understanding biomechanical causes and …vk276ss2747/... · · 2011-09-22musculoskeletal atlas...
TRANSCRIPT

UNDERSTANDING BIOMECHANICAL CAUSES AND FUNCTIONAL
MECHANISM OF TREATMENT FOR STIFF-KNEE GAIT IN CEREBRAL
PALSY
A DISSERTATION
SUBMITTED TO THE DEPARTMENT OF MECHANICAL ENGINEERING
AND THE COMMITTEE ON GRADUATE STUDIES
OF STANFORD UNIVERSITY
IN PARTIAL FULFILLMENT OF THE REQUIREMENTS
FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
Melanie Diane Fox
May 2011

http://creativecommons.org/licenses/by-nc/3.0/us/
This dissertation is online at: http://purl.stanford.edu/vk276ss2747
© 2011 by Melanie Diane Fox. All Rights Reserved.
Re-distributed by Stanford University under license with the author.
This work is licensed under a Creative Commons Attribution-Noncommercial 3.0 United States License.
ii

I certify that I have read this dissertation and that, in my opinion, it is fully adequatein scope and quality as a dissertation for the degree of Doctor of Philosophy.
Scott Delp, Primary Adviser
I certify that I have read this dissertation and that, in my opinion, it is fully adequatein scope and quality as a dissertation for the degree of Doctor of Philosophy.
Thomas Andriacchi
I certify that I have read this dissertation and that, in my opinion, it is fully adequatein scope and quality as a dissertation for the degree of Doctor of Philosophy.
Jessica Rose
Approved for the Stanford University Committee on Graduate Studies.
Patricia J. Gumport, Vice Provost Graduate Education
This signature page was generated electronically upon submission of this dissertation in electronic format. An original signed hard copy of the signature page is on file inUniversity Archives.
iii

iv
ABSTRACT
Many children with cerebral palsy walk with a stiff knee gait, or a reduction
and delay in swing phase knee flexion, which causes tripping or energy-inefficient
compensatory movements. Since over-activity of the rectus femoris muscle is
frequently implicated as the cause, a common treatment is transfer of the distal end of
the rectus femoris from its insertion on the patella to a location behind the knee.
Outcomes, though positive on average, vary among individuals, with some patients
demonstrating unimproved or worsened knee flexion postoperatively. This variability
is due in part to insufficient understanding of the biomechanical causes of stiff-knee
gait and the functional effects of surgical treatment. The goal of this dissertation was
to clarify the causes of stiff-knee gait and examine the biomechanical mechanism of
improvement following rectus femoris transfer surgery.
Swing-phase rectus femoris activity is commonly thought to cause of stiff-knee
gait, despite evidence that many patients have excessive knee extension moments in
preswing rather than swing phase. We compared the effects of preswing to swing
phase activity of the rectus femoris on peak knee flexion in swing by creating and
analyzing musculoskeletal simulations of subjects with stiff-knee gait. We found that
in six out of ten subjects preswing rectus femoris activity had at least a 90% higher
effect on peak knee flexion than swing phase rectus femoris activity, suggesting that
preswing rectus femoris activity is an important factor limiting knee flexion in some
subjects and should be examined to better determine the factors leading to stiff-knee
gait.

v
To understand how other muscles, besides rectus femoris, may limit knee
flexion in stiff-knee gait, it is first necessary to understand how muscles coordinate
successful swing phase knee flexion in unimpaired gait and how muscle contributions
change with walking speed, since many stiff-knee subjects walk slowly. We analyzed
simulations of unimpaired subjects walking at different speeds to determine the
muscles that accelerated and decelerated knee flexion prior to swing. We found that
preswing knee flexion acceleration was achieved primarily by the hip flexor muscles
with help from biceps femoris short head, suggesting that weakness in these muscles
may contribute to stiff-knee gait. Vasti and soleus decelerated knee flexion, suggesting
over-activity in these muscles may contribute to stiff-knee gait.
We also investigated the mechanism of improvement following rectus femoris
transfer surgery. We altered the geometry of rectus femoris and simulated the
dynamics of the swing phase of subjects with stiff-knee gait after different surgical
procedures. Analysis of the simulations demonstrated that knee flexion may be
improved with a reduction of the knee extension moment generated by the rectus
femoris, even if the muscle is not converted to a knee flexor.
This dissertation clarifies preswing rectus femoris activity as a cause of stiff-
knee gait, demonstrates the functional mechanism of improvement following transfer
surgery, and informs future research investigating other potential contributors to stiff-
knee gait.

vi
ACKNOWLEDGEMENTS
I have been indescribably blessed to have had the guidance, support, and
collaboration of so many talented and wonderful people during my time at Stanford.
First, I am incredibly grateful to my advisor, Scott Delp, whose excellent guidance,
genuine concern, and infectious confidence make him an outstanding mentor. It has
truly been a privilege to work with him.
I am also extremely thankful for the collaborations of my co-authors, Allison
Arnold, Sylvia Ounpuu, and Jeffrey Reinbolt, who made this work possible and
contributed to the high quality of research. I owe a particular debt of gratitude to
Jeffrey Reinbolt, not only for his vital contributions to the work presented here, but
also for his early guidance and patience and his continued support. I am appreciative
of the excellent work done by Mike Schwartz and May Liu, who collected the data
and created the simulations which we analyzed in Chapter 5 of this dissertation. I am
also thankful for the generosity of my NMBL labmates who have always been willing
to give excellent feedback and ask tough questions on presented research. The NMBL
group is full of remarkable talent, and I am grateful to have had the chance to learn in
this environment. I am especially grateful to Kat Steele, Jen Hicks, and Chand John
for their valuable input, brainstorming, and research help.
I would like to acknowledge the funding sources that made this research
possible. A National Science Foundation Graduate Fellowship and grants from the
National Institutes of Health, including R01 HD046814, T32GM63495, and
T15LM7033 funded this research. The musculoskeletal images used in Chapter 2 are

vii
taken with permission from the University of Washington Musculoskeletal Atlas: A
Musculoskeletal Atlas of the Human Body by Carol Teitz, M.D. and Dan Graney,
Ph.D.
My time at Stanford would not have been nearly as fun or fulfilling without
Mandy Koop, Melinda Cromie, and Ariel Dowling. I could always count on these
ladies for everything from late night robot building sessions to crazy adventure races
to spiritual support. Finally, I am exceedingly grateful to my parents, Art and Mary,
and my sisters, Cristina and Lisa, for their love and support. I appreciate their endless
supply of encouragement, as well as their discernment in recognizing those times
when not to ask how research was going. Though over 3000 miles away, they stood
beside me throughout this journey.

viii
CONTENTS Abstract .......................................................................................................................... iv
Acknowledgements ....................................................................................................... vi
List of Tables .................................................................................................................. x
List of Figures ................................................................................................................ xi
1 Introduction ................................................................................................................. 1
1.1 Focus of the Dissertation ...................................................................................... 3
1.2 Significance .......................................................................................................... 4
1.3 Thesis Overview ................................................................................................... 8
2 Background ................................................................................................................ 10
2.1 Knee Flexion in Normal Gait .............................................................................. 10
2.2 Stiff-Knee Gait in Children with Cerebral Palsy: Causes and Treatments ......... 11
2.2.1 Rectus femoris over-activity ........................................................................ 12
2.2.2 Vasti over-activity ........................................................................................ 30
2.2.3 Ankle mechanics .......................................................................................... 32
2.2.4 Insufficient hip flexion moment ................................................................... 35
2.2.5 Hamstrings over-activity .............................................................................. 36
2.2.6 Crouch gait ................................................................................................... 39
2.2.7 Other potential causes .................................................................................. 42
2.3 Methodology: Using Simulation to Understand Muscle Function .................... 43
2.3.1 Simulation of Gait ........................................................................................ 44
2.3.2 Analysis of Simulations ............................................................................... 46
3 Importance of Preswing Rectus Femoris Activity in Stiff-knee Gait ........................ 49
3.1 Abstract ............................................................................................................... 49
3.2 Introduction ......................................................................................................... 50
3.3 Methods .............................................................................................................. 52
3.4 Results ................................................................................................................. 59
3.5 Discussion ........................................................................................................... 60
4 Mechanisms of Improved Knee Flexion After Rectus Femoris Transfer Surgery .... 67
4.1 Abstract ............................................................................................................... 67

ix
4.2 Introduction ......................................................................................................... 68
4.3 Methods .............................................................................................................. 70
4.4 Results ................................................................................................................. 77
4.5 Discussion ........................................................................................................... 79
5 Contributions of Muscles and Passive Dynamics to Swing Initiation Over a Range of Walking Speeds ............................................................................................................ 83
5.1 Abstract ............................................................................................................... 83
5.2 Introduction ......................................................................................................... 84
5.3 Methods .............................................................................................................. 86
5.4 Results ................................................................................................................. 90
5.5 Discussion ........................................................................................................... 94
6 Preliminary work: rectus femoris velocities before and after rectus femoris lengthening surgery ...................................................................................................... 99
6.1 Introduction ......................................................................................................... 99
6.2 Methods .............................................................................................................. 99
6.3 Results ............................................................................................................... 103
6.4 Discussion ......................................................................................................... 106
7 Conclusion ............................................................................................................... 110
7.1 Summary ........................................................................................................... 110
7.2 Future work ....................................................................................................... 112
8 References ............................................................................................................... 114

x
LIST OF TABLES Table 3.1 Descriptive values for stiff-knee and able-bodied subjects......................... 53
Table 3.2 Rectus femoris electromyography deviations among subjects................... 55
Table 6.1 Multivariate model predicting change in knee range of motion after surgery.........................................................................................................................105

xi
LIST OF FIGURES Figure 2.1 Rectus femoris muscle…………………………………. ……………..... 13
Figure 3.1 Simulation of gait during preswing through early swing ……………..... 56
Figure 3.2 Method used to determine increase in peak knee flexion when rectus femoris activity was eliminated during preswing or early swing………………….… 58
Figure 3.3 Bar graph of increases in peak knee flexion caused by eliminating rectus femoris activity during preswing or early swing …………........................................ 60
Figure 3.4 EMG from two subjects with varying levels of preswing and early swing rectus femoris activity.……………………………………………………………….. 62
Figure 4.1 Knee flexion over gait cycle of stiff-knee subjects ……………..........… 72
Figure 4.2 Illustrations and moment arms of musculoskeletal models of rectus femoris transfer………………………………………………………………………. 75
Figure 4.3 Illustrations of peak knee flexion resulting from simulations of rectus femoris transfers………………………………………….......................................... 76
Figure 4.4 Bar graph of increase in peak knee flexion after simulated rectus femoris transfers……………………………………………………….................................... 78
Figure 4.5 Bar graph of change in peak knee flexion due to hip and knee moments of rectus femoris……………………………………………………………………..….. 78
Figure 5.1 Comparison of experimental and simulated knee kinematics and depiction of superposition of perturbation analysis for one subject……………………………. 89
Figure 5.2 Change in knee kinematics with walking speed……………..………….. 91
Figure 5.3 Bar graph of contributions of leg muscles and passive dynamics to knee flexion acceleration…………………………………………...................................... 93
Figure 5.4 Bar graph of contributions of individual muscle on preswing leg to knee flexion acceleration ...……………………………………….......................................94
Figure 6.1 Average knee flexion angles of subject and control groups ...................100
Figure 6.2 Estimated rectus femoris musculotendon lengths and velocities for a representative subject compared to average of controls ……....................................102
Figure 6.3 Preoperative peak rectus femoris lengthening velocities versus change in peak rectus femoris lengthening velocities after surgery ...........................................104
Figure 6.4 Preoperative peak rectus femoris lengthening velocities versus change in knee range of motion after surgery .……………………………...............................104

1
1 INTRODUCTION
Walking is an important skill of daily living that enables independent mobility
in diverse environments and provides myriad health benefits. Particularly for children,
ambulation plays an important role in social development. Walking is also indicative
of general health. In fact, walking speed is predictive of mortality in older adults
(Studenski et al., 2011).
Gait pathologies limit or impair mobility, which can reduce quality of life and
lead to secondary health problems. Left untreated, gait pathologies can become worse
over time and result in the inability to ambulate due to prohibitive energy cost or
intolerable pain.
Determining effective treatment of gait abnormalities is challenging.
Currently, treatment plans are typically based on interpretation of gait and physical
exam data and clinical intuition. These data are often insufficient to identify the
underlying causes of gait impairments. Patients frequently exhibit many gait
abnormalities simultaneously, and it is not always possible to determine which
abnormal findings are primary pathologies and which are secondary compensations.
The functional mechanisms of treatments are not well understood. As a result,
treatment outcomes are highly variable. Many available treatments for children with
cerebral palsy target muscles, such as strengthening, surgical lengthening, and surgical
transfer, but without rigorous understanding of how individual muscles contribute to
whole body movement, treatment may target the wrong muscles or alter them
inappropriately. Understanding the biomechanics of impaired gait and the functional

2
effects of treatments will help clinicians match appropriate treatments to individual
patients, leading to improved and more consistent outcomes.
Computer simulation is a powerful tool for understanding muscle function
during walking. Simulation of walking allows estimation of quantities that cannot be
measured, such as how much a muscle contributes to motion at all joints. It is also
valuable in elucidating cause and effect relationships, such as determining whether
inappropriate activity of a particular muscle could cause an observed gait abnormality.
This dissertation uses computer simulation to examine stiff-knee gait, a
common abnormal gait pattern among children with spastic cerebral palsy. Cerebral
palsy is a neuromuscular disorder resulting from non-progressive damage to the
developing brain which can lead to impaired motor control, abnormal muscle
physiology, and bone deformities. Stiff-knee gait is the inability to properly flex the
knee during the swing phase of gait. This makes it difficult to clear the toe from the
ground and frequently results in tripping or energy-inefficient compensatory motions,
which may make walking unsafe or exceedingly difficult (Mattsson and Brostrom,
1990; Lage et al., 1995; Abdulhadi et al., 1996). The cause is generally thought to be
inappropriate activity of the rectus femoris muscle (Sutherland et al., 1975; Waters et
al., 1979; Perry, 1987; Sutherland et al., 1990; Renshaw et al., 1995); however, it is
unclear at what point in the gait cycle activity from this muscle most impacts swing
phase knee flexion. The most common treatment for stiff-knee gait is surgical transfer
of the rectus femoris muscle in an effort to convert it from a knee extensor to a knee
flexor. However, outcomes of the surgery are variable and the mechanism by which
the surgery causes improvement in some subjects is unclear. Additionally, it is not

3
known whether other biomechanical factors may contribute to stiff-knee gait, since it
has been difficult to determine how muscles coordinate successful swing phase in
unimpaired gait.
1.1 FOCUS OF THE DISSERTATION
The first goal of this dissertation was to identify the biomechanical factors
contributing to stiff-knee gait in children with cerebral palsy. We clarified the
understanding of a currently accepted cause of stiff-knee gait by creating subject-
specific simulations of subjects with stiff-knee gait and altering the simulated muscle
excitations of rectus femoris during preswing and early swing. We demonstrated that
rectus femoris activity during preswing, though not traditionally recognized as a cause,
contributed to stiff-knee gait in many subjects. We also analyzed simulations of eight
unimpaired subjects walking at four speeds to quantify how muscles and passive
dynamics coordinate successful preparation for swing phase. This identified other
potential causes of stiff-knee gait in children with cerebral palsy.
The second goal of this dissertation was to reconcile conflicting experimental
evidence about the functional effect of surgical transfer of the rectus femoris. We
modeled the transferred geometry of the muscle and simulated its effect on knee
flexion in subjects with stiff-knee gait to show that even if the muscle is not converted
to a knee flexor, as intended, substantial improvement in knee flexion may be attained
through reduction of the muscle’s hindering knee extension effect.

4
1.2 SIGNIFICANCE
The work presented in this dissertation contributes significantly to both the
clinical and biomechanical communities. To the clinical community, this work further
clarifies the causes of stiff-knee gait. It provides modified clinical indications for
surgical transfer of rectus femoris and it proposes other possible causes of stiff-knee
gait in cerebral palsy to direct future research. Additionally, this work clarifies the
mechanism of improvement following rectus femoris transfer surgery. This informs
surgical technique and postoperative rehabilitation that could lead to improved
outcomes.
This work also contributes to the biomechanical community. It provides a
comprehensive understanding of how muscles and passive dynamics contribute to
coordinating a successful swing phase in unimpaired gait at different walking speeds.
This adds to the general understanding of muscle function in normal gait.
Additionally, we have provided new computational tools and methods for
investigating human walking.
The primary contributions of the research presented in this dissertation are:
• Creation of ten muscle-actuated simulations that accurately represent the
dynamics of children with cerebral palsy walking with a stiff-knee gait
We created some of the first muscle-actuated simulations of impaired gait. These
simulations allowed us to compare the effects of preswing and early swing rectus
femoris activity on swing phase knee flexion in stiff-knee gait. They also allowed
us to simulate the effects of different types of rectus femoris transfer surgery on

5
knee flexion. Additionally, they provide a resource for future investigation of
muscle function and compensations in stiff-knee gait and are available at
https://simtk.org/home/stiffknee.
• Identification of preswing rectus femoris activity as a contributor to stiff-knee
gait
Swing phase rectus femoris activity has classically been implicated as the cause of
stiff-knee gait. However, investigation of joint moments in individuals with
cerebral palsy walking in a stiff-knee gait showed that many patients had high
knee extension moments in preswing rather than the swing phase of gait (Goldberg
et al., 2006). Our study demonstrated that preswing rectus femoris activity can
contribute to stiff-knee gait and in many subjects, had a more limiting effect on
knee flexion in swing than swing phase rectus femoris activity. This finding helps
to clarify surgical indications for rectus femoris treatment. Traditionally, only
swing phase EMG has been considered an indication for rectus femoris treatment,
but our results suggest that preswing rectus femoris activity may also be an
indication for rectus femoris treatment. Additionally, comparing patient rectus
femoris EMG to speed-matched unimpaired EMG may assist in diagnosing
improper preswing rectus femoris activity since unimpaired individuals typically
exhibit preswing rectus femoris activity at free and fast speeds, while patients with
stiff-knee gait tend to walk at slower speeds. It should, however, be noted that
surface EMG may be subject to cross-talk from the vasti (Barr et al., 2010).

