universita’ degli studi di torino biologia …lavsrl.it/uploads/pdf/tesi alessandra maraffino...
TRANSCRIPT

UNIVERSITA’ DEGLI STUDI DI TORINO
SCUOLA DI SCIENZE DELLA NATURA
Tesi di Laurea Magistrale in
BIOLOGIA DELL’AMBIENTE
Classe di laurea 6/S
LA SIMBIOSI MICORRIZICA (MICOSAT F) IN ZEA MAYS: RILEVANZE BIOCHIMICHE ED ANALISI BROMATOLOGICA DI ALIMENTI
DERIVATI.
Candidato: Relatore: Alessandra Maraffino Mariaconcetta Bruzzoniti
Anno Accademico…2012/2013……..

1.INTRODUZIONE
Il lavoro scientifico riportato in queste pagine vuole descrivere con oggettività le
osservazioni raccolte in circa sei mesi di analisi chimica volta alla rilevazione di
significative differenze nelle determinazioni di diverse specie chimiche tra Zea mays
coltivata in maniera tradizionale e la medesima coltivata in micorrize. Il corpus del lavoro
sperimentale sarà trattato nella seconda parte del testo mentre si è provveduto a redigere
una breve sinossi introduttiva (prima parte) al fine di fornire alcune nozioni basilari
necessarie all’inquadramento delle ipotesi e quindi della tesi.
1.1 Zea mays
Il mais (Zea mays L.) fa parte della sottofamiglia Maydeae, della grande famiglia delle
Poaceae (o Gramineae) e della classe delle Monocotiledoni. Sulla base dei reperti fossili, si
stima che questo gruppo di Poaceae derivi da un ancestrale comune presente 55-70 milioni
di anni fa, verso la fine del regno dei dinosauri. La parola Zea è di origine greca e significa
“vivere” mentre, il nome della specie mays è riconducibile alla parola indiana “mahiz” o
“marisi” che significa “pane di vita”. Il mais è organizzato nel genere Zea, un gruppo di
piante native del Messico e dell’America centrale. Questo genere comprende specie
selvatiche, conosciute come “teosinte” (termine derivato da “teocintli” degli indiani
Nahuátl che significa “seme degli dei”), annuali e perenni. La specie coltivata (Zea mays
L.) deriva dall’addomesticamento di Zea parviglumis iniziato circa 10.000 anni fa nella
valle del fiume Balsas nel sud del Messico. Si trovano anche generi selvatici affini, come
Tripsacum, diffusi dal nord al sud America, che formano cespugli in zone umide, intorno a
insenature o corsi d’acqua. Negli Stati Uniti, Tripsacum dactyloides è impiegata come
foraggio per il bestiame.

Figura 1 - Zea Mays; prime rappresentazioni del mais. Immagine tratta da Mattioli P.A., I discorsi, Venezia 1712
La pianta del mais presenta molte caratteristiche comuni alle altre Poaceae:
• il fusto o culmo distinto in nodi e internodi;
• una singola foglia a ciascun nodo e le foglie distribuite sul culmo in due file
opposte o distiche;
• ogni foglia consiste in una lamina espansa collegata a una guaina che avvolge il
culmo.
I nodi basali hanno la tendenza a formare ramificazioni o culmi di accestimento (polloni) e
sviluppano radici avventizie. E’ una pianta monoica con fiori maschili, verdastri riuniti in
pannocchie terminali; i fiori femminili sono raggruppati in grosse spighe cilindriche
all’ascella delle foglie e sono rivestiti da grandi brattee membranose; hanno stili filiformi,
giallastri, molto lunghi che fuoriescono dalle brattee. I frutti sono cariossidi, ovoidi o
poliedriche, appressate le une alle altre in 8-10 serie longitudinali; le cariossidi sono dure,
lucenti, di colore variabile (giallo, rossastro, bruno, violaceo o bianco).1
In ogni caso è importante approfondire i meccanismi fisiologici di crescita della pianta
(Zea mays spp.), e quindi l’apporto di nutrienti nonché dell’azoto ai fini di verificare
eventuali effetti positivi che potenzialmente possono essere considerati nell’ambito
dell’ottimizzazione delle tecniche colturali del mais e quindi della produttività
agroalimentare. Nei paragrafi successivi verranno presi in considerazione i vari aspetti
biochimici legati alla fisiologia vegetale della pianta, a partire dal ciclo biogeochimico
dell’azoto.

1.2. Il Ciclo dell’Azoto
Molti composti biochimici presenti nelle cellule contengono azoto. Per esempio, l’azoto si
trova nei nucleosidi ortofosfati e negli amminoacidi che formano le unità di base
rispettivamente degli acidi nucleici e delle proteine. Più abbondanti dell’azoto nelle piante
troviamo solo gli elementi carbonio, ossigeno e idrogeno. La maggior parte degli
ecosistemi agricoli e naturali mostra incrementi notevoli nella produttività dopo
concimazione con azoto inorganico, dimostrando l’importanza di questo elemento.
L’azoto è presente nella biosfera in diverse forme. L’atmosfera contiene grandi quantità di
azoto molecolare (N2), circa il 78% del volume totale. Per la maggior parte, questa grande
quantità di azoto non è direttamente disponibile per gli organismi viventi. L’acquisizione di
azoto dall’atmosfera richiede la rottura di un triplo legame covalente (N≡N) per produrre
ammoniaca (NH3) o nitrato (NO3-). Queste reazioni, conosciute meglio con il nome di
fissazione dell’azoto, possono avvenire per mezzo di processi sia naturali sia industriali.
In condizioni di alta temperatura (circa 200°C) ed elevata pressione (circa 200atm) e in
presenza di un catalizzatore metallico (di solito Fe) l’azoto molecolare si combina con
l’idrogeno per formare l’ammoniaca. Queste condizioni estreme sono necessarie per
vincere l’elevata energia di attivazione della reazione. Questa reazione di fissazione
dell’azoto, chiamata processo di Haber-Bosch (noto anche semplicemente come processo
Haber, ove inizialmente si utilizzava come catalizzatore una miscela di osmio e uranio), è il
punto di partenza per la produzione di un gran numero di composti per l’industria e per
l’agricoltura. La produzione industriale mondiale di fertilizzanti azotati ammonta diverse
migliaia di tonnellate annue2.
I processi naturali in grado di fissare l’azoto (circa 190 x 1012g/anno) sono i seguenti:3
• Fulmini. Della quantità totale fissata circa l’8% è dovuta ai fulmini che causano la
formazione di radicali liberi ossidrilici, a partire dal vapor d’acqua e dall’ossigeno
contenuti nell’atmosfera; queste specie reattive attaccano l’azoto molecolare e
formano acido nitrico (HNO3), che precipita al suolo con la pioggia.
• Reazioni fotochimiche. Circa il 2% dell’azoto fissato deriva da reazioni
fotochimiche che coinvolgono ossido nitrico gassoso (NO) e ozono (O3), che
producono acido nitrico.
• Fissazione biologica dell’azoto. Il rimanente 90% deriva dalla fissazione biologica
dell’azoto, praticata da alcuni batteri detti azotofissatori, che grazie all’azione di

enzimi (nitrogenasi) trasformano l’azoto gassoso in ammoniaca la quale, nel mezzo
acquoso del loro citoplasma, passa in soluzione formando ioni ammonio (NH4+).
Da un punto di vista agronomico, la fissazione biologica dell’azoto è una fonte importante
di azoto poiché la richiesta di azoto per le produzioni vegetali non potrebbe essere ottenuta
solamente attraverso l’uso di fertilizzanti chimici.4
Altri processi che intervengono nel ciclo dell’azoto sono:
Ammonificazione. Rappresenta un processo attraverso cui sono resi disponibili nel terreno
ioni ammonio, a partire dai resti vegetali e animali e dai prodotti di rifiuto animali come
feci e urina. Questi ultimi sono decomposti da batteri e funghi del suolo che ne estraggono
le proteine e gli amminoacidi necessari per la propria crescita espellendo ioni ammonio che
sono riutilizzabili dalle piante. In questo modo è operato un efficiente “riciclaggio”.
Nitrificazione. Un gruppo di batteri nitrificanti (Nitrosomonas) trasforma gli ioni ammonio
in ioni nitrito (NO2-), che non tendono ad accumularsi perché un altro gruppo (Nitrobacter)
provvede a ossidarli immediatamente a ioni nitrato (NO3-), la forma in cui le piante
assumono dal terreno la maggior parte dell’azoto necessario.
Assimilazione. Gli ioni ammonio e gli ioni nitrato assorbiti dalle radici delle piante sono
assimilati e utilizzati per la biosintesi di amminoacidi e proteine. Le piante sono la sola
fonte di azoto diretta o indiretta per tutti gli animali lungo le catene alimentari.
Denitrificazione. È il processo che chiude il ciclo dell’azoto, grazie all’azione di un gruppo
di batteri detti denitrificanti che operano nel terreno in assenza di ossigeno, trasformando i
nitrati in azoto molecolare che ritorna nell’atmosfera (con questa reazione ricavano
l’ossigeno necessario per la propria respirazione).
Tra tutti i nutrienti essenziali per le piante, l’azoto è quello che maggiormente tende a
scarseggiare. Tra le cause principali vi è l’elevata solubilità dei sali di ammonio e dei
nitrati che li rende facilmente allontanabili per dilavamento ad opera delle precipitazioni,
con conseguente accumulo negli ecosistemi acquatici (in particolare quelli lacustri). Nei
terreni agricoli le perdite di azoto sono rilevanti sia perché il suolo è più esposto al
dilavamento, sia perché una parte dell’azoto viene continuamente sottratta ai terreni con le
piante stesse quando si effettua il raccolto. Per questo motivo è necessario impiegare
ingenti quantità di fertilizzanti azotati. L’eccesso di questi composti, insieme a quello dei
fosfati (anch’essi usati abbondantemente come fertilizzanti) si riversa per dilavamento nei
corsi d’acqua e si accumula nei laghi causando un tipo di inquinamento noto come
eutrofizzazione che causa un anomalo sviluppo di alghe. Come conseguenza si determina
un’intensificazione dei processi di decomposizione, che comportano un forte consumo di

ossigeno che ha come effetto la drastica riduzione o la morte di molte specie
dell’ecosistema lacustre.5
Elevate concentrazioni di ammonio, accumulate nei tessuti vegetali, risultano tossiche sia
per la pianta sia per gli animali, che di queste si nutrono. L’ammonio dissipa il gradiente
protonico transmembrana indispensabile per il trasporto degli elettroni sia della fotosintesi
sia della respirazione come anche per l accumulo di metaboliti nel vacuolo. Poiché elevate
concentrazioni di ammonio sono pericolose, gli animali hanno sviluppato una tipica
avversione al suo odore. Le piante assimilano azoto attraverso i siti di assorbimento, quali
radici e peli radicali e accumulano rapidamente qualsiasi eccesso nei vacuoli evitando così
i suoi effetti tossici sulle membrane o sul citosol.
A differenza dell’ammonio, le piante possono accumulare elevate quantità di nitrato e
possono traslocarlo da tessuto a tessuto senza effetti dannosi. Comunque se gli animali o le
persone consumano vegetali con alto contenuto di nitrato, possono sviluppare
metemoglobinemia, ovvero l’ossidazione del Fe(II) a Fe(III) nell’eme dell’Hb che
determina cianosi quando la metaemoglobina supera livelli ematici di 1.5g/l. Si ha grave
ipossia tissutale quando si presentano livelli di metaemoglobina pari al 30% (traslazione
della curva di dissociazione dell’O2 a sx). Animali e uomo possono anche convertire il
nitrato in nitrosammine (CYP450), che sono composti cancerogeni.
1.3 Assimilazione del nitrato
Le radici delle piante assorbono attivamente il nitrato dalla soluzione del suolo tramite
numerosi cotrasportatori nitrato-protone a bassa ed elevata affinità6.
Le piante assimilano la maggior parte del nitrato assorbito, in composti organici azotati. Il
primo passaggio di questo processo è la riduzione citosolica del nitrato in nitrito7 .
L’enzima nitrato reduttasi (NR), una metalloflavoproteina (EC 1.7.1.1. - 3), catalizza la
reazione generale:
dove NAD(P)H indica NADH o NADPH. In generale le isoforme della NR isolate nelle
piante superiori, nei funghi e nelle alghe sfruttano la cooperazione con i piridin nucleotidi
(NADH o NADPH) secondo lo schema generale:
NR-NADH
(EC 1.7.1.1)
NR-NAD(P)H
(EC 1.7.1.2)
NR-NADPH
(EC 1.7.1.3)
La forma più comune di NR nelle piante superiori utilizza solo NADH come donatore di
elettroni, mentre un’altra forma dell’enzima si trova principalmente in tessuti non verdi
come le radici, e può utilizzare sia NADH sia NADPH8.
Figura 2 - stuttura chimica del NADP
La NR è una metalloflavoproteina contenente un eme-Fe ed un atomo di Mo complessato
con una pterina formante il c.d. Complesso molibdopterinico (Mo-MPT). La NR catalizza
una serie molto vasta di reazioni, in particolare NADH-dipendenti. L'enzima catalizza la
Piante
Funghi
Piante
Funghi
Alghe
Funghi

riduzione del NO3- a NO2- attraverso il flusso elettronico generato dalla formazione di
complessi catalitici tra i cofattori ed il metallo prostetico, secondo l'ordine NADH → FAD
→ Mo → NO3-. Dal punto di vista strutturale l'NR è costituito da due omodimeri di massa
~ 100kD, ognuno dei quali contenente un gruppo FAD, un eme-Fe ed un gruppo Mo-MPT.
Va tenuto conto che esistono almeno 40 tipi di sequenze polipeptidiche codificate per la
NR. La NR è costituita da cinque domini strutturali distinti: Mo-MPT, interfaccia dimerica,
citocromo b (cyt b), FAD, NADH. Nella miscela di reazione quando interagiscono FAD-
NADH si ha la formazione del c.d. Frammento della citocromo b reduttasi (CbR), e
quando questo va ad interagire a sua volta con il cyt b, si forma il frammento della
citocromo c reduttasi (CcR). Regioni caratteristiche della catena polipeptidica sono la N-
terminale, ricca di residui amminoacidici acidi, la Hinge 1 che contiene il sito di
fosforilazione dell'enzima (in genere un residuo di Ser) e la Hinge 2 che contiene il sito per
le proteinasi. Il dominio Mo-MPT e l'interfaccia dimerica sono localizzati in prossimità ad
enzimi di membrana come la solfito ossidasi (SOX – EC 1.8.2.1), che catalizza la reazione
Figura 3 - Subunità della NR (EC 1.7.1.3.) dove è visibile la Mo-MPT. Il rendering elaborato pone in evidenza la struttura secondaria della proteina. Per il gruppo Mo-MPT si è utilizzato il modello CPK.

