley, 1967), oxen (Findlay and
Thompson, 1968), cats (Feldberg and Myers, 1963, 1964a, 1964b), dogs
(Feldberg, Hellon and Myers, 1966), and monkeys (Feldberg, Hellon and
Lotti, 1967); and both hyperthermia and hypothermia in rats (Feldberg
and Lotti, 1967; Myers and Yaksh, 1968; Slater and Turnbull, 1969).
Since catecholamines are known to cross the blood brain barrier
with difficulty (Axelrod, Weil-Malherbe and Tomchlck, 1959; Weil-Malherbe,
Witby and Axelrod, 1961; Samorajski and Marks, 1962), it could be

3
justifiably argued that peripheral administration of catecholamines
produces hyperthermia by peripheral mechanisms whereas intracerebroven-
trlcular injection produces hyperthermia or hypothermia by central
mechanisms. On the other hand, dextroamphetamine, a noncatechol sympa
thomimetic amine, readily penetrates the blood brain barrier (Young and
Gordon, 1962; Fuller and Hines, 1967; Rosso, Dolfini and Franchl, 1968;
Malckel et al., 1969) and simultaneously produces central and peripheral
actions. Hence, a similar statement can not be made about dextroam
phetamine with respect to its body temperature effects and the contro
versy persists. The present project was conducted in order to help
resolve the controversy regarding the relative roles of central and pe
ripheral mechanisms in dextroamphetamine-Induced body temperature changes
in rats.

CHAPTER 2
THE EFFECT OF INTRAVENTRICULAR!^ AND INTRAPERITONEAL ADMINISTERED DEXTROAMPHETAMINE ON BODY TEMPERATURE OF RATS
Although numerous published investigations involving the periph
eral injection of amphetamine exist, relatively few investigations have
been reported in which the drug has been administered intracerebrally.
Intraventricular, intracisternal or intrahypothalamic administration of
amphetamine has been reported to produce lethargy, mydriasis, tachypnea
and panting (Gaddum and Vogt, 1956), excitation (Leimdorfer, 1950), ano
rexia (Booth, 1968), inhibition of phenylbenzoquinone-induced writhing,
piloerection, excitation (high dose) and depression (low dose) (Davis
and Horlington, 1964). However, a survey of the literature failed to
reveal reports concerning the influence of intraventricularly adminis
tered amphetamine on body temperature.
Since peripherally administered amphetamine has been hypothe
sized to cause hyperthermia by a central mechanism (Euler, 1961; Belenky
and Vitolinat 1962; Valzelli et al., 1966, 1968) and since proposed
thermoregulatory sites in the hypothalamus are in close proximity to
cerebrospinal fluid in the third cerebral ventricle (Bllgh, 1966b), it
appeared logical to use the intraventricular route of administration as
a means of studying the role of the central nervous system In ampheta
mine-induced body temperature changes. In the present study the tempera
ture responses to intraventricularly and Intraperitoneally administered
dextroamphetamine are compared. It was hoped that small doses of
4

5
dextroamphetamine given by intraventricular injection can produce prima
rily central actions with little or no peripheral actions.
Materials and Methods
Animals and Housing
Male Sprague-Dawley rats (Holtzman Co., Madison, Wisconsin),
weighing 225 to 275 g, were used throughout this Investigation and were
housed at 25 ±1°C with free access to water and food (Canine Checkers,
Ralston Purina Co.). Except for an Initial series of observations in
which restraint was employed (see Fig. 1), animals were removed from
their home cages for only brief periods during experimental manipula
tions such as injections or body temperature determinations. Animals
employed for the study reported in Fig. 1 were restrained in plastic rat
holders throughout the entire period of temperature determination. In
studies which Involved altered ambient temperatures, animals were trans
ferred to the new temperature environment In their home cages* Animals
were pre-conditioned at ambient temperatures of 30 + l°C, 6 ± 1°C and
20 +1°C for 1, 20 and 48 hours, respectively. The room in which experi
mental animals were housed was illuminated with incandescent lights from
6 A.M. to 6 P.M. daily and experiments were consistently performed be
tween 11 A.M. and 5 P.M.
Administration of Drugs
Drugs were administered by Intraventricular, Intraperitoneal and
subcutaneous injections. Intraventricular injections were made by means
of permanently Implanted cannulas in the right lateral cerebral

ventricle (Grunden and Linburn, 1969). Specifications of the materials
and details of the implantation and injection techniques are presented
in Appendix A. Details concerning the preparation of drug solutions for
this investigation are presented in Appendix B.
Measurement of Body Temperature
Body temperature was monitored by means of a telethermometer
(Yellow Springs Instrument Co.) with the thermistor probe inserted in
the rectum to a depth of 5 cm. Thirty-five seconds were allowed for
equilibration. Since handling and the procedures of injection are known
to cause alterations in body temperature (Brown and Julian, 1968), ani
mals were routinely subjected to body temperature determinations 3 times
at 30-minute intervals prior to dextroamphetamine or saline administra
tion. The third body temperature determination, made immediately before
injection, served as the reference temperature for subsequent body tem
perature changes.
Assay of Tritiated Dextroamphetamine
Rat6 were injected intraventricularly with 100 meg of tritiated
dextroamphetamine sulfate or intraperitoneally with 2 mg/kg of tritiated
dextroamphetamine sulfate. Thirty, 60 and 120 minutes after injection
the animals were killed and brain tissue was assayed for tritiated dex
troamphetamine sulfate. The assay was performed as described by Fuller
and Hines (1967). Details of the materials and procedure are presented
In Appendix C.

Surgical Techniques
Ablation of the central nervous system. Animals were anesthe
tized with methohexital and prepared by tracheal intubation for artifi
cial respiration. A large needle was quickly passed through the orbital
cavity into the brain and down the spinal column. The pithed animals
were then attached to a respirator (model V5KG, E & M Instrument Co.)
and respired at a rate of 66 cycles per minute with a 1:1 inspiration:
expiration ratio and an applied pressure of 15 cm water. The ambient
temperature was maintained at 26+0.2°C during this series of experi
ment s.
Adrenalectomy. Bilateral adrenalectomy was performed as de
scribed by D'Amour, Blood and Belden (1965). Animals were allowed to
recover at least two weeks before they were employed for experimental
studies.
Statistical Procedures
Data were statistically analyzed by means of Student's t_ test
(Snedecor and Cochran, 1967). Details of the calculations are presented
in Appendix D.
Results
Body Temperature Changes after Intraventricular Dextroamphetamine in Restrained Rats
Four meg of intraventricularly administered dextroamphetamine
resulted in hyperthermia in restrained rats, whereas 12, 20, or 100 meg
produced dose-related hypothermia (Fig. 1).

8
Body Temperature Changes after Intraventricular Dextroamphetamine in Unrestrained Rats
Four meg of intraventricularly administered dextroamphetamine
produced a modest but significant (P<0.01) increase in body temperature
at 30 minutes (Fig. 2). One hundred meg of dextroamphetamine produced
hypothermia which reached a peak at 30 minutes, showed partial recovery
at 60 minutes and returned to control levels at 120 minutes. In Com
parison, 400 meg of dextroamphetamine produced hypothermia no greater
than that produced by 100 meg at the 30-minute time period. However,
the body temperature of animals treated with the larger dose returned to
pre-injection levels more rapidly (in 60 minutes) and was elevated above
normal at 120 minutes.
Body Temperature Changes after Intraperitoneal Dextroamphetamine in Unrestrained Rats
Intraperitoneal administration of 100 meg of dextroamphetamine
produced no change in body temperature; whereas 200 meg and 400 meg pro
duced hyperthermia at 30 minutes postadministratlon (Table 1).
Brain Concentrations of Dextroamphetamine Sulfate
The brain concentrations of dextroamphetamine sulfate at se
lected times after intraventricular (100 meg) or intraperitoneal
(2 mg/kg) administration of dextroamphetamine sulfate are presented in
Fig. 3. At 30 minutes the brain concentrations for the 2 routes of ad
ministration were equivalent. By 60 minutes postadministratlon, the
mean brain concentration of dextroamphetamine after Intraventricular in
jection had decreased markedly, whereas the mean brain concentration
after Intraperitoneal injection was not significantly different

9
(P>0.05) from the 30-rainute value. By 120 minutes, the mean brain con
centration of dextroamphetamine after intraventricular injection was
still significantly less (P<0.01) than that after intraperitoneal
Injection, although both values were less than 20 per cent of the re
spective 30-minute values. Since these respective doses of dextroam
phetamine resulted in equivalent brain concentrations at 30 minutes
postadministration, they were chosen for further studies and will be re
ferred to as "equivalent" doses.
Comparison of Body Temperature Changes after Equivalent Intraperitoneal and Intraventricular Doses of Dextroamphetamine at Various Ambient Temperatures
Intraperitoneal administration of dextroamphetamine (2 mg/kg)
resulted in hyperthermia which reached a peak effect at 60 minutes
(Fig. 4). In a separate study involving the use of a series of ambient
temperatures (Fig. 5), the same dose of dextroamphetamine produced an
elevation of body temperature of approximately 1.1°C and 0.4°C at ambi
ent temperatures of 25°C and 20°C, respectively. In contrast, at an
ambient temperature of 6°C intraperitoneal dextroamphetamine injection
caused pronounced hypothermia. Control animals injected intraperito-
neally with saline solution showed relatively stable body temperatures
at these different environmental temperatures.
At an ambient temperature of 25°C, intraventricular injection of
dextroamphetamine produced hypothermia (Fig. 6). This hypothermic ef
fect was augmented by an ambient temperature of 6°C but was antagonized
by an ambient temperature of 30°C. Control animals given intraventricu
lar injections of saline showed moderate hyperthermia at an ambient

10
temperature of 30°C but showed no change In body temperature at ambient
temperatures of 25°C or 6°C.
Effects of Adrenergic Blocking Drugs on Body Temperature Responses to Dext roamphet amine
Propranolol In a dose of 10 mg/kg failed to block hypothermia
induced by intraventricular administration of dextroamphetamine at an
ambient temperature of 25°C (Fig. 7). On the other hand, it antagonized
the hyperthermic effect of peripherally administered dextroamphetamine
at an ambient temperature of 25°C and caused peripherally administered
dextroamphetamine to produce a moderate hypothermia, rather than hyper
thermia, at an ambient temperature of 20°C (Fig. 8).
Phenoxybenzamine in a dose of 10 mg/kg diminished by approxi
mately 50 per cent the hypothermia produced by intraventricular adminis
tration of dextroamphetamine at an ambient temperature of 25°C (compare
Figs. 7 and 6). Similar to propranolol, phenoxybenzamine also antago
nized the hyperthermic effect of intraperitoneal dextroamphetamine at an
ambient temperature of 25°C and reversed the temperature response pro
duced by intraperitoneal dextroamphetamine at an ambient temperature of
20°C (Fig. 9).
Effect of Pithing on Body Temperature Response to Dextroamphetamine
Destruction of the central nervous system of rats results in a
continuous fall in body temperature. Intraperitoneal administration of
dextroamphetamine perceptibly retarded this process (Fig. 10).

11
Effect of Adrenalectomy on Body Temperature Response to Dextroamphetamine
Bilateral adrenalectomy exerted no change on body temperature of
rats at an ambient temperature of 25°C. However, intraperitoneal admin
istration of dextroamphetamine In adrenalectomlzed rats failed to pro
duce hyperthermia (Fig. 11).
Discussion
Catecholamines administered intraventricularly to rats in low
doses produce moderate hyperthermia, whereas larger doses produce hypo
thermia (Feldberg and Lotti, 1967; Myers and Yaksh, 1968; Slater and
Turnbull, 1969). In the present study Involving restrained rats in
jected intraventricularly with dextroamphetamine, the lowest dose of
dextroamphetamine produced hyperthermia, whereas the larger doses (up to
100 meg) produced hypothermia. Similar results were obtained with in
traventricular injections of dextroamphetamine in unrestrained rats
(this and all subsequent body temperature studies were performed on un
restrained rats in order to permit possible correlation with data ob
tained from locomotor activity studies reported in a later chapter).
Very large doses of intraventricularly administered dextroamphetamine
produced three unexpected effects: (1) 400 meg of dextroamphetamine
produced no greater hypothermia than 100 meg, (2) hypothermia was of
shorter duration with the larger dose, and (3) with the larger dose a
secondary hyperthermia became apparent at 2 hours postadministratlon.
These anomalous effects on body temperature might be explained on the
basis of interplay between central hyperthermic and hypothermic mecha
nisms and peripheral hyperthermic mechanisms. Centrally mediated

12
O SALINE (N-3) • 4meg AMPHETAMINE {N-2> A I2mcg AMPHETAMINE (N«2) • 20meg AMPHETAMINE (N* 2) • 100meg AMPHETAMINE (N«3)
24
TIME (MINUTES)
Figure 1. Effect of Intraventricular Amphetamine on Body Temperature of Restrained Rats at an Ambient Temperature of 25°C
Dextroamphetamine sulfate or 0.9% NaCl solution was injected into the lateral cerebral ventricle in a volume of 5 mcl at time 0. The amount of dextroamphetamine is expressed as the sulfate salt.

13
O SALINE (N<7) * 4meg AMPHETAMINE (N«4) • 100meg AMPHETAMINE (N»5) • 400meg AMPHETAMINE (N>5)
U
z>
£ K ItJ a 3 UJ l->-o o m - 0 . 5
UJ o z X u
- 1 . 5
2.0
SO 60
TIME (MINUTES)
Figure 2. Effect of Intraventricular Amphetamine on Body Temperature of Unrestrained Rats at an Ambient Temperature of 25 C
Dextroamphetamine sulfate or 0.9% NaCl solution was Injected into the lateral cerebral ventricle in a volume of 5 mcl at time 0. The amount of dextroamphetamine is expressed as the sulfate salt. Four meg produced significant (P<0.01) hyperthermia at 30 minutes, whereas 100 and 400 meg produced significant (P<0.001) hypothermia at 30 minutes. By 60 minutes the value for 400 meg is not significantly different (P>0.05) from the value for saline-treated rats but is significantly different (P<0.05) from value for 100 meg. By 120 minutes the value for 400 meg is significantly higher (P<0.05) than control. Vertical lines indicate + S.E.M.

Table 1
Effect of Intraperitoneal Amphetamine on Body Temperature of Rats at an Ambient Temperature of 25°C
TREATMENT® CHANGE IN BODY TEMPERATURE (°C) (30 minutes after treatment)
Saline -0.13 + 0.16 (N=5)
100 meg Amphetamine -0.06 ± 0.09 (N=5 )
200 meg Amphetamine +0.46 + 0.06b
(N=4)
AOO meg Amphetamine +0.66 + 0.10c
(N=5)
a Dextroamphetamine sulfate (dose expressed as salt) or 0.9% NaCl solution was injected intraperitoneally in a volume of 2 ml/kg.
b Significantly different from saline treatment (P<0.05)
c Significantly different from saline treatment (P<0.01)

15
2.5
2.0
1.9
o o E 1.0
0.5
• AMPHETAMINE ip |N"B) A AMPHETAMINE ivt(N>6)
30 i
60 120 TIME(MINUTES)
Figure 3. Concentration of Amphetamine in Rat Brain Following Intraperitoneal and Inttfaventricular Administration at an Ambient Temperature of 25°C
Intraperitoneally (ip)» dextroamphetamine sulfate (2 mg/kg) was injected in a volume of 2 ml/kg at time 0. Intraventriculariy (ivt), dextroamphetamine sulfate (100 njcg) was injected in a volume of 5 mcl at time 0. The ordinate indicates ntcg of dextroamphetamine sulfate per g of tissue. Values for the 2 routes are not significantly different (P>0„5) at 30 minutes. At 60 and 120 minutes the values were significantly different from each other (P<0.001 and P<0.01, respectively). Vertical lines Indicate + S.E.M.

16
O SALINE (N>8)
• AMPHETAMINE (N* 7)
Ili
UJ
+ 0.5
-0.5
30 60
TIME (MINUTES)
Figure A. Effect of Intraperitoneal Amphetamine on Body Temperature of Rats at an Ambient Temperature of 25°C
Dextroamphetamine sulfate (2 rog/kg) or 0.9% NaCl solution was injected intraperitoneally in a volume of 2 ml/kg at time 0. Dextroamphetamine produced significant (P<0.001) hyperthermia at all time periods shown. Vertical lines indicate + S.E.M.

17
+ I .5
HI CC O I-< CC UJ 0. z LJ
>-Q O ffi
UJ o z « X o
+ I .0
+ 0 5
-0.5
- I 0
- 1.5
-2.0
O SALINE ,25*C (N>6) • AMPHETAMINE, 25*C (N-5)
& SALINE, 20'C <N«8> A AMPHETAMINE, Z0*C(N*8)
• SALINE, 6*C (N>5)
• AMPHETAMINE, 6*C (N>7)
-i 60 30
TIME( MINUTES)
Figure 5* Effect of Intraperitoneal Amphetamine on Body Temperature of Rats at Various Ambient Temperatures
Dextroamphetamine sulfate (2 mg/kg) or 0.9% NaCl solution was injected lntraperitoneally in a volume of 2 ml/kg at time 0. At ambient temperatures of 25° and 6 C dextroamphetamine-treated animals showed significant (P<0.001) alterations in body temperature 30 and 60 minutes after injection compared to the corresponding saline controls. At an ambient temperature of 20°C dextroamphetamine-treated animals showed a significant increase in body temperature 30 and 60 minutes after injection compared to saline controls (P<0.05 and P<0.001, respectively). Vertical lines indicate + S.E.M.

18
+ 0 5
0 SALINE, 30°C (N»4) • AMPHETAMINE, 30* C(N«4|
A SALINE, 25*C (N-7) * AMPHETAMINE, 25*C (N-S>
• SALINE, 6* C (N» 5) • AMPHETAMINE, 6*C(N"6)
30 60
TIME (MINUTES)
Figure 6. Effect of Intraventricular Amphetamine on Body Temperature of Rats at Various Ambient Temperatures
Dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution was injected into the lateral cerebral ventricle in a volume of 5 mcl at time 0. At ambient temperatures of 25° and 6°C dextroamphetamine-treated animals showed significant (P<0.001) reductions in body temperature compared to saline controls. Vertical lines indicate * S.E.M.

19
+ 0.5 -
O PHENOXYBENZAMINE + SALINE (N-4» • PHENOXYBENZAMINE + AMPHETAMINE tN-SI
A PROPRANOLOL + A PROPRANOLOL +•
SALINE (N«4) AMPHETAMINE (N-6)
60
TIME (MINUTES) 120
Figure 7. Effect of Phenoxybenzamine and Propranolol on Hypothermia Produced by Intraventricular Amphetamine in Rats at an Ambient Temperature of 25°C
Dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution was injected into the lateral cerebral ventricle in a volume of 5 mcl at time 0. Phenoxybenzamine hydrochloride (10 mg/kg) was injected intraperitoneally 5 hours before dextroamphetamine or saline; propranolol hydrochloride (10 mg/kg) was injected subcutaneously 30 minutes before dextroamphetamine or saline. In both phenoxybenzamine and propranolol pretreated animals, dextroamphetamine produced significant (P<0.001) hypothermia at 30 minutes postadministration. Vertical lines indicate t S.E.M.

20
O SALINE, 25°C (N = 3) • AMPHETAMINE, 25"C (N=3)
A SALINE, 20°C (Na4) ^ • AMPHETAMINE, 20°C (N-4)
UJ a: o
£ + 0 . 5 a: UJ CL 5 UJ I-
5 o CQ
z - 0 . 5
UJ e> z < X o
60 30
TIME (MINUTES)
Figure 8. Effect of Propranolol on Hyperthermia Produced by Intraperitoneal Amphetamine in Rats at Ambient Temperatures of 25° and 20 C
Dextroamphetamine sulfate (2 mg/kg) or 0.9% NaCl solution was Injected intraperltoneally in a volume of 2 ml/kg at time 0. Propranolol hydrochloride (10 mg/kg) was injected subcutaneously 30 minutes before dextroamphetamine or saline. At an ambient temperature of 20°C dextro-amphetamlne-treated animals showed a significant decrease in body temperature 30 and 60 minutes after injection compared to saline controls (P<0.001 and P<0,05, respectively). At an ambient temperature of 25°C dextroamphetamine-treated animals showed no significant (P>0.05) change in body temperature compared to saline controls. Vertical lines indicate ± S.E.M.

21
O SALINE, 25*C (N«5) • AMPHETAMINE, 25'C (N-5)
A SALINE, 20#C (N-4)
A AMPHETAMINE, 20*C (N»4)
U t. + 10
Ul oc
a 00 UJ +0 5 Q. 5 Ul I-
>-Q O CD
Z
UJ e> z < X
- 0 5
o
30 60
TIME (MINUTES)
Figure 9. Effect of Phenoxybenzamine on Hyperthermia Produced by Intra peritoneal Amphetamine in Rats at Ambient Temperatures of 25° and 20 C
Dextroamphetamine sulfate (2 mg/kg) or 0.9% NaCl solution was injected intraperitoneally in a volume of 2 mg/kg at time 0. Phenoxybenzamine hydrochloride (10 mg/kg) was injected intraperitoneally 5 hours before dextroamphetamine or saline. At an ambient temperature of 20°C dextroamphetamine -treated animals showed a significant (P<0.05) decrease in body temperature 60 minutes after injection compared to saline control. At an ambient temperature of 2S°C dextroamphetamine-treated animals showed a significant (P<0.05) increase in body temperature 30 minutes after injection compared to saline control. Vertical lines indicate t S.E.M.

22
SALINE (N-S) AMPHETAMINE (N>5)
U • r Ui ac => Si oc UJ a. z UJ l—
-2
>-O o ffi z
UI CD z x -4 O
-5
10 20 60 30 40 50
TIME AFTER PITHING(MINUTES)
k Figure 10. Effect of Intraperitoneal Amphetamine on Body Temperature Change in Pithed Rats at an Ambient Temperature of 26°C
Dextroamphetamine sulfate (2 ml/kg) or 0.9% NaCl solution was injected intraperitoneally In a volume of 2 ml/kg immediately after pithing. Dextroamphetamine significantly retarded the process of hypothermia due to pithing at the 40-, 50- and 60-minute time periods (P<0.01) P<0.01 and P<0.05, respectively). Vertical lines indicate + S.E.M.