6
• Development of musculoskeletal models of three different types of rectus
femoris transfer
Modeling the transferred geometry of the rectus femoris muscle allowed
evaluation of the mechanism of knee flexion improvement following surgery.
These models offer the opportunity to answer a broad range of questions of clinical
interest regarding biomechanical effect of the surgery. For example, they could be
used to investigate other aspects of transfer surgery, such as effect on the muscle’s
force-length relationship, they could be combined with models of surgical
treatment to other muscles to investigate multilevel surgery, and they could be
combined with a finite element approach and a model of scar tissue to investigate
force transmission in scarred transfers as well as the role fascial connections play
in force transmission.
• Quantitative evidence that the mechanism of improved swing phase knee
flexion following rectus femoris transfer is reduction of the muscle’s knee
extension moment
Rectus femoris transfer was developed with the intention of converting the
muscle’s knee extension moment, which hinders swing phase knee flexion, to a
knee flexion moment by relocating its insertion behind the knee (Perry, 1987).
However, experimental evidence has shown that the transferred muscle still
generated an extension moment in many subjects (Riewald and Delp, 1997;
Asakawa et al., 2002). We modeled the transferred muscle and simulated the effect
on swing phase knee flexion to determine that the mechanism of improved swing

7
phase knee flexion following transfer was reduction of the muscle’s knee
extension moment. This is clinically significant because it suggests that less
invasive methods of reducing the muscle’s knee extension moment, such as
lengthening surgery, may be comparably effective. It also suggests that
mechanisms of reducing scar tissue formation between the rectus femoris and the
underlying vasti has the potential to improve surgical efficacy.
• Modification of an existing algorithm to quantify contributions of Coriolis
and centrifugal accelerations to joint motion
An existing algorithm allowed quantification of joint accelerations induced by
forces from muscles and gravity while accounting for the portion of the ground
reaction force generated by each (Liu et al., 2006a). We modified this algorithm to
enable quantification of joint accelerations induced by Coriolis and centrifugal
accelerations along with their contributions to the ground reaction force. With this
addition, all system forces were accounted for allowing confidence in the analysis
by summing all contributors to knee angular acceleration and comparing the total
to the measured knee angular acceleration. Furthermore, since our analysis
quantified the contributors at different speeds, calculation of velocity-dependent
terms was an important factor.
• Generation of a quantitative comprehensive understanding of how muscles
and passive dynamics accelerate the knee into flexion during preswing

8
While rectus femoris over-activity is the most commonly accepted cause of stiff-
knee gait, variability in treatment outcomes and research in the stroke population
suggest that other factors may contribute to stiff-knee gait. Understanding how
muscles and passive dynamics contribute to swing initiation in unimpaired gait is
necessary to identifying possible causes of an individual’s stiff-knee gait.
Additionally, it is necessary to understand how these contributions may change
with speed, since many subjects with stiff-knee gait tend to walk at slower speeds.
This analysis provides a framework for investigating other possible causes of stiff-
knee gait. It also adds to the understanding of muscle function in normal gait.
1.3 THESIS OVERVIEW
This dissertation is comprised of five subsequent chapters. Chapter 2 presents a
summary of the literature surrounding stiff-knee gait and presents the key clinical
questions. It also contains an explanation of the perturbation technique used to analyze
muscle function. Chapters 3 through 5 are self-contained journal articles, resulting in
some redundant presentation of introduction and methodology. Chapter 3 (Reinbolt et
al. (2009), published in the Journal of Biomechanics) presents a study using muscle-
actuated simulations of stiff-knee gait to demonstrate that rectus femoris activation in
preswing contributes to stiff-knee gait. Chapter 4 (Fox et al. (2009), published in the
Journal of Biomechanics) describes the creation of musculoskeletal models of
transferred rectus femoris and simulation of resulting knee flexion to identify
reduction of the muscle’s knee extension moment as the mechanism of improvement
following surgery. Chapter 5 (Fox et al. (2010), published in the Journal of

9
Biomechanics) details the use of a modified perturbation algorithm to quantify
contributions from muscles and passive dynamics to knee flexion acceleration during
double support at different walking speeds. Chapter 7 summarizes the main
conclusions of the research and suggests future directions for study.
In appreciation of the invaluable contributions of my collaborators to this research, I
use the pronoun “we” throughout this dissertation to refer to the multiple coauthors of
each study. The principle contributors to each of the presented studies include:
Chapter 3: Jeffrey Reinbolt, Allison Arnold, Sylvia Ounpuu, Scott Delp
Chapter 4: Jeffrey Reinbolt, Sylvia Ounpuu, Scott Delp
Chapter 5: Scott Delp

10
2 BACKGROUND
2.1 KNEE FLEXION IN NORMAL GAIT
The gait cycle is divided into phases defined by foot contact with the ground.
Each leg undergoes a period of stance, in which the foot is in contact with the ground,
and a period of swing, in which the foot is off of the ground. At the beginning of
single limb stance, the foot contacts the ground and the knee undergoes mild flexion,
to approximately 20 degrees, absorbing energy from the impact. The leg extends
during mid-stance, acting as a strut while the weight of the body passes over it. As the
opposite leg comes into contact with the ground at approximately 50% of the gait
cycle, the double support phase begins, an important period in preparation for swing
phase. During the double support, or preswing, phase the leg flexes rapidly reaching a
maximum knee flexion velocity around toe-off at 60% of the gait cycle, as the foot
leaves the ground, of approximately 340 degrees per second (Goldberg et al., 2006).
The knee reaches maximum flexion of approximately 60 degrees during swing phase
at approximately 10% of the gait cycle after toe-off. Sufficient knee flexion velocity at
toe-off is required for the knee to achieve adequate flexion to clear the toe from the
ground without stubbing the toe or necessitating out-of-plane compensatory motions.
Rectus femoris is normally active briefly at toe-off in normal gait which is
thought to prevent excess knee flexion in swing at free and fast walking speeds and to
contribute to hip flexion (Perry, 1992). Nene et al. (1999) found a linear relationship
between the angular acceleration of the shank and the amount of rectus femoris
activity in unimpaired gait.

11
2.2 STIFF-KNEE GAIT IN CHILDREN WITH CEREBRAL PALSY: CAUSES AND
TREATMENTS
Cerebral palsy is diagnosed in approximately 3 out of every 1,000 children
each year (CDC, 2004). This condition is a result of damage to the motor control areas
of the brain during development. Children with cerebral palsy commonly exhibit loss
of selective motor control, muscle weakness, exaggerated muscle stretch reflexes,
shortened muscle-tendon units, and subsequent bone deformities, which can make
walking very difficult. One of the most common walking abnormalities among
individuals with cerebral palsy is “stiff-knee gait,” or the inability to properly flex the
knee during the swing phase of gait (Wren et al., 2005a). Though precise kinematic
definitions differ stiff-knee gait has been identified by abnormal swing phase knee
kinematics including diminished and delayed peak knee flexion, reduced knee range
of motion, and difficulty with toe-clearance (Gage et al., 1987; Sutherland et al., 1990;
Kerrigan et al., 1991; Sutherland and Davids, 1993; Kerrigan et al., 1999; Rodda et al.,
2004; Wren et al., 2005a). This condition can lead to injury from tripping or energy-
inefficient compensatory movements such as circumduction, vaulting, upward pelvis
tilt, and pelvic lag to clear the stiff leg from the floor (Mattsson and Brostrom, 1990;
Lage et al., 1995; Abdulhadi et al., 1996). Stiff-knee gait is common not only in
cerebral palsy, but also in stroke, spinal cord injury, and traumatic brain injury
populations. Much of the literature investigating causes of stiff-knee gait has studied
these other populations. Stiff-knee gait is one of the most common gait abnormalities
in ambulatory children with spastic cerebral palsy (Wren et al., 2005a), making it a
valuable area of research aimed at improving treatments outcomes.

12
There has been much investigation into stiff-knee gait aimed at characterizing
the condition, probing the causes, investigating functional effects of treatments, and
assessing clinical treatment outcomes. Yet, the question of how to improve treatment
outcomes of patients with stiff-knee gait remains unanswered. Why do some knees
remain stiff after treatment? Failed treatment outcomes are either due to inaccurate
identification of the cause or ineffective treatment. To improve outcomes we must
understand more thoroughly both the causes of stiff-knee gait and the functional
effects of treatments.
2.2.1 RECTUS FEMORIS OVER-ACTIVITY
Rectus femoris over-activity has been considered historically to be the cause of
stiff-knee gait and remains the most commonly cited and treated cause of stiff-knee
gait (Sutherland et al., 1975; Waters et al., 1979; Perry, 1987; Sutherland et al., 1990;
Renshaw et al., 1995). In unimpaired gait, the rectus femoris is active briefly at toe-
off, which is thought to prevent excess knee flexion in swing and flex the hip (Perry,
1987; Perry, 1992; Nene et al., 1999; Schwartz et al., 2008). Nene et al. (1999)
showed that in unimpaired gait, the magnitude of rectus femoris activity was linearly
related to the angular acceleration of the shank in early swing. It is thought that
inappropriate activity of the rectus femoris, either in magnitude or in timing, could
result in an excessive knee extension moment and restrict knee flexion in swing
(Perry, 1987). Over-activity of the rectus femoris may be due to a spastic reflex
response triggered by rapid knee flexion near toe-off (Jonkers et al., 2006). Others
have suggested that the spastic response of the rectus femoris could be triggered when

13
the hip reaches maximum extension near terminal stance (Silfverskiold, 1923). In
stroke patients, others have suggested that a multi-joint, heteronymous stretch reflex
triggered by extension of hip extensors may result in inappropriate activation of the
knee extensors (Lewek et al., 2007). These authors found that imposed hip extension
resulted in greater reflex responses of the rectus femoris and vatus lateralis in subjects
with stroke as compared to controls and was correlated with decreased knee flexion
during swing (Lewek et al., 2007).
Figure 2. 1 Rectus femoris, part of the quadriceps muscle group, is a bi-articular muscle generating a hip flexion moment and a knee extension moment. (Copyright 2003-2004, University of Washington. All rights reserved including all photographs and images. No re-use, re-distribution or commercial use without prior written permission of the authors and the University of Washington.)

14
The timing of rectus femoris activity in preswing has been identified as a
contributor to stiff-knee gait. However, rectus femoris over-activity during swing is
generally cited as the cause of stiff-knee gait and is considered as an indication for
surgical treatment (Sutherland et al., 1990; Chung et al., 1997; Miller et al., 1997;
Yngve et al., 2002; Saw et al., 2003; Kay et al., 2004; Muthusamy et al., 2008). Rectus
femoris activity during preswing, before toe-off, has not historically been considered
to cause stiff-knee gait because the rectus femoris is active during this time in
unimpaired subjects at self-selected walking speeds (Perry, 1992; Nene et al., 1999;
Schwartz et al., 2008). However, many subjects with stiff-knee gait walk at slow
speeds and the rectus femoris is not typically active during preswing in unimpaired
subjects walking at slow speeds (Schwartz et al., 2008). Additionally, a recent study
has shown that many subjects with cerebral palsy who walk with a stiff-knee gait
exhibit excessive knee extension moments during preswing, not swing (Goldberg et
al., 2006). Also, it has been observed clinically that inappropriate swing phase rectus
femoris activity exists in patients who do not exhibit a stiff-knee gait (DeLuca et al.,
1997).
Musculoskeletal simulation has provided evidence that preswing rectus
femoris activity can contribute to stiff-knee gait. Simulations of unimpaired gait
demonstrated that sufficient knee flexion velocity at toe-off is necessary to achieve
adequate knee flexion during swing (Piazza and Delp, 1996; Anderson et al., 2004).
Reduced knee flexion velocity at toe-off is exhibited in many individuals with stiff-
knee gait (Goldberg et al., 2006) and can be caused by preswing rectus femoris
activity (Goldberg et al., 2004). Finally, simulations of stiff-knee gait have suggested

15
that restoring knee flexion velocity at toe-off to normal values can restore peak knee
flexion in swing (Goldberg et al., 2003) and that preswing rectus femoris activity is at
least as influential as early swing rectus femoris activity in limiting knee flexion in
swing (Reinbolt et al., 2008, chapter 3 of this dissertation).
Experimental evidence has also demonstrated that preswing rectus femoris
activity can contribute to stiff-knee gait. Electrical stimulation of the rectus femoris
during treadmill walking in unimpaired subjects demonstrated that excessive rectus
femoris activity during preswing caused greater reduction in swing phase peak knee
flexion than excessive rectus femoris activity during swing (Hernandez et al., 2010).
Following injection of botulinum toxin injection into the rectus femoris in adults with
stiff knee gait due to stroke or traumatic brain injury, subjects showed an increase in
knee flexion velocity at toe-off suggesting that preswing rectus femoris activity had
been contributing to stiff-knee gait by limiting knee flexion velocity at toe-off
(Robertson et al., 2009). A group of subjects with cerebral palsy with rectus femoris
activity in preswing demonstrated improved peak knee flexion after rectus femoris
transfer surgery leading the authors to suggest that either rectus femoris activity in
preswing was contributing to stiff-knee gait, or another concomitant surgery had
improved stiff-knee gait (Miller et al., 1997). Collectively, this research suggests that
rectus femoris over-activity during preswing can limit peak knee flexion in swing.
Rectus femoris transfer
Rectus femoris transfer, proposed by Perry (1987) and Gage et al. (1987), is
the most common treatment for stiff-knee gait (Chambers, 2001). The surgical

16
procedure consists of release of the distal end of the muscle with its component of the
quadriceps tendon, dissection away from the underlying vasti, displacement of the
distal end either medially or laterally (depending on intended transfer site), and
reattachment (Gage et al., 1987; Sutherland et al., 1990; Patrick, 1996; Chambers et
al., 1998). Common transfer sites are medial to sartorius, gracilis, or semitendinosus or
lateral to the iliotibial band.
Mechanism of surgical effect
Rectus femoris transfer was developed with the intention of converting the
rectus femoris to a knee flexor while preserving its hip flexion moment (Perry, 1987;
Sutherland et al., 1990). Experimental study of transfer in cadavers combined with
musculoskeletal modeling showed that rectus femoris had a knee flexion moment arm
after transfer to semitendinosus, gracilis, or sartorius, and an insignificant moment arm
after transfer to the iliotibial band (Delp et al., 1994). However, further investigation
of rectus femoris after transfer has suggested that it is not converted to a knee flexor.
Riewald and Delp (1997) stimulated the transferred muscle in four subjects and found
that it generated a knee extension torque in all subjects. A dynamic imaging study
(Asakawa et al., 2002) measured the motion of the transferred muscle during passive
knee movement and found that the transferred rectus femoris displaced with the knee
extensors, meaning that it still acted as a knee extensor after transfer, though its
velocity relative to the knee extensors, signifying its capacity for knee extension, was
diminished by the surgery. A static imaging study (Asakawa et al., 2004; Gold et al.,
2004) found evidence of scar tissue between rectus femoris and the underlying vasti.

17
They also found that the transferred rectus femoris muscle path from origin to
insertion showed an angular deviation, greater than 35 degrees, in most cases. Force
generated by the rectus femoris may be transferred to the vasti through the scar tissue
connection, resulting in a knee extension moment. Finally, a simulation study
demonstrated that if scarred transfer of rectus femoris reduces its knee extension
capacity by half, improvements in peak knee flexion in swing similar to clinically
reported improvement following isolated rectus femoris transfer (Hemo et al., 2007)
may still be achieved (Fox et al., 2009, chapter 4 of this dissertation). These
simulations also showed that simulated force output of rectus femoris after transfer to
the knee flexors, assuming no scarring, was reduced due to the muscle’s conversion
from operating eccentrically to concentrically, since it was stretched when anterior to
the knee but shortened when posterior to the knee during knee flexion at toe-off (Fox
et al., 2009, chapter 4 of this dissertation).
Though it is clear that reducing scar tissue formation is helpful in achieving
good outcomes following rectus femoris transfer, it is unclear how surgical technique
and postoperative rehabilitation affect the formation of scar tissue . Lipaphayom and
Prasongchin (2011) intra-operatively examined the transferred rectus femoris in three
knees during subsequent surgery 7 – 60 months following the rectus femoris transfer.
In contrast to the previously described imaging studies, they observed minimal scar
tissue formation and a smoothly gliding muscle path and elicited knee flexion by
manual pull on the rectus femoris tendon, which they attributed to surgical technique.
In contrast, surgical revision of rectus femoris transfer presented as a case study for a
single patient found significant scar tissue formation after 4 years (Johnson et al.,

18
2011). Improvement in swing phase knee flexion was achieved in this patient after
release of scar tissue and lengthening of the rectus femoris. Some advocate the use of
continuous passive movement machines and early mobilization postoperatively, but
there are no studies comparing the effects of different postoperative rehabilitation
strategies.
Outcomes
Outcomes following rectus femoris transfer surgery are positive on average,
but variable among individuals. As rectus femoris transfer may be performed to treat
diminished peak knee flexion in swing or to prevent a reduction in peak knee flexion
following hamstrings treatment for crouch gait (DeLuca et al., 1997), knee range of
motion is a relevant outcome metric to compare outcomes from studies with and
without concomitant hamstrings surgeries. All rectus femoris outcome data available
for review reported an average improvement in knee range of motion, with the
exception of Carney et al. (2006) which reported no significant difference on average
in 29 limbs. Among the subject groups that showed an average significant
postoperative improvement, the average amount of improvement in knee range of
motion during gait was 12.8 degrees with a range of 4 degrees to 36 degrees. Other
outcome metrics, though less commonly reported, included decreased mean knee
extensor moment in stance (Adolfsen et al., 2007) and increased knee flexion velocity
surrounding toe-off (Hadley et al., 1992; Muthusamy, 2006).
Different effects were observed following rectus femoris transfer with versus
without concomitant hamstrings lengthening. On average among studies in which