Figura 4 - Subunità della NR (EC 1.7.1.1.) dove è visibile il FAD. Il rendering elaborato pone in
evidenza la struttura secondaria della proteina. Il FAD è rappresentato con lo scheletro molecolare.
Il citocromo associato alla reazione della SOX è il c-551. In generale possiamo dire che la
NR esiste in forme ossidate e ridotte, in particolare lo stato redox dell'enzima dipende da
quello dei suoi cofattori e gruppi prostetici, infatti riscontriamo per il FAD 3 stati redox, 2
per il Fe, 3 per il Mo. L'enzima (omodimero) può assumere dai 12 ai 18 stati redox. La
corrente elettronica che si genera per la riduzione del NO3- durante la catalisi enzimatica è
determinata dai potenziali di riduzione di inizio/fine della reazione, in particolare E°=-
320mV per l'NADH ed E°=420mV per il NO3-, corrispondente ad un ∆E°=740mV, ovvero
una variazione di energia libera per la reazione di riduzione del NO3- pari a ∆G°=-
34.2kcal/mol (-143kJ/mol), che risponde sostanzialmente a dire che la reazione è
termodinamicamente irreversibile (equilibrio spostato totalmente vs. i prodotti)9.
Figura 5 - Struttura chimica del FAD

Il meccanismo catalitico dell’enzima fin qui brevemente descritto, come già detto, culmina
con il trasferimento di elettroni dal gruppo MO-MPT al NO3-, come schematizzato in
figura 6.
Figura 6 - Struttura chimica della Molibdopterina
La NR è la principale proteina contenente molibdeno dei tessuti vegetali e, un sintomo
della carenza da molibdeno è l’accumulo di nitrato, derivante dalla diminuzione
dell’attività della nitrato reduttasi10.
Il dominio della NR che lega il FAD, accetta due elettroni dal NADH o dal NADPH. Gli
elettroni passano quindi attraverso il dominio dell’eme per giungere al complesso
molibdeno, dove sono trasferiti al nitrato.
Figura 7 - Ciclo catalitico della nitrato reduttasi
E’ stato dimostrato che il nitrato è presente all’interno delle cellule delle piante in due pool
differenti: un pool di stoccaggio e un pool metabolico. Il nitrato presente nel pool

metabolico è direttamente disponibile per la piante e induce l’attività della NR. La luce, il
glucosio ed altri fattori ambientali possono influire sulla nitrato reduttasi. E’ stato osservato
che sottoponendo la pianta alla luce e calcolando la concentrazione di nitrito dopo 6 e dopo
12 ore dalla presenza della luce, la produzione di nitrito cessa in entrambe i casi dopo 4 ore;
aggiungendo al terreno il nitrato, la produzione di nitrito riprende e continua per almeno 3
ore. Ciò dimostra che l’enzima rimane attivo e il raggiungimento del plateaux nella
produzione di nitrito è dovuto ad una reale deplezione di nitrato nel pool metabolico. Dopo
12 ore di illuminazione la produzione totale di nitrito è circa due volte maggiore rispetto
alla produzione di nitrito dopo 6 ore11.
Figura 8 - Cinetica della NR in condizioni di differente illuminazione Inoltre nelle foglie illuminate, si è osservato un incremento dell attività della nitrato
reduttasi, all’aumentare del nitrato nel pool metabolico (8).
Velocità di reazio
Tempo/[h]
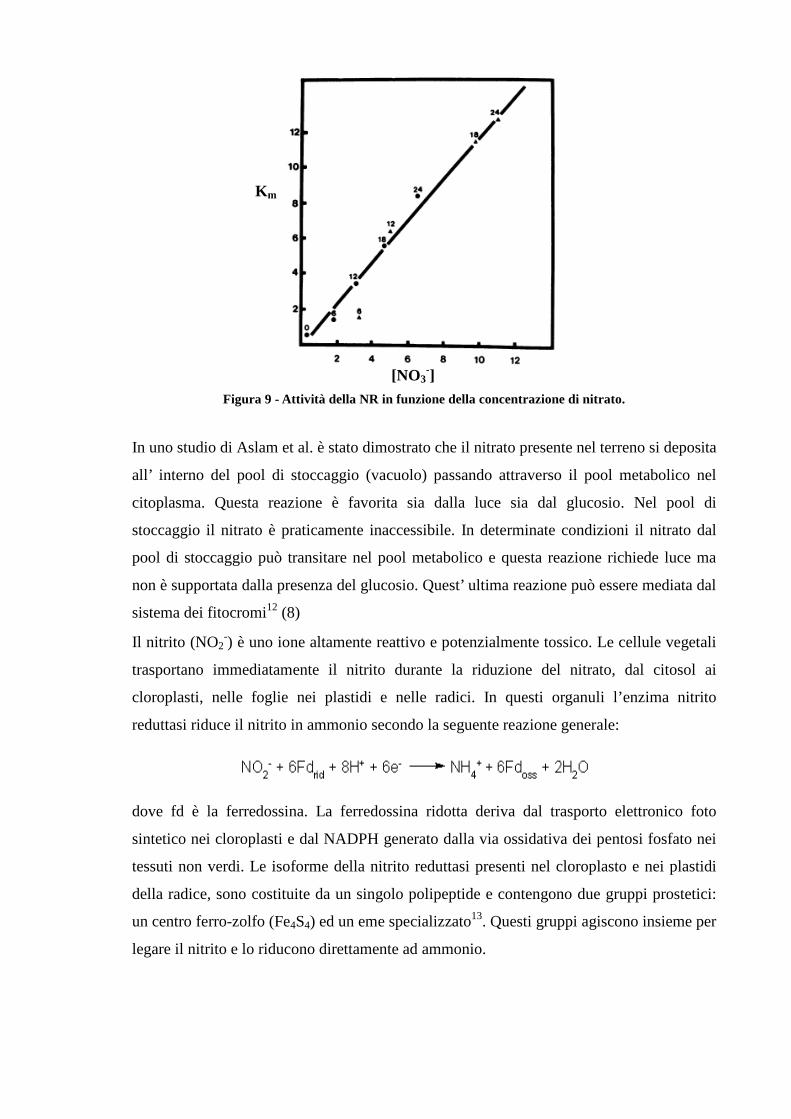
Figura 9 - Attività della NR in funzione della concentrazione di nitrato.
In uno studio di Aslam et al. è stato dimostrato che il nitrato presente nel terreno si deposita
all’ interno del pool di stoccaggio (vacuolo) passando attraverso il pool metabolico nel
citoplasma. Questa reazione è favorita sia dalla luce sia dal glucosio. Nel pool di
stoccaggio il nitrato è praticamente inaccessibile. In determinate condizioni il nitrato dal
pool di stoccaggio può transitare nel pool metabolico e questa reazione richiede luce ma
non è supportata dalla presenza del glucosio. Quest’ ultima reazione può essere mediata dal
sistema dei fitocromi12 (8)
Il nitrito (NO2-) è uno ione altamente reattivo e potenzialmente tossico. Le cellule vegetali
trasportano immediatamente il nitrito durante la riduzione del nitrato, dal citosol ai
cloroplasti, nelle foglie nei plastidi e nelle radici. In questi organuli l’enzima nitrito
reduttasi riduce il nitrito in ammonio secondo la seguente reazione generale:
dove fd è la ferredossina. La ferredossina ridotta deriva dal trasporto elettronico foto
sintetico nei cloroplasti e dal NADPH generato dalla via ossidativa dei pentosi fosfato nei
tessuti non verdi. Le isoforme della nitrito reduttasi presenti nel cloroplasto e nei plastidi
della radice, sono costituite da un singolo polipeptide e contengono due gruppi prostetici:
un centro ferro-zolfo (Fe4S4) ed un eme specializzato13. Questi gruppi agiscono insieme per
legare il nitrito e lo riducono direttamente ad ammonio.
Km
[NO3-]

Figura 10 - Rendering dell'enzima nitrito reduttasi (EC 1.7.2.1.) dove è visibile la struttura secondaria del polipeptide nonchè la struttura molecolare dei cofattori.
Anche se non vi è accumulo di composti azotati o di stati redox intermedi, una piccola
percentuale (0,02-0,2%) di nitrato ridotto è liberata come ossido nitroso (N2O), un gas da
effetto serra14.
La nitrito reduttasi è codificata nel nucleo e sintetizzata come precursore che reca un
peptide di transito all’ N-terminale che la indirizza verso il plastidio15.
Elevate concentrazioni di NO3- e la luce inducono la trascrizione dell’mRNA della nitrito
reduttasi. L’accumulo di prodotti finali del processo come l’asparagina e la glutammina,
reprime questa induzione.
In molte piante, quando le radici ricevono piccole quantità di nitrato, esso è ridotto prima
nelle radici. Come aumenta la disponibilità di nitrato una quantità via via superiore di
nitrato è trasferita al germoglio e qui assimilata16.
Anche in condizioni simili di disponibilità di nitrato il bilancio fra il metabolismo del
nitrato fra radici e germoglio, misurato come proporzione di attività di nitrato reduttasi in
ognuno di questi due organi o dalla concentrazione relativa di nitrato e azoto ridotto del
succo xilematico, varia da specie a specie.
Al fine di individuare il range di stabilità delle specie anioniche e relativi equilibri di
ossidoriduzione dell’azoto nel suolo, è necessario prendere in considerazione l’analisi di
potenziali di riduzione in fuzione del pH. Ciò è possibile procedendo al calcolo delle curve
E = f(pH) a partire dalla relazione di Nerst per il potenziale elettrodico di semicella. Le

curve vengono successivamente graficate nel tradizionale diagramma di Pourbaix E[V]/pH
che si riporta di seguito.
Figura 11 - Diagrmma di pourbaix per le specie chimiche dell'azoto nel suolo.
1.4 Assimilazione dell'ammonio
Le cellule vegetali evitano la tossicità dell’ammonio convertendo rapidamente l’ammonio
generato dall’assimilazione del nitrato o dalla foto respirazione in amminoacidi. La via
principale per questa conversione coinvolge l’azione sequenziale della glutammina
sintetasi e della glutammato sintasi.
La glutammina sintetasi unisce l’ammonio con il glutammato per formare la glutammina:
Questa reazione richiede l’idrolisi di ATP e coinvolge uno ione bivalente come Mg2+,
Mn2+ o Co2+ come cofattore. Le piante possiedono due classi di GS, una nel citosol e
l’altra nei plastidi delle radici o nei cloroplasti del germoglio. Le forme citosoliche sono
espresse nei semi in via di germinazione o nei fasci vascolari di radici e germogli e
producono glutammina per il trasporto intracellulare dell’azoto. La GS dei plastidi delle
radici produce azoto ammidico per il consumo locale; la GS dei cloroplasti del germoglio
riassimila l’NH4+ della foto respirazione17.

La luce e la quantità di carboidrati alternano l’espressione delle forme plastidiali
dell’enzima, ma hanno uno scarso effetto sulle forme citosoliche.
Elevate quantità di glutammina plastidiale stimolano l’attività della glutammato sintasi
(GOGAT). Questo enzima trasferisce il gruppo ammidico della glutammina al 2-
oxoglutarato, formando due molecole di glutammato. Le piante contengono due tipi di
GOGAT: una accetta elettroni dal NADH, l’altra accetta gli elettroni dalla ferredossina
(Fd):
La forma NADH dipendente dell’enzima (NADH-GOGAT) è localizzata nei plastidi dei
tessuti non foto sintetici quali le radici o i fasci vascolari delle foglie in via di sviluppo.
Nelle radici la NADH-GOGAT è coinvolta nell’ assimilazione dell’NH4+ assorbito dalla
rizosfera (il suolo vicino alle radici); nei fasci vascolari delle foglie in via di sviluppo la
NADH-GOGAT assimila la glutammina traslocata dalle radici alle foglie senescenti.
Figura 12 - Rendering della glutammato sintasi dove sono visibili le subunità nonché la struttura
secondaria del polipeptide.
La forma della glutammato sintasi ferredossina dipendente (Fd-GOGAT) si trova nei
cloroplasti e serve per il metabolismo dell’azoto liberato durante la foto respirazione. La
quantità di proteina e l’attività dell’enzima aumentano con l’aumentare della luce. Le
radici, in particolare quelle nutrite con nitrato, hanno la Fd-GOGAT nei plastidi. La Fd-
GOGAT delle radici serve probabilmente per incorporare la glutammina generata durante
l’assimilazione del nitrato.

