universidade federal do rio de janeirolivros01.livrosgratis.com.br/cp093450.pdf · (alcian-blue...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
LEONARDO CAMPOS MONÇÃO RIBEIRO
O PAPEL DOS RECEPTORES P2X7 EM MODELO MURINO DE LESÃO PULMONAR AGUDA
Rio de Janeiro 2008

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

2
LEONARDO CAMPOS MONÇÃO RIBEIRO
O PAPEL DOS RECEPTORES P2X7 EM MODELO MURINO DE LESÃO PULMONAR AGUDA
Dissertação de Mestrado submetida ao Programa de Pós-Graduação em Ciências Morfológicas visando a obtenção do grau de Mestre em Ciências Morfológicas
Orientador: Prof Dra Christina Maeda Takiya
(ICB/UFRJ) Co-orientador: Prof. Dr. Robson Coutinho-Silva
(IBCCF-UFRJ)
Rio de Janeiro 2008

3
LEONARDO CAMPOS MONÇÃO RIBEIRO O papel dos receptores P2X7 em modelo murino de lesão pulmonar aguda
Dissertação de Mestrado submetida ao Programa de Pós-graduação em Ciências Morfológicas visando a obtenção do grau de Mestre em Ciências
Morfológicas
Orientador: Prof Dra Christina Maeda Takiya Prof. Associado do Instituto de Ciências Biomédicas
(ICB/UFRJ)
Co-orientador: Prof. Dr. Robson Coutinho-Silva Prof. Associado do Instituto de Biofísica Carlos Chagas Filho
(IBCCF-UFRJ)

4
Monção-Ribeiro, Leonardo Campos O papel dos receptores P2X7 em modelo murino de lesão pulmonar aguda Rio de Janeiro: UFRJ/CCS/ICB/Instituto de Ciências Biomédicas,2008 p.130 Dissertação de Mestrado em Ciências Morfológicas 1. Receptores P2X7 2. ALI/ARDS 3. inflamação

5
Este trabalho foi realizado nos laboratórios de Patologia Celular, no
Instituto de Ciências Biomédicas e nos laboratórios de Imunofisiologia e
Fisiologia pulmonar no Instituto de Biofísica Carlos Chagas Filho com os
auxílios financeiros concedidos pela FAPERJ, CNPq e PRONEX

6
AGRADECIMENTOS
A Deus.
À minha família. Ao meu filho que esta com 10 semanas na barriga da minha esposa
Flávia Aline, meu Amor.
À prof. Christina Maeda Takiya e ao prof. Robson Coutinho-Silva, meus orientadores e
responsáveis pela minha paixão que é a pesquisa.
A todos que direta ou indiretamente participaram desta dissertação ou da minha vida.

7
RESUMO Tem sido verificada a importância da sinalização purinérgica na imunidade inata e em
processos patológicos. Neste trabalho verificamos o papel dos receptores P2X7 em modelo
murino de lesão pulmonar aguda. Camundongos C57BL/6 selvagens e nocautes para o
receptor P2X7 foram instilados com lipopolissacarídeo - LPS (E. coli-60mg/50µL) ou salina
(50 µL), constituindo os grupos (LPS-KO, LPS-WT, CTRL-KO e CTRL-WT). Os animais
foram sacrificados 24 horas após a instilação sendo realizados estudos histológicos,
imunohistoquimicos e histomorfométricos. A partir do fluido do lavado bronco-alveolar
(BALF) foram dosados óxido nítrico, IL-β e lactato dehidrogenase (LDH) além da
realização de testes de função pulmonar (elastância estática e dinâmica, pressões resistivas
e viscoelásticas e o deltaP total) obtidos pelo método de oclusão ao final da inspiração. A
expressão dos receptores P2X7 foi aumentada no grupo LPS-WT em comparação com os
animais WT (p<0,05). Houve atenuação da lesão pulmonar nos animais LPS-KO com
diminuição significativa da inflamação pulmonar (células mononucleares e
polimorfonucleares), ausência de mastócitos e decréscimo de áreas de colapso pulmonar
(p<0,05), assim como na quantidade de NO, IL1β e LDH secretados no BALF nos animais
LPS-KO em comparação com animais KO (p>0,05). Apesar de não haver modificação do
número de macrófagos (F4/80+) no grupo LPS-KO em comparação com animais CTRL-
KO, o número de macrófagos ativados (BSL+) foi semelhante ao dos animais CTRL-KO.
As tramas de fibras colágenas, elásticas e fibronectina depositada nos animais LPS-KO são
quantitativamente semelhantes as do grupo CTRL-KO (p>0,05). A imunoexpressão de
TGF-β e MMP-9 aumentou em todos os grupos LPS quando comparados aos seus
controles, mas o aumento visto no grupo LPS-KO foi menor que o do grupo LPS-WT

8
(p<0,001). MMP-2 por outro lado, apesar de ter aumentado significativamente nos animais
LPS-WT, não foi modulado no animal LPS-KO quando comparado com o CTRL-KO. Os
animais LPS-KO não apresentaram alterações na função pulmonar (p>0.05). Em conclusão
podemos dizer que o receptor P2X7 participa na lesão pulmonar aguda induzida pelo LPS
atuando no recrutamento de neutrófilos e mastócitos, na ativação macrofágica e no
remodelamento da matriz extracelular.
Palavras-chaves: lesão pulmonar aguda; sinalização purinérgica, remodelamento pulmonar,
função pulmonar.

9
ABSTRACT The importance of the purinergic signalling in the mantainance of the innate immunity and
in pathological process has been object of extensive research.
We have investigated the roles of P2X7R activation in mice model of acute lung injury
(ALI). C57BL/6 wild-type and P2X7R knockout mice were intratracheally instillated with
LPS (n=6) (E. coli-60mg/50µL- LPS-KO e LPS-WT) or saline (n=5) (50 µL, C-KO e C-
WT) (LPS-KO, LPS-WT, CTRL-KO e CTRL-WT). Lung histology (amount of elastic,
collagen fibers, fibronectin and mast cells in the alveolar septa), the expression of P2X7R,
F4/80, BSL-1 lectin TGF-β and MMPs 2 and 9, mechanical parameters (static and dynamic
elastances, resistive and viscoelastic/inhomogeneous pressures, and deltaPtot obtained by
end-inflation occlusion method) and biochemical analysis of lactate dehydrogenase (LDH),
nitric oxide (NO) and IL-1β secretion in bronchoalveolar lavage fluid (BALF) were
analyzed after 24 hours. Histomorphometric analysis of lung parenchyma demonstrated
diminished polymorphonuclear and mononuclear cells (p<0.05), absence of mast cells
(alcian-blue pH2.5) and decrease in alveolar collapse (p<0.05) in LPS-KO group . The
LPS-KO and C-KO animals did not change the amount of collagen and elastic fibers,
fibronection immunoexpression as well as secretion of LDH, NO and IL-1β in BALF
(p>0.05). The expression of P2X7R was increased in LPS-WT group when compared with
WT animals (p<0.05). Macrophages (F4/80+) did not change after LPS treatment in LPS
(WT and KO) groups (p>0.05), however the macrophage activation (BSL-1+ lectin) was
abolished in LPS-KO group (p<0.05). The expression of TGF-β was increased in all LPS
groups compared with controls, but the increase showed by LPS-KO group was smaller
(p<0,001). The animals LPS-KO did not have altered pulmonary functions (p>0.05).Thus

10
the P2X7R participates in ALI on the recruitment of neutrophils and mast cells, in
macrophage activation and in extracellular matrix remodeling.
Keywords: acute lung injury, purinergic signaling, pulmonary remodeling, pulmonary
function.

11
LISTA DE ILUSTRAÇÕES
FIGURA 1. Esquema mostrando sistema respiratório. .................................................. 23
FIGURA 2. Esquema mostrando o receptor P2X7 e suas interações com componentes
celulares ............................................................................................................................... 32
FIGURA 3. Esquema mostrando protocolo experimental. ............................................ 41
FIGURA 4. Montagem experimental da obtenção e coleta de dados consistindo de ... 54
FIGURA 5. Representação esquemática dos traçados de fluxo, volume, pressão em
função do tempo, obtidos a partir da oclusão da via aérea ao final da inspiração....... 55
FIGURA 6. Detecção e Quantificação por imunohistoquímica (densidade de
superfície) dos receptores P2X7 . ...................................................................................... 58
FIGURA 7. Arquitetura pulmonar dos animais estudados evidenciados pela técnica de
hematoxilina e eosina. ........................................................................................................ 61
TABELA 1. Parâmetros Morfométricos e Celularidade no Parênquima Pulmonar... 63
FIGURA 8. Quantificação de nitrito no BAL de camundongos selvagens e nocautes
com e sem tratamento com LPS por 24 horas ................................................................. 64
FIGURA 9. Quantificação da presença da enzima lactato desidrogenase (LDH) no
BAL de camundongos selvagens e nocautes com e sem tratamento com LPS por 24
horas..................................................................................................................................... 65
FIGURA 11. Detecção imunohistoquímica e quantificação da População de
Macrófagos F4/80. .............................................................................................................. 68
FIGURA 12. Caracterização e quantificação dos macrófagos pulmonares ativados
pela deteção da isolectina Griffonia simplicifora. ........................................................... 70

12
FIGURA 13. Caracterização e quantificação dos mastócitos presentes no parênquima
pulmonar pela técnica histoquímica para alcian blue pH 2.5. ...................................... 73
FIGURA 14. Fibras colágenas no parênquima pulmonar evidenciadas pela coloração
.............................................................................................................................................. 76
de picrosirius. ...................................................................................................................... 76
FIGURA 15. Fibras elásticas no parênquima pulmonar. Evidenciadas pela coloração
Orceína. ............................................................................................................................... 79
FIGURA 16. Densidade de imunoexpressão da fibronectina no parênquima pulmonar.
.............................................................................................................................................. 81
FIGURA 17. Quantificação das metaloproteinases 2 e 9 por imunohistoquímica e da
citocina TGF-β por densidade de superfície. ................................................................... 83
FIGURA 18. Avaliação Funcional do Pulmão. Gráficos de elastância estática (A)
dinâmica do pulmão (B) de pressão resistiva (C) e de pressão viscoelástica (D) e de
variação de pressão total do pulmão (E). ......................................................................... 85
FIGURA 19. Esquema mostrando resumo dos resultados obtidos na dissertação .... 100

13
LISTA DE TABELAS
TABELA 1. Parâmetros Morfométricos e Celularidade no Parênquima Pulmonar... 63

14
SUMÁRIO
AGRADECIMENTOS........................................................................................................... 6
RESUMO ............................................................................................................................... 7
ABSTRACT ........................................................................................................................... 9
LISTA DE ILUSTRAÇÕES ................................................................................................ 11
1 INTRODUÇÃO................................................................................................................. 18
2 REVISÃO BIBLIOGRÁFICA......................................................................................... 21
2.1 PULMÃO .................................................................................................................. 21
2.1.1 LESÃO E REPARO PULMONAR..................................................................... 24
2.2 LESÃO PULMONAR AGUDA (ALI) E O MODELO DO
LIPOPOLISSACARÍDEO BACTERIANO (LPS) ...................................................... 27
2.3 RECEPTORES P2X7.................................................................................................. 30
2.3.1 PULMÃO E RECEPTORES P2X7 ..................................................................... 32
2.3.2 INFLAMAÇÃO E RECEPTORES P2X7 ........................................................... 33
2.3.3 LPS E RECEPTORES P2X7 ............................................................................... 35
3 OBJETIVOS..................................................................................................................... 38
3.1 OBJETIVO GERAL................................................................................................... 38
3.2 OBJETIVOS ESPECIFICOS ..................................................................................... 38
4 MATERIAIS E MÉTODOS............................................................................................. 40
4.1 INSTILAÇÃO DO LPS ........................................................................................... 40
4.2 PROCEDIMENTO CIRÚRGICO.............................................................................. 40
4.3 PROTOCOLO EXPERIMENTAL ............................................................................ 41
4.4 LAVADO BRONCOALVEOLAR (BAL) ................................................................ 41

15
4.5 PROCESSAMENTO HISTOLÓGICO...................................................................... 42
4.6 IMUNOHISTOQUÍMICA E HISTOQUÍMICA ...................................................... 44
4.6.1 ANTICORPOS UTILIZADOS .......................................................................... 44
4.6.2 IMUNOHISTOQUÍMICA .................................................................................. 45
4.6.3 HISTOQUÍMICA PARA A DETECÇÃO E QUANTIFICAÇÃO DOS
MACRÓFAGOS PULMONARES ATRAVÉS DA MARCAÇÃO DE RADICAIS
CARBOIDRÁTICOS ................................................................................................... 47
4.7 HISTOMORFOMETRIA........................................................................................... 48
4.8 ANÁLISE ESTATÍSTICA........................................................................................ 49
4.9 MECÂNICA RESPIRATÓRIA ................................................................................. 49
5 RESULTADOS ................................................................................................................. 57
5.1 TRATAMENTO COM LPS AUMENTOU SIGNIFICATIVAMENTE
EXPRESSÃO DOS RECEPTORES P2X7 NO PARÊNQUIMA PULMONAR............. 57
5.2 ANIMAIS P2X7-/-
APRESENTARAM INFLAMAÇÃO ATENUADA E MENOS
ZONAS DE COLAPSO APÓS O TRATAMENTO COM LPS...................................... 60
5.3 AUSÊNCIA DO RECEPTOR P2X7 INIBIU A SECREÇÃO DO NITRITO (NO),
DA ENZIMA LACTATO DESIDROGENASE (LDH) E DA CITOCINA IL-1β NO
BAL DE ANIMAIS NOCAUTESPARA O RECEPTOR P2X7 APÓS O
TRATAMENTO COM LPS............................................................................................. 64
5.4 INIBIÇÃO DA ESTIMULAÇÃO DE MACRÓFAGOS, MAS NÃO DA
POPULAÇÃO DE MACRÓFAGOS F4/80+ EM ANIMAIS P2X7-/-
APÓS O
TRATAMENTO COM LPS............................................................................................. 66
SE ALTERARAM NOS ANIMAIS P2X7-/-
APÓS O TRATAMENTO COM LPS....... 75

16
5.7 OS ANIMAIS P2X7-/-
APÓS O TRATAMENTO COM LPS APRESENTARAM
DIMINUIÇÃO DA IMUNOREATIVIDADE PARA TGF-β, AUMENTO DA
IMUNOREATIVIDADE PARA MMP- 9 E AUSÊNCIA DE MODIFICAÇÃO DA
QUANTIDADE DE IMUNOREATIVIDADE PARA MMP-2 ..................................... 81
6 DISCUSSÃO..................................................................................................................... 87
7 CONCLUSÕES................................................................................................................. 99
8 REFERÊNCIAS .............................................................................................................. 102

17
INTRODUÇÃO

18
1 INTRODUÇÃO
Moléculas sinalizadoras são essenciais para o início e manutenção das reações
inflamatórias. Recentemente, nucleotídeos extracelulares como ATP e UTP, têm sido
considerados como uma nova classe de moléculas sinalizadoras que desempenham um
papel na inflamação (DI VIRGILIO E COLS., 2001; BOURS E COLS., 2006).
Nucleotídeos são liberados nos sítios de inflamação como resultado do dano celular,
secretados por células apresentadoras de antígeno, macrófagos e linfócitos (LA SALA E
COLS., 2003).
Virtualmente, todos os tipos celulares expressam receptores de superfície para
nucleotídeos sinalizadores (BURNSTOCK & KNIGHT 2004). Dentre os receptores para
nucleotídeos, o grupo dos receptores P2 compreende os receptores P2Y acoplados a
proteínas-G (P2YR) e os receptores P2X (P2XR), os quais são canais iônicos intrínsecos
(BURNSTOCK & KNIGHT 2004; DI VIRGILIO E COLS., 2001).
Receptores P2X são ativados por ATP extracelular e já foram clonados 7 sub-tipos
de receptores desta família P2X1–7 (BURNSTOCK & KNIGHT 2004).
O ATP extracelular pode ser secretado por células infectadas e submetidas a estresse
(FERRARI E COLS., 1997; WANG E COLS., 2004; BURNSTOCK 2006). O ATP quando
age através do receptor P2X7 pode levar a estimulação da secreção de citocinas pró-
inflamatórias, tais como IL-1β (GALLUCCI & MATZINGER 2001; BOURS E COLS.,
2006). Em diferentes tipos celulares, os receptores P2X7 levam a produção e secreção de
mediadores inflamatórios, tais como: IL-1β, IL6, TNF, IL18, monocyte chemoattractant
protein-1 (MCP-1) (BOURS E COLS., 2006; FERRARI E COLS., 2006), clivagem de
metaloproteases, CD23, CD27 e selectina-L (GU E COLS., 1998; 2006).

19
A Síndrome do Estresse Respiratório Agudo (ARDS) foi primeiramente descrita em
1967 por Ashbaugh e colaboradores. Ela continua sendo até os dias de hoje uma importante
causa de morbidade e mortalidade, sendo caracterizada por um processo inflamatório
pulmonar agudo com apoptose de células epiteliais e intersticiais e edema intra-alveolar,
seguida pela fibrose (ATABAI & MATTHAY, 2002). O lipopolissacarídeo (LPS) é um
componente da parede celular de bactérias, sendo, portanto, um estímulo para a iniciação
local de inflamação aguda. A instilação intratraqueal de LPS em animais tem sido
considerada como um modelo experimental de ARDS (BRIGHAM & MEYRICK., 1986),
servindo como um modelo de doença pulmonar obstrutiva crônica (COPD) (BRASS E
COL.S, 2003; GEORGE E COLS., 2001).
Neste trabalho, estamos interessados particularmente em entender como os
receptores P2X7 estão envolvidos com a patogenia da ARDS, visto que desempenham
funções em diversos processos biológicos, como apoptose, desenvolvimento da imunidade
e controle da infecção microbiana (COUTINHO-SILVA E COLS., 2001; DI VIRGILIO,
1995; DI VIRGILIO E COLS., 2001; CHEN & BROSNAN, 2006; LISTER E COLS.,
2007; FRANCHI E COLS., 2007).

20
REVISÃO BIBLIOGRÁFICA

21
2 REVISÃO BIBLIOGRÁFICA
2.1 PULMÃO
O sistema respiratório, constituído pelos pulmões e por uma seqüência de vias
aéreas, funcionam fornecendo oxigênio para as células do corpo e eliminando dióxido de
carbono das células do corpo. O sistema respiratório está dividido em duas porções: a
porção condutora e a porção respiratória. A porção condutora situada dentro e fora dos
pulmões, transporta ar do meio externo para os pulmões. A porção respiratória, localizada
estritamente dentro dos pulmões (GARTNER & HIATT 2007), funciona na troca efetiva de
oxigênio por dióxido de carbono.
A árvore brônquica começa na bifurcação da traquéia, com a formação dos
brônquios primários, direito e esquerdo, que se arborizam (formando ramos que diminuem
gradualmente de tamanho). Ela é constituída por vias aéreas localizadas fora dos pulmões
(brônquios primários, brônquios extrapulmonares e por vias localizadas dentro dos
pulmões: os brônquios intrapulmonares (secundários e terciários), bronquíolos, bronquíolos
terminais e bronquíolos respiratórios. Cada brônquio intrapulmonar constitui a via aérea
que supre um lobo do pulmão. Estas vias aéreas são semelhantes aos brônquios primários,
com exceção de que os anéis cartilaginosos em C são substituídos por placas irregulares de
cartilagem hialina, que envolvem completamente a luz dos brônquios intrapulmonares.
Os brônquios secundários são os ramos diretos dos brônquios primários que se
dirigem para os lobos pulmonares. Em humanos, o pulmão esquerdo tem dois lobos e,
portanto, tem dois brônquios secundários; o pulmão direito tem três lobos e, portanto, tem

22
três brônquios secundários. Ao entrarem nos lobos pulmonares, os brônquios secundários
subdividem-se em ramos menores, os brônquios terciários. Cada brônquio terciário se
arboriza, e vai para uma secção definida de tecido pulmonar denominada segmento
broncopulmonar. Cada pulmão tem dez segmentos broncopulmonares, que estão
completamente separados um do outro por elementos de tecido conjuntivo (GARTNER &
HIATT 2007).
Os bronquíolos não possuem cartilagem em sua parede, têm menos de 1mm de
diâmetro e possuem células de clara em seu revestimento epitelial. Cada bronquíolo fornece
ar para o lóbulo pulmonar. O revestimento epitelial dos bronquíolos varia de colunar
simples ciliado, com células caliciformes ocasionais, nos bronquíolos maiores, a células
cubóides simples, células de clara ocasionais e ausência de células caliciformes, nos
bronquíolos menores (GARTNER & HIATT 2007). Os bronquíolos terminais formam a
região menor e mais distal da porção condutora do sistema respiratório. Estas estruturas
trazem ar para os ácinos pulmonares, subdivisões do lobo pulmonar. Os bronquíolos
terminais se ramificam, dando origem aos bronquíolos respiratórios.
A porção respiratória do sistema respiratório é composta por bronquíolos
respiratórios, dutos alveolares, sacos alveolares e alvéolos. Os bronquíolos respiratórios são
a primeira região do sistema respiratório em que pode ocorrer a troca de gases, sua estrutura
é semelhante à dos bronquíolos terminais, mas sua parede está interrompida pela presença
de estrutura de paredes delgadas, semelhantes a bolsas, denominadas alvéolos, nos quais
pode haver trocar gasosas. Ao se ramificarem, o diâmetro dos bronquíolos respiratórios
torna-se mais estreito e aumenta sua população de alvéolos. Após ramificar-se várias vezes,
cada bronquíolo respiratório termina em um duto alveolar (FIGURA 1).
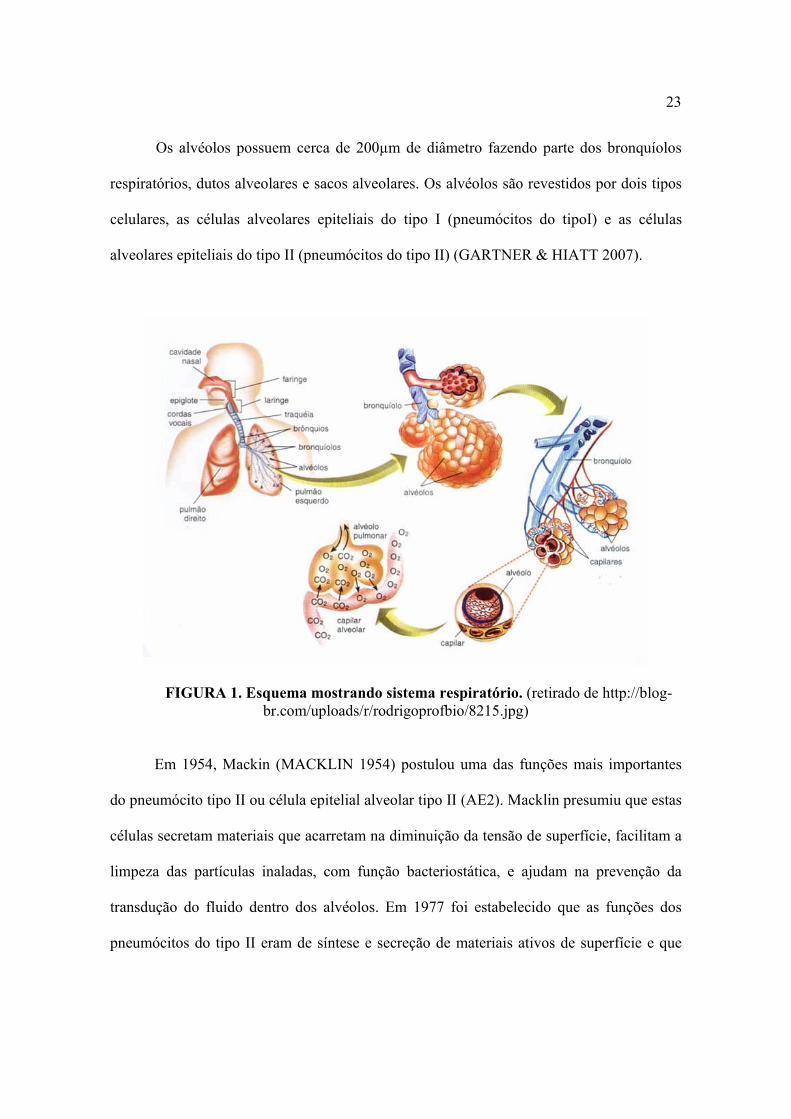
23
Os alvéolos possuem cerca de 200µm de diâmetro fazendo parte dos bronquíolos
respiratórios, dutos alveolares e sacos alveolares. Os alvéolos são revestidos por dois tipos
celulares, as células alveolares epiteliais do tipo I (pneumócitos do tipoI) e as células
alveolares epiteliais do tipo II (pneumócitos do tipo II) (GARTNER & HIATT 2007).
FIGURA 1. Esquema mostrando sistema respiratório. (retirado de http://blog-br.com/uploads/r/rodrigoprofbio/8215.jpg)
Em 1954, Mackin (MACKLIN 1954) postulou uma das funções mais importantes
do pneumócito tipo II ou célula epitelial alveolar tipo II (AE2). Macklin presumiu que estas
células secretam materiais que acarretam na diminuição da tensão de superfície, facilitam a
limpeza das partículas inaladas, com função bacteriostática, e ajudam na prevenção da
transdução do fluido dentro dos alvéolos. Em 1977 foi estabelecido que as funções dos
pneumócitos do tipo II eram de síntese e secreção de materiais ativos de superfície e que