19
hamstrings lengthening was performed with rectus femoris transfer, peak knee flexion
in swing was retained while knee extension in stance was increased, resulting in
improved knee range of motion during gait. Following rectus femoris transfer without
hamstrings lengthening, on average peak knee flexion in swing was increased while
knee extension in stance was decreased to a lesser extent or unchanged, resulting in
improved knee range of motion during gait (Miller et al., 1997; Carney et al., 2006;
Hemo et al., 2007). In addition, Miller et al. (1997) reported an increase in knee
flexion at initial contact when rectus femoris transfer was performed without
hamstrings lengthening.
Change in the timing of peak knee flexion in swing is also an important metric
of the success of rectus femoris transfer surgery. All group averages reported in the
studies examined showed either improvement in timing in peak knee flexion in swing
or no significant change. Among the studies that reported a significant improvement,
the average improvement in timing of peak knee flexion was 3.5% of the gait cycle.
Timing of peak knee flexion in swing has been reported as percent of gait cycle or
percent of swing phase. Van der Linden et al. (2003) have suggested reporting the
outcome as percent of gait cycle since the duration of swing phase may change
following treatment. For the studies reporting timing of peak knee flexion as a
percentage of swing phase, these outcomes have been converted to percent of gait
cycle assuming a swing phase duration of 40% of the gait cycle to allow comparison
across all studies.
Reports of long term outcomes following rectus femoris transfer are few and
varied. The largest study of long-term rectus femoris transfer outcomes reported that

20
peak knee flexion and knee range of motion in swing were relatively stable from years
1 to 3 postoperatively in 50 limbs (Moreau et al., 2005). Similarly, Adolfsen et al.
(2007) found improvements after one year to be maintained after 4 years in 9 limbs.
Saw et al. (2003) also reported maintained improvement in swing phase knee flexion
after 4.6 years in 26 limbs, but reported a decrease in total knee range of motion
during gait. In this study, 9 limbs of 5 subjects developed crouch and required
subsequent hamstrings lengthening; many subjects in this study did not receive
concomitant hamstrings lengthening with the original rectus femoris transfer.
Early investigation hypothesized that the choice of rectus femoris transfer site
may provide additional benefit by correcting transverse plane rotational deformities
(Gage et al., 1987). In specific, rectus femoris transfer medially to sartorius or
semimembranosus was hypothesized to create an external rotation moment at the knee
which would improve excessive internal rotation, while lateral transfer to the iliotibial
band was hypothesized to improve excessive external rotation. No significant
rotational effect has been observed after transfer either medially or laterally (Gage et
al., 1987; Nene et al., 1993; Ounpuu et al., 1993a). In the sagittal plane, no significant
difference in transfer sites has been reported comparing changes in peak knee flexion
in swing, knee range of motion, and knee extension at in stance (Ounpuu et al., 1993a;
Chambers et al., 1998). Some studies have reported differences in a single outcome
metric among transfer sites (peak knee flexion in swing or knee extension in stance),
but have suggested that this may be due to disparate proportions of concomitant
hamstrings lengthenings among the different groups (Muthusamy, 2006; Hemo et al.,
2007; Muthusamy et al., 2008). One study, with relatively equal proportions of

21
hamstrings lengthenings in each group, reported greater improvement in peak knee
flexion after transfer to gracilis compared to sartorius (Chung et al., 1997). They
suggest that this improvement may be a result of their revised technique in transferring
rectus femoris to gracilis, although average improvement in knee range of motion in
this study is less than that reported by Ounpuu et al. (1993a).
Limitations with current outcome studies following rectus femoris transfer
include that many studies do not correct for inclusion of data from two limbs per
subject, many studies do not account for differences in walking speed, some studies
statistically evaluate postoperative values without statistically comparing preoperative
values, and some studies do not take into account changes in height in evaluating
temporal-spatial parameters.
Although rectus femoris spasticity is often considered an indication for rectus
femoris surgery, there have been few studies investigating whether rectus femoris
spasticity actually decreases following rectus femoris surgery. The few studies that
have been conducted have used the Duncan Ely test, rather than a more quantitative
measurement, to assess spasticity. Subjects undergoing rectus femoris transfer did not
all have positive preoperative Duncan Ely tests (86%, Kay et al., 2004; 100%, Hemo
et al., 2007; 42%,Koca et al., 2009). The studies that have reported change in Duncan
Ely scores after surgery report variable results between 43% and 85% rates of
spasticity improvement (50%, Kay et al., 2004; 51%, Adolfsen et al., 2007; 85%,
Hemo et al., 2007; 34%, Koca et al., 2009), suggesting that spasticity, as measured by
Duncan Ely test, may improve in some but not all patients after rectus femoris
transfer. Kay et al. (2004) found that in a group that maintained positive Duncan Ely

22
tests before and after surgery, spasticity decreased, measured by Ashworth scale and
angle at which hip rise occurred during Duncan Ely test. More quantitative
measurements of changes in spasticity following surgery and correlation with surgical
outcome are warranted.
Clinical indications
Surgical indications for rectus femoris transfer differ by institution, but
generally include diminished peak knee flexion, delayed peak knee flexion, reduced
knee range of motion, and impaired foot clearance. Abnormal rectus femoris activity,
such as prolonged activity into swing, or continuous activity throughout the gait cycle,
and rectus femoris spasticity, measured during physical exam by a Duncan Ely test
(Bleck, 1987) are also taken into account.
There are limitations to the current clinical indicators. First, clinical measures
of spasticity may be unreliable. The most common clinical measure of rectus femoris
spasticity is the Duncan Ely test. In this test, the patient lies prone with the hips
extended while the knee is gradually flexed to 130 degrees. If the hip flexes, causing
the buttocks to rise from the table, the rectus femoris is considered spastic. There is
concern that the Ely test may not isolate rectus femoris activity, as iliopsoas activity
may also result in a positive test (Sutherland et al., 1975; Perry et al., 1976). Chambers
et al. (1998) found that the Duncan Ely test had no predictive value for abnormal
rectus femoris activity during gait. Marks et al. (2003) also questioned the relationship
between the Duncan Ely test and kinematic and electromyographic indicators of stiff-
knee gait, though they caution that the high prevalence of rectus femoris dysfunction

23
in their subject population may have affected their results. Another limitation of
current assessment for spasticity during clinical exam is that it may not correlate with
spasticity during gait. Sutherland et al. (1975) noticed anecdotally that a large number
of spastic patients demonstrated a positive Duncan Ely test yet did not exhibit
functional limitations during gait. Kerrigan et al. (1999) observed patients with
quadriceps spasticity on static evaluation that had normal activity during walking. One
possible explanation is that subjects may walk slowly enough to avoid eliciting a
spastic rectus femoris response during gait.
A more useful evaluation of the utility of Duncan Ely may be its predictive
value of outcome after treatment. Some studies have found Duncan Ely to have no
predictive value for surgical outcome (Goldberg et al., 2006; Muthusamy, 2006;
Muthusamy et al., 2008). In contrast, Kay et al. (2004) found Duncan Ely may be a
helpful predictor of outcome after rectus femoris transfer since knee range of motion
and timing of peak knee flexion improved on average only in the group of subjects
with a positive Duncan Ely tests. However, these are average improvements, and
outcomes may vary for individual patients. Other measures of spasticity, including the
Ashworth and modified Ashworth scales, the Tardieu scale, and the pendulum test, do
not isolate rectus femoris spasticity. Better measures of spasticity are required to
evaluate the effects of rectus femoris spasticity on stiff-knee gait and outcomes after
rectus femoris transfer.
Another limitation of current clinical indicators for rectus femoris transfer is
that the usefulness of preoperative rectus femoris activity, measured by EMG, is
unclear. Rectus femoris activity during swing has been observed in conjunction with

24
normal knee kinematics (DeLuca et al., 1997). Several studies reported that rectus
femoris EMG does not have predictive value for surgical outcomes (Chambers et al.,
1998; Saw et al., 2003; Muthusamy, 2006; Muthusamy et al., 2008). In contrast,
Miller et al. (1997) suggested that rectus femoris EMG does have predictive value
since a group with swing phase rectus femoris EMG had greater average improvement
in peak knee flexion than a group with normal rectus femoris EMG; however, they did
not report differences in preoperative peak knee flexion between the groups, and there
was a small number of subjects in each group. Barr et al. (2010) has cautioned using
surface EMG to measure rectus femoris activity, as it can be subject to crosstalk from
the vasti, particularly during a crouch gait at fast speeds.
Additional clinical indicators have been explored to identify subjects that need
rectus femoris transfer. Reduced preoperative knee range of motion during gait has
been suggested as a predictor for positive outcomes after rectus femoris transfer
(Ounpuu et al., 1993b; Chung et al., 1997; Niiler et al., 2007), though Chambers et al.
(1998) found no relationship between preoperative and postoperative knee range of
motion. Knee flexion velocity at toe-off has been suggested as a possible indicator for
rectus femoris surgery (Muthusamy et al., 2008), though some report greater
improvements in subjects with low knee flexion velocities (Muthusamy, 2006), while
others report greater improvement with high knee flexion velocities (Reinbolt et al.,
2009). Neither of these studies accounted for confounding variables, such as overall
severity, in their analyses, which may contribute to the discrepancy. Goldberg et al.
(2006) found that increases in knee flexion velocity at toe-off were associated with
improvements in stiff-knee gait. Vasti over-activity has been investigated as a

25
contraindication for rectus femoris transfer. Chambers et al. (1998) and Sutherland et
al. (1990) found no difference in outcomes among subjects with cerebral palsy with
and without concomitant abnormal vasti activity. In contrast, Waters et al. (1979)
reported that the amount of improvement in stroke patients with stiff-knee gait
following release of rectus femoris, and vastus intermedius in some subjects, was
dependent on the component of the quadriceps that showed inappropriate EMG in
swing.
Rectus femoris release
Rectus femoris release was developed before rectus femoris transfer as a
treatment for stiff-knee gait, with the intent of improving knee flexion in swing
(Silfverskiold, 1923; Sutherland et al., 1975). It was originally performed as a
treatment for hip flexion contracture (Duncan, 1955; Cottrell, 1963; McMulkin et al.,
2005), and secondarily knee flexion deformity (Duncan, 1955; Cottrell, 1963). Rectus
femoris release may be performed distally or proximally. Concern over the negative
effects of proximal release on hip and pelvis motion (Sutherland et al., 1975) led to the
adoption of distal release (Perry, 1987). The surgery involves dissection of the rectus
femoris away from the underlying vasti, and dissection of the tendon from its
insertion, either proximally or distally. Gage et al. (1987) proposed that inferior results
following rectus femoris release may be due to the muscle retaining its ability to
generate a knee extension moment, possibly due to reattachment to the quadriceps
through scar tissue formation.

26
Outcomes following rectus femoris release are inferior to rectus femoris
transfer. Studies of outcomes after distal rectus femoris release, with many subject
receiving concomitant hamstring lengthening, have reported no changes in knee range
of motion during gait (Ounpuu et al., 1993b; Chambers et al., 1998) and either no
change (Chambers et al., 1998) or a decrease (Ounpuu et al., 1993b) in peak knee
flexion after lengthening. Sutherland et al. (1975) reported increased knee flexion in
swing in six of eight subjects following proximal rectus femoris release, though
outcomes were variable and improvements limited. After distal rectus femoris release
without hamstring lengthening, Sutherland et al. (1990) reported an average increase
in peak knee flexion in swing and knee range of motion during gait, though both were
smaller than improvements after rectus femoris transfer. Studies reported no
improvement on average in timing of peak knee flexion in swing following release
(Sutherland et al., 1990; Ounpuu et al., 1993b; Chambers et al., 1998). Some studies
report that release was accompanied by an increase in crouch (Sutherland et al., 1975),
though others have not observed this (Sutherland et al., 1990; Chambers et al., 1998).
In an adult stroke population undergoing proximal rectus femoris release without
hamstrings lengthening (Waters et al., 1979), average improvement in peak knee
flexion (9.6 degrees) was comparable to rectus femoris distal release combined with
hamstrings lengthening in patients with cerebral palsy (9.1 degrees, Chambers et al.,
1998). The degree of improvement in this stroke population was dependent on the
component of quadriceps that showed inappropriate EMG in swing (i.e., if rectus
femoris was the only active head, improvements were greater).

27
Rectus femoris lengthening
The literature describing rectus femoris lengthening to treat stiff-knee gait is
sparse. Others have described rectus femoris lengthening in an effort to treat hip or
knee flexion deformities (Matsuo et al., 1987; Guerado and de la Varga, 2001), but
there is no journal article describing rectus femoris lengthening to treat stiff-knee gait.
In rectus femoris intramuscular lengthening surgery, incisions are made into the
aponeurosis, the portion of the tendon connected to the muscle fibers. The intended
effect is reduction of muscle spasticity by decreasing the lengthening velocity of the
muscle fibers. The procedure is less invasive than rectus femoris transfer and requires
less dissection, potentially allowing reduced postoperative pain, earlier postoperative
mobilization, and reduction of complications.
The effectiveness of this surgery on the treatment of stiff-knee gait is currently
unclear as there has been only a preliminary report of outcomes. A conference abstract
has reported outcomes following rectus femoris intramuscular lengthening in 72 knees
of 43 subjects, most of which received concomitant hamstrings lengthening (Cruz et
al., 2009). The authors conclude that outcomes are similar to rectus femoris transfer
with an average 2% of gait cycle improvement in timing of peak knee flexion and an
average 2-degree improvement in knee range of motion during gait, though there was
variability among subjects. Rectus femoris lengthening may be an attractive
alternative to rectus femoris transfer if it proves to be as effective, but additional study
is needed to quantify its effectiveness.

28
Neuromuscular block
Injection of neuromuscular toxins into the rectus femoris may be used for
temporary evaluation of treatment options or for longer-term treatment. Temporary
treatments, such as lidocaine, are injected into the vicinity of the target nerve to block
activation of the muscle. For more long term treatment, phenol may be used to destroy
the nerve fibers or botulinum toxin may be injected to inhibit the release of
acetylcholine from the neuromuscular junction (Burgen et al., 1949).
Almost all outcome studies of nerve or motor point block of rectus femoris
have been conducted in the adult population. The only study reporting outcomes after
botulinum toxin injection to rectus femoris in subjects with cerebral palsy found no
appreciable change in dynamic knee range of motion in eight subjects (Chambers,
2001). In stroke, early reports of nerve block to treat stiff-knee gait were generally
unsuccessful in improving knee flexion and left some patients too unstable to walk
(Mooney and Goodman, 1969; Treanor, 1969). More recently, Albert et al. (2002)
performed femoral nerve block in 12 subjects with stiff-knee gait due to CNS injury
and also reported no improvement in gait parameters. However, the majority of
outcome reports following nerve block of rectus femoris report positive impact on
stiff-knee gait. Average improvement in peak knee flexion has been reported
following lidocaine (15 degrees, Sung and Bang, 2000; 11 degrees, Robertson et al.,
2009), phenol (9 degrees, Sung and Bang, 2000) , and botulinum toxin injections (5
degrees, Stoquart et al., 2008; 8 degrees, Robertson et al., 2009). Chantraine et al.
(2005) found an average 5 degree increase in knee range of motion among 6 subjects
after motor branch block with lidocaine. Caty et al. (2008) reported increased peak

29
knee flexion by an average of 5 deg in 20 stroke subjects after botulinum toxin into a
combination of rectus femoris, semitendinosus, and triceps surae.
Most studies report average improvements in knee flexion velocity at toe-off
after injection of lidocaine (0.9 degrees per percent gait cycle, Sung and Bang, 2000;
43 degrees per second, Stoquart et al., 2008), phenol (0.47 degrees per percent gait
cycle, Sung and Bang, 2000), and botulinum toxin (30 degrees per second, Stoquart et
al., 2008; 53 degrees per second, Robertson et al., 2009). In contrast, Chantraine et al.
(2005) found no improvement on average in knee flexion velocity at toe-off among six
subjects after motor branch block with lidocaine. No improvement was measured in
maximum preswing knee moment among subjects (Chantraine et al., 2005) or peak
moments at hip, knee, or ankle (Robertson et al., 2009). However, Robertson et al.
(2009) described two subjects who had excessive preoperative knee extension
moments in preswing that were maintained following botulinum toxin injection, yet
they were able to attain normal peak knee flexion. Duncan Ely scores improved after
nerve block in stroke subjects (Chantraine et al., 2005; Caty et al., 2008; Stoquart et
al., 2008).
Some undesired secondary effects after injection of neuromuscular blocks,
though infrequent, have been reported including weakness of quads or knee buckling
during stair climbing (Sung and Bang, 2000), and one subject whose gait changed
from crouch to jump knee with no improvement in swing phase knee flexion
(Chantraine et al., 2005). Another secondary effect reported after Botulinum toxin
injection into rectus femoris was a reduction in vastus lateralis and biceps femoris
activity (Stoquart et al., 2008). There is debate over whether results of nerve block of

30
rectus femoris depend on activity present in other heads of the quadriceps. Sung and
Bang (2000) reported preswing to midswing quadriceps EMG activity confined to
rectus femoris seemed to result in more improved outcomes in 31 subjects, while
Chantraine et al. (2005) found no relationship in 6 subjects. Severity of quadriceps
spasticity, measured by Ashworth scale, did not influence nerve block effect (Sung
and Bang, 2000). There is also disagreement over whether preoperative knee range of
motion influences outcome following nerve block. Stoquart et al. (2008) reported 4
out of 19 subjects who had less than 10 degrees of knee range of motion before
injection did not improve, whereas Caty et al. (2008) found that higher dosage
injection of botulinum toxin into multiple muscles improved swing phase knee flexion
in subjects with less than 10 degrees of knee range of motion.
2.2.2 VASTI OVER-ACTIVITY
Over-activity of the vasti is a less commonly proposed cause of stiff-knee gait
in patients with cerebral palsy. In cerebral palsy, rectus femoris is more frequently
over-active during gait than the vasti (Csongradi et al., 1979; Sutherland et al., 1990;
DeLuca et al., 1997; Chambers et al., 1998). However, over-activity of the vasti could
increase the knee extensor moment and decrease toe-off velocity at toe-off, similar to
over-activity of the rectus femoris. Musculoskeletal simulation of unimpaired gait has
identified the vasti as having a large potential to decrease knee flexion velocity during
double support (Goldberg et al., 2004).
Experimental evidence suggesting vasti over-activity as a cause of stiff-knee
gait is sparse. Gage et al. (1987) stated that reflex activity in vastus medialis may