L’ammonio può essere assimilato attraverso vie alternative. La glutammato deidrogenasi
(GDH) catalizza la reazione reversibile capace di sintetizzare o deaminare il glutammato (v.
Fig.4).
Una forma NADH dipendente della GDH si trova nei mitocondri e una forma NADPH è
localizzata nei cloroplasti degli organi foto sintetici. Sebbene entrambe le forme siano
relativamente abbondanti, non può sostituire la via GS-GOGAT per l’assimilazione
dell’ammonio e la funzione principale della DGH è la deaminazione del glutammato
durante la riallocazione dell’azoto.
Una volta assimilato nella glutammina e nel glutammato, l’azoto è incorporato in altri
amminoacidi attraverso reazioni di transamminazione. Queste reazioni vengono portate a
termine da enzimi conosciuti come amminotransferasi. Un esempio è la aspartato
amminotransferasi che catalizza la seguente rezione:
in cui l’ammino gruppo del glutammato è trasferito sul gruppo carbossilico dell’ aspartato.
L’aspartato è un amminoacido che prende parte dello shuttle malato-aspartato per trasferire
equivalenti di riduzione dal mitocondrio e dal cloroplasto al citosol e nel trasporto del
carbonio dal mesofillo alla guaina del fascio nella fissazione del carbonio nelle piante C4.
Tutte le reazione di transamminazione richiedono come cofattore il piridossal fosfato
(vitamina B6).
Le amminotrasferasi si trovano nel citoplasma, nei cloroplasti, nei mitocondri, nei
gliossisomi e nei perossisomi. Le amminotrasferasi localizzate nei cloroplasti possono
avere un ruolo importante nella biosintesi degli amminoacidi, poiché quando le foglie o i
cloroplasti isolati sono esposti ad anidride carbonica marcata, avviene una rapida
incorporazione del biossido di carbonio radioattivo nel glutammato, nell’aspartato,
nell’alanina nella serina e nella glicina.
Le persone e la maggior parte degli animali non sono in grado di sintetizzare certi
amminoacidi, come istidina, isoleucina, leucina, lisina, metionina, fenilalanina, treonina,
triptofano, valina e arginina (solo i giovani, gli adulti possono sintetizzare arginina), e
devono così ottenere questi cosiddetti amminoacidi essenziali dalla loro dieta. Invece le
piante sintetizzano tutti i 20 amminoacidi presenti nelle proteine. Come riportato in

precedenza, gli ammino gruppi contenenti azoto derivano da reazioni di transamminazione
con la glutammina o con il glutammato. Lo scheletro carbonioso degli amminoacidi deriva
dal 3-fosfoglicerato, dal fosfoenolpiruvato o dal piruvato generati durante la glicolisi o dal
2-oxoglutarato o dall’ossalacetato generati nel ciclo dell’acido citrico.
1.5 Fissazione biologica dell’azoto
La fissazione biologica dell’azoto è responsabile della maggior parte della fissazione
dell’N2 atmosferico in ammonio.
Alcuni batteri possono convertire l’azoto atmosferico in ammonio. La maggior parte di
questi procarioti azoto-fissatori vive libera nel suolo di solito in modo indipendente da altri
organismi. Pochi di essi formano associazioni simbiontiche con le piante superiori in cui il
procariote fornisce direttamente alla pianta ospite l’azoto fissato in cambio di altri nutrienti
e carboidrati.
I sistemi enzimatici utilizzati per la fissazione dell’azoto sono identici in quasi tutti i batteri,
nonostante le differenze esistenti tra questi, e necessitano di cinque requisiti essenziali:
1. La nitrogenasi, la proteina che catalizza la riduzione dell’N2 ad ammonio.
2. Un potente riducente, come la ferrodossina ridotta (o flavodossina).
3. L’ATP.
4. Le condizioni anaerobiche.
5. I meccanismi di regolazione.
I principali meccanismi di regolazione sono due:
• L’ADP inibisce l’attività della nitrogenasi; quindi quando il rapporto ATP/ADP
diminuisce, la fissazione dell’azoto è inibita.
• L’NH 4+ reprime l’espressione dei geni nif, i geni che codificano per le proteine del
sistema di fissazione dell’azoto. Fino ad oggi, sono stati identificati 20 geni nif.
La repressione dell’espressione del gene nif, da parte dell’ammonio, il principale prodotto
della fissazione dell’azoto, è un sistema efficiente ed efficace per sospendere questo
processo metabolico quando il suo prodotto finale non è necessario18.
Poiché la fissazione dell’azoto coinvolge il trasferimento di grandi quantità di energia, gli
enzimi della nitrogenasi possiedono siti che facilitano lo scambio di elettroni ad alta
energia. L’ossigeno essendo un forte accettore di elettroni può danneggiare questi siti e
disattivare irreversibilmente la nitrogenasi, così l’azoto deve essere fissato in condizioni
anaerobiche.

I batteri azoto fissatori liberi sono aerobi (Azotobacter, Gloeothece) facoltativi o anaerobi i
quali possono essere sia fotosintetici (Rodospirillum), sia non fotosintetici (Clostridium).
Ognuno di questi organismi azoto fissatori può agire in condizioni anaerobiche o in
presenza di ossigeno può crescere in un ambiente anaerobico interno alla pianta ospite.
Nei cianobatteri per esempio, le condizioni sono create in cellule specializzate dette
eterocisti. Si tratta di cellule con parete ispessita e prive del fotosistema due, il quale
sviluppa ossigeno nei cloroplasti; in questo modo riescono a non produrre ossigeno19
(Burris 1976).
1.6 La fissazione simbiontica dell’azoto
Come già precedentemente detto, alcuni batteri possono formare associazioni simbiontiche
con le piante superiori, traendo un reciproco vantaggio. I procarioti simbionti che fissano
l’azoto dimorano nei noduli radicali. I noduli radicali sono il risultato dell’associazione
simbiotica tra radici di leguminose e batteri appartenenti, per la maggior parte, ai generi
Rhizobium e Bradyrhizobium, capaci di fissare l'azoto atmosferico e cederlo alla pianta in
cambio di carbonio in forma organica.
In natura, le radici delle piante vivono quasi sempre in simbiosi con funghi del suolo,
formando le micorrize. In queste associazioni il fungo fornisce la pianta di fosfati e altri
minerali, e la protegge da diversi tipi di stress e da organismi patogeni, mentre la pianta
cede zuccheri al fungo. Molti funghi, inoltre, possono completare il loro ciclo vitale solo se
formano un'associazione micorrizica. Le micorrize si dividono in due grandi gruppi, le
endomicorrize e le ectomicorrize. Nelle ectomicorrize il fungo simbionte avvolge le radici
della pianta ospite formando il mantello o micoclena e penetra tra le cellule epidermiche e
corticali, mantenedosi sempre all'esterno delle pareti cellulari, formando il reticolo di

Hartig .
Le associazioni ectomicorriziche si stabiliscono tra piante arboree forestali (castagni, faggi,
ecc.) e funghi Basidio- ed Ascomiceti come ad es. i Boleti, le Amanite e i tartufi che solo in
simbiosi formano i corpi fruttiferi.
Figura 13 - sezione longitudinale al MO di radice ectomicorrizata
Nelle endomicorrize arbuscolari i funghi simbionti, appartenenti al phylum
Glomeromycota, penetrano all'interno delle cellule epidermiche e corticali della radice,
sempre avvolti dalla membrana plasmatica dell'ospite. Le ife del suolo prendono contatto
con la superficie della radice, su cui formano appressori, e quindi penetrano nella
corteccia dove, esternamente, formano avvolgimenti nelle cellule (i gomitoli) e, più in
profondità, gli arbuscoli, strutture intracellulari ramificate ad alberello, specializzate negli
scambi tra i due simbionti.
Circa l'80% delle piante, tra cui la quasi totalità delle piante erbacee, comprese molte
piante orticole (pomodori, graminacee, ecc. ), e alberi da frutta formano questo tipo di
simbiosi

Figura 14 - sezione longitudinale al MO di radice endomicorrizata
Figura 15 - sezione trasversale comparativa dei tessuti micorrizati arbuscolari ed ectomicorrizati
nonchè non trattati Gli arbuscoli rappresentano lo stadio finale di crescita dei funghi coinvolti in questa
associazione simbiotica poiché dopo 4-10 giorni di simbiosi collassano e la pianta li
degrada completamente lasciando spazio ad una seconda possibile associazione. Durante la
colonizzazione i funghi occupano una porzione molto grande delle cellule corticali della
pianta ma le loro strutture si separano dal protoplasto della cellula vegetale grazie ad una
membrana, la membrana periarbuscolare. Questa membrana circonda completamente gli
arbuscoli aumentando la superficie di assorbimento della membrana plasmatica della
cellula vegetale20.

PARTE SPERIMENTALE – Svolta presso il Laboratorio di Chimica
Merceologica della LAV srl sotto la direzione del Dott. Riccardo Ariotti.
2. FLUSSO METABOLICO DELL’AZOTO ED ANALISI
BROMATOLOGICA DEL LATTE
2.1 Inquadramento generale
Il lavoro sperimentale qui di seguito riportato è volto ad individuare i caratteri biochimici
relativi al metabolismo dell’azoto in Zea Mays coltivata con il sistema Micosat F, ovvero
in simbiosi micorrizica, e a valutare dal punto di vista dell’analisi bromatologica le
variazioni nutrizionali del latte prodotto da allevamenti bovini alimentati con insilato
prodotto a partire da Zea Mays micorrizata.
Il Micosat F® rappresenta sostanzialmente una associazione simbiotica che si instaura tra le
radici di molte piante e funghi AM presenti nel suolo. Questa associazione sinergica è
costituita da un consorzio microbiologico composto da micelio e spore di funghi
micorrizici, funghi saprofiti e batteri della rizosfera agronomicamente utili.
In particolare, come esempio, riportiamo qui di seguito, la formulazione di MICOSAT F
wp semi, utilizzato per la micorizzazione di cereali nella proporzione di 0,8-1,2 kg/ha:
100 g di prodotto contengono un inoculo misto di:
Funghi simbionti:
32 g di radici micorizzate e triturate, spore e ife di funghi simbionti dei generi: Glomus
coronatus GO01 e GU53, Glomus caledonium GM24, Glomus intraradices GB67 e GG32,
Glomus mosseae GP11 e GC11, Glomus viscosum GC41, capaci di punti di ingresso nelle
radici dell’ospite in percentuale minima del 30%;
Altri microrganismi:

Ingredienti biologici attivi 5,7% e in misura minima di 4,25x108 C.F.U./g (di cui 0,8%
Thricoderma spp. in percentuale minima del 3x106 C.F.U./g);
Batteri della rizosfera:
Bacillus subtilis BR48;
Funghi saprofiti:
Beauveria spp. BB48, Trichoderma harzianum TH01, Trichoderma atroviride TA28,
Supporti inerti q.b. a 100.
Lo studio del sistema biologico così descritto è rivolto principalmente all’analisi del flusso
metabolico dell’azoto, che come evincibile dal lavoro di Tian et al. (2010) è responsabile
della maggiore attività proteica della pianta e quindi dell’espressione di determinati geni
codificanti enzimi diversi anche nell’ambito della biosintesi dei carotenoidi (β−carotene).
In tal senso la determinazione dell’azoto totale presente nelle foglie di ZeaMays
micorrizata e di controllo ha dimostrato che Zea Mays coltivata in regime di simbiosi
presenta una quantità di azoto totale kjeldhal nelle foglie di circa il 10% in più rispetto al
controllo. Tale fatto indica confermati i dati degli autori succitati circa le deduzioni sulla
cascata metabolica della glutamina, a livello dell’assimilazione del nitrato dal suolo.
Per quanto concerne le caratteristiche nutrizionali del latte ottenuto da allevamenti
alimentati con insilato prodotto a partire da Zea Mays micorrizato, sono stati determinati i
titoli di vitamina A e vitamina E, nonché il frazionamento degli acidi grassi su due pool:
micorrizato e controllo. Il primo ha prevalso per quantitativo vitaminico, mentre vi è una
differente distribuzione delle quantità relative di determinati acidi grassi tra “micorrizato” e
“controllo”. Si rimanda pertanto alla trattazione dei dati quantitativi riportata nei paragrafi
seguenti.
2.2 Analisi delle foglie di Mais
Si è proceduto inizialmente alla ricostruzione del campione globale a partire dai campioni
elementari, secondo la tabella di campionamento numero 1. La ricostruzione è avvenuta
per omogenizzazione dei campioni elementari e, al fine di ottimizzare la dispersione
granulometrica del macinato, si è provveduto alla setacciatura con porosità del crivello
n.20 (scala Sieve), ciò al fine di eliminare o comunque rendere trascurabile l’effetto
matrice per l’analisi spettroscopica NIR.
Foglie =Campione
2a.1 2a.2 2a.3 2b.1 2b.2 2b.3 2

5a.1 5a.2 5a.3 5b.1 5b.2 5b.3 5
8a.1 8a.2 8a.3 8b.1 8b.2 8b.3 6
11a.1 11a.2 11a.3 11b.1 11b.2 11b.3 11
14a.1 14a.2 14a.3 14b.1 14b.2 14b.3 14
17a.1 17a.2 17a.3 17b.1 17b.2 17b.3 17
20a.1 20a.2 20a.3 20b.1 20b.2 20b.3 20
23a.1 23a.2 23a.3 23b.1 23b.2 23b.3 23
26a.1 26a.2 26a.3 26b.1 26b.2 26b.3 26
29a.1 29a.2 29a.3 29b.1 29b.2 29b.3 29
32a.1 32a.2 32a.3 32b.1 32b.2 32b.3 32
35a.1 35a.2 35a.3 35b.1 35b.2 35b.3 35
38a.1 38a.2 38a.3 38b.1 38b.2 38b.3 38
41a.1 41a.2 41a.3 41b.1 41b.2 41b.3 41
44a.1 44a.2 44a.3 44b.1 44b.2 44b.3 44
47a.1 47a.2 47a.3 47b.1 47b.2 47b.3 47
Tabella 1 - Campionamento Foglie di mais
Il campione globale così realizzato è stato suddiviso in tre aliquote destinate
rispettivamente alla analisi tradizionale Kjeldhal, alla analisi spettroscopica NIR e al
controcampione per la conservazione. Al fine di poter costruire il modello per l’analisi dei
dati spettroscopici a seguito del processamento dei campioni suddetti con lo spettrometro
NIR, sono stati analizzati 10 campioni (5 come controllo e 5 micorrizzati) utilizzando il
metodo tradizionale Kjeldahl per la determinazione dell’azoto totale, a norma ISO
1871:2009. Qui di seguito viene descritto il metodo utilizzato in laboratorio per
determinare l’azoto organico presente all’ interno delle foglie di mais.
2.3 Determinazione dell’azoto totale con il metodo Kjeldahl 2.3.1 Principio del metodo
Il metodo prevede la digestione di un’aliquota del campione utilizzando acido solforico
concentrato in presenza di un catalizzatore per trasformare l’azoto organico in solfato
d’ammonio. Il prodotto finale della digestione viene addizionato di idrossido di sodio (in
eccesso) spostando l’equilibrio acido-base dell’ammonio verso l’ammoniaca, che viene
distillata in eccesso di acido borico al 4%. L’ammoniaca viene poi titolata con una
soluzione standard di acido cloridrico, determinando il tenore di azoto totale.
2.3.2 Reagenti e soluzioni di riferimento concentrate
2.3.2.1 Acido solforico concentrato, ρ = 1,83g/ml – 1,84g/ml;