24
serviriam como progenitor para as células AE1, as quais formam a barreira ar-sangue
(MASON 1977).
O pulmão é freqüentemente exposto à partículas do ambiente como micróbios,
partículas poluentes e substâncias que causam alergia. O macrófago alveolar é a célula
inicialmente responsável pelo reconhecimento, ingestão e limpeza de partículas inaladas
(BOWDEN 1987). Quando as partículas inaladas de microorganismos são opsonizadas por
anticorpos e/ou complemento, os macrófagos alveolares (MAs) podem se ligar mais
facilmente e fagocitar estas substancias via os receptores Fc (SWANSON & BAER 1995).
Os MAs são células imunes efetoras residentes nos espaços alveolares e vias aéreas
de condução. São responsáveis pela ativação de células inflamatórias e pela limpeza
pulmonar (HOCKING & GOLDE 1979). São responsáveis também por ingerir partículas
que podem ser o resultado de uma situação inflamatória aguda à também uma simples
ingestão e limpeza acarretando quase nenhuma inflamação (BECKER E COLS., 1989;
KOBZIK E COLS., 1990; ZHANG E COLS., 1993). Esta capacidade fagocítica é
especialmente importante para a limpeza de partículas inertes e partículas sem nenhum
risco de efeitos. Assim, os receptores que participam no reconhecimento das partículas
inaladas pelos MAs é o primeiro passo importante na manutenção da homeostasia
pulmonar.
2.1.1 LESÃO E REPARO PULMONAR
A inflamação é o evento inicial que precede a fibrose em diversas doenças
pulmonares, incluindo a fibrose pulmonar idiopática. Apesar da fibrose tecidual ser

25
resultante de um processo inflamatório agudo ou sub-agudo, já é conhecido que a
inflamação nem sempre vai resultar em fibrose (TRACEY 2002; NATHAN 2002). As
questões chave nas observações a cerca dos mecanismos de remodelamento das vias aéreas
são: (1) quais são os iniciadores inflamatórios específicos? e (2) qual é a seqüência de
eventos que culmina na fibroproliferação (início da fibrose)?
Em função da sua diversidade, as células epiteliais que revestem as vias aéreas são
particularmente bem adaptadas à proteção da mucosa em resposta a vários tipos de injúrias
(tabaco, poluentes, vírus), e bactérias, dentre várias outras causas. Estas células executam
uma série de funções críticas nas defesas imunes inatas. Na superfície do epitélio aéreo
(brônquios e bronquíolos) existem células especializadas incluindo as células colunares
ciliadas, células produtoras do muco, células de clara e células basais (GARTNER &
HIATT 2007). As células aéreas podem rapidamente mudar suas estruturas e funções para
se adaptar a mudanças do ambiente ou para efetuar o reparo epitelial após injúria
(GARTNER & HIATT 2007).
O epitélio que reveste as vias aéreas serve como uma barreira contra
microorganismos e moléculas agressivas. Suas funções incluem: a limpeza mecânica do
muco; adequação da homeostasia do transporte de íons e água, antioxidantes, antiproteases
e como uma barreira celular uma vez que apresentam junções intercelulares. Todas essas
funções são fundamentais para a proteção e manutenção da integridade do epitélio, a qual
pode ser rapidamente destruída após inflamação ou infecção (PARK E COLS., 2006). Em
uma situação normal, é sabido que as células ciliadas são células terminalmente
diferenciadas e que não são capazes de se dividirem, além de serem muito sensíveis a
injúrias. Entretanto, foi observado que essas células ciliadas podem se transdiferenciar em

26
células escamosas dentro de poucas horas após dano bronquiolar com o objetivo de manter
a integridade epitelial após a diferenciação (PARK E COLS., 2006).
Um dos passos mais importantes para a reparação pulmonar é a rápida e eficiente re-
epitelização da membrana basal desnuda após a injúria. O reparo epitelial eficiente pode
inibir o desenvolvimento da fibrose pulmonar, desde que esteja presente uma camada
intacta de células epiteliais suprindo a proliferação e deposição da matriz extracelular
efetuada pelos fibroblastos (ADAMSON E COLS., 1990).
Diversos mecanismos estão envolvidos no reparo epitelial alveolar. Células
alveolares epiteliais do tipo II (cuboidais) representam as células progenitoras que
regeneram o epitélio alveolar após injúria (ADAMSON & BOWDEN 1974; UHAL 1997;
FEHRENBACH 2001).
A ativação de quimiocinas, interleucinas, fatores de crescimento e fatores
estimuladores de colônias têm sido freqüentemente descritas durante estágios iniciais da
inflamação e durante a resposta quimiotática do epitélio aéreo (MESSAGE & JOHNSTON
2004; CHUNG 2001; KHAIR E COLS., 1996). Todos esses fatores são secretados por
células mesenquimais, endoteliais e macrófagos, mas também são secretados pelas células
epiteliais durante injúria e reparo. O TGF-β1 modula a composição da matriz provisória
(LECHAPT-ZALCMAN E COLS., 2006). As células epiteliais migram e estão envolvidas
com o aumento do reparo aéreo in vitro, via aumento da MMP-2. Este aumento da
metaloprotease em resposta ao TGF-β1 pode promover o reparo aéreo em condições de
homeostasia, enquanto que a MMP-9 pode ser crítica em um contexto inflamatório
caracterizado por um processo de remodelamento prolongado (LECHAPT-ZALCMAN E
COLS., 2006).

27
Outros fatores que estão envolvidos nos processos de lesão e reparação pulmonar
são as espécies reativas de oxigênio (ROS). Biologicamente, são considerados importantes:
radicais anions superóxidos (O2), peróxido de hidrogênio (H2O2), radicais hidroxila (OH) e
ácidos (HOCl). As espécies reativas de nitrogênio incluindo peroxinitrato (ONOO),
também tem sido implicadas na oxidação (nitração) de proteínas e lipídeos
(ISCHIROPOULOS 1998). As espécies reativas de oxigênio e nitrogênio podem acarretar a
injúria tecidual através de vários mecanismos, incluindo: (1) dano direto ao DNA
resultando em mutações; (2) peroxidação lipídica com a formação de moléculas vasoativas
e pró-inflamatórias; (3) oxidação de proteínas que alteram a atividade protéica (FIALKOW
E COLS., 1993;1994;1997), levando à liberação de proteases e à inativação de
antioxidantes e enzimas antiproteases (GADEK & PACHT 1996) e (4) alteração de fatores
de transcrição como proteína-1 e fator nuclear β (NF) κ- β, levando a um aumento na
expressão de genes pró-inflamatórios (HADDAD E COLS., 2000).
No metabolismo normal, as células a fim de neutralizar e conter os efeitos das ROS
liberadas expressa antioxidantes endógenos como, superóxido dismutase, catalase e
glutationa peroxidase (FINK 2002). Entretanto, estes anti-oxidantes são rapidamente
utilizados durante respostas inflamatórias agudas.
2.2 LESÃO PULMONAR AGUDA (ALI) E O MODELO DO LIPOPOLISSACARÍDEO BACTERIANO (LPS)
A Injúria Pulmonar Aguda (ALI) e a sua forma mais severa, a Síndrome do Estresse
Respiratório Agudo (ARDS) são importantes causas de morbidade e mortalidade no mundo
(WARE & MATTHAY 2000). Uma das características mais marcante da ALI/ARDS é o

28
acúmulo de fluido rico em proteínas, ou seja, edema no compartimento alveolar pulmonar
(WARE & MATTHAY 2000), causada pelo aumento do fluxo de fluidos nos espaços
aéreos e a diminuição do fluxo fora dos espaços aéreos. A Síndrome do Estresse
Respiratório Agudo representa um conceito fisiopatológico no qual as anormalidades
pulmonares são devidas a uma lesão pulmonar aguda (ASHBAUGH E COLS., 1967;
PONTOPPIDAN E COLS., 1972).
O lipopolissacarídeo bacteriano (LPS) ou endotoxina é uma mistura de fragmentos
de paredes celulares externas das bactérias gram-negativas. Ele é vital para a integridade
estrutural, funcional bacteriana e atua como um potente ativador de macrófagos. O LPS
induz inflamação local e sistêmica, pois é reconhecido pelas células locais que vão atuar na
tentativa de eliminá-lo e, como conseqüência, ocorre a liberação de fatores mediadores da
inflamação iniciando o processo de alteração da permeabilidade da membrana alvéolo-
capilar. Conseqüentemente, ocorre um grande afluxo de neutrófilos para o tecido pulmonar,
acarretando assim, uma liberação de várias enzimas proteolíticas e citocinas que estão
diretamente ligadas à agressão pulmonar (OPAL 2007).
A lesão pulmonar aguda (ALI), de manifestação grave, e a Síndrome do Estresse
respiratório agudo (ARDS) são igualmente definidas como disfunção grave da troca gasosa
e de anormalidades radiográficas peitorais (BERNARD E COLS., 1994). ARDS e ALI
podem ocorrer na presença de vários eventos, tais como, sepse, trauma ou transplante de
órgãos (ATABAI & MATTHAY 2002). Inflamação intra-pulmonar e endotelial e dano
epitelial culminando nas respostas reparativas, são componentes que estão envolvidos na
ALI e na progressão para a ARDS (BERNARD E COLS., 1994; MATTHAY E COLS.,
2003).

29
A incidência da ALI ainda é incerta, mesmo nos EUA, onde o Instituto Nacional de
Saúde, em 1972, a estimou em 75 casos/100 mil habitantes ao ano. Porém, estudos mais
recentes encontraram números bem mais baixos, de 1,5 a 15/100 mil habitantes ao ano
(BERNARD E COLS., 1994; WARE & MATTHAY 2000; LUCE 1998; GARBER E
COLS., 1996; ZILBERBERG & EPSTEIN, 1998; HUDSON & STEINBERG 1999;
ATABAI & MATTHAY 2002, VILLAR & SLUTSKY 1989; LUHR E COLS., 1999;
ARROLIGA E COLS., 2002; BERSTEN E COLS., 2002). O número de estudos
epidemiológicos vem crescendo nos últimos anos após a American-European Consensus
Conference on ARDS de 1994, ter redefinido os critérios diagnósticos. No Brasil, ainda não
temos estudos populacionais.
As características mais clássicas da ALI são o aumento da permeabilidade capilar,
edema intersticial e alveolar, influxo da circulação de células inflamatórias e a formação de
membranas hialinas (são fragmentos celulares, resultantes da morte das células alveolares
do tipo I e de fibrina depositada no espaço alveolar) (BERNARD E COLS., 1994;
MATTHAY E COLS., 2003). O aumento da permeabilidade leva ao edema pulmonar e a
um balanço defeituoso na manutenção do fluxo dentro dos espaços aéreos e nos
mecanismos biológicos da sua remoção. A gravidade da ALI depende do balaço entre as
injúrias alveolares epiteliais e/ou as injúrias vasculares endoteliais e os seus mecanismos de
reparo.
O LPS é reconhecido através da sua interação com proteínas séricas (proteína
LBP/receptor CD14/TLR4), ou com proteínas do surfactante pulmonar (VERNOOY E
COLS., 2002). Os receptores Toll like (TLRs) são expressos na maioria dos tecidos
animais, sendo que, os humanos e os camundongos possuem 10 subtipos. Contudo, os
camundongos possuem dois receptores adicionais onze e doze que não estão representados

30
em humanos (BEUTLER 2004). Os receptores TLRs reconhecem não somente bactérias e
fungos, mas também protozoários (CAMPOS E COLS., 2001) e vírus (HOEBE E COLS.,
2003; ALEXOPOULOU E COLS., 2001), sendo que o TLR4 reconhece mais
especificamente o LPS (POLTORAK E COLS., 1998). O TLR4 é um dos onze parálogos
identificados em mamíferos, dez dos quais foram descritos (CHUANG & ULEVITCH
2000;2001).
2.3 RECEPTORES P2X7
O conceito de que o ATP é uma molécula sinalizadora extracelular em adição ao
papel que possui no metabolismo intracelular e como sendo uma molécula energética, é
aceito a algum (GORDON 1986). Com o desenvolvimento de novas tecnologias e métodos
de estudo, a sinalização purinérgica tem sido estabelecida principalmente como um
sinalizador rápido envolvido na neurotransmissão, mas também tem ganhado bastante
atenção em tecidos animais e em diversos processos biológicos, incluindo proliferação
celular (BURNSTOCK & KNIGHT 2004). Nos 30 anos desde que a sinalização
purinérgica foi primeiramente proposta, o conceito de purinoreceptores têm sido
transformado primariamente da idéia de alvos terapêuticos para a prática clínica
(BURNSTOCK 2002). Conseqüentemente seus subtipos de receptores para nucleotídeos
extracelulares tem sido clonados em várias espécies de mamíferos. Os purinoreceptores
tiveram sua classificação original baseada na sua farmacologia e função e foram
primariamente divididos em receptores P1, tendo como a adenosina como ligante, e em
receptores P2, tendo o ATP e ADP como os principais ligantes. Os receptores P2 foram
mais tarde subdivididos em P2X e P2Y (RALEVIC & BURNSTOCK 1998). Existem, até
hoje, sete receptores P2X e oito receptores P2Y clonados e caracterizados.

31
Os receptores P2X são formados por três subunidades. Cada subunidade consistindo
em dois domínios transmembrana. São receptores de canais catiônicos não seletivos. Sob
condições fisiológicas normais, quando ativados, resultam em um influxo de Na+ e Ca2+ e
saída de K+ através da membrana celular levando a despolarização da membrana plasmática
(COUTINHO-SILVA E COLS, 1996).
Os receptores P2X7 (FIGURA 2) são bastante expressos em monócitos,
macrófagos, células dendríticas e linfócitos (COUTINHO-SILVA E COLS., 1999; 2001;
DI VIRGILIO 2001). Estão envolvidos na eliminação de micobactérias e clamídias em
macrófagos infectados (LAMMAS E COLS., 1997; COUTINHO-SILVA E COLS., 2003).
Os receptores P2X7 foram identificados no pulmão de ratos, camundongos e em humanos
(YAMAMOTO E COLS., 2004; SCHWIEBERT & ZSEMBERY 2003; LEWIS & EVANS
2001; SIM E COLS., 2004) inclusive em células epiteliais alveolares (CHEN E
COLS.,2004). A exposição desses receptores ao ATP extracelular leva a abertura de um
canal seletivo de cátions, que permite um influxo de Ca+ e Na+ e a saída de K+. A
estimulação com certas concentrações de ATP, na maioria das células que apresentam o
receptor P2X7, acarreta a formação de bolhas apicais membranares (VIRGINIO E COLS.,
1999) e morte celular (CHOW E COLS., 1997; DI VIRGILIO E COLS., 2001;
COUTINHO-SILVA E COLS., 1999).
Análises proteômicas identificaram 11 proteínas que estão associadas com o
receptor P2X7 em ratos, incluindo laminina α3, integrina β2, β-actina, α-actinina, MAGuK,
fosfatidilinositol-4 quinase e três proteínas de choque térmico (HSP70, HSP71 e HSP90)
(KIM E COLS., 2001) (FIGURA 2). Tem sido sugerido que esses receptores são parte de
um complexo multi-protéico, que facilita a comunicação entre a matriz extracelular, o
citoesqueleto de actina e cascatas de sinalização intracelular.

32
FIGURA 2. Esquema mostrando o receptor P2X7 e suas interações com componentes celulares. (modificado de KIM E COLS., 2001)
2.3.1 PULMÃO E RECEPTORES P2X7
Atualmente pouco é conhecido sobre a distribuição dos receptores P2X e P2Y no
sistema respiratório. E sabe-se que apesar deste fato, o epitélio aéreo tem sido um dos
tecidos mais estudados com o objetivo de se analisar os efeitos de agonistas purinérgicos
(DAVIS 2002; KUNZELMANN E COLS., 2003; HAYASHI E COLS., 2005).
É sabido que as células epiteliais alveolares liberam ATP e possuem tanto
receptores P2X quanto P2Y (SCHWIEBERT & ZSEMBERY, 2003; NOVAK 2003;

33
TAYLOR E COLS., 1999). Os receptores P2X4, por exemplo, já foram identificados em
cultura primária de células epiteliais alveolares, derivadas tanto de vias aéreas quanto de
epitélio alveolar (SOTO E COLS., 1996; TAYLOR E COLS., 1999). Outras isoformas
também foram encontradas no epitélio respiratório (LEPZIGER 2003; TAYLOR E COLS.,
1999). Os receptores P2X7 e P2X4 são utilizados como um novo marcador para células
epiteliais alveolares do tipo I (CHEN E COLS., 2004; QIAO E COLS 2003).
Os receptores P2X7 já foram identificados em: células endoteliais da artéria
pulmonar de ratos e humanos (YAMAMOTO E COLS., 2004; NOVAK 2003), no pulmão
de camundongos (LEWIS & EVANS, 2001) e em células alveolares epiteliais de rato
(CHEN E COLS., 2004).
2.3.2 INFLAMAÇÃO E RECEPTORES P2X7
A inflamação é um processo que pode ser desencadeado por microorganismos
invasores, estes estimulam a imunidade adaptativa através de um número quase infinito de
antígenos e a imunidade inata através de um repertório muito mais simples e conservado
(JANEWAY & MEDZHITOV 2002). Estes repertórios são desencadeados pelos padrões
moleculares associados à patógenos (PAMPs) (JANEWAY & MEDZHITOV 2002). Os
PAMPs sensibilizam as células inflamatórias inicialmente durante a infecção, é um
estímulo potente para a imunidade inata e são referidos como sinais de perigo exógenos.
Entretanto, todos os imunologistas sabem que o estresse celular ou dano tecidual
desencadeia a inflamação mesmo na ausência de patógenos. É sabido hoje em dia que as
células imunes reagem à injúria induzida pela liberação de moléculas que são normalmente
localizadas dentro das células (MATZINGER 2002). Fazendo parte do que chamamos de

34
sinais endógenos. Por este motivo, elas também são conhecidas como “padrões moleculares
associados ao dano” (DAMPs) (KORNBLUTH & STONE 2006). Contudo, tanto os
PAMPs quanto os DAMPs são necessários para iniciar uma resposta imune eficiente.
Moléculas sinalizadoras são essenciais para a manutenção das reações inflamatórias,
como citado anteriormente. Elas estão envolvidas no recrutamento de leucócitos,
mastócitos e células apresentadoras de antígenos ao sítio de inflamação. Atuam na ativação
de vasos e no prolongamento de outras respostas inflamatórias. Com o advento do
conhecimento acerca do ATP fazendo papel de uma molécula inflamatória (DI VIRGILIO
E COLS., 2001), essa molécula é secretada nos sítios de inflamação como resultante da
resposta inflamatória.
O ATP extracelular é secretado por células infectadas e submetidas a estresse. Em
virtude deste fato, é considerado atualmente como um marcador de “sinal de perigo” uma
vez que, atuando através do receptor P2X7 pode levar a estimulação da secreção de
citocinas pró-inflamatórias (GALLUCCI & MATZINGER 2001; BOURS E COLS., 2006;
HAAG E COLS., 2007). Uma das habilidades mais notáveis do ATP extracelular é a
capacidade de permeabilizar temporariamente a membrana plasmática de certas células,
fato observado primeiramente em mastócitos peritoneais (COCKCROFT & GOMPERTS
1979). Outro papel na inflamação é a constatação de que o ATP também induz o
processamento e a liberação de citocinas pró-inflamatórias (particularmente IL-1 e IL-6)
pela ação de receptores P2X7 (NORTH 2002; BULANOVA, E COLS., 2005; FERRARI E
COLS., 2006). Em animais P2X7-/-, macrófagos não foram capazes de secretar IL-1β em
resposta ao ATP (SOLLE E COLS., 2001). Foi mostrado também que camundongos P2X7-/-

35
desenvolveram uma artrite reduzida no modelo após injeção de colágeno (LABASI E
COLS., 2002) e atenuação da lesão obstrutiva renal (GONCALVES E COLS., 2006).
A ativação da sinalização intracelular dos receptores P2X7 pode ter efeitos
estimulatórios na síntese e secreção de citocinas. Em diferentes tipos celulares, os
receptores P2X7 levam a produção e secreção de mediadores inflamatórios, tais como: IL-1,
IL6, TNF, IL18, monocyte chemoattractant protein-1 (MCP-1) (BOURS E COLS., 2006;
FERRARI E COLS., 2006; HEWINSON E COLS., 2008; TAKENOUCHI E COLS.,
2008) clivagem de metaloproteases, CD23, CD27, selectina-L, e matriz (GU E COLS.,
1998; 2006). Em mastócitos, a ativação dos receptores causa um aumento na expressão de
IL-4, IL-6, IL-13 e TNF-α (BULANOVA E COLS., 2005; PELEGRIN E COLS., 2008).
Em macrófagos, a ativação destes receptores aumenta os níveis de mRNA para óxido
nítrico sintase induzível (iNOS) e TNF-α causados por lipopolissacarídeos (LPS)
(TONETTI E COLS., 1994; 1995; HU E COLS., 1998).
2.3.3 LPS E RECEPTORES P2X7
Diversos estudos tem mostrado que os nucleotídeos extracelulares podem modular a
ação do LPS sob a função macrofágica (TANKE E COLS., 1991; PROCTOR E COLS.,
1994; TONETTI E COLS., 1995; DENLINGER E COLS., 1996; 1998; HUMPHREYS &
DUBYAK 1996; FERRARI E COLS., 1997; HU E COLS., 1998; SPERLAGH E COLS.,
1998; PERREGAUX E COLS., 2000; SOLLE E COLS., 2001). Entretanto, o papel
específico do clearance da Mycobacterium tuberculosis e os receptores para nucleotídeos
ainda envolvidos no processo ainda não esta claro (LAMMAS E COLS., 1997; SIKORA E
COLS., 1999). Os receptores P2X7 tem sido descritos como potenciadores da produção de

36
citocinas inflamatórias induzidas pelo LPS, citocinas essas que são extremamente
importantes para a fisiopatologia do choque séptico (WATTERS E COLS., 2001; DI
VIRGILIO E COLS., 2001). Outros estudos mostraram que macrófagos tratados com
agonistas ou antagonistas do receptor P2X7 podem modular a produção de IL-1β e NO
(TONETTI E COLS., 1995; DENLINGER E COLS., 1996; FERRARI E COLS., 1997;
HU E COLS., 1998). Adicionalmente, macrófagos estimulados pelo LPS em camundongos
nocautes para o receptor P2X7 produzem níveis menores de IL-1β e IL-6 após a co-
administração de ATP (SOLLE E COLS., 2001). O co-tratamento com o 2-methylthio-ATP
protege camundongos da injeção letal de LPS e promove uma redução dos níveis de TNF-α
e IL-1α (PROCTOR E COLS., 1994).
O LPS por si só é capaz de promover a síntese e a externalização de IL-1β, mas a
subseqüente adição de ATP as amostras tratadas com LPS aumentam a externalização da
citocina de dez a trinta vezes (PERREGAUX E COLS., 2000).
Estudos recentes indicam um mecanismo similar pelos ligantes TLRs para ATP
induzindo a ativação da caspase 1 em macrófagos peritoneais murinos (MARIATHASAN
E COLS., 2004; YAMAMOTO E COLS., 2004).