31
contribute to stiff-knee gait, but provided no surgical results. In the stroke population,
Kerrigan et al. (1991) observed prevalent inappropriate vasti activity in preswing and
early swing and concluded that this prevalence implied a relationship with stiff-knee
gait; however neither a correlation nor causal relationship was examined. Also in the
stroke population, Waters et al. (1979) performed tenotomy of one or 2 heads of the
quadriceps, based on inappropriate muscle activity during swing. They found that
improvement in swing phase peak knee flexion was larger when all heads of the quads
with inappropriate activity were released, suggesting that vasti may contribute to stiff-
knee gait. However, it should be noted that the group with inappropriate muscle
activity in fewer heads of the quadriceps may have been less involved, which may
have contributed to the difference in outcomes between the groups. In opposition to
this evidence, Chambers et al. (1998) found no difference in outcome of subjects with
cerebral palsy after rectus femoris surgery (transfer in most, release in others) between
a group with only abnormal rectus femoris activity alone and a group with abnormal
activity in both rectus femoris and vastus lateralis.
The vasti are not a common target of treatment for stiff-knee gait in cerebral
palsy, and there is no quantitative report of outcomes following vasti treatment in
cerebral palsy. It has been suggested that surgical treatment of the vasti may
compromise knee stability in stance (Sutherland et al., 1975; Waters et al., 1979).
Namdari et al. (2010) performed rectus femoris transfer with vastus lengthening in 37
subjects with stiff-knee gait due to stroke or traumatic brain injury and reported an
average increase of 25 degrees in peak knee flexion in 21 subjects. Hebela and Keenan
(2004) described fractional lengthening of over-active vasti in subjects with upper

32
motor neuron syndromes, but did not report quantitative outcomes. Waters et al.
(1979) performed tenotomy of one or two heads of the quadriceps in unilateral stiff-
knee stroke subjects, based on inappropriate muscle activity during swing. They
reported variable changes in swing phase peak knee flexion with an average of 10
degrees and a range of -21 to 30 degrees.
2.2.3 ANKLE MECHANICS
Another proposed cause of stiff-knee gait is abnormal ankle mechanics. Two
mechanisms have been proposed describing how abnormal ankle mechanics may limit
knee flexion in swing. The first is walking in equinus, or excessive ankle
plantarflexion. Kerrigan et al. (2001a) reported that normal subjects performing toe-
walking experienced a significant reduction in peak knee flexion (from 59 to 42
degrees), suggesting that toe-walking may contribute to stiff-knee gait. They also
acknowledged that not all patients who toe-walk have stiff-knee gait.
The second mechanism proposed in the literature is that insufficient ankle
plantarflexion moment before toe-off may limit swing phase knee flexion through
dynamic coupling. Many stiff-knee subjects have diminished ankle plantarflexion
moments (Kerrigan et al., 2001b; Goldberg et al., 2006; Robertson et al., 2009).
However, Goldberg et al. (2006) noted that ankle plantarflexion moments during
double support did not correlate with knee flexion velocity at toe-off in 23 subjects
with stiff-knee gait and cerebral palsy. It is unclear whether low plantarflexion
moments might be due to weak gastrocnemius or weak soleus which have potentially
contradictory effects on knee motion.

33
The effect of preswing plantarflexion moment on swing phase knee flexion is
unclear. Though simulation studies of normal gait agree that soleus activity in
preswing accelerates the knee into extension (Yamaguchi and Zajac, 1990; Neptune et
al., 2001; Fox and Delp, 2010, chapter 5 of this dissertation), simulation evidence for
gastrocnemius function has been more variable. Fox and Delp (2010, chapter 5 of this
dissertation) found that gastrocnemius induced a small knee flexion acceleration,
though its effect on the knee was variable, while Yamaguchi and Zajac (1990)
reported a large knee flexion effect. Neptune et al. (2001) found that gastrocnemius
contributed to knee extension acceleration in preswing. Since the bi-articular
gastrocnemius generates both a plantarflexion moment that induces knee extension
acceleration and a knee flexion moment that induces knee flexion acceleration, its
action is sensitive to the muscle’s ankle and knee moment arms, body position, and
foot contact model, which varied among the studies.
Experimental study using surface electrodes to stimulate muscle during gait
(Stewart et al., 2007) found that soleus caused ankle plantarflexion and knee extension
while gastrocnemius caused ankle dorsiflexion and knee flexion during preswing in
five adult subjects. Riley and Kerrigan (1999) used simulation to investigate the effect
of plantarflexion moment on the knee in stiff-knee stroke subjects, but did not account
for the effect of the portion of the ground reaction force that was induced by the ankle
moment on the knee.
Although the net effect of the plantarflexors may be to extend the knee, studies
have suggested that they play a role in swing initiation. It has been observed that hip
flexors compensate during preswing when plantarflexors are weak due to stroke

34
(Nadeau et al., 1999) or absent due to amputation (Zmitrewicz, 2007). Neptune et al.
(2008) suggested that gastrocnemius contributes to swing initiation by delivering
energy to the preswing leg. Van der Krogt et al. (2010) created forward dynamic
simulations of stable cyclic walking and found that magnitude of the push off impulse
increased swing phase knee flexion. Evidence for ankle plantarflexion moment deficiency as a cause of stiff-knee
gait has been presented in the stroke population. Kerrigan and Glenn (1994) conducted
an interventional study in a spinal cord injury patient who walked with a stiff-knee
gait. The patient was treated with an ankle-foot orthosis to resist dorsiflexion and an
exercise program to strengthen gastrocnemius and soleus. After two weeks, the patient
showed improved peak knee flexion (from 22 to 35 degrees) though still diminished
from normal, while walking speed remained unchanged. Plantarflexion strength was
not shown to be increased by manual muscle test, but ankle plantarflexion moment in
preswing was increased. It is unclear whether the follow-up gait analysis was
conducted with or without the prescribed ankle-foot orthosis. In another study,
Kerrigan et al. (1991) also observed delayed heel rise in 21 of 23 subjects with stiff-
knee gait due to stroke or head injury. The authors proposed that delayed heel rise may
be indicative of plantarflexor weakness and proposed a relationship between delayed
heel rise and peak knee flexion in swing, though no quantitative correlation was
performed. Since no intervention was performed, it is unclear whether delayed heel
rise was a cause of stiff-knee gait, a compensation, or an unrelated abnormality. More
research is needed to understand the functions of soleus and gastrocnemius in swing

35
initiation and to determine whether dysfunction of either muscle can contribute to
stiff-knee gait.
2.2.4 INSUFFICIENT HIP FLEXION MOMENT
Insufficient hip flexion moment has been proposed as a cause of stiff-knee gait
in stroke patients (Kerrigan et al., 1999). Through dynamic coupling, a hip flexion
moment induces a knee flexion acceleration. Simulation studies agree that adequate
preswing hip flexion moment is important for knee flexion in swing in normal gait
(Yamaguchi and Zajac, 1990; Piazza and Delp, 1996; Neptune et al., 2008; Fox and
Delp, 2010, chapter 5 of this dissertation). The preswing hip flexion moment is not
generally diminished among stiff-knee subjects, but there is variability among
subjects. All 23 subjects with stiff-knee gait and cerebral palsy in a study by Goldberg
(2006) had both preswing and swing phase hip flexion moments that were within two
standard deviations of normal. In fact, many subjects had excessive hip flexion
moments in double support, which the authors suggested could be due to a delay in
peak hip flexion moment which normally occurs before double support. In populations
of subjects with stiff knee gait due to stroke (n = 10, Robertson et al., 2009) and TBI
(n = 20, Kerrigan et al., 2001b) preswing hip moments were within normal limits.
If insufficient hip flexion moment is a contributor to stiff-knee gait, possible
interventions may include hip flexor strengthening exercises, biofeedback focused on
hip flexion, and strategies to electrically stimulate the hip flexors (Kerrigan and Riley,
1998). Kerrigan and Riley (1998) reported that in a single stiff-knee stroke subject
who underwent daily hip flexor strengthening, knee flexion during swing improved

36
nine degrees after two months; however, the authors did not provide quantitative
evidence that hip flexor strength increased.
2.2.5 HAMSTRINGS OVER-ACTIVITY
Two other mechanisms have been proposed by which hamstrings over-activity
may contribute to stiff-knee gait including 1) hamstrings over-activity could generate
an excessive hip extension moment and 2) hamstrings over-activity could limit knee
velocity during swing.
Kerrigan et al. (1999) postulated that inappropriate hamstrings activity during
preswing or early swing could contribute to stiff-knee gait in stroke subjects by
generating an excessive hip extension moment. Musculoskeletal simulation suggests
that the effect of hamstrings activity on knee motion may vary throughout the gait
cycle. Simulation studies have shown that the hamstrings contribute to knee extension
during single limb support (Arnold et al., 2005; Hicks et al., 2008) and early swing
(Arnold et al., 2007b) in unimpaired gait and crouch gait (Hicks et al., 2008) and,
therefore, may limit knee extension in swing during these periods. In contrast, during
double support, hamstring activity contributes to knee flexion velocity at toe-off in
normal gait (Goldberg et al., 2004). More work is required to investigate the
contributions of hamstrings to knee flexion in impaired gait. It should be recognized
that interpretation of bi-articular muscle activity in simulations is dependent on the
modeled ratio of moment arms of the muscle at each joint and the foot floor contact
model used during the analysis.

37
It has also been proposed that hamstrings spasticity may limit knee flexion in
swing indirectly. Tuzson et al. (2003) showed that maximum knee flexion velocity
(occurring in preswing) was correlated with maximum knee extension velocity
(occurring in terminal swing) in 18 children with cerebral palsy (r = .94, p < .001). It
has been suggested that stable walking requires this symmetry (Mcgeer, 1990). If so, it
is possible that a limitation of knee extension velocity, due to hamstrings over-activity
in terminal swing, may cause a patient to walk with reduced knee flexion velocity at
toe-off to maintain symmetry, resulting in a stiff-knee gait. Tuzson et al. (2003)
measured the minimum knee angular velocities at which a spastic response was
elicited or spastic threshold velocity, in the quadriceps and hamstrings muscle groups
of subjects with cerebral palsy. In subjects with spastic responses in both the
quadriceps and hamstrings muscle groups, the lower spastic threshold velocity of the
two muscle groups was correlated with knee angular velocity during walking (Cheung
et al., 2003; Tuzson et al., 2003). These results suggest that stiff-knee subjects may
reduce their knee flexion velocity at toe-off to match the spastic threshold of their
hamstrings in terminal swing.
Whether either mechanism enables the hamstrings to contribute to stiff-knee
gait, there has been a lack of clinical evidence of hamstrings involvement in stiff-knee
gait. Kerrigan et al. (1991) compared kinematics between two groups of stroke
subjects (one with preswing hamstring activity and one without) with stiff-knee gait,
most of whom had inappropriate rectus femoris activity, and did not find a difference
in preswing or peak knee flexion between the groups. The only clinical evidence
supporting hamstrings as a cause of stiff-knee gait is in the stroke population. Kerrigan

38
and Riley (1998) performed a temporary intramuscular neurolysis of the hamstrings in
one stiff-knee stroke subject with inappropriate hamstrings activity which resulted in a
five degree increase in peak knee flexion in swing.
Although there is a lack of understanding of how hamstrings may contribute to
stiff-knee gait, it has been repeatedly reported clinically that hamstrings lengthening
can induce stiff-knee gait. There have been many reports of stiff-knee induced by
hamstrings lengthening without rectus femoris surgery (Gage et al., 1987; Thometz et
al., 1989; Damron et al., 1993; Nicholson, 2000). Van der Linden et al. (2003)
reported peak knee flexion was decreased on average in 32 limbs of 18 subjects with
hamstrings lengthening without rectus femoris surgery, but when walking speed was
accounted for, the postoperative peak knee flexion was at least 100% of normal peak
knee flexion for slower speeds, suggesting the importance of accounting for walking
speed when reporting outcomes. Hsu et al. (1990) reported 10 out of 49 hamstrings
lengthening patients developed stiff-knee gait postoperatively, but also reported that
all cases of stiff-knee gait resolved with physical therapy. Goldberg et al. (2004)
proposed that hamstrings lengthening could lead to stiff-knee gait by reducing the
force output of the hamstrings which they showed contributed to knee flexion velocity
at toe off.
Though there are no commonly employed treatments for hamstrings as a cause
of stiff-knee gait in cerebral palsy, Kerrigan et al. (1999) suggested if reduced hip
flexion and power generation are observed in concert with inappropriate hamstrings
activity, a temporary motor point block may be used in the hamstrings to determine
their involvement in stiff-knee gait. If the motor point block improves swing phase

39
knee flexion, they recommend stretching exercises for the hamstrings. They
recommend against permanent block to the hamstrings due to concern for stance phase
stability.
2.2.6 CROUCH GAIT
Although the precise nature of the interaction between crouch gait and stiff-
knee gait is not well-defined, there is much evidence that the two conditions may be
related. Many authors have observed quadriceps and hamstrings spasticity frequently
occur together (Csongradi et al., 1979; Gage et al., 1984; Gage, 1990). Outcomes
following treatment for crouch and stiff-knee gait also appear to be related. Csongradi
et al. (1979) observed both the development of crouch following rectus femoris release
and the development of hyperextended knee gait following hams transfer or
lengthening. Many studies confirm the finding of stiff-knee gait resulting from
hamstrings lengthening without concomitant rectus femoris transfer (Gage et al., 1987;
Thometz et al., 1989; Hsu and Li, 1990; Damron et al., 1993; Nicholson, 2000).
Studies of hamstrings lengthening with rectus femoris transfer report improved knee
range of motion and knee extension in stance (Hadley et al., 1992; Nene et al., 1993;
Rethlefsen et al., 1999; Yngve et al., 2002; Carney and Oeffinger, 2003; Park et al.,
2009).
It has been suggested that crouch gait may play a role in contributing to stiff
knee gait. Three mechanisms have been proposed by which crouch gait may contribute
to stiff-knee gait. The first mechanism proposes hamstrings spasticity, a commonly

40
cited cause of crouch gait, may contribute directly to stiff knee gait. The evidence for
this mechanism has been presented previously in this article.
The second mechanism proposes that excessive quadriceps activity in
preswing, either of the vasti or rectus femoris, necessary to achieve the high knee
extension moments required to hold the body upright in a crouched posture, may limit
the knee flexion acceleration necessary in preswing to achieve normal knee flexion in
swing. Lin et al. (2000) observed an increase in both preswing knee extension moment
and rectus femoris activity in subjects with cerebral palsy walking in a crouch gait,
defined as increased knee flexion throughout stance phase, as compared to subjects
with cerebral palsy not walking with a crouch gait pattern. Goldberg et al. (2006) also
noted excessive knee extension moments in double support in 17 of 23 subjects with
stiff knee gait. They reported that some subjects walked with excessive stance phase
knee flexion, though they did not report the incidence of preoperative crouch gait.
They also noted that decreases in the average knee extension moment in double
support were associated with improvements in stiff-knee gait.
One argument against this proposed mechanism is that unimpaired subjects
walking in a voluntary crouch gait (van der Krogt et al., 2007) or a crouch gait
induced by exoskeleton (Matjacic and Olensek, 2007) do not exhibit reduced or
delayed peak knee flexion during swing. One counter argument to this is that it is not
known whether unimpaired individuals may be able to compensate in ways that
individuals with cerebral palsy may not, therefore achieving adequate swing phase
knee flexion despite decreased knee flexion acceleration in preswing. Another counter
argument is that muscle relaxation rates in individuals with cerebral palsy may be

41
delayed compared to typically developing individuals, which may prolong the
excessive knee extension moment and interfere with the transition to knee flexion
acceleration. Tammik et al. (2008) reported significantly longer relaxation times in
quadriceps of children with spastic diplegia compared to age-matched controls.
Downing et al. (2009) also found a decreased rate of net knee extension moment
relaxation in subjects with cerebral palsy compared to able-bodied controls.
Clinical evidence of the relationship between excessive knee extension
moments in crouch and stiff knee gait is inconclusive. Both Gage et al. (1987) and
Goldberg et al. (2006) observed residual stance phase knee flexion in subjects with
poor outcomes following rectus femoris transfer, but it is not clear from either study
whether the residual crouch posture was causing the stiff-knee gait or rather was
coexisting with the stiff-knee gait.
The final mechanism proposes that the passive dynamics of crouch gait can
result in diminished knee flexion in swing. This mechanism was proposed by van der
Krogt et al. (2010) who created a simulation of cyclic walking using a two-
dimensional simple model with a passive swing knee. They initially set both knees at a
certain degree of knee flexion, locked the stance knee, and applied an external “push-
off” impulse to the swing limb. They also employed methods to prevent swing limb
hyperextension and represent foot contact. Some limitations of this approach included
non physiological kinematics such as a rigid ankle, no knee flexion velocity at toe-off,
and zero degrees of knee flexion throughout stance in the normal simulation. They
noticed that with increasing degrees of initial knee flexion angle, or crouch severity,
the simulated peak knee flexion in swing was increasingly diminished. They explained

42
that this was due to a decrease in the contribution of gravitational force to knee flexion
during swing with more crouched postures.
Earlier simulation studies investigated gravitational contribution to swing
phase knee flexion (Anderson et al., 2004) and knee flexion acceleration (Arnold et
al., 2007b) in normal gait using a three-dimensional model. These studies found that
gravity did not contribute substantially to peak knee flexion during swing because it
accelerated all of the segments of the swing limb, including the swing-limb side of the
pelvis, downward nearly uniformly. They recognized that using a model in which the
trajectory of the hip was prescribed, would have resulted in a substantial contribution
from gravity knee extension. The model created by van der Krogt et al. (2010) could
be considered a fixed hip flexion trajectory, since the stance leg was locked and two-
dimensional motion did not allow for pelvic list, however their results describe gravity
inducing knee flexion during swing rather than knee extension as proposed by
Anderson et al. (2004) and Arnold et al. (2007b). It is hard to generalize the results of
the simple model study to human walking in the presence of many non-physiologic
assumptions and simplifications.
2.2.7 OTHER POTENTIAL CAUSES
It is possible that other biomechanical abnormalities may contribute to stiff-
knee gait. The effects of patella alta on the moment arm of the quadriceps has been
studied (Ward et al., 2005; Sheehan et al., 2008; Luyckx et al., 2009) but it remains
unclear whether patella alta affects the moment arm of the knee extensors and could
contribute to stiff-knee gait. Others have suggested that femoral anteversion could lead

43
to increased rectus femoris stretch and cause stiff-knee gait (Piccinini et al., 2009;
Cimolin et al., 2010), but the effect has not been adequately investigated.
2.3 METHODOLOGY: USING SIMULATION TO UNDERSTAND MUSCLE
FUNCTION
Modeling and simulation are valuable tools for understanding muscle function
in gait. Dynamic simulation allows us to quantify the contribution of muscle forces to
joint motion, which can be unintuitive since muscles contribute to motion of joints that
they do not span and bi-articular muscles can accelerate joints in a direction opposite
of that assumed by anatomy (Zajac and Gordon, 1989). Simulation is also a valuable
tool in representing the extremely complex interactions that are necessary to generate
motion. Neural excitations from the brain or spinal cord activate muscles by causing a
diffusion of calcium ions in muscle fiber cells. Activated muscle fibers then generate
force which is dependent upon the level of activation, the length of muscle fibers, the
velocity at which the muscle fibers contract or lengthen, and other factors. Fiber forces
are transmitted across tendons to the bones. Muscle forces are applied across joints
creating joint moments. Joint moments affect the motion of all the joints in the body,
resulting in motion. In addition, reaction forces occur when the body contacts objects
in the environment. Simulation allows representation and investigation of these
complex interactions.