2.3.2.2 Catalizzatore (compresse di HgO);
2.3.2.3Soluzione di acido borico al 4%w/V;
2.3.2.4 Indicatore: Tashiro;
2.3.2.5 Perossido di Idrogeno non inferiore in titolo al 30%w/w;
2.3.2.6 Idrossido di sodio in titolo non inferiore al 30%w/w;
2.3.2.7 Solfato di Potassio (in compresse o polvere cristallina);
2.3.2.8 Acido cloridrico 0,1M;
2.3.2.9 Solfato d’Ammonio con purezza non inferiore al 99,9%;
2.3.2.10 Triptofano con purezza non inferiore al 99,9%.
2.3.3 Apparecchiatura
2.3.2.1 Bilancia Analitica (precisione 0,001mg);
2.3.2.2 Termoreattore;
2.3.2.3 Unità di distillazione;
2.3.2.5 Buretta e vetreria da laboratorio;
2.3.2.6 Palline di vetro;
2.3.2.7 Carta da peso e spatole;
2.3.2.8 Provettoni da digestione.
2.3.4 Procedimento e modalità operative
Le operazioni effettuate rispettano le indicazioni fornite integralmente nella norma ISO
1871:2009.
Si pesano 5g di campione su bilancia analitica (1.1.3.1), servendosi di carta da peso e spa-
tole (1.1.3.7), trasferendoli quantitativamente nel provettone per la digestione (1.1.3.8);
Lavorando sotto cappa, dopo aver posizionato i provettoni (1.1.3.8) negli idonei spazi nel
termoreattore, si introducono i seguenti reattivi nell’ordine: n.2 compresse di K2SO4/HgO
(3,5g/0,175g) o equivalenti in polvere (1.1.2.7), 5ml di H2O2 (1.1.2.5), 20ml di H2SO4
(1.1.2.1);
Dopo aver posizionato l’apparecchio per l’aspirazione dei fumi sopra i provettoni, si pro-
cede ad azionare il termoreattore già precedentemente impostato su idoneo programma
Temperatura/tempo come stabilito dalla ISO 1871:2009, in modo tale da procedere alla di-
gestione ad una temperatura di 430°C per 45’;

Dopo raffreddamento del sistema, si procede alla distillazione utilizzando l’unità di distil-
lazione (1.1.3.3), avendo cura di aggiungere 50ml di H2O, all’interno di ciascun provettone
e posizionando il medesimo nella idonea sede dell’unità di distillazione (1.1.3.3);
Prima di procedere con la distillazione, si aliquotano 25ml di H3BO3 (1.1.2.3) e alcune
gocce di indicatore tashiro (1.1.2.4) in una beuta da 250ml, avendo cura di riporla nell’ i-
donea sede di distillazione.
Si procede quindi ad avviare la distillazione impostando la durata della corsa (7 minuti) e
la concentrazione di NaOH (50ml).
Dopo la distillazione, si procede alla titolazione dell’ammoniaca aggiungendo nell’apposita
buretta (1.1.3.5) HCl (0,1M) osservando il viraggio della soluzione da verde a viola.
2.3.5 Calcolo ed espressione del risultato
I risultati vengono espressi come percentuale di azoto totale calcolato attraverso la
seguente relazione,
( )1000
10014% 01
××××−=
m
cVVN t
Dove,
V1= volume di titolante impiegato per il campione
V0= volume di titolante impiegato per la prova in bianco
ct= titolo dell’acido cloridrico
m= massa del campione sottoposto all’analisi
2.4 Analisi spettroscopica nel vicino infrarosso (NIR) ed evidenze
sperimentali
Dopo aver analizzato dieci campioni (cinque micorrizati e cinque di controllo) attraverso
il metodo Kjeldhal si è proceduto all’analisi spettroscopica nel vicino infrarosso al fine di
ottenere un modello semplice e rapido per ricavare la percentuale di azoto organico
presente all’ interno di ogni campione di foglie di mais.
Nella spettroscopia di riflettenza nel vicino infrarosso il campione solido finemente
suddiviso viene irradiato (come sorgenti si usano comunemente lampade a tungsteno) con
una o più strette bande di radiazioni di lunghezza d’ onda compresa tra 1 e 2,5 µm. Si ha
una riflettenza diffusa perché la radiazione penetra nello strato superficiale delle particelle,

promuove le vibrazioni della molecola dell’analita e quindi viene diffusa in tutte le
direzioni; si ottiene così uno spettro di riflettenza che dipende dalla composizione del
campione. Lo spettro ha in ordinata il logaritmo del reciproco della riflettenza R, dove R è
il rapporto dei poteri radianti delle radiazioni riflesse dal campione, rispetto la riflettenza di
un campione standard come solfato di bario o ossido di magnesio finemente suddivisi.
Gli strumenti per la misura della riflettenza diffusa sono di produzione corrente; alcuni
usano più filtri di interferenza per produrre strette bande di radiazione mentre altri sono
provvisti di monocromatore. La superficie interna della sfera di integrazione che circonda
il campione è rivestita di solfato di bario, un materiale che assicura una riflessione diffusa
quasi perfetta. La radiazione riflessa del campione arriva al rivelatore (i rilevatori sono
generalmente dei fototubi a solfuro di piombo) dopo riflessioni multiple all’interno della
sfera.21
In particolare, lo strumento con cui è stata svolta l’analisi in laboratorio è un FT-NIR
Bruker Optics MPA equipaggiato con sfera d’integrazione a riflettenza diffusa, il quale
consente la scansione dei campioni e la registrazione di spettri nell’intervallo di numeri
d’onda fra 12000 cm-1 e 3600 cm-1 pari a 833-2778 nm, con una risoluzione di 32 cm-1.
La procedura analitica consisteva nel riporre in una capsula tipo Petri di quarzo,
debitamente pulita con etanolo assoluto, circa 10g di campione ben distribuiti sulla
superficie della capsula stessa: la setacciatura è stata necessaria per eliminare potenziali
interferenze dovute ad eventuale disomogeneità granulometrica. Va infatti ricordato che
l’uniforme distribuzione granulometrica del solido è essenziale per eliminare le
interferenze dovute alla differente distribuzione dell’intensità radiante attraverso le
particelle di solido. Successivamente per la verifica del modello, ovvero della curva di
taratura costruita con i dati ottenuti mediante analisi chimica tradizionale, il laboratorio ha
utilizzato la cross-validation ove si costruisce la curva di taratura con tutti gli standard
(valori ottenuti dal processamento Kjeldahl) ad eccezione di uno il quale viene testato con
il modello di regressione. Si ripete questo procedimento più volte lasciando fuori dalla
curva di taratura uno standard sempre differente ottenendo così una valutazione media
della performance del modello.
Le percentuali di azoto dei 10 campioni di foglie di mais (5 micorrizzati e 5 controlli)
ottenute attraverso il metodo Kjeldhal sono riportate qui di seguito nella tabella 2. E’ stata
anche calcolata la percentuale di proteine presente nel campione moltiplicando il titolodi
azoto per il fattore di correzione pari a 6,25.
% Azoto % Proteine

Nº Camp MIC CONTR MIC CONTR 6 1,87 11,69 32 1,92 12,20 11 1,57 9,83 17 1,96 12,32 38 1,77 11,06 47 1,59 9,95 5 2,15 13,49 20 1,50 9,11 26 1,57 9,82 44 1,48 9,25
Tabella 2 - % di azoto e proteine relative ai campioni di foglie di mais di cui alla Tab.1 con il metodo
Kjieldhal.
Per quanto riguarda l’analisi spettroscopica nel vicino infrarosso, è stato riportato in
Errore. L'origine riferimento non è stata trovata., il tipico spettro NIR ottenuto dai
campioni precedentemente succitati. Inoltre, nella tabella 3 sono state riportate le
percentuali di azoto di tutti i campioni suddivisi in micorrizzati e non, ottenuti attraverso i
modelli predittivi (sono stati elaborati con il NIR due modelli prendendo in esame
separatamente i campioni micorizzati e i campioni di controllo) rappresentati dalle curve di
taratura in figura 3 e 4.
Figura 16 - Spettro FT-NIR relativo a campione di foglie di mais
campioni N_%_sp
MIC CONTR 14 1,91

17 1,75 38 1,81 41 1,87 8 1,87
11 1,57 32 1,95 2 1,29
23 2,01 29 2,20 44 1,18 5 2,15
20 1,1 26 1,57 47 1,59
Tabella 3 - % di azoto relative a campione di foglie di mais ottenute con la NIR
.
2. Analisi del latte
Dopo aver analizzato le foglie di mais, si è provveduto alla determinazione della vitamina
A (retinolo) e della vitamina E (d,l-α-tocoferolo) nella matrice alimentare “latte crudo”
proveniente da allevamenti bovine alimentati con insilato tradizionale e prodotto a partire
da mais micorrizato. Queste vitamine lipofile sono state determinate attraverso un metodo
interno creato sulla base del Rapporto ISTISAN 96/34. Il metodo utilizzato per la
determinazione consta di tre fasi operative: l’idrolisi, l’estrazione con solvente e l’iniezione
in HPLC. Sono stati analizzati in totale 45 campioni di latte e qui di seguito viene riportato
nel dettaglio il metodo utilizzato.
2.1 Determinazione quantitativa del retinolo e del d,l – α – tocoferolo nel
latte crudo
2.1.1 Principio del metodo La determinazione della vitamina A e della vitamina E, rispettivamente il (2E,4E,6E,8E)-
3,7-dimetil-9-(2,6,6-trimetilcicloes-1-enil)nona- 2,4,6,8-tetraen-1-olo ed il 2,5,7,8-
tetrametil-2-(4,8,12-trimetiltridecil)-3,4-diidrocromen-6-olo, avviene attraverso idrolisi
basica e successiva estrazione con CH2Cl2, rivelazione spettrofotometrica a λmax=285nm
dopo separazione in RP-HPLC su SP C18.

2.1.2 Soluzioni d’uso
2.1.2.1 Alcool etilico 95º
2.1.2.2 Soluzione di ascorbato di sodio al 10%
2.1.2.3 Soluzione di idrossido di potassio a 50%
2.1.2.4 Idrossido di potassio 0,5 N
2.1.2.5 Soluzione di solfuro di sodio 0,5 N in glicerina al 70%
2.1.2.6 Diclorometano
2.1.2.7 Fase mobile metanolo-acqua 98:2
2.1.2.8 Alcool metilico
2.1.3 Strumenti ed apparecchiature di laboratorio 2.1.3.1 Piastra riscaldante
2.1.3.2 Evaporatore rotante
2.1.3.3 Filtri a membrana da 0,45 µm
2.1.3.4 Bilancia analitica
2.1.3.5 Cromatografo liquido ad alta risoluzione con rivelatore a 290 nm
2.1.3.6 Colonna HPLC C18 da 25 cm x 4 mm
2.1.3.7 Vetreria da laboratorio
2.1.4 Procedimento
2.1.4.a Idrolisi Basica
Le operazione si conducono al riparo dalla luce utilizzando vetreria ambrata o realizzando
involucri con fogli di alluminio.
Con una pipetta graduata di precisione (classeA) e divisione 0,1ml si prelevano
esattamente 10 ml di campione e si pongono in un pallone da evaporazione da 250 ml di
vetro pyrex. Si aggiungono successivamente 36 ml di alcool etilico (2.1.2.1), 2 ml della
soluzione di ascorbato di sodio (2.1.2.2), 6 ml di idrossido di potassio al 50%w/v (2.1.2.3)
e 2 ml di solfuro di sodio (2.1.2.5).
I palloni da evaporazione si pongono successivamente in bagnomaria bollente con
refrigerante a ricadere per 30 min ± 1min .

2.1.4.b Estrazione in CH2Cl2 Dopo raffreddamento, la miscela di idrolisi si trasferisce quantitativamente all’ interno di
imbuti separatori aggiungendovi 100 ml di diclorometano (2.1.2.6) e 120 ml di idrossido di
potassio 0,5 N (2.1.2.4).
Dopo aver tappato e agitato l’imbuto separatore, si lasciano separare le due fasi
raccogliendo la fase apolare (diclorometano) inferiore direttamente in un atro imbuto
separatore. Ivi si aggiungono 5 ml di idrossido di potassio 0,5 N (2.1.2.4).
Dopo aver tappato e agitato l’imbuto separatore, si lasciano separare le fasi e si raccoglie la
parte inferiore in un altro imbuto separatore.
Operando come in precedenza, si lava la fase apolare con 40 ml di acqua distillata fino alla
scomparsa dell’alcalinità (3 volte).
Figura 17- imbuto separatore contenente la parte diclorometanica inferiore
2.1.4.c Rivelazione in HPLC Dopo aver effettuato i 3 lavaggi, la fase apolare inferiore si trasferisce quantitativamente in
un pallone da evaporazione e si essicca su evaporatore rotante (2.1.3.2) a T=40°C.