37
OBJETIVOS

38
3 OBJETIVOS
3.1 OBJETIVO GERAL
Estudar o envolvimento da sinalização pelo ATP na via de ativação dos receptores
P2X7 no modelo de lesão pulmonar aguda pelo LPS.
3.2 OBJETIVOS ESPECIFICOS
1- Caracterizar a lesão pulmonar induzida pela instilação intratraqueal de LPS em animais
selvagens e nocautes para o receptor P2X7.
2- Caracterizar a imunoexpressão dos receptores P2X7 na lesão pulmonar aguda induzida
pelo LPS
3- Avaliar o recrutamento e ativação de células inflamatórias na lesão pulmonar aguda em
animais selvagens e nocautes para o receptor P2X7.
4- Quantificar no lavado broncoalveolar (BAL) a presença de Interleucina 1β, TGF-β, de
óxido nítrico (NO) e a lise celular através da dosagem da enzima lacto dehidrogenase em
animais selvagens e nocautes para o receptor P2X7.
5- Quantificar as mudanças das tramas de fibras elásticas, colágenas e de fibronectina no
parênquima pulmonar em animais selvagens e nocautes para o receptor P2X7.
6- Quantificar no parênquima pulmonar a imunoexpressão das metaloproteases-2 e -9 em
animais selvagens e nocautes para o receptor P2X7.
7- Analisar os parâmetros da mecânica respiratória em animais selvagens e nocautes para o
receptor P2X7.

39
MATERIAIS E MÉTODOS

40
4 MATERIAIS E MÉTODOS
4.1 INSTILAÇÃO DO LPS
A utilização e o protocolo de experimentos foram aprovados pelo Comitê de Ética
do Centro de Ciências da Saúde (CCS) da Universidade federal do Rio de Janeiro. Foram
utilizados camundongos C57/BL6 nocautes para o gene do receptor P2X7 (P2X7-/-) e
selvagens, de ambos os sexos, oriundos dos laboratórios de Patologia Celular (ICB) e de
Imunofisiologia (IBCCF) da Universidade Federal do Rio de Janeiro, com 8-10 semanas de
idade e peso aproximado de 20g. A lesão pulmonar foi induzida através da instilação de
LPS (E. coli O55:B5, SIGMA, USA), por via intratraqueal, em dose única (60 µg), diluída
em 50 µL de solução salina estéril.
Os camundongos P2X7 (-/-) gerados inicialmente pelo acasalamento de
camundongos 129/Ola com camundongos C57BL/6 mice. A prole foi acasalada por 12
gerações com camundongos C57BL/6 (COUTINHO-SILVA E COLS., 2003), sendo
gentilmente cedidas pelo Dr James Mobley (PGRD, Pfizer Inc, Groton, CT, USA). Os
animais P2X7 (-/-) apresentam deleção da região do T1527 to T1605, localizado no exon
13.
4.2 PROCEDIMENTO CIRÚRGICO
Os animais foram anestesiados com ketamina® (35mg/Kg) e xilazina® (9mg/Kg) por
via intraperitoneal. Na região cervical foi realizada incisão da pele e divulsão da
musculatura para exposição da traquéia. Posteriormente o LPS foi instilado através de

41
seringa de insulina (50µL). Após este procedimento, a pele e o tecido celular subcutâneo
foram suturados com fio de nylon 4-0.
4.3 PROTOCOLO EXPERIMENTAL
Após 24 horas da instilação de LPS, os animais foram anestesiados e eutanasiados
por secção medular. Os grupos foram divididos seguindo o esquema (FIGURA 3). Para
cada grupo foram utilizados cinco animais (n = 5).
FIGURA 3. Esquema mostrando protocolo experimental.
4.4 LAVADO BRONCOALVEOLAR (BAL)
Após eutanásia, os animais foram colocados em decúbito ventral. A pele e toda
camada muscular adjacente à traquéia foram removidas. Uma cânula de polietileno foi

42
inserida na traquéia e um volume de 1mL de solução salina (PBS pH 7.4) à 37o C +
heparina (10 UI / mL) instilado e aspirado em cada animal.
O material obtido pelo BAL foi centrifugado a 250g durante cinco minutos a 4ºC
(Fanem®, São Paulo, Brasil). O sobrenadante foi coletado e congelado à -4°C para análises
posteriores (LDH e nitrito).
A dosagem IL-1β foi determinada usando-se o kit comercial ELISA, com o uso de
anticorpo monoclonal e policlonal contra camundongo. A detecção limite foi de 10pg/ml,
estando de acordo com o protocolo da empresa (R&D Systems Ltda., USA). Citocinas
recombinantes de camundongos foram utilizadas em todos os ensaios.
4.5 PROCESSAMENTO HISTOLÓGICO
Para a análise histopatológica os animais foram previamente anestesiados e com a
traquéia exposta foi realizada a sua oclusão ao final da expiração. Esta manobra tem por
finalidade manter o volume de reserva expiratório e do volume residual, mantendo assim a
capacidade residual funcional, ou seja, impedir o colabamento dos pulmões. Logo após este
procedimento os animais foram eutanasiados.
Os pulmões foram fixados em formol tamponado a 10% por um período de 12
horas. Após fixação estes foram processados em séries crescentes de etanol e banhos de
xileno (100%) objetivando desidratação e diafanização do material. Em seguida os mesmos
foram impregnados em parafina. Cortes de 4 µm do tecido pulmonar foram obtidos por
microtomia em Micrótomo Leica (USA) e corados pela técnica de Hematoxilina-Eosina
(HE).

43
A trama colagênica dos granulomas foi observada através da coloração pelo Sirius
red modificado para microscopia confocal (DOLBER & SPACH, 1993). Após a
desparafinização e hidratação do material com três banhos de xilol e álcoois decrescentes
(100 / 95 / 70%), respectivamente, as lâminas foram lavadas em água destilada por duas
vezes de 5 minutos. Posteriormente as lâminas foram postas na solução de ácido
fosfomolíbdico (0,2%) por 1 minuto. Após o ácido, as lâminas foram colocadas na solução
de picrosirius (Sirius red F3 BA - 0,2g + Solução saturada de ácido pícrico [Dissolver 1 a
1,5g de ácido pícrico em 100ml de água destilada quente, Deixando esfriar e filtrado no dia
seguinte] - 200ml) por 90 minutos. Após 90 minutos, as lâminas foram lavadas com ácido
clorídrico 0,01N durante 2 minutos.e em álcool 70% por 45 segundos. Após este processo
as lâminas foram banhadas em banhos crescentes de álcoois e em três banhos de xileno
puro, todos por um período de 5 minutos. Finalmente, as lâminas foram montadas em
Entellan®.
As fibras elásticas foram coradas pelo método da orceína. Após a desparafinização e
hidratação do material com três banhos de xilol e álcoois decrescentes (100 / 95 / 70%),
respectivamente, as lâminas foram lavadas em água destilada por duas vezes de 5 minutos.
Posteriormente as lâminas foram postas na solução de orceína (orceína -1g +Etanol -100ml
+ ácido Clorídrico concentrado -.0,7ml) por 30 minutos. Após este processo, as lâminas
foram diferenciadas em álcool clorídrico 0,1% rapidamente, e lavadas em água destilada
por 2 vezes de 3 minutos. Após a lavagem, as lâminas foram contracoradas com solução de
ácido pícrico saturado (Dissolver em 100mL de água destilada quente 1,5g de Ácido
Pícrico, deixando esfriar e filtrado no dia seguinte) por 30 segundos. Após este processo as
lâminas foram banhadas em banhos crescentes de álcoois e em três banhos de xileno puro,
todos por um período de 5 minutos. Finalmente, as lâminas foram montadas em Entellan®.

44
Os mastócitos foram corados pela técnica de alcian blue pH 2,5. Após a
desparafinização e hidratação do material com três banhos de xilol e álcoois decrescentes
(100 / 95 / 70%), respectivamente, as lâminas foram lavadas em água destilada por duas
vezes de 5 minutos. Posteriormente, as lâminas foram colocadas na solução de alcian blue
pH 2,5 (alcian blue GX-1g+ ácido acético 3% -100mL) por 5 minutos. Após a solução as
lâminas foram lavadas em água estilada rapidamente e contracoradas com vermelho neutro
0,5% por 3 minutos. Após o vermelho neutro, as lâminas foram lavadas em água destilada
rapidamente e banhadas em banhos crescentes de álcoois e em três banhos de xileno puro,
todos por um período de 5 minutos. Finalmente, as lâminas foram montadas em Entellan®.
4.6 IMUNOHISTOQUÍMICA E HISTOQUÍMICA 4.6.1 ANTICORPOS UTILIZADOS
Foram utilizados os seguintes anticorpos primários:
1- Anticorpo policlonal coelho contra o receptor P2X7 extracelular (Alamone Labs,
-Jerusalem, Israel cat. APR008, na diluição de 1/200.
2- Anticorpo policlonal coelho contra pan-TGF-β da R&D Systems, USA – cat.
AB00NA), na diluição de 1/200.
3- Anticorpo policlonal cabra contra a MMP-2 (C-19) da Santa Cruz
Biotechnologies, CA, USA, cat. sc-6838, na diluição de 1/100.
4- Anticorpo policlonal cabra contra a MMP-9 (C-20) da Santa Cruz
Biotechnologies, CA, USA, cat. sc-6840, na diluição de 1/100.

45
5- Anticorpo monoclonal rato contra o antígeno F4/80 da Serotec, USA, cat.
MCA497, na diluição de 1/200.
6 - Anticorpo policlonal coelho contra fibronectina da DAKOCYTOMATION,
Carpinteria, CA, USA, cat. A245, na diluição de 1/800.
4.6.2 IMUNOHISTOQUÍMICA
Cortes de parafina foram desparafinizados e hidratados com três banhos de xilol e
álcoois decrescentes (100 / 95 / 70%) por 5 minutos, respectivamente. Os resíduos
aldeídicos foram inibidos pelo Bórax 3% com dois banhos de 15 minutos. As lâminas
foram lavadas em água destilada em dois banhos de cinco minutos. Após duas lavagens em
tampão fosfato salino (PBS) pH 7,4, a peroxidase endógena foi inibida com H2O2 3%
(Reagen) em metanol por 20 minutos, seguindo-se de dois banhos de PBS pH 7.4 por cinco
minutos cada. Recuperação antigênica foi realizada com tampão citrato pH. 6,0 no steamer
por 30 minutos, na temperatura de 96 oC, exceto para o F4/80, onde os cortes foram
submetidos a digestão enzimática com solução a 0,1% de tripsina, 0,01% de cloreto de
cálcio, no Tris-HCl (Trypsin tablets, SIGMA ), pH 7,6, a 38oC, por 30 minutos.
Posteriormente os cortes histológicos foram submetidos a bloqueio das ligações
inespecíficas dos anticorpos sendo realizado com a solução de PBS-BSA (albumina sérica
bovina, cat. A9647, SIGMA) 10%: leite molico 8% (v/v), exceto para o anticorpo anti-
P2X7 onde as lâminas foram incubadas com PBS-BSA 5%, soro normal de carneiro 10%,
por 30 minutos, seguido por três lavagens de cinco minutos cada em PBS. Foi realizado um
outro bloqueio das ligações inespecíficas com PBS-BSA5% + soro normal de camundongo
10% por uma hora.

46
Após estes procedimentos, as lâminas foram incubadas com os anticorpos diluídos
na solução de PBS/BSA 3% - Triton 0,1% - Tween 0,05%, permanecendo até o próximo
dia, em câmara úmida, a 4 oC.
No dia seguinte, após alcançar a temperatura ambiente, as lâminas foram lavadas
duas vezes em Tampão PBS pH 7.4 e uma vez em PBS - Tween 0,25%, por 10 minutos
cada e incubadas com o anticorpo secundário respectivo: anti -IgG de coelho feita em
cabra, biotinilada (BA-1000, Vector Laboratories, USA), na diluição de 1:50; anti -IgG de
rato feita em cabra, biotinilada, (BA-1000, Vector, USA) na diluição de 1:100, anti -IgG de
cabra feita em cabra, biotinilada (BA-5000, Vector Laboratories, USA) na diluição de
1:50, por uma hora. Ao final desse período e após duas lavagens com PBS - Tween 0,25%,
as lâminas foram incubadas com a Estreptavidina - Peroxidase (E-8386, Sigma, USA)
diluída 1:50 em PBS por 1 hora e reveladas com o substrato cromógeno diaminobenzidina -
DAB (K-3468, DAB líquida, DAKO, Carpinteria, CA, USA) por 10 a 15 minutos. Após 2
lavagens com tampão PBS pH 7.4 - Tween 20 0,25% por cinco minutos e uma lavagem em
água destilada por 10 minutos, as lâminas foram contra-coradas com hematoxilina de
Harris diluída (1:3) e montadas em Entellan® (Merck).
Como controles negativos da reação, os cortes histológicos foram incubados com
soro não imune de rato, coelho ou cabra ao invés do anticorpo primário respectivo. Para o
P2X7 o controle negativo consistiu na incubação das lâminas com a solução do anticorpo
diluído contendo o peptídeo P2X7 (Alamone) na proporção de 1:2, preparado com
antecedência de 1 hora.

47
4.6.3 HISTOQUÍMICA PARA A DETECÇÃO E QUANTIFICAÇÃO DOS MACRÓFAGOS PULMONARES ATRAVÉS DA MARCAÇÃO DE RADICAIS CARBOIDRÁTICOS
A fim de analisar e quantificar os macrófagos pulmonares ativados foi utlizada a
técnica histoquímica para a detecção da lectina Bandeiracea Griffonia Simplicifolia
isoagglutinin, que é um marcador de macrófago ativado (MADDOX E COLS., 1982 a e b)
Após a desparafinização e hidratação do material com três banhos de xilol e álcoois
decrescentes (100 / 95 / 70%) e banhos em água destilada respectivamente, os resíduos
aldeídicos foram inibidos pelo bórax 5 % por 20 minutos, seguindo-se a inibição da
peroxidase endógena com H2O2 3% (Reagen) em metanol por 20 minutos e banhos
seguidos de tampão PBS pH 7.4 por cinco minutos cada. Realizou-se o bloqueio das
ligações inespecíficas com PBS - BSA 3% por uma hora. Após este procedimento, as
lâminas foram incubadas com a lectina Bandeiracea Griffonia Simplicifolia conjugada à
biotina (B-1105, Vector Laboratories, USA) em câmara úmida na concentração de 1:100
em PBS-BSA 1% + Solução traço de metais, contendo Cloreto de Cálcio, Cloreto de
Magnésio, Cloreto de Manganês + Azida de Sódio durante a noite a 4°C. No dia seguinte,
após alcançar a temperatura ambiente, as lâminas foram lavadas em PBS pH 7.4 - Tween
20 0,25%, incubadas com a Estreptavidina - Peroxidase (E-8386, Sigma, USA) diluída 1:50
em PBS por 40 minutos e revelada pela DAB (K-3468, DAB líquida, DAKO, Carpinteria,
CA, USA) por 10 a 15 minutos. Após duas lavagens com água destilada por cinco minutos,
as lâminas foram contra-coradas com Hematoxilina de Harris (diluída 1:3) e montadas em
Entellan® (Merck).

48
4.7 HISTOMORFOMETRIA
O estudo morfométrico foi realizado através de um sistema de captura de imagens
constituído de câmara fotográfica digital EVOLUTION (Media Cybernetics, USA)
acoplada a microscópio de luz E 500 NIKON (Japão) e a um computador contendo o
programa de análise de imagens IMAGEPRO–PLUS 5.0 Media Cybernetics, USA).
Imagens digitalizadas de alta qualidade (2048 x 1536 pixels buffer) foram obtidas das
lâminas coradas pela hematoxilina-eosina, orceína, picrosírius, alcian blue. griffonia
simplicifora (BSL-1), F4/80, P2X7, TGF-β, Fibronectina, MMP-2, MMP-9, utilizando a
lente objetiva de 40x.
Os resultados foram expressos como percentual de imunoexpressão (superfície
reativa / superfície total examinada X 100) nas análises da imunoexpressão de TGF-β,
P2X7, MMP-2 e MMP-9; para a análise de mastócitos, o resultado foi expresso como o
número absoluto de células/µm2 ou número absoluto de células/campo para células reativas
para o BSL-1 e F4/80. Para as fibras colágenas e elásticas, os resultados foram expressos
como densidade de superfície das fibras /µm2.
Para a quantificação da celularidade no parênquima pulmonar foi realizado o
método de retículo de 100 pontos e 50 linhas acoplado a um microscópio convencional. A
fração de áreas alveolares colapsadas e normais foram realizadas utilizando a técnica de
contagem de pontos (SANTOS E COLS., 2006).

49
4.8 ANÁLISE ESTATÍSTICA
As variáveis foram analisadas descritivamente, sendo submetidas a um teste de
normalidade. As variáveis que apresentaram distribuição normal foram analisadas pelos
testes paramétricos T DE STUDENT, para amostras independentes (two tailed). Para a
comparação de mais de dois grupos, foi utilizado o teste ONE WAY ANOVA e quando
obtido o efeito principal foi empregado pós-teste de Tukey. Para a análise de variáveis que
não apresentaram distribuição normal, foram utilizados os testes não paramétricos
KRUSKAL-WALLIS e MANN-WHITNEY para análise de dois grupos e análise de mais
de dois grupos, respectivamente. Em todas as análises realizadas foi admitido p < 0,05 (5%)
como significativo.
4.9 MECÂNICA RESPIRATÓRIA
A mecânica foi realizada no laboratório de fisiologia pulmonar no IBCCF-UFRJ.
Esta técnica nós permite analisar a função pulmonar (fisiologia pulmonar) dos animais
como um todo, que juntamento com os dados morfológicos nos permite ter uma analise
mais fidedígna acerca do assunto abordado neste trabalho. Após 24 horas de LPS, os
animais foram sedados com diazepam (1 mg i.p.), em seguida pesados (balança Filizola,
modelo BR, Indústrias Filizola SA, SP, Brasil) e, então, anestesiados com pentobarbital
sódico (20 mg/kg i.p.).
Depois de anestesiados, os animais foram colocados em uma pequena mesa sob
foco cirúrgico em decúbito dorsal, sendo seus membros fixados por esparadrapo. Foi
realizada traqueotomia com introdução de cateter Jelco 20G com 32 mm de comprimento e

50
0,8 mm de diâmetro interno, sendo a cânula fixada à traquéia por meio de fios de algodão.
Os animais foram paralisados com trietiliodeto de galamina (2 mg/kg, através de
administração intra-abdominal).
Os camundongos foram, então, acoplados à prótese ventilatória por um ventilador
de fluxo constante (Samay VR15, Universidad de la Republica, Montevideu, Uruguai) e
ventilados com freqüência de 100 incursões respiratórias por minuto e um volume corrente
(VT) de 0,2 mL.
Após a adaptação ao respirador, os animais foram submetidos à incisão cirúrgica
por tesoura na linha média do abdômen estendendo-se a região torácica, sobre o esterno. A
pele foi rebatida por tração laterial. A seguir, a musculatura abdominal foi incisada até o
bordo inferior das costelas até atingir a linha axilar anterior, bilateralmente. Com a
cavidade abdominal aberta, foi possível visualizar o diafragma, que foi perfurado e
secionado segundo a mesma orientação da abertura da parede abdominal. Imediatamente
antes da perfuração do diafragma foi instalada pressão positiva ao final da expiração
(PEEP) de 2 cmH2O (SALDIVA E COLS., 1992). Após a retirada do diafragma, a parede
torácica foi removida.
O ventilador foi ajustado previamente para gerar, quando desejado, uma pausa de
cinco segundos ao final da inspiração. Foram tomados cuidados especiais na manutenção
do volume (VT = 0,2 mL) e fluxo (V’= 1 mL/s) constantes em todos os animais, a fim de
evitar os efeitos de diferentes fluxos, volumes e duração da inspiração nas variáveis
medidas (KOCHI E COLS., 1988 a; 1988 b).
O tubo traqueal foi conectado a um pneumotacógrafo para pequenos animais, como
descrito por Mortola e Noworaj (MORTOLA & NOWORAJ 1983), sendo o respirador
acoplado à outra extremidade do pneumotacógrafo. O pneumotacógrafo é constituído por

51
uma cânula metálica com duas saídas laterais conectadas a um transdutor diferencial de
pressão, Validyne MP 45-2 (Engineering Corp, Northridge, CA, EUA), para medida de
fluxo aéreo e volume corrente. Através de outra saída lateral, a via aérea foi conectada a um
transdutor diferencial de pressão Validyne MP45-2 (Engineering Corp, Northridge, CA,
EUA) para medida da pressão traqueal (Ptr).
A inexistência de mudanças abruptas no diâmetro do circuito (da traquéia até a
extremidade do pneumotacógrafo) evitou erros de medida de resistência ao fluxo (LORING
& MEAD, 1982; CHANG E COLS., 1981). Os transdutores conectados ao
pneumotacógrafo e ao tubo traqueal registraram os sinais de V’ e Ptr, respectivamente.
Em seguida, os sinais foram convertidos a digitais por um conversor analógico-
digital de 12-bitz (DT-2801A, Data Translation, Malboro, MA, EUA), e amostrados a uma
freqüência de 200 Hz. Os sinais foram armazenados em microcomputador, utilizando-se o
software LABDAT (RHT-InfoData, Montreal, Canadá) e gravados em disquetes
magnéticos para posterior análise (off line), que foi realizada pelo programa ANADAT
(RHT-InfoData, Montreal, Canadá). A montagem experimental está demonstrada
esquematicamente na figura 4.
Os parâmetros da mecânica respiratória foram determinados 24 horas após a
instilação LPS ou salina. A mecânica respiratória foi obtida através da captação de 10
ciclos respiratórios, pelo método da oclusão ao final da inspiração (BATES E COLS., 1985
a e b), que permite analisar separadamente os componentes elásticos, resistivos e
viscoelástico e/ou inomogêneo do sistema respiratório.
Após a oclusão das vias aéreas ao final da inspiração, sob fluxo constante, ocorre
uma queda súbita da PL até um ponto de inflexão (Pi) a partir do qual o decaimento da

52
pressão assume caráter mais lento, atingindo um platô em sua porção terminal. Esta fase de
platô corresponde à pressão de retração elástica do pulmão (Pel). A diferença de pressão
(∆P1) que caracteriza a queda rápida inicial, representada pela diferença entre a pressão
máxima inicial (Pmax) e o ponto a partir do qual a queda se torna mais lenta (Pi),
corresponde ao componente viscoso pulmonar. A segunda variação de pressão (∆P2),
representada pela queda lenta, do Pi ao platô (Pel), reflete a pressão dissipada para vencer
os componentes viscoelásticos (“stress relaxation”) e/ou inomogêneo (“pendelluft”) do
tecido pulmonar. A soma de ∆P1 e ∆P2 fornece a variação total de pressão no pulmão
(∆Ptot). A elastância estática (Est) do pulmão pode, então, ser obtida dividindo-se Pel pelo
volume corrente (FIGURA 5).
As seguintes fórmulas foram utilizadas na análise da mecânica pulmonar:
∆P1 = Pmax – Pi
∆P2 = Pi – Pel
∆Ptot = ∆P1 + ∆P2
Est = Pel / VT
Onde:
∆P1 = variação de pressão relativa ao componente viscoso pulmonar
∆P2 = variação de pressão relativa ao componente viscoelástico e/ou inomogêneo
pulmonar
∆Ptot = variação total de pressão pulmonar
Pmax = pressão pulmonar máxima atingida

53
Pi = pressão pulmonar no ponto de inflexão
Pel = pressão de retração elástica pulmonar
Est = elastância estática do pulmão
VT = volume corrente
A resistência total do equipamento (Req), incluindo a cânula traqueal, foi
previamente aferida através da aplicação de fluxos de ar ao sistema, com concomitante
registro das variações de pressão (∆P). Uma vez que R = ∆P / V’, a resistência do
equipamento corresponde ao coeficiente angular da curva ∆PxV’. A Req, constante até
fluxos de 26 mL/s (bem acima da faixa de fluxo utilizada no presente experimento) foi de
0,12 cmH2O/mL/s. A variação de pressão determinada pelo equipamento (∆Peq = Req.V’)
foi subtraída das pressões resistivas do pulmão, de tal forma que os resultados representam
as propriedades mecânicas intrínsecas.