44
2.3.1 SIMULATION OF GAIT
In this dissertation, we used a model with 12 rigid segments, 21 degrees of
freedom and 92 muscle-tendon actuators to represent the human musculoskeletal
system (Delp et al., 1990). The position and orientation of the pelvis segment with
respect to ground were defined by six degrees of freedom. The head, arms and torso
were represented by a single rigid segment articulating with the pelvis by a ball-and-
socket joint. Each hip was modeled as a ball-and-socket joint, each knee as a planar
joint with tibiofemoral and patellofemoral kinematics defined by knee flexion angle
(Delp et al., 1990), and each ankle and subtalar joint as revolute joints (Inman, 1976).
Muscle paths were represented by line segments, with multiple lines of action for
muscles with broad attachments. Wrapping surfaces and via points represented
interaction of the muscle path with bone surfaces and soft tissue. Muscle force-
generating properties were represented with a lumped parameter model that accounted
for muscle force as a function of the muscle fiber length, muscle fiber velocity, tendon
length, and activation.
Simulations were generated using a four-step process. First, the
musculoskeletal model is scaled to match the subject’s anthropometry. This includes
scaling the segment lengths, masses, and inertias. Muscle and tendon lengths are also
scaled such that the ratio of muscle length to tendon length is maintained. Next,
inverse kinematics is applied to determine the set of joint angle trajectories over time
that minimize the least-squares distance between model markers and experimental
markers over the duration of the motion. Forces generated by the feet contacting the
ground, known as ground reaction forces, are measured during experimental data

45
collection. Due to inaccuracies in the model and errors in experimental measurement
of body motion and ground reaction forces, the model’s motion is not dynamically
consistent with the measured ground reaction force, i.e., maF ≠ . To make the
simulation dynamically consistent, a residual elimination algorithm is implemented,
which solves for the joint moments required to track the model’s motion while
measured ground reaction forces are applied, allowing small deviations in kinematics
to eliminate dynamic inconsistencies (Thelen and Anderson, 2006). An alteration on
this algorithm is residual reduction algorithm, which reduces rather than eliminates the
non-physical residual forces and moments resulting from dynamic inconsistency (Delp
et al., 2007). This modified algorithm, which was used in Chapter 5, prevents
excessive flexion and rotation of the torso segment that can be induced by the residual
elimination algorithm. The next step is to determine the individual muscle excitations
that generate the net joint moments that drive the model’s motion. This is
accomplished using the computed muscle control algorithm (Thelen et al., 2003). This
algorithm applies a feedback controller and an optimizer to choose the set of muscle
excitations that minimizes the sum of squared muscle activations that sufficiently
tracks the motion. The end result is a set of muscle excitation trajectories over time
that accurately reproduces the measured motion when applied to the musculoskeletal
model in concert with the ground reaction forces. The simulated muscle excitations are
compared to experimentally collected muscle electromyographic data to determine
how well the simulated muscle activity represents the measured muscle activity. If
inconsistencies are observed, simulated muscle excitations are constrained and

46
computed muscle control is reapplied. This step is repeated until the resulting
kinematics, kinetics, and muscle activity satisfactorily represent the experimental data.
2.3.2 ANALYSIS OF SIMULATIONS
In all three of the studies presented in this dissertation, forward dynamic
simulation was used to create altered simulations after making changes to muscles in
the unaltered simulation in order to analyze muscle function. This was done in
different contexts, with different simulations, to answer specific clinical questions. In
all cases, altering muscle force in a simulation requires a foot-floor contact model to
account for the resulting change in the ground reaction force. In all of these studies,
linear and rotational spring-damper elements were used to model the change in the
ground reaction force. The spring-damper elements were attached to the center of
pressure of the foot, or the point at which the resultant of the ground reaction force is
applied. As the change in the muscle force alters the path of the foot in the altered
simulation, the spring-damper elements apply a force and torque, proportional to
deviations in foot position and velocity, to the foot in the perturbed simulation that
pull it toward the position of the foot in the unaltered simulation. A scaling function is
used to apply these spring-damper forces smoothly as the foot transitions in and out of
contact with the ground and as the foot rotates in and out of foot-flat position.
The studies outlined in chapters 3 and 4 perform large changes to muscles in the
altered simulation and compare the resulting kinematics to the unaltered simulation to
determine the effect on knee motion. In contrast, the study presented in chapter 5
employs very small perturbations to muscle force in order to more precisely quantify

47
contributions of muscles to knee motion at specific time points throughout the gait
cycle. This technique is known as perturbation analysis (Liu et al., 2006b). Although
we used this method, other analysis methods have been employed to investigate
muscle function in gait (Anderson and Pandy, 2003; Hamner et al., 2010; Lin et al.,
2011). Using perturbation analysis, the contribution of the force of a muscle m to the
acceleration of a joint at time it , )( im tx&& , is determined by calculating the sensitivity,
m
m
Fx
∂∂ && , of the joint acceleration to force generated by muscle m and then scaling it by
the force produced by the muscle in the unaltered simulation, mF :
mm
im FFx
tx∂
∂=
&&&& )(
The sensitivity of the joint acceleration to force generated by the muscle of interest is
defined by:
m
imimm
m FtFxtFFx
Fx
Δ−Δ+
=∂
∂ ),(),( &&&&&&
where it is the current time in the unaltered simulation, x&& is the joint acceleration, mF
is the force generated by muscle m, and mFΔ is the perturbation applied to the muscle
force. This forward difference method is an exact expression for the sensitivity since
acceleration and force are linearly related. The component of the ground reaction force
generated in response to the muscle’s perturbation is accounted for by the foot-floor
contact model described previously. This sensitivity is calculated by incrementing the
force of a single muscle m in the simulation by a constant force perturbation, chosen to

48
be mFΔ = 1.0 N, integrating the simulation forward in time over a short interval, and
quantifying the resulting change in knee motion.
The change in joint angular acceleration induced by the muscle perturbation is
estimated assuming a constant acceleration over the short interval, picked to be 20ms,
of the forward integration. The acceleration is estimated using equations of constant
acceleration that are based on changes in position. This process was repeated for each
muscle in the model. A similar approach was utilized to quantify the contribution of
gravitational acceleration to the knee angular acceleration.
A unique contribution of this dissertation was the development of an algorithm to
quantify the contributions of Coriolis and centrifugal accelerations to knee angular
acceleration. This procedure involved setting all forces in the simulation to zero, such
that the motion resulting after forward integration was due only to the initial velocities
of the coordinates in the model and the resulting Coriolis and centrifugal accelerations.
The contribution, )( iv tx&& , of Coriolis and centrifugal accelerations to knee angular
acceleration at time ti was then defined by:
[ ]m
imimimmiv F
ttFxtFxttFFxtx
ΔΔ−−Δ+Δ+
=),(),(),(2
)(&
&&
With the development of the algorithm to calculate contributions of Coriolis and
centrifugal accelerations to joint acceleration, all contributions in the system to joint
acceleration can be quantified. Summing all contributions to joint acceleration in the
system and comparing to the actual acceleration of the joint of interest allows one
check of the appropriateness of the algorithm implementation and the foot-floor
contact model.

49
3 IMPORTANCE OF PRESWING RECTUS FEMORIS
ACTIVITY IN STIFF-KNEE GAIT 3.1 ABSTRACT
Stiff-knee gait is characterized by diminished and delayed knee flexion during
swing. Rectus femoris transfer surgery, a common treatment for stiff-knee gait, is
often recommended when a patient exhibits prolonged activity of the rectus femoris
muscle during swing. Treatment outcomes are inconsistent, in part, due to limited
understanding of the biomechanical factors contributing to stiff-knee gait. This study
used a combination of gait analysis and dynamic simulation to examine how activity
of the rectus femoris during swing, and prior to swing, contribute to knee flexion. A
group of muscle-actuated dynamic simulations was created that accurately reproduced
the gait dynamics of ten subjects with stiff-knee gait. These simulations were used to
examine the effects of rectus femoris activity on knee motion by eliminating rectus
femoris activity during preswing and separately during early swing. The increase in
peak knee flexion by eliminating rectus femoris activity during preswing (7.5 ± 3.1°)
was significantly greater on average (paired t-test, p = 0.035) than during early swing
(4.7 ± 3.6°). These results suggest that preswing rectus femoris activity is at least as
influential as early swing activity in limiting the knee flexion of persons with stiff-
knee gait. In evaluating rectus femoris activity for treatment of stiff-knee gait,
preswing as well as early swing activity should be examined.

50
3.2 INTRODUCTION
Stiff-knee gait is a debilitating consequence of cerebral palsy characterized by
diminished knee motion and delayed peak knee flexion during swing. Each year, three
out of every 1000 children manifest one or more of the symptoms of cerebral
palsy(CDC, 2004). Approximately 48–79% of all individuals with cerebral palsy are
ambulatory (Stanley, 2000). Stiff-knee gait is one of the most common gait
abnormalities in ambulatory children with spastic cerebral palsy (Wren et al., 2005b).
Many individuals with stiff-knee gait frequently trip or perform inefficient
compensatory movements due to inadequate toe clearance (Sutherland and Davids,
1993).
Distal transfer of the rectus femoris is a common surgical treatment for stiff-
knee gait (Gage et al., 1987; Perry, 1987). Though increased vasti and decreased
iliopsoas activity have been identified as potential causes of stiff-knee gait (Goldberg
et al., 2004), the limited knee flexion is usually attributed to abnormal prolongation of
rectus femoris activity into early swing phase (Sutherland et al., 1975; Waters et al.,
1979; Gage et al., 1987; Perry, 1987; Sutherland et al., 1990; Sutherland and Davids,
1993). Rectus femoris transfer surgery is intended to decrease the muscle’s ability to
extend the knee while preserving its ability to generate hip flexion moment (Gage et
al., 1987; Perry, 1987; Asakawa et al., 2002), which promotes knee flexion (Piazza
and Delp, 1996; Kerrigan and Riley, 1998; Asakawa et al., 2002). Several studies have
reported that rectus femoris transfer typically improves knee flexion (Gage et al.,
1987; Sutherland et al., 1990; Ounpuu et al., 1993a; Ounpuu et al., 1993b; Rethlefsen

51
et al., 1999). However, less positive outcomes related to swing phase peak knee
flexion have more recently been reported in some patients (Yngve et al., 2002).
Outcomes of surgical treatments for stiff-knee gait are inconsistent, in part, due to
insufficient understanding of the biomechanical factors contributing to stiff-knee gait.
Although rectus femoris transfer is thought to improve knee flexion by decreasing
knee extension moment, Goldberg et al. (2003) found that many subjects with stiff-
knee gait did not walk with abnormally large knee extension moments during early
swing, but they walked with abnormally low knee flexion velocity at toe-off. Goldberg
et al. (2006) subsequently reported that many subjects with stiff-knee gait walked with
abnormally large knee extension moments during double support, which were
correlated with low knee flexion velocity at toe-off. Moreover, most subjects with
favorable outcomes following surgery walked with decreased knee extension moments
during double support and corresponding increased knee flexion velocities at toe-off
(Goldberg et al., 2006). These results suggest that knee extension moment, which is
influenced by rectus femoris activity, prior to toe-off, rather than after toe-off, may be
a more prevalent contributor to stiff-knee gait than previously thought. A better
understanding of when rectus femoris activity contributes to stiff-knee gait is
necessary to refine clinical indications for rectus femoris transfer surgery.
This study used dynamic simulation, in combination with gait analysis, to
evaluate the relative importance of preswing (i.e., the period immediately prior to toe-
off) rectus femoris activity as a biomechanical factor contributing to diminished knee
flexion in subjects with stiff-knee gait. We hypothesized that rectus femoris activity
during preswing has a greater impact on peak knee flexion than rectus femoris activity

52
during early swing (i.e., the period from toe-off to peak knee flexion) in subjects with
stiff-knee gait. We tested this hypothesis by simulating the elimination of rectus
femoris activity during preswing and separately during early swing for a group of ten
subjects with cerebral palsy walking with stiff-knee gait and computing the resulting
changes in knee flexion. Identifying the function of rectus femoris activity during
preswing and early swing in subjects with stiff-knee gait contributes to our
understanding of this gait abnormality and provides insights needed to improve
treatment planning.
3.3 METHODS
The subjects in this study underwent gait analysis at Connecticut Children’s
Medical Center in Hartford, CT. Gait analysis data, including three-dimensional joint
angles, ground reaction forces and moments, and surface electromyographic (EMG)
recordings from preamplifier electrodes, were collected as a routine part of treatment
planning. Our inclusion criteria (Goldberg et al., 2006) required that each subject (i)
subsequently underwent rectus femoris transfer surgery as a correctional treatment for
stiff-knee gait, (ii) was between 6 and 17 years of age prior to surgery, (iii) had not
undergone a selective dorsal rhizotomy, and (iv) walked without orthoses or other
assistance. Ten subjects were identified and categorized as exhibiting stiff-knee gait in
at least one limb preoperatively (Table 3.1).

53
Table 3.1 Descriptive values for 10 subjects with stiff-knee gait and 15 able-bodied subjects

54
Four gait parameters (Goldberg et al., 2006) were used to determine whether a
subject walked with stiff-knee gait: peak knee flexion in swing phase (Gage et al.,
1987; Sutherland et al., 1990), knee range of motion in early swing (Goldberg et al.,
2003), total knee range of motion (Gage et al., 1987; Ounpuu et al., 1993b; Ounpuu et
al., 1993a), and timing of peak knee flexion during swing phase (Sutherland et al.,
1990; Ounpuu et al., 1993b; Ounpuu et al., 1993a). A limb was classified as ‘‘stiff’’ if
three or more of these measures were more than two standard deviations below (or
above in the case of the timing measure) the average control value. Control data were
collected from 15 able-bodied subjects of approximately the same average age, height,
and weight as the subjects with cerebral palsy (Table 3.1). Surface EMG data were not
used to include or exclude subjects from this study. However, all of the subjects with
stiff-knee gait did exhibit abnormal rectus femoris activity (Table 3.2). All subjects
gave informed consent for the collection of their gait data. Mutual institutional
approval was obtained for retrospective analysis of these data. The data analysis
included the creation of subject-specific dynamic simulations.