23
O SALINE (N-6) • AMPHETAMINE (N-5)
A SALINE, Adrtnalcclomiztd (N«3) A AMPHETAMINE, Adrenol«ctomiz6d IN"4)
O
UJ
5
£ 0. 2 UJ H
+0.5
5 3 Z
UJ o z < X o
-0.5
60
TIME (MINUTES)
Figure 11. Effect of Adrenalectomy on Hyperthermia Produced by Intraperitoneal Amphetamine at an Ambient Temperature of 25°C
Dextroamphetamine sulfate (2 mg/kg) or 0.9% NaCl solution was injected intraperitoneally in a volume of 2 ml/kg at time 0. Body temperature changes for dextroamphetamine and saline in non-adrenalectomized animals are taken from Fig. 5. Body temperature response to dextroamphetamine in adrenaiectomlzed animals is not significantly different (P>0.5) from saline control. Vertical lines indicate + S.H.M.

hyperthermic and hypothermic mechanisms have been demonstrated by sev
eral investigators (Keller, 1963; Jacobson and Squires, 1964; Rewerskl
and Jori, 1967). The present study suggests that both of these central
mechanisms are activated by dextroamphetamine and catecholamines but
that the hyperthermic mechanism is more sensitive and may be activated
by low doses of sympathomimetic amines, whereas the hypothermic mecha
nism requires larger doses. Once Initiated, however, the hypothermic
response appears to mask the centrally-induced hyperthermic response.
Although intraventricular doses of 100 meg and 400 meg of dex
troamphetamine both produce hypothermia, the shorter duration and
secondary hyperthermia associated with the larger dose may be due to
peripherally-induced hyperthermia consequent to escape of dextroampheta
mine into the systemic circulation. This explanation seems plausible
since a peripheral hyperthermic mechanism which responds to sympathomi
metic amines has been reported (Gordon et al., 1966; Bernard!, Paglialunga
and Jori, 1968; Cowell and Davey, 1968; Cowell, 1969; Gessa, Clay and
Brodie, 1969). The failure of 100 meg of dextroamphetamine intraven
tricular^ to produce secondary hyperthermia may have resulted from
inadequate escape into the systemic circulation. This suggestion of
inadequate escape is supported by a lack of hyperthermia following In
traperitoneal administration of 100 meg of dextroamphetamine and the
production of hyperthermia following intraperitoneal administration of
both 200 meg and 400 meg of dextroamphetamine. The above hypothesis im
plies that dextroamphetamine in adequate doses, regardless of route of
administration, may simultaneously activate central hyperthermic, cen
tral hypothermic and peripheral hyperthermic mechanisms.

25
To teBt this proposed central-peripheral Interaction of dextro
amphetamine, body temperature changes In rats were studied following the
Intraperitoneal Injection of a dose of dextroamphetamine (2 mg/kg) which
yielded the same brain concentration of drug as that produced by intra
ventricular administration of 100 meg of the amine. Since body tempera
ture represents a balance between heat production by the body and heat
loss from the body (Du Bois, 1939; Mount, 1960; Hammel and Hardy» 1963;
Borison and Clark, 1967; Hemingway and Price, 1968), altered ambient
temperatures were employed for the purpose of modifying this balance.
Variation of the ambient temperature produced no changes In body tem
perature of control animals injected lntraperitoneally with normal sa
line. In comparison, intraperitoneal Injection of dextroamphetamine
caused an elevation or a reduction of body temperature which was depen
dent upon the environmental temperature. Furthermore, intraventricular
injection of an equivalent dose of dextroamphetamine produced a reduc
tion of body temperature at all ambient temperatures tested. These
observations suggest that systemic injection of dextroamphetamine simul
taneously activates both hyperthermic and hypothermic mechanisms, thus
allowing the ambient temperature to become an important determinant of
body temperature. On the other hand, intraventricular injection of dex
troamphetamine, at the dose employed, predominantly activates central
hypothermic mechanisms, thus alteration of ambient temperature does not
qualitatively affect body temperature changes.
Adrenergic blocking agents were more effective In antagonizing
hyperthermia induced by peripherally Injected dextroamphetamine than hy
pothermia Induced by intraventrlcularly injected dextroamphetamine. At

26
an ambient temperature of 20°C propranolol and phenoxybenzamlne not only
prevented hyperthermia induced by peripheral administration of dextroam
phetamine but modified the response so that hypothermia ensued. This
"reversal" of body temperature change is presumably the result of re
moval of peripheral opposition (i.e., hyperthermia) to the central hypo
thermic component of dextroamphetamine action.
Pithing produced a precipitous fall In body temperature which
was effectively retarded by peripheral administration of dextroampheta
mine. This observation further supports the concept of a peripheral hy
perthermic action of dextroamphetamine since the central nervous system
is obliterated in pithed animals.
Because peripheral administration of epinephrine produced hyper
thermia (Masek and Raskova, 1964; Feldberg and Lottl, 1967) and because
dextroamphetamine has been shown to release epinephrine from the adrenal
gland (Rubin and Jaanus, 1966; Harvey, Sulkowski and Weenlg, 1968), it
was Important to Investigate the role which endogenous epinephrine might
play in hyperthermia produced by peripherally administered dextroam
phetamine. Since adrenalectomy blocked the hyperthermic response to
dextroamphetamine It is suggested that epinephrine liberated from the
adrenal medulla plays an important role in dextroamphetamine-induced hy
perthermia (perhaps through the release of free fatty acids). Because
adrenocortlcosterolds potentiate free fatty acid release by epinephrine
(Ingle, 1956; Reshef and Shapiro, 1960; Shafrlr and Steinberg, 1960),
the possibility exists that total adrenalectomy may interfere with dex
troamphetamine-induced hyperthermia through deprivation of corticoster
oids. However, since adrenalectomized rats showed normal growth without

27
supplementation with exogenous corticosteroids or saline and since rats
are known to have accumulations of tissues'similar to adrenocortical
tissue (Gorbman and Bern, 1962), it appears that there is no deficiency
of corticosteroids. Therefore, lack of adrenal epinephrine appears im
portant in preventing dextroamphetamine-induced hyperthermia.
The data presented in this section support the hypothesis that
dextroamphetamine may simultaneously activate central hyperthermic, cen
tral hypothermic and peripheral hyperthermic mechanisms.

CHAPTER 3
BEHAVIORAL AND PHYSIOLOGICAL FACTORS ASSOCIATED WITH DEXTROAMPHETAMINE-INDUCED BODY TEMPERATURE CHANGES
Numerous behavioral and physiological changes occur in associa
tion with alteration of body temperature (Du Bois, 1939; Euler, 1961).
Both peripheral and intracerebral administrations of amphetamine, which
alter body temperature, produce many behavioral, physiological and bio
chemical changes. Peripheral administration of amphetamine has been re
ported to produce stimulation of motor activity (Greenblatt and Osterberg,
1961; Moore, 1964; Rech, 1964; Stein, 1964; Morpurgo and Theobald, 1965;
Smith, 1965; Wenzel and Broadie, 1966; Clark, Blackman and Preston,
1967), stereotyped sniffing and gnawing (Mennear, 1965; Randrup and
Scheel-Kruger, 1966: Ernest, 1967; Fog, Randrup and Pakkenberg, 1967;
Morpurgo and Theobald, 1967; Randrup and Munkvad, 1967; Arnfred and
Randrup, 1968), elevation of oxygen consumption (Gyermek, 1950; Obal,
Mozes and Erdei, 1955; Zalls, Lundberg and Knutson, 1967), hypoglycemia
(Moore, Sawdy and Shaul, 1965; Moore, 1966a, 1966b; Clark et al., 1967),
hyperglycemia (Mantegazza, Naimzada and Rlva, 1968b), increased fat mo
bilization (Fasslna, 1966; Gessa, Clay and Brodie, 1969), depletion of
glycogen stores (Moore, 1966a) and alterations of tissue and plasma
electrolyte concentrations (Moore, 1966c). Intraventricular, intracls-
ternal or intrahypo thai antic administration of amphetamine has been re
ported to produce lethargy, mydriasis, tachypnea and panting (Gaddum and
Vogt, 1956), excitation (Leimdorfer, 1950), anorexia (Booth, 1968),
28

29
inhibition of phenylbenzoqulnone-induced writhing, piloerection, excita
tion (high dose) and depression (low dose) (Davis and Horlington, 1964).
i Two of the above parameters have been widely studied as causal
factors in drug-induced body temperature changes. Some investigators
have suggested that increased oxygen consumption or motor activity is
responsible for hyperthermia after administration of amphetamine or
other central nervous system stimulants (Sokoloff et al., 1957; Hardinge
and Peterson, 1964; Morpurgo and Theobald, 1965; Clark et al., 1967;
Zalis et al., 1967). Other investigators report poor correlation be
tween body temperature changes and increases in oxygen Consumption or
motor activity (Allen and Marley, 1967; Mount and Willmott, 1967;
Mantegazza et al.» 1968a; Turner and Spencer, 1968; Gessa, Cho, Clay,
Tagllamonte and Brodle, 1969). In view of this controversy, It was
considered important to study behavioral and physiological responses
which occurred In association with alterations of body temperature fol
lowing intraventricular injection of dextroamphetamine. Another justi
fication for such evaluations is provided by the apparent association
between behavioral and physiological events and alterations of body tem
perature during heating or cooling of the hypothalamus. For example,
elevation of hypothalamic temperature has been reported to produce de
pression (Hemingway et al., 1940; Euler, 1961). Kahn (1904) reported an
"almost narcotic effect" from heating carotid blood. Local hypothalamic
heating also causes peripheral vasodilation (Hemingway et al., 1940) and
reduction of body temperature (Magoun et al., 1938; Andersson, Ekman,
Gale and Sundsten, 1963; Andersson, Gale and Ohga, 1963; Hammel et al.,
1963; Edinger, Elsenman and Brobeck, 1969). On the other hand,

30
hypothalamic cooling causes peripheral vasoconstriction (Kruger et al.,
1959; Hamrnel, Hardy and Fusco, 1960; Andersson et al., 1964), elevation
of oxygen consumption (Jacobson and Squires, 1964) and elevation of body
temperature (Andersson, Ekman, Gale and Sundsten, 1963; Andersson, Gale
and Ohga, 1963). Since intraventricular injection of dextroamphetamine
has been shown to reduce body temperature it was considered important to
investigate the effect of Intraventricular injection of dextroampheta
mine on oxygen consumption and spontaneous activity. In addition,
stereotyped sniffing induced by peripheral administration of dextroam
phetamine was compared with the response induced by intraventricular ad
ministration of dextroamphetamine.
Materials and Methods
Where applicable, materials and methods used in Chapter 2 were
also used in this section. All studies in this section were performed
at an ambient temperature of 25°C. Techniques unique to this section
are described below.
Measurement of Oxygen Consumption
Oxygen consumption was determined with a Minute Oxygen Uptake
Servo Equipment Spirometer (model 160, Custom Engineering and Develop
ment Co*, St. Louis, Missouri). Animals were placed into the upper por
tion of a glass desiccator which was maintained at 25 t1°C by passing
cooled air (21-22°C) over the sealed chamber. The consumed oxygen was
replaced by the spirometer through a tubing connected to the desiccator
lid, and expired carbon dioxide and water were absorbed by potassium
hydroxide solution (187L) and a calcium sulfate-soda lime mixture.

31
Animals were protected from these chemicals by two layers of copper
screen (18 mesh). Oxygen consumption was measured for two consecutive
15-minute intervals, beginning 5 minutes after injection of saline or
dextroamphetamine. The rate of oxygen consumption was expressed as
ml/kg/mln.
Measurement of Spontaneous Activity
Spontaneous motor activity of individual rats was recorded with
an Ultra-Sonic/Electrostatic Grid Motion Detector (Alton Electronics Co.,
Gainesville, Florida). The home cage was placed in a sound-Insulated
box, and the transmitting and receiving transducers were positioned in
the box but outside of the cage. Sensitivity of the motion detector was
adjusted to record head movements but not breathing movements. Sponta
neous activity was continously recorded by means of a digital counter.
Cumulative spontaneous activity was noted at the end of each 10-mlnute
interval during a 1 hour period following saline or dextroamphetamine
injection.
Quantification of Sniffing Behavior
Evaluation of sniffing behavior was made by observing the number
of seconds per 60-second time period that the animal was engaged in
sniffing activity. Sniffing activity was defined as exploratory move
ments associated with alternating dorsi- and ventro-flexion of the neck.
Results
Approximately 5 minutes after dextroamphetamine was administered
lntraventricularly animals displayed a typical posture, of approximately

32
15 minutes duration, characterized by an elongation of the body with the
ventral surface of the body flattened against the floor of the cage.
Animals showed little mobility during this period. This effect was not
seen after intraperitoneal administration of dextroamphetamine or after
intraventricular or intraperitoneal administration of saline.
Effects of Dextroamphetamine on Oxygen Consumption
Oxygen consumption was significantly (P<0.05) increased (25%)
during the second 15-minute time interval following intraperitoneal ad
ministration of dextroamphetamine (2 mg/kg), although no alteration of
oxygen consumption was seen during the first 15-mlnute measurement (Fig.
12). Oxygen consumption was significantly (P<0.05) decreased (18%)
during the first 15-minute time interval following intraventricular ad
ministration of dextroamphetamine (100 meg), but oxygen consumption dur
ing the second 15-minute time interval was not different from control
values.
Effects of Dextroamphetamine on Spontaneous Motor Activity
Intraperitoneally administered dextroamphetamine increased cumu
lative spontaneous activity approximately 300 per cent during the 60-
mlnute observation (Fig. 13). The spontaneous activity of control
animals was highest during the initial (exploratory) 10 minute counting
interval and then declined with each succeeding interval to a very low
level* In contrast, the spontaneous activity of drug-treated animals
showed a high level (comparable to the initial value for control ani
mals) which was sustained throughout the 60-minute observation. In the

33
final measurement Intervals, activity of the drug-treated animals was
greater than for saline controls.
Intraventricularly administered dextroamphetamine had very lit
tle effect on cumulative spontaneous activity (reduction of 16%) during
the 60-minute observation period (Fig. 14). The spontaneous activity of
control animals was highest during the Initial 10-minute counting inter
vals and then declined with each succeeding interval to a very low
level. In contrast, the spontaneous activity of drug-treated animals
was low (25% that of the corresponding control value) during the initial
10-minute interval. This level of activity was maintained throughout
the 60-minute observation, however, and during the final measurement in
tervals activity of the drug-treated animals was greater than for saline
controls.
Evaluation of Sniffing Behavior
The administration of dextroamphetamine by either intraperito
neal or intraventricular injection stimulated sniffing behavior (Fig.
15). Intraperitoneal injection of dextroamphetamine caused almost con
tinuous sniffing behavior during the 30- and 60-minute time periods.
This response was diminished approximately 50 per cent at 120 minutes
postadmlnistration. Intraventricular injection of dextroamphetamine
also stimulated sniffing behavior during the 30- and 60-minute time peri
ods, but to a lesser degree. At 120 minutes postadmlnistration, the
sniffing activity had subsided and the animals treated intraventricu
larly with dextroamphetamine behaved no differently than saline-injected
controls. Testing at an ambient temperature of 6°C, pretreatment with

34
propranolol, or adrenalectomy did not alter the sniffing behavior pro
duced by intraperitoneal dextroamphetamine (Table 2).
Discussion
It has long been recognized that constancy of body temperature
in homeothermic animals is maintained by a balance between heat genera
tion and heat loss (Du Bols, 1939; Mount, 1960; Hammel and Hardy, 1963;
Borison and Clark, 1967; Hemingway and Price, 1968). Changes in oxygen
consumption rate and in spontaneous motor activity are evaluated in the
present study since these functions are associated with changes in heat
generative processes. The rate of oxygen consumption, which is a
reflection of the overall metabolic rate, is increased when heat gen
erative processess are stimulated and decreased when heat generative
processes are depressed.
Intraperitoneal administration of dextroamphetamine, which in
duces hyperthermia, caused an increase in oxygen consumption. These
observations are in agreement with the reports of Gyermek (1950) and
Zalis et al. (1967) and imply that a causal relationship may exist
between oxygen consumption rate and body temperature changes. That
increased oxygen consumption may be an important factor in dextroam-
phetamlne-induced hyperthermia is further suggested by the report that
adrenalectomy, which prevented dextroamphetamine-induced hyperthermia
(see previous Chapter), reduced the elevation of oxygen consumption
caused by administration of racemic amphetamine (Gyermek, 1950). Fur
thermore, in the present investigation intraventricular administration
of dextroamphetamine produced both hypothermia and a decrease in oxygen

35
40
SO
ZO
I 0
O* JC
INTRAPERITONEAL
ill rfl
10
I
Jl
10 MLINE AMPHETAMINE •AUttE AMPHETAMINE
«o
Jo
O " o
Ld (9 >o >-
O
10
INTRAVENTRICULAR
10
\h
•ALINE AMPHETAMINE
(»; 9-20 mfo )
k
to
h
•AUHK AMPHETAMINE
(1:20-59 mlit)
Figure 12. Effect of Intraperitoneal and Intraventricular Amphetamine on Oxygen Consumption Rate of Rats at an Ambient Temperature of 25°C
Intraperitoneally, dextroamphetamine sulfate (2 mg/kg) or 0.9% NaCl solution was injected in a volume of 2 ml/kg. Intraventricularly, dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution was injected in a volume of 5 mcl. Oxygen consumption rate was determined during 2 consecutive 15-minute intervals, 5 to 20 minutes and 20 to 35 minutes after Injection (t: 5-20 min and t: 20-35 min, respectively). Number in box equals N. Adenotes significant difference from corresponding saline control (P<0.05). Vertical lines indicate + S.E.M.

36
• SALINE CN-5)
AMPHETAMINE (N>6)
10
9
? 8 O
X 7
CO h-Z z> o o
o <
6
5
4
> 3
2
I » #
r*
V.
ft
V.
K<
J: \V
i;
& o-lb 10-20 20-30 30-4o 40-30 90-60
TIME(MINUTES)
Figure 13. Effect of Intraperitoneal Amphetamine on Spontaneous Activity of Rats at an Ambient Temperature of 25°C
Dextroamphetamine sulfate (2 mg/kg) or 0.9% NaCl solution was injected intraperitonally in a volume of 2 ml/kg at time 0. Spontaneous activity was cumulated for each 10-minute interval for I hour, o denotes significant difference from saline control (P<0.01). • denotes significant difference from saline control (P<0.001). Vertical lines indicate + S.E.M.
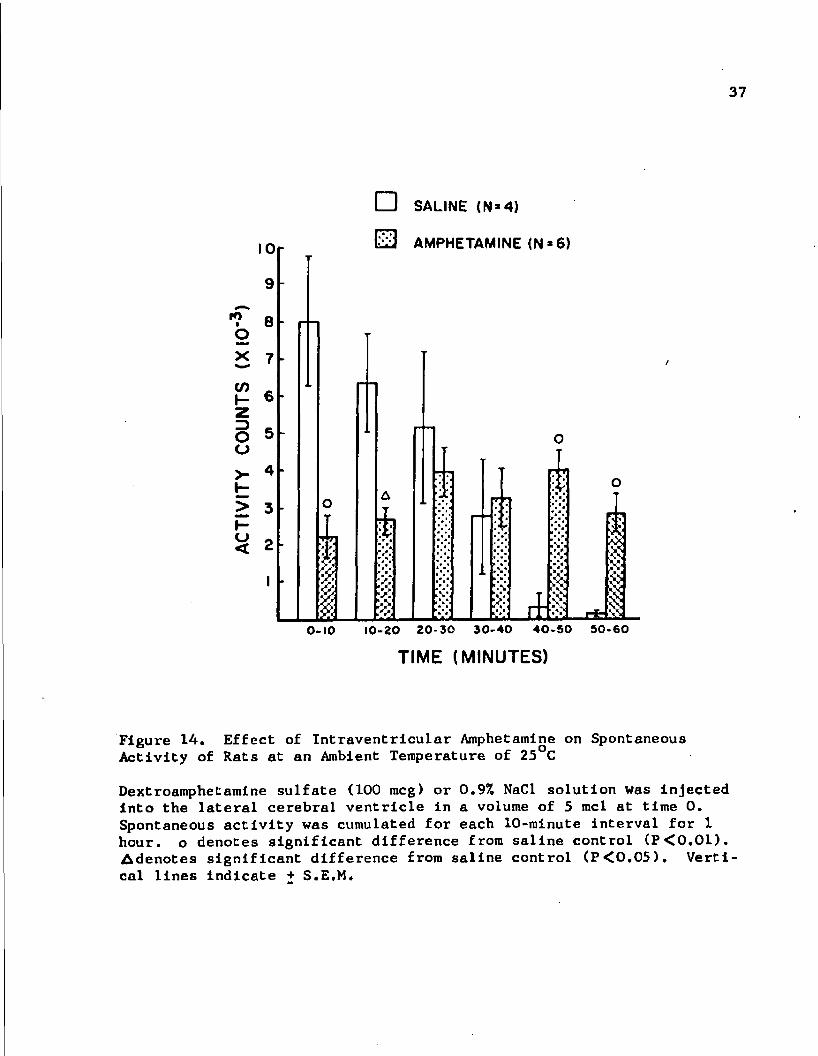
37
10
9
8
>< 7
ro
O
CO I- 1
O 5 o
>- 4 K
> 3 I-<-> 0 < 2
I
I
0-10
• SALINE (N»4)
AMPHETAMINE (N»6)
y.
& $
A 3 $
1 10-20 20-30 30-40 40-50 50-60
TIME (MINUTES)
Figure 14. Effect of Intraventricular Amphetamine on Spontaneous Activity of Rats at an Ambient Temperature of 25 C
Dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution was injected Into the lateral cerebral ventricle In a volume of 5 mcl at time 0. Spontaneous activity was cumulated for each 10-minute interval for 1 hour, o denotes significant difference from saline control (P<0.01). Adenotes significant difference from saline control (P<0.05). Vertical lines indicate + S.E.M.