54
12 13 11
FIGURA 4. Montagem experimental da obtenção e coleta de dados consistindo de: 1 - Cilindro de ar comprimido. 2 - Rotâmero de agulha. 3 - Ventilador de fluxo inspiratório constante com duas válvulas solenóides. 4 - Pneumotacógrafo. 5 - Peça T para medida de pressão nas vias aéreas. 6 - Cânula traqueal. 7 - Mesa cirúrgica. 8 - Transdutor de pressão esofagiana. 9 - Transdutor de pressão traqueal. 10 - Transdutor diferencial de pressão para medida de fluxo. 11 - Scireq-24 12 - Conversor analógico-digital de 12 bits. 13 - Microcomputador.

55
∆P1 ∆P2
1,25 1,00
0,5
0
-0,5
-1,00 -1,25
0,20
0,10
0,00
PL (c
mH
2O)
Pel
Pmax Pi
FIGURA 5. Representação esquemática dos traçados de fluxo, volume, pressão em função do tempo, obtidos a partir da oclusão da via aérea ao final da inspiração.
Pmax = pressão máxima alcançada; Pi = ponto de inflexão; Pel = pressão de retração elástica e VT = volume corrente. O platô foi alcançado após uma pausa inspiratória de cinco s. Após a oclusão das vias aéreas, há uma queda rápida na PL (∆P1) que corresponde a Pmax – Pi, pressão dissipada para vencer o componente viscoso do pulmão, seguida por uma queda lenta (∆P2), pressão dissipada para vencer os componentes viscoelástico e/ou inomogêneo do pulmão, até um ponto de equilíbrio elástico, representada pela pressão de retração elástica pulmonar (Pel). A linha de base do registro de pressão corresponde à pressão positiva ao final da expiração (PEEP) de 2 cmH2O.

56
RESULTADOS

57
5 RESULTADOS
5.1 TRATAMENTO COM LPS AUMENTOU SIGNIFICATIVAMENTE EXPRESSÃO DOS RECEPTORES P2X7 NO PARÊNQUIMA PULMONAR
Com o objetivo de se caracterizar se os receptores P2X7 estão presentes no tecido
pulmonar após a indução da lesão pulmonar aguda, imunohistoquímica foi realizada
fazendo-se o uso de anticorpo policlonal para os receptores P2X7 subunidade extracelular, o
qual revelou grandes diferenças na imunorreatividade nos grupos selvagens. Os animais
selvagens controle apresentaram marcação presente para o receptor de P2X7 em todos os
tipos celulares do parênquima pulmonar, com mais freqüência em células epiteliais
alveolares, cuja densidade de reatividade foi de (14,2 ± 4,2). Já para os animais selvagens
tratados, a reatividade também foi vista nos mesmos tipos celulares marcação porém se
tornou mais proeminente devido ao maior número de células inflamatórias presentes
(FIGURAS 6 A, B). A densidade de reatividade para o receptor P2X7 foi de (33,0 ± 2,6).
Houve diferença significativa entre os dois grupos (p<0,05) (FIGURA 6 D). Os animais
nocautes tratados ou não pelo LPS não apresentaram marcação para o receptor (FIGURA 6
C).

58
FIGURA 6. Detecção e Quantificação por imunohistoquímica (densidade de superfície) dos receptores P2X7 .
(A – C) Imunohistoquímica usando-se anticorpo para os receptores P2X7. (A) Animal selvagem sem tratamento com LPS (P2X7+/+ - CTRL). A maioria das células imunoreativas estão presentes no septo alveolar , células epiteliais e inflamatórias. (B) Animal selvagem tratado com LPS (LPS- P2X7+/+). Aumento da imunoexpressão em conseqüência ao aumento do número de células inflamatórias. (C) Controle negativo- animal nocaute controle. (D) Gráfico da quantificação dos receptores P2X7 por densidade de superfície. (*p<0.05). Colunas pretas são de animais após o tratamento com LPS por 24 horas. Barras 60µm. (inserts- aumento de 650x).

59

60
5.2 ANIMAIS P2X7
-/- APRESENTARAM INFLAMAÇÃO ATENUADA E MENOS
ZONAS DE COLAPSO APÓS O TRATAMENTO COM LPS
Os animais normais que não receberam tratamento com LPS, tanto selvagens como
nocautes para o receptor P2X7, apresentaram estrutura pulmonar preservada com raros
macrófagos presentes nos espaços alveolares (FIGURA 7, A e C). Após 24 horas de
instilação com LPS os animais selvagens apresentaram uma inflamação nos septos
alveolares (alveolite) com presença de mononucleares e polimorfonucleares, enquanto que
nos animais nocautes a alveolite era menos acentuada. (FIGURA 7, B e D; TABELA 1).
Notou-se também, zonais focais de colapso do parênquima.
Os animais selvagens tratados apresentaram congestão capilar, uma discreta
hiperplasia alveolar e um aumento no número de células inflamatórias no espaço
peribronquiolar, com predominância de mononucleares. Estas alterações também foram
vistas nos animais nocautes, porém em pequena quantidade.
Para a aferição de mudanças relativas da superfície dos espaços aéreos
considerando-se que as zonas de colapso representam espaços aéreos colabados
(diminuição da superfície) foi realizada morfometria da superfície dos espaços aéreos. Não
foram consideradas zonas do parênquima pulmonar situadas externamente, ou seja, ao lado
da pleura, pois muitas vezes refletem alterações devido a coleta do pulmão.
A histomorfometria através do uso de retículo demonstrou que os animais nocautes
após o tratamento com LPS por 24 horas apresentaram um número menor de áreas
colapsadas alveolares quando comparadas aos animais selvagens após o tratamento
(p<0,05) (TABELA 1).

61
FIGURA 7. Arquitetura pulmonar dos animais estudados evidenciados pela técnica de hematoxilina e eosina.
(A ) e (C) Animais sem tratamento (CTRL), P2X7+/+ e P2X7-/ -, respectivamente. Animal P2X7+/+ com estrutura pulmonar preservada (A) enquanto que, em animal P2X7-/ - nota-se discreta inflamação no septo alveolar. (B) e (D) Animais tratados com LPS (CTRL), P2X7+/+ e P2X7-/-, respectivamente, exibindo edema dema perivascular (seta) e inflamação em septo alveolar. Notar presença de inflamação mais branda no animal nocaute (B), em relação ao WT (B). Barras: 120µm. (insertos - aumento de 400x).

62

63
O número de polimorfonucleares totais analisados (a partir dos cortes histológicos)
presentes no grupo P2X7+/+ CTRL foi de (8,8 ± 2,0), já o grupo P2X7
+/+ LPS foi de (27,0 ±
2,8). Nos animais nocautes o grupo P2X7-/- CTRL foi de (9,3 ± 1,1) e o P2X7
-/- LPS foi de
(17,6 ± 1,9) (Tabela 1).
TABELA 1. Parâmetros Morfométricos e Celularidade no Parênquima Pulmonar
Grupo Area Normal
(%)
Colapso Alveolar
(%)
PMN (%) MN (%) Celularidade Total (%)
P2X7R+/+ 91.2 ± 4.2 a 8.8 ± 2.0 a 8.8 ± 2.0 a 21.1±1.6 a 29.9±2.5 a
P2X7R-/- 87.8 ± 1.8 a,c 12.2 ± 1.8 a 9.3 ± 1.1 a 21.5±2.0 a 30.8±2.6 a
LPS-P2X7R+/+ 75.4 ± 6.6 b,c 24.6 ± 6.6 b 27.0 ± 2.8 b 12.1±1.1 b 39.0±2.6 b
LPS-P2X7R-/- 83.2 ± 1.7 c 16.8 ± 1.7 a,b 17.6 ± 1.9 c 16.1±1.5 c 33.7±1.2 a,b
Os valores estão apresentados como media (±SD) de 6 a 9 animais em cada grupo. Os dados foram obtidos em 10 campos aleatórios em cada animal. Letras diferentes indicam diferenças significantes entre os grupos (p<0.05). PMN: área de células polimorfonucleares, MN: área de células mononucleares, Celularidade Total: fração de celularidade total.

64
5.3 AUSÊNCIA DO RECEPTOR P2X7 INIBIU A SECREÇÃO DO NITRITO (NO), DA ENZIMA LACTATO DESIDROGENASE (LDH) E DA CITOCINA IL-1β NO BAL DE ANIMAIS NOCAUTESPARA O RECEPTOR P2X7 APÓS O TRATAMENTO COM LPS
A análise de nitrito, que é uma medida indireta para se chegar aos níveis de NO,
apresentou comportamento semelhante aos resultados obtidos pela análise da enzima LDH,
isto é, os animais selvagens não liberaram uma quantidade de nitrito após o tratamento com
LPS. Os animais selvagens apresentaram uma diferença significativa após o tratamento
com LPS por 24 horas em relação a todos os outros grupos (p<0.05) (4,9±0,1 (P2X7+/+
CTRL) para (5,6 ± 0,5) (P2X7+/+ LPS) e (4,6 ± 0,3) (P2X7
-/- CTRL) para (4,5 ± 0,3) (P2X7-
/- LPS) (FIGURA 8).
FIGURA 8. Quantificação de nitrito no BAL de camundongos selvagens e nocautes com e sem tratamento com LPS por 24 horas. Seguindo o mesmo padrão obtido para a análise de LDH, os níveis da liberação de nitrito em animais após o tratamento com LPS por 24 horas foi apenas observado nos animais selvagens, indicando uma atenuação da lesão nos animais nocautes. (*p<0,05 quando comparado com todos os outros grupos).
65
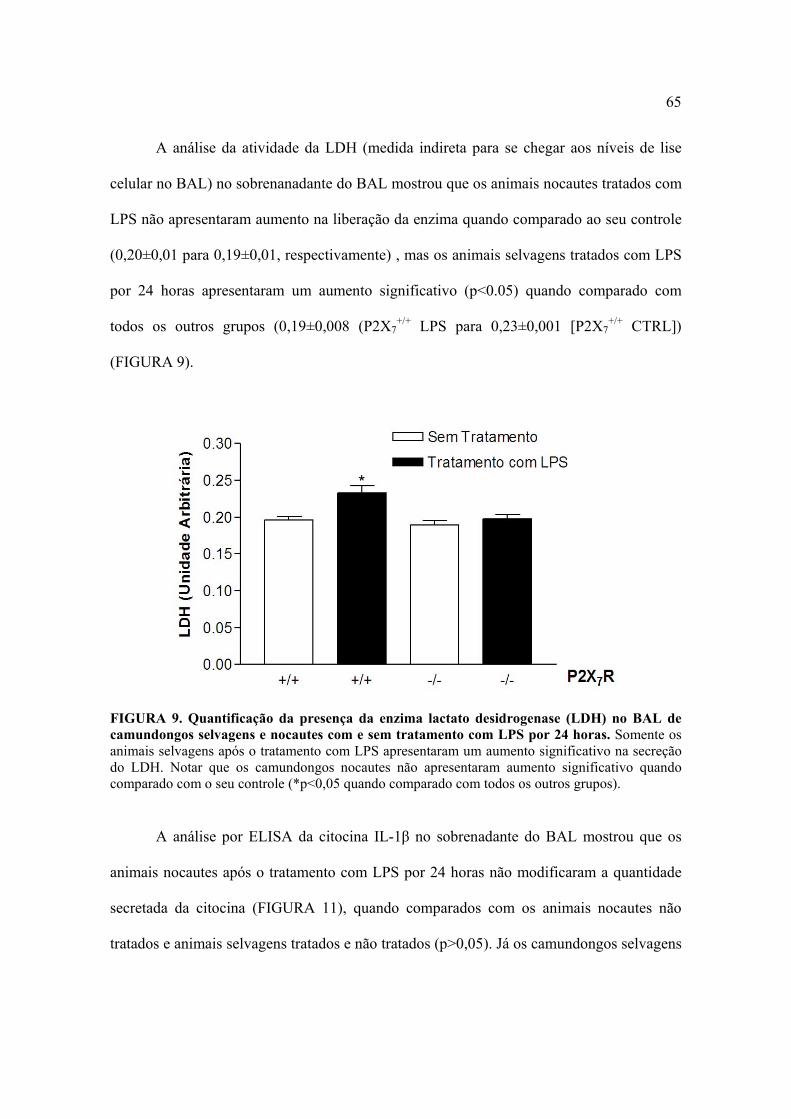
A análise da atividade da LDH (medida indireta para se chegar aos níveis de lise
celular no BAL) no sobrenanadante do BAL mostrou que os animais nocautes tratados com
LPS não apresentaram aumento na liberação da enzima quando comparado ao seu controle
(0,20±0,01 para 0,19±0,01, respectivamente) , mas os animais selvagens tratados com LPS
por 24 horas apresentaram um aumento significativo (p<0.05) quando comparado com
todos os outros grupos (0,19±0,008 (P2X7+/+ LPS para 0,23±0,001 [P2X7
+/+ CTRL])
(FIGURA 9).
FIGURA 9. Quantificação da presença da enzima lactato desidrogenase (LDH) no BAL de camundongos selvagens e nocautes com e sem tratamento com LPS por 24 horas. Somente os animais selvagens após o tratamento com LPS apresentaram um aumento significativo na secreção do LDH. Notar que os camundongos nocautes não apresentaram aumento significativo quando comparado com o seu controle (*p<0,05 quando comparado com todos os outros grupos).
A análise por ELISA da citocina IL-1β no sobrenadante do BAL mostrou que os
animais nocautes após o tratamento com LPS por 24 horas não modificaram a quantidade
secretada da citocina (FIGURA 11), quando comparados com os animais nocautes não
tratados e animais selvagens tratados e não tratados (p>0,05). Já os camundongos selvagens
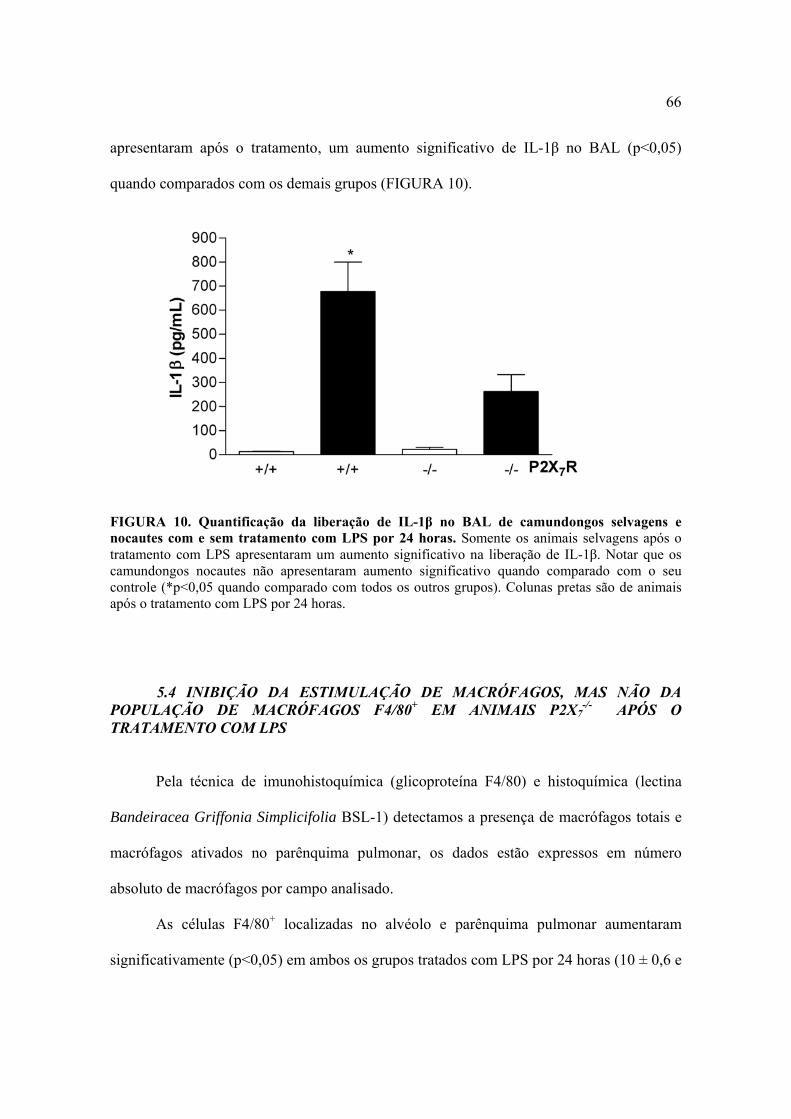
66
apresentaram após o tratamento, um aumento significativo de IL-1β no BAL (p<0,05)
quando comparados com os demais grupos (FIGURA 10).
FIGURA 10. Quantificação da liberação de IL-1β no BAL de camundongos selvagens e nocautes com e sem tratamento com LPS por 24 horas. Somente os animais selvagens após o tratamento com LPS apresentaram um aumento significativo na liberação de IL-1β. Notar que os camundongos nocautes não apresentaram aumento significativo quando comparado com o seu controle (*p<0,05 quando comparado com todos os outros grupos). Colunas pretas são de animais após o tratamento com LPS por 24 horas.
5.4 INIBIÇÃO DA ESTIMULAÇÃO DE MACRÓFAGOS, MAS NÃO DA POPULAÇÃO DE MACRÓFAGOS F4/80+ EM ANIMAIS P2X7
-/- APÓS O
TRATAMENTO COM LPS
Pela técnica de imunohistoquímica (glicoproteína F4/80) e histoquímica (lectina
Bandeiracea Griffonia Simplicifolia BSL-1) detectamos a presença de macrófagos totais e
macrófagos ativados no parênquima pulmonar, os dados estão expressos em número
absoluto de macrófagos por campo analisado.
As células F4/80+ localizadas no alvéolo e parênquima pulmonar aumentaram
significativamente (p<0,05) em ambos os grupos tratados com LPS por 24 horas (10 ± 0,6 e

67
10 ± 0,4, nos grupos P2X7+/+ LPS (FIGURA 11 B) e P2X7
-/- LPS (FIGURA 11 D),
respectivamente) em relação aos controles (6,7 ± 0,4 e 7,6 ± 0,4), nos grupos P2X7+/+
(FIGURA 11 A) e P2X7-/- (FIGURA 11 C), respectivamente) (FIGURA 11 E). Já as células
BSL-1+ aumentaram significativamente no grupo P2X7+/+ LPS (16 ± 0,8) (FIGURA 12 B)
em relação a todos os outros grupos (4,5 ± 0,3; 4,5 ± 0,4 e 4,9 ± 0,4; nos grupos P2X7+/+
CTRL (FIGURA 12 A), P2X7-/-CTRL (FIGURA 12 C) e P2X7
-/- LPS (FIGURA 12 D),
respectivamente) (p<0.001) (FIGURA 12 E).

68
FIGURA 11. Detecção imunohistoquímica e quantificação da População de Macrófagos F4/80.
(A – D) Imunohistoquímica usando o anticorpo F4/80 (macrófagos-seta). (A e C) Animais sem tratamento (CTRL), P2X7+/+ e P2X7-/-, respectivamente. Raros macrófagos positivos (setas) presentes no septo alveolar. (B e D) Animais tratados com LPS (CTRL), P2X7+/+ e P2X7-/-, respectivamente. Diversos macrófagos (setas) septais e alveolares reativos encontrados. Barras 90µm. Insertos, aumento de 450x. (E) Representação gráfica da densidade de superfície de células imunoreativas para F4/80. (*p<0.05 e **p<0.001 quando comparado com todos os outros grupos).

69

70
FIGURA 12. Caracterização e quantificação dos macrófagos pulmonares ativados pela deteção da isolectina Griffonia simplicifora.
(A – D) Deteção histoquímica da lectina BSL-1 (macrófagos ativados- seta). (A) Animal não tratado P2X7
+/+ CTRL. Os macrófagos reativos (setas) são pequenos e arredondados, presentes somente em raros septos alveolares. (B) Animal P2X7
+/+ tratado com LPS exibindo macrófagos reativos para BSL-1 (setas), com tamanho maior que os vistos no animal controle e presentes nos septos alveolares, espaços peribronquico e vascular e em espaços aéreos. (C) Animal P2X7
-/-, CTRL. Semelhante ao animal não tratado P2X7+/+ CTRL porém com um pouco
mais de células inflamatórias (setas) em septos alveolares. (D) Animal P2X7
-/- tratado com LPS. Semelhante ao animal CTRL respectivo. Barras: 90µm. Insertos, aumento de 480x. (E) Representação gráfica da densidade de superfície de células reativas para a lectina BSL-1. (*p<0.05 e **p<0.001 quando comparado com todos os outros grupos).