55
Table 3. 2 Description of abnormal rectus femoris activity in subjects
A three-dimensional, full-body musculoskeletal model with 21 degrees of
freedom and 92 muscle–tendon actuators formed the foundation of each simulation
(Fig. 3.1). The position and orientation of the pelvis relative to ground was defined
with 6 degrees of freedom. The head, arms, and torso were represented as a rigid
segment connected with the pelvis by a ball-and-socket joint (Anderson and Pandy,
1999). The remaining lower extremity joints were modeled as follows: each hip as a
ball-and-socket joint, each knee as a planar joint with tibiofemoral and patellofemoral
translational constraints as a function of knee flexion (Delp et al., 1990), and each
ankle and subtalar joints as revolute joints (Inman, 1976). Each muscle–tendon
actuator was modeled as a Hill-type muscle in series with tendon based on
musculotendon parameters from Delp et al. (1990). The musculoskeletal model and
corresponding dynamic simulation code were produced using SIMM and the
Dynamics Pipeline (Delp and Loan, 2000) along with SD/FAST (Parametric

56
Technology Corporation, Waltham, MA). The musculoskeletal model was used in
conjunction with gait analysis data to create subject-specific dynamic simulations.
Figure 3.1 Muscle-actuated dynamic simulation of a subject’s gait during the period of preswing through early swing. A three-dimensional, full-body musculoskeletal model with 21 degrees of freedom and 92 muscle-tendon actuators was used in conjunction with the subject’s gait analysis data to create each subject-specific simulation. The dynamic simulation is shown at the initiation of preswing (left), just following toe-off (center), and at the termination of early swing (right). Each subject-specific simulation was used to conduct simulation experiments to evaluate the relative importance of preswing rectus femoris activity as a biomechanical factor contributing to the subject’s diminished knee flexion.
A muscle-actuated dynamic simulation of each subject was created using a
four-step process. First, the musculoskeletal model was scaled to represent the

57
experimentally measured size of each subject. Second, inverse kinematics analysis
was utilized to obtain values of generalized coordinates for the model that closely
matched the experimentally measured kinematics of each subject. Third, a residual
elimination algorithm (Thelen and Anderson, 2006) was applied to achieve dynamic
consistency between the model’s motions and the experimentally measured ground
reactions of each subject, by adjusting pelvis translations and back rotations. Fourth,
computed muscle control (Thelen et al., 2003) was implemented to determine an
optimal set of muscle activities that produced forward simulations and that were
generally consistent with the experimentally measured kinematics and EMG patterns
of each subject. Constraints were placed on the muscle activity of each simulation
based on the recorded EMG. For example, when activity was recorded for rectus
femoris during early swing, the simulated rectus femoris was required to have activity
during this time as well. This four-step process was used to create simulations for each
subject’s preoperative gait during the period of preswing through peak knee flexion in
swing. The simulated joint angles reproduced the subjects’ measured hip, knee, and
ankle angles within 31. The subject-specific dynamic simulations were used to
conduct subsequent simulation experiments.
The simulation of each subject was altered to examine the effects of rectus
femoris activity on knee motion. In particular, the activity of rectus femoris was
eliminated during preswing and separately during early swing, creating two new
simulations per subject, to determine the muscle’s relative importance to peak knee
flexion for each case (Fig. 3.2). By observing the changes in peak knee flexion
between the new and unperturbed simulations, the muscle’s contribution to knee
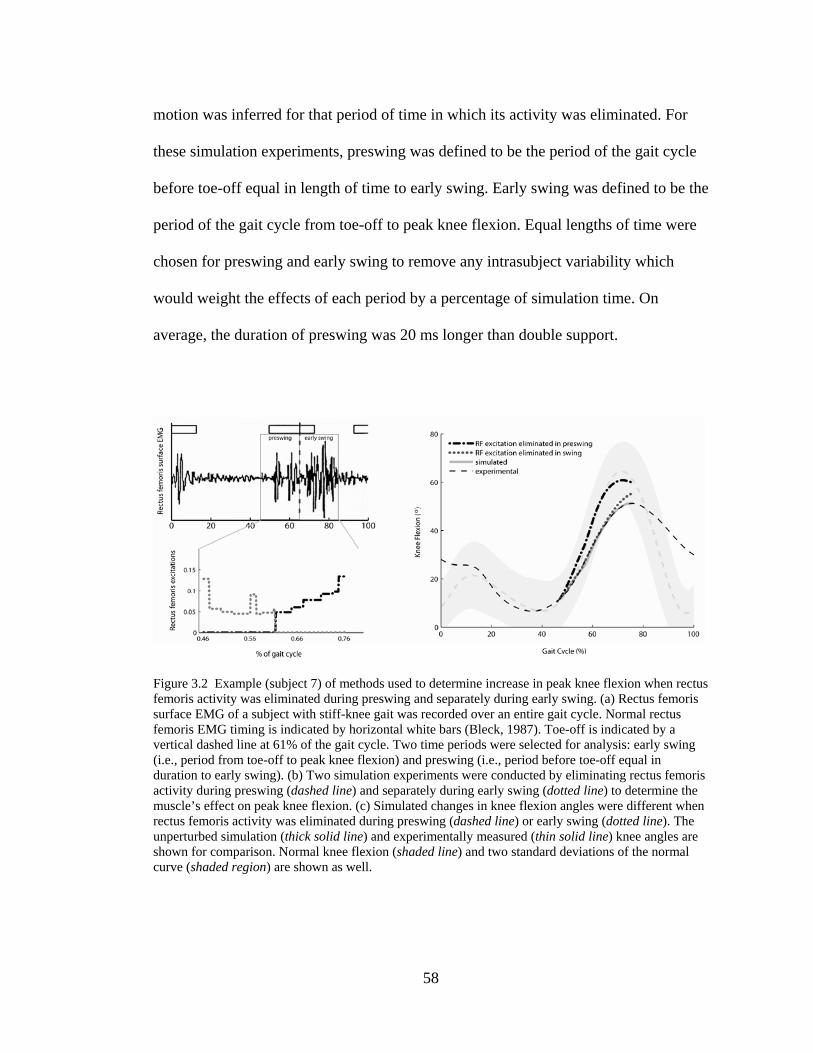
58
motion was inferred for that period of time in which its activity was eliminated. For
these simulation experiments, preswing was defined to be the period of the gait cycle
before toe-off equal in length of time to early swing. Early swing was defined to be the
period of the gait cycle from toe-off to peak knee flexion. Equal lengths of time were
chosen for preswing and early swing to remove any intrasubject variability which
would weight the effects of each period by a percentage of simulation time. On
average, the duration of preswing was 20 ms longer than double support.
Figure 3.2 Example (subject 7) of methods used to determine increase in peak knee flexion when rectus femoris activity was eliminated during preswing and separately during early swing. (a) Rectus femoris surface EMG of a subject with stiff-knee gait was recorded over an entire gait cycle. Normal rectus femoris EMG timing is indicated by horizontal white bars (Bleck, 1987). Toe-off is indicated by a vertical dashed line at 61% of the gait cycle. Two time periods were selected for analysis: early swing (i.e., period from toe-off to peak knee flexion) and preswing (i.e., period before toe-off equal in duration to early swing). (b) Two simulation experiments were conducted by eliminating rectus femoris activity during preswing (dashed line) and separately during early swing (dotted line) to determine the muscle’s effect on peak knee flexion. (c) Simulated changes in knee flexion angles were different when rectus femoris activity was eliminated during preswing (dashed line) or early swing (dotted line). The unperturbed simulation (thick solid line) and experimentally measured (thin solid line) knee angles are shown for comparison. Normal knee flexion (shaded line) and two standard deviations of the normal curve (shaded region) are shown as well.

59
We evaluated our hypothesis regarding the relative importance of preswing
and early swing rectus femoris activity by conducting a paired t-test at the 0.05
significance level. A one-tailed test was used due to a priori expectation about
directionality (i.e., rectus femoris activity during preswing has a greater impact on
peak knee flexion than rectus femoris activity during early swing in subjects with stiff-
knee gait). The null hypothesis was that the difference in peak knee flexion change
between the preswing and the early swing simulations was zero. The test was
performed against the right-tailed alternative hypothesis that peak knee flexion
increased more, on average, in the simulations when rectus femoris activity was
eliminated during preswing than during early swing.
3.4 RESULTS
Peak knee flexion increased more (p = 0.035), on average, when rectus femoris
activity was eliminated during preswing than during early swing in our simulations
(Fig. 3.3). Peak knee flexion increased 7.5±3.1° when activity was eliminated during
preswing and 4.7±3.6° when eliminated during early swing. Peak knee flexion
increased more for the preswing case than for the early swing case in the majority of
subject simulations. For six subjects (1, 4, 5, 7, 9, and 10), the increase in peak knee
flexion was 90% higher or more for the preswing case than for the early swing case.
For three subjects (2, 6, and 8), the increase in peak knee flexion was similar (within
10%) for the preswing case and the early swing case. For the remaining subject (3),
the increase in peak knee flexion was substantially lower (37%) for the preswing case
than for the early swing case.

60
Figure 3.3 Increase in peak knee flexion caused by eliminating rectus femoris activity during preswing and separately during early swing in simulations of ten subjects with stiff-knee gait (ordered by increasing unperturbed peak knee flexion). The increase was determined by eliminating rectus femoris activity in a forward dynamic simulation and computing the change in peak knee flexion compared with the unperturbed value (Fig. 3.2). The changes in simulated knee motion give insight into the biomechanical contribution of rectus femoris for that period of time in which its activity was eliminated. 3.5 DISCUSSION
Rectus femoris transfer surgery is often performed to treat stiff-knee gait when
a patient exhibits prolonged activity of the rectus femoris into early swing. However,
Goldberg et al. (2006) suggested that rectus femoris activity prior to toe-off may also
contribute to stiff-knee gait by causing abnormally large knee extension moments and
corresponding low knee flexion velocity at toe-off. Our results confirm that preswing
rectus femoris activity is at least as important as early swing activity and, for some
subjects with stiff-knee gait, may limit knee flexion more than activity in early swing.
The subject population in this study was diverse, and findings may not generalize to
all subjects with stiff-knee gait. In evaluating rectus femoris activity for treatment of
stiff-knee gait, preswing and early swing EMG should be examined. It should be

61
noted, however, that surface EMG may be subject to cross-talk from the vasti (Barr et
al., 2010).
There are several possible biomechanical explanations why preswing rectus
femoris activity may limit knee flexion more than early swing activity. First, impaired
motor control may cause varying levels of preswing and early swing rectus femoris
activity (Fig. 3.4). In fact, preswing and early swing EMG activity of the rectus
femoris varies considerably in children with stiff-knee gait (Miller et al., 1997).
Excessive preswing activity (Fig. 3.4a) may result in above normal muscle force that
limits knee flexion. Second, the delay between muscle excitation and muscle force
generation suggests that preswing rectus femoris activity may cause forces that persist
into early swing phase and limit knee flexion. This delay in electromechanical
coupling has been reported to be between 30 and 100 ms (Cavanagh and Komi, 1979),
which is roughly 25–75% of the duration of preswing or early swing for the subjects in
this study. Third, musculoskeletal geometry and multibody dynamics may cause
varying magnitudes of preswing and early swing muscle forces that produce joint
motion. The transmission of muscle force to joint motion depends on the muscle’s
moment arm, which varies during movement. In fact, rectus femoris has large
potential during double support to decrease peak knee flexion velocity (Goldberg et
al., 2004). Preswing activity may result in a potentially large knee extension moment
that limits knee flexion.

62
Figure 3.4 Two subjects with varying levels of preswing and early swing rectus femoris activity. (a) Subject 8 had more rectus femoris activity in preswing compared to early swing. (b) Subject 3 had less rectus femoris activity in preswing compared to early swing.
There are several possible reasons why simulated increases in peak knee
flexion varied across subjects. First, EMG patterns and simulated muscle activity
varied across subjects. Constraints were placed on the muscle activity of each
simulation based on the recorded EMG. As a result, elimination of high rectus femoris
activity led to large simulated increases in peak knee flexion. For example, subject 3
had more activity in early swing compared to preswing (Fig. 3.4b); consequently, peak
knee flexion increased more when rectus femoris activity was eliminated during early
swing than during preswing (Fig. 3.3). The presence of high vasti activity in lieu of
rectus femoris activity may have attenuated the simulated increases in peak knee
flexion. Second, joint motion and body mass properties varied across subjects and
these can affect the change in knee motion caused by rectus femoris activity. For
example, improper positioning of the foot before toeoff may dramatically decrease the
ankle power required for proper knee flexion. Third, the duration of the simulation
times varied across subjects and long simulations may have produced large changes in
knee flexion. Given two simulations differing only in length of time, a longer

63
simulation eliminating rectus femoris activity increases peak knee flexion more than a
shorter simulation because the inhibitory knee extension moment, in part due to rectus
femoris muscle force, is reduced for a longer period of time.
By carefully defining the duration of preswing to equal the duration of early
swing, the intra-subject results were not contaminated by the effects of a long
perturbation being compared with those of a short perturbation. The period of double
support is defined by vertical ground reaction force measurements. Knee flexion
motion along with ground reactions defines early swing. There was significant
variation between double support and early swing durations for each subject (Table
3.1). For example, double support was roughly 54 ms shorter than early swing for
subject 2. If we had not controlled the duration over which we perturbed RF activity in
the simulations, then this difference would have allowed the early swing perturbations
to affect the model motion for 60% more time than double support perturbations. For
this reason, preswing was defined to be the period before toe-off equal in length of
time to early swing.
The simulated elimination of rectus femoris activity was not intended to
represent the activity of able-bodied control subjects. Although rectus femoris activity
for an individual subject is repeatable, there are significant differences across subjects
(Arsenault et al., 1986). In some cases using surface electrodes, a bi-phasic pattern
(i.e., one main burst during swing-to-stance transition and the second main burst
during stance-to-swing transition) can be observed (Murray et al., 1984; Shiavi, 1985;
Arsenault et al., 1986). In other cases using fine wire electrodes, no muscle activity or
a brief, weak burst is observed during stance-to-swing transition (Perry, 1987). In a

64
study with both types of electrodes, cross-talk from the underlying vasti contaminated
the EMG of rectus femoris recorded with surface electrodes (Nene et al., 2004).
Rather than simulating rectus femoris activity of able-bodied control subjects, the
simulations in this study allowed the muscle’s contribution (i.e., importance) to knee
flexion to be determined for that period of time in which its activity was eliminated.
The muscle-actuated simulations of stiff-knee gait developed in this study had
several limitations. First, the model used in this study was scaled to represent the size
and mass properties of each subject, but not individual impairments (e.g., skeletal
deformities, muscle contractures, and spasticity). Second, the simulations did not
explicitly model arm motions, which may have minimally affected the motions of
other body segments. Third, the model utilized muscle parameters representative of an
able-bodied adult, whereas the subjects in this study were children with neuromuscular
abnormalities. Fourth, the forces produced by muscles in our simulations may not
have accurately represented the forces generated by individual subjects even though
the net joint moments were representative of each subject. Although the increases in
peak knee flexion reported may change if we made different modeling assumptions,
our conclusions regarding the relative importance of preswing and early swing activity
would be unlikely to change significantly because the same assumptions would be
simulated across both time periods.
Our finding that preswing rectus femoris activity is an important
biomechanical factor contributing to diminished knee flexion in subjects with stiff-
knee gait is consistent with the findings of others. Several studies have shown that
swing-phase initial conditions are important in generating knee flexion during normal

65
gait with (Piazza and Delp, 1996) and without (Mochon and McMahon, 1980; Mena et
al., 1981) muscle activity. More recently, (Goldberg et al., 2003) demonstrated the
importance of swing-phase initial conditions, particularly knee flexion velocity at toe-
off, in stiff-knee gait. Our finding supports these studies because preswing muscle
forces generate initial conditions for swing phase (e.g., knee flexion velocity at toe-
off). In particular, excessive rectus femoris force during double support has the
potential to decrease knee flexion velocity at toe-off in normal gait (Goldberg et al.,
2004). Some studies have reported subjects with stiff-knee gait exhibit a below normal
knee flexion velocity at toe-off (Granata et al., 2000; Goldberg et al., 2003), and others
have simulated the proportional relationship between knee flexion velocity at toe-off
and swing-phase knee flexion for normal gait (Piazza and Delp, 1996) and stiff-knee
gait (Goldberg et al., 2003). Our finding of the cause–effect relationship between
preswing rectus femoris activity and swing-phase knee flexion is consistent with these
studies as well. The current work demonstrates the impact of rectus femoris activity on
swing-phase knee flexion and provides a direct comparison of preswing and early
swing importance for a number of subjects with stiff-knee gait.
Many subjects with stiff-knee gait walked with excessive knee flexion in
stance phase (e.g., crouch gait). This results in larger than normal knee extension
moments during double support. Large knee moments generated by the knee extensors
are necessary to support the body (McNee et al., 2004), but diminish knee flexion
velocity at toe-off and reduce peak knee flexion (Goldberg et al., 2006). Recent
analyses (Goldberg et al., 2006) suggest that the improvements in stiff-knee gait are
associated with sufficient decreases in excessive knee extension moments during

66
double support and corresponding increases in knee flexion velocity at toe-off. Further
analyses are necessary to determine if the correction of excessive knee flexion in
stance may diminish the excessive knee extension moments in double support. If so,
excessive rectus femoris excitation may no longer be necessary for body support.
Correcting excessive knee flexion in stance may increase knee flexion in swing.
The combination of gait analysis and dynamic simulation in this study identified the
importance of preswing rectus femoris activity in stiff-knee gait. This result indicates
that excessive preswing rectus femoris activity is a biomechanical factor contributing
to diminished knee flexion in subjects with stiff-knee gait. While gait analysis tools
alone are useful for characterizing stiff-knee gait, dynamic simulation provides an
additional, valuable tool for investigating its underlying biomechanical causes and the
mechanisms leading to improvement following treatment.

67
4 MECHANISMS OF IMPROVED KNEE FLEXION AFTER
RECTUS FEMORIS TRANSFER SURGERY 4.1 ABSTRACT
Rectus femoris transfer is frequently performed to treat stiff-knee gait in
subjects with cerebral palsy. In this surgery, the distal tendon is released from the
patella and re-attached to one of several sites, such as the sartorius or the iliotibial
band. Surgical outcomes vary, and the mechanisms by which the surgery improves
knee motion are unclear. The purpose of this study was to clarify the mechanism by
which the transferred muscle improves knee flexion by examining three types of
transfers. Muscle-actuated dynamic simulations were created of ten children
diagnosed with cerebral palsy and stiff-knee gait. These simulations were altered to
represent surgical transfers of the rectus femoris to the sartorius and the iliotibial band.
Rectus femoris transfers in which the muscle remained attached to the underlying vasti
through scar tissue were also simulated by reducing but not eliminating the muscle’s
knee extension moment. Simulated transfer to the sartorius, which converted the
rectus femoris’ knee extension moment to a flexion moment, produced 32° ± 8°
improvement in peak knee flexion on average. Simulated transfer to the iliotibial band,
which completely eliminated the muscle’s knee extension moment, predicted only
slightly less improvement in peak knee flexion (28° ± 8°). Scarred transfer
simulations, which reduced the muscle’s knee extension moment, predicted
significantly less (p < 0.001) improvement in peak knee flexion (14° ± 5°).
Simulations revealed that improved knee flexion following rectus femoris transfer is

68
achieved primarily by reduction of the muscle’s knee extension moment. Reduction
of scarring of the rectus femoris to underlying muscles has the potential to enhance
knee flexion.
4.2 INTRODUCTION
Stiff-knee gait, characterized by diminished and delayed peak knee flexion in
swing (Sutherland and Davids, 1993), is one of the most common gait problems in
children with cerebral palsy (Wren et al., 2005b). Insufficient knee flexion during
swing can lead to tripping and falling, and energy-inefficient compensatory
movements. Despite the prevalence of stiff-knee gait, its causes are not well
understood. Several factors may contribute to stiff-knee gait (Kerrigan and Glenn,
1994; Piazza and Delp, 1996; Riley and Kerrigan, 1998; Kerrigan et al., 1999;
Goldberg et al., 2003), but over-activity of the rectus femoris muscle is considered a
primary cause (Waters et al., 1979; Perry, 1987; Sutherland et al., 1990). Treatments
including rectus femoris transfer surgery and botulinum toxin injection aim to alter the
function of this muscle. A rectus femoris transfer relocates the insertion of the rectus
femoris from the patella to a more posterior site to augment knee flexion (Gage et al.,
1987; Perry, 1987).
Average improvement in knee flexion after rectus femoris transfer is positive
but variable. Most studies report an average increase in peak knee flexion after
transfer between 7-10° (Gage et al., 1987; Ounpuu et al., 1993b; Chambers et al.,
1998; Saw et al., 2003; Moreau et al., 2005). Some studies report increases in peak
knee flexion between 12-26° (Sutherland et al., 1990; Miller et al., 1997; Hemo et al.,

