understanding organism energy allocations in response to climate change – ideas for approaches to...
TRANSCRIPT

Understanding organism energy allocations in response to climate change
– ideas for approaches to ‘systems biology’ modelling?
Chris Hauton
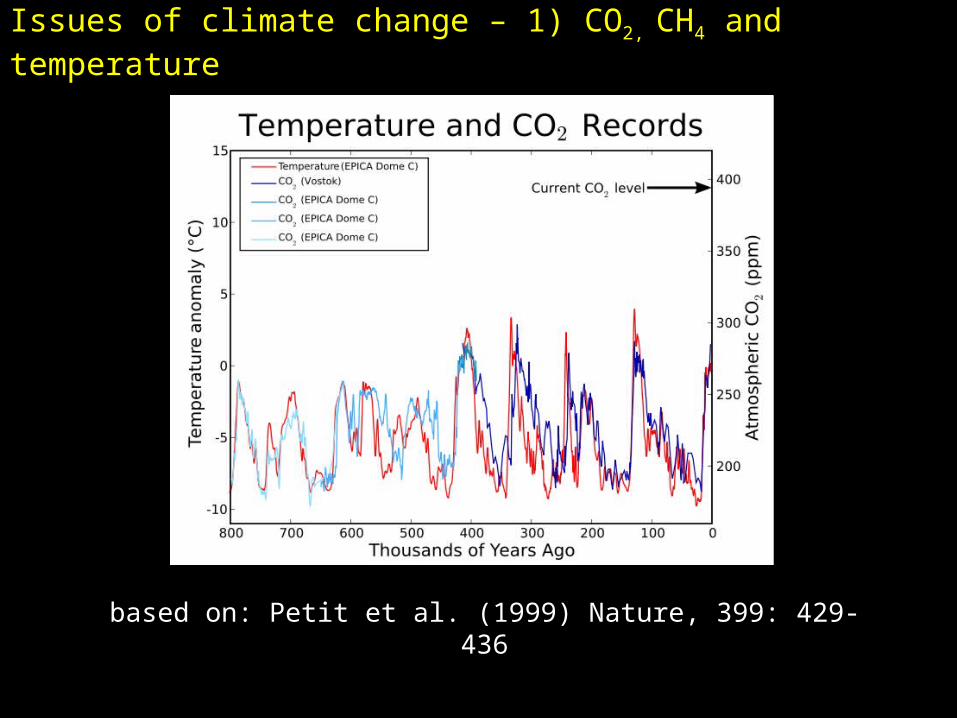
Issues of climate change – 1) CO2, CH4 and temperature
based on: Petit et al. (1999) Nature, 399: 429-436
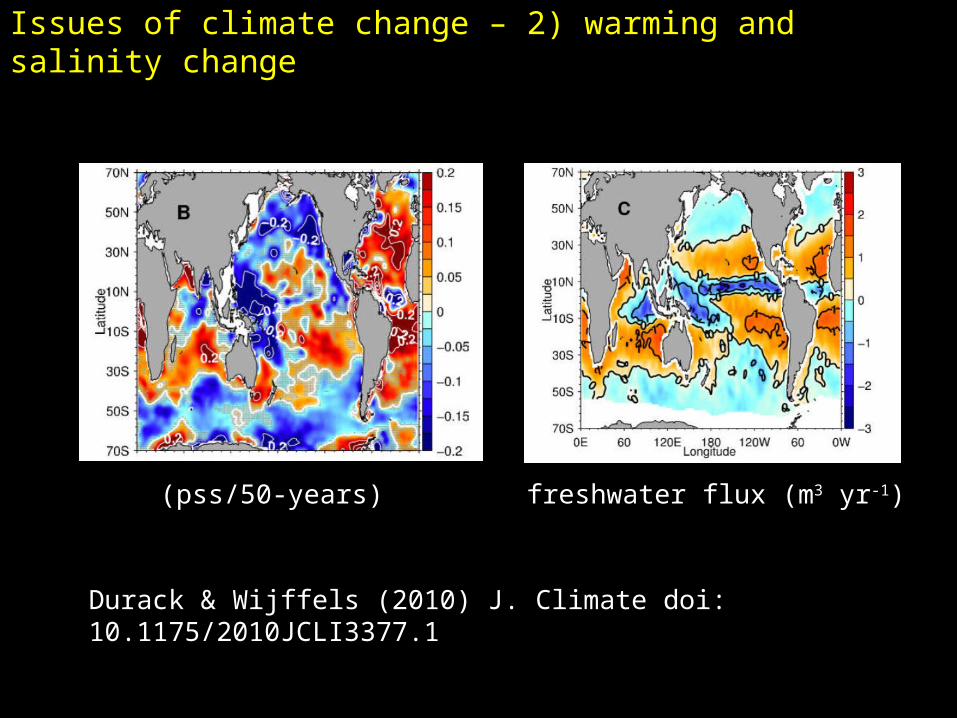
Issues of climate change – 2) warming and salinity change
Durack & Wijffels (2010) J. Climate doi: 10.1175/2010JCLI3377.1
(pss/50-years) freshwater flux (m3 yr-1)