71

72
5.5 AUSÊNCIA DO RECRUTAMENTO DE MASTÓCITOS EM ANIMAIS P2X7-/-
APÓS O TRATAMENTO COM LPS
Com o objetivo de se caracterizar a presença dos mastócitos no parênquima
pulmonar dos animais estudados, as lâminas foram coradas com o corante alcian blue pH.
2,5 como descrito nos materiais e métodos.
Os mastócitos podem ser vistos em animais normais nas zonas de interface
da mucosa com o meio ambiente. Portanto, os animais CTRL apresentam uma pequena
quantidade de mastócitos vistos em regiões peribronquicas ou vasculares além do septo
alveolar (FIGURA 13 E).
No grupo CTRL a densidade mastocitária foi de (0,0010 ± 0,0008) e (0,0100 ± 0,0006) nos
animais P2X7+/+ e P2X7
-/- CTRL, respectivamente (FIGURA 13 A e C). No grupo LPS 24
horas o número de mastócitos/µm2 aumentou de (0,0200 ± 0,0006) para (0,0100 ± 0,0003)
nos animais P2X7+/+ e P2X7
-/- LPS, respectivamente (FIGURA 13 B e D).

73
FIGURA 13. Caracterização e quantificação dos mastócitos presentes no parênquima pulmonar pela técnica histoquímica para alcian blue pH 2.5.
(A – D). histoquímica (alcian blue pH. 2,5 – mastócitos corados em rosa (seta). (A e C) animais sem tratamento com LPS (CTRL), P2X7
+/+ e P2X7-/-, respectivamente. Barra: 100
µm. (B e D) animais tratados com LPS por 24 horas, P2X7+/+ e P2X7
-/-, respectivamente. Barras: 100 µm. (E) Representação gráfica da densidade de superfície de mastócitos (***p<0,001 quando comparado com todos os outros grupos).

74

75
5.6 AS FIBRAS ELÁSTICAS, COLÁGENAS E FIBRONECTINA NÃO SE ALTERARAM NOS ANIMAIS P2X7
-/- APÓS O TRATAMENTO COM LPS
Os animais P2X7+/+ e P2X7
-/- apresentaram rede colágena presente em torno dos
brônquios, vasos sanguíneos e em menor quantidade nos septos alveolares (FIGURAS 14 A
e B), não se notando diferenças entre a distribuição mas parecendo ser menos pronunciada
nos animais nocautes. Com o tratamento pela LPS, verificou-se um aumento desta trama,
mais visível nos animais P2X7+/+ (FIGURA 14 C). Já os animais nocautes tratados não
exibiram modificações na distribuição das fibras colágenas. Porém, quando da sua
quantificação verificou-se que os animais P2X7+/+ - LPS exibiram aumento significativo na
densidade de superfície para o colágeno. A densidade de colágeno no grupo P2X7+/+ - LPS
foi de (12,30 ± 0,80) em relação ao grupo P2X7+/+ (6,10 ± 0,20), diferença esta significante
(p<0,05) (FIGURA 14 E). No animal P2X7-/- - CTRL, o nível da densidade de colágeno
foi de (6,10 ± 0,20) passando a (9,70 ± 0,60) após o tratamento (p>0,05) (FIGURA 14 E).

76
FIGURA 14. Fibras colágenas no parênquima pulmonar evidenciadas pela coloração
de picrosirius.
(A) Os animais P2X7
+/+CTRL exibiram uma trama colágena presente em torno dos brônquios, paredes vasculares e no septo alveolar.. (B) P2X7
-/- CTRL. Aspecto semelhante ao do animal P2X7+/+ CTRL.
(C) P2X7+/+ LPS 24Hs . Trama colágena mais exuberante que o visto no CTRL.
(D) P2X7-/- LPS 24Hs. Aspecto semelhante ao dos animais P2X7
+/+CTRL. Barras: 220µm. Insertos, aumento de 480x. (E) Representação gráfica da densidade de superfície do colágeno; resultados expressos em percentagem. (***p<0.001).

77

78
As fibras elásticas, nos animais P2X7+/+ se mostraram presentes nas paredes dos
brônquios e vasos sanguíneos assim como nos septos alveolares. Os animais P2X7+/+
tratados com LPS exibiram uma diminuição da rede de fibras elásticas
Os animais P2X7-/- apresentaram a rede de fibras elásticas preservada (FIGURA 15),
não havendo mudanças na quantidade das fibras nos animais tratados em relação aos
animais não tratados com LPS. Já os animais selvagens apresentaram diminuição da rede de
fibras elásticas, uma das características da ALI, ou seja, destruição das fibras. No grupo
CTRL, a quantidade das fibras elásticas foram (14,90 ± 0,10) e (10,90 ± 0,10) nos animais
P2X7+/+ e P2X7
-/-, respectivamente (FIGURA 15). Já no grupo tratado com LPS por 24
horas as fibras elásticas aumentaram de (7,70 ± 0,30) para (10,20 ± 0,40) nos animais
P2X7+/+ e P2X7
-/-, respectivamente (FIGURA 15).
A fibronectina está presente nos septos alveolares nos animais P2X7+/+. Não houve
alteração na distribuição de fibronectina nos animais LPS-P2X7+/+. A densidade de
imunoexpressão da fibronectina nos animais LPS-P2X7-/- foi de 0,168±0,010 e é
semalhante ao do animal controle (P2X7-/-) (0,166 ± 0,010) (p>0,05). Os animais LPS-
P2X7+/+ exibiram imunoreatividade de (0,266 ± 0,013) que foi significativamente diferente
da encontrada nos animais controles (P2X7+/+), (0,172 ± 0,001) (p<0,05). (FIGURA 16).

79
FIGURA 15. Fibras elásticas no parênquima pulmonar. Evidenciadas pela coloração Orceína.
(A) Rede de fibras elásticas presente na parede de vasos sanguíneos e de forma discontínua em septos alveolares em animais P2X7
+/+ CTRL. (C) Animais P2X7
-/- CTRL – fibras elásticas presentes com a mesma distribuição da vista em animais P2X7
+/+ CTRL. (B) Rede de fibras elásticas se tornam mais proeminentes nos P2X7+/+ LPS
24Hs. (D) Animais P2X7
-/- LPS 24Hs. Barras: 220µm. Insertos, aumento de 480x. (E) Gráfico da quantificação (densidade de superfície) das fibras elásticas representados em percentagem. (*p<0.001).

80

81
+/+ +/+ -/- -/-0.0
0.1
0.2
0.3
P2X7R
* Sem TratamentoTratamento com LPS
Imun
oesp
ress
ãoFi
bron
ectin
a( µ
m2
)
FIGURA 16. Densidade de imunoexpressão da fibronectina no parênquima pulmonar. Representação gráfica da densidade de superfície da fibronectina (imunohistoquímica). (*p<0,05).
5.7 OS ANIMAIS P2X7
-/- APÓS O TRATAMENTO COM LPS APRESENTARAM
DIMINUIÇÃO DA IMUNOREATIVIDADE PARA TGF-β, AUMENTO DA IMUNOREATIVIDADE PARA MMP- 9 E AUSÊNCIA DE MODIFICAÇÃO DA QUANTIDADE DE IMUNOREATIVIDADE PARA MMP-2
A imunoreatividade para a citocina TGF-β foi evidenciada principalmente em
células inflamatórias, mas também em células de clara e células epiteliais em ambos os
animais estudados (P2X7+/+ e P2X7
-/-).
Diferenças significativas foram encontradas em todos os grupos estudados. Para os
animais selvagens foi observado um aumento de (6,0 ± 3,0 para 25,0 ± 1,1), nos grupos
P2X7+/+ CTRL e P2X7
+/+ LPS, respectivamente (p<0.001) (FIGURA 17 C). Os animais
P2X7-/- CTRL exibiram uma menor imunorreatividade para TGF-β, mas também comum

82
exibiram aumento significativo após o tratamento com o LPS. As densidades da
imunoreatividade foram de (11,0 ± 4,0 para 14,0 ± 5,0), nos grupos P2X7-/- CTRL e P2X7
-/-
LPS, respectivamente (p<0.05) (FIGURA 17 C).
Na imunohistoquímica para as metaloproteases 2 e 9 (MMP-2 e MMP-9) foram
observadas imunoreatividade presente principalmente em células mononucleares
(macrófagos) e em polimorfonucleares. As células bronquiolares também demonstraram
reatividade para as metaloproteases. A quantificação da imunoreatividade para a MMP-9
demonstrou que os animais P2X7+/+ LPS e os P2X7
-/- LPS apresentaram um aumento na
imunoexpressão (0,037 ± 0,003 e 0,037 ± 0,004, respectivamente), significativamente
diferente da dos animais controles respectivos (p<0,05) (FIGURA 17 A). Ambos os
animais após o tratamento com LPS por 24 horas apresentaram um aumento significante
quando comparados com os seus respectivos animais controles (0,021 ± 0,003 e 0,019 ±
0,002, respectivamente) (p<0,05) (FIGURA 17 A).
Já a quantificação da imuexpressão da MMP-2 demonstrou que apenas os animais
P2X7+/+ LPS (0,041± 0,004) apresentaram um aumento significativo na imunoexpressão
quando comparados ao seu controle (0,009± 0,001) (p>0,05) (FIGURA 17 B). Não houve
aumento significativo apresentados pelos grupos de animais nocautes (0,031 ± 0,002
(P2X7-/- LPS) e 0,028 ± 0,004 (P2X7
-/-)) (p<0,05) (FIGURA 17 B).

83
FIGURA 17. Quantificação das metaloproteinases 2 e 9 por imunohistoquímica e da citocina TGF-β por densidade de superfície. Gráficos da quantificação (densidade de superfície) da imunohistoquímica para MMP-9 (A), MMP-2 (B) e TGF-� (C). (*p<0.05; **p<0,001).

84
5.8 OS ANIMAIS P2X7-/-
APRESENTARAM FUNÇÃO PULMONAR NORMAL APÓS O TRATAMENTO COM LPS
A elastância estática foi aumentada de (29,0 ± 2,0) no grupo P2X7+/+ CTRL para
(58,0 ± 0,7) no grupo P2X7+/+ LPS) significativamente nos animais selvagens tratados com
LPS por 24 horas (p<0.001), fato não observado nos animais nocautes (de 29±1,4 no grupo
P2X7-/- CTRL para (36,0 ± 3,4) no grupo P2X7
-/- LPS) (FIGURA 18). Já para a análise da
elastância estática observou-se o mesmo padrão de aumento nos animais testados. Um
aumento significativo (p<0.05) foi observado nos animais selvagens tratados com LPS de
(6,0 ± 0,7) no grupo P2X7+/+ CTRL para (12,5 ± 1,2) no grupo P2X7
+/+ LPS. Nos animais
nocautes a diferença não foi significativa de (7,3 ± 0,3) no grupo P2X7-/- CTRL para (8,2 ±
0,4) no grupo P2X7-/- LPS (FIGURA 18).
O ∆P1 foi significativamente maior somente nos animais nocautes tratados com
LPS por 24 horas (p<0.05) de (0,40 ± 0,05) no grupo P2X7-/- CTRL para (0,60 ± 0,04) no
grupo P2X7-/- LPS (FIGURA 18), nos animais selvagens com o tratamento com LPS não foi
observado significância (de 0,67±0,07 no grupo P2X7+/+ CTRL para (0,61 ± 0,1) no grupo
P2X7+/+ LPS (FIGURA 18).
O ∆P2 obteve o padrão inverso, onde foi observado um aumento significativo
(p<0.001) nos animais selvagens tratados com LPS por 24 horas de (1,2 ± 0,1) no grupo
P2X7+/+ CTRL para (2,4 ± 0,2) no grupo P2X7
+/+ LPS (FIGURA 18). Nos animais
selvagens não foi observado significância no aumento de (1,4 ± 0,06) no grupo P2X7-/-
CTRL para (1,6 ± 0,1) no grupo P2X7-/- LPS (FIGURA 18).
O ∆Ptotal aumentou significativamente de (2,0 ± 0,2) para (3,0 ± 0,2) (p<0.05) nos
animais selvagens CTRL e LPS, respectivamente (FIGURA 18). Nos animais nocautes

85
aumentou de (1,8 ± 0,1) para (2,2 ± 0,1), nos grupos CTRL e LPS, respectivamente
(FIGURA 18).
FIGURA 18. Avaliação Funcional do Pulmão. Gráficos de elastância estática (A) dinâmica do pulmão (B) de pressão resistiva (C) e de pressão viscoelástica (D) e de variação de pressão total do pulmão (E). Camundongos selvagens e nocautes com e sem tratamento com LPS por 24 horas. (*p<0.05). Colunas pretas – animais após o tratamento com LPS por 24 horas, colunas brancas – animais controles.

86
DISCUSSÃO

87
6 DISCUSSÃO
Lipopolissacarídeo (LPS) é um antígeno da superfície celular de bactérias. Quando
aplicado in vivo suscita uma resposta inflamatória. Um dos primeiros eventos é a ativação
de macrófagos que, através de processo mediado por receptor leva a liberação de uma série
de citocinas incluindo o TNF-α (SU E COLS., 1995; RIETSCHEL E COLS., 1994). Esta
citocina leva ao recrutamento de neutrófilos e a infiltração no espaço aéreo (FAFFE E
COLS., 2000). A inflamação pulmonar é freqüentemente verificada no lavado
broncoalveolar constituído principalmente de células mononucleares incluindo
monócitos/macrófagos e linfócitos além de neutrófilos. É bem conhecido o fato de que há
diferenças da resposta ao LPS dependendo das cepas de camundongos (CORTELING E
COLS., 2002). No camundongo C57BL/6, após inalação intranasal, a resposta inflamatória
induzida pelo LPS é menor que a vista nos animais BALB/c. Segundo estes autores, após
24 horas, neutrófilos são as únicas células vistas no BAL, havendo uma quantidade seis
vezes menor de neutrófilos, em comparação com camundongos BALB/C (CORTELING E
COLS., 2002).
Nosso modelo animal utiliza uma dose alta de LPS (60 µg) por via intratraqueal, em
comparação com outros estudos, porém capaz de induzir um processo inflamatório
intrapulmonar após 24 horas de instilação, associado a remodelamento estrutural refletida
nas provas funcionais pulmonares.
Tendo em vista que este modelo de doença é caracterizado por um processo
inflamatório que, eventualmente leva a agressão dos tecidos, foi o modelo escolhido para a

88
verificação do envolvimento da sinalização purinérgica, mais especificamente do receptor
P2X7 na inflamação e lesão tecidual.
Pudemos evidenciar a importância dos receptores P2X7, na patogênese da lesão
induzida pelo LPS neste estudo. Através da realização de técnica imunohistoquímica para a
evidenciação do receptor P2X7, foi possível validar o modelo em estudo. Os animais
nocautes não exibiram imunoreatividade para o receptor enquanto que os animais WT
exibem imunoreatividade em células inflamatórias assim como no epitélio pulmonar como
já descrito por outros autores (CHEN E COLS., 2004; TAYLOR E COLS., 1999). Além
disso, pudemos demonstrar que o tratamento com LPS por 24 horas, leva a aumento
significativo da imunoexpressão dos receptores P2X7 nos animais selvagens. Estes dados
confirmam o papel destes receptores como sinalizadores do stress celular (SELSZNER E
COLS., 2004; HAAG E COLS., 2007). Sabe-se que após a ativação dos receptores P2X7,
este induz a liberação de diversas citocinas que influenciam a resposta imune adaptativa
(DI VIRGILIO E COLS., 2001; SOLLE E COLS., 2001). Neste trabalho os animais
nocautes apresentaram diminuição da secreção/imunoexpressão das citocinas aqui
analisadas (IL-1β e TGF- β), resultados que corroboram os dados de Solle e colaboradores
(2001). Estes autores demonstraram que a magnitude da resposta imune adaptativa em
resposta ao estímulo com LPS é diminuída em camundongos nocautes para o receptor P2X7
in vivo (SOLLE E COLS., 2001). Adicionalmente, macrófagos tratados com LPS ativam
seus receptores P2X7 (via liberação de ATP) que induz a secreção de IL-1β e diversas
outras citocinas (PELEGRIN E COLS., 2008; SOLLE E COLS., 2001).
A citocina IL-1β age recrutando e ativando populações de neutrófilos e macrófagos
que migram para o sítio de inflamação para combate-la (WARD 1994). Em adição à
locomoção, fagocitose e atividades microbicidas, macrófagos residentes e infiltrantes

89
secretam uma variedade de citocinas e quimiocinas que resultam na quimiotaxia de
leucócitos ao foco inflamatório e aumentam a resposta imune pela cooperação com outros
componentes do sistema imune (UNANUE & ALLEN, 1987). Os mecanismos moleculares
do LPS induzindo a ativação de macrófagos já tem sido extensamente investigados
(FUJIHARA E COLS., 2003). Estudos anteriores têm mostrado que os macrófagos
ativados expressam grupos carboidráticos que não são encontrados em macrófagos não
estimulados e são detectados utilizando-se lectinas (MADDOX E COLS., 1982A; 1982;
PETRYNIAK E COLS., 1986; TABOR E COLS., 1989). Para se examinar o papel dos
receptores P2X7 na infiltração e ativação macrofágica na patogênese da inflamação
pulmonar em resposta ao LPS, macrófagos foram marcados com anticorpo para a
glicoproteína F4/80 e com a lectina BSL-1 (Bandeiraea Simplicifolia Lectin I). Para a
marcação com o anticorpo F4/80, nossa hipótese era de que os macrófagos alveolares e
intersticiais poderiam ser identificados pela expressão da glicoproteína F4/80
(MCKNIGHT E COLS., 1996). A lectina BSL-1 (glicoproteína GSI) tem sido descrita
como sendo marcador direto para macrófagos ativados (MADDOX E COLS., 1982a;
1982b; PETRYNIAK E COLS., 1986; TABOR E COLS., 1989). Nós observamos que a
extensão da população de macrófagos (F4/80 positivos) não é dependente dos receptores
P2X7 em animais tratados com LPS por 24 horas. Estes dados combinados com a
histoquímica para a lectina BSL-1, demonstram haver uma importante queda na ativação de
macrófagos nos animais nocautes para o receptor P2X7 tratados com LPS quando
comparados com todos os grupos (300%) (p<0.001).
Sabe-se também que o conceito clássico da ativação macrofágica se dá pela
interação dos macrófagos com agentes inflamatórios (LPS) ou material provenientes de
necrose celular, ou ainda, citocinas. Macrófagos ativados produzem uma variedade enorme

90
de citocinas pró-inflamatórias, espécies reativas de oxigênio e de nitrogênio, proteases etc.,
os quais mantêm e perpetuam o processo inflamatório (GOERDT E COLS., 1999;
GOERDT 1999; BARNES 2000). Os macrófagos também liberam uma variedade de
mediadores antiinflamatórios, como IL-10, o qual de forma parácrina suprime a ativação
macrofágica e a produção de mediadores inflamatórios. Contudo, não podemos esquecer
que macrófagos expressam diversos receptores P2X e P2Y, incluindo P2Y1, P2Y2 e P2X7
(COUTINHO-SILVA E COLS., 2005) e estes receptores possuem papel na modulação da
ação do LPS (DI VIRGILIO E COLS., 2001). Por exemplo, o 2-methylthio-ATP (agonista
dos receptores P2X e P2Y) inibe a liberação in vivo de IL-1β e TNF- α e protege os
camundongos da morte por LPS (DI VIRGILIO E COLS., 2001; PROCTOR E COLS.,
1994; SAK & WEBB 2002). Portanto, adicionalmente aos achados deste trabalho, existem
diversas evidências que suportam a idéia de que os receptores P2X7 podem estar envolvidos
na ativação de macrófagos pelo LPS (DENLINGER E COLS., 1996; 2001; WATTERS E
COLS., 2001; HU E COLS., 1998 ).
Adicionalmente ao IL-1β, neste trabalho a citocina TGF-β se apresentou diminuída
nos animais nocautes após o tratamento com LPS quando comparados com os animais
selvagens após o tratamento. Várias são as fontes de TGF-β após uma agressão. TGF-α é
secretado por uma série de células, inclusive as células inflamatórias como macrófagos,
mastócitos e células epiteliais pulmonares (MALIZIA E COLS., 2008; HOSTTLER E
COLS., 2008; MALAVIA E COLS., 2008). Sabe-se que a ativação dos receptores P2X7
leva ao aumento da expressão do RNAm de TGF-β, efeito este dependente da via de
signalização PKC/MAPK, aspecto visto em astrócitos (WANG E COLS., 2003).
A diminuição junto com a IL-1β pode ter sido mais um estímulo à atenuação de
praticamente todos os parâmetros inflamatórios analisados à estarem atenuados, porque

91
sabe-se que o TGF-β medeia a quimiotaxia de diferentes tipos de células inflamatórias,
como, monócitos, macrófagos, mastócitos e linfócitos T (WAHL E COLS., 1987; ALLEN
E COLS., 1990; ADAMS E COLS., 1991; GRUBER E COLS., 1994). Podendo ainda,
estimular a expressão de moléculas de adesão célula-célula como a molécula-1 de adesão
intercelular (ICAM-1) nas células endoteliais (SUZUKI E COLS., 1994). TGF-β pode
também induzir a expressão de citocinas como o próprio IL-1β por monócitos e contribuir
assim para o processo inflamatório (WAHL E COLS., 1987; BOMBARA & IGNOTZ
1992). Contudo o TGF-β desempenha um papel crítico no desenvolvimento da inflamação
e fibrose pulmonar após a ALI (GIRI E COLS., 1993; PITTET E COLS., 2001). Portanto,
existem bastantes evidências de que o TGF-β está envolvido na inflamação. Finalmente, já
foi mostrado em cultura de astrócitos que os receptores P2X7 medeiam a secreção de TGF-
β1 (WANG E COLS., 2003).
A população de mastócitos nos animais nocautes após o tratamento com LPS não
foi recrutada. Estes são conhecidos como sendo uma das maiores fontes de citocinas e
fatores de crescimento e têm sido implicados em várias desordens fibróticas e inflamatórias
(WASSERMAN 1980). Assim, nossos dados sugerem que o receptor P2X7 além de induzir
ativação e degranulação em mastócito (BULANOVA E COLS., 2005; MYRTEK &
IDZKO 2007; SCHULMAN E COLS., 1999; LEE E COLS., 2001; JAFFAR & PEARCE
1990) está também, de alguma forma, associado com a migração dos mesmos em sítios
inflamatórios.
Tem sido verificado que os receptores para nucleotídeos tem sido implicados na
modulação da produção de superóxidos, peróxido de hidrogênio e outras espécies reativas
de oxigênio por diversas células do sistema imune (WARD E COLS., 1988; KUHNS E
COLS., 1988; KUROKI &. MINAKAMI 1989; SEIFERT E COLS., 1989; SUH E COLS.,

92
2001; MURPHY E COLS., 1993; DICHMANN E COLS., 2000; FERRARI E COLS.,
2000; PARVATHENANI E COLS., 2003; GUERRA E COLS., 2007). Com respeito à
sinalização dos receptores P2X7, ja é bastante conhecido sobre o envolvimento de ROS
nestes processos (NOGUCHI E COLS., 2008; TSAI E COLS., 2007; MASON E COLS.,
2004; CRUZ E COLS., 2007; KIM E COLS., 2007) , como também foi descrito que a
ativação do NFkB pelo P2X7 é sensível a antioxidantes (FERRARI E COLS., 1997),
sugerindo que ROS contribuem para estes processos.
O LPS induz a geração de espécies reativas de oxigênio (ROS) que é uma das
maiores causas da citotoxicidade. ROS pode causar a ativação de fatores de transcrição
sensitivos à redução como o NF-κB e AP-1 o qual transcrevem diversos genes
inflamatórios (BHATTACHARYY E COLS., 2004). No pulmão, estudos prévios
mostraram que a iNOS é aumentada durante a indução de lesão pulmonar induzido pelo
LPS e que a superprodução de NO via esta enzima é em detrimento da função pulmonar
(NUMATA E COLS., 1998). A presença de NO, a partir do lavado broncoalveolar, foi
quantificada de modo indireto pela presença de nitrito (AMBER E COLS., 1988; TSAI E
COLS., 1999). Os animais selvagens tratados com LPS liberaram uma quantidade
significativa de nitrito, fato que não ocorreu com os animais nocautes. Ou seja, nos animais
nocautes, o LPS induz uma lesão mais branda. Também podemos supor que o edema
peribronquiolar mais leve ou ausente nestes animais é conseqüente a inibição da liberação
de NO que finalmente reduz a inflamação como um todo. Neste contexto, foi observado
que na ausência do receptor P2X7, a inflamação e o edema foram reduzidos.
Adicionalmente, na ocorrência da inflamação é sabido que, na presença de uma resposta
inflamatória exuberante, a capacidade microbicida das células inflamatórias que migraram
para o foco de lesão é suprimida pela alta produção de óxido nítrico (CROSARA-

93
ALBERTO E COLS., 2002; BENJAMIN E COLS., 2002; 2000). Ou seja, esse óxido
nítrico inicialmente é liberado em quantidades fisiológicas, mas quando sua produção é
exageradamente aumentada pela ativação da sintase induzível do óxido nítrico (iNOS) ele
passa então, a inibir a atividade microbicida das células inflamatórias, principalmente dos
neutrófilos (CROSARA-ALBERTO E COLS., 2002; BENJAMIN E COLS., 2002;2000).
Estes resultados também podem ser relacionados com a idéia de que macrófagos RAW
264.7O pré-tratados com antagonistas do receptor P2X7 adenosina 5’trifosfato oxidado
(oATP) inibe a produção de NO e a expressão de iNOS e atenua a ativação de NFkB e
ERK-1,2 após a estimulação com LPS (HU E COLS., 1998). Estes receptores podem
também como citado anteriormente modular a produção de TNF-α e IL-1β e NO
(GUERRA E COLS., 2003; PFEIFFER E COLS., 2007).
Quanto ao remodelamento pulmonar após o tratamento com LPS pudemos
demonstrar que os animais selvagens apresentaram diminuição da trama de fibras elásticas
e aumento da rede colágena e da fibronectina. Estas alterações estão relacionadas ao
aumento progressivo da lesão pulmonar (JANOFF,1985; NEGRI E COLS., 2002; ORITO
E COLS., 2006; POSTMA & TIMENS 2006). Sabe-se que com a degradação das fibras
elásticas formam-se fragmentos de elastina que podem recrutar uma segunda população de
células inflamatórias ao pulmão, levando assim ao aumento da lesão pulmonar,
principalmente pelo recrutamento de macrófagos, que por sua vez liberam mais fatores
inflamatórios e enzimas proteolíticas (HAUTAMAKI E COLS., 1997). Em adição também
existe a liberação de espécies reativas (estresse oxidativo) e a apoptose de células
estruturais pulmonares (MACNEE 2005; OWEN 2005).