38
O SALINE ip (N*8) • AMPHETAMINE ip (N• 10)
£ SALINE ivt (N*9) • AMPHETAMINE ivl (N 9)
UJ 30
60
TIME( MINUTES) 120
Figure 15. Effect of Intraperitoneal and Intraventricular Amphetamine on Sniffing Behavior of Rats at an Ambient Temperature of 25°C
Intraperltoneally (ip), dextroamphetamine sulfate (2 mg/kg) or 0.9X NaCl solution was Injected in a volume of 2 ml/kg at time 0. Intraventricular^ (lvt), dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution was injected Into the lateral cerebral ventricle in a volume of 5 tncl at time 0. Intraperitoneal dextroamphetamine produced significant (P<0.001) enhancement of sniffing behavior at all time periods. Intraventricular dextroamphetamine produced significant enhancement at the 30- and 60-minute time periods (P<0.001 and P<0.01, respectively). Vertical lines indicate + S.E.M.

39
Table 2
Effect of Ambient Temperature of 6°C, Propranolol and Adrenalectomy on Sniffing Behavior Following Intraperitoneal Amphetamine in Rats
TREATMENT® SNIFFING SCORE (seconds in 60)
Saline (25°C)c 3 + 2b
(N=8 )
Amphetamine (25°C) 56+1 ( N= 10 )
Saline (6 C) 2*1 ( N=3 )
Amphetamine (6°C) 58+2 (N=7)
Propranolol + Saline (25°C) 1+1 (N=3 )
Propranolol + Amphetamine (25°C) 56+1 (N=4)
Adrenalectomy + Saline (25°C) 2+1 (N=3 )
Adrenalectomy + Amphetamine (25°C) 56 J 1 (N=4)
a Dextroamphetamine sulfate (2 rag/kg) or 0.97. NaCl solution was injected intraperitoneally in a volume of 2 ml/kg 30 minutes before sniffing score evaluation. Propranolol hydrochloride (10 mg/kg) was injected subcutaneously 30 minutes before saline or dextroamphetamine.
b Value represents mean sniffing score i- S.E.M.
c Ambient temperature

40
consumption. This last observation provides an additional basis for
suggesting that the body temperature effects of dextroamphetamine are at
least partially due to its actions on oxygen consumption rate.
Spontaneous motor activity of test animals is presumably the re
sult of interaction between environmental factors (i.e., manipulations,
process of injection, familiarization with the new environment of the
counting box) and dextroamphetamine treatment. Hence, the difference
between each test result and its corresponding control result should
represent the drug effect, a positive value Indicating stimulation and a
negative value indicating depression of spontaneous motor activity.
Consideration of these calculated values reveal that intraperitoneal in
jection of dextroamphetamine caused stimulation of spontaneous motor ac
tivity which developed gradually and became pronounced and sustained.
In comparison, intraventricular injection of dextroamphetamine caused a
biphasic response. Spontaneous motor activity was initially depressed,
gradually returned to near normal by the third and fourth observation
periods, and was moderately stimulated during the remainder of the test.
These spontaneous motor activity observations appear to correlate with
dextroamphetamine-induced body temperature changes noted In the previous
Chapter. It was noted that at an environmental temperature of 25°C In
traperitoneal Injection of dextroamphetamine produced hyperthermia which
reached a peak value at 30 minutes postadministration and was sustained.
On the other hand, intraventricular injection of dextroamphetamine pro
duced hypothermia which reached a peak value at 30 minutes postadmlnis-
tration and was partially recovered at 60 minutes postadministration.

41
It is also noteworthy that moderate local heating of the hypo
thalamus causes sedation (Magoun et al.» 1938; Andersson, Gale and Ohga,
1963; Hammel et al., 1963; Edinger et al., 1969). These observations
lend further support to the concept of correlation between depression
of spontaneous activity and the production of hypothermia.
Sniffingp a characteristic dextroamphetamine-induced behavioral
effect, can be readily evaluated without disturbing the animal. This
parameter was quantified during some of the body temperature investiga
tions. Both intraventricularly and intraperitoneally administered dex
troamphetamine produced an increase in sniffing behavior, the duration
of which appeared to correspond with the sojourn of dextroamphetamine in
brain tissue (see Fig. 4). Sniffing behavior seems to be unrelated to
changes in body temperature since treatments which prevent hyperthermia
produced by peripherally injected dextroamphetamine did not affect snif
fing behavior. This 1b not unexpected since sniffing behavior is re
ported to be a centrally-mediated phenomenon (Laursen, 1962; Ernest and
Smelik, 1966; Fog, 1967), whereas as pointed out in Chapter 2, dextroam
phetamine -induced hyperthermia may result from activation of peripheral
mechanisms.
Of the 3 physiological and behavioral parameters which were
evaluated in this section, oxygen consumption rate and spontaneous motor
activity show the best correlations with dextroamphetamine-induced tem
perature changes. Since changes in spontaneous motor activity may be at
least partially responsible for corresponding changes in oxygen consump
tion rate, it is not surprising that they both correlate with body
temperature changes produced by dextroamphetamine. In view of the

apparently similar effects produced by moderate local heating of the
hypothalamus and Intraventricular Injection of dextroamphetamine (1.
sedation and hypothermia), the hypothalamus Is suggested as an lmpor
tant site for the hypothermic action of dextroamphetamine.

CHAPTER 4
THE ROLE OF CENTRAL MONOAMINES IN DEXTROAMPHETAMINE-INDUCED HYPOTHERMIA
The sympathomimetic activity of amphetamine is prevalently
thought to be mediated through the libration of norepinephrine (Burn
and Rand, 1958; Burn, I960; Trendelenberg» 1963; Harvey et al., 1968).
The role of norepinephrine in the action of amphetamine on the central
nervous system is less clearly established (Van Rossum, Van der Schoot
and Hurkmans, 1962; Moore and Lariviere, 1963, Moore, 1964; Stein, 1964;
Carlsson, Linqvist, Dahlstrom, Fuxe and Masuoka, 1965; Hanson, 1966;
Mennear and Rudzik, 1966; Corrodi, Fuxe and Hokfelt, 1967; Littleton,
1967; Morpurgo and Theobald, 1967). Since dextroamphetamine releases
norepinephrine from brain tissue following intraventricular administra
tion (Carr and Moore, 1969a) and peripheral administration (Moore,
1963a, 1963b, 1964; Smith, 1965; Littleton, 1967; Fuxe and Ungerstedt,
1968; Gropetti and Costa, 1969; Welch and Welch, 1969), and since intra
ventricular administration of norepinphrlne causes alterations of body
temperature (Feldberg and Lottl, 1967; Myers and Yaksh, 1968) similar to
those following intraventricular administration of dextroamphetamine
(see Chapter 2), it would appear that norepinephrine may be a central
mediator In the hypothermic action of dextroamphetamine.
Another catecholamine, dopamine, has been implicated as a media
tor of various behavioral effects of dextroamphetamine (Van Rossum and
Hurkmans, 1964; Ernest and Smelik, 1966; Randrup and Scheel-Kruger,
43

44
1966; Ernest, 1967; Fog, 1967). Administered intraventricularly it
caused hyperthermia (Myers and Yaksh, 1968). Dextroamphetamine has been
shown to release dopamine into the ventricular perfusate after intraven
tricular administration (Carr and Moore, 1969b). Peripheral administra
tion of dextroamphetamine has been reported to increase (Smith, 1965;
Lewander, 1968a; Littleton, 1967; Welch and Welch, 1969) and decrease
(Lewander, 1968b; Welch and Welch, 1969) brain concentrations of this
catecholamine. The increase was seen following Injection of small doses
of dextroamphetamine whereas the decrease was seen following injection
of large or repeated doses.
A third monoamine which might be a mediator of dextroampheta-
mine-induced hypothermia is 5-hydroxytryptamine. This amine has been
reported to produce hypothermia when it is administered by the intra
ventricular route to rats (Feldberg and Lotti, 1967; Myers and Yaksh,
1968). In addition, brain concentrations of 5-hydroxytryptamine have
been reported to be elevated by dextroamphetamine (McLean and McCartney,
1961; Smith, 1965; Welch and Welch, 1969).
Different types of studies have been used to relate alterations
in body temperature with endogenous chemicals. For example, drugs which
alter brain norepinephrine, dopamine and 5-hydroxytryptamine levels were
administered and body temperature was subsequently monitored (Ingenito
and Bonnycastle, 1967a; Somerville and Whittle, 1967; Shellenberger and
Elder, 1967, 1968). In general, studies of this type have shown poor
correlations between whole brain amine levels and alteration of body
temperature.

45
Another approach has been to expose animals to altered environ
mental temperature and monitor tissue levels or turnover of suspected
mediators. These studies report norepinephrine synthesis In peripheral
tissues to be enhanced at reduced environmental temperatures (Ollverio
and Stjarne, 1965; Corrodi and Malmfors, 1966; Goldstein and Nakajima,
1966; Gordon et al., 1966). Some studies have reported little or no
change In metabolism of brain norepinephrine and 5-hydroxytryptamine
during cold exposure (Corrodi, et al., 1967; Reid et al., 1968; Ingenito
and Bonnycastle, 1967b; Ingenito, 1968). Others have reported the con
centration of brain norepinephrine to be reduced in cold-stressed ani
mals (Levi and Maynert, 1964; Maynert and Levi, 1964). With heat
stress, the metabolism of brain norepinephrine and 5-hydroxytryptamine
is increased (Corrodi et al., 1967; Shellenberger and Elder, 1967; Reid
et al., 1968; Simmonds and Iversen, 1969).
An additional approach has involved direct administration of the
proposed mediator. Work of this type has gained wide acceptance since
Feldberg and Myers (1964a) proposed the concept that the body tempera
ture is related to a balance between 5-hydroxytryptamine and norepineph
rine in the hypothalamus.
Two of the principles described above will be applied in the
present investigation. In one approach, tissue levels of catecholamines
and/or 5-hydroxytryptamine will be reduced by commonly used depleting
agents* and dextroamphetamine will then be administered intraventricu
lar^ to determine the effect of monoamine defecit on the usual hypo
thermic effect. This investigation will attempt to establish which, if
any, of these monoamines are important in mediating the body temperature

46
response Induced by intraventricular administration of dextroampheta
mine. The drugs used for depleting tissue catecholamines and 5-hydroxy
tryptamine are Ro 4-1284, p-chlorophenylalanine, and a combination of
alpha-methyl-m-tyrosine with alpha-methyl-p-tyrosine. Ro 4-1284, a ben-
zoquinolizine, produces a rapid reduction of brain norepinephrine, dopa
mine and 5-hydroxytryptamine levels. The levels of these monoamines are
maximally reduced from 1 to 3 hours postadministration and are recovered
to approximately 85 per cent of normal by 12 hours postadministration
(Pletscher, 1957; Quinn, Shore and Brodie, 1959; Pletscher, Burkard and
Gey, 1964; Carlsson and Lindqvist, 1966; Pletscher and Da Prada, 1966;
Pletscher and Bartholini, 1967; Jobe, 1970). p-Chlorophenylalanine, a
hydroxylase inhibitor which is more selective for tryptophan hydroxylase
than for tyrosine hydroxylase (Koe and Weissman, 1966), produces a
marked reduction of brain 5-hydroxytryptamine levels but only a modest
reduction of brain catecholamines from 1 to 4 days postadministration
(Koe and Weissman, 1966; Jequier, Lovenberg and Sjoerdsma, 1967;
Lovenberg, Jequier and Sjoerdsma, 1968; Sato et al., 1967; Welch and
Welch, 1967; Jobe, 1970). Alpha-methyl-m-tyrosine, 12 hours postadmin
istration, reduced brain norepinephrine and dopamine levels to less than
60 per cent of normal while leaving brain 5-hydroxytryptamine at near
normal levels (Hess et al., 1961; Porter, Totaro and Leiby, 1961;
Carlsson, Falck and Hillarp, 1962; Carlsson, Dahlstrom, Fuxe and Hillarp,
1965; Fuxe, 1965; Shore, Alpers and Busfield, 1966; Jobe, 1970). Alpha-
methyl-p-tyrosine, a tyrosine hydroxylase inhibitor, reduced brain nor
epinephrine and dopamine levels (Nagatsu, Levitt and Udenfriend, 1964;
Levitt et al., 1965; Rech, Borys and Moore, 1966; Spector, Sjoerdsma and

Udenfriend, 1965; Weissman and Koe, 1967; Jobe, 1970). The combination
of alpha-methyl-m«tyrosine and repeated doses of alpha-methyl-p-tyrosine
has been reported to produce marked lowering of brain norepinephrine and
dopamine with no effect on brain 5-hydroxytryptaraine (Jobe, 1970).
Another approach used in this section of the investigation in
volves a study of the effect of dextroamphetamine administration on
brain monoamine levels and its relationship to dextroamphetamine-induced
hypothermia. Repeated systemic administrations of dextroamphetamine at
short intervals cause the development of tachyphylaxis to peripheral re
sponses* a phenomenon which is associated with decreases in peripheral
catecholamine levels (Burn and Rand, 1958; Cession-Fossion, 1963; Harvey
et al., 1968). Analogous studies to observe the effect of repeated in
traventricular injections of dextroamphetamine on centrally mediated
hypothermia and on brain monoamine levels may provide additional infor
mation regarding the relative Importance of the various monoamines in
dextroamphetamine-induced hypothermia.
Materials and Methods
Where applicable, materials and methods used in Chapter 2 were
also used in this section. All studies in this section were performed
at an ambient temperature of 25°C. Techniques unique to this section
are described below.
Effects of Monoamine-depleting Agents on Dextroamphetamine-induced Body Temperature Changes
Monoamine-depleting agents were Injected into rats, an appro
priate interval of time was allowed to elapse, dextroamphetamine was

48
injected intraventricularly or Intraperitoneally, and body temperature
was determined. Simultaneous depletion of brain norepinephrine, dopa
mine and 5-hydroxytryptamine was accomplished by administration of
Ro 4-1284 (10 mg/kg) 1 hour before testing with dextroamphetamine.
Twelve-hour pretreatment with the benzoquinollzine was employed to de
termine the body temperature effect of dextroamphetamine at a time when
monoamines have recovered to near normal levels. Whole brain monoamine
levels were assayed by the spectrophotofluorometric technique described
below. Depletion of brain 5-hydroxytryptamine (with modest depletion of
catecholamines) was induced by intraperitoneal injection of p-chloro-
phenylalanine (300 mg/kg) 3 days before testing with dextroamphetamine.
Simultaneous depletion of both norepinephrine and dopamine was achieved
by combination treatment with a single dose of alpha-methyl-m-tyrosine
(500 mg/kg) and repeated doses of alpha-methyl-p-tyrosine (80 mg/kg).
The former drug was injected 12 hours and the latter drug was Injected
12, 8 and 4 hours before testing with dextroamphetamine.
Brain Monoamine Levels after Single Intraventricular or Intraperitoneal Administration of Dextroamphetamine
Rats were injected intraventricularly or intraperitoneally with
saline or dextroamphetamine solutions as described previously. Whole
brain concentrations of norepinephrine, dopamine, or 5-hydroxytryptamine
of some animals were analyzed 30 minutes postadministration by the spec
trophotof luorometric technique described below. Various parts of brain
from other animals were analyzed for monoamine concentration by the his-
tochemical fluorescence technique described below.

49
Effects of Repeated Intraventricular Injections of Dextroamphetamine on Hypothermic Response
Rats were injected intraventricularly with saline or dextroam
phetamine and body temperature responses were recorded at various times.
When the body temperature of dextroamphetamine-treated animals returned
to preinjectlon levels, saline and dextroamphetamine treatments were re
peated in control and test animals, respectively, and body temperature
was again monitored. This procedure was repeated until hypothermia was
nonsignificant.
Brain Monoamine Levels at Peak Hypothermic Effect Induced by the Last of Three Repeated Intraventricular Injections of Dextroamphetamine
Rats were administered 3 intraventricular injections of saline
or dextroamphetamine solution at 2-hour intervals (corresponding to time
for recovery from hypothermia). Thirty minutes after the final injec
tion, a time which corresponds to peak hypothermia, the anterior hypo
thalamic nucleus periventricularis and nucleus caudatus putamen were
analyzed by the histochemical fluorescence technique described below.
Brain Monoamine Levels at Time of Recovery of Body Temperature after Repeated Intraventricular Injections of Dextroamphetamine
Rats were administered up to 4 intraventricular injections of
saline or dextroamphetamine solution at 2-hour intervals between injec
tions 1, 2 and 3 and at a 1-hour interval between Injections 3 and 4.
These time intervals, including the 1-hour interval after the fourth in
jection, correspond to the time of recovery from hypothermia Induced by
multiple injections of dextroamphetamine. Fluorescence in the anterior
hypothalamic nucleus periventricularis and nucleus caudatus putamen was

50
analyzed at the following times: (1) before dextroamphetamine or saline
injection, (2)2 hours after the second dextroamphetamine or saline in
jection, and (3) 1 hour after the fourth dextroamphetamine or saline
injection.
Spectrophotofluorometric Assay of Catecholamines and 5-Hydroxytryptamine
A spectrophotofluorometric technique was used for biochemical
determinations of whole brain norepinephrine, dopamine and 5-hydroxy-
tryptamine. The extraction technique was a modification of the methods
described by Wiegand and Perry (1961) and Mead and Finger (1961). Assay
procedures for detection of norepinephrine, dopamine and 5-hydroxytryp
tamine were similar to those of Shore and Olin (1958), Carlsson and
Waldeck (1958) and Bogdanski et al. (1956), respectively. A detailed
account of the spectrophotofluorometric procedure is presented in Appen
dix £.
Histochemical Fluorescence Technique
The histochemical fluorescence technique for viewing norepineph
rine, dopamine and 5-hydroxytryptamine used in the present work is a
modification of the method reported by Falck and Owman (1965). The
areas of the brain employed in the present studies included the anterior
hypothalamic nucleus periventricularis, the fluorescence of which is
primarily norepinephrine (Falck, 1964; Dahlstrom, Fuxe and Hillarp, 1965;
Friede, 1966); the nucleus caudatus putamen, the fluorescence of which
is primarily due to dopamine (Fuxe, 1965); and the globus pallidus, the
fluorescence of which is primarily due to 5-hydroxytryptamine (Fuxe,

51
Hokfelt and Ungerstedt, 1968). A description of the histochemical tech
nique Is presented in Appendix F.
Results
Effects of Monoamine-depleting Agents on Dextroamphetamine-induced Body Temperature Changes
One-hour pretreatment with Ro 4-1284 reduced whole brain norepi
nephrine, dopamine and 5-hydroxytryptamine by 59, 72 and 73 per cent,
respectively (Table 3). This pretreatment antagonized the hypothermic
effect of intraventricularly administered dextroamphetamine (Table 4,
compare Fig. 16 with Fig. 2) and the hyperthermic effect of intraperito-
neally administered dextroamphetamine (compare Fig. 17 with Fig. 4).
Antagonism of the hypothermic action of dextroamphetamine was no longer
apparent 12 hours after pretreatment (Table 4). Pretreatment with
p-chlorophenylalanine did not affect the body temperature change induced
by intraventricular administration of dextroamphetamine (Table 4).
Catecholamine depletion by combination pretreatment with alpha-methyl-m-
tyroslne and alpha-methyl-p-tyrosine partially antagonized both the
hypothermic effect of intraventricularly administered dextroamphetamine
(compare Fig. 16 with Fig. 2) and the hyperthermic effect of intraperi-
toneally administered dextroamphetamine (compare Fig. 17 with Fig. 4).
Brain Monoamine Levels after Single Intraventricular or Intraperitoneal Administration of Dextroamphetamine
Intraventricular or intraperitoneal administration of dextroam
phetamine produced no significant (P>0.1) alteration in whole brain
concentration of norepinephrine, dopamine or 5-hydroxytryptamine 30

52
minutes after injection (Table 5). However, fluorescence in the ante
rior hypothalamic nucleus periventricularis was noticeably reduced 30
minutes after injection and was normal by 60 minutes postadministration
(Fig. 18). Administration of dextroamphetamine by either intraventricu
lar or intraperitoneal injection produced no change in fluorescence in
the nucleus caudatus putamen or globus pallidus 30 minutes after treat
ment (Fig. 19).
Effects of Repeated Intraventricular Injections of Dextroamphetamine on Hypothermic Response
Repeated intraventricular injections of dextroamphetamine pro
duced a peak hypothermic response, 30 minutes after each Injection,
which was progressively less pronounced (Fig. 20). The fifth injection
of dextroamphetamine produced a hypothermic response in test animals
which was not significantly different (P>0.1) from that of correspond
ing saline-treated controls (V, Fig. 20). It was also observed that
recovery of body temperature to preinjection level required 2 hours fol
lowing the first and second dextroamphetamine injections. This recovery
time was reduced to 1 hour following the third, fourth and fifth Injec
tions of dextroamphetamine.
Brain Monoamine Levels at Peak Hypothermic Effect Induced by the Last of Three Repeated Intraventricular Injections of Dextroamphetamine
Analysis of brain tissue 30 minutes after the last of 3 repeated
intraventricular injections of dextroamphetamine revealed reduction in
fluorescence in the anterior hypothalamic nucleus periventricularis
(compare A with B, Fig. 21). Triple intraventricular injection of dex
troamphetamine appears to be more effective in reducing fluorescence in

53
the hypothalamus than a single intraventricular injection of dextroam
phetamine (compare B» Fig. 21 with B, Fig. 18). In the case of the
nucleus caudatus putamen, 3 repeated intraventricular injections of dex
troamphetamine appear to have no effect on fluorescence (compare A1 with
B', Fig. 21).
Brain Monoamine Levels at Time of Recovery of Body Temperature Following Repeated Intraventricular Injections of Dextroamphetamine
Analysis of brain tissue at a time when body temperature had
recovered to preinjection levels following repeated intraventricular in
jections of dextroamphetamine revealed a progressive reduction of fluo
rescence in the anterior hypothalamic nucleus periventricularis (compare
A, B* and C1, Fig. 22). Fluorescence In the hypothalamus of controls
was also progressively reduced by repeated intraventricular injections
of saline solution (compare A, B and C, Fig. 22), but the reduction In
fluorescence was more pronounced in dextroamphetamlne-treated animals.
In contrast, fluorescence In the nucleus caudatus putamen was unchanged
by repeated intraventricular Injections of saline or dextroamphetamine
solution (Fig. 23).
Discussion
Ro 4-1284 markedly depleted norepinephrine, dopamine and 5-hy-
droxytryptamine and antagonized the hypothermic action of intraventricu-
larly administered dextroamphetamine. This antagonism was lost at a
time when brain levels of these monoamines were apparently recovered to
near normal. These observations imply that deficiency of 1 or more of
these monoamines may be responsible for the reduced effect of

54
Table 3
Effect of Ro 4-1284 on Whole Brain Norepinephrine, Dopamine and 5-Hydroxytryptamine Concentrations in Rats
at an Ambient Temperature of 25°C
TREATMENT3 NOREPINEPHRINE DOPAMINE 5-HYDROXYTRYPTAMINE (mcg/g tissue) (mcg/g tissue) (mcg/g tissue)
Saline 0.33 ± 0.02b 0.86 0.03 0.52 ± 0.03
Ro 4-1284 0.08 ± 0.01c 0.24 t 0.03° 0.14 + 0.02c
a Ro 4-1284 (10 mg/kg) or 0.9% NaCl solution was injected intraperito-neally 1 hour before sacrifice.
b Values represent mean concentration ± S.E.M. N equals 4 for all determinations.
c Significantly different from corresponding values In saline-treated animals (P<0.01).