Successivamente il film lipidico che si forma sul fondo del pallone è ripreso con 2 o 3 ml
di metanolo (2.1.2.8).
Servendosi di siringa da 5ml e microfiltro da 0,22µm (2.1.3.3) si preparano i vials che
verranno posizionati all’ interno dell’HPLC (2.1.3.5) dove verranno iniettati esattamente 25
µl di estratto. La fase mobile che viene utilizzata per l’analisi in HPLC contiene metanolo
e acqua rispettivamente in rapporto 98:2 (2.1.2.7).
a) Preparazione delle soluzioni standard ed elaborazione della curva di taratura
Le soluzioni standard sono state preparate a partire da soluzioni standard già pronte in
commercio.
Dopo aver preparato le soluzioni standard a concentrazione nota si è proceduto all’
elaborazione della curva di taratura ottenuta utilizzando un software specifico, mathcad.
Si è proceduto a costruire la retta ai minimi quadrati al fine di interpolare i punti originati
dalla correlazione tra l’altezza del picco cromatografico con la concentrazione delle
relative soluzioni standard processate. I grafici ottenuti sono riportati in figura 6. Qui di
seguito sono anche riportate le concentrazioni standard di vitamina A e di vitamina E alle
diverse diluizioni che sono state utilizzate per l’elaborazione della curva di taratura (tabella
4 e 5).
[Vit_A]/[ug/100g] Altezza_del_picco_cromatografico
10 627
50 3997
75 6074
100 8363
[Vit_E]/[ug/100g] Altezza_del_picco_cromatografico
38 52
190 353
284 575
380 798

2.1.5 Espressione del risultato ed evidenze sperimentali La determinazione delle concentrazioni di vitamina A e vitamina E è stata effettuata con il
metodo della retta di calibrazione esposto al paragrafo precedente. Pertanto il calcolo del
titolo di vitamina si ricava a partire dalla forma implicita della rappresentazione analitica
del modello lineare, ottenendo la concentrazione teorica, la quale va corretta per il
coefficiente di diluizione. Quest’ultimo è ricavato a partire dai volumi di diluizione
utilizzati nel protocollo analitico.
dove è il coefficiente di diluizione, in cui Vr è il volume di ripresa finale dal
pallone dopo essicazione su E.R. (2 o 3ml a seconda dei casi) e VI è il volume di campione
sottoposto all’analisi (10ml), hi è l’altezza del picco cromatografico, ci è la concentrazione
di vitamina teorica nel campione, cf è la concentrazione teorica corretta per il coefficiente
di diluizione, m è il coefficiente angolare della retta di calibrazione e q la sua intercetta.
Nel grafico riportato nel paragrafo “Discussione dei Risultati” si riportano i valori medi dei
titoli di vitamine ottenuti nei pool di “controllo” e “micorrizati”, a partire da una
popolazione totale di n.50 campioni di latte crudo.
Tabella 5 - Curva di taratura Vitamina A
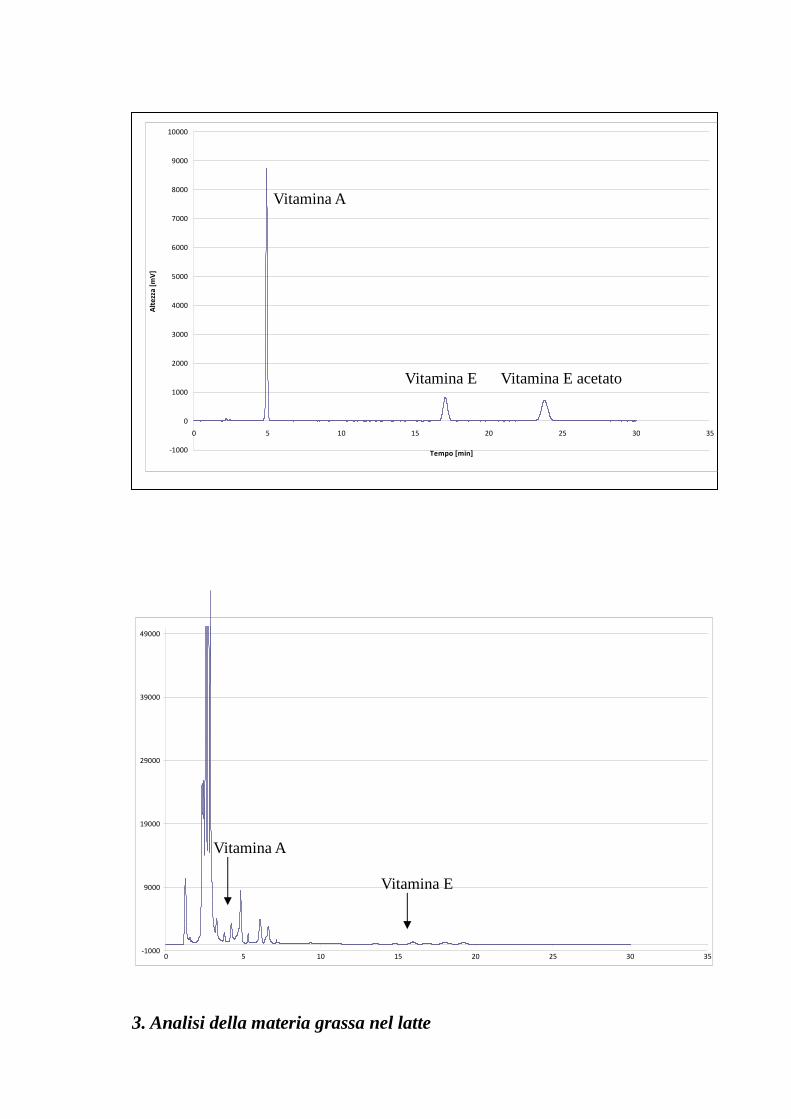
-1000
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
0 5 10 15 20 25 30 35
Tempo [min]
Alt
ezz
a [
mV
]
Vitamina A
Vitamina E Vitamina E acetato
-1000
9000
19000
29000
39000
49000
0 5 10 15 20 25 30 35
3. Analisi della materia grassa nel latte
Vitamina A
Vitamina E

Dopo aver determinato la quantità di vitamina A e di vitamina E, si è proceduto con la
determinazione della quantità di grasso presente all’ interno del campione di latte. Per
l’analisi del grasso è stato messo a punto un metodo interno basato sull’ estrazione dei
grassi dal latte secondo il tradizionale metodo Rose – Gottlieb. Successivamente si è
provveduto all’ esterificazione del grasso utilizzando metanolo e acido solforico, al fine di
ottenere gli esteri metilici degli acidi grassi. Gli esteri metilici volatili sono stati frazionati
in Gas Cromatografia e rilevati con la tecnica (FID). Sono stati analizzati in totale 16
campioni di latte e qui di seguito viene riportato nel dettaglio il metodo utilizzato.
3.1 Determinazione degli acidi grassi nel latte
Il metodo per la determinazione degli acdi grassi nel latte si articola in due fasi: a)
l’estrazione del grasso con il metodo di Rose – Gottlieb:
3.1.2 Principio del metodo
Il contenuto di materia grassa è determinato attraverso l’estrazione da una soluzione
ammoniaco alcoolica del latte con etere etilico ed etere di petrolio, successiva
evaporazione dei solventi e pesata del residuo. Gli acidi grassi estratti vengono
derivatizzati in metilesteri utilizzando metanolo e acido solforico. La determinazione è
stata eseguita in GC-FID.
3.1.3 Reattivi
3.1.3.1 Soluzione di ammoniaca al 30% (m/v) di NH3. Densità a 20ºC circa 0,91 g/ml
3.1.3.2 Alcool etilico 96%
3.1.3.3 Etere etilico esente da perossidi
3.1.3.4 Etere di petrolio distillato tra 30 ºC – 60 ºC
3.1.3.5 Metanolo
3.1.3.6 Acido solforico
3.1.3.7 Esano
3.1.4 Apparecchiature
3.1.4.1 Bilancia analitica
3.1.4.2 Stufa termostatata

3.1.4.3 Evaporatore rotante
3.1.4.4 Vetreria da laboratorio
3.1.4.5 Palline di vetro
3.1.4.6 Bagnetto termostatato
3.1.5 Procedimento
a) Determinazione del grasso
Si sono trasferite in un pallone de evaporazione due o tre perle di vetro (3.1.4.5)
lasciando a essiccare i palloni in stufa ( 3.1.4.2) per circa un ora; dopo il raffreddamento si
pesano i palloni con la precisione di 0,1 mg.
Successivamente si pesano in un cilindro da 100 ml, 10 g di campione e si travasano in un
altro pallone da evaporazione. In seguito si aggiungono 1,5 ml di soluzione di ammoniaca
al 30% (3.1.3.1) e 10 ml di alcool etilico (3.1.3.2) e si mescola accuratamente.
Successivamente si aggiungono 25 ml di etere etilico (3.1.3.3) e 25 ml di etere di petrolio
(3.1.3.4), si tappa il pallone e si agita energicamente per circa un minuto. In seguito si
travasa il contenuto del pallone all’ interno di un imbuto separatore e si lascia a riposo
finchè lo strato superiore divenga limpido e si separi nettamente.
Successivamente si travasa con cura lo strato superiore, nel pallone precedentemente tarato.
Si procede ad una seconda estrazione ripetendo le operazioni descritte a partire dall’
aggiunta dei solventi, utilizzando però soltanto 15 ml di etere etilico (3.1.3.3) e 15 ml di
etere di petrolio (3.1.3.4).
In seguito l’estratto si porta a secco utilizzando l’ evaporatore rotante (3.1.4.3) .
Dopo il raffreddamento a temperatura ambiente, si pesa il pallone con l’ approssimazione
di 0,1 mg.

Figura 18 - Separazione; la fase limpida superiore contiene il grasso estratto
b) Esterificazione
Dopo aver estratto il grasso con il metodo precedentemente succitato, si solubilizzano i
grassi aggiungendo all’ interno del pallone precedentemente tarato, 2 o 3 ml di etere di
petrolio (3.1.3.4).
Successivamente si travasa il grasso solubilizzato all’ interno di provette di vetro le quali si
lasciano essiccare in “bagno maria” a 70 ºC.
Dopo il raffreddamento, si addizionano 3 ml di metanolo (3.1.3.5) e 100 ul di acido
solforico concentrato (3.1.3.6) e dopo aver tappato e agitato con cura le provette, si
depongono in “bagno maria” per 60 minuti a 80 ºC. Dopo il raffreddamento, si aggiunge 1
ml di esano (3.1.3.7), si agita e si separa il surnatante.
c) Separazione in gas cromatografia
Il surnatante precedentemente separato, è stato trasferito all’interno di vials di vetro
ambrato. Successivamente si è proceduto al frazionamento degli acidi grassi iniettando nel
gas cromatografo su colonna capillare polimetilsilossanica (apolare), 1 ul di estratto. Si
sono utilizzati come gas carrier He, N2, e per l’alimentazione della fiamma del FID aria

(70%N,30%O2) e H2. Il cromatogramma tipo per i campioni di controllo è riportato in
figura ___ mentre il cromatogramma tipo per i campioni micorrizati è riportato in
figura____.
La discussione relativa alla distribuzione delle diverse specie di acidi grassi nei pool di
“controllo” e “micorrizati” viene riportata nel paragrafo “Discussione dei risultati”.
d) Preparazione delle soluzioni standard
La determinazione degli acidi grassi in GC-FID viene espressa in percentuale sul grasso,
pertanto le soluzioni standard preparate a partire da uno standard certificato presente in
commercio hanno avuto utilizzo prettamente qualitativo al fine di determinare l’RT per
ognuno dei 37 metil esteri considerati nell’ analisi. A tal fine sono state preparate tre
soluzioni in esano (3.1.3.7) a concentrazioni progressive rispettivamente di 0,1-10-50 ppm.
In figura xy viene riportato il cromatogramma della soluzione standard a 10 ppm.
e) Espressione del risultato
Il contenuto di sostanze grasse del campione, espresso in percentuale, è stato calcolato in
base alla seguente formula:
Sostanze grasse totali (g/100g) = [(M1 – M2)/ S] x 100
M1 = peso in grammi del pallone M contenente la materia grassa dopo l’ operazione a).
M2 = peso in grammi dl pallone M (vuoto) dopo essiccamento dello stesso.
S = aliquota del campione in grammi utilizzata per l’analisi.
3.2 Evidenze sperimentali
La percentuale di grasso dei 16 campioni di latte analizzati, che è stata determinata
attraverso il metodo di Rose –Gottlieb è stata riportata qui di seguito nella tabella 6.
Numero Campioni Grassi [g/100g]
19 3,2

31 4,8
32 4,2
34 5,8
35 4,8
36 6,8
33 4,5
30 5,3
4 4,7
5 5,1
24 4,6
25 4,08
27 3,2
29 3,5 Tabella 6 - Quantità di grasso [g/100g] estratto attraverso il metodo di Rose-Gottli

RT Area Area% Altezza Altezza% A/H Nome 6571 810771 7.10 252822 8.08 3.21 ottanoico 7995 12480 0.11 4220 0.13 2.96 nonanoico 9284 98318 0.86 31285 1.00 3.14 7-nonenoico 9416 2593108 22.72 824196 26.34 3.15 decanoico 11088 16271 0.14 4317 0.14 3.77 undecanoico 12999 5886 0.05 1949 0.06 3.02 fenolo 13215 12391 0.11 3945 0.13 3.14 6-nonenoico 13386 1095536 9.60 282123 9.02 3.88 dodecanoico (laurico) 15733 15828 0.14 4198 0.13 3.77 tridecanoico 17207 12403 0.11 3281 0.10 3.78 12-metil-tridecanoico 17773 99502 0.87 26915 0.86 3.70 11-ottadecenoico 18105 1924066 16.86 483145 15.44 3.98 tetradecanoico 19646 35774 0.31 10013 0.32 3.57 12-metil-tetradecanoico 21618 8739 0.08 2758 0.09 3.17 14-meti-pentadecanoico 21945 86050 0.75 23403 0.75 3.68 7-esadecenoico 22442 3419150 29.95 868857 27.78 3.94 esadecanoico (palmitico) 23643 6033 0.05 2120 0.07 2.85 14-metil-esadecanoico 23811 11235 0.10 3279 0.10 3.43 pentacosanoico 23917 5769 0.05 1426 0.05 4.05 13-ottadecenoico 24371 15784 0.14 4088 0.13 3.86 15-metil-esadecanoico 25653 38178 0.33 10610 0.34 3.60 9,12-octadecadienoico 25797 716288 6.28 181129 5.79 3.95 10-ottadecenoico 25892 26965 0.24 10012 0.32 2.69 9-ottadecenoico 25960 41802 0.37 9018 0.29 4.64 15-ottadecenoico 26273 260707 2.28 68887 2.20 3.78 16-metileptadecanoico 29283 14461 0.13 3523 0.11 4.10 14-metil-butylesadecanoico
Tabella 7 - Acidi grassi riscontrati nel cromatogramma di Fig.19
Figura 19 - Cromatogramma acidi grassi, micorrizato.
Figura 20 - Cromatogramma acidi grassi, controllo

RT area area% altezza altezza% A/H Nome 6,559 683137 0,34 221929 0.86 3.08 ottanoico 9,278 64426 0,03 22448 0.09 2.87 4-decenoico 9411 1553333 0,77 466667 1.82 3.33 decanoico
13406 3258941 1,61 856050 3.33 3.81 dodecanoico 17781 302586 0,15 84006 0.33 3.60 11-dodecenoico 18182 24623813 12,19 4511886 17.58 5.46 tetradecanoico 19468 84504 0,04 24283 0.09 3.48 pentadecanoico 19648 207905 0,10 58033 0.23 3.58 12-metil-tetradecanoico 21621 75284 0,04 20053 0.08 3.75 14-metil-pentadecanoico 21965 706757 0,35 170528 0.66 4.14 11-ottadecenoico 22620 79530208 39,42 7109512 27.69 11.19 esadecanoico (palmitico) 23659 189341 0,09 53411 0.21 3.54 15-metil-esadecanoico 23829 285261 0,14 73685 0.29 3.87 14-metil-esadecanoico 23917 118533 0,06 36167 0.14 3.28 9-eptadecenoico 24384 321871 0,16 89119 0.35 3.61 eptadecanoico (margarico) 25694 1158137 0,57 219165 0.85 5.28 7,10-esadecadienoico 25952 57179894 28,31 6376272 24.84 8.97 10-ottadecenoico 26415 30803675 15,25 5092287 19.84 6.05 ottadecanoico (stearico) 26665 211415 0,10 55361 0.22 3.82 10,11- ottadecadienoico 28585 68662 0,03 20668 0.08 3.32 5,8,11,14-eicosatetraenoico (arachidonico) 29283 98797 0,05 24331 0.09 4.06 13-eicosenoico 29775 82058 0,04 24742 0.10 3.32 18-metilnonadecanoico
Tabella 8 - Acidi grassi riscontrati nel cromatogramma di Fig.20
3.3 Discussione dei risultati
Come si può osservare dal grafico n tot riportato qui di seguito, la cultivar di mais
micorrizzata presenta una percentuale di azoto totale significativamente superiore rispetto
alla cultivar di mais non trattata. In particolare la cultivar micorrizata ha un titolo medio
percentuale di azoto totale contenuto a livello fogliare pari al 9,9% in più rispetto alla
media delle piante coltivate in modo tradizionale.