69
2007). Even studies that report no significant improvement (Hadley et al., 1992;
Ounpuu et al., 1993a; Rethlefsen et al., 1999) or an average decrease (Yngve et al.,
2002; Carney and Oeffinger, 2003) in peak knee flexion in swing find that patients
exhibit an average increase in knee range of motion, typically when rectus femoris
transfers are performed in conjunction with hamstrings lengthenings. There is limited
understanding of how the transferred rectus femoris affects knee motion and a lack of
consensus regarding the effects of transfer site. Several studies have reported no
difference among transfer sites on peak knee flexion improvement (Ounpuu et al.,
1993a; Muthusamy, 2006), whereas others have suggested that outcome is dependent
on transfer site (Chung et al., 1997; Hemo et al., 2007).
The mechanism by which rectus femoris transfer may increase knee flexion is
unclear. The transfer was originally intended to convert the muscle from a knee
extensor to a knee flexor (Perry, 1987), and a study on cadavers showed that the rectus
femoris has a knee flexion moment arm after transfer to the sartorius or
semitendinosus (Delp et al., 1994). However, examination of postoperative patients
revealed that the transferred muscle generates a knee extension moment upon
electrical stimulation (Riewald and Delp, 1997). In vivo dynamic imaging confirmed
that the muscle is not converted to a knee flexor but that the knee extension capacity
of the rectus femoris is diminished after surgery (Asakawa et al., 2002). In these same
subjects, magnetic resonance imaging also revealed the formation of scar tissue
between the rectus femoris and the vasti postoperatively. This connective tissue may
allow force to be transmitted from the rectus femoris to the vasti, resulting in the
rectus femoris producing a net knee extension moment despite transfer of its distal

70
tendon to the sartorius or semitendinosus (Asakawa et al., 2004). A proposed
mechanism of improvement in knee flexion after rectus femoris transfer is reduction
of the muscle’s knee extension moment with preservation of its hip flexion moment
(Delp et al., 1994; Riewald and Delp, 1997; Asakawa et al., 2002), which promotes
knee flexion through dynamic coupling (Piazza and Delp, 1996; Kerrigan and Riley,
1998). However, there is little evidence indicating whether this mechanism is likely to
increase knee flexion in patients with stiff-knee gait following rectus femoris surgery.
The purpose of this study was to investigate the mechanisms of improved knee flexion
after rectus femoris transfer by comparing the changes in knee flexion predicted by
simulating transfers to sartorius, transfers to iliotibial band, scarred rectus femoris
transfers, and botulinum toxin injection of the rectus femoris. This study evaluated the
relative importance of the transferred muscle’s hip and knee moments by analyzing
subject-specific simulations of children with stiff-knee gait.
4.3 METHODS
Muscle-actuated simulations were created to investigate the mechanism by
which rectus femoris transfer alters muscle function. We created simulations that
reproduced the gait dynamics of ten subjects with stiff-knee gait prior to treatment.
The rectus femoris was then altered to simulate the effects of different treatments.
Resulting changes in peak knee flexion were compared.

71
Subjects
All ten subjects in this study were diagnosed with spastic cerebral palsy and
classified as exhibiting stiff-knee gait (Fig. 4.1). To be classified as “stiff” the
subject’s knee motion was outside a normal range by more than two standard
deviations for at least three of four gait parameters: (1) peak knee flexion angle, (2)
range of knee flexion in early swing (from toe-off to peak knee flexion), (3) total
range of knee motion, and (4) timing of peak knee flexion in swing (Goldberg et al.,
2006). Control data was collected from 15 typical children of approximately the same
average age, height, and weight as the subjects with stiff-knee gait. Each stiff-knee
subject underwent a physical exam and gait analysis at Connecticut Children’s
Medical Center in Hartford, CT. All subjects met the following selection criteria: (i)
underwent rectus femoris transfer surgery to treat stiff-knee gait, (ii) were 6 - 17 years
of age at surgery, (iii) had not undergone a selective dorsal rhizotomy, and (iv) walked
without orthoses or other assistance. The group of subjects had an average age of 10.6
years. Rectus femoris was transferred distally to sartorius in all subjects.
Retrospective analysis of these data was performed with approval of participating
institutions.

72
Figure 4.1 Average experimental knee flexion (in more stiff limb) of ten subjects with stiff-knee gait (thick line) ± one standard deviation (dark shaded region), compared to average knee flexion of typical subjects (thin line) ± one standard deviation (light shaded region).
Creating subject-specific simulations
The musculoskeletal system was represented by a three-dimensional model
with 21 degrees of freedom and 92 muscle-tendon actuators. The position and
orientation of the pelvis segment with respect to ground was defined with six degrees
of freedom. A single rigid segment articulating with the pelvis by a ball and socket
joint represented the head, arms, and torso. Each hip was modeled as a ball and socket
joint, each knee as a planar joint with tibiofemoral and patellofemoral kinematics
defined by knee flexion angle (Delp et al., 1990), and each ankle and subtalar joint as
revolute joints (Inman, 1976). The musculoskeletal model and corresponding
dynamic simulation code were produced using SIMM (Delp and Loan, 2000) and
SD/FAST (Parametric Technology Corporation, Waltham, MA).
The musculoskeletal model was scaled to match the size and weight of each
subject. Next, we solved for the joint angles that minimized the distances between

73
virtual markers on the model and experimentally measured marker positions. Then, a
residual elimination algorithm (Thelen and Anderson, 2006) was applied to make
model kinematics consistent with measured ground reaction forces by adjusting pelvis
translations and back angles. Finally, computed muscle control (Thelen et al., 2003)
was utilized to determine a set of muscle excitations that produced a forward
simulation of the subject’s preoperative kinematics. The muscle excitations were
constrained to be consistent with measured EMG patterns. Sagittal knee, hip, and
ankle angles were within 3° of measured preoperative angles. Simulations represented
a portion of the gait cycle from preswing to peak knee flexion, centered around toe-
off.
Comparing effects of simulated treatments on knee flexion
Three types of rectus femoris transfers and a botulinum toxin injection into the
rectus femoris were simulated for each subject. The three different types of rectus
femoris transfer investigated included transfer to the sartorius, transfer to the iliotibial
band, and a transfer in which rectus femoris was scarred to the underlying vasti. The
preoperative musculoskeletal model for each subject (Fig. 4.2a) was altered to
represent each transfer scenario. The tendon slack length of the transferred muscle
was adjusted to keep muscle fibers operating near preoperative length ranges. The set
of muscle excitations from each subject’s preoperative simulation was applied to each
transfer model to simulate the effects of the surgery.
To simulate a transfer of the rectus femoris to the sartorius the rectus femoris
insertion was relocated to the effective insertion of the sartorius in the model (Fig.

74
4.2b). Even though the effective insertion of the sartorius is more proximal and
posterior than the anatomical insertion, the simulated rectus femoris transfer closely
approximates moment arms of transferred muscles measured experimentally by Delp
et al. ((1994); compare model to experiment in Fig. 4.2b). To simulate the negligible
knee extension moment arm (0-5 mm) measured by Delp et al. (1994) after transfer to
the iliotibial band, the muscle’s insertion was fixed on the femur, transforming it into a
uniarticular hip flexor. To simulate the effects of a transfer surgery in which the
rectus femoris becomes scarred to the underlying vasti we reduced the knee extension
moment arm of rectus femoris by half (Fig. 4.2d). Although the model for the scarred
transfer does not resemble a typical transfer surgery, it represents the net effect of a
scarred rectus femoris transfer on the muscle’s knee extension capacity based on the
subject who showed the least reduction in rectus femoris knee extension capacity in
the study of Asakawa et al. (2002). To simulate the effects of injection of botulinum
toxin into the rectus femoris, which decreases active muscle force by inhibiting the
release of acetylcholine from the neuromuscular junction (Burgen et al., 1949), we
applied the set of muscle excitations in the preoperative simulation to the preoperative
model but eliminated rectus femoris excitation.

75
Figure 4.2 Illustrations of the rectus femoris muscle (top panels) and its moment arm at the knee averaged over 20-60 degrees of knee flexion (bottom panels). The moment arms of the models (black bars) are compared with moment arms measured experimentally (grey bars) by Delp et al. (1994) in cadaver specimens in the (a) preoperative condition, after (b) transfer to the sartorius, and after (c) transfer to the iliotibial band. The muscle insertions shown in models (b) and (c) are effective insertions used to calculate moment arms (see Methods). The muscle’s knee extension moment arm in the (d) scarred transfer model is compared to its moment arm calculated from in vivo measurements of the muscle’s displacement in the patient from Asakawa et al. (2002) with the minimum reduction in knee extension moment arm after rectus femoris transfer.
After creating simulations of each treatment for each subject, the
improvements in peak knee flexion, measured at the point of preoperative peak knee
flexion, were quantified (Fig. 4.3). The average amounts of peak knee flexion
improvement for all subjects were compared among treatments, using a two-tailed,
paired t-test (p < 0.05) with Bonferroni correction for multiple comparisons.

76
Figure 4.3 Peak knee flexion resulting from simulation of (a) preoperative gait, (b) RF transfer to sartorius, (c) RF transfer to iliotibial band, (d) scarred RF transfer, and (e) botulinum toxin injection for a single subject.
Comparing influence of hip and knee moments of rectus femoris on knee flexion
To investigate the relative importance of the hip and knee moments of
transferred rectus femoris on peak knee flexion in swing, we analyzed four additional
simulations for each subject to represent conditions under which the hip and knee
moments of the muscle were separately eliminated or preserved. The simulations of
each subject’s preoperative gait served as the first condition that preserved both hip
and knee moments of the muscle. A second model was created for each subject in
which the muscle’s knee extension moment was eliminated, while its hip flexion
capacity after transfer was preserved. We preserved the transferred muscle’s hip
flexion capacity by maintaining its hip flexion moment arm and adjusting tendon slack
length to approximate the range of fiber operating lengths in the preoperative
simulation. A third model was created for each subject in which the muscle’s hip

77
flexion moment was eliminated while its knee extension moment was preserved. A
fourth model was created for each subject in which rectus femoris was removed from
the model, eliminating both hip and knee moments.
The set of muscle excitations found for each subject’s preoperative simulation was
applied to each model to simulate the effects of each condition on knee flexion. We
compared the changes in peak knee flexion from the preoperative condition using a
two-tailed, paired t-test (p < 0.05) with Bonferroni correction.
4.4 RESULTS
The largest improvement in peak knee flexion (32° ± 8°) occurred after
simulated transfer of the rectus femoris to sartorius (Fig. 4.4). Transfer of the muscle
to the iliotibial band predicted a smaller (p < 0.001) improvement in peak knee flexion
(28° ± 8°). Simulated scarred rectus femoris transfer resulted in an average
improvement in peak knee flexion of 14° ± 5°, which was significantly less (p <
0.001) than the average improvement from unscarred transfer to either the sartorius or
the iliotibial band. Simulated botulinum toxin injection to the rectus femoris, which
eliminated the muscle’s active hip flexion and knee extension moments, resulted in an
average improvement of 12° ± 5° in knee flexion.

78
Figure 4.4 Average increases (and one standard deviation) in peak knee flexion of the subject group for each of the simulated treatments.
The knee extension moment of rectus femoris was more influential than its hip
flexion moment on peak knee flexion (Fig. 4.5). All subjects showed larger (p <
0.001) increase in peak knee flexion (28° on average) when knee extension moment
was eliminated compared to decrease in peak knee flexion (-8° on average) when hip
flexion moment was eliminated. In the absence of the muscle’s knee extension
moment, average improvement in peak knee flexion was 3° greater (p < 0.001) with
hip flexion moment than without hip flexion moment.
Figure 4.5 Average increases (and one standard deviation) in peak knee flexion of the subject group for simulations in which knee extension or hip flexion moments of rectus femoris were independently eliminated.

79
4.5 DISCUSSION
Our results suggest that the primary mechanism by which rectus femoris
transfer improves knee flexion is the reduction of the rectus femoris’ knee extension
moment. Previous observations that the surgery diminishes knee extension moment
but generally does not convert the rectus femoris to a knee flexor (Riewald and Delp,
1997; Asakawa et al., 2002) support this finding.
A secondary mechanism of improvement is preservation of some of the
muscle’s hip flexion moment, which induces knee flexion (Piazza and Delp, 1996;
Riley and Kerrigan, 1998). However, it is likely the hip flexion moment generated by
the rectus femoris is diminished after transfer. The hip flexion moment generated by
the rectus femoris could not be preserved in simulations in which the muscle’s
insertion was relocated. In the preoperative simulation, the rectus femoris lengthened
due to the patella translating distally on the femur with knee flexion. In the transfer
simulations in which the rectus femoris was relocated from the patella the muscle
shortened. Concentric muscle contraction in the transfer simulations produced less
force than the eccentric muscle contraction in the preoperative simulations; this
reduced the hip flexion moment generated by the rectus femoris in transfer
simulations. This suggests that when the rectus femoris is transferred from its
insertion on the patella the muscle’s hip flexion moment may not be preserved,
reducing the contribution of the muscle’s hip flexion moment to knee flexion and
increasing the importance of reducing, or eliminating, the muscle’s knee extension
moment.

80
Our results show that rectus femoris transfers to either the sartorius or the
iliotibial band predicted greater improvements in peak knee flexion than a scarred
transfer, suggesting that methods to reduce postoperative scarring of the muscle to
underlying vasti may be beneficial. Transfer to the sartorius predicted only slightly
greater peak knee flexion in swing (approximately 4°) than transfer to the iliotibial
band, even though the simulated transfer to the sartorius converted the muscle to a
knee flexor. Although the rectus femoris had a knee flexion moment arm in the model
of transfer to the sartorius, the amount of force the rectus femoris produced was
diminished after transfer since the change in muscle path caused the muscle to shorten
rather than lengthen while contracting. Transfer to semitendinosus or gracilis, which
have larger knee flexion moment arms than sartorius, may result in larger changes in
knee flexion. Transfer to sartorius was investigated in this study because of its
prevalence as a surgical option (Gage et al., 1987; Ounpuu et al., 1993a; Saw et al.,
2003; Moreau et al., 2005; Hemo et al., 2007).
Simulated botulinum toxin injection to the rectus femoris predicted less peak
knee flexion improvement than transfer of the muscle. This is likely due to the
preservation of detrimental passive knee extension moments (i.e., the knee extension
moment generated by stretch of the rectus femoris even when it is inactive) and
elimination of helpful active hip flexion moments. The simulated result may
overestimate anticipated clinical improvement since injection of botulinum toxin is
unlikely to achieve complete elimination of active muscle force. Stoquart et al. (2008)
reported a 5° average increase in peak knee flexion after botulinum toxin injection into
rectus femoris in adults with stiff-knee gait due to stroke. Other studies have reported

81
no significant change (Chantraine et al., 2005) and 9° average improvement (Sung and
Bang, 2000) in peak knee flexion after motor branch block of the rectus femoris in
patients with stiff-knee gait.
Simulations of rectus femoris transfers to the sartorius and the iliotibial band
resulted in larger peak knee flexion improvements than those reported clinically or
observed in the postoperative kinematics of our subjects (2° ± 13°) measured
approximately one year postoperatively. This is likely to occur because the rectus
femoris may not produce the intended knee flexion moment due to scarring of the
transferred muscle to underlying soft tissues (Asakawa et al., 2004). Also, the subjects
in this study were treated with other surgeries that may affect knee flexion. All ten
subjects received hamstrings lengthenings and many received additional bony or soft-
tissue surgeries. We have not attempted to model the effects of concomitant surgeries
in our subject group because our goal was to isolate the effects of rectus femoris
transfer surgery. Our simulations of scarred rectus femoris transfers predict
improvements in knee flexion of 14° on average, which is similar to a clinical report
of 12° of increase after isolated rectus femoris transfer (Hemo et al., 2007).
Several assumptions were made in implementing the gait simulations. First,
the muscle excitations used to drive the treatment simulations were assumed to remain
unchanged from preoperative simulations. While some muscle excitations are likely
to change, Patikas et al. (2007) found no significant changes in rectus femoris activity
after multilevel surgery, including rectus femoris transfer. Secondly, muscle
parameters prescribed in these models were based on data for typical adults, which are
unlikely to be accurate representations of muscles in children with cerebral palsy.

82
Additionally, our subject-specific models were scaled to the sizes of each subject and
accurately represented the kinematics and kinetics measured preoperatively for each
subject, but did not replicate skeletal deformities or contractures which may have been
present. Also, we cannot be certain that the muscle forces produced in our simulations
accurately represented the forces generated by individual subjects in vivo, although the
net joint moments and muscle excitations in the preoperative simulations were
consistent with those measured in the gait lab. Finally, we modeled the net effect of a
scarred transfer by reducing the average moment arm of rectus femoris at the knee by
half. This is an approximation of the function of a scarred transfer, and may not
represent secondary effects of scarring or variations in the actual moment arm of the
transferred muscle throughout a range of knee flexion. Although different modeling
assumptions may have resulted in different peak knee flexion improvements, the
relationships of improvement in peak knee flexion among treatments would not be
likely to change since the same assumptions were used in each treatment simulation.
Our dynamic simulations of individual subjects with stiff-knee gait revealed
that substantial improvement in peak knee flexion in swing after rectus femoris
transfer may be obtained by reducing the muscle’s knee extension moment.
Preserving the muscle’s hip flexion moment may provide some additional
improvement, but was less influential than decreasing the muscle’s knee extension
moment. Surgeries that intend to convert the rectus femoris to a knee flexor may
instead only reduce its knee extension moment, possibly due to scarring (Asakawa et
al., 2002). Reducing postoperative scarring of the rectus femoris to underlying tissues
may improve postoperative peak knee flexion in swing.