Table 4
Effect of p-Chlorophenylalanine and Ro 4-1284 on Hypothermia Produced by Intraventricular Amphetamine at an
Ambient Temperature of 25°C
PRETREATMENT CHANGE IN BODY TEMPERATURE (°C) (30 minutes after amphetamine)3
Saline (1 hr)c -2.05 ±0.15b
(N=4)
Ro 4-1284 (1 hr)c -0.40 t 0.19e
(N=4)
Saline (12 hr)c -1.72 + 0.17 (N=4)
Ro 4-1284 (12 hr)c -1.45 ± 0.13 (N=6)
Vehicle (3 da)d -2.00 ± 0.18 (N=3)
p-Chlorophenylalanine (3 da)d -1.62 ± 0.27 (N=10)
a Dextroamphetamine sulfate (100 meg) was injected into the lateral cerebral ventricle in a volume of 5 mcl.
b Mean change in body temperature + S.E.M.
c Ro 4-1284 (10 mg/kg) or 0.9% NaCl solution was injected intraperitoneally 1 or 12 hours before dextroamphetamine .
d p-Chlorophenylalanine (300 mg/kg) or suspension vehicle (0.4% Tween 80, 1% carboxymethylcellulose, 0.97. NaCl) was injected intraperitoneally 3 days before dextroamphetamine.
e Value taken from data in Fig. 16. Significantly different from saline control (P<0.001)

56
A R04-I284 4 SALINE (N-4) A R04-L264 + AMPHETAMINE (N-4)
• O.MMT and aMPT + SALINE (N-4) • o»MMT ond aMPT + AMPHETAMINE (N-5) U
+ 0.5 Ui oz
6 ac UJ a. 2 UJ H
>•
§ "0 5 CO
z UJ o z < X o
60 30
TIME(MINUTES)
Figure 16. Body Temperature Response to Intraventricular Amphetamine at an Ambient Temperature of 25°C in Rats Pretreated with Ro 4-1284 or a Combination of Alpha-methyl-m-tyrosine (aMMT) and Alpha-methyl-p-tyrosine (aMPT)
Ro 4-1284 (10 mg/kg) was injected intraperitoneally 1 hour before saline or amphetamine. aMMT (500 mg/kg) was injected intraperitoneally 12 hours before saline or amphetamine; aMPT (80 mg/kg) was injected intraperitoneally 12, 8 and 4 hours before saline or amphetamine. Dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution was injected into the lateral cerebral ventricle in a volume of 5 mcl at time 0. Vertical lines indicate ± S.E.M.

57
O o
UJ cc. 3
5 a: UJ o. 2E UJ
>-o o ffi
UJ o z < u
A R04-IZ84 + SALINE (N-3) A R04-I284 + AMPHETAMINE (N»7> • ammf and ampt + SALINE (N>3) • am ml and ampt + AMPHETAMINE <N«6>
+ 0.5
-0.5
i 30
—i 60
TIME (MINUTES)
Figure 17. Body Temperature Response to Intraperitoneal Amphetamine at an Ambient Temperature of 25°C in Rats Pretreated with Ro 4-1284 or a Combination of Alpha-methyl-m-tyrosine (aMMT) and Alpha-methyl-p-tyrosine (aMPT)
Ro 4-1284 (10 mg/kg) was injected intraperitoneally I hour before saline or amphetamine. aMMT (500 mg/kg) was injected intraperitoneally 12 hours before saline or amphetamine; aMPT (80 mg/kg) was injected intraperitoneally 12, 8 and 4 hours before saline or amphetamine. Dextroamphetamine sulfate (2 mg/kg) or 0.97a NaCl solution was injected intraperitoneally in a volume of 2 ml/kg at time 0. Vertical lines indicate ± S.E.M.

58
Table 5
Effect of Intraventricular and Intraperitoneal Amphetamine on Whole Brain Norepinephrine, Dopamine and 5-Hydroxytryptamine Concentrations in Rats at an Ambient Temperature of 25°C
TREATMENT NOREPINEPHRINE DOPAMINE 5-HYDROXYTRYPTAMINE (mcg/g tissue) (mcg/g tissue) (mcg/g tissue)
Saline (ivt)a 0.36 ± 0.03c 0.91 ± 0.06 0.49 ± 0.03
Amphetamine (ivt)a 0.40 + 0.02 1.01 + 0.04 0.50 + 0.02
Saline <ip)b 0.37 + 0.02 0.78 + 0.06 0.44 ± 0.02
Amphetamine (ip)b 0.42 + 0.03 1.01 + 0.11 0.48 + 0.01
a Dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution was injected into the lateral cerebral ventricle in a volume of 5 mcl 30 minutes before sacrifice.
b Dextroamphetamine sulfate (2 mg/kg) or 0.9% NaCl solution was injected lntraperitoneally in a volume of 2 ml/kg 30 minutes before sacrifice.
c Mean concentration ± S.E.M. N equals 4 to 6 for each determination.

Figure 18. Effect of Intraventricular and Intraperitoneal Amphetamine on Fluorescence of Anterior Hypothalamic Nucleus Periventricularis in the Rat at an Ambient Temperature of 25°C
Intraventricularly (ivt), dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution in a volume of 5 mcl was injected in A, B, C and D. Intraperitoneally (ip), dextroamphetamine sulfate (2 mg/kg) or 0.9% NaCl solution in a volume of 2 mg/kg was injected in A', B', C* and D'.
Key to figure:
A 30 min after ivt saline B 30 min after ivt amphetamine C 60 min after ivt amphetamine D 120 min after ivt amphetamine
A' 30 min after ip saline B* 30 min after ip amphetamine C1 60 min after ip amphetamine D1 120 min after ip amphetamine

59
IV T IP
A
B
c
D
Figure 18. Effect of Amphetamine on Hypothalamic Fluorescence

Figure 19. Effect of Intraventricular and Intraperitoneal Amphetamine on Fluorescence of Nucleus Caudatus Putamen (NCP) and Globus Pallidus (GP) in the Rat at an Ambient Temperature of 25°C
Intraventricularly (ivt), dextroamphetamine sulfate (1Q0 meg) or 0,9% NaCl solution in a volume of 5 mcl was injected in A, Bt C and D 30 minutes before sacrifice. Intraperitoneally (ip), dextroamphetamine sulfate (2 mg/kg) or 0.9% NaCl solution in a volume of 2 ml/kg was injected in A1, B', C' and D1 30 minutes before sacrifice.
Key to figure:
A ivt saline B ivt amphetamine
A' ip saline B' ip amphetamine
C ivt saline D ivt amphetamine
C' D'
ip saline ip amphetamine

60
IVT IP
A
e
NC!It
c
Figure 19. Amphetamine on Nucleus Caudatus Putamen and Globus Pallidus

61
+ 0 . 5
o
LJ £E
5 00 UJ a. Z UJ l-
>-o o CO
tu o
< X u
HI IS
- 0 . 5
- I .0
- I .5
3
1 1
t
-2.0 I
$3
>X
A-F
V* 'A
#
i
I i-S
<F
t£J AMPHETAMINE (N-8>
• SALINE (N-4)
Figure 20. Peak Hypothermic Responses to Repeated Intraventricular Injections of Amphetamine at an Ambient Temperature of 25°C
Dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution in a volume of 5 mcl was injected into the lateral cerebral ventricle at 2-hour intervals for doses 2 and 3 and at 1-hour Intervals for doses 4 and 5. These times correspond to the time of body temperature recovery from dextroamphetamine -induced hypothermia. Values in the figure represent the mean body temperature change + S.E.M. 30 minutes after saline or amphetamine injection. All hypothermic effects except that following injection V are significantly different (P<0.001) from saline control.

62
A
B
Figure 21. Fluorescence in Anterior Hypothalamic Nucleus Periventricularis and Nucleus Caudatus Putamen at Time of Peak Hypothermia after Multiple Intraventricular Injections of Amphetamine at an Ambient
0 Temperature of 25 C
Dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution in a volume of 5 mel was injected into the lateral cerebral ventricle at 2-hour intervals for 3 injections. These times correspond to the time of body temperature recovery from dextroamphetamine-induced hypothermia. Animals were sacrificed 30 minutes after third injection.
Key to figure:
A hypothalamus (saline) A' caudate nucleus (saline) B hypothalamus (amphetamine) B' caudate nucleus (amphetamine)

A
B
c c'
Figure 22. Fluorescence in Anterior Hypothalamic Nucleus Periventricularis at Time of Body Temperature Recovery after Multiple Intraventricular Injections of Amphetamine at an Ambient Temperature of 25°C
63
Dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution in a volume of 5 mel was injected into the lateral cerebral ventricle at 2-hour intervals for doses 2 and 3 and at 1-hour interval for dose 4. These times, including 1 hour after the fourth dose, correspond to the time of body temperature recovery from dextroamphetamine-induced hypothermia.
Key to figure:
A no injection
B 2 hours after second saline injection B' 2 hours after second amphetamine injection
c 1 hour after fourth saline injection C' 1 hour after fourth amphetamine injection

A
B a'
c
Figure 23. Fluorescence in Nucleus Caudatus Putamen at Time of Body Temperature Recovery after Multiple Intraventricular Injections of Amphetamine at an Ambient Temperature of 25°C
Dextroamphetamine sulfate (100 meg) or 0.9% NaCl solution in a volume
64
of 5 mel was injected into the lateral cerebral ventricle at 2-hour intervals for doses 2 and 3 and at 1-hour interval for dose 4. These times, including 1 hour after the fourth dose, correspond to the time of body temperature recovery from dextroamphetamine-induced hypothermia.
Key to figure:
A no injection
B 2 hours after second saline injection B' 2 hours after second amphetamine injection
c 1 hour after fourth saline injection C' 1 hour after fourth amphetamine injection

dextroamphetamine on body temperature. Marked reduction of brain 5-hy
droxy try pt amine concentrations with slight reduction of catecholamine
levels (1.e., p-chlorophenylalanine pretreatment) failed to produce an
tagonism of the hypothermic effect produced by intraventricular injec
tion of dextroamphetamine. In contrast, depletion of catecholamines
without depletion of 5-hydroxytryptamine (I.e., alpha-methyl-ni-tyroslne
and alpha-methyl-p-tyrosine pretreatment) caused approximately AO per
cent reduction In the hypothermic response induced by intraventricular
injection of dextroamphetamine. These data suggest that the catechola
mines may participate in mediating the central hypothermic action of
dextroamphetamine and that 5-hydroxytryptamine may be of little or no
Importance In this regard.
A number of published observations lend support to the pre
sumption that a central hypothermic action of dextroamphetamine is me
diated through the release of endogenous catecholamines. For example,
it has been demonstrated that intraventricular injection of norepineph
rine can cause hypothermia (Feldberg and Lottl, 1967; Myers and Yaksh,
1968) and that injection of dextroamphetamine into the cerebrospinal
space releases norepinephrine into the cerebrospinal fluid (Carr and
Moore, 1969a). Also, dextroamphetamine releases catecholamines in such
a manner as to form metabolites which are produced primarily by cate
chol -o-methyl transferase rather than by monoamine oxidase (Glowlnskl,
Snyder and Axelrod, 1965; Glowlnskl, Axelrod and Iversen, 1966; Fuxe and
Ungerstedt, 1968), suggesting that these released endogenous catechola
mines have access to receptor sites In the brain.

66
If catecholamines are to be accepted as possible mediators of
dextroamphetamine-lnduced hypothermia, it is important to correlate al
teration in concentration or turnover of catecholamines in brain tissue
with reduction in body temperature. Thirty minutes after intraventricu
lar or intraperitoneal administration of dextroamphetamine) spectropho-
tofluorometric analyses showed no change in the concentration of whole
brain norepinephrine, dopamine and 5-hydroxytryptamine. However, histo-
chemlcal fluorescence analyses of specific areas of the brain demonstra
ted a reduction of norepinephrine in the anterior hypothalamic nucleus
periventricularis, but no change in the concentration of dopamine in the
nucleus caudatus putamen or of 5-hydroxytryptamine in the globus palli-
dus. The anterior region of the hypothalamus is considered important in
thermoregulation (Meyer, 1913; Teague and Ranson, 1936; Jacobson and
Squires, 1961; Bligh, 1966b), and reduction of norepinephrine in this
area correlates with peak hypothermia induced by intraventricular injec
tion of dextroamphetamine. The observation that intraperitoneal admin
istration of dextroamphetamine also reduces norepinephrine levels in the
hypothalamus appears to coincide with the postulation that systemic ad
ministration not only activates peripheral hyperthermic mechanisms but
also central hypothermic mechanisms (see Chapter 2).
Analyses of the nucleus caudatus putamen and globus pallidus
were based on the assumption that if dopamine and/or 5-hydroxytryptamine
were mediators of dextroamphetamine in production of hypothermia,
changes in amine level in these areas would reflect similar changes in
brain areas involved with thermoregulation, On the basis of this as
sumption, there is no change in the concentration of dopamine or

67
5-hydroxytryptamlne in thermoregulatory areas during dextroamphetamine -
induced hypothermia. Of the 3 amines under consideration, it would ap
pear that norepinephrine is the most likely candidate as a mediator for
hypothermia produced by dextroamphetamine.
The importance of endogenous norepinephrine in the central hypo
thermic action of dextroamphetamine is further supported by the observa
tions involving repeated intraventricular injections of dextroamphetamine
at close intervals. Each injection of drug produced a fall in body tem
perature which was less pronounced than the preceding response. After
the fifth injection the hypothermic response was nonsignificant. In
conjunction with the body temperature study, 1 set of hlstochemlcal
fluorescence observations revealed that norepinephrine concentration in
the hypothalamus was less 30 minutes after the third intraventricular
injection of dextroamphetamine than 30 minutes after a single intraven
tricular injection of dextroamphetamine. A sequence of 3 injections
also gave rise to less reduction in body temperature than did the sin
gle injection. As in the case following a single injection of dextro
amphetamine, repeated Injections produced no change in the concentration
of dopamine in the nucleus caudatus putamen. Another set of observa*
tions revealed that successive intraventricular administrations of dex
troamphetamine, after recovery from hypothermia, caused a progressive
decline of norepinephrine concentration in the hypothalamus but no
change in the concentration of dopamine in the nucleus caudatus puta-
men. The progressive depletion of norepinephrine appears to correlate
well with the progressive decrease in hypothermic response.

68
The observations reported in this Chapter do not completely rule
out dopamine or 5-hydroxytryptamine as mediators for dextroamphetamine
in the production of hypothermia. There is a possibility that dopamine
and/or 5-hydroxytryptamine concentrations may be reduced in brain areas
involved in thermoregulation and yet such changes may not be reflected
by analyses of the nucleus cadatus putamen or the globus pallidus.
Also, the turnover rate of certain blochemicals has shown correlation
with physiological phenomena in the absence of absolute change in con
centration (Garattini, Glacolone and Valzelli, 1967; Engel et al., 1968;
Reid et al*, 1968; Slmmonds and Iversen, 1969). Such a relationship may
exist for dopamine or 5-hydroxytryptamine with regard to dextroampheta
mine -induced hypothermia. However, of the 3 monoamines considered, the
evidence presented in this study provides most support for norepineph
rine aB a mediator for dextroamphetamine-lnduced hypothermia.

CHAPTER 5
GENERAL DISCUSSION
Although the hyperthermic effect of systemically administered
amphetamine has been uniformly reported by numerous investigators, the
mechanism by which hyperthermia is produced remains controversial. Some
Investigators claim the effect to be central in origin (Brodie et al.,
1959; Euler, 1961; Belenky and Vitolina, 1962; Valzelli et al., 1966,
1968), whereas others claim the effect to be peripheral in origin
(Gessa, Clay and Brodie, 1969; Slater and Turnbull, 1969). Since the
drug is distributed throughout the body (Young and Gordon, 1962; Fuller
and Hines, 1967; Rosso et al., 1968; Maickel et al., 1969), its effect
on body temperature may be due to simultaneous actions on central and
peripheral tissues. In the present investigation the effects of intra
ventricular^ and intraperitoneally injected dextroamphetamine on body
temperature were studied. It was hoped that this investigation would
reveal more definitive information concerning the actions of dextroam
phetamine in producing body temperature changes. It was anticipated
that small intraventricular injections of dextroamphetamine would evoke
a relatively pure central action.
Intraventricular administration of graded doses of dextroam
phetamine to rats produced alterations in body temperature which do not
appear to describe a conventional dose-response relationship. For ex
ample, at an ambient temperature of 25°C, a small dose (4 meg) produced
69

a transient elevation of body temperature, whereas an intermediate dose
{100 meg) produced hypothermia and a large dose (400 meg) produced Ini
tial hypothermia followed by secondary hyperthermia. These unusual
dose-response observations may be tentatively explained on the basis of
central and peripheral actions of dextroamphetamine. The central ac
tions of dextroamphetamine may involve 1 or both of 2 proposed hypo
thalamic temperature regulating centers, i.e. the heat gain center and
the heat loss center (Meyer, 1913; Teague and Ranson, 1936; Jacobson and
Squires, 1961; Keller, 1963; Bligh, 1966a, 1966b). It is postulated
that the heat gain center is more sensitive than the heat loss center
and that the small dose of lntraventrlcularly administered dextroam
phetamine may activate the former without activating the latter. It is
further postulated that the intermediate dose may activate both hypo
thalamic centers, but the effects of the heat loss center mask the
effects of the heat gain center.
Support for the concept that the intermediate dose of dextroam
phetamine activates the heat loss center is provided by preliminary
observations in this laboratory. It was noted that the surface tempera
ture of the rat tail is increased at a time when body temperature is
reduced by lntraventrlcularly administered dextroamphetamine. Activa
tion of the heat loss center is reported to cause peripheral vasodila
tion, increased surface temperature and augmented loss of body heat
(Andersson, Grant and Larson, 1956; Andersson and Persson, 1957;
Andersson, Persson and Strom, 1960). The initial hypothermia following
intraventricular administration of the large dose of dextroamphetamine
may be explained on the same basis as the hypothermia produced by the

71
intermediate dose of dextroamphetamine. The secondary hyperthermia is
probably caused by escape of dextroamphetamine Into the systemic circu
lation to activate peripheral hyperthermic mechanisms.
It was observed in the current investigation that intraperito
neal injection of 100 meg of dextroamphetamine produced no change in
body temperature, whereas intraperitoneal injection of 20Q and 400 meg
of dextroamphetamine produced hyperthermia. These observations suggest
that the temperature effects of small and intermediate do$es of intra
ventricular^ injected dextroamphetamine are mainly, if n<>t solely,
central in origin. However, because the central hypothermic effect
masks the central hyperthermic effect, the central action of dextroam
phetamine may be considered predominantly hypothermic in nature. It was
also shown in the current investigation that intraperitoneal administra
tion of 2 mg/kg of dextroamphetamine produced a brain concentration of
dextroamphetamine equivalent to that produced by intraventricular admin
istration of the intermediate dose (100 meg) of dextroamphetamine. Pe
ripheral administration of this equivalent dose of dextroamphetamine
caused hyperthermia despite a brain concentration of dextvoamphetamine
which can produce hypothermia. On the basis of these observations It
may be postulated that systemlcally administered dextroamphetamine si
multaneously activated peripheral hyperthermic and central hypothermic
mechanisms. At an ambient temperature of 25°C the peripheral hyperther
mic effect exceeded the central hypothermic effect and hyperthermia was
observed. Plausibility of such a postulatlon is strengthened by reports
that hypothermia induced by intraventricular injection of norepinephrine
Is antagonized by peripheral administration of dextroamphetamine

72
(Brittaln, 1966) or norepinephrine (Cowell and Davey, 1968). If the
concept that systemic dextroamphetamine simultaneously activates periph
eral hyperthermic and central hypothermic mechanisms is correct, it
should be possible to unmask the hypothermic action of peripherally in
jected dextroamphetamine by overcoming the hyperthermia through en
hancing the dissipation of body heat.
Alterations in ambient temperature influence the rate of body
heat loss (Euler, 1961; Borison and Clark, 1967; Hemingway and Price,
1968). It can be expected that higher ambient temperatures would reduce
the rate of body heat dissipation and lower ambient temperatures would
enhance the rate of body heat dissipation. Intraventricular injection
of the intermediate dose of dextroamphetamine (100 meg) consistently
caused hypothermia which varied with the ambient temperatures employed
in the test. In contrast, intraperitoneal injection of the matching
dose of dextroamphetamine (2 mg/kg) caused hyperthermia at 25° and 20°C
but caused hypothermia at an ambient temperature of 6°C. In comparison,
the body temperature of control animals was constant at these 3 ambient
temperatures. It may be logically rationalized that at the higher ambi
ent temperatures (25° and 20°C) heat dissipation was low, the hyperther
mic action of dextroamphetamine predominated and hyperthermia was
observed. At the lowest ambient temperature (6°C) heat dissipation was
greatly enhanced, hypothermic action of dextroamphetamine predominated
and hypothermia was observed. These studies provide further support to
the concept that peripherally administered dextroamphetamine simultane
ously activates central hypothermic and peripheral hyperthermic mecha
nisms.