Figura 21 - Istogramma relativo alla % di N totale nei campioni micorrizzati e di controllo
Sembrerebbe quindi che l’associazione simbiotica tra le radici delle piante di mais e i
funghi, porti ad un incremento della disponibilità di azoto, elemento minerale
fondamentale per la crescita delle piante. In effetti molti studi confermano questa ipotesi e
riconoscono che le piante con queste simbiosi, migliorino la loro nutrizione minerale,
registrando un effetto positivo sulla loro crescita e cedendo in cambio zuccheri al fungo.
In particolare, uno studio sulle simbiosi AM afferma la presenza di trasportatori attivi sulla
membrana plasmatica del fungo e rileva la presenza di un gradiente protonico nello spazio
periarbuscolare che separa la pianta dalla membrana plasmatica del fungo. Questo
gradiente protonico è generato dall’ enzima H+-ATPasi e partecipa al co-trasporto dei
carboidrati dall’apoplasto all’ interno delle cellule delle ife fungine. Il glucosio trasportato
all’interno delle cellule del fungo viene poi trasformato in trealosio o glicogeno; il glucosio
può essere direttamente utilizzato per la biosintesi dei lipidi o entrare nella via dei pentosi
fosfato; i lipidi ed il glicogeno sono poi trasferiti nel micelio extraradicale. Inoltre per
quanto concerne lo ione nitrato, le simbiosi AM giocano un ruolo attivo in quanto sono
capaci di liberare il nitrato complessato nella materia organica all’ interno del suolo,
cedendolo alla pianta.22
Inoltre grazie alla simbiosi micorriza, la pianta risulta essere più resistente a stress biotici o
abiotici, aumenta la tolleranza alla mancanza di acqua o alla presenza di inquinanti e porta
a una riduzione della sensibilità ai comuni agenti patogeni23 (Smith, Read 2008). In
particolare, le simbiosi AM aumentano la disponibilità di elementi minerali (es. fosforo e
azoto) per la pianta, controllano la qualità delle comunità vegetali aumentandone la

biodiversità e produttività e vengono perciò considerate dei biofertilizzatori naturali.
Sappiamo infatti che le piante micorrizate hanno una duplice via per procurarsi il prezioso
fosforo (fig. 2): da una parte lo assumono direttamente tramite i loro trasportatori presenti
nelle cellule epidermiche radicali, dall’altra l’assunzione è indiretta in quanto mediata dai
funghi simbionti che sono caratterizzati da trasportatori del fosfato ad alta efficienza
localizzati nel micelio extraradicale24 (Smith et al., 2011; Gomez Ariza et al., 2009).
Una recente scoperta (Kiers et al., 2011) dimostra che tra le piante e i loro funghi simbionti
si stabilisce un livello di cooperazione così raffinato che la pianta seleziona, tra quelli
presenti nella rizosfera, i funghi AM più efficienti e cooperativi nel cederle il prezioso
elemento25.
Figura 22 - Lo schema illustra come l’assunzione del P segua una duplice strada nelle piante portatrici di funghi micorrizici arbuscolari (AM). Da una parte la pianta esprime i suoi trasportatori di P (qui illustrati usando la terminologia per Solanum lycopersicum): LePT1 e LePT2 che sono espressi nelle cellule epidermiche. Nelle radici micorrizate il fungo attiva il suo trasportatore (GmosPT) che gli permette di assumere il P dal suolo e di convogliarlo sotto forma di granuli di polifosfato dentro la corteccia della radice dove il P è rilasciato alla pianta. Qui vengono attivati dei trasportatori vegetali la cui espressione è particolarmente innalzata nelle cellule che ospitano gli arbuscoli (LePT3, LePT4, LePT5). In altre piante, come Medicago truncatula, l’espressione del suo trasportatore MtPT4 è esclusiva delle cellule arbuscolate.
Inoltre alcuni studi di genomica hanno permesso di decifrare il funzionamento delle cellule
sito degli scambi tra pianta e fungo. Le moderne piattaforme tecnologiche permettono di
studiare l’espressione globale dei geni di un individuo (ad esempio di una pianta in
determinate condizioni). Attraverso l’uso di microarray si è potuto dimostrare come una
pianta di Lotus, una leguminosa modello, risponde alla presenza dei funghi AM regolando

nella radice l’espressione di più di 500 geni (Guether et al., 2009a) e che molti di questi
codificano per proteine coinvolte nel trasporto di P, N, ammonio e amminoacidi26 (Guether
et al., 2009a,b, Guether et al., 2011). Inoltre la possibilità di usare una tecnica assai
sofisticata (Laser microdissector) che permette di studiare popolazioni omogenee di cellule
dimostra come le cellule in cui il fungo forma arbuscoli (le strutture ramificate tipiche dei
funghi AM) sono probabilmente quelle che esprimono il maggior numero di geni di
trasporto, confermando l’ipotesi che le cellule arbuscolate siano la sede più importante
degli scambi tra i due partner (fig. n).
Figura 23 - Lo schema illustra come il fungo AM (in blu) colonizzi la radice della pianta ospite formando delle strutture ramificate (gli arbuscoli) nelle cellule più profonde della corteccia. Le analisi di trascrittomica condotte usando cellule arbuscolate (riquadro in alto) e confrontandole con simili cellule parenchimatiche non colonizzate (riquadro sotto) dimostrano che molti geni della pianta che codificano per trasportatori di ammonio, di fosfato, di aminoacidi, e di zolfo sono localizzati sulla membrana periarbuscolare che avvolge il fungo. Grazie a questi trasportatori, la cui funzionalità è stata dimostrata, la pianta prende gli elementi minerali dal fungo e li usa per il suo metabolismo.
Nel 2010 Tian et al. hanno approfondito la caratterizzazione genica e la relativa
espressione alla base del flusso metabolico dell’azoto nell’ ambito di simbiosi con
Glomeromycota. Attualmente il modello comune di assorbimento dell’azoto dal suolo nelle
micorrize arbuscolari è basato sulla sintesi di arginina nel micelio extraradicale e quindi il
suo trasferimento nel micelio intraradicale dove subisce biotrasformazione al fine di

rendere disponibile l’azoto per i diversi usi metabolici. L’analisi del meccanismo e la
regolazione del trasferimento di azoto dal fungo alla pianta, ove è risaputo attualmente che
11 geni sono coinvolti in questa attività grazie alla loro identificazione dal Glomus
intraradices, va diretta allo studio delle diverse isoforme enzimatiche presenti nelle
micorrize. A questo fine è stato sequenziato il cDNA dal micelio extraradicale e
successivamente è stata ottenuta la sequenza codificante per i geni degli enzimi coinvolti
nell’assorbimento dell’azoto e il suo relativo metabolismo nei funghi. La caratterizzazione
funzionale è stata eseguita utilizzando anticorpi specifici per la regione di poliistidina
inserzionata sulle sequenze codificanti espresse in Saccharomyces Cerevisiae. Come già
riportato in precedenza il viene trasferito dal suolo al micelio extraradicale attraverso
la nitrato transferasi (NT) e successivamente si riduce a nitrito (nitrato reduttasi-EC
1.7.1.1./2/3) ed ancora ad ammonio (nitrito reduttasi- EC 1.7.2.1.). Attraverso le due
isoforme dell’enzima glutammina sintetasi (GS1 e GS2) lo ione ammonio viene
incorporato nel glutammato per formare glutammina. Le due isoforme presentano una Km ,
rispettivamente di 1,87mM e 3,80mM; la Km o costante di Michaelis, indica la
concentrazione di substrato presente alla metà della velocità di reazione massima
catalizzata dall’enzima; questo sta ad indicare quindi che l’isoforma GS1 ha una cinetica
di reazione più rapida rispetto all’isoforma GS2, poiché a parità di velocità la GS1
converte una quantità maggiore di substrato cioè di glutammato. Successivamente, la
glutammina, attraverso l’azione sequenziale di tre enzimi rispettivamente, la carbamoil
fosfato sintasi (CPS), l’arginina succinato sintasi (ASS) e l’ arginina succinato liasi (AL), si
converte in arginina. In seguito l’arginina viene traslocata nel micelio intraradicale dove
subisce l’azione dell’ enzima arginasi (CAR1) che la scinde in urea e ornitina. L’ornitina
può subire l’azione di due enzimi: l’ornitina ammino trasferasi (OAT) che la converte in
glutammato o l’ornitina decarbossilasi che la trasforma in putrescina. L’urea invece,
attraverso l’enzima ureasi (URE) libera lo ione ammonio il quale viene trasportato dal
micelio intraradicale al citosol della cellula corticale rendendosi quindi disponibile per il
metabolismo della pianta.27 Le reazioni qui descritte sono schematizzate nella Figura ___.

Figura 24 - Vie metaboliche nella micorriza/ospite.

Per quanto concerne invece l’analisi della vitamina A e della vitamina E, come si può
osservare dal grafico n__, il contenuto vitaminico del latte crudo ottenuto da vacche
alimentate con insilato proveniente da coltivazioni micorrizate è nettamente superiore a
quello proveniente da vacche alimentate in modo tradizionale. Nell’istogramma che segue,
si evince che il titolo di Vitamina A e di Vitamina E del pool micorrizato eccede quello del
pool di controllo rispettivamente del 23,9% e del 12,4%.
Figura 25 - Risultanze sperimentali Vitamina A ed E.
La ragione di questa differenza va probabilmente ricercata partendo dalla biosintesi della
vitamina A. La vitamina A è una vitamina liposolubile e in natura si trova in diverse forme.
Con il termine di vitamina A vengono indicati sia il retinolo che i suoi analoghi,
detti retinoidi, di cui si conoscono almeno 1500 tipi diversi, tra naturali e sintetici. Il
precursore della vitamina A è il β-carotene il quale appartiene alla famiglia dei carotenoidi,
una classe di pigmenti organici che possono essere rinvenuti nelle piante o in altri
organismi fotosintetici, come le alghe ed alcune specie di batteri. Esistono oltre 600 tipi di
carotenoidi conosciuti; vengono normalmente suddivisi in due classi: i caroteni (che sono
idrocarburi quindi privi di ossigeno) e le xantofille (che invece lo contengono). Sono
pigmenti accessori che nella fotosintesi consentono di assorbire lunghezze d'onda differenti
rispetto alla clorofilla e proteggono quest'ultima dalla foto ossidazione. La nostra
trattazione riguarderà principalmente la sintesi del β-carotene in quanto come
precedentemente succitato è il precursore della vitamina A.