83
5 CONTRIBUTIONS OF MUSCLES AND PASSIVE
DYNAMICS TO SWING INITIATION OVER A RANGE OF WALKING SPEEDS 5.1 ABSTRACT
Stiff-knee gait is a common walking problem in cerebral palsy characterized
by insufficient knee flexion during swing. To identify factors that may limit knee
flexion in swing, it is necessary to understand how unimpaired subjects successfully
coordinate muscles and passive dynamics (gravity and velocity-related forces) to
accelerate the knee into flexion during double support, a critical phase just prior to
swing that establishes the conditions for achieving sufficient knee flexion during
swing. It is also necessary to understand how contributions to swing initiation change
with walking speed, since patients with stiff-knee gait often walk slowly. We
analyzed muscle-driven dynamic simulations of eight unimpaired subjects walking at
four speeds to quantify the contributions of muscles, gravity, and velocity-related
forces (i.e. Coriolis and centrifugal forces) to preswing knee flexion acceleration
during double support at each speed. Analysis of the simulations revealed
contributions from muscles and passive dynamics varied systematically with walking
speed. Preswing knee flexion acceleration was achieved primarily by hip flexor
muscles on the preswing leg with assistance from biceps femoris short head. Hip
flexors on the preswing leg were primarily responsible for the increase in preswing
knee flexion acceleration during double support with faster walking speed. The hip
extensors and abductors on the contralateral leg and velocity-related forces opposed
preswing knee flexion acceleration during double support.

84
5.2 INTRODUCTION
The biomechanical causes of diminished and delayed swing-phase knee
flexion, or stiff-knee gait, in children with cerebral palsy are unclear, making it
difficult to determine appropriate treatment. Over-activity of the rectus femoris is
commonly thought to be the primary cause of stiff-knee gait (Perry, 1987; Sutherland
et al., 1990), yet many patients do not improve after rectus femoris transfer (Hadley et
al., 1992; Ounpuu et al., 1993a; Rethlefsen et al., 1999; Yngve et al., 2002; Carney
and Oeffinger, 2003), a surgery aimed at reducing the muscle’s knee extension
moment, suggesting that there may be other causes in some cases. Other proposed
causes of stiff-knee gait include over-activity of the vasti (Waters et al., 1979;
Kerrigan et al., 1991), weakness of the hip flexors (Kerrigan and Riley, 1998), and
weakness of the ankle plantarflexors (Kerrigan and Glenn, 1994). A better
understanding of the factors that contribute to stiff-knee gait will allow clinicians to
employ treatment strategies that address underlying causes.
To determine the cause of an individual’s stiff-knee gait, it is necessary to
understand how muscles and passive dynamics (gravity and velocity-related forces)
contribute to swing initiation in normal gait. Preswing has been identified as a key
portion of the gait cycle affecting swing-phase peak knee flexion because the muscle
forces produced during preswing determine the knee flexion velocity at toe-off, which
is highly correlated to swing-phase peak knee flexion (Mochon and McMahon, 1980;
Piazza and Delp, 1996; Goldberg et al., 2003; Reinbolt et al., 2008). However, a
thorough understanding of the biomechanical factors that accelerate the knee during

85
this period does not exist. Understanding these factors can be challenging because
muscles that do not cross the knee can still accelerate the knee due to dynamic
coupling (Zajac and Gordon, 1989). Modeling and simulation tools are valuable in
analyzing gait dynamics because they enable quantification of the effects of muscles,
gravity, and velocity-related forces on knee flexion acceleration.
It is necessary to understand how contributions to swing initiation change with
walking speed, since patients with stiff-knee gait often walk more slowly than
typically developing children. Walking speed affects kinematics, kinetics, and muscle
activity during gait (Andriacchi et al., 1977; Murray et al., 1984; Kirtley et al., 1985;
Shiavi et al., 1987; Stansfield et al., 2001b; Stansfield et al., 2001a; Hof et al., 2002;
van der Linden et al., 2002; den Otter et al., 2004; Nymark et al., 2005; Cappellini et
al., 2006; Stansfield et al., 2006; Schwartz et al., 2008). The contributions of muscles
and passive dynamics to support and progression of the body’s mass center (Liu et al.,
2008; Neptune et al., 2008) and knee flexion in swing (Arnold et al., 2007a) change
with walking speed. Thus, it is essential to understand how contributions of muscles
and passive dynamics to swing initiation may change with walking speed.
Comparison of contributors to swing initiation between a child with stiff-knee gait and
a typically developing child walking at a similar speed will enable discrimination
between differences due to pathology or walking speed.
The objectives of this study were to identify the major contributors to preswing
knee flexion acceleration during double support and to determine how these
contributions change with walking speed.

86
5.3 METHODS
We analyzed simulations created and tested by Liu et al. (2008) using
OpenSim software (Delp et al., 2007). The software and simulations are freely
available at http://simtk.org. Liu et al. (2008) created these simulations to quantify
muscle contributions to support and progression during walking. In this study, we
have analyzed the simulations to determine muscular and passive contributions to knee
flexion acceleration during double support. The double support period of simulations
of eight unimpaired subjects walking at four speeds was analyzed. The subjects’ ages
ranged from 7 to 18 years with a mean of 12.9 years. Protocols for collection and
processing of gait data, including ground reaction forces, kinematics, and
electromyographic (EMG) recordings, were reported by Schwartz et al. (2008).
Walking trials for each subject were assigned post-hoc to categories of very slow,
slow, free, and fast speeds as described by Liu et al. (2008) using a non-
dimensionalized walking speed leggLvv /*= , where v is absolute walking velocity,
Lleg is leg length, and g is gravitational acceleration (Hof, 1996). Average walking
speeds were 0.54 m/s for very slow, 0.75 m/s for slow, 1.15 m/s for free, and 1.56 m/s
for fast.
The musculoskeletal model and procedures for creating and testing the
simulations is described in detail elsewhere (Liu et al., 2008). Briefly, a generic
musculoskeletal model (Delp et al., 1990; Thelen and Anderson, 2006) with 23
degrees of freedom and 92 muscle-tendon actuators was scaled to match each
subject’s anthropometry. Subtalar and metatarsophalangeal joints were locked at

87
neutral anatomical angles. Dynamic inconsistency between the measured ground
reaction forces and the model kinematics was resolved by applying small external
forces and torques (i.e. residuals) to the pelvis and making small adjustments to the
model’s mass properties and kinematics (Delp et al., 2007). Computed muscle control
(Thelen et al., 2003), with constraints on muscle excitations applied as necessary, was
used to find a set of actuator excitations that when applied to the model in concert with
external ground reaction forces would both track the experimental kinematics and be
generally consistent with experimental and literature-reported EMG patterns. We
verified that the excitation patterns from the simulations at the different walking
speeds generally scaled with speed as reported in the literature (Hof et al., 2002; den
Otter et al., 2004; Cappellini et al., 2006; Schwartz et al., 2008).
In each simulation, we quantified the contributions of individual muscles,
gravity, and velocity-related forces to preswing knee flexion acceleration using a
perturbation analysis (Liu et al., 2006a). This analysis independently calculated the
contribution from each force (individual muscles, gravity, or velocity-related) and was
repeated for all contributors in the simulation at 10 ms intervals throughout double
support. For each muscle, we added 1 N to the force produced by an individual
muscle in the simulation, integrated forward the equations of motion for a 10 ms
period, and observed the resulting change in preswing knee flexion angle. The
resulting change in preswing knee flexion angle was used to calculate the preswing
knee flexion acceleration generated by 1 N of muscle force, assuming that the
acceleration generated by the muscle over the short 10 ms integration period was
constant. The preswing knee flexion acceleration generated by 1 N of muscle force

88
was multiplied by the muscle’s force in the unperturbed simulation to quantify the
muscle’s contribution to preswing knee flexion acceleration at the beginning of the 10
ms period. Translational and rotational spring-dampers applied to the center of
pressure of each foot accounted for changes in the ground reaction force induced by
the muscle perturbation. A similar technique was applied to determine the
contribution of gravity to preswing knee flexion acceleration. To quantify
contributions from velocity-related forces, the model was set in its original
configuration and given its original velocities at the start of every 10 ms period. No
muscle, gravity, or ground reaction forces were applied; only reaction forces from the
spring-dampers representing foot-floor contact were applied. We integrated forward
the equations of motion for a 10 ms period and observed the resulting change in
preswing knee flexion angle. The contribution of velocity-related forces to preswing
knee flexion acceleration was calculated from this change in preswing knee flexion
angle, assuming that the acceleration generated by the velocity-related forces over the
short 10 ms integration period was constant. All contributions (muscle, gravity, and
velocity-related) were averaged over the period of double support in each simulation.
To test the validity of our method, we verified that the sum of all calculated
contributions to preswing knee flexion acceleration was in agreement with the
unperturbed preswing knee flexion acceleration at each time point of analysis
throughout double support (Fig. 5.1). This suggests that our model of foot-floor
contact is a reasonable representation of the constraints on the foot during double
support.

89
Figure 5.6 Knee flexion angle and acceleration of subject 5, a representative subject, for the simulation (thick gray line) compared to experimentally measured values (thin black line) during the free speed trial. The simulation closely tracked experimental knee flexion. Shaded region represents ± one standard deviation from the mean of the free speed trials of all eight subjects. Black dots represent the sum of all calculated contributions from the perturbation analysis at each step. Overlap of the black dots with the thick gray line indicates that the sum of contributors to preswing knee flexion acceleration calculated by the perturbation analysis closely approximated the knee flexion acceleration of the simulation.
We performed a one-way repeated measures analysis of variance (SPSS Inc.,
Chicago, IL) to determine if walking speed had a significant effect on the average
contributions of muscles, gravity, and velocity to preswing knee flexion acceleration.
For data that violated sphericity assumptions, a Huynh-Feldt epsilon correction was
applied. When speed was determined to significantly affect a contributor, we
analyzed the within-subject repeated contrasts to determine if there was a significant

90
difference between successive speed pairs (i.e., very slow to slow, slow to free, free to
fast). The significance level for all tests was α ≤ 0.05.
5.4 RESULTS
In preparation for swing, the preswing knee is strongly accelerated into flexion
during double support. Most of the flexion acceleration occurs before the toe leaves
the ground, resulting in a peak knee flexion velocity around toe-off (Fig. 5.2A).
Achieving a sufficient knee flexion velocity at toe-off is crucial to achieving sufficient
peak knee flexion in swing. Knee flexion velocity at toe-off increased with walking
speed (p < 0.05; Fig. 5.2B). This was achieved by an increase in average knee flexion
acceleration during double support with increased walking speed (p < 0.01; Fig. 5.2C).
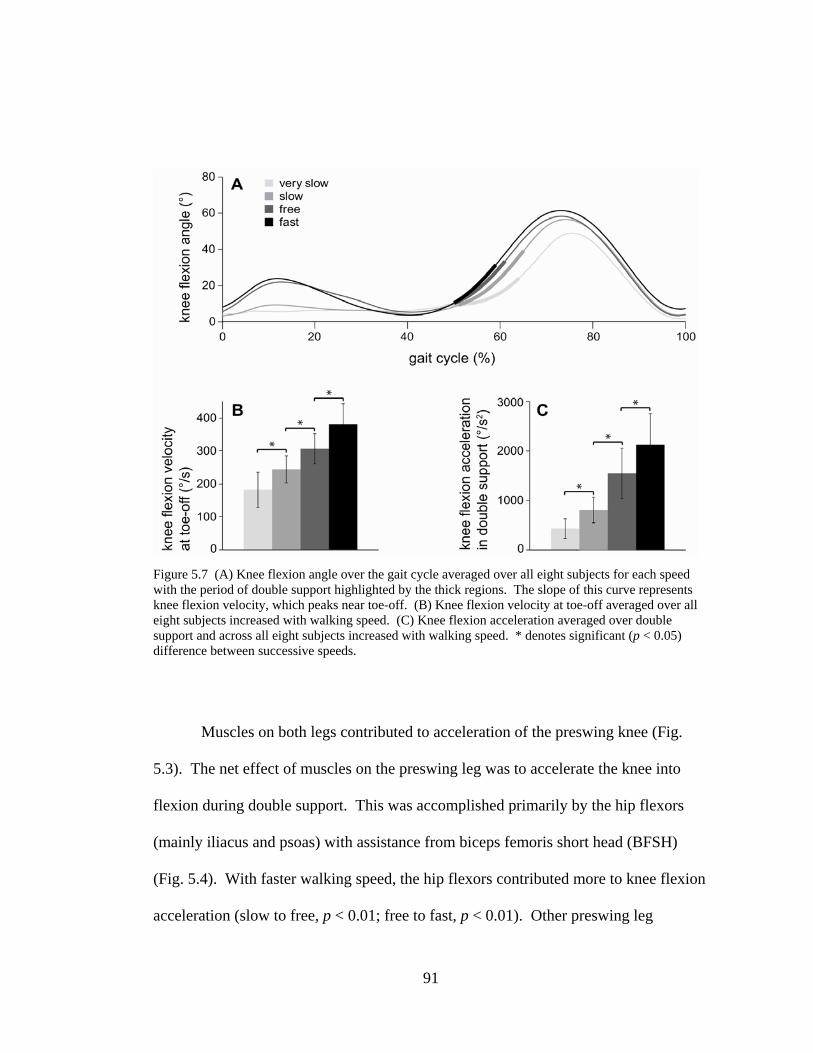
91
Figure 5.7 (A) Knee flexion angle over the gait cycle averaged over all eight subjects for each speed with the period of double support highlighted by the thick regions. The slope of this curve represents knee flexion velocity, which peaks near toe-off. (B) Knee flexion velocity at toe-off averaged over all eight subjects increased with walking speed. (C) Knee flexion acceleration averaged over double support and across all eight subjects increased with walking speed. * denotes significant (p < 0.05) difference between successive speeds.
Muscles on both legs contributed to acceleration of the preswing knee (Fig.
5.3). The net effect of muscles on the preswing leg was to accelerate the knee into
flexion during double support. This was accomplished primarily by the hip flexors
(mainly iliacus and psoas) with assistance from biceps femoris short head (BFSH)
(Fig. 5.4). With faster walking speed, the hip flexors contributed more to knee flexion
acceleration (slow to free, p < 0.01; free to fast, p < 0.01). Other preswing leg

92
muscles, including gastrocnemius and the ankle dorsiflexors (DF), also contributed to
preswing knee flexion acceleration, but made small contributions relative to the
preswing hip flexors. Some muscles on the preswing leg decelerated knee flexion,
including the uniarticular plantarflexors (UPF) (mainly soleus), vasti, rectus femoris,
and the hip extensors and abductors. The net effect of muscles on the contralateral leg
at all speeds was to decelerate preswing knee flexion. This was primarily due to hip
extensors and abductors on the contralateral leg. Hip flexors on the contralateral leg
opposed the extension effect, but to a lesser extent. Contributions from back muscles
and residual forces and torques varied across subjects, but were small on average.
Average residual contributions to knee flexion acceleration across subjects at each
speed were -417 °/s2 for very slow, -170 °/s2 for slow, -1646 °/s2 for free, and -939 °/s2
for fast.
The contributions of passive dynamics during double support varied
systematically with walking speed (Fig. 5.3). Gravity accelerated the knee into flexion
(Movie 1) with a relatively constant magnitude across speeds, though slightly greater
at free and fast speeds (p < 0.01). Velocity-related (Coriolis and centrifugal) forces
mildly decelerated preswing knee flexion with an increasing effect with faster walking
speed (very slow to slow, p < 0.001; slow to free, p < 0.001; free to fast, p = 0.012).
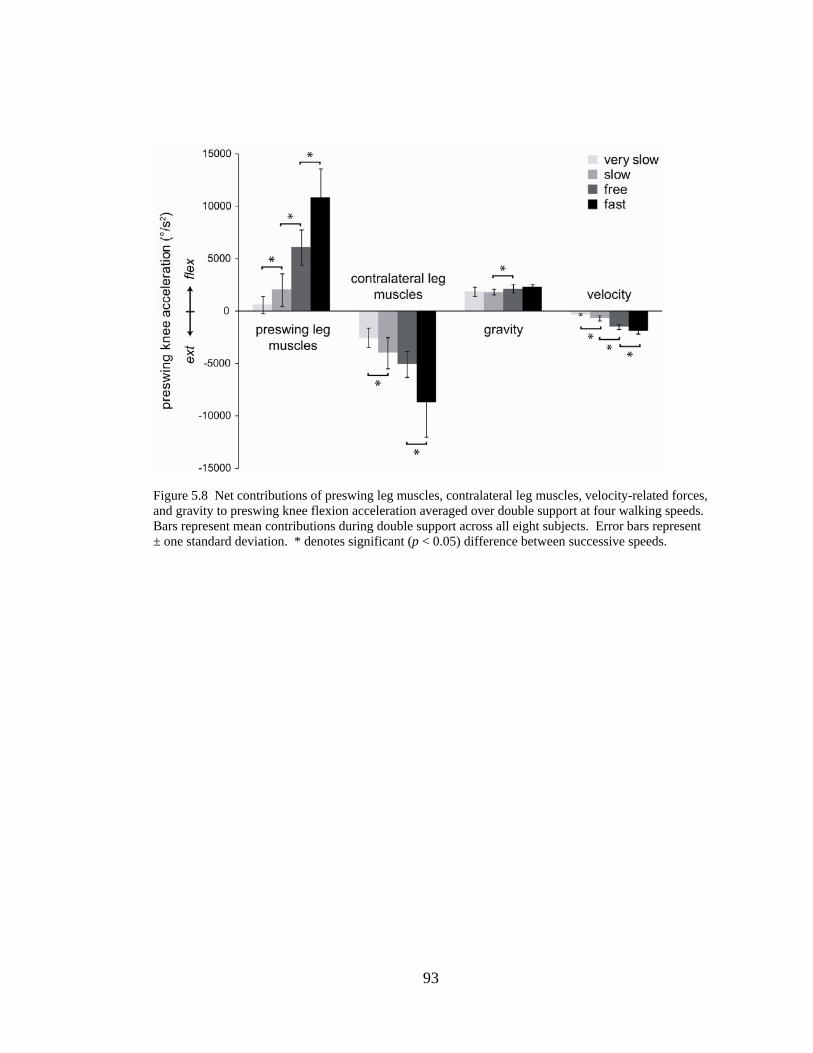
93
Figure 5.8 Net contributions of preswing leg muscles, contralateral leg muscles, velocity-related forces, and gravity to preswing knee flexion acceleration averaged over double support at four walking speeds. Bars represent mean contributions during double support across all eight subjects. Error bars represent ± one standard deviation. * denotes significant (p < 0.05) difference between successive speeds.