Issues of climate change – 3) CO2 and seawater acidification
‘the other CO2 problem’
Doney et al. (2009) Annual Reviews - Marine Science, 1
CO2 (g) CO2 (aq)
CO2 (aq) + H2O (aq) HCO3- (aq) + H+ (aq)
CaCO3 (s) Ca 2+ (aq) + CO3
2- (aq)
H+ (aq) + CO32-
(aq) HCO3-
(aq)
3a) bicarbonate buffering…
carbonate dissolution…

Issues of climate change – 3) CO2 and seawater acidification
‘the other CO2 problem’
from Pörtner et al. (2004)
3b) disruption to acid base balance in osmoconformers

Issues of climate change – 3) CO2 and seawater acidification
‘the other CO2 problem’
3b) disruption to acid base balance in osmoconformers (Hauton et al., 2009)
Treatment
Control Nom. pH 7.8Nom. pH 7.6
CO
PIE
S m
RN
A.
g t
ota
l RN
A-1
0
2e+6
4e+6
6e+6
hsp70 gene
4.0e+6
2.4e+7
4.4e+7
6.4e+7
gapdh gene
*

Issues of climate change – 3) CO2 and seawater acidification
‘the other CO2 problem’
3a) reduced availability of carbonate ions causing a reduction in calcification (Riebesell, 2000; Sciandra et al., 2003; Gazeau et al. 2007) but species/experimental differences? (Iglesias-Rodriguez et al., 2008)
3b) direct impact on the intracellular pH (pHi) of many species (Seibel & Walsh, 2003), impacting normal protein synthesis (Kwast & Hand, 1996), respiratory function (Spicer & Taylor, 1994; Pörtner et al., 2004) and immune function (Bibby et al., 2008)
disruption to the physiology and performance of marine species (Kurihara et
al., 2004; Berge et al., 2006; Spicer et al., 2007)

All estimates are subject to uncertainty; variation with region, latitude and depth (IPCC Fourth Assessment Report)
– T of +2 to +4 oC– pH of -0.4 to -0.5 units– S of -0.05 psu
Issues of climate change – predictions by 2100?
However…
coastal and estuarine environments extremely variable
species which may have evolved strategies to accommodate extreme episodic low pH
Ringwood & Keppler, 2002
Attrill et al., 1999

Requirements for modelling
– stakeholders and policy makers require predictions
– research efforts must be directed towards outputs which have stakeholder relevance
– large-scale EU and UK research programmes have a significant component for integrating predictive models at different levels of biological organization