La biosintesi dei carotenoidi (figura), ovvero dei tetraterpeni fa parte della via biogenetica
del mevalonato, sino alla formazione dell’isopentenil pirofosfato (IPP) dal quale passando
per il geranilgeranil pirofosfato si giunge al fitene per poi arrivare al β-carotene.
Il primo passaggio della biosintesi dei carotenoidi implica la condensazione di due molecole di
GGPP ad opera della fitoene sintasi (PSY – EC2.5.1.32) che si compie in due tappe portando
prima alla formazione di prefitoene pirofosfato(PPPP) e quindi di fitoene. La reazione di
condensazione porta alla perdita dell’idrogeno e del gruppo pirofosforico in C-1’ della stessa
molecola. Quindi si ha il distacco del gruppo pirofosforico in C-1 del PPPP seguito da un
riaggiustamento C-1’ con la conseguente formazione del fitoene, che può risultare 15-cis
oppure tutto in trans. Successivamente una serie di quattro ossidazioni (ad opera di fitoene
desaturasi EC 1.3.99.30) portano alla formazione dei doppi legami coniugati convertendo il
fitoene (incolore) in fitofluene (incolore) in ς-carotene (giallo), in neurosporene (arancio) e
licopene (rosso). L’azione della licopene ciclasi (EC 5.5.1.18/19) porta alla formazione del
carotenoide biciclico β-carotene. Negli ultimi stadi della via biosintetica si formano le
xantofille.
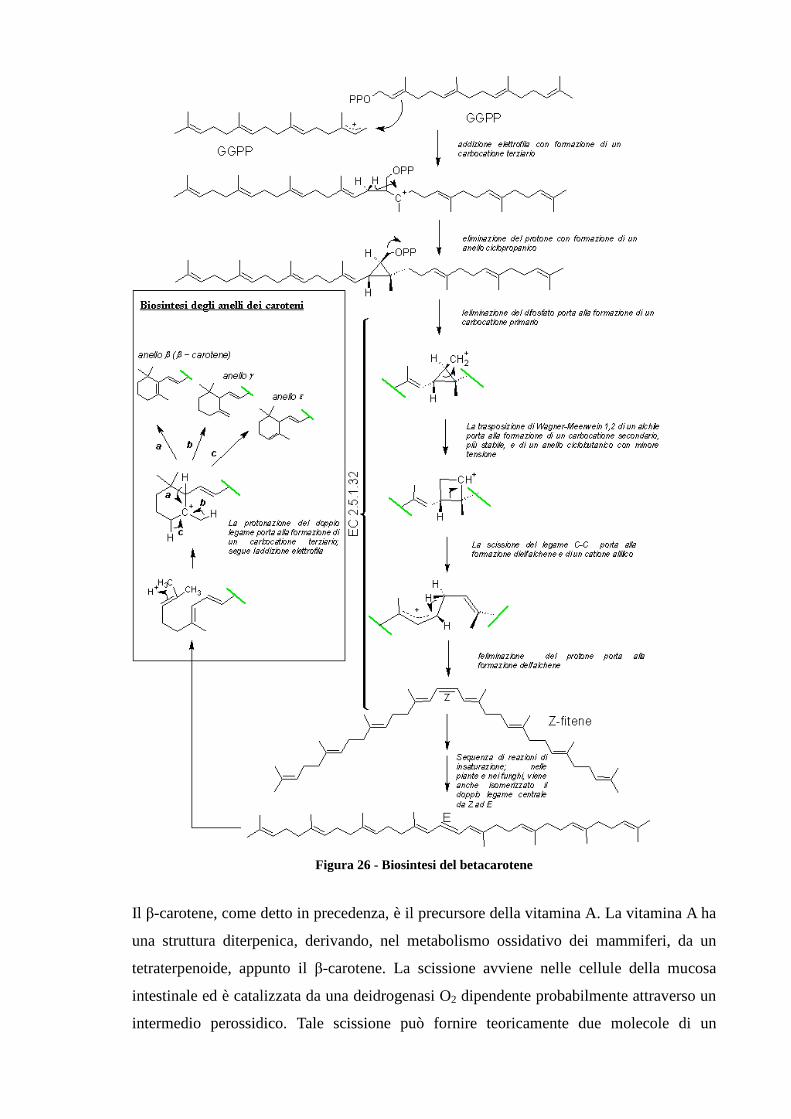
Figura 26 - Biosintesi del betacarotene
Il β-carotene, come detto in precedenza, è il precursore della vitamina A. La vitamina A ha
una struttura diterpenica, derivando, nel metabolismo ossidativo dei mammiferi, da un
tetraterpenoide, appunto il β-carotene. La scissione avviene nelle cellule della mucosa
intestinale ed è catalizzata da una deidrogenasi O2 dipendente probabilmente attraverso un
intermedio perossidico. Tale scissione può fornire teoricamente due molecole di un

intermedio aldeidico, il retinale, che viene successivamente ridotto ad alcool, il retinolo.
Sebbene il β-carotene scisso a livello del legame centrale può dare origine a due molecole
di retinolo, ci sono prove che la scissione può avvenire anche a livello di altri doppi legami,
e viene chiamata in questo caso, scissione acentrica. Un ulteriore accorciamento della
catena produce poi il retinale, ma viene prodotta una sola molecola per ogni molecola di β-
carotene.
Figura 27 - Biosintesi della Vitamina A
Parallelamente, per quanto concerne la vitamina E, è importante prendere in
considerazione la via biosintetica, al fine di correlare le rilevanze sperimentali con quanto
già conosciuto nell’ambito metabolico delle piante. La vitamina E fa parte della classe dei
tocoferoli. Questi ultimi sono spesso ritrovati nei cloroplasti e la loro biosintesi ha molte
caratteristiche in comune con la sintesi dei plastochinoni con una reazione aggiuntiva di
ciclizzazione che coinvolge l’anello idrochinonico e la catena laterale terpenoidica per dare
un anello cromano. Perciò i tocoferoli non sono in effetti chinoni, ma sono certo correlati ai
plastochinoni. La catena isoprenoidica aggiunta, derivante dal fitil difosfato contiene solo 4
unità isopreniche e tre dei doppi legami attesi hanno subito riduzione. Ancora una volta la
decarbossilazione dell’acido omogentisico è contemporanea alla reazione di alchilazione.
Gli stadi di C-metilazione con SAM (S-adenosilmetionina) e la ciclizzazione
dell’idrochinone a γ-tocoferolo sono stati stabiliti come mostrato nello schema biosintetico
sotto riportato. Si noti che uno dei metili sul nucleo aromatico deriva dall’omogentisato
mentre l’altro proviene dalla SAM.

Figura 28 - Biosintesi della Vitamina E
Le evidenze sperimentali ottenute dall’analisi quantitativa della vitamina A e della
vitamina E nel latte crudo di vacche alimentate con insilato di mais micorrizato, potrebbero
essere correlate ad una maggiore disponibilità di azoto presente nella cultivar micorizzata
rispetto alla cultivar non trattata. I risultati di questo studio hanno infatti molti riscontri in
letteratura in quanto alcuni studi evidenziano come la disponibilità di azoto determini una
variazione nel contenuto di pigmenti fotosintetici quali, la clorofilla a, la clorofilla b, la
luteina, il β- carotene, la neoxantina, la viola xantina, la zeaxantina e l’antera xantina. Uno
studio condotto sulla Phragmites autralis (comune canna di palude), evidenzia come il
contenuto di pigmenti foto sintetici dipenda dalla posizione delle foglie lungo il culmo.
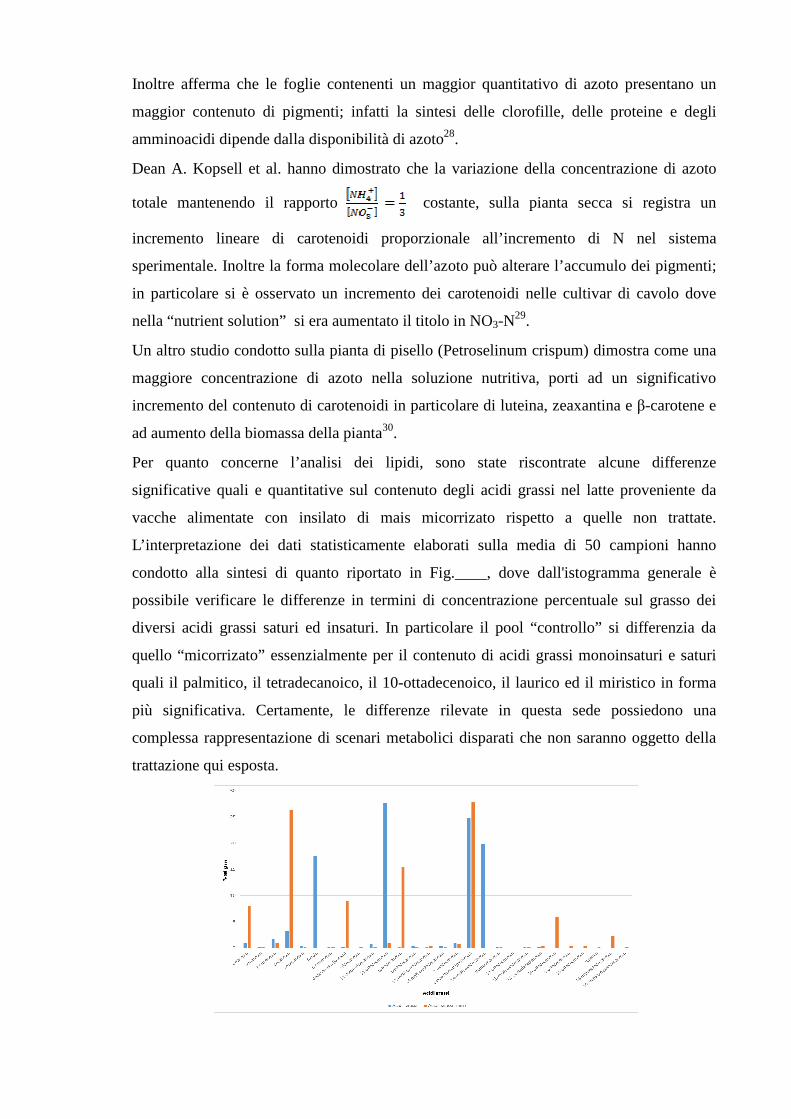
Inoltre afferma che le foglie contenenti un maggior quantitativo di azoto presentano un
maggior contenuto di pigmenti; infatti la sintesi delle clorofille, delle proteine e degli
amminoacidi dipende dalla disponibilità di azoto28.
Dean A. Kopsell et al. hanno dimostrato che la variazione della concentrazione di azoto
totale mantenendo il rapporto costante, sulla pianta secca si registra un
incremento lineare di carotenoidi proporzionale all’incremento di N nel sistema
sperimentale. Inoltre la forma molecolare dell’azoto può alterare l’accumulo dei pigmenti;
in particolare si è osservato un incremento dei carotenoidi nelle cultivar di cavolo dove
nella “nutrient solution” si era aumentato il titolo in NO3-N29.
Un altro studio condotto sulla pianta di pisello (Petroselinum crispum) dimostra come una
maggiore concentrazione di azoto nella soluzione nutritiva, porti ad un significativo
incremento del contenuto di carotenoidi in particolare di luteina, zeaxantina e β-carotene e
ad aumento della biomassa della pianta30.
Per quanto concerne l’analisi dei lipidi, sono state riscontrate alcune differenze
significative quali e quantitative sul contenuto degli acidi grassi nel latte proveniente da
vacche alimentate con insilato di mais micorrizato rispetto a quelle non trattate.
L’interpretazione dei dati statisticamente elaborati sulla media di 50 campioni hanno
condotto alla sintesi di quanto riportato in Fig.____, dove dall'istogramma generale è
possibile verificare le differenze in termini di concentrazione percentuale sul grasso dei
diversi acidi grassi saturi ed insaturi. In particolare il pool “controllo” si differenzia da
quello “micorrizato” essenzialmente per il contenuto di acidi grassi monoinsaturi e saturi
quali il palmitico, il tetradecanoico, il 10-ottadecenoico, il laurico ed il miristico in forma
più significativa. Certamente, le differenze rilevate in questa sede possiedono una
complessa rappresentazione di scenari metabolici disparati che non saranno oggetto della
trattazione qui esposta.

Figura 29 - Istogramma relativo al confronto tra frazionamento degli acidi grassi nei campioni micorrizati e di controllo
4. CONCLUSIONE
Le evidenze sperimentali ottenute dall’analisi dei campioni di foglie di mais micorrizate
rispetto a quelle non trattate hanno riscontrato un effettivo incremento del titolo di azoto
organico presente all’interno delle foglie della pianta di mais. Anche l’analisi
bromatologica del latte crudo proveniente da allevamenti di bovine alimentate con insilato
di mais micorrizato ha evidenziato un significativo aumento del contenuto di vitamina A e
di vitamina E e una differente distribuzione del contenuto degli acidi grassi, rispetto al latte
proveniente da bovine alimentate con mais non trattato. Queste differenze potrebbero
essere attribuite al ruolo sostanziale della simbiosi micorrizica tra le radici della pianta di
mais e il consorzio microbiologico costituito dal Micosat F. Infatti è stato dimostrato come
l’associazione simbiontica tra i due organismi permetta il loro ciclo vitale vivendo a stretto
contatto e traendo benefici reciproci sia di natura nutrizionale che di altro tipo. Molti studi
dimostrano come le fonti naturali di fosfato e nitrato stanno diminuendo in modo sensibile
mentre l’uso si fertilizzanti continua ad aumentare per mantenere alto il livello di
produttività. In questo contesto i funghi AM potrebbero giocare un ruolo fondamentale nel
promuovere un agricoltura sostenibile, basata fondamentalmente sulla limitazione dei
fertilizzanti, sul rispetto degli equilibri microbiologici e sulla conservazione della struttura
del suolo.
Per quanto concerne l’analisi sul contenuto vitaminico del latte crudo, svariati studi
sperimentali dimostrano come la vitamina E abbia un’azione antiperossidante proteggendo
quindi dalla perossidazione gli acidi grassi insaturi, la vitamina A e i caroteni. I fosfolipidi,
costituenti le membrane cellulari, sono ricchi di acidi grassi insaturi e questo spiega come
la deficienza di vitamina E si ripercuota sulla integrità strutturale e funzionale delle
membrane. Inoltre alcuni studi hanno dimostrato che nei ratti maschi, una deficienza di
vitamina E produce un alterazione degenerativa irreversibile dell’epitelio germinale e
quindi sterilità. Nei ratti femmine la avitaminosi E determina aborto spontaneo, dovuto
tuttavia ad alterazioni reversibili; la somministrazione di vitamina E ripristina infatti la
capacità di condurre a termine la gravidanza. Inoltre nei conigli, cavie e scimmie la
avitaminosi E produce una grave distrofia muscolare associata a creatinuria (comparsa
nelle urine di creatinina, una sostanza particolarmente presente nei muscoli, dove riveste
un'importante funzione di conservazione dell'energia). Quindi la presenza di un maggior
quantitativo di vitamina E nel latte crudo analizzato in questo studio, potrebbe avere un

effetto positivo anche sulle bovine e quindi potrebbe evitare l’insorgenza di fenomeni di
sterilità o problemi legati a disturbi dell’apparato muscolo-scheletrico. Per quanto concerne
invece l’analisi sul contenuto di grasso all’interno del latte crudo, si potrebbe supporre che
un aumento di grasso nel latte potrebbe portare a fenomeni di bioaccumulo e quindi
potrebbe scatenare possibili effetti negativi sulla salute dell’uomo.
In conclusione, le osservazioni sperimentali riassunte in questo lavoro costituiscono
l’analytical core dello studio interdisciplinare relativo alle ricadute in campo
agroalimentare conseguente all’utilizzo di Micosat F nelle colture di Zea Mays. Pertanto
sarà necessario effettuare ulteriori indagini circa gli aspetti ancora sconosciuti in campo
bromatologico ed agronomico.