94
O remodelamento pulmonar nos animais nocautes após o tratamento com LPS
demonstrou-se inalterado quando comparado aos seus controles, havendo uma preservação
das trama colágena e elástica bem como a manutenção da fibronectina no tecido pulmonar.
Possivelmente estes dados expliquem os dados normais da função pulmonar (mecânica
respiratória) neste grupo.
Além disso, foi possível demonstrar que os animais nocautes apresentam uma trama
elástica menor que a vista nos animais selvagens e uma trama colágena maior que a dos
grupos controle selvagem (p<0,05). Estes dados ainda não tinham sido verificados nestes
animais podendo estar modificando parâmetros fisiológicos referentes a arquitetura
pulmonar normal. Provavelmente estes dados refletem a baixa expressão da citocina TGF-β
verificada em nosso estudo. É bem conhecido o fato que esta citocina é considerada como
prófibrótica (WILLIS & BOROK 2007; DANIELS E COLS., 2004; YEHUALAESHET E
COLS., 2000; ROMAN E COLS., 2005), como visto em diversos modelos experimentais
(SHEN E COLS., 2008; CUTRONEO E COLS., 2007; KITOWSKA E COLS., 2008;
VYAS-READ E COLS., 2007; XISTO E COLS., 2005).
Os efeitos verificados em relação as proteínas da MEC podem estar correlacionados
com a expressão de metaloproteases. Já foi verificado que as MMPs 2 e 9 estão implicadas
no remodelamento pulmonar após tratamento com LPS (COIMBRA E COLS., 2006;
CARNEY E COLS., 2001). Tanto o LPS quanto as citocinas próinflamatórias aumentam a
síntese de MMP-2 e a MMP-9 (gelatinases) por uma série de células inflamatórias.
Também se encontram elevadas na septicemia e ARD humanas (NAKAMURA E COLS.,
1998; CORBEL E COLS., 2000) assim como em animais (COIMBRA E COLS., 2006)

95
Nossos resultados estão condizentes com os dados da literatura. As MMP-2 e -9
estão aumentados após introdução de LPS sendo que a ausência de modulação da MMP-2
pode ser devida a sua regulação pelo TGF-β (LECHAPT-ZALCMAN E COLS., 2006).
Além disso, podemos explicar pela imunoexpressão da MMP-2 que nos animais
nocautes apresentou-se inalterada após o tratamento com LPS por 24 horas, indicando uma
possível mudança na interação célula-célula e célula-matriz que finalmente afeta a
migração e ativação celular (PARKS & SHAPIRO 2001). Ou possivelmente pela
diminuição do seqüestro de polimorfonucleares para dentro do interstício (CORRY E
COLS., 2002). Apesar da MMP-9 ter se mantido aumentada nos animais nocautes após o
tratamento com LPS e estar envolvida em diversos processos inflamatórios pulmonares ,
incluindo a ALI (WINKLER E COLS., 2003; SANTOS E COLS., 2006; RICOU E COLS.,
1996; RENCKENS E COLS., 2006), estes conseguiram manter uma atenuação da
inflamação pelos diversos parâmetros inflamatórios analisados que demonstraram-se
inalterados após o tratamento com LPS.
A mecânica pulmonar foi analisada pelo método da oclusão ao final da inspiração
(BATES E COLS., 1985; 1988). Após a oclusão ocorre uma queda rápida da pressão
traqueal até um ponto de inflexão, e a partir daí ocorre uma queda lenta que decorre até um
platô, o qual equivale à pressão de retração elástica do pulmão. A partir desses pontos de
pressão puderam-se calcular as variáveis estudadas no presente trabalho. A ∆Ptot, L,
correspondente à variação total de pressão pulmonar, refletindo as modificações nos
componentes resistivos e viscoelásticos do pulmão como um todo. Os animais selvagens
apresentaram um aumento do ∆Ptot após o tratamento com LPS por 24 horas (p<0.05), fato
não observado nos animais nocautes. A ∆P1, L, correspondente à dissipação de energia no

96
componente viscoso do pulmão, ou seja, a variação de pressão necessária para vencer o
componente viscoso das vias aéreas centrais. A ∆P1 nos animais selvagens CTRL e
tratados com LPS não obtiveram diferença significativa, estando de acordo com trabalhos
anteriores (RACKOW E COLS., 1988; FERRARI-BALIVIERA E COLS., 1989; PIPER E
COLS., 1996). Fato oposto foi observado com os animais nocautes tratados com LPS
quando comparados aos animais CTRL. Isto pode ser explicado pelo fato de que os animais
nocautes tratados com LPS por 24 horas apresentaram uma média do ∆P1 no mesmo
patamar dos animais selvagens CTRL e nos animais tratados com LPS. Contudo, este
aumento caracterizado nos animais nocautes pode ser explicado por alterações de fase
aguda que ocorre nos primeiros momentos da lesão por LPS (MENEZES E COLS., 2005).
Podemos, também relacionar estes dados à redução das vias aéreas em função do colapso
alveolar. Os componentes elásticos do pulmão ou ∆P2, L, reflete a variação de pressão
necessária para vencer os componentes viscoelásticos do pulmão nas vias aéreas
periféricas, traduzindo condições do parênquima pulmonar. O ∆P2 dos animais selvagens
de acordo com os achados histológicos apresentaram com tratamento com LPS por 24
horas, um aumento significativo (p<0.01) na sua variação. Corroborando com a piora geral
dos camundongos selvagens em relação aos camundongos nocautes, pois este componente
pulmonar tem relação com a estrutura pulmonar, a qual foi afetada. A elastância estática do
pulmão ou EST, L reflete a rigidez pulmonar e representa o inverso da complacência, ou
seja, mede a capacidade de distensão do pulmão. Este parâmetro mostrou que os animais
nocautes mesmo com o tratamento com o LPS não obtiveram diferença significativa na
EST e na DE (elastância dinâmica), fato não observado nos animais selvagens.

97

98
CONCLUSÕES

99
7 CONCLUSÕES
1- O LPS, após 24 horas, por instilação intratraqueal, induziu um processo inflamatório
pulmonar predominantemente do tipo mononuclear e remodelamento pulmonar.
2- A imunoexpressão do receptor P2X7 está aumentada significativamente no pulmão, na
lesão induzida pelo LPS, o que está de acordo com o conceito de ser um sinalizador de
estresse celular.
3- O receptor P2X7 parece estar envolvido no recrutamento de neutrófilos e mastócitos, mas
não no de macrófagos F4/80+ no modelo da lesão pulmonar induzida pelo LPS.
4- O receptor P2X7 parece estar envolvido na ativação de macrófagos na lesão aguda
pulmonar induzida pelo LPS.
5- A atenuação da lesão pulmonar induzida pela ausência do receptor P2X7 provavelmente
se deve não só à ausência do NO (menor estresse oxidativo), como também a menor
produção de fatores pró-inflamatórios, como o IL-1 e prófibróticos, como o TGF-β.
6- A atenuação da lesão pulmonar induzida pela ausência do receptor P2X7 também está
relacionada a diminuição/inibição do remodelamento pulmonar (tramas colágena e elástica
mantida, ausência de deposição de fibronectina e ausência de modulação de MMP-2).
7- A diminuição/inibição do remodelamento pulmonar pode ter sido o responsável pela
manutenção dos parâmetros funcionais em níveis normais nos animais nocautes.
8- Concluímos com um resumo esquemático de todos os resultados apresentados aqui.

100
FIGURA 19. Esquema mostrando resumo dos resultados obtidos na dissertação.

101
REFERÊNCIAS

102
8 REFERÊNCIAS ADAMS, D.H., HATHAWAY, M., SHAW, J., BURNETT, D., ELIAS, E. & STRAIN,
A.J. (1991). Transforming growth factor-beta induces human T lymphocyte
migration in vitro. J Immunol, 147, 609-12.
ADAMSON, I.Y. & BOWDEN, D.H. (1974). The pathogenesis of bloemycin-induced
pulmonary fibrosis in mice. Am J Pathol, 77, 185-97.
ADAMSON, I.Y., HEDGECOCK, C. & BOWDEN, D.H. (1990). Epithelial cell-fibroblast
interactions in lung injury and repair. Am J Pathol, 137, 385-92.
ALEXOPOULOU, L., HOLT, A.C., MEDZHITOV, R. & FLAVELL, R.A. (2001).
Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like
receptor 3. Nature, 413, 732-8.
ALLEN, J.B., MANTHEY, C.L., HAND, A.R., OHURA, K., ELLINGSWORTH, L. &
WAHL, S.M. (1990). Rapid onset synovial inflammation and hyperplasia induced
by transforming growth factor beta. J Exp Med, 171, 231-47.
AMBER, I.J., HIBBS, J.B., JR., TAINTOR, R.R. & VAVRIN, Z. (1988). Cytokines induce
an L-arginine-dependent effector system in nonmacrophage cells. J Leukoc Biol, 44,
58-65.
ARROLIGA, A.C., GHAMRA, Z.W., PEREZ TREPICHIO, A., PEREZ TREPICHIO, P.,
KOMARA, J.J., JR., SMITH, A. & WIEDEMANN, H.P. (2002). Incidence of
ARDS in an adult population of northeast Ohio. Chest, 121, 1972-6.
ASHBAUGH, D.G., BIGELOW, D.B., PETTY, T.L. & LEVINE, B.E. (1967). Acute
respiratory distress in adults. Lancet, 2, 319-23.
ATABAI, K. & MATTHAY, M.A. (2002). The pulmonary physician in critical care. 5:

103
Acute lung injury and the acute respiratory distress syndrome: definitions and
epidemiology. Thorax, 57, 452-8.
BARNES, P.J. (2000). Chronic obstructive pulmonary disease. N Engl J Med, 343, 269-80.
BATES, J.H., BACONNIER, P. & MILIC-EMILI, J. (1988). A theoretical analysis of
interrupter technique for measuring respiratory mechanics. J Appl Physiol, 64,
2204-14.
BATES, J.H., ROSSI, A. & MILIC-EMILI, J. (1985). Analysis of the behavior of the
respiratory system with constant inspiratory flow. J Appl Physiol, 58, 1840-8.
BECKER, S., DEVLIN, R.B. & HASKILL, J.S. (1989). Differential production of tumor
necrosis factor, macrophage colony stimulating factor, and interleukin 1 by human
alveolar macrophages. J Leukoc Biol, 45, 353-61.
BENJAMIM, C.F., FERREIRA, S.H. & CUNHA, F.Q. (2000). Role of nitric oxide in the
failure of neutrophil migration in sepsis. J Infect Dis, 182, 214-23.
BENJAMIM, C.F., SILVA, J.S., FORTES, Z.B., OLIVEIRA, M.A., FERREIRA, S.H. &
CUNHA, F.Q. (2002). Inhibition of leukocyte rolling by nitric oxide during sepsis
leads to reduced migration of active microbicidal neutrophils. Infect Immun, 70,
3602-10.
BERNARD, G.R., ARTIGAS, A., BRIGHAM, K.L., CARLET, J., FALKE, K., HUDSON,
L., LAMY, M., LEGALL, J.R., MORRIS, A. & SPRAGG, R. (1994). Report of the
American-European Consensus conference on acute respiratory distress syndrome:
definitions, mechanisms, relevant outcomes, and clinical trial coordination.
Consensus Committee. J Crit Care, 9, 72-81.
BERSTEN, A.D., EDIBAM, C., HUNT, T. & MORAN, J. (2002). Incidence and mortality
of acute lung injury and the acute respiratory distress syndrome in three Australian

104
States. Am J Respir Crit Care Med, 165, 443-8.
BEUTLER, B. (2004). Inferences, questions and possibilities in Toll-like receptor
signalling. Nature, 430, 257-63.
BHATTACHARYYA, J., BISWAS, S. & DATTA, A.G. (2004). Mode of action of
endotoxin: role of free radicals and antioxidants. Curr Med Chem, 11, 359-68.
BOMBARA, C. & IGNOTZ, R.A. (1992). TGF-beta inhibits proliferation of and promotes
differentiation of human promonocytic leukemia cells. J Cell Physiol, 153, 30-7.
BOURS, M.J., SWENNEN, E.L., DI VIRGILIO, F., CRONSTEIN, B.N. & DAGNELIE,
P.C. (2006). Adenosine 5'-triphosphate and adenosine as endogenous signaling
molecules in immunity and inflammation. Pharmacol Ther, 112, 358-404.
BOWDEN, D.H. (1987). Macrophages, dust, and pulmonary diseases. Exp Lung Res, 12,
89-107.
BRASS, D.M., SAVOV, J.D., GAVETT, S.H., HAYKAL-COATES, N. & SCHWARTZ,
D.A. (2003). Subchronic endotoxin inhalation causes persistent airway disease. Am
J Physiol Lung Cell Mol Physiol, 285, L755-61.
BRIGHAM, K.L. & MEYRICK, B. (1986). Endotoxin and lung injury. Am Rev Respir Dis,
133, 913-27.
BULANOVA, E., BUDAGIAN, V., ORINSKA, Z., HEIN, M., PETERSEN, F., THON, L.,
ADAM, D. & BULFONE-PAUS, S. (2005). Extracellular ATP induces cytokine
expression and apoptosis through P2X7 receptor in murine mast cells. J Immunol,
174, 3880-90.
BULANOVA, E., BUDAGIAN, V., ORINSKA, Z., HEIN, M., PETERSEN, F., THON, L.,
ADAM, D. & BULFONE-PAUS, S. (2005). Extracellular ATP induces cytokine
expression and apoptosis through P2X7 receptor in murine mast cells. J Immunol,

105
174, 3880-90.
BURNSTOCK, G. (2002). Potential therapeutic targets in the rapidly expanding field of
purinergic signalling. Clin Med, 2, 45-53.
BURNSTOCK, G. (2006). Pathophysiology and therapeutic potential of purinergic
signaling. Pharmacol Rev, 58, 58-86.
BURNSTOCK, G. & KNIGHT, G.E. (2004). Cellular distribution and functions of P2
receptor subtypes in different systems. Int Rev Cytol, 240, 31-304.
CAMPOS, M.A., ALMEIDA, I.C., TAKEUCHI, O., AKIRA, S., VALENTE, E.P.,
PROCOPIO, D.O., TRAVASSOS, L.R., SMITH, J.A., GOLENBOCK, D.T. &
GAZZINELLI, R.T. (2001). Activation of Toll-like receptor-2 by
glycosylphosphatidylinositol anchors from a protozoan parasite. J Immunol, 167,
416-23.
CARNEY, D.E., MCCANN, U.G., SCHILLER, H.J., GATTO, L.A., STEINBERG, J.,
PICONE, A.L. & NIEMAN, G.F. (2001). Metalloproteinase inhibition prevents
acute respiratory distress syndrome. J Surg Res, 99, 245-52.
CHANG, H.K., DELAUNOIS, L., BOILEAU, R. & MARTIN, R.R. (1981). Redistribution
of pulmonary blood flow during experimental air embolism. J Appl Physiol, 51,
211-7.
CHEN, L. & BROSNAN, C.F. (2006). Regulation of Immune Response by P2X7 Receptor.
Crit Rev Immunol, 26, 499-513.
CHEN, Z., JIN, N., NARASARAJU, T., CHEN, J., MCFARLAND, L.R., SCOTT, M. &
LIU, L. (2004). Identification of two novel markers for alveolar epithelial type I and
II cells. Biochem Biophys Res Commun, 319, 774-80.
CHOW, S.C., KASS, G.E. & ORRENIUS, S. (1997). Purines and their roles in apoptosis.

106
Neuropharmacology, 36, 1149-56.
CHUANG, T. & ULEVITCH, R.J. (2001). Identification of hTLR10: a novel human Toll-
like receptor preferentially expressed in immune cells. Biochim Biophys Acta, 1518,
157-61.
CHUANG, T.H. & ULEVITCH, R.J. (2000). Cloning and characterization of a sub-family
of human toll-like receptors: hTLR7, hTLR8 and hTLR9. Eur Cytokine Netw, 11,
372-8.
CHUNG, K.F. (2001). Cytokines in chronic obstructive pulmonary disease. Eur Respir J
Suppl, 34, 50s-59s.
COCKCROFT, S. & GOMPERTS, B.D. (1979). Activation and inhibition of calcium-
dependent histamine secretion by ATP ions applied to rat mast cells. J Physiol, 296,
229-43.
COIMBRA, R., MELBOSTAD, H., LOOMIS, W., PORCIDES, R.D., WOLF, P., TOBAR,
M. & HOYT, D.B. (2006). LPS-induced acute lung injury is attenuated by
phosphodiesterase inhibition: effects on proinflammatory mediators,
metalloproteinases, NF-kappaB, and ICAM-1 expression. J Trauma, 60, 115-25.
CORBEL, M., BOICHOT, E. & LAGENTE, V. (2000). Role of gelatinases MMP-2 and
MMP-9 in tissue remodeling following acute lung injury. Braz J Med Biol Res, 33,
749-54.
CORRY, D.B., RISHI, K., KANELLIS, J., KISS, A., SONG LZ, L.Z., XU, J., FENG, L.,
WERB, Z. & KHERADMAND, F. (2002). Decreased allergic lung inflammatory
cell egression and increased susceptibility to asphyxiation in MMP2-deficiency. Nat
Immunol, 3, 347-53.
CORTELING, R., WYSS, D. & TRIFILIEFF, A. (2002). In vivo models of lung neutrophil

107
activation. Comparison of mice and hamsters. BMC Pharmacol, 2, 1.
COUTINHO-SILVA, R., ALVES, L.A., DE CARVALHO, A.C., SAVINO, W. &
PERSECHINI, P.M. (1996). Characterization of P2Z purinergic receptors on
phagocytic cells of the thymic reticulum in culture. Biochim Biophys Acta, 1280,
217-22.
COUTINHO-SILVA, R., ALVES, L.A., SAVINO, W. & PERSECHINI, P.M. (1996). A
cation non-selective channel induced by extracellular ATP in macrophages and
phagocytic cells of the thymic reticulum. Biochim Biophys Acta, 1278, 125-30.
COUTINHO-SILVA, R., OJCIUS, D.M., GORECKI, D.C., PERSECHINI, P.M.,
BISAGGIO, R.C., MENDES, A.N., MARKS, J., BURNSTOCK, G. & DUNN,
P.M. (2005). Multiple P2X and P2Y receptor subtypes in mouse J774, spleen and
peritoneal macrophages. Biochem Pharmacol, 69, 641-55.
COUTINHO-SILVA, R., PERFETTINI, J.L., PERSECHINI, P.M., DAUTRY-VARSAT,
A. & OJCIUS, D.M. (2001). Modulation of P2Z/P2X(7) receptor activity in
macrophages infected with Chlamydia psittaci. Am J Physiol Cell Physiol, 280,
C81-9.
COUTINHO-SILVA, R., PERSECHINI, P.M., BISAGGIO, R.D., PERFETTINI, J.L.,
NETO, A.C., KANELLOPOULOS, J.M., MOTTA-LY, I., DAUTRY-VARSAT, A.
& OJCIUS, D.M. (1999). P2Z/P2X7 receptor-dependent apoptosis of dendritic
cells. Am J Physiol, 276, C1139-47.
COUTINHO-SILVA, R., STAHL, L., RAYMOND, M.N., JUNGAS, T., VERBEKE, P.,
BURNSTOCK, G., DARVILLE, T. & OJCIUS, D.M. (2003). Inhibition of
chlamydial infectious activity due to P2X7R-dependent phospholipase D activation.
Immunity, 19, 403-12.