73
An additional study which served to dissociate the simultaneous
hyperthermic and hypothermic actions of systemically administered dex
troamphetamine involved the use of propranolol and phenoxybenzamine at
ambient temperatures of 20° and 25°C. Propranolol and phenoxybenzamine
are adrenergic blocking drugs which have the ability to antagonize
amphetamine-induced hyperthermia (Wolf and George, 1964; Mantegazza
et al., 1968a). It was found in the current study that the adrenergic
blocking drugs exerted no overt effect on the body temperature of con
trol animals at either ambient temperature. Also, at an ambient tem
perature of 25°C they were more effective in antagonizing hyperthermia
caused by peripheral injection of dextroamphetamine than hypothermia
caused by intraventricular injection of dextroamphetamine. But at 20°C,
an ambient temperature at which intraperitoneal injection of dextroam
phetamine typically causes hyperthermia, pretreatment with the adrenergic
blocking drugs caused dextroamphetamine to produce hypothermia. The
"reversal" of body temperature is apparently due to inhibition of the
peripheral hyperthermic effect of dextroamphetamine, which resulted in
unmasking of the concurrent central hypothermic action of dextroampheta
mine. These observations further strengthen the concept that system
ically administered dextroamphetamine simultaneously activates peripheral
hyperthermic and central hypothermic mechanisms.
Since dextroamphetamine is distributed throughout the body,
Including the central nervous system, It was important to substantiate
the concept that the hyperthermic effect of dextroamphetamine is due to
its action on peripheral tissues. A study, in which the central nervous
system of rats was destroyed, showed that intraperitoneal injection of

74
dextroamphetamine effectively retarded the rate of fall in body tempera
ture following ablation of the brain and spinal cord. This study pro
vided evidence that dextroamphetamine produces hyperthermia, at least
in part, by acting on peripheral tissues. Another study, in which the
hyperthermic action of dextroamphetamine was examined, involved consid
eration of the role of the adrenal glands in mediating the hyperthermic
action of dextroamphetamine. Peripheral administration of epinephrine
is reported to cause hyperthermia in rats (Masek and Raskova, 1964;
Feldberg and Lotti, 1967). This action must be due mainly, if not ex
clusively, to peripheral mechanisms because catecholamines do not readily
cross the blood brain barrier (Axelrod et al., 1959; Weil-Malherbe
et al., 1961; Samorajski and Marks, 1962). Since dextroamphetamine
releases epinephrine from the adrenal glands (Harvey et al., 1968; Rubin
and Jaanus, 1966) and since the present study showed that adrenalectomy
prevented the hyperthermic effect of intraperltoneally administered dex
troamphetamine, it could be concluded that the hyperthermic response in
duced by dextroamphetamine is, to a large extent, mediated peripherally
by the release of epinephrine from the adrenal glands. Also, it has
been reported that peripheral neuronal norepinephrine is responsible for
mediating the hyperthermic action of dextroamphetamine (Gessa, Clay and
Brodle, 1969). Although it is not known which catecholamine is rela
tively more important in mediating the hyperthermic action of dextroam
phetamine, there is convincing evidence that the hyperthermic action of
dextroamphetamine is indirect and is primarily peripheral In origin.
Various physiological and behavioral responses are known to
occur in association with induced temperature changes (Du Bols, 1939;

75
Euler, 1961)* Because some of these responses may be Involved In tem
perature regulation, a study was undertaken to determine if there Is a
correlation between certain of these physiological or behavioral re
sponses and dextroamphetamlne-induced body temperature changes. It
was found that peripheral administration of dextroamphetamine at an
ambient temperature of 25°C induced hyperthermia and caused an Increase
In spontaneous motor activity, oxygen consumption and sniffing behavior.
Intraventricular administration of dextroamphetamine produced hypother
mia and caused a reduction of spontaneous motor activity and oxygen con
sumption, but a moderate increase in sniffing behavior. The initial
impression was that changes In spontaneous motor activity and oxygen
consumption appeared to correlate well with changes in body temperature,
whereas sniffing behavior did not correlate with changes in body tem
perature. Subsequently, it was observed that intraperitoneal injection
of dextroamphetamine in normal rats at an ambient temperature of 6°C and
in adrenalectomlzed rats at an ambient temperature of 25°C produced a
reduction in body temperature and no change in body temperature, respec
tively. Yet, on the basis of overt observations both procedures appeared
to increase spontaneous motor activity. These later observations seem to
contradict the initial findings and seem to imply a lack of correlation
between spontaneous motor activity and dextroamphetamine-induced body
tenqierature changes. However, It should be recalled that body tempera
ture Is the result of heat generating and heat dissipating processes and
that systemically administered dextroamphetamine simultaneously activates
both processes. Therefore, enhancement of a process may result in over
riding of the opposing process, whereas inhibition of a process may

76
result in domination by the opposing process. From these considera
tions » it appears logical to suggest that various physiological re
sponses, such as increase or decrease in spontaneous motor activity and
Increase or decrease in oxygen consumption, may be more appropriately
correlated with heat generation or heat dissipation rather than with
changes in body temperature per se.
Because the administration of dextroamphetamine is reported to
modify the brain concentration of norepinephrine (Moore, 1964; Smith,
1965J Fuxe and Ungerstedt, 1968; Gropetti and Costa, 1969), dopamine
(Smith, 1965; Littleton, 1967; Lewander, 1968a, 1968b; Welch and Welch,
1969) and 5-hydroxytryptamine (McLean and McCartney, 1961; Smith, 1965;
Welch and Welch, 1969) and because body temperature is modified by
intraventricular administration of norepinephrine (Feldberg and Lotti,
1967; Myers and Yaksh, 1968), dopamine (Myers and Yaksh, 1968) and 5-hy
droxytryptamine (Feldberg and Lotti, 1967; Myers and Yaksh, 1968), it is
reasonable to speculate that 1 or more of these biogenic amines may be
lnqjortant in mediating the central body temperature effects of dextroam
phetamine. The relative importance of the 3 biogenic amines was exam
ined by means of a study in which chemical agents were employed to
deplete individual or combinations of the monoamines by inhibiting their
synthesis or by enhancing their release before testing with dextroam
phetamine. It was observed that release and depletion of all 3 monoa
mines with the benzoqulnollzlne Ro 4-1284 antagonized the hypothermic
action of intraventricularly injected dextroamphetamine. Depletion of
5-hydroxytryptamine with the tryptophan hydroxylase inhibitor p-chloro-
phenylalanine failed to antagonize the hypothermic effect of

77
lntraventricularly administered dextroamphetamine. On the other hand,
depletion of norepinephrine and dopamine with the combined use of alpha-
methyl -m-tyrosine , an agent which releases catecholamines, and alpha-
methyl -p -tyrosine, an agent which inhibits tyrosine hydroxylase,
decreased dextroamphetamine-induced hypothermia by 40 per cent. These
observations suggest that the central hypothermic action of dextroam
phetamine is mediated, at least in part, by 1 or both of the catechola
mines and that 5-hydroxytryptamine is not involved.
An additional study was performed to substantiate the possible
role of 1 or both of the catecholamines as central mediators of dex
troamphetamine -induced hypothermia. In this study histochemical fluo
rescence analyses were made of specific areas of the brain following
administration of dextroamphetamine. Thirty minutes after intraven
tricular or intraperitoneal administration, a time when peak hypothermic
activity may be expected, dextroamphetamine caused a reduction of nor
epinephrine In the anterior hypothalamic nucleus, but no change in the
concentration of dopamine in the nucleus caudatus putaraen or of 5-hy
droxytryptamine in the globus pallidus. Each of the amines exists as
the predominant amine in each of the respective areas examined (Fuxe,
1965; Frlede, 1966; Fuxe, Hokfelt and Ungerstedt, 1968). It was assumed
that changes In the fluorescence of a given area reflects mainly a
change of the predominant amine. Furthermore, It was assumed that if
these amines were mediators of dextroamphetamine in the production of
hypothermia, changes in amine level In any of these areas would reflect
similar changes in brain areas Involved with thermoregulation. Because
norepinephrine was the only 1 of the 3 amines whose concentration was

78
reduced, it was postulated that norepinephrine is Important In mediating
the central hypothermic action of dextroamphetamine. The postulation
Is particularly appealing because norepinephrine is the predominant
monoamine in the anterior hypothalamus and because this region is con
sidered to be important in thermoregulation (Meyer, 1913; Teague and
Ranson, 1936; Jacobson and Squires, 1961; Bllgh, 1966b). Further sup
port for this concept was provided by repeated intraventricular injec
tions of dextroamphetamine at close intervals. Repeated intraventricular
injections of dextroamphetamine resulted in a progressive development
of tachyphylaxis to dextroamphetamlne-lnduced hypothermia. These in
jections also caused a progressive reduction in the concentration of
norepinephrine in the anterior hypothalamus but no change in the con
centration of dopamine in the caudate nucleus. Hence, there appears to
be a correlation between changes In the degree of hypothermia and the
relative concentration of norepinephrine in the anterior hypothalamus.
This correlation further supports the postulation that norepinephrine is
important in mediating the central hypothermic action of dextroampheta
mine.
The findings of the present investigation may be summarized as
follows:
1. The central action of dextroamphetamine on body temperature is pre
dominantly hypothermic. There is a central hyperthermic component which
is believed to be readily masked by the hypothermic component.
2. The peripheral action of dextroamphetamine on body temperature is
hyperthermic.
3. The central hypothermic action of dextroamphetamine is believed to

79
be mediated to a large extent by norepinephrine, whereas the peripheral
hyperthermic action Is mediated at least partially by adrenal epineph
rine.
4. Various physiological or behavioral responses, such as motor ac
tivity and oxygen consumption, Induced by dextroamphetamine which may
participate in thermoregulation are probably more appropriately corre
lated with heat generation or heat dissipation rather than with changes
in body temperature per se.
5* Dextroamphetamine, administered systemically» is distributed through
out the body and simultaneously activates central hypothermic and periph
eral hyperthermic mechanisms. Consequently, the direction and extent
of body temperature change after systemic dextroamphetamine administra
tion depends on extraneous factors which may Increase or decrease the
rate of heat dissipation.

APPENDIX A
INTRACEREBROVENTRICULAR TECHNIQUE
Equipment
A. Implantation apparatus: The apparatus consists of 2 basic com-
ponents, the animal securing device and the cannula implantation
component. The securing device is composed of a wooden platform
and a metal incisor bar. The animal is secured to the platform
and the upper incisors are placed over the bar. The incisor bar
is attached to the base of a ring stand (E & M Instrument Co.)
and is perpendicular to the saggital plane of the animal. A
myograph tension adjustor (E & M Instrument Co.) Is positioned
on the ring stand so that adjustment moves a horizontally sus
pended bar in a vertical plane. A pair of curved forceps Is
attached to the horizontal bar with a Drinker clamp (A. H.
Thomas Co., Inc.). The forceps are used to hold the cannula.
Vertical adjustment of the cannula is accomplished by manipu
lation of the muscle tension adjustor. Lateral adjustment is
accomplished by rotation of the entire muscle tension adjustor
on the metal shaft of the ring stand.
B. Dental drill: model number 73, Foredom Electric Co., Inc.
C. Doriot hand piece: model 42, Foredom Electric Co., Inc.
D. Dental burr: number 700 uarbide, number 3 carbide, S. S. White
Co.
80

81
E. Cannula: The cannula is composed of 2 basic components, the
outer guide which directs the tip of the injecting needle into
the lateral ventricle and the inner stainless steel shaft which
is removed when the injecting needle is inserted. The shaft
maintains an open passage way for insertion of the injecting
needle and prevents leakage of cerebrospinal fluid. The outer
guide is made from a 23 gauge hypodermic needle. A small high
speed rotary motor fitted with a beveled grinding stone is used
to reduce the size of the hub and length of the shaft. When
completed the overall length of the cannula (shaft and hub) is
8 mm and the length of the shaft is 5 mm. The hub is ground so
that 2 flat surfaces are opposite each other. A groove approxi
mately 1 mm wide and 0.5 mm deep completely encircles the hub
and is positioned 0.5 mm above the juncture of the shaft and the
hub. The flat surfaces facilitate proper attachment to the can
nula holding component of the implantation apparatus. The
groove assures firm attachment to the orothodontic resin. The
inner stainless steel shaft is made from a 30 gauge hypodermic
needle. The dimensions of this needle are reduced with the same
equipment as described above. When completed the overall length
(shaft and hub) Is 10 nun and the length of the shaft is 8 mm.
The grinding procedure closed the lumen of the shaft. The re
duced hub is used to facilitate removal and replacement of the
inner shaft.
F. Injecting needle: The injecting needle is made from a blunt, 30
gauge 8 mm hypodermic needle. The size of the hub is reduced to

82
a length of 3 mm. The width at the juncture of the shaft and
the hub Is 2.5 mm. From this point the hub is beveled so that
It is generally cone shaped. A 15 inch polyethylene tube
(PE 20) is forced onto the cone and cemented in place with Epoxy
resin. The open end of the polyethylene tube is attached to a
10 mcl syringe (Hamilton Co., Inc.).
II. Materials
A. Iodochlorhydroxyquin powder: Vioform, Ciba Pharmaceutical
Products, Inc.
B. Methohexital: Brevital Sodium, Eli Lilly and Co.
C. Orthodontic resin: L. D. Caulk Co.
111. Implantation of cannula
A. Anesthetize a 180-220 g rat by intravenous administration of
methohexital.
B. Clip hair from dorsal surface of head.
C. Make mldsaggltal incision from the eyes to lambda.
D. Retract skin to provide ample working area.
E. Remove periosteum from exposed skull by scraping.
F. Drill a hole through the skull at a point 1.5 to 2.0 mm lateral
and 1.0 mm caudal to the bregma with a number 700 dental burr.
G. Approximately 9.5 mm from this hole drill 3 additional holes so
that they describe a triangle with the original hole (drilled in
step III.F) in the center. Keep all holes at least 1.5 mm
from the saggital suture since excessive bleeding occurs if the
saggital sinus is ruptured.

Enlarge all 4 holes with a number 3 dental burr. Avoid rupture
of the dura mater.
Place a stainless steel machine screw (0-80 X 0.32 cm, 1/8 inch)
in each hole drilled in step III.G. Screws should not penetrate
beyond ventral surface of skull bone.
Insert a sharp 26 gauge hypodermic needle through the center
hole and penetrate the dura mater. Avoid excessive damage to
brain tissue.
Mount a cannula on the holding component of the implantation
apparatus.
Position animal and cannula so that the cannula is perpendicular
to the skull at the point of the center hole.
Lower the tip of cannula 4 mm below the dorsal surface of the
skul1.
Quickly cement the cannula In place by application of a small
amount of orthodontic resin to the base of the exposed cannula
so that contact is made with the skull and at least 1 screw.
Walt 20 to 30 seconds and carefully detach holding component
from cannula.
Immediately elevate holding component and rotate away from ani
mal.
Apply additional resin so that contact is made between cannula
and all 3 stainless steel screws.
Allow 2 to 3 minutes for resin to harden.
Apply a small quantity of lodochlorhydroxyquin powder to ex
posed subdermal tissue*

84
T. House animals individually.
U. Allow at least 5 days for recovery.
IV. Intraventricular administration of drugs
A. Remove inner shaft from cannula.
B. Insert injecting needle into cannula opening.
C. Inject solution at the rate of 10 mcl per minute.
D. Slowly remove injecting needle.
E. Quickly replace inner shaft of cannula.
V. Confirmation of proper implantation
A. Inject 5 mcl of permanent black ink through cannula.
B. Decapitate animal and remove brain.
C. Dissect away the cortex near the path of the cannula.
D. Uniform staining of lateral ventricle wall suggests proper Im
plantation of cannula.
E. Perform saggital section.
P. Proper implantation is further assured if ink outlines third and
fourth ventricles.

APPENDIX B
PREPARATION OF DRUGS
D,L-Alpha-methyl-m-tyrosine (Nutritional Biochemicals Corp.)
Type of preparation: Solution
Vehicle: 0.9% NaCl solution
Solution strength: 500 mg/10 ml
D,L-Alpha-methyl-p-tyrosine (Regis Chemical Co.)
Type of preparation: Suspension
Vehicle: 0.9% NaCl solution
Dose: 80 mg/kg
Suspension strength: 80 mg/5 ml
Dextroamphetamine sulfate (Dexedrine Sulfate, Smith, Kline & French Labs.) for intraventricular use
Type of preparation: Solution
Vehicle: Deionized water (solution adjusted to osmolarity equal to 0.9% NaCl)
Dose: 4 meg (as salt)
Solution strength: 4 meg/5 mcl
Dose: 12 meg (as salt)
Solution strength: 12 mcg/5 mcl
85

Dose: 20 meg (as salt)
Solution strength: 20 meg/mel
Dose: 100 meg (as salt)
Solution strength: 100 mcg/5 mcl
Vehicle: Delonlzed water
Dose: 400 meg (as salt)
Solution strength: 400 mcg/5 mcl
Dextroamphetamine sulfate (Dexedrlne Sulfate, Smith, Kline & French Labs.) for intraperitoneal use
Type of preparation: Solution
Vehicle: 0.9% NaCl solution
Dose: 2 mg/kg
Solution strength: 2 mg/2 ml
D,L-p-chlorophenylalanlne (Chas. Pfizer & Company, Inc.)
Type of preparation: Suspension
Vehicle: 0.4% Tween 80, 1% carboxymethylcellulose, 0.9% NaCl
Dose: 300 mg/kg
Suspension strength: 300 mg/5 ml
Phenoxybenzamine hydrochloride (Dlbenzyllne Injection, Smith, Kline French Labs.)
Type of preparation: Solution
Vehicle: Delonlzed water (1 part Dlbenzyllne Injection diluted with 9 parts dionized water)

Dose: 10 mg/kg
Solution strength: 10 mg/2 ml
Propranolol hydrochloride (Inderal, Ayerst Laboratories)
Type of preparation: Solution
Vehicle: 0.9% NaCl solution
Dose: 10 mg/kg
Solution strength: 10 mg/2 ml
Ro 4-1284 (Hoffman-LaRoche, Inc.)
Type of preparation: Solution
Vehicle: 0.9% NaCl solution
Dose: 10 mg/kg
Solution strength: 10 mg/5 ml

APPENDIX C
ASSAY FOR BRAIN DEXTROAMPHETAMINE
1. Equipment
A. Centrifuge: Servall, type SS-3, Ivan Sorvall, Inc.
B. Glass Homogenizer: Duall, size C, Kontes Glass Co.
C. Liquid scintillation counting system: model 314, Packard
Instrument Co.
D. Mechanical shaker: Eberbach Corporation
E. Scintillation counting vial: Spectravial, model 3334 Nylon
vial. New England Nuclear
II. Materials
A. Benzene: analytical reagent grade, Mallinckrodt Chemical Works
B. Dextroamphetamine-H^ (G) sulfate: New England Nuclear
C. 2,5-Diphenyloxazole (PPO): scintillation grade, Packard
Instrument Co.
D. Hydrochloric acid: analytical reagent grade, Mallinckrodt
Chemical Works
E. Perchloric acid: analyzed reagent grade, J. T. Baker
Chemical Co.
F. 2,2-p-Phenylenebis-(5-phenyloxazole) (POPOP): scintillation
grade, Packard Instrument Co.
G. Sodium hydroxide: analyzed reagent grade, J. T. Baker
Chemical Co.
88

89
H. Toluene: analytical reagent grade, Mallinckrodt Chemical Works
III. Solutions
A. Isotope solutions
1. Dextroamphetamine stock solutions for standards
a. 10 meg standard: Ten meg of dextroamphetamine sulfate
is contained in each 0.1 ml of solution. The tritium
label is diluted 1:10,000 in this solution; the vehicle
is 0.01N HC1.
b. 3 meg standard: Three meg of dextroamphetamine sulfate
is contained in each 0.1 ml of solution. The tritium
label is diluted 1:10,000 in this solution; the vehicle
is 0.01N HC1.
c. 1 meg standard: One meg of dextroamphetamine sulfate is
contained in each 0.1 ml of solution. The tritium label
is diluted 1:10,000 in this solution; the vehicle is
0.01N HC1.
2. Dextroamphetamine stock solutions for injection
a. Solution for intraperitoneal injection: One mg/ml of
dextroanqphetamine sulfate is contained in the solution
for injection. The tritium label is diluted 1:10,000;
the vehicle is 0.9% NaCl solution.
b. Solution for intraventricular injection: Twenty mg/ml
of dextroamphetamine sulfate is contained in the solu
tion for injection. The tritium label is diluted
1:10,000; osmolarlty of the solution Is adjusted to

90
equal that of 0.9% NaCl solution.
B. Solutions for extraction procedure
1. Hydrochloric acid: 0.1N
2. Perchloric acid: 30% w/v
3. Sodium hydroxide: ION
4. Washed benzene: Keep an excess of deionized water in
container of benzene.
C. Scintillator solution
1. Place 2.0 g FPO into glass-stoppered container.
2. Add 50 mg POFOP and 500 ml toluene
3. Protect from light and store at 5°C.
IV. Procedure
A. Decapitate animal and dissect out brain.
B. Expose lateral ventricles with saggital incisions through cortex
approximately 2 mm lateral to midline.
C. Flood ventricles and outer surface of brain with 25 ml of cold
0.9% NaCl.
D. Drain surface liquid from brain and freeze brain on dry ice.
Store at -30°C until step E below.
E. Weigh tissue to nearest 10 mg.
F. Homogenize tissue with A volumes of 0.1N HC1 in motor-driven
chilled glass homogenizer. Assume brain tissue has a density of
1 g/ml. For standards substitute 0.1 ml of appropriate stock
solution (step 1II.A.1) per g of brain tissue for the same
amount of 0.1N HC1.