APPENDICE
In questa appendice al lavoro sperimentale vengono riepilogati i principali dati di
validazione dei metodi utilizzati per la determinazione dei parametri seguenti: Azoto Totale
Kjeldahl, Vitamina A, Vitamina E ed acidi grassi. I parametri riportati sono l'incertezza di
misura associata al metodo, la ripetibilità, la riproducibilità, il recupero, la deviazione
standard, l'intervallo di fiducia ed il coefficiente di variazione associato alla media dei
valori ottenuti durante le procedure di validazione dei metodi di analisi chimica.
A.1. Determinazione dell'Azoto Totale Kjeldhal – ISO 1871:2009
Parametro Descrizione Valore
Xm Valore medio su dieci
determinazioni 0,50g/100g
U Incertezza estesa di misura
associata al metodo 2,50%
σ Deviazione standard sulla popolazione considerata
0,14
CV Coefficiente di variazione 0,68
IF Intervallo di fiducia ±0,08
LOD Limite minimo di determinazione
0,5g/100g
LOQ Limite minimo di quantificazione
0,5g/100g
R% Recupero percentuale 92,30%
sR Scarto Tipo di riproducibilità 0,14
sr Scarto Tipo di ripetibilità 0,23
r Coefficiente di Ripetibilità 4,6
TAB. A1
Le forma analitiche delle grandezze riportate nella Tab.A1 si riportano nella Tab.A2.
L’incertezza estesa di misura riferita al campione sottoposto ad analisi chimica si ricava
attraverso un approccio metrologico a partire dall’incertezza del metodo ricavata di seguito in
questo paragrafo. Bisognerà dapprima ottenere l’incertezza composta moltiplicando le singole

componenti dell’incertezza per il fattore di ripetibilità, ovvero ( )[ ] ( )
m
Nyuyu cnc && =
dove m è il
numero di prove ripetute per la determinazione del campione incognito, N il numero di
ripetizioni eseguite per il calcolo della ripetibilità del metodo. In tal senso si procede al calcolo
dell’incertezza estesa relativa alla misura y considerando che l’incertezza estesa definita come
( )[ ]eff
tyuU nce ν,95.0⋅= &, deve essere calcolata tenendo conto dei gradi di libertà del sistema,
stimabili con la formula di Welch-Satterthwaite ∑
=
i i
i
ceff
v
u
u4
4
&
&ν
. Il risultato sarà espresso
come eUy ± .
A.2. Determinazione della Vitamina A e della Vitamina E – M.I.M. 01032013/A
Parametro Descrizione Valore
Xm Valore medio su dieci
determinazioni 1,0ug/100g
U Incertezza estesa di misura
associata al metodo 2,80%
σ Deviazione standard sulla popolazione considerata
CV Coefficiente di variazione
IF Intervallo di fiducia
LOD Limite minimo di determinazione
0,20g/100g
LOQ Limite minimo di quantificazione
0,55g/100g
R% Recupero percentuale 90,15%
sR Scarto Tipo di riproducibilità
sr Scarto Tipo di ripetibilità
r Coefficiente di Ripetibilità
TAB. A2
Per quanto concerne i parametri sopra riportati la metodologia di calcolo è la medesima: media
su dieci misure; per la riproducibilità 4 repliche eseguite da uno stesso operatore e 10
ripetizioni.
L'incertezza di misura associata al singolo campione viene calcolata come riportato nel
paragrafo precedente.

Essendo quest'ultimo un metodo strumentale si riporta qui di seguito a titolo esemplificativo
l'approccio metrologico per il calcolo dell'incertezza estesa di misura associata al metodo. Calcolo della media:
Xm=1,0ug/100g
Calcolo della deviazione standard
( )097,0
11
2
=−
−=∑
=
n
xxs
n
ii
r
Calcolo della stima dell’incertezza
( ) 031.02
==n
sxu r
r
( ) ( ) 310126.106.5
031.0 −⋅===x
xuxu r
rc
1) Valutazione del contributo all’incertezza della bilancia
Incertezza estesa dello strumento (certificato LAT) Ue(bilancia) = 0.001
( ) ( ) 3
%1.92;2
100.1 −
==
⋅==lfk
e
k
bUbu
( ) ( ) 4100.110
−⋅==g
bubu gr
( ) 44 10414.1100.12 −− ⋅=⋅⋅=buc
2) Valutazione del contributo all’incertezza del cromatografo
Errore dello strumento = ± 0.255%, essendo gli estremi dell’incertezza verosimili, ricaviamo l’incertezza tipo del-
la componente relativa al cromatografo utilizzando una distribuzione rettangolare, ovvero
( ) 31047.11003
255.0 −⋅=⋅
=UFLCuc

3) Valutazione del contributo all’incertezza del materiale di riferimento
La preparazione delle soluzioni di lavoro di Vitamina A e Vitamina E (standard secondario) a
partire dalle soluzioni standard di Vitamina A e Vitamina E certificate ad una purezza del
99,9% comporta la presenza di un errore in ordine alle diverse diluizioni. Va pertanto
considerata l’incertezza relativa alle diluizioni effettuate con la pipetta di precisione, che
calcoliamo essere,
Labmate – Errore = ±0.15% mlpur
43
100.93
105.1)( −
−
⋅=⋅= e rapportando al volume si
ottiene ( ) 3
4
100.91.0
100.9 −−
⋅=⋅=ml
mlpuc
( ) ( ) ( ) ( ) ( ) 2232322 101.1100.9108.5 −−− ⋅=⋅+⋅=+= RupuRifu ccc
da cui deriva l’incertezza estesa, ( ) ( ) 11.0=⋅⋅= RifuxkRU ce
Lo standard di OTA (materiale di riferimento secondario) ha una incertezza estesa assoluta di
0.01µg/ml, per cui ripercorrendo il ragionamento già fatto in precedenza ricaviamo
l’incertezza della componente relativa al materiale di riferimento, ovvero
3103.1503
11.0)( −⋅=
⋅=RFuc
4) Valutazione del contributo all’incertezza delle pipette
Labmate 96/B Errore = + 0.15%
upip1 = 0.0015/√3 = 0.0009 ml
u°pip1= upip1/V1= 0.0009/1 ml = 0.0009
Labmate 53/B Errore = - 0.22%
Upip2 = 0.0022/√3 = 0.0013 ml
u°pip2= upip2/V2= 0.0012/0.2 ml = 0.006

5) Valutazione del contributo all’incertezza della retta di taratura
Per la valutazione del contributo relativo alla retta di taratura si fa riferimento alla comune
letteratura scientifica nell’ambito dell’analisi numerica e trattamento dei dati. La formula
utilizzata per il calcolo dell’errore di interpolazione ai minimi quadrati è la seguente
( ) ( )( )[ ]( )
−
−−−−
−= ∑
∑∑
2
2
2/ 2
1
xx
yyxxyy
Nxyε
Dove
N = numero di dati utilizzati per la costruzione della retta, mentre y è la variabile dipendente
ed x quella indipendente.
( )nnx
taru xyc
∑⋅
=2
/1 ε
Dove n = valori di incertezza di taratura star, ottenendo in tal modo
( ) 4103.7 −⋅=taruc
5) Valutazione del contributo dell’accuratezza all’incertezza
Il contributo dell’accuratezza all’incertezza del metodo viene calcolata considerando il range di
accettabilità di ± 10%. In particolar modo si considera un Bias (scostamento sistematico) pari
al 10% del valore nominale di OTA presente nel materiale di riferimento secondario (tenore in
OTA 50µg/ml certificato d’analisi sigma-aldrich, effettuando prove su campioni fortificati).
( ) 3101.7250
5.0 −⋅=⋅
=⋅
=nx
BiasBiasuc
Calcolo dell’incertezza combinata del metodo
(I contributi da utilizzare sono ricavati al termine di ogni sottoparagrafo)
( ) ( ) ( ) ( ) ( ) ( ) ( ) 32222222 108.921)( −⋅=++++++= biasutarupupuRFuUFLCubuOTAu cccccccc

Calcolo dell’incertezza estesa associata al metodo (approccio olistico)
( ) ( ) mlgOTAukOTAU ce /02.0 µ=⋅=

BIBLIOGRAFIA
1 Renzo Angelini (2007)
Il mais
Bayer CropScienze
2 FAO/UNIDO/Word Bank Working Group on Fertilizers. (1996) Current World Fertilizer Situation and
Outlook 1994/95-2000/2001. Food and Agricolture Organization of the United Nations, Rome.
3 (Schle-Singer 1997): 4 FAOSTAT (2001) Agricolture Data. Food and Agricolture Organization of the United Nations, Rome. 5 istituto italiano edizioni atlas 6 Crawford, N. M., and Forde, B. J. (2002)
Molecular and developmental biology of inorganic nitrogen nutrition.
In: The Arabidopsis Book, Someville, C. and Meyerowitz,E., eds. American Society of Plant
Physiologists, Rockville, MD, pp. Doi/10.1199/tab.0011.
7 Oaks, A. (1994)
Primary nitrogen assimilation in higher plants and its regulation
Can. J. Bot. 72: 739-750.
8 Warner, R. L., and Kleinhofs, A. (1992)
Genetics and molecular biology of nitrate metabolism in higher plant.
Physiol. Plant. 85: 245-252.
9 Wilbur H. Campbel. Nitrate reductasi structure, function and regulation: Bridging and Gap between
Biochemistry and Physiology. Plant Physiol. Plant Mol. Biol. 1999. 50:277-303
10 Mendel, R.R. (2005)
Molybdenum: Biological activity and metabolism.
Dalton Transanction 2005: 3404-3409.
11 Muhammad Aslam, Ann Oaks and Ray C. Huffaker
Effect of light and glucose on the induction of nitrate reductase and the distribuction of nitrate in etiolated
barley leaves.

Plant Physiol (1976) 58, 588-591.
12 13 Siegel L. M., and Wilkerson, J. Q. (1989)
Structure and function of spinach ferredoxin-nitrite reductase.
In: Molecular and Genetics Aspects of Nitrate Assimilation, Oxford, pp 263-283.
14 Smart, D. R., and Bloom, A. J. (2001)
Wheat leaves emit nitrous oxide during nitrate assimilation.
PNAS 98: 7875-7878.
15 WRAY, J. L. – 1993 - Molecular biology, genetic and regulation of nitrite reductasi in higher plants.
Physiol. Plant. 89: 607-612.
16 MARSCHNER, H. – 1995- Mineral Nutrition of Higher Plants. 2nd ed. Accademic Press, London.
17 LAM, H. M., COSCHIGANO, K. T., OLIVEIRA, I. C., MELO OLIVEIRA, R., and CORUZZI, G. M. -
1996 - The molecular genetics of nitrogen assimilation into amino acidsin higher plants. Annu. Rev. Plant
Physiology Plant Mol. Biol. 47: 569-593.
18 PETER ATKINS -1995- Chimica inorganica. Zanichelli 19 BURRIS, R.H. – 1976 – Nitrogen fixation. Plant Biochemistry 3rd ed pp. 887-908 20 BETTINA HAUSE, THOMAS FESTER – 2005 – Molecular and cell biology of arbuscular mycorrhizal symbiosis. Planta 221:184-196. 21 SMOOG, LEARY – 1995- Chimica analitica strumentale. Edises 22 BETTINA HAUSE, THOMAS FESTER – 2005 – Molecular and cell biology of arbuscular mycorrhizal symbiosis. Planta 221:184-196. 23 SMITH S.E., READ D.J. - 2008 - Mycorrhizal Symbiosis. 3rd ed., Academic Press, San Diego, CA, USA. http://www.amazon.com/Mycorrhizal-Symbiosis-Third-Sally-Smith/product-reviews/ 0123705266/ref=sr_1_1_cm_cr_acr_img?ie=UTF8&showViewpoints=1 24 SMITH S.E., JAKOBSEN I., GROENLUND M., SMITH F.A. - 2011 - Roles of arbuscular mycorrhizas in plant phosphorus nutrition: interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phos- phorus acquisition. Plant Physiology. 156, 1050-1057. GÓMEZ-ARIZA J., BALESTRINI R., NOVERO M., BONFANTE P. - 2009 - Cell-specific gene expression of phosphate transporters in mycorrhizal tomato roots. Biology and Fertility of Soils, 45, 845-853. 25 KIERS E.T., DUHAMEL M., BEESETTY Y., MENSAH J.A., FRANKEN O., VERBRUGGEN E., FELLBAUM C.R., KOWALCHUK G.A., HART M.M., BAGO A. , PALMER T. M., WEST S. A., VANDENKOORNHUYSE P., JANSA J., BÜCKING H. - 2011 - Reciprocal Rewards Stabilize Cooperation in the Mycorrhizal Symbiosis. Science, 333, 880-882. 26 GUETHER M., NEUHÄUSER B., BALESTRINI R., DYNOWSKI M., LUDEWIG U., BONFANTE P. - 2009 - A mycorrhizal-specific ammonium transporter from Lotus japonicus acquires nitrogen released by arbuscular mycorrhizal fungi. Plant Physiology, 150, 73-83. GUETHER M., VOLPE V., BALESTRINI R., REQUENA N., WIPF D., BONFANTE P. - 2011 - LjLHT1.2 - a mycorrhiza-inducible plant amino acid transporter from Lotus japonicus. Biology and Fertility of Soils, (in press). ISSN:0178-2762 IF:2.156 27 CHUNJIE TIAN, BETH KASIBORSKI, RAMAN KOUL, PETER J.LAMMERS, HEIKE BUCKING, YAIR SHACHAR-HILL - 2010 – Regulation of the Nitrogen Transfer Pathway in the Arbuscular Mycorrhizal Symbiosis: Gene Characterization and the Coordination of Expression with Nitrogen Flux. Plant Physiology Vol.153 pp 1175-1187. 28 INDRA LIPPERT, HARDY ROLLETSCHEK, JOHANNES-GUNTER KOHL - 2001 – Photosynthetic pigments and efficiencies of two Phragmites australis stands in different nitrogen availabilities. Aquatic

botany 69 359-365. 29 DEAN A. KOPSELL, DAVID E KOPSELL AND JOANNE CURRAN-CELENTANO -2007- Carotenoid pigments in Kale are influenced by nitrogen concentration and form. Journal of the Scienze of Food and Agriculture 87: 900-907 30 CHENARD CH, KOPSELL DA and KOPSELL DE – 2005 – Nitrogen concentration affects nutrient and carotenoid accumulation in parsley. J. Plant Nutr 28: 285-297.