108
CROSARA-ALBERTO, D.P., DARINI, A.L., INOUE, R.Y., SILVA, J.S., FERREIRA,
S.H. & CUNHA, F.Q. (2002). Involvement of NO in the failure of neutrophil
migration in sepsis induced by Staphylococcus aureus. Br J Pharmacol, 136, 645-
58.
CRUZ, C.M., RINNA, A., FORMAN, H.J., VENTURA, A.L., PERSECHINI, P.M. &
OJCIUS, D.M. (2007). ATP activates a reactive oxygen species-dependent
oxidative stress response and secretion of proinflammatory cytokines in
macrophages. J Biol Chem, 282, 2871-9.
CUTRONEO, K.R., WHITE, S.L., PHAN, S.H. & EHRLICH, H.P. (2007). Therapies for
bleomycin induced lung fibrosis through regulation of TGF-beta1 induced collagen
gene expression. J Cell Physiol, 211, 585-9.
DANIELS, C.E., WILKES, M.C., EDENS, M., KOTTOM, T.J., MURPHY, S.J., LIMPER,
A.H. & LEOF, E.B. (2004). Imatinib mesylate inhibits the profibrogenic activity of
TGF-beta and prevents bleomycin-mediated lung fibrosis. J Clin Invest, 114, 1308-
16.
DARVILLE, T., WELTER-STAHL, L., CRUZ, C., SATER, A.A., ANDREWS, C.W., JR.
& OJCIUS, D.M. (2007). Effect of the purinergic receptor P2X7 on Chlamydia
infection in cervical epithelial cells and vaginally infected mice. J Immunol, 179,
3707-14.
DAVIS, C.W. (2002). Regulation of mucin secretion from in vitro cellular models.
Novartis Found Symp, 248, 113-25; discussion 125-31, 277-82.
DENLINGER, L.C., FISETTE, P.L., GARIS, K.A., KWON, G., VAZQUEZ-TORRES, A.,
SIMON, A.D., NGUYEN, B., PROCTOR, R.A., BERTICS, P.J. & CORBETT, J.A.
(1996). Regulation of inducible nitric oxide synthase expression by macrophage

109
purinoreceptors and calcium. J Biol Chem, 271, 337-42.
DENLINGER, L.C., FISETTE, P.L., SOMMER, J.A., WATTERS, J.J., PRABHU, U.,
DUBYAK, G.R., PROCTOR, R.A. & BERTICS, P.J. (2001). Cutting edge: the
nucleotide receptor P2X7 contains multiple protein- and lipid-interaction motifs
including a potential binding site for bacterial lipopolysaccharide. J Immunol, 167,
1871-6.
DENLINGER, L.C., GARIS, K.A., SOMMER, J.A., GUADARRAMA, A.G., PROCTOR,
R.A. & BERTICS, P.J. (1998). Nuclear translocation of NF-kappaB in
lipopolysaccharide-treated macrophages fails to correspond to endotoxicity:
evidence suggesting a requirement for a gamma interferon-like signal. Infect
Immun, 66, 1638-47.
DI VIRGILIO, F. (1995). The P2Z purinoceptor: an intriguing role in immunity,
inflammation and cell death. Immunol Today, 16, 524-8.
DI VIRGILIO, F., CHIOZZI, P., FERRARI, D., FALZONI, S., SANZ, J.M., MORELLI,
A., TORBOLI, M., BOLOGNESI, G. & BARICORDI, O.R. (2001). Nucleotide
receptors: an emerging family of regulatory molecules in blood cells. Blood, 97,
587-600.
DICHMANN, S., IDZKO, M., ZIMPFER, U., HOFMANN, C., FERRARI, D.,
LUTTMANN, W., VIRCHOW, C., JR., DI VIRGILIO, F. & NORGAUER, J.
(2000). Adenosine triphosphate-induced oxygen radical production and CD11b up-
regulation: Ca(++) mobilization and actin reorganization in human eosinophils.
Blood, 95, 973-8.
DOLBER, P.C. & SPACH, M.S. (1993). Conventional and confocal fluorescence
microscopy of collagen fibers in the heart. J Histochem Cytochem, 41, 465-9.

110
FAFFE, D.S., SEIDL, V.R., CHAGAS, P.S., GONCALVES DE MORAES, V.L.,
CAPELOZZI, V.L., ROCCO, P.R. & ZIN, W.A. (2000). Respiratory effects of
lipopolysaccharide-induced inflammatory lung injury in mice. Eur Respir J, 15, 85-
91.
FEHRENBACH, H. (2001). Alveolar epithelial type II cell: defender of the alveolus
revisited. Respir Res, 2, 33-46.
FERRARI, D., CHIOZZI, P., FALZONI, S., HANAU, S. & DI VIRGILIO, F. (1997).
Purinergic modulation of interleukin-1 beta release from microglial cells stimulated
with bacterial endotoxin. J Exp Med, 185, 579-82.
FERRARI, D., IDZKO, M., DICHMANN, S., PURLIS, D., VIRCHOW, C., NORGAUER,
J., CHIOZZI, P., DI VIRGILIO, F. & LUTTMANN, W. (2000). P2 purinergic
receptors of human eosinophils: characterization and coupling to oxygen radical
production. FEBS Lett, 486, 217-24.
FERRARI, D., PIZZIRANI, C., ADINOLFI, E., LEMOLI, R.M., CURTI, A., IDZKO, M.,
PANTHER, E. & DI VIRGILIO, F. (2006). The P2X7 receptor: a key player in IL-1
processing and release. J Immunol, 176, 3877-83.
FERRARI, D., WESSELBORG, S., BAUER, M.K. & SCHULZE-OSTHOFF, K. (1997).
Extracellular ATP activates transcription factor NF-kappaB through the P2Z
purinoreceptor by selectively targeting NF-kappaB p65. J Cell Biol, 139, 1635-43.
FERRARI-BALIVIERA, E., MEALY, K., SMITH, R.J. & WILMORE, D.W. (1989).
Tumor necrosis factor induces adult respiratory distress syndrome in rats. Arch
Surg, 124, 1400-5.
FIALKOW, L., CHAN, C.K. & DOWNEY, G.P. (1997). Inhibition of CD45 during
neutrophil activation. J Immunol, 158, 5409-17.

111
FIALKOW, L., CHAN, C.K., GRINSTEIN, S. & DOWNEY, G.P. (1993). Regulation of
tyrosine phosphorylation in neutrophils by the NADPH oxidase. Role of reactive
oxygen intermediates. J Biol Chem, 268, 17131-7.
FIALKOW, L., CHAN, C.K., ROTIN, D., GRINSTEIN, S. & DOWNEY, G.P. (1994).
Activation of the mitogen-activated protein kinase signaling pathway in neutrophils.
Role of oxidants. J Biol Chem, 269, 31234-42.
FINK, M.P. (2002). Role of reactive oxygen and nitrogen species in acute respiratory
distress syndrome. Curr Opin Crit Care, 8, 6-11.
FRANCHI, L., KANNEGANTI, T.D., DUBYAK, G.R. & NUNEZ, G. (2007). Differential
requirement of P2X7 receptor and intracellular K+ for caspase-1 activation induced
by intracellular and extracellular bacteria. J Biol Chem, 282, 18810-8.
FUJIHARA, M., MUROI, M., TANAMOTO, K., SUZUKI, T., AZUMA, H. & IKEDA, H.
(2003). Molecular mechanisms of macrophage activation and deactivation by
lipopolysaccharide: roles of the receptor complex. Pharmacol Ther, 100, 171-94.
GADEK, J.E. & PACHT, E.R. (1996). The interdependence of lung antioxidants and
antiprotease defense in ARDS. Chest, 110, 273S-277S.
GALLUCCI, S. & MATZINGER, P. (2001). Danger signals: SOS to the immune system.
Curr Opin Immunol, 13, 114-9.
GARBER, B.G., HEBERT, P.C., YELLE, J.D., HODDER, R.V. & MCGOWAN, J.
(1996). Adult respiratory distress syndrome: a systemic overview of incidence and
risk factors. Crit Care Med, 24, 687-95.
GARTNER, L.P. & HIATT, J.L. (2007). Color textbook of histology. Philadelphia, PA:
Saunders/Elsevier.
GEORGE, C.L., JIN, H., WOHLFORD-LENANE, C.L., O'NEILL, M.E., PHIPPS, J.C.,

112
O'SHAUGHNESSY, P., KLINE, J.N., THORNE, P.S. & SCHWARTZ, D.A.
(2001). Endotoxin responsiveness and subchronic grain dust-induced airway
disease. Am J Physiol Lung Cell Mol Physiol, 280, L203-13.
GIRI, S.N., HYDE, D.M. & HOLLINGER, M.A. (1993). Effect of antibody to
transforming growth factor beta on bleomycin induced accumulation of lung
collagen in mice. Thorax, 48, 959-66.
GOERDT, S. & ORFANOS, C.E. (1999). Other functions, other genes: alternative
activation of antigen-presenting cells. Immunity, 10, 137-42.
GOERDT, S., POLITZ, O., SCHLEDZEWSKI, K., BIRK, R., GRATCHEV, A.,
GUILLOT, P., HAKIY, N., KLEMKE, C.D., DIPPEL, E., KODELJA, V. &
ORFANOS, C.E. (1999). Alternative versus classical activation of macrophages.
Pathobiology, 67, 222-6.
GONCALVES, R.G., GABRICH, L., ROSARIO, A., JR., TAKIYA, C.M., FERREIRA,
M.L., CHIARINI, L.B., PERSECHINI, P.M., COUTINHO-SILVA, R. & LEITE,
M., JR. (2006). The role of purinergic P2X7 receptors in the inflammation and
fibrosis of unilateral ureteral obstruction in mice. Kidney Int, 70, 1599-606.
GORDON, J.L. (1986). Extracellular ATP: effects, sources and fate. Biochem J, 233, 309-
19.
GRUBER, B.L., MARCHESE, M.J. & KEW, R.R. (1994). Transforming growth factor-
beta 1 mediates mast cell chemotaxis. J Immunol, 152, 5860-7.
GU, B., BENDALL, L.J. & WILEY, J.S. (1998). Adenosine triphosphate-induced shedding
of CD23 and L-selectin (CD62L) from lymphocytes is mediated by the same
receptor but different metalloproteases. Blood, 92, 946-51.
GU, B.J. & WILEY, J.S. (2006). Rapid ATP-induced release of matrix metalloproteinase 9

113
is mediated by the P2X7 receptor. Blood, 107, 4946-53.
GUERRA, A.N., FISETTE, P.L., PFEIFFER, Z.A., QUINCHIA-RIOS, B.H., PRABHU,
U., AGA, M., DENLINGER, L.C., GUADARRAMA, A.G., ABOZEID, S.,
SOMMER, J.A., PROCTOR, R.A. & BERTICS, P.J. (2003). Purinergic receptor
regulation of LPS-induced signaling and pathophysiology. J Endotoxin Res, 9, 256-
63.
HAAG, F., ADRIOUCH, S., BRASS, A., JUNG, C., MOLLER, S., SCHEUPLEIN, F.,
BANNAS, P., SEMAN, M. & KOCH-NOLTE, F. (2007). Extracellular NAD and
ATP: Partners in immune cell modulation. Purinergic Signal, 3, 71-81.
HAAG, F., ADRIOUCH, S., BRASS, A., JUNG, C., MOLLER, S., SCHEUPLEIN, F.,
BANNAS, P., SEMAN, M. & KOCH-NOLTE, F. (2007). Extracellular NAD and
ATP: Partners in immune cell modulation. Purinergic Signal, 3, 71-81.
HADDAD, J.J., OLVER, R.E. & LAND, S.C. (2000). Antioxidant/pro-oxidant equilibrium
regulates HIF-1alpha and NF-kappa B redox sensitivity. Evidence for inhibition by
glutathione oxidation in alveolar epithelial cells. J Biol Chem, 275, 21130-9.
HAUTAMAKI, R.D., KOBAYASHI, D.K., SENIOR, R.M. & SHAPIRO, S.D. (1997).
Requirement for macrophage elastase for cigarette smoke-induced emphysema in
mice. Science, 277, 2002-4.
HAYASHI, T., KAWAKAMI, M., SASAKI, S., KATSUMATA, T., MORI, H.,
YOSHIDA, H. & NAKAHARI, T. (2005). ATP regulation of ciliary beat frequency
in rat tracheal and distal airway epithelium. Exp Physiol, 90, 535-44.
HEWINSON, J., MOORE, S.F., GLOVER, C., WATTS, A.G. & MACKENZIE, A.B.
(2008). A key role for redox signaling in rapid P2X7 receptor-induced IL-1 beta
processing in human monocytes. J Immunol, 180, 8410-20.

114
HOCKING, W.G. & GOLDE, D.W. (1979). The pulmonary-alveolar macrophage (first of
two parts). N Engl J Med, 301, 580-7.
HOEBE, K., DU, X., GEORGEL, P., JANSSEN, E., TABETA, K., KIM, S.O., GOODE,
J., LIN, P., MANN, N., MUDD, S., CROZAT, K., SOVATH, S., HAN, J. &
BEUTLER, B. (2003). Identification of Lps2 as a key transducer of MyD88-
independent TIR signalling. Nature, 424, 743-8.
HOSTETTLER, K.E., ROTH, M., BURGESS, J.K., GENCAY, M.M., GAMBAZZI, F.,
BLACK, J.L., TAMM, M. & BORGER, P. (2008). Airway epithelium-derived
transforming growth factor-beta is a regulator of fibroblast proliferation in both
fibrotic and normal subjects. Clin Exp Allergy.
HU, Y., FISETTE, P.L., DENLINGER, L.C., GUADARRAMA, A.G., SOMMER, J.A.,
PROCTOR, R.A. & BERTICS, P.J. (1998). Purinergic receptor modulation of
lipopolysaccharide signaling and inducible nitric-oxide synthase expression in
RAW 264.7 macrophages. J Biol Chem, 273, 27170-5.
HUDSON, L.D. & STEINBERG, K.P. (1999). Epidemiology of acute lung injury and
ARDS. Chest, 116, 74S-82S.
HUMPHREYS, B.D. & DUBYAK, G.R. (1996). Induction of the P2z/P2X7 nucleotide
receptor and associated phospholipase D activity by lipopolysaccharide and IFN-
gamma in the human THP-1 monocytic cell line. J Immunol, 157, 5627-37.
IDZKO, M., HAMMAD, H., VAN NIMWEGEN, M., KOOL, M., WILLART, M.A.,
MUSKENS, F., HOOGSTEDEN, H.C., LUTTMANN, W., FERRARI, D., DI
VIRGILIO, F., VIRCHOW, J.C., JR. & LAMBRECHT, B.N. (2007). Extracellular
ATP triggers and maintains asthmatic airway inflammation by activating dendritic
cells. Nat Med, 13, 913-9.

115
ISCHIROPOULOS, H. (1998). Biological tyrosine nitration: a pathophysiological function
of nitric oxide and reactive oxygen species. Arch Biochem Biophys, 356, 1-11.
JAFFAR, Z.H. & PEARCE, F.L. (1990). Histamine secretion from mast cells stimulated
with ATP. Agents Actions, 30, 64-6.
JANEWAY, C.A., JR. & MEDZHITOV, R. (2002). Innate immune recognition. Annu Rev
Immunol, 20, 197-216.
JANOFF, A. (1985). Elastases and emphysema. Current assessment of the protease-
antiprotease hypothesis. Am Rev Respir Dis, 132, 417-33.
KHAIR, O.A., DAVIES, R.J. & DEVALIA, J.L. (1996). Bacterial-induced release of
inflammatory mediators by bronchial epithelial cells. Eur Respir J, 9, 1913-22.
KIM, M., JIANG, L.H., WILSON, H.L., NORTH, R.A. & SURPRENANT, A. (2001).
Proteomic and functional evidence for a P2X7 receptor signalling complex. Embo J,
20, 6347-58.
KIM, S.Y., MOON, J.H., LEE, H.G., KIM, S.U. & LEE, Y.B. (2007). ATP released from
beta-amyloid-stimulated microglia induces reactive oxygen species production in an
autocrine fashion. Exp Mol Med, 39, 820-7.
KITOWSKA, K., ZAKRZEWICZ, D., KONIGSHOFF, M., CHROBAK, I.,
GRIMMINGER, F., SEEGER, W., BULAU, P. & EICKELBERG, O. (2008).
Functional role and species-specific contribution of arginases in pulmonary fibrosis.
Am J Physiol Lung Cell Mol Physiol, 294, L34-45.
KOBZIK, L., GODLESKI, J.J. & BRAIN, J.D. (1990). Selective down-regulation of
alveolar macrophage oxidative response to opsonin-independent phagocytosis. J
Immunol, 144, 4312-9.
KOCHI, T., OKUBO, S., ZIN, W.A. & MILIC-EMILI, J. (1988). Chest wall and

116
respiratory system mechanics in cats: effects of flow and volume. J Appl Physiol,
64, 2636-46.
KOCHI, T., OKUBO, S., ZIN, W.A. & MILIC-EMILI, J. (1988). Flow and volume
dependence of pulmonary mechanics in anesthetized cats. J Appl Physiol, 64, 441-
50.
KORNBLUTH, R.S. & STONE, G.W. (2006). Immunostimulatory combinations:
designing the next generation of vaccine adjuvants. J Leukoc Biol, 80, 1084-102.
KUHNS, D.B., WRIGHT, D.G., NATH, J., KAPLAN, S.S. & BASFORD, R.E. (1988).
ATP induces transient elevations of [Ca2+]i in human neutrophils and primes these
cells for enhanced O2- generation. Lab Invest, 58, 448-53.
KUNZELMANN, K. & MALL, M. (2003). Pharmacotherapy of the ion transport defect in
cystic fibrosis: role of purinergic receptor agonists and other potential therapeutics.
Am J Respir Med, 2, 299-309.
KUROKI, M. & MINAKAMI, S. (1989). Extracellular ATP triggers superoxide production
in human neutrophils. Biochem Biophys Res Commun, 162, 377-80.
LA SALA, A., FERRARI, D., DI VIRGILIO, F., IDZKO, M., NORGAUER, J. &
GIROLOMONI, G. (2003). Alerting and tuning the immune response by
extracellular nucleotides. J Leukoc Biol, 73, 339-43.
LABASI, J.M., PETRUSHOVA, N., DONOVAN, C., MCCURDY, S., LIRA, P.,
PAYETTE, M.M., BRISSETTE, W., WICKS, J.R., AUDOLY, L. & GABEL, C.A.
(2002). Absence of the P2X7 receptor alters leukocyte function and attenuates an
inflammatory response. J Immunol, 168, 6436-45.
LAMMAS, D.A., STOBER, C., HARVEY, C.J., KENDRICK, N., PANCHALINGAM, S.
& KUMARARATNE, D.S. (1997). ATP-induced killing of mycobacteria by human

117
macrophages is mediated by purinergic P2Z(P2X7) receptors. Immunity, 7, 433-44.
LECHAPT-ZALCMAN, E., PRULIERE-ESCABASSE, V., ADVENIER, D., GALIACY,
S., CHARRIERE-BERTRAND, C., COSTE, A., HARF, A., D'ORTHO, M.P. &
ESCUDIER, E. (2006). Transforming growth factor-beta1 increases airway wound
repair via MMP-2 upregulation: a new pathway for epithelial wound repair? Am J
Physiol Lung Cell Mol Physiol, 290, L1277-82.
LECHAPT-ZALCMAN, E., PRULIERE-ESCABASSE, V., ADVENIER, D., GALIACY,
S., CHARRIERE-BERTRAND, C., COSTE, A., HARF, A., D'ORTHO, M.P. &
ESCUDIER, E. (2006). Transforming growth factor-beta1 increases airway wound
repair via MMP-2 upregulation: a new pathway for epithelial wound repair? Am J
Physiol Lung Cell Mol Physiol, 290, L1277-82.
LEE, Y.H., LEE, S.J., SEO, M.H., KIM, C.J. & SIM, S.S. (2001). ATP-induced histamine
release is in part related to phospholipase A2-mediated arachidonic acid metabolism
in rat peritoneal mast cells. Arch Pharm Res, 24, 552-6.
LEIPZIGER, J. (2003). Control of epithelial transport via luminal P2 receptors. Am J
Physiol Renal Physiol, 284, F419-32.
LEWIS, C.J. & EVANS, R.J. (2001). P2X receptor immunoreactivity in different arteries
from the femoral, pulmonary, cerebral, coronary and renal circulations. J Vasc Res,
38, 332-40.
LISTER, M.F., SHARKEY, J., SAWATZKY, D.A., HODGKISS, J.P., DAVIDSON, D.J.,
ROSSI, A.G. & FINLAYSON, K. (2007). The role of the purinergic P2X7 receptor
in inflammation. J Inflamm (Lond), 4, 5.
LORING, S.H. & MEAD, J. (1982). Action of the diaphragm on the rib cage inferred from
a force-balance analysis. J Appl Physiol, 53, 756-60.

118
LUCE, J.M. (1998). Acute lung injury and the acute respiratory distress syndrome. Crit
Care Med, 26, 369-76.
LUHR, O.R., ANTONSEN, K., KARLSSON, M., AARDAL, S., THORSTEINSSON, A.,
FROSTELL, C.G. & BONDE, J. (1999). Incidence and mortality after acute
respiratory failure and acute respiratory distress syndrome in Sweden, Denmark,
and Iceland. The ARF Study Group. Am J Respir Crit Care Med, 159, 1849-61.
MACKLIN, C.C. (1954). The pulmonary alveolar mucoid film and the pneumonocytes.
Lancet, 266, 1099-1104.
MACNEE, W. (2005). Pulmonary and systemic oxidant/antioxidant imbalance in chronic
obstructive pulmonary disease. Proc Am Thorac Soc, 2, 50-60.
MADDOX, D.E., GOLDSTEIN, I.J. & LOBUGLIO, A.F. (1982). Griffonia simplicifolia I
lectin mediates macrophage-induced cytotoxicity against Ehrlich ascites tumor. Cell
Immunol, 71, 202-7.
MADDOX, D.E., SHIBATA, S. & GOLDSTEIN, I.J. (1982). Stimulated macrophages
express a new glycoprotein receptor reactive with Griffonia simplicifolia I-B4
isolectin. Proc Natl Acad Sci U S A, 79, 166-70.
MALAVIA, N.K., MIH, J.D., RAUB, C.B., DINH, B.T. & GEORGE, S.C. (2008). IL-13
induces a bronchial epithelial phenotype that is profibrotic. Respir Res, 9, 27.
MALIZIA, A.P., KEATING, D.T., SMITH, S.M., WALLS, D., DORAN, P.P. & EGAN,
J.J. (2008). Alveolar Epithelial Cell Injury with Epstein Barr Virus Upregulates
TGF{beta}1 Expression. Am J Physiol Lung Cell Mol Physiol.
MARIATHASAN, S., NEWTON, K., MONACK, D.M., VUCIC, D., FRENCH, D.M.,
LEE, W.P., ROOSE-GIRMA, M., ERICKSON, S. & DIXIT, V.M. (2004).
Differential activation of the inflammasome by caspase-1 adaptors ASC and Ipaf.