Existing approaches – ecosystem models
European Regional seas Ecosystem Model
version run by PML
tends to treat organisms as black boxes
efforts to refine this to reflect the different performance of different species
keystone species and ecosystem engineers
requires inputs from organism life history models
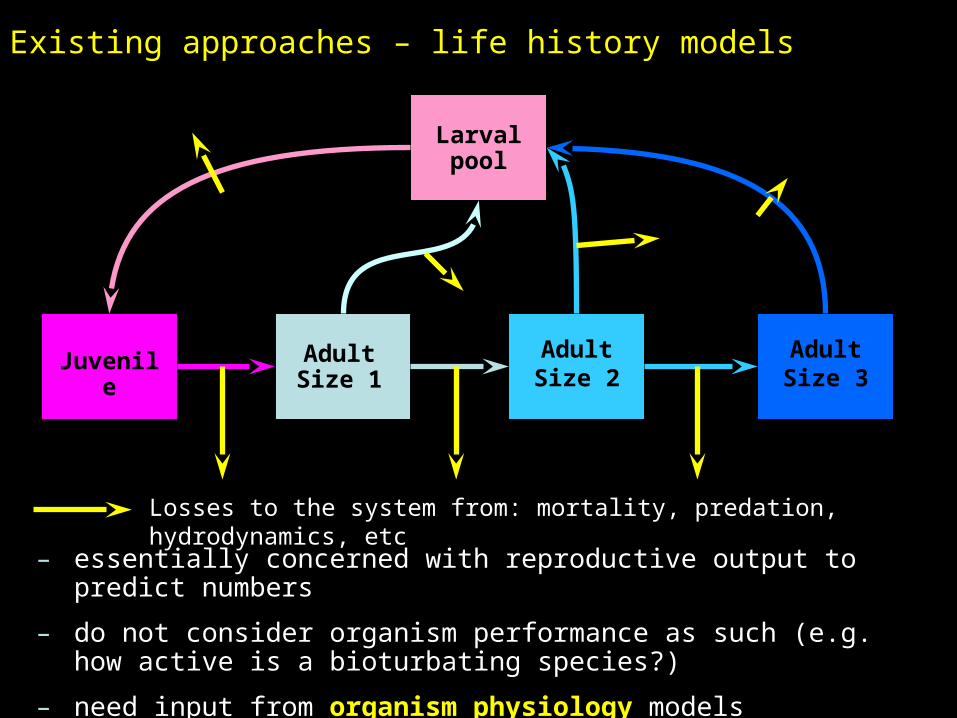
Existing approaches – life history models
Adult Size 1
Adult Size 2
Adult Size 3
Juvenile
Larval pool
Losses to the system from: mortality, predation, hydrodynamics, etc
– essentially concerned with reproductive output to predict numbers
– do not consider organism performance as such (e.g. how active is a bioturbating species?)
– need input from organism physiology models
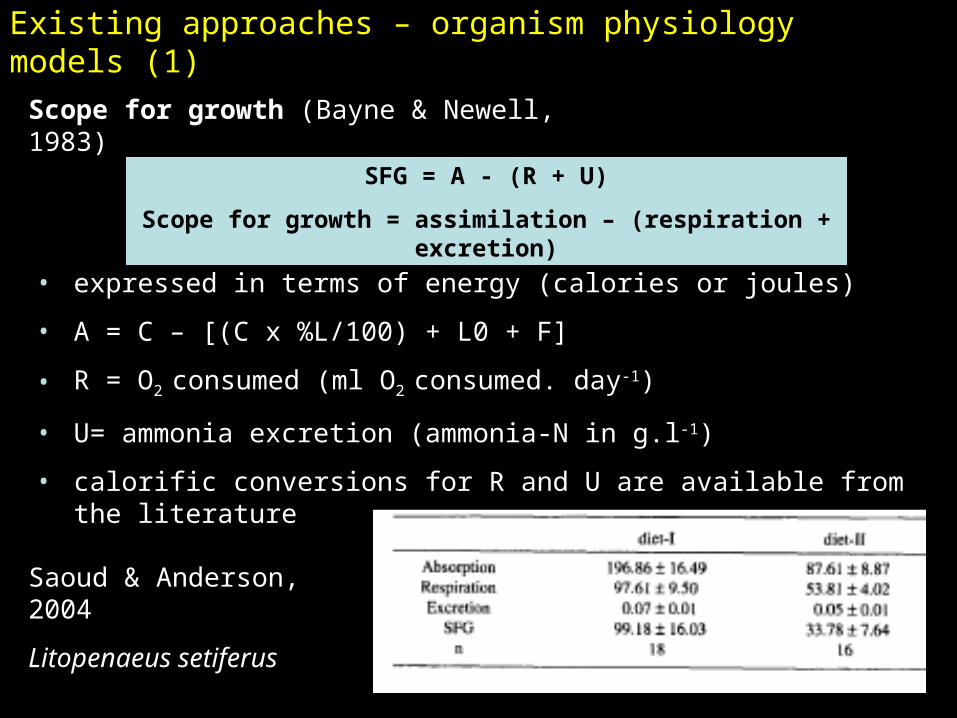
Existing approaches – organism physiology models (1)
Scope for growth (Bayne & Newell, 1983)
SFG = A - (R + U)
Scope for growth = assimilation – (respiration + excretion)
• expressed in terms of energy (calories or joules)
• A = C – [(C x %L/100) + L0 + F]
• R = O2 consumed (ml O2 consumed. day-1)
• U= ammonia excretion (ammonia-N in g.l-1)
• calorific conversions for R and U are available from the literature
Saoud & Anderson, 2004
Litopenaeus setiferus

SFG models are regarded as ‘net production models’– empirically determined sequence for nutrition and resource
allocation based on allometry– assume that assimilated energy is immediately available for
maintenance, the rest is used for growth or stored as reserves
Dynamic energy budget (DEB) models (Kooijman, 2000)
from Muller et al. (2010)
Existing approaches – organism physiology models (2)
DEB models – assimilated energy is stored in reserves which are then used for
maintenance, growth, development and reproduction– do not use allometric relationships, feeding rate is proportional to
surface area, maintenance scales to body volume – ‘aim’ is for a generic theory of energy budgets

Existing approaches – organism physiology models (2)
(DEB) models for the Pacific oyster (Pouvreau et al., 2006)
Existing approaches – organism physiology models (2)
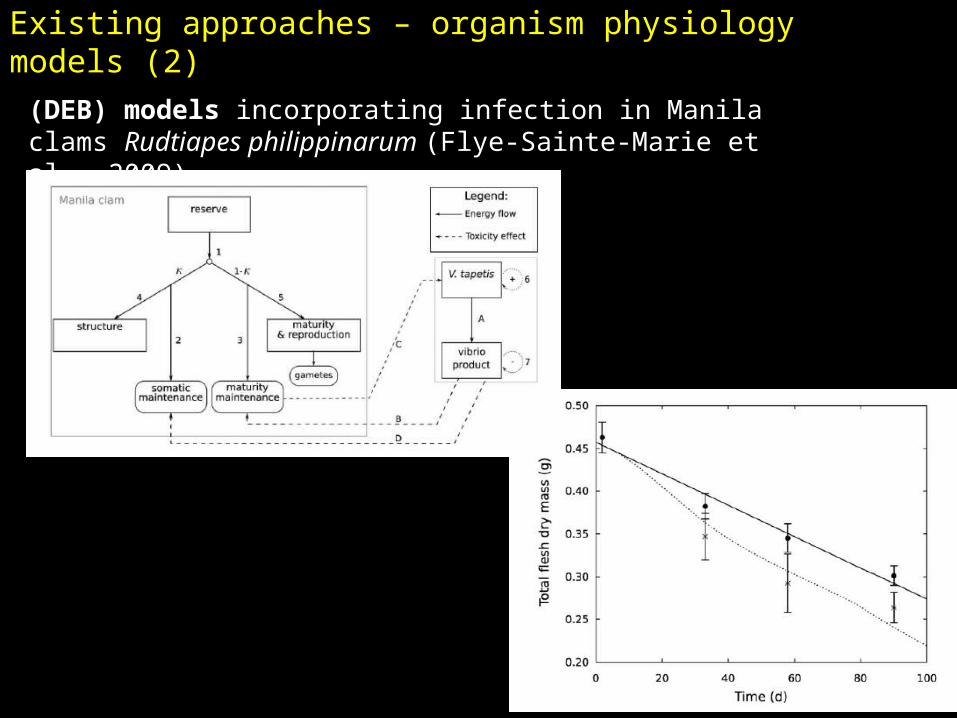
(DEB) models incorporating infection in Manila clams Rudtiapes philippinarum (Flye-Sainte-Marie et al., 2009)

Existing approaches – organism physiology models (2)
(DEB) models incorporating heavy metal pollution in bivalves (Muller et al., 2010)

Existing approaches – organism physiology models (2)
(DEB) models (oyster Crassostrea gigas model in STELLA™)
Organism life history
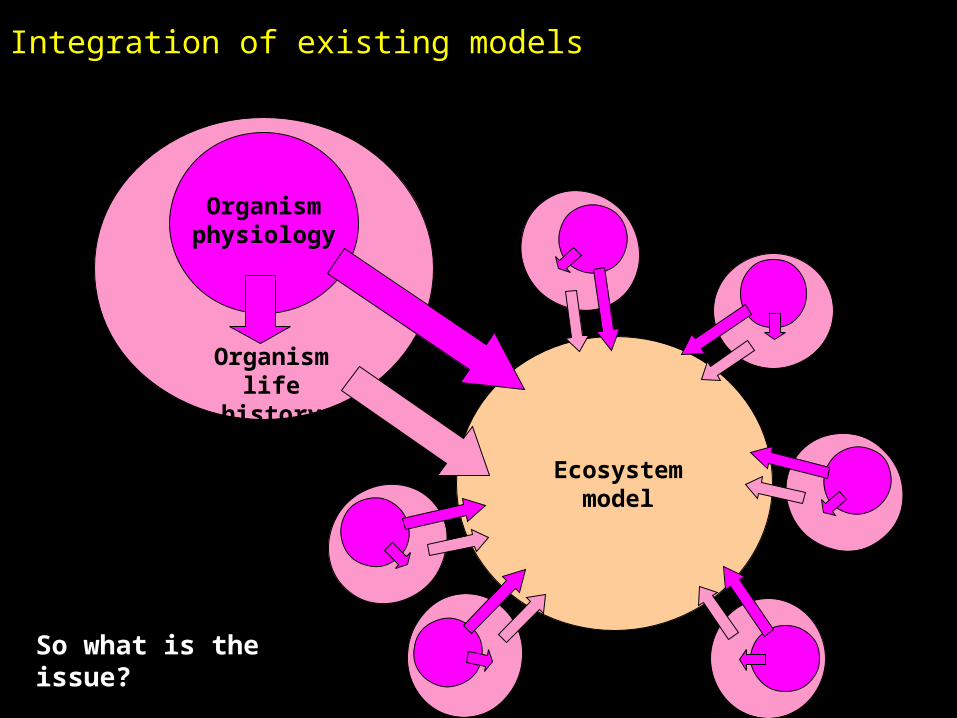
Integration of existing models
Organism physiology
Ecosystem model
So what is the issue?

Limitation of DEB and SFG models (1) – a personal view
Ecosystem model
1) measurements of the impacts of perturbation in pCO2, temperature and salinity are revealing sub-lethal changes in processes within ‘somatic maintenance’
e.g. acid base balance, protein turnover, osmoregulation, immune function

Limitation of DEB and SFG models (1) – a personal view
Ecosystem model
1) measurements of the impacts of perturbation in pCO2, temperature and salinity are revealing sub-lethal changes in processes within ‘somatic maintenance’
e.g. acid base balance, protein turnover, osmoregulation, immune function 2) some are (presently) unpredictable or not intuitive, but will have an impact on an organism performance when additional factors (e.g. pathogens) are added to the environment
3) current models, despite intentions, appear species or condition specific – a parsimonious solution is desired which accounts for all environmental perturbation and which is truly generically applicable

Limitation of DEB and SFG models (2) – a personal view
Ecosystem model
4) at critical life stages (e.g. larvae, juveniles) or in some species (polychaetes, small crustaceans or gastropods) only certain types of physiological measurements are possible
- protein expression, gene expression, enzyme activities, rates of protein turnover
- there is no convention on converting these to energy equivalents
- a need to cope with rates and proportions or relative quantities?

Summary– an end user need to predict the effects of future scenarios on
marine ecosystems
– working towards this by developing our understanding of organism
physiology from laboratory studies and small-scale lab and field
mesocosm experiments
– results indicate non linear and indirect effect of perturbations from
temperature and pCO2, acting in isolation and synergy
– have yet to incorporate in these experiments issues such as infection, pollution or salinity as multiplexed drivers
– reliance on integrated modelling from organism physiology to life
history and thence ecosystem models
– the challenge of incorporating disparate and high resolution
datasets (which are planned or being collected) into organism
level models (DEB or SFG) or alternatives… (process algebra?)
– a need to be generic