91
G. Add 30% perchloric acid to make a final concentration of 6%
perchloric acid. Mix thoroughly.
H. Transfer contents of homogenlzer to a 60 ml glass-stoppered
bottle.
I. Centrifuge at 1000 X gravity for 8 minutes.
J. Transfer 1 ml of supernatant to a second 60 ml glass-stoppered
bottle containing 0.2 ml 10N NaOH and 3 ml washed benzene.
K. Shake for 10 minutes on mechanical shaker at 220 oscillations
per minute.
L. Centrifuge at 1000 X gravity for 4 minutes.
M. Transfer 2 ml of benzene (upper) phase to scintillation counting
vial containing 10 ml of scintillator solution.
N. Quantitate the tritium decay with a liquid scintillation
counting system.
0. Calculate dextroamphetamine concentration In brain tissue by the
method described in V below.
Calculation of dextroamphetamine concentration in brain
A. Determine theoretical decays for entire brain
Theoretical decays (X) = decays per minute recorded for sample (A) (B) (C)
where
A» ^ .1 (2/3) (0.4) (0.95) [(6.25) (brain weight in g)J
The term in brackets represents the fraction of the whole
sample transferred to benzene. The 2/3 represents the

92
fraction transferred from benzene to scintillation solution.
The 0.4 and 0.95 represent approximate counting efficiency
and extraction efficiency, respectively.
B = the dilution factor for the tritium label (1/10,000)
C = correction factor for decomposed dextroamphetamine sulfate
This value reflects a decomposition of approximately 1.5%
per month. If 1 month had elapsed since dextroamphetamine-H3
sulfate was assayed by the manufacturer, then the value for
C would be 0.985.
B. Determine amount of dextroamphetamine sulfate in brain tissue
Amount of dextroamphetamine sulfate = 5 mce 5.36 x 107 6
The 5.36 x 10^ represents the theoretical decays of 1 meg of
dextroamphetamine-H sulfate.
C. Determine concentration of dextroamphetamine sulfate in brain
tissue
dextroamphetamine sulfate = amount of dextroamphetamine sulfate brain weight in g

APPENDIX D
STATISTICAL PROCEDURES
Student's t^ Test
Calculations were performed according to the following formula
|Xy _ iz|
= ~ 2 i_±_ + Jjfn*
where
p (-t -a
t = the calculated Student's t_ value
x = the mean value of a treatment
2 Sp— the pooled variance
n ° the number of replicates per sample
and
y and z = the particular treatments in question.
93

94
The standard error of the mean (S.E.M.) was calculated according
to the following formula:
S.E.M. - bX ' X>2"1 | n(n - 1) I
where
X = the value of a replicate within a sample
x = the mean of a sample
and
n = the number of replicates per sample.
Numerical data In this work are reported as the mean + S.E.M.
In line drawings and bar graphs the S.E.M. is indicated by a vertical
bracketed line. The difference between 2 means is reported to be
significant only when the calculated Student's t value is larger than
that value of the Student's t_ distribution where the probability (P)
of a larger t_ value is less than 0.05.

APPENDIX E
SPECTROPHOTOFLUOROMETRIC ASSAY OF 5-HYDROXY-TRYPTAMINE, NOREPINEPHRINE AND DOPAMINE
I. Equipment
A. Glass homogenizers: Duall, size D, Kontes Glass Co.
B. Centrifuge: Servall type SS-3, Ivan Sorvall, Inc.
C. Cells: fused quartz, 5 ml., American Instrument Co.
D. Spectrophotofluorometer: Aminco-Bowman, American Instrument Co
(photomultiplier tube, IP21, slit size, 3/16"; slit arrangement
No. 5)
E. Dopamine resin columns: The dopamine resin columns are made
from 14 mm glass tubing with a constriction on one end which is
10 mm long and has a 5 mm Inside diameter. A sintered glass
column 5 mm in diameter and 2.5 mm In length is fused into the
distal end of the constriction. The overall length of the dopa
mine resin column is 120 mm with the top 110 mm serving as a
reservoir for solutions and the lower 10 mm constriction serv
ing as a resin compartment.
II. Materials
A. L-Ascorbic acid: reagent grade, Fisher Scientific Co.
B. 1-Butanol: reagent grade, Fisher Scientific Co.
C. Dopamine hydrochloride: Calblochem
95

96
D. Glacial acetic acid: analytical reagent grade, Malllnckrodt
Chemical Works
E. n-Heptane: pure grade, Phillips Petroleum Co.
F. 5-Hydroxytryptamine creatinine sulfate: Calbiochem
G. Hydrochloric acid: analytical reagent grade, Malllnckrodt
Chemical Works
H. Iodine: analytical reagent grade, Malllnckrodt Chemical Works
I. Norepinephrine bltartrate, monohydrate: Winthrop Laboratories
J. Parafilm "M": American Can and Co.
K. Potassium chloride: analytical reagent grade, Malllnckrodt
Chemical Works
L. Potassium iodide: analytical reagent grade, Malllnckrodt
Chemical Works
M. Potassium phosphate, monobasic: analytical reagent grade,
Malllnckrodt Chemical Works
N. Resin: Dowex 50W-X4 resin, 200-400 mesh, hydrogen ionic form,
J. T. Baker Chemical Co.
0. Sodium acetate, trihydrate: analytical reagent grade,
Malllnckrodt Chemical Works
P. Sodium hydroxide: analytical reagent grade, Malllnckrodt
Chemical Works
Q. Sodium phosphate, dibasic: analytical reagent grade,
Malllnckrodt Chemical Works
R. Sodium phosphate, dibasic, heptahydrate: analytical reagent
grade, Malllnckrodt Chemical Works

97
S. Sodium phosphate, monobasic, monohydrate: analytical reagent
grade, Mallinckrodt Chemical Works
T. Sodium sulfite, heptahydrate: analyzed reagent grade,
J. T. Baker Chemical Co.
U. Sodium thiosulfate, pentahydrate: analytical reagent grade,
Mallinckrodt Chemical Works
III. Preparation of cation exchange resin, sodium ionic form (Moore and
Stein, 1951)
A. Place 1 pound of moist Dowex 50W-X4 resin into a Buchner funnel.
B. Wash with 4N HC1 until filtrate is almost colorless*
C. Wash resin twice with distilled water.
D. Wash resin with 2N NaOH until filtrate is alkaline.
E. Place the resin in about 3 times its volume of IN NaOH and sus
pend over a steam bath.
F. Heat approximately 3 hours and shake periodically.
G. Allow resin to settle.
H. Decant and discard liquid.
I. Add 3 volumes of hot IN NaOH and heat with shaking as in steps
III.E-H above.
J. Repeat step III.l 3 times.
K. Wash resin with distilled water until filtrate is neutral.
L. Pass resin through a 120 mesh screen with 6000 to 8000 ml of
distilled water.
M. Collect the screened resin and store as the moist sodium salt in
an air-tight container.

98
IV. Preparation of solutions
A. Stock solutions for monamlne standards: Solutions may be
stored several weeks If refrigerated and protected from light.
1. 5-Hydroxytryptamine: Weigh 23.007 mg of 5-hydroxytryptamlne
creatinine sulfate and dilute to 100 ml vlth 0.01N HCl.
This solution contains 100 meg of 5-hydroxytryptamlne base
per ml.
2. Norepinephrine: Weigh 19.935 mg of norepinephrine bitar-
trate, monohydrate and dilute to 100 ml with 0.01N HCl.
This solution contains 100 meg of norepinephrine base per
ml.
3. Dopamine: Weigh 12.38 mg of dopamine hydrochloride and
dilute to 100 ml with 0.01N HCl. This solution contains
100 meg of dopamine base per ml.
B. Salt-saturated butanol solution
1. Place 2500 ml of 1-butanol into a 4000 ml separatory funnel.
2. Add 1000 ml of 0.01N HCl.
3. Shake vigorously for 2 minutes; allow phase separation.
4. Discard lower phase.
5. Place approximately 120 g of NaCl into a 500 ml glass-
stoppered flask.
6. Add approximately 300 ml of the butanol phase from step 3
above.
7. Agitate on mechanical shaker for 30 minutes.
8. Allow phase separation.

99
9. Decant and store top layer (salt-saturated butanol) in air
tight glass-stoppered flasks.
C. Solutions for assay of norepinephrine and dopamine
1. Acetate buffer
a. Place 1 volume of 2N acetic acid in glass-stoppered
flask.
b. Add 2 volumes of 2N sodium acetate.
c. Mix thoroughly and store at 5°C.
2. Norepinephrine iodine solution
a. Dissolve 1.27 g of iodine in 100 ml of absolute ethanol.
b. Store at 5°C in light-resistant glass-stoppered flask.
3. Sodium thiosulfate solution
a. Dissolve 1.24 g of sodium thiosulfate, pentahydrate in
100 ml of deionized water. Properly prepared solutions
are assured when 1.875 + 0.025 ml of the sodium thio
sulfate solution decolorizes 1.00 ml of the norepineph
rine iodine solution.
4. Alkaline ascorbate solution
a. Dissolve 50 mg of ascorbic acid in 5 ml of deionized
water.
b. Add 10 ml of 5N NaOH; mix carefully.
5. Dopamine buffer number 1
a. Place 2.5940 g of monobasic sodium phosphate, mono-
hydrate and 1.8700 g of dibasic sodium phosphate, hepta-
hydrate into a 1000 ml volumetric flask.
b. Dilute to 1000 ml with deionized water.

100
6. Dopamine buffer number 2
a. Place 1.8440 g of monobasic sodium phosphate mono-
hydrate, 4.2500 g of dibasic sodium phosphate
heptahydrate, and 74.560 g of potassium chloride Into
a 1000 ml volumetric flask.
b. Dilute to 1000 ml with deionlzed water.
7. Dopamine buffer number 3
a. Place 4.259 g of dibasic sodium phosphate and 9.520 g
of monobasic potassium phosphate into a 1000 ml volume
tric flask.
b. Dilute to 1000 ml with deionlzed water.
8. Dopamine iodine solution
a. Dissolve 5.00 g of K1 in 5.0 ml of deionlzed water.
b. Add 0.2540 g of iodine.
c. Agitate until solution is achieved.
d. Dilute to 100 ml with deionlzed water.
e. Store at 5°C in a light-resistant bottle.
9. Alkaline sodium sulfite solution
a. Dissolve 0.5040 g of sodium sulfite heptahydrate in
1 ml of deionlzed water.
b. Dilute to 10 ml with 5N NaOH.
V. Procedure
A. Preparation of glassware
1. Quartz cells
a. Handle cells with plastic-tipped tongs.

101
b. Rinse cells several times with delonlzed water. *
c. Soak in concentrated nitric acid for at least 10
minutes.
d. Remove from acid, rinse with delonlzed water, and
place in hot-air oven to dry.
e. Remove from oven and allow to cool before use.
2. Pipettes
a. Rinse inside and outside of pipette with distilled water.
b. Immerse pipettes in concentrated sulfuric acid contain
ing an excess of sodium nitrate. Store for at least 2
hours in this solution.
c. Remove pipettes from acid and wash with tap water.
d. Place in automatic pipette washer and rinse with tap
water for at least 2 hours.
e. Flush each pipette with judicious quantities of delon
lzed water.
f. Dry pipettes in hot-air oven and store in dust-free
area.
3. Dopamine resin columns
a. Rinse column with delonlzed water to remove resin from
sintered glass.
b. Reverse-flush columns with delonlzed water several
times.
c. Dry in hot-air oven and store In dust-free area.
4. Other glassware
a. Thoroughly rinse with tap water.

102
b. Clean with Tig soap, warm water, and brush.
c. Rinse with tap water.
d. Immerse In dilute HCl for at least 2 hours,
e. Rinse with large quantities of delonlzed water.
f. Dry in hot-air oven and store in a dust-free area.
B. Preliminary preparations for extraction of 5-hydroxytryptamine,
norepinephrine and dopamine
1. Prepare room for assay.
a. Adjust room temperature to approximately 22°C.
b. Remove dust from room.
2. Prepare salt-saturated butanol bottles.
a. Place 25 ml of Bait-saturated butanol into 12 60-ml
ground-glass stoppered bottles.
b. Add 2 g of NaCl to each bottle.
c. Add 3 ml of 0.01N HCl to the reagent blank bottle.
d. Add 2.5 ml of 0.01N HCl to the low standard bottle.
e. Add 2 ml of 0.01N HCl to the high standard bottle.
f. Bottles containing only salt and butanol are called
sanple bottles and will each receive 3 ml of homogenized
brain tissue (see step V.C.5).
g. Bottles may be stored for several days at 5°C.
h. Throughout the assay procedure maintain bottles at 5°C
or less whenever they are not in use.
3. Prepare heptane bottles.
a. Place 20 ml of n-heptane into 12 60-ml glass-stoppered
bottles.

103
b. Add 3 ml of 0.01N HCl to each bottle.
Extraction of 5-hydroxytryptamine, norepinephrine and dopamine
1. Decapitate animal.
2. Remove whole brain, rinse with distilled water, and blot
with filter paper.
3. Immediately weigh brain tissue to nearest 10 nig.
4. Homogenize brain in motor-driven chilled glass homogenlzers
with 2 volumes of cold 0.01N HCl. For this step the density
of the brain is assumed to be 1 g/ml.
5. Transfer 3 ml of the homogenate to the appropriate sample
bottle.
6. Secure lid, cover bottle with light-resistant black plastic,
and store on ice until step 9.
7. To prepare monoamine standard bottles transfer 1 ml of the
5-hydroxytryptamine and dopamine stock solutions and 0.5 ml
of the norepinephrine stock solution to a volumetric flask
and dilute to 100 ml with 0.01N HCl. This solution contains
1 meg each of 5-hydroxytryptamine and dopamine base and 0.5
meg of norepinephrine base per ml.
8. Transfer 0.5 ml of this diluted solution to the low-standard
bottle and 1 ml to the high-standard bottle. The low-
standard bottle now contains 0.5 meg each of 5-hydroxytryp-
tamlne and dopamine base and 0.25 meg of norepinephrine
base. The high standard bottle contains 1.0 meg each of
S-hydroxytryptamlne and dopamine base and 0.5 meg of
norepinephrine base.

104
9. Agitate bottles for 10 minutes on mechanical shaker.
10. Centrifuge for 5 minutes at a relative centrifugal force
of 600 X gravity.
11. Transfer 20 ml of the butanol (upper) phase to the corre
sponding heptane bottle.
12. Repeat steps 9 and 10.
13. Aspirate and discard most of the butanol-heptane (upper)
phase. The remaining 3 ml aqueous phase is the aqueous acid
extract.
D. Determination of 5-hydroxytryptamine concentration
1. Transfer 1 ml of the aqueous acid extract (see step V.C.13)
into a quartz cell.
2. Add 0.3 ml of concentrated HC1.
3. Agitate gently until homogeneity is achieved.
4. Determine fluorescence emission intensity with spectrophoto-
fluororaeter. Use an activation wavelength of 300 nm and a
detection wavelength of 545 nm.
5. Calculate 5-hydroxytryptamine concentration as described in
step VI.
E. Determination of norepinephrine concentration
1. Transfer 0.6 ml of aqueous acid extract from each bottle
(see step V.C.13) to an appropriate small test tube.
2. Collect the residual extract from each sample bottle used in
step E.l above.
3. Transfer 0.6 ml of the pooled residual extract to a small

105
test tube. This tube contains extract to be used as the
brain blank.
4. Add 0.2 ml of acetate buffer to each small test tube.
Agitate thoroughly.
5. Add 0.02 ml of norepinephrine iodine solution to each small
test tube except the 1 reserved for the brain blank. Agi
tate thoroughly.
6. After exactly 6 minutes add 0.04 ml of sodium thlosulfate
solution to each small test tube except the brain blank.
7. Immediately add 0.2 ml of alkaline ascorbate solution, pre
pared within 10 minutes prior to use, to each small tube
except brain blank.
8. Add 0.04 ml of sodium thlosulfate solution to brain blank.
Agitate thoroughly.
9. Add 0.02 ml of norepinephrine iodine solution to brain
blank. Agitate thoroughly.
10. Immediately add 0.2 ml of freshly prepared alkaline ascor
bate solution to brain blank. Agitate thoroughly.
11. Expose all tubes to strong fluorescent light for 45 minutes.
12. Transfer to quartz cells.
13. Determine fluorescence emission Intensity with spectrophoto-
fluoroneter. Use an activation wavelength of 400 nm and a
detection wavelength of 510 nm.
14. Calculate norepinephrine concentration as described in
step VI.
F. Determination of dopamine concentration

106
1. Place a lightly packed 2 mm column of Dowex 50W-X4, sodium
Ionic form Into each dopamine resin column.
2. Wash each column with 2 ml of deionlzed water (care should
be taken to avoid disturbing the resin when liquids are
added).
3. Place 1 ml of appropriate aqueous acid extract (see step
V.C.13) on the resin column.
4. Repeat step 2 above.
5. Discard solutions that pass through resin column.
6. Elute 3,4-dihydroxyphenylalanine by addition of dopamine
buffer number 1 to the column.
7. Discard eluate.
8. Position clean tubes for collection of dopamine containing
eluate in step 9 below.
9. Place 8 ml of dopamine buffer number 2 on the column and
collect eluate which contains dopamine.
10. Thoroughly agitate contents of collection tubes and transfer
3 ml of each solution into a separate silica test tube.
11. Add 0.5 ml of dopamine buffer number 3 to each silica test
tube.
12. Add 0.3 ml of deionlzed water to each silica test tube.
Agitate thoroughly.
13. Add 0.05 ml of dopamine iodine solution to each silica test
tube. Agitate thoroughly.
14. Exactly 5 minutes later add 0.5 ml of freshly prepared alka
line sodium sulfite solution. Agitate thoroughly.

107
15. Exactly 5 minutes after addition of sulfite solution add
0.6 ml of 5N acetic acid. Agitate thoroughly.
16. Seal each test tube with parafilm.
17. Expose to strong fluorescent light for 17 to 22 hours.
18. Transfer approximately 1.5 ml of solution from each silica
tube into a separate quartz cell.
19. Determine fluorescence emission intensity with spectrophoto-
fluorometer. Use an activation wavelength of 338 nm and a
detection wavelength of 376 nm.
20. Calculate dopamine concentration as described In step VI.
Calculation of monoamine concentration in brain
A >= The photomultlplier microphotometer reading for the high stand
ard minus the reagent blank reading
B = The photomultlplier microphotometer reading for the low standard
minus the reagent blank reading
C • The number of micrograms of monoamine in the low standard before
extraction
D « The photomultlplier microphotometer reading for each sample
minus the appropriate blank. Calculations for dopamine and
5-hydroxytryptemine employed the reagent blank rather than the
tissue blank since It has been shown In our laboratory that
Concentration of monoamine
where

108
there is no difference between the 2 values. Calculations for
norepinephrine employed the tissue (brain) blank.

APPENDIX F
HISTOCHEMICAL FLUORESCENCE TECHNIQUE FOR DIRECT OBSERVATION OF NOREPINEPHRINE,
DOPAMINE AND 5-HYDROXYTRYPTAMINE
Equipment
A. Isopentane cooling apparatus: This apparatus consists of 2
compartments. The outer compartment (Fluo-War Flask, 4000 ml,
Virtis Co., Inc.) contains liquid nitrogen. The Inner compart
ment (stainless steel beaker, 600 ml) suspends isopentane In the
liquid nitrogen. A small copper basket (14 mesh, 55 mm diameter,
35 mm height) is used to confine the tissue while freezing.
B. Temporary storage container for brain tissue: Fluo-War Flask,
8000 ml, Virtis Co., Inc.
C. Aluminum foil container: The container Is made from 3 thick
nesses of aluminum foil and is 63 mm long, 35 mm wide and 15 mm
deep.
D. Plastic tray: The bottom of the tray is made of polyethylene
plastic 6 ram thick, 136 mm long and 89 mm wide. The walls of
the tray which are 2.5 mm thick and 25 mm high describe an inner
compartment for brain tissue and a surrounding outer compartment
for granular phosphorus pentoxlde. The outer compartment is 83
mm wide and 121 mm long, whereas the inner compartment is 51 mm
wide and 81 mm long.
E. Glass lyophylization chamber: Freeze-drying flask with ground

110
glass flange, 1000 ml, Vlrtls Co., Inc. The top has a vented
19/38 ground glass outer joint. The chamber is held horizon
tally by a wooden rack.
F. Dry-air box: The box is made from stainless steel and is 600 mm
high, 500 mm long and 376 mm wide. It has a glass window for
viewing and 2 arm ports fitted with long surgical gloves for
manipulation of tissue.
G. Wire rack: The rack consists of an 18 mesh copper screen bottom
(130 mm diameter) on which 8 wire cylinders (15 mm height, 25 mm
diameter) are attached.
H. Paraformaldehyde humidifying chamber: Glass desiccator, 250 mm
inside diameter, Kimax, Owens-Illinois Glass Co. The proper
sulfuric acid solution is placed In the bottom of the desic
cator. Paraformaldehyde powder is held in a crystallization
dish (170 mm diameter, 100 mm height) which rests inside the
desiccator.
I. Formaldehyde reaction chamber: A glass desiccator (170 mm di
ameter) is fitted with a two-piece wooden clamp In order to
secure the lid during heating.
J. Constant temperature oven: Isotemp Oven, Fisher Scientific Co.
K. Vacuum pump: Duo Seal Vacuum Pump, Model 1405, Welch Scientific
Co. A 3-way glass stopcock connects the pump, lyophylizatlon
chamber and air intake. The air intake is fitted with a silica
gel column. A vacuum gauge (McLeod Gauge, Virtis Co., Inc.) is
placed between the pump and the stopcock. High vacuum rubber
tubing and metal hose clamps are used for the various

Ill
connections. A 19/38 ground glass joint connects the vacuum
system to the lid of the glass lyophylization chamber.
L. Embedding chamber: The embedding chamber consists of 2 halves
joined together with a 24/40 ground glass connector. The lower
half is a 125 ml flat-bottom boiling flask. The upper half,
which Is mounted with a vacuum outlet, has a chamber positioned
for short-term storage of brain tissue. A photograph of this
apparatus is shown by Falck and Owman (1965).
M. Metal cylinder: The metal cylinder is made from a polished
steel, nickel plated cork borer and is approximately 200 mm long
and 18 mm In diameter.
N. Tissue embedding medium: Paraplast, Van Waters and Rogers, Inc.
0. Microtome: American Optical Co.
P. Slide warmer: Chicago Surgical and Electrical Co.
Q. Glass slides: 1 mm, Corning Glass Works.
R. Cover glass: No. 0, Corning Glass Works.
S. Fluorescence microscope: Ortholux, model 250, E. Leltz, Inc.
T. Film: Black and white photographs were made with Kodak Tri-X
Fan 35 mm film.
11. Materials and solutions
A. High vacuum grease (silicone lubricant): Dow Corning Co.
B. Humidified paraformaldehyde: A 10 mm layer of paraformaldehyde
(Fisher Scientific Co.) Is placed in the crystallization dish
and subjected to a relative humidity of 58.3%, 70.4% or 97.5% in
the humidifying chamber for at least 5 days prior to use. These

112
humidities were obtained by use of sulfuric acid solutions
with densities equal to 1.3 g/ml, 1.25 g/ml or 1.05 g/ml,
respectively.
C. Isopentane: pure grade, Phillips Petroleum Company
D. Phosphorus pentoxide, granular: analyzed reagent grade, J. T.
Baker Chemical Co.
E. Phosphorus pentoxide, powder: Mallinckrodt Chemical Works
F. Silica gel: 6 to 16 mesh, Fisher Scientific Co.
III. Procedure
A. Lyophylization step
1. Decapitate animal.
2. Remove brain (steps 1 and 2 should require less than 2
minutes).
3. Remove brain area containing desired portion of hypothala
mus, nucleus caudatus putamen and globus pallidus. With a
scalpel make an anterior coronal section at the intersection
of the optic nerves and a posterior coronal section approxi
mately A mm from the first cut.
4. Place the brain tissue on a numbered copper plate so that
the anterior surface makes contact with the plate.
5. Stain the posterior side (now facing upward) with approxi
mately 1 mcl of eosln dye.
6. Drop tissue and plate into isopentane cooled with liquid
nitrogen. Both solid and liquid isopentane should be
present during the freezing procedure.

113
7. After 1 minute transfer tissue and plate to a 10 ml beaker
filled with liquid nitrogen.
8. For temporary storage place the 10 ml beaker in a large
container of liquid nitrogen. Tissues may be stored for
at least 24 hours In liquid nitrogen.
9. Transfer tissue and plate to the aluminum foil container
filled with liquid nitrogen. Care should be taken to keep
tissue covered with liquid nitrogen.
10. Fill outer compartment of the plastic tray with granular
phosphorus pentoxide,
11. Place the aluminum foil container into the Inner compartment
of the plastic tray.
12. Insert the plastic tray into the glass lyophylization
• chamber which has been stored In a freezing unit at less
than -45°C.
13. Place a thin film of high vacuum grease on the ground glass
lip of the chamber and its lid.
14. Grease and attach the vacuum pump connection to chamber lid.
15. Press lid into position on lyophylization chamber and at the
same time activate vacuum pump.
16. After the liquid nitrogen and residual lsopentane have
vaporized reduce the chamber pressure to less than 0.005
mm Hg.
17. Continue lyophylization for 3 days at a temperature of
approximately -30°C.
Formaldehyde treatment step

1. Remove glass chamber from freezing unit and slowly
adjust temperature of chamber to approximately 35°C.
2. Slowly release vacuum by allowing room air to enter the
chamber after passing through 6 X 0.5 inch silica gel
column.
3. Remove chamber lid and quickly transfer plastic tray to
dry-air box.
4. Carefully transfer the tissue to the appropriate basket
in the wire rack.
5. Place paraformaldehyde into formaldehyde reaction chamber
which has been heated to 80°C. Use the humidified paraform
aldehyde prepared in 58.3% relative humidity for development
of catecholamines. For development of 5-hydroxytryptamlne
use the humidified paraformaldehyde prepared in 70.4% rela
tive humidity.
6. Transfer wire rack from dry-air box to reaction chamber.
7. Secure reaction chamber lid and place in constant tempera
ture oven at 80°C for 1 hour.
8. Remove reaction chamber from oven and allow to cool in a
stream of room air for 30 minutes.
9. Slowly remove reaction chamber lid.
10. If 5-hydroxytryptamine is being developed repeat steps 5
through 9 above using a humidified paraformaldehyde prepared
at 91.57. relative humidity. If catecholamines are being de
veloped omit this step and proceed directly to step 11 below,
11. Transfer wire rack to vacuum desiccator containing powdered

115
phosphorus pentoxide. If overnight storage is desired re
duce desiccator pressure to less than 0.05 mm Hg and protect
from light. Before removing tissue from desiccator increase
internal pressure as specified in step 2 above.
Embedding step
1. Place an appropriate amount of tissue embedding medium into
lower half of embedding chamber. Melt in a water bath
heated to 60°C.
2. Transfer a tissue into the upper half of the embedding
chamber, connect upper portion to lower half of chamber and
reduce pressure to less than 0.05 mm Hg.
3. Tip the specimen into the liquefied tissue embedding medium
by tilting the chamber and tapping the upper half. Care
should be taken to avoid coating the inside wall of the
upper half of the chamber with liquefied embedding medium.
4. Infiltrate for 20 minutes.
5. Release vacuum and remove bottom half of chamber from water
bath.
6. Trap tissue (stained side up) inside the metal cylinder.
7. Cool lower half of chamber and cylinder with dry ice for
20 minutes.
8. Remove metal cylinder from chamber. The metal cylinder now
contains the tissue surrounded by solidified tissue embedd
ing medium.
9. Remove the tissue embedding medium and tissue from the
cylinder with aid of a glass rod. Tissues can now be stored

at room temperature for several weeks if protected from
light.
D. Fluorescence viewing step
1. Mount the embedded tissue on microtome so that sectioning
begins at the unstained surface.
2. Section tissue at a thickness of 10 microns.
3. When sectioning reaches a depth of 140 microns from the
anterior (unstained) surface, collect sections and place
on a dry slide. This portion corresponds to the section
which is 5780 microns anterior to the frontal zero plane
as described by Konig and Klippel (1963).
4. Transfer the slide to a slide warmer, maintained at 60° to
65°C, to allow the paraplast to melt and the sections f".o
spread. Remove from heat and allow to solidify.
5. Place cover glass over tissue and view with the aid of a
fluorescence microscope. With the third ventricle as a
guide, the anterior hypothalamic nucleus periventricularls
is viewed 0.8 to 0.9 mm dorsal to the ventral surface of the
brain. Intense green to yellow-green discrete fluorescence
appears in varicosities of nerve fibers in this region. The
nucleus caudatus putamen is viewed approximately 3 mm lat
eral to the third ventricle and 5.5 mm dorsal to the ventral
surface of the optic chiasm. This nucleus is characterized
by diffuse green to yellow-green fluorescence which con
trasts with the dark brain regions surrounding it. The
globus pallldus is found approximately 3 mm lateral to the

117
third ventricle and 3 ram dorsal to the ventral surface of
the optic chiasm. Discrete yellow fluorescence is seen in
this region.
6. Photograph the various areas soon after exposure to the
activating light since loss of some fluorescence occurs
after about 3 minutes.

LIST OF REFERENCES
Allen, D. J. and Marley, E.: Effect of sympathoniimetlc and allied amines on temperature and oxygen consumption in chickens. Brit. J. Pharmacol. 31: 290-312, 1967.
Andersson, B. , Ekman, L., Gale, C. C. and Sundsten, J. W. : Control of thyrotrophic hormone (TSH) secretion by the "heat loss center." Acta Physiol. Scand. 59: 12-33, 1963.
Andersson, B., Gale, C, C., Hokfelt, B. and Ohga, A.: Relation of preoptic temperature to the function of the sympathico-adrenomedu1lary system and the adrenal cortex. Acta Physiol. Scand. 61^: 182-191, 1964.
Andersson, B., Gale, C. C. and Ohga, A.: Supression by thyroxine of the thyroidal response to local cooling of the "heat loss center." Acta Physiol. Scand. 59: 67-73, 1963.
Andersson, B., Grant, R. and Larson, S.: Central control of heat loss mechanisms in the goat. Acta Physiol. Scand. 37^ 261-280, 1956.
Andersson, B. and Persson, N.: Pronounced hypothermia elicited by prolonged stimulation of the "heat loss centre" in unanesthetized goats. Acta Physiol. Scand. 41: 277-282, 1957.
Andersson, B., Persson, N. and Strom, L.: Post-stimulatory electrical activity in the preoptic "heat loss centre" concomitant with persistent thermoregulatory response. Acta Physiol. Scand. 50: 54-61, 1960.
Arnfred, T. and Randrup, A.: Cholinergic mechanism in brain inhibiting amphetamine-induced stereotyped behaviour. Acta Pharmacol. Toxicol. 26: 384-394, 1968.
Askew, B. M.: Hyperpyrexia as a contributory factor in the toxicity of amphetamine in aggregated mice. Brit. J. Pharmacol. JJh 2 45-257, 1962.
Axelrod, J., Weil-Malherbe, H. and Tomchick, R.: The physiological disposition of H-3-epinephrine and its metabolite metanephrine. J. Pharmacol. Exp. Therap. 127: 251-256, 1959.
Belenky, M. L. and Vitolina, M.: The pharmacological analysis of the hyperthermia caused by phenamine (amphetamine). Int. J. Neuro-pharmacol. 1^ 1-7, 1962.
118

119
Bernardi, D., Paglialunga, S. and Jori, A.: Peripheral and central components in the hyperthermic effect of deslpramine in reserpinized rats. J. Phartn. Pharmacol. 20: 204-209, 1968.
Bligh, J.: Effects on temperature of monoamines injected into the lateral ventricle of sheep. J. Physiol. (London) 185: 46-47, 1966a.
Bligh, J.: The thermosensitivity of the hypothalamus and thermoregulation in mammals. Biol. Rev. 41: 317-367, 1966b.
Bogdanski, D. F., Pletscher, A., Brodie, B. B. and Udenfriend, S.: Identification and assay of serotonin in brain. J. Pharmacol. Exp. Therap. U7: 82-88, 1956.
Booth, D. A.: Amphetamine anorexia by direct action on the adrenergic feeding system of rat hypothalamus. Nature 217: 869-870, 1968.
Borison, H. L. and Clark, W. G.: Drug actions on thermoregulatory mechanisms. Advance. Pharmacol. 5: 129-212, 1967.
Brittain, R. T.: The intracerebral effects of noradrenaline and its modification by drugs in the mouse. J. Pharm. Pharmacol. 113: 621-623, 1966.
Brittain, R. T. and Handley, S. L.: Temperature changes produced by the injection of catecholamines and 5-hydroxytryptamine into the cerebral ventricles of the conscious mouse. J. Physiol. (London) 192: SOS-SIS, 1967.
Brodie, B. B. , Spector, S. and Shore, P. A.: Interaction of drugs with norepinephrine in the brain. Pharmacol. Rev. 1A: 548-564, 1959.
Brown, A. M. and Julian, T.: The body temperature response of two inbred strains of mice to handling, saline and amphetamine. Int. J. Neuro-pharmacol. 1\ 531-541, 1968.
Burn, J. H.: Tyramine and other amines as noradrenaline-releasing substances. Iji Adrenergic Mechanisms, ed. by J. R. Vane, G. E. W. Wolstenholme and M. O'Connor, pp. 326-336, Little, Brown and Company, Boston, 1960.
Burn, J. H. and Rand, M. J.: Action of sympathomimetic amines in animals treated with reserplne. J. Physiol. (London) 144: 314-336, 1958.
Carlsson, A., Dahlstrom, A., Fuxe, K. and Hillarp, N.-A.: Failure of reserplne to deplete noradrenaline neurons of <*-roethylnoradrenaline formed from e(-methyl DOPA. Acta Pharmacol. Toxicol. 22_: 270-276, 1965.
Carlsson, A., Falck, B. and Hillarp, N.-A.: Cellular localization of brain monoamines. Acta Physiol. Scand. 5<S: suppl. 196, pp. 1-28, 1962.

120
Carlsson, A. and Lindqvist, M.: The interference of tetrabenazine, benzquinamide and prenylamine with the action of reserpine. Acta Pharmacol. Toxicol. 24: 112-120, 1966.
Carlsson, A., Lindqvist, M., Dahlstrom, A., Fuxe, K. and Masuoka, D.: Effects of the amphetamine group on intraneuronal brain amines in vivo and in vitro. J, Phar. Pharmacol. 17_: 521-524, 1965.
Carlsson, A, and Waldeck, B.: A fluorometric method for the determination of dopamine (3-hydroxytyramine). Acta Physiol. Scand. 44: 293-298, 1958.
Carr, L. A. and Moore, K. E.: Norepinephrine: release from brain by d-amphetamine iri vivo. Science 164: 322-323, 1969a.
Carr, L. A. and Moore, K. E.: Amphetamine-Induced release of catecholamines from cat brain as demonstrated by cerebroventricular perfusion. Fed. Proc. 28: 796, 1969b.
Cession-Fossion, A.: Hypertensive properties of amphetamine and tyramine in the rat. Arch. Int. Physiol. 71_: 382-392, 1963.
Cheymol, J. and Levassort, C«: Hyperthermic amphetaminique et curarisa-tion par d-tubocurarine. Annu. Pharmacol. Franc. 15^: 103-106, 1957.
Clark, W. C., Blackman, H. J. and Preston, J. E.: Certain factors in aggregated mice d-amphetamine toxicity. Arch. Int. Pharmacodyn. 170: 350-363, 1967.
Cooper, K. E., Cranston, W, I. and Honour, A. J.: Effects of intraventricular and intrahypothalamic Injection of noradrenaline and 5-hydroxytryptamine on body temperature in conscious rabbits. J. Physiol. (London) 181: 852-864, 1965.
Corrodi, H., Fuxe, K. and Hokfelt, T.: A possible role played by central monoamine neurons in thermo-regulation. Acta Physiol. Scand. 71: 224-232, 1967.
Corrodi, H. and Malmfors, T.: The effect of nerve activity on the depletion of the adrenergic transmitter by inhibitors of noradrenaline snythesis. Acta Physiol. Scand. 67: 352-357, 1966.
Cowell, P.: The interactions of noradnamine and imipramine-like antidepressant drugs. Brit. J. Pharmacol. 36: 85-96, 1969.
Cowell, P. and Davey, M. J.: The reversal of the central effects of noradrenaline by antidepressant drugs in mice. Brit. J. Pharmacol. 34; 159-168, 1968.

121
Dahi strom, A., Fuxe, K. and Hi Harp, N.-A.: Site of action of reserplne. Acta Pharmacol. Toxicol. 22_: 277-292, 1965.
D'Amour, F. E., Blood, F. R. and Belden, D. A., Jr.: Manual for Laboratory Work in Mammalian Physiology, third edition, pp. 89-92, The University of Chicago Press, Chicago, 1965.
Davis, R. A. and Horlington, M.: Effects of reserpine pre-treatment on the protective action of amphetamine and phenoxypropazine in the phenylbenzoquinone-induced writhing syndrome in mice. Nature 201: 306-307, 1964.
Du Bois, E. F.: Heat loss from the human body. Bull. N. Y. Acad. Med. 15: 143-173, 1939.
Edinger, H. M., Eisenman, J. S. and Brobeck, J. R.: Thermosensitive units in the posterior hypothalamus. Fed. Proc, 28: 713, 1969.
Engel, J. Hanson, L. C. F., Roos, B.-E. and Strombergsson, L.-E.: Effect of electroshock on dopamine metabolism in rat brain. Psychopharma-cologia 13: 140-144, 1968.
Ernest, A. M.: Mode of action of apomorphine and dexamphetamine on gnawing compulsion in rats. Psychopharmacologia 10: 316-323, 1967.
Ernest, A. M. and Smelik, P. G.: Site of action of dopamine and apomorphine on compulsive gnawing behavior in rats. Experientia 22: 837-839, 1966.
Essex, H. F.: Further observations of certain responses of tolerant and control animals to massive doses of epinephrine. Amer. J. Physiol. 171: 78-86, 1952.
Euler, C. von: Physiology and pharmacology of temperature regulation. Pharmacol. Rev. 1J: 361-398, 1961.
Euler, U. S. von, Linder, E. and Myrin, S. D.: Uber die fiebererregende wirkung des adrenalius. Acta Physiol. Scand. 5^: 85-86, 1943.
Falck, B.: Cellular localization of monoamines. Progr. Brain Res. 8: 28-44, 1964.
Falck, B. and Owman, C.: A detailed methodological description of the fluorescence method for the cellular demonstration of biogenic amines. Acta Univ. Lund. 2: 5-17, 1965.
Fasslna, G.: Azlone di farmaci anoressanti sugll acldi grassi liberi plasmatic!. Arch. Int. Pharmacodyn. 161: 410-422, 1966.

122
Feldberg, W., Hellon, R. F. arid Lottl, V. J.: Temperature effects produced in dogs and monkeys by injections of monoamines and related substances into the third ventricle. J. Physiol. (London) 191: 501-515, 1967.
Feldberg, W., Hellon, R. F. and Myers, R. D.: Effects on temperature of monoamines injected into the cerebral ventricles of anesthetized dogs. J, Physiol. (London) 186: 413-423, 1966.
Feldberg, W. and Lotti, V. J.: Temperature responses of monoamines and an inhibitor of MAO injected into the cerebral ventricles of rats. Brit. J. Pharmacol. 31: 152-161, 1967.
Feldberg, W. and Myers, R. D.: A new concept of temperature regulation by amines in the hypothalamus. Nature 200: 1325, 1963.
Feldberg, W. and Myers, R. D.: Effects on temperature of amines injected into the cerebral ventricles. A new concept of temperature regulation. J. Physiol. (London) 173: 226-237, 1964a.
Feldberg, W. and Myers, R. D.: Temperature changes produced by amines injected into the cerebral ventricles during anesthesia. J. Physiol. (London) 175: 464-468, 1964b.
Findlay, J. D. and Thompson, G. E.: The effect of Intraventricular injections of noradrenaline, 5-hydroxytryptamine, acetylcholine and tranylcypromine on the ox (Bos Taurus) at different environmental temperatures. J. Physiol. (London) 194: 809-816, 1968.
Fog, R. L.: Role of corpus striatum in typical behavioral effects in rats produced by both amphetamine and neuroleptic drugs. Acta Pharmacol. Toxicol. 25_: suppl. 4, p. 59, 1967.
Fog, R. L., Randrup, A. and Pakkenberg, H.: Aminergic mechanisms in corpus striatum and amphetamine-induced stereotyped behavior, Psychopharmacologia _11: 179-183, 1967.
Friede, R. L.: Topographic Brain Chemistry, pp. 282-316, Academic Press Inc., New York, 1966.
Fuller, R. W. and Hines, C. W.: d-Amphetamine levels in brain and other tissues of isolated and aggregated mice. Biochem. Pharmacol. 16: 11-16, 1967.
Fuxe, K.: Evidence for the existence of monoamine neurons in the central nervous system. III. The monoamine nerve terminal. Z. Zellforsch. 65: 573-596, 1965.
Fuxe, K., Hokfelt, T. and Ungerstedt, U.: Localization of indolealkyl-amines in CNS. Advance. Pharmacol. 6A: 235-251, 1968*

123
Fuxe, K. and Ungerstedt, U.: Hlstochemical studies on the effect of (+)-amphetamine, drugs of the imipramine group and tryptamine on central catecholamine and 5-hydroxytryptamine neurons after intraventricular injection of catecholamines and 5-hydroxytryptamine. Europe. J. Pharmacol. 4: 135-144, 1968.
Gaddum, J. H. and Vogt, M.: Some central actions of 5-hydroxytryptamine and various antagonists. Brit. J. Pharmacol. 1_1: 175-179, 1956.
Garattlni, S., Giacalone, E. and Valzelli, L.: Isolation, aggressiveness and brain 5-hydroxytryptamine turnover. J. Phami. Pharmacol. 19: 338-339, 1967.
Gessa, G. L., Cho, A. K., Clay, G, A,, Tagliamonte, A. and Brodie, B. B.: Possible role of p-hydroxynorephedrine in development of tolerance to d-amphetamine. Fed. Proc. 28: 796, 1969.
Gessa, G. L., Clay, G. A. and Brodie, B. B.: Evidence that hyperthermia produced by d-amphetamine is caused by a peripheral action of the drug. Life Sci. 8: 135-141, 1969.
Glowinski, J., Axelrod, J. and Iversen, L. L.: Regional studies of catecholamines in the rat brain. IV. Effects of drugs on the disposition and metabolism of H -norepinephrine and H^-dopamine. J. Pharmacol. Exp. Therap. 153: 30-41, 1966.
Glowinski, J., Snyder, S. H. and Axelrod, J.: Subcellular localization of H3 -norepinephrine in the rat brain and the effect of drugs. J. Pharmacol. Exp. Therap. 152: 282-2 92, 1965.
Goldstein, M. and Nakajima, K.: The effect of dlsulfiram on the biosynthesis of catecholamines during exposure of rats to cold. Life Sci. 5: 175-179, 1966.
Gorbman, A. and Bern, H. A,: A Textbook of Comparative Endocrinology, p. 306, John Wiley and Sons Inc., New York, 1962.
Gordon, R. Spector, S., Sjoerdsma, A. and Udenfriend, S.: Increased synthesis of norepinephrine and epinephrine during exercise and exposure to cold. J. Pharmacol. Exp. Therap. 153: 440-447, 1966.
Greenblatt, E. N. and Osterberg, A. C.: Correlations of activating and lethal effects of excitatory drugs in grouped and isolated mice. J. Pharmacol. Exp. Therap. 131: 115-119, 1961.
Gropetti, A. and Costa, E.: d-Amphetamine (A): metabolites and depletion of brain and heart norepinephrine in guinea pig and rat. Fed. Proc. 28: 795, 1969.

124
Grunden, L. R. and Linburn, G. E.: Permanent cannular-injection system for intracerebral injections in small animals. J. Pharm. Sci. 58: 1147-1148, 1969.
Gyermek, L.: The action of aktedron (phenylisopropylamine) on gas respiration (oxygen consumption) in adrenalectomized and thyroxine-treated animals. Arch. Exp. Pathol. Pharmakol. 209: 27-34, 1950.
Haas, H. T. A.: Askorbinsaure und Fieber. Arch. Exp. Pathol. Pharmakol. 192: 331-360, 1939.
Haffner, F.: Zur pharmakologle und praxis der stimulanten. Klin. Wochensch. 17: 1310-1311, 1938.
Hammel, H. T. and Hardy, J. D.: A gradient type of calorimeter for measurement of thermoregulatory responses in the dog. In Temperature: Its Measurement and Control in Science and Industry, vol. 3, part 3 (Biology and Medicine), ed. by J. D. Hardy, pp. 31-42, Reinhold Publishing Corporation, New York, 1963.
Hammel, H. T., Hardy, J. D. and Fusco, M. M.: Thermoregulatory responses to hypothalamic cooling in unanesthetized dogs. Amer. J. Physiol. • 198: 481-486, 1960.
Hammel, H. T., Jackson, D. C., Stolwijk, J. A., Hardy, J. D. and Stromme, S. B.: Temperature regulation by hypothalamic proportional control with an adjustable set-point. J. Appl. Physiol. 18^: 1146-1154, 1963.
Hanson, L. C. F.: Evidence that the central action of amphetamine is mediated via catecholamines. Psychopharmacologia 9: 78-80, 1966.
Hardlnge, M. G. and Peterson, D. I.: The effect of forced exercise on body temperature and amphetamine toxicity. J. Pharmacol. Exp. Therap. 145: 47-51, 1964.
Harrison, J. W. E., Ambrus, C. M. and Ambrus, J. L.: Tolerance of rats toward amphetamine and methamphetamine. J. Amer. Pharm. Ass. 41: 539-541, 1952.
Harvey, S. C., Sulkowskl, T. S. and Weenig, D. J.: Effect of amphetamines on plasma catecholamines. Arch. Int. Pharmacodyn. 172: 301-322, 1968.
Hemingway, A. and Price, W. M.: The autonomic nervous system and regulation of body temperature. Anesthesiology ̂ 9: 693-701, 1968.
Hemingway, A., Rasmussen, T., Wikoff, H. and Rasmussen, A. T.: Effects of heating hypothalamus of dogs by diathermy. J. Neurophyslol. 3: 329-338, 1940.

125
Hess, S. M., Connamacher, R. H. Ozaki, M, and Udenfriend, S.: The effects of <X-methyl -dopa and o(-methyl -meta-tyrosine on the metabolism of norepinephrine and serotonin iri vivo. J. Pharmacol. Exp. Therap. 134: 129-138, 1961.
Ingenito, A. J.: Norepinephrine levels in various areas of rat brain during cold acclimation. Proc. Soc. Exp. Biol. Med. 127: 74-77, 1968.
Ingenito, A. J. and Bonnycastle, D. D.: On the relationship between drug-induced changes in brain amines and body temperature. Can. J. Physiol. Pharmacol. 45^ 723-732, 1967a.
Ingenito, A. J. and Bonnycastle, D. D.: The effect of exposure to heat and cold upon rat brain catecholamine and 5-hydroxytryptamine levels. Can. J. Physiol. Pharmacol. 45; 733-743, 1967b.
Ingle, D. J.: The role of the adrenal cortex in homeostasis. Pediatrics (New York) 17: 407-413, 1956.
Jacobson, F. H. and Squires, R. D.: Chronic deficits of temperature regulation produced by lesions in the preoptic regions of cats. Fed. Proc. 20: 215, 1961.
Jacobson, F. H. and Squires, R. D.: Thermoregulatory responses to varied preoptic temperature during heat and cold exposure. Fed. Proc. 23: 566, 1964.
Jequler, E., Lovenberg, W. and Sjoerdsma, A.: Tryptophan hydroxylase inhibition: the mechanism by which p-chlorophenylalanlne depletes rat brain serotonin. Mol. Pharmacol. 3: 274-278, 1967.
Jobe, P. C.: Relationship of Brain Amine Metabolism to Audiogenic Seizure in the Rat. pp. 20-90, Ph.D. dissertation, The University of Arizona, 1970.
Jorl, A., Pagllalunga, S. and Garattini, S.: Effect of antidepressant and adrenergic blocking drugs on hyperthermia induced by reserpine. Arch. Int. Pharmacodyn. 165: 384-393, 1967.
Kahn, R. H.: Uber die erwarmung des carotidenblutes. Arch. Anat. Physiol. 28: 81-134, 1904.
Keller, A. D.: Temperature regulation disturbances in dogs following hypothalamic ablations. Iii Temperature: Its measurement and Control in Science and Industry, vol. 3, part 3 (Biology and Medicine), ed. by J. D. Hardy, pp. 571-584, Reinhold Publishing Corporation, New York, 1963.

126
Klissiunls, M. N. and Dosl, 1.: Effects of promethazine on phenyliso-propy1amine-induced fever in rabbits. Therapie 14: 745-746, 1959.
Koe, B. K. and Weissman, A.: p-Chlorophenylalanine: a specific depletor of brain serotonin. J. Pharmacol. Exp. Therap. 154: 499-316, 1966.
Konig, J. F. R. and Klippel, R. A.: The Rat Brain, pp. 1-161, The Williams and Wilkins Co., Baltimore, 1963.
Kruger, F. J., Kundt, H. W., Hensel, H. and Pruck, K.: Das verhalten der hautdurchblutung bei hypothalamuskuhlung an der wachen katze. Pflugers Arch. Ges. Physiol. 269: 240-247, 1959.
Laursen, A. M.: Movements evoked from the region of the caudate nucleus in cats. Acta Physiol. Scand. 54: 175-184, 1962.
Leimdorfer, A.: The action of sympathomimetic amines on the central nervous system and the blood sugar: relation of chemical structure to mechanism of action. J. Pharmacol. Exp. Therap. 98: 62-71, 1950.
Lessln, A. W. and Parkes, M. W.: The hypothermic and sedative action of reserpine in the mouse. J. Pharm. Pharmacol. 9: 657-662, 1957.
Levi, R. and Maynert, E. W.: The subcellular localization of brain-stem norepinephrine and 5-hydroxytryptamine in stressed rats. Biochem. Pharmacol. 13_: 615-621, 1964.
Levitt, M., Spector, S., Sjoerdsma, S. and Udenfriend, 5.: Elucidation of the rate-limiting step in norepinephrine biosynthesis in the perfused guinea pig heart. J. Pharmacol, Exp. Therap. 148: 1-8, 1965.
Lewander, T.: Effects of amphetamine on urinary and tissue catecholamines in rats after inhibition of its metabolism with desipramine. Europe. J. Pharmacol. 5: 1-9, 1968a.
Lewander, T.: Urinary excretion and tissue levels of catecholamines during chronic dl-amphetamine intoxication of the rat. Psycho-pharmacologia 13: 387-393, 1968b.
Littleton, J. M.: The interaction of dexamphetamine with inhibitors of noradrenaline biosynthesis in rat brain iri vivo. J. Pharm. Pharmacol. 19.: 414-415, 1967.
Lovenberg, W. Jequier, E. and Sjoerdsma, A.: Tryptophan hydroxylatlon In mammalian systems. Advance. Pharmacol. 6A: 21-36, 1968.
Magoun, H. W., Harrison, F., Brobeck, J. R. and Ranson, S. W.: Activation of heat loss mechanisms by local heating of the brain. J. Neurophysiol. JL: 101-114, 1938.

127
Malckel, R. P., Cox, R. H., Jr., Miller, F. P., Segal, D. S. and Russell, R. W.: Correlation of brain levels of drugs with behavioral effects. J. Pharmacol. Exp. Therap. 165: 216-224, 1969.
Mantegazza, P., Naimzada, K. M. and Riva, M.: Effects of propranolol on some activities of amphetamine. Europe. J. Pharmacol. 4: 25-30, 1968a.
Mantegazza, P., Naimzada, K. M. and Riva, M.: Activity of amphetamine in hypothyroid rats. Europe. J. Pharmacol. 5: 10-16, 1968b.
Masek, K. and Raskova) H.: Catecholamlne-induced endotoxin-like fever. In Biochemical and Neurophysiological Correlation of Centrally Acting Drugs, ed. by E. Trabucchi, R. Paoletti and N. Canal, pp. 219-224, Pergamon Press, New York, 1964.
Maynert, E. W. and Levi, R.: Stress-induced release of brain norepinephrine and its inhibition by drugs. J. Pharmacol. Exp. Therap. 143: 90-96, 1964.
McLean, J. R. and McCartney, M.: Effect of d-amphetamine on rat brain noradrenaline and serotonin. Proc. Soc. Exp, Biol. Med. 107: 77-79, 1961.
Mead, J. A. R. and Finger, K. F.: A single extraction method for the determination of both norepinephrine and serotonin in brain. Bio-chem. Pharmacol. <5: 52-53, 1961.
Mennear, J H.: Interactions between central cholinergic agents and amphetamine in mice. Psychopharmacologia 7: 107-114, 1965.
Mennear, J. H. and Rudzik, A. D.: The effects of amine depleting agents on the toxicity of amphetamine in aggregated mice. Life Scl. 5: 349-356, 1966.
Meyer, H. H.: Theorle des flebers und seine behandlung. Zentbl. Ges. Inn. Med. 6: 385-386, 1913.
Moore, K. E.: Endogenous catecholamines and amphetamine toxicity in aggregated mice. Fed. Proc. 22^: 310, 1963a.
Moore, K. E.: Toxicity and catecholamine releasing actions of d- and 1-amphetamine in isolated and aggregated mice. J. Pharmacol. Exp. Therap. 142: 6-12, 1963b.
Moore, K. E.: The role of endogenous norepinephrine in the toxicity of d-amphetamine In aggregated mice. J. Pharmacol. Exp. Therap. 144: 45-51, 1964.

128
Moore, K. E.: Amphetamine toxicity in hyperthyroid mice: effects on blood glucose and liver glycogen. Biochem. Pharmacol. 15: 353-360, 1966a.
Moore, K. E.: Effects of (+)-amphetamine on lactic acid concentrations in the tissues of aggregated mice. J. Phartn. Pharmacol. 18: 196-197, 1966b.
Moore, K. E.: Effects of d-amphetamine on plasma and tissue electrolyte concentrations of aggregated and of hyperthyroid mice. Proc. Soc. Exp. Biol. Med. 122: 292-295, 1966c.
Moore, K. E. and Lariviere, E. W.: Effects of d-amphetamine and restraint on the content of norepinephrine and dopamine in rat brain. Biochem. Pharmacol. 12: 1283-1288, 1963.
Moore, K. E., Sawdy, L. C. and Shaul, S. R.: Effects of D-amphetamine on blood glucose and tissue glycogen levels of isolated and aggregated mice. Biochem. Pharmacol. 14: 197-204, 1965.
Moore, S. and Stein, W. H.: Chromatography of amino acids on sulfonated polystyrene resins. J. Biol. Chem. 192: 663-681, 1951.
Morpurgo, C. and Theobald, W.: Influence of imipramine-like compounds and chlorpromazine on the reserpine-hypothermia in mice and the amphetamine-hyperthermia in rats. Med. Pharmacol. Exp. 12: 226-232, 1965.
Morpurgo, C. and Theobald, W.: Pharmacological modifications of the amphetamine-induced hyperthermia in rats. Europe J. Pharmacol. 2: 287-294, 1967.
Mount, L. E.: The influence of huddling and body size on the metabolic rate of the young pig. J. Agric. Sci. 55/. 101-105, 1960.
Mount, L. E. and Willmott, J. V.: The relation between spontaneous activity, metabolic rate and the 24 hour cycle in mice at different environmental temperatures. J. Physiol. (London) 190: 371-380, 1967.
Myers, R. D. and Yaksh, T. L.: Feeding and temperature responses in the unrestrained rat after injections of cholinergic and aminergic substances into the cerebral ventricles. Physiol. Behav. 3: 917-928, 1968.
Nagatsu, T., Levitt, M. and Udenfriend, S.: Tyrosine hydroxylase--the initial step in norepinephrine biosynthesis. J. Biol. Chem. 239: 2910-2917, 1964.

129
Obal, F., Mozes, M. and Erdei, P.: Role of the nervous system In the action of calorlgenic substances with different modes of action. Acta Physiol. Acad. Sci. Hung. 7: 245-249, 1955.
Oliverio, A. and Stjarne, L.: Acceleration of noradrenaline turnover in the mouse heart by cold exposure. Life Sci. 4: 2339-2343, 1965.
Pletscher, A.: Release of 5-hydroxytryptamine by benzoquinolizine derivatives with a sedative action. Science, 126: 507, 1957.
Pletscher, A. and Bartholinl, G.: Drug-induced changes of uptake and metabolism of 5-hydroxytryptamine by brain slices. Med. Pharmacol. Exp. 16: 432-440, 1967.
Pletscher, A., Burkard, W. P. and Gey, K. F.: Effect of monoamine releasers and decarboxylase inhibitors on endogenous 5-hydroxy-Indole derivatives in brain. Biochem. Pharmacol. 13: 385-390, 1964.
Pletscher, A. and Da Prada, M.: Veranderung des cerebralen dopamin-metabollsmus durch psychopharmaca. Helv. Physsio. Pharmacol. Acta 24: C45-C47, 1966.
Porter, C. C., Totaro, J. A. and Leiby, C. M.: Some biochemical effects of ot-methyl-3,4-dihydroxyphenylalanine and related compounds in mice. J. Pharmacol. Exp. Therap. 134; 139-145, 1961.
Qulnn, G. P., Shore, P. A. and Brodie, B. B.: Biochemical end pharmacological studies of Ro 1-9569 (tetrabenazlne), a non-indole tranqullizlng agent with reserpine-like effects. J. Pharmacol. Exp. Therap. 127: 103-109, 1959.
Randrup, A. and Munkvad, I.: Stereotyped activities produced by amphetamine in several animal species and man. Psychopharmacologia 11: 300-310, 1967.
Randrup, A. and Scheel-Kruger, J.: Dlethyldithiocarbamate and amphetamine stereotype behaviour. J. Pharm. Pharmacol. l£k 752, 1966.
Rech, R. H.: Antagonism of reserplne behavioral depression by d-ampheta-mlne. J. Pharmacol. Exp. Therap. 146: 369-376, 1964.
Rech, R. H., Borys, H. D. and Moore, K. E.: Alterations In behavior and brain catecholamine levels in rats treated with oC-methyltyrosine. J. Pharmacol. Exp. Therap. 153: 412-419, 1966.
Reid, W. D., Volicer, L. Smookler, H., Beaven, M. A. and Brodie, B. B.: Brain amines and temperature regulation. Pharmacology 1^ 329-344, 1968.

130
Reshef, L. and Shapiro, B.: Effect of epinephrine, cortisone, and growth hormone on release of une&terlfied fatty acids by adipose tissue in vitro. Metabolism 9: 551-555, 1960.
Kewerski, W. and Jori, A.: Bilateral lesion of anterior hypothalamus and regulation of body temperature in rats. Med. Pharmacol. Exp. 16: 355-364, 1967.
Rosso, R., Dolfini, L. and Franchi, G.: Metabolism of amphetamine in tumor bearing rats. Biochera. Pharmacol. 17: 633-634, 1968.
Rubin, R. P. and Jaanus, S. D.: A study of the release of catecholamines from the adrenal medulla by indirectly acting sympathomimetic amines. Arch. Pharmakol. Exp. Pathol. 254: 125-137, 1966.
Samorajskl, T. and Marks, B. H.: Localization of tritiated norepinephrine in mouse brain. J. Hlstochem. Cytochem. 10: 392-399, 1962.
Sato, T. L. Jequier, E., Lovenberg, W. and Sjoerdsma, A.: Characterization of a tryptophan hydroxylating enzyme from malignant mouse cells. Europe. J. Pharmacol. _1: 18-25, 1967.
Shafrir, E. and Steinberg, D.: The essential role of the adrenal cortex in the response of plasma free fatty acids, cholesterol, and phos-pholipides to adrenaline injection. J. Clin. Invest. 3_9_: 310-319, 1960.
Shellenberger, M. K. and Elder, J. T.: Changes in rabbit core temperature accompanying alterations in brainstem monoamine concentrations. J. Pharmacol. Exp. Therap. 158: 219-226, 1967.
Shellenberger, M. K. and Elder, J. T.: Alterations in rabbit core temperature produced by alpha-methyl metatyrosine, Europe. J. Pharmacol. 3: 196-202, 1968.
Shore, P. A., Alpers, H. S. and Busfield, D.: On the mechanism of norepinephrine depletion by reserpine, metaramino1 and related compounds and antagonism by monoamine oxidase inhibition. JLn Mechanisms of Release of Biogenic Amines, ed. by U. S. von Euler, S. Rosell and B. Uvnas, pp. 319-329, Pergamon Press, New York, 1966.
Shore, P. A. and Olin, J. S.: Identification and chemical assay of norepinephrine in brain and other tissues. J. Pharmacol. Exp. Therap. 122: 295-300, 1958.
Slmmonds, M. A. and Iversen, L. L.: Thermoregulation: effects of environmental temperature on turnover of hypothalamic norepinephrine. Science 163: 473-474, 1969.

131
Slmonyl, J. and Szentgyorgyi, D.: The effect of acedron (phenylisopro-pylamine) and thyroxine on the body temperature. Arch. Int. Fharmacodyn. 80: 1-14, 1949.
Slater, P. and Turnbull, M. J.: Hypothermia following intraventricular injection of homocarnosine in the rat. Life Sci. 8: 441-447, 1969.
Smith, C. B.: Effects of d-amphetamine upon brain amine content and locomotor activity of mice. J. Pharmacol. Exp. Therap. 147: 96-102, 1965.
Snedecor, G. W. and Cochran, W. G.: Statistical Methods, pp. 44-51, Iowa State University Press, Ames, Iowa, 1967.
Sokoloff, L. Perlin, S., Kornetsky, C. and Kety, S. S.: The effects of d-lyserglc acid diethylamide on central circulation and overall metabolism. Ann. N. Y. Acad. Sci. 66^: 468-477, 1957.
Somerville, A. R. and Whittle, B. A.: The interrelation of hypothermia and depletion of noradrenaline, dopamine and 5-hydroxytryptamine from brain by reserpine, p-chlorophenylalanine, and «<-methylmeta-tyrosine. Brit. J. Pharmacol. 3_1: 120-131, 1967.
Spector, S., Sjoerdsma, A. and Udenfriend, S.: Blockade of endogenous norepinephrine synthesis by oc-methyl-tyrosine, an inhibitor of tyrosine hydroxylase. J. Pharmacol. Exp. Therap. 147: 86-95, 1965.
Stein, L.: Self-stimulation of the brain and the central stimulant action of amphetamine. Fed. Proc. 23_: 836-850, 1964.
Teague, R. S. and Ranson, S. W.: The role of the anterior hypothalamus in temperature regulation. Amer. J. Physiol. 117: 562-570, 1936.
Tedeschl, R. E., Tedeschi, D. H., Ames, P. L., Cook, L., Mattis, P. A. and Fellows, E. J.: Some pharmacological observations on tranylcypromine (SKF trans-385); a potent inhibitor of monoamine oxidase. Proc. Soc. Exp. Biol. Med. 102: 380-381, 1959.
Trendelenburg, U.: Supersensitivity and subsensitivity, to sympathomimetic amines. Pharmacol. Rev. 15: 225-276, 1963.
Turner, T. A. R. and Spencer, P. S. J.: Effect of pretreatment with monoamine oxidase inhibitors or (+)-amphetamine on leptazol convulsions in mice and rats. J. Pharm. Pharmacol. 20: 122S-125S, 1968.
Valzelli, L. Consolo, S. and Morpurgo, C.: Influence of imlpramlne-like drugs on the metabolism of amphetamine. In Antidepressant Drugs, ed. by S. Garattinl and M. N. G. Dukes, pp. 61-69, Excerpta Medlca Foundation, Amsterdam, 1966.

132
Valzelli, L., Dolflni, E., Tansella, M. and Garattini, S.: Activity of centrally acting drugs on amphetamine metabolism. J. Pharm. Pharmacol. 20: 595-599, 1968.
Van Rossum, J. M. and Hurkmans, J. A. Th.: Mechanism of action of psychomotor stimulant drugs. Significance of dopamine in locomotor stimulant action. Int. J. Neuropharmacol. 3: 227-239, 1964.
Van Rossum, J. M., Van der Schoot, J. B. and Hurkmans, J. A. Th.: Mechanism of the action of cocaine and amphetamine in the brain. Experientia 18: 229-231, 1962.
Weil-Malherbe, H., Witby, L. G. and Axelrod, J.: The uptake of circulating (3H) norepinephrine by the pituitary gland and various areas of the brain. J. Neurochem. 8: 55-64, 1961.
Weissman, A. and Koe, B. K.: Contrasting locomotor effects of catecholamine releasers and tyrosine hydroxylase inhibitors in MAO-inhibited mice. Psychopharmacologia 11: 282-286, 1967.
Welch, A. S. and Welch, B. L.: Effect of p-chlorophenylalanine on brain noradrenaline in mice. J. Pharm. Pharmacol. 1£: 632-633, 1967.
Welch, B. L. and Welch, A. S.: Brain norepinephrine and dopamine: rapid elevation and subsequent stimulus-facilitated depletion by d-ampheta mine, Fed. Proc. 28^: 796, 1969.
Wenzel, D. G. and Broadie, L. L.: Effects of group size and cage design on voluntary running activity of amphetamine-treated mice. Arch. Int. Pharmacodyn. 159: 154-160, 1966.
Wiegand, R. G. and Perry, J. E.: Effect of 1-dopa and N-methyl-N-benzyl -2-propynylamlne.HCl on DOPA, dopamine, norepinephrine, epinephrine and serotonin levels in mouse brain. Biochem. Pharmacol. T_\ 181-186 1961.
Wolf, H. H. and George, D. J.: Drug-induced hyperthermia and amphetamine toxicity. J. Pharm. Scl. 52: 748-752, 1964.
Young, R. L. and Gordon, M. W.: The disposition of amphetaminein rat brain. J. Neurochem. 9: 161-167, 1962.
Zalls, E. G., Lundberg, G. D. and Knutson, R. A.: The pathophysiology of acute amphetamine poisoning with pathologic correlation. J. Pharm. Pharmacol. 158: 115-127, 1967.