119
Nature, 430, 213-8.
MASON, H.L. (1987). Evolving health care system: research issues and challenges. Am J
Pharm Educ, 51, 396-9.
MASON, H.S., BOURKE, S. & KEMP, P.J. (2004). Selective modulation of ligand-gated
P2X purinoceptor channels by acute hypoxia is mediated by reactive oxygen
species. Mol Pharmacol, 66, 1525-35.
MATTHAY, M.A., ZIMMERMAN, G.A., ESMON, C., BHATTACHARYA, J.,
COLLER, B., DOERSCHUK, C.M., FLOROS, J., GIMBRONE, M.A., JR.,
HOFFMAN, E., HUBMAYR, R.D., LEPPERT, M., MATALON, S., MUNFORD,
R., PARSONS, P., SLUTSKY, A.S., TRACEY, K.J., WARD, P., GAIL, D.B. &
HARABIN, A.L. (2003). Future research directions in acute lung injury: summary
of a National Heart, Lung, and Blood Institute working group. Am J Respir Crit
Care Med, 167, 1027-35.
MATZINGER, P. (2002). The danger model: a renewed sense of self. Science, 296, 301-5.
MCKNIGHT, A.J., MACFARLANE, A.J., DRI, P., TURLEY, L., WILLIS, A.C. &
GORDON, S. (1996). Molecular cloning of F4/80, a murine macrophage-restricted
cell surface glycoprotein with homology to the G-protein-linked transmembrane 7
hormone receptor family. J Biol Chem, 271, 486-9.
MENEZES, S.L., BOZZA, P.T., NETO, H.C., LARANJEIRA, A.P., NEGRI, E.M.,
CAPELOZZI, V.L., ZIN, W.A. & ROCCO, P.R. (2005). Pulmonary and
extrapulmonary acute lung injury: inflammatory and ultrastructural analyses. J Appl
Physiol, 98, 1777-83.
MERCER, R.R. & CRAPO, J.D. (1990). Spatial distribution of collagen and elastin fibers
in the lungs. J Appl Physiol, 69, 756-65.

120
MESSAGE, S.D. & JOHNSTON, S.L. (2004). Host defense function of the airway
epithelium in health and disease: clinical background. J Leukoc Biol, 75, 5-17.
MORTOLA, J.P. & NOWORAJ, A. (1983). Two-sidearm tracheal cannula for respiratory
airflow measurements in small animals. J Appl Physiol, 55, 250-3.
MURPHY, J.K., LIVINGSTON, F.R., GOZAL, E., TORRES, M. & FORMAN, H.J.
(1993). Stimulation of the rat alveolar macrophage respiratory burst by extracellular
adenine nucleotides. Am J Respir Cell Mol Biol, 9, 505-10.
MYRTEK, D. & IDZKO, M. (2007). Chemotactic activity of extracellular nucleotideson
human immune cells. Purinergic Signal, 3, 5-11.
NAKAMURA, T., EBIHARA, I., SHIMADA, N., SHOJI, H. & KOIDE, H. (1998).
Modulation of plasma metalloproteinase-9 concentrations and peripheral blood
monocyte mRNA levels in patients with septic shock: effect of fiber-immobilized
polymyxin B treatment. Am J Med Sci, 316, 355-60.
NATHAN, C. (2002). Points of control in inflammation. Nature, 420, 846-52.
NEGRI, E.M., HOELZ, C., BARBAS, C.S., MONTES, G.S., SALDIVA, P.H. &
CAPELOZZI, V.L. (2002). Acute remodeling of parenchyma in pulmonary and
extrapulmonary ARDS. An autopsy study of collagen-elastic system fibers. Pathol
Res Pract, 198, 355-61.
NOGUCHI, T., ISHII, K., FUKUTOMI, H., NAGURO, I., MATSUZAWA, A., TAKEDA,
K. & ICHIJO, H. (2008). Requirement of reactive oxygen species-dependent
activation of ASK1-p38 MAPK pathway for extracellular ATP-induced apoptosis in
macrophage. J Biol Chem, 283, 7657-65.
NORTH, R.A. (2002). Molecular physiology of P2X receptors. Physiol Rev, 82, 1013-67.
NOVAK, I. (2003). ATP regulation of epithelial Cl- channels--new challenges? J Physiol,

121
547, 1.
NUMATA, M., SUZUKI, S., MIYAZAWA, N., MIYASHITA, A., NAGASHIMA, Y.,
INOUE, S., KANEKO, T. & OKUBO, T. (1998). Inhibition of inducible nitric
oxide synthase prevents LPS-induced acute lung injury in dogs. J Immunol, 160,
3031-7.
OPAL, S.M. (2007). The host response to endotoxin, antilipopolysaccharide strategies, and
the management of severe sepsis. Int J Med Microbiol, 297, 365-77.
ORITO, K., HASHIDA, M., HIRATA, K., KUROKAWA, A., SHIRAI, M. & AKAHORI,
F. (2006). Effects of single intratracheal exposure to chlorhexidine gluconate on the
rat lung. Drug Chem Toxicol, 29, 1-9.
OWEN, C.A. (2005). Proteinases and oxidants as targets in the treatment of chronic
obstructive pulmonary disease. Proc Am Thorac Soc, 2, 373-85; discussion 394-5.
PARK, K.S., WELLS, J.M., ZORN, A.M., WERT, S.E., LAUBACH, V.E., FERNANDEZ,
L.G. & WHITSETT, J.A. (2006). Transdifferentiation of ciliated cells during repair
of the respiratory epithelium. Am J Respir Cell Mol Biol, 34, 151-7.
PARKS, W.C. & SHAPIRO, S.D. (2001). Matrix metalloproteinases in lung biology.
Respir Res, 2, 10-9.
PARVATHENANI, L.K., TERTYSHNIKOVA, S., GRECO, C.R., ROBERTS, S.B.,
ROBERTSON, B. & POSMANTUR, R. (2003). P2X7 mediates superoxide
production in primary microglia and is up-regulated in a transgenic mouse model of
Alzheimer's disease. J Biol Chem, 278, 13309-17.
PELEGRIN, P., BARROSO-GUTIERREZ, C. & SURPRENANT, A. (2008). P2X7
receptor differentially couples to distinct release pathways for IL-1beta in mouse
macrophage. J Immunol, 180, 7147-57.

122
PERREGAUX, D.G., MCNIFF, P., LALIBERTE, R., CONKLYN, M. & GABEL, C.A.
(2000). ATP acts as an agonist to promote stimulus-induced secretion of IL-1 beta
and IL-18 in human blood. J Immunol, 165, 4615-23.
PETRYNIAK, J., HUARD, T.K., NORDBLOM, G.D. & GOLDSTEIN, I.J. (1986). Lectin
binding studies on murine peritoneal cells: physicochemical characterization of the
binding of lectins from Datura stramonium, Evonymus europaea, and Griffonia
simplicifolia to murine peritoneal cells. Arch Biochem Biophys, 244, 57-66.
PFEIFFER, Z.A., GUERRA, A.N., HILL, L.M., GAVALA, M.L., PRABHU, U., AGA,
M., HALL, D.J. & BERTICS, P.J. (2007). Nucleotide receptor signaling in murine
macrophages is linked to reactive oxygen species generation. Free Radic Biol Med,
42, 1506-16.
PIPER, R.D., PITT-HYDE, M., LI, F., SIBBALD, W.J. & POTTER, R.F. (1996).
Microcirculatory changes in rat skeletal muscle in sepsis. Am J Respir Crit Care
Med, 154, 931-7.
PITTET, J.F., GRIFFITHS, M.J., GEISER, T., KAMINSKI, N., DALTON, S.L., HUANG,
X., BROWN, L.A., GOTWALS, P.J., KOTELIANSKY, V.E., MATTHAY, M.A.
& SHEPPARD, D. (2001). TGF-beta is a critical mediator of acute lung injury. J
Clin Invest, 107, 1537-44.
POLTORAK, A., SMIRNOVA, I., HE, X., LIU, M.Y., VAN HUFFEL, C., MCNALLY,
O., BIRDWELL, D., ALEJOS, E., SILVA, M., DU, X., THOMPSON, P., CHAN,
E.K., LEDESMA, J., ROE, B., CLIFTON, S., VOGEL, S.N. & BEUTLER, B.
(1998). Genetic and physical mapping of the Lps locus: identification of the toll-4
receptor as a candidate gene in the critical region. Blood Cells Mol Dis, 24, 340-55.
PONTOPPIDAN, H., GEFFIN, B. & LOWENSTEIN, E. (1972). Acute respiratory failure

123
in the adult. 3. N Engl J Med, 287, 799-806.
POSTMA, D.S. & TIMENS, W. (2006). Remodeling in asthma and chronic obstructive
pulmonary disease. Proc Am Thorac Soc, 3, 434-9.
PROCTOR, R.A., DENLINGER, L.C., LEVENTHAL, P.S., DAUGHERTY, S.K., VAN
DE LOO, J.W., TANKE, T., FIRESTEIN, G.S. & BERTICS, P.J. (1994).
Protection of mice from endotoxic death by 2-methylthio-ATP. Proc Natl Acad Sci
U S A, 91, 6017-20.
QIAO, R., ZHOU, B., LIEBLER, J.M., LI, X., CRANDALL, E.D. & BOROK, Z. (2003).
Identification of three genes of known function expressed by alveolar epithelial type
I cells. Am J Respir Cell Mol Biol, 29, 98-105.
RACKOW, E.C., ASTIZ, M.E., JANZ, T.G. & WEIL, M.W. (1988). Absence of
pulmonary edema during peritonitis and shock in rats. J Lab Clin Med, 112, 264-9.
RALEVIC, V. & BURNSTOCK, G. (1998). Receptors for purines and pyrimidines.
Pharmacol Rev, 50, 413-92.
RENCKENS, R., ROELOFS, J.J., FLORQUIN, S., DE VOS, A.F., LIJNEN, H.R., VAN'T
VEER, C. & VAN DER POLL, T. (2006). Matrix metalloproteinase-9 deficiency
impairs host defense against abdominal sepsis. J Immunol, 176, 3735-41.
RICCIARDOLO, F.L., STERK, P.J., GASTON, B. & FOLKERTS, G. (2004). Nitric oxide
in health and disease of the respiratory system. Physiol Rev, 84, 731-65.
RICOU, B., NICOD, L., LACRAZ, S., WELGUS, H.G., SUTER, P.M. & DAYER, J.M.
(1996). Matrix metalloproteinases and TIMP in acute respiratory distress syndrome.
Am J Respir Crit Care Med, 154, 346-52.
RIETSCHEL, E.T., KIRIKAE, T., SCHADE, F.U., MAMAT, U., SCHMIDT, G.,
LOPPNOW, H., ULMER, A.J., ZAHRINGER, U., SEYDEL, U., DI PADOVA, F.

124
& ET AL. (1994). Bacterial endotoxin: molecular relationships of structure to
activity and function. Faseb J, 8, 217-25.
RINALDO, J.E. (1986). Mediation of ARDS by leukocytes. Clinical evidence and
implications for therapy. Chest, 89, 590-3.
ROMAN, J., JEON, Y.J., GAL, A. & PEREZ, R.L. (1995). Distribution of extracellular
matrices, matrix receptors, and transforming growth factor-beta 1 in human and
experimental lung granulomatous inflammation. Am J Med Sci, 309, 124-33.
SAK, K. & WEBB, T.E. (2002). A retrospective of recombinant P2Y receptor subtypes and
their pharmacology. Arch Biochem Biophys, 397, 131-6.
SALDIVA, P.H., ZIN, W.A., SANTOS, R.L., EIDELMAN, D.H. & MILIC-EMILI, J.
(1992). Alveolar pressure measurement in open-chest rats. J Appl Physiol, 72, 302-
6.
SANTOS, F.B., NAGATO, L.K., BOECHEM, N.M., NEGRI, E.M., GUIMARAES, A.,
CAPELOZZI, V.L., FAFFE, D.S., ZIN, W.A. & ROCCO, P.R. (2006). Time course
of lung parenchyma remodeling in pulmonary and extrapulmonary acute lung
injury. J Appl Physiol, 100, 98-106.
SCHULMAN, E.S., GLAUM, M.C., POST, T., WANG, Y., RAIBLE, D.G., MOHANTY,
J., BUTTERFIELD, J.H. & PELLEG, A. (1999). ATP modulates anti-IgE-induced
release of histamine from human lung mast cells. Am J Respir Cell Mol Biol, 20,
530-7.
SCHWIEBERT, E.M. & ZSEMBERY, A. (2003). Extracellular ATP as a signaling
molecule for epithelial cells. Biochim Biophys Acta, 1615, 7-32.
SEIFERT, R., BURDE, R. & SCHULTZ, G. (1989). Activation of NADPH oxidase by
purine and pyrimidine nucleotides involves G proteins and is potentiated by

125
chemotactic peptides. Biochem J, 259, 813-9.
SELMAN, M. & PARDO, A. (2006). Role of epithelial cells in idiopathic pulmonary
fibrosis: from innocent targets to serial killers. Proc Am Thorac Soc, 3, 364-72.
SELZNER, N., SELZNER, M., GRAF, R., UNGETHUEM, U., FITZ, J.G. & CLAVIEN,
P.A. (2004). Water induces autocrine stimulation of tumor cell killing through ATP
release and P2 receptor binding. Cell Death Differ, 11 Suppl 2, S172-80.
SHEN, Z.J., ESNAULT, S., ROSENTHAL, L.A., SZAKALY, R.J., SORKNESS, R.L.,
WESTMARK, P.R., SANDOR, M. & MALTER, J.S. (2008). Pin1 regulates TGF-
beta1 production by activated human and murine eosinophils and contributes to
allergic lung fibrosis. J Clin Invest, 118, 479-90.
SIKORA, A., LIU, J., BROSNAN, C., BUELL, G., CHESSEL, I. & BLOOM, B.R. (1999).
Cutting edge: purinergic signaling regulates radical-mediated bacterial killing
mechanisms in macrophages through a P2X7-independent mechanism. J Immunol,
163, 558-61.
SIM, J.A., YOUNG, M.T., SUNG, H.Y., NORTH, R.A. & SURPRENANT, A. (2004).
Reanalysis of P2X7 receptor expression in rodent brain. J Neurosci, 24, 6307-14.
SOLLE, M., LABASI, J., PERREGAUX, D.G., STAM, E., PETRUSHOVA, N.,
KOLLER, B.H., GRIFFITHS, R.J. & GABEL, C.A. (2001). Altered cytokine
production in mice lacking P2X(7) receptors. J Biol Chem, 276, 125-32.
SOTO, F., GARCIA-GUZMAN, M., GOMEZ-HERNANDEZ, J.M., HOLLMANN, M.,
KARSCHIN, C. & STUHMER, W. (1996). P2X4: an ATP-activated ionotropic
receptor cloned from rat brain. Proc Natl Acad Sci U S A, 93, 3684-8.
SPERLAGH, B., HASKO, G., NEMETH, Z. & VIZI, E.S. (1998). ATP released by LPS
increases nitric oxide production in raw 264.7 macrophage cell line via P2Z/P2X7

126
receptors. Neurochem Int, 33, 209-15.
SU, G.L., SIMMONS, R.L. & WANG, S.C. (1995). Lipopolysaccharide binding protein
participation in cellular activation by LPS. Crit Rev Immunol, 15, 201-14.
SUH, B.C., KIM, J.S., NAMGUNG, U., HA, H. & KIM, K.T. (2001). P2X7 nucleotide
receptor mediation of membrane pore formation and superoxide generation in
human promyelocytes and neutrophils. J Immunol, 166, 6754-63.
SUZUKI, Y., TANIGAKI, T., HEIMER, D., WANG, W., ROSS, W.G., MURPHY, G.A.,
SAKAI, A., SUSSMAN, H.H., VU, T.H. & RAFFIN, T.A. (1994). TGF-beta 1
causes increased endothelial ICAM-1 expression and lung injury. J Appl Physiol,
77, 1281-7.
SWANSON, J.A. & BAER, S.C. (1995). Phagocytosis by zippers and triggers. Trends Cell
Biol, 5, 89-93.
TABOR, D.R., LARRY, C.H. & JACOBS, R.F. (1989). Differential induction of
macrophage GSIB4-binding activity. J Leukoc Biol, 45, 452-7.
TAKENOUCHI, T., IWAMARU, Y., SUGAMA, S., SATO, M., HASHIMOTO, M. &
KITANI, H. (2008). Lysophospholipids and ATP mutually suppress maturation and
release of IL-1 beta in mouse microglial cells using a Rho-dependent pathway. J
Immunol, 180, 7827-39.
TANKE, T., VAN DE LOO, J.W., RHIM, H., LEVENTHAL, P.S., PROCTOR, R.A. &
BERTICS, P.J. (1991). Bacterial lipopolysaccharide-stimulated GTPase activity in
RAW 264.7 macrophage membranes. Biochem J, 277 ( Pt 2), 379-85.
TAYLOR, A.L., SCHWIEBERT, L.M., SMITH, J.J., KING, C., JONES, J.R.,
SORSCHER, E.J. & SCHWIEBERT, E.M. (1999). Epithelial P2X purinergic
receptor channel expression and function. J Clin Invest, 104, 875-84.

127
TONETTI, M., STURLA, L., BISTOLFI, T., BENATTI, U. & DE FLORA, A. (1994).
Extracellular ATP potentiates nitric oxide synthase expression induced by
lipopolysaccharide in RAW 264.7 murine macrophages. Biochem Biophys Res
Commun, 203, 430-5.
TONETTI, M., STURLA, L., GIOVINE, M., BENATTI, U. & DE FLORA, A. (1995).
Extracellular ATP enhances mRNA levels of nitric oxide synthase and TNF-alpha
in lipopolysaccharide-treated RAW 264.7 murine macrophages. Biochem Biophys
Res Commun, 214, 125-30.
TRACEY, K.J. (2002). The inflammatory reflex. Nature, 420, 853-9.
TSAI, S.H., LIN-SHIAU, S.Y. & LIN, J.K. (1999). Suppression of nitric oxide synthase
and the down-regulation of the activation of NFkappaB in macrophages by
resveratrol. Br J Pharmacol, 126, 673-80.
TSAI, T.L., CHANG, S.Y., HO, C.Y. & KOU, Y.R. (2007). Activation of P2X
Purinoceptors by ATP subsequent to an increase in reactive oxygen species causes
enhanced laryngeal reflex reactivity in a rat model of extraoesophageal reflux. Gut.
UHAL, B.D. (1997). Cell cycle kinetics in the alveolar epithelium. Am J Physiol, 272,
L1031-45.
UNANUE, E.R. & ALLEN, P.M. (1987). The basis for the immunoregulatory role of
macrophages and other accessory cells. Science, 236, 551-7.
VERNOOY, J.H., DENTENER, M.A., VAN SUYLEN, R.J., BUURMAN, W.A. &
WOUTERS, E.F. (2002). Long-term intratracheal lipopolysaccharide exposure in
mice results in chronic lung inflammation and persistent pathology. Am J Respir
Cell Mol Biol, 26, 152-9.
VILLAR, J. & SLUTSKY, A.S. (1989). The incidence of the adult respiratory distress

128
syndrome. Am Rev Respir Dis, 140, 814-6.
VIRGINIO, C., MACKENZIE, A., NORTH, R.A. & SURPRENANT, A. (1999). Kinetics
of cell lysis, dye uptake and permeability changes in cells expressing the rat P2X7
receptor. J Physiol, 519 Pt 2, 335-46.
VYAS-READ, S., SHAUL, P.W., YUHANNA, I.S. & WILLIS, B.C. (2007). Nitric oxide
attenuates epithelial-mesenchymal transition in alveolar epithelial cells. Am J
Physiol Lung Cell Mol Physiol, 293, L212-21.
WAHL, S.M., HUNT, D.A., WAKEFIELD, L.M., MCCARTNEY-FRANCIS, N., WAHL,
L.M., ROBERTS, A.B. & SPORN, M.B. (1987). Transforming growth factor type
beta induces monocyte chemotaxis and growth factor production. Proc Natl Acad
Sci U S A, 84, 5788-92.
WANG, C.M., CHANG, Y.Y. & SUN, S.H. (2003). Activation of P2X7 purinoceptor-
stimulated TGF-beta 1 mRNA expression involves PKC/MAPK signalling pathway
in a rat brain-derived type-2 astrocyte cell line, RBA-2. Cell Signal, 15, 1129-37.
WANG, X., ARCUINO, G., TAKANO, T., LIN, J., PENG, W.G., WAN, P., LI, P., XU,
Q., LIU, Q.S., GOLDMAN, S.A. & NEDERGAARD, M. (2004). P2X7 receptor
inhibition improves recovery after spinal cord injury. Nat Med, 10, 821-7.
WARD, P.A. (1994). Oxygen radicals, cytokines, adhesion molecules, and lung injury.
Environ Health Perspect, 102 Suppl 10, 13-6.
WARD, P.A., CUNNINGHAM, T.W., MCCULLOCH, K.K. & JOHNSON, K.J. (1988).
Regulatory effects of adenosine and adenine nucleotides on oxygen radical
responses of neutrophils. Lab Invest, 58, 438-47.
WARE, L.B. & MATTHAY, M.A. (2000). The acute respiratory distress syndrome. N Engl
J Med, 342, 1334-49.

129
WASSERMAN, S.I. (1980). The lung mast cell: its physiology and potential relevance to
defense of the lung. Environ Health Perspect, 35, 153-64.
WATSON, R.W., REDMOND, H.P. & BOUCHIER-HAYES, D. (1994). Role of
endotoxin in mononuclear phagocyte-mediated inflammatory responses. J Leukoc
Biol, 56, 95-103.
WATTERS, J.J., SOMMER, J.A., PFEIFFER, Z.A., PRABHU, U., GUERRA, A.N. &
BERTICS, P.J. (2002). A differential role for the mitogen-activated protein kinases
in lipopolysaccharide signaling: the MEK/ERK pathway is not essential for nitric
oxide and interleukin 1beta production. J Biol Chem, 277, 9077-87.
WILLIS, B.C. & BOROK, Z. (2007). TGF-beta-induced EMT: mechanisms and
implications for fibrotic lung disease. Am J Physiol Lung Cell Mol Physiol, 293,
L525-34.
WINKLER, M.K., FOLDES, J.K., BUNN, R.C. & FOWLKES, J.L. (2003). Implications
for matrix metalloproteinases as modulators of pediatric lung disease. Am J Physiol
Lung Cell Mol Physiol, 284, L557-65.
XISTO, D.G., FARIAS, L.L., FERREIRA, H.C., PICANCO, M.R., AMITRANO, D.,
LAPA, E.S.J.R., NEGRI, E.M., MAUAD, T., CARNIELLI, D., SILVA, L.F.,
CAPELOZZI, V.L., FAFFE, D.S., ZIN, W.A. & ROCCO, P.R. (2005). Lung
parenchyma remodeling in a murine model of chronic allergic inflammation. Am J
Respir Crit Care Med, 171, 829-37.
YAMAMOTO, M., YAGINUMA, K., TSUTSUI, H., SAGARA, J., GUAN, X., SEKI, E.,
YASUDA, K., AKIRA, S., NAKANISHI, K., NODA, T. & TANIGUCHI, S.
(2004). ASC is essential for LPS-induced activation of procaspase-1 independently
of TLR-associated signal adaptor molecules. Genes Cells, 9, 1055-67.

130
YEHUALAESHET, T., O'CONNOR, R., BEGLEITER, A., MURPHY-ULLRICH, J.E.,
SILVERSTEIN, R. & KHALIL, N. (2000). A CD36 synthetic peptide inhibits
bleomycin-induced pulmonary inflammation and connective tissue synthesis in the
rat. Am J Respir Cell Mol Biol, 23, 204-12.
ZHANG, Y., LEE, T.C., GUILLEMIN, B., YU, M.C. & ROM, W.N. (1993). Enhanced IL-
1 beta and tumor necrosis factor-alpha release and messenger RNA expression in
macrophages from idiopathic pulmonary fibrosis or after asbestos exposure. J
Immunol, 150, 4188-96.
ZILBERBERG, M.D. & EPSTEIN, S.K. (1998). Acute lung injury in the medical ICU:
comorbid conditions, age, etiology, and hospital outcome. Am J Respir Crit Care
Med, 157, 1159-64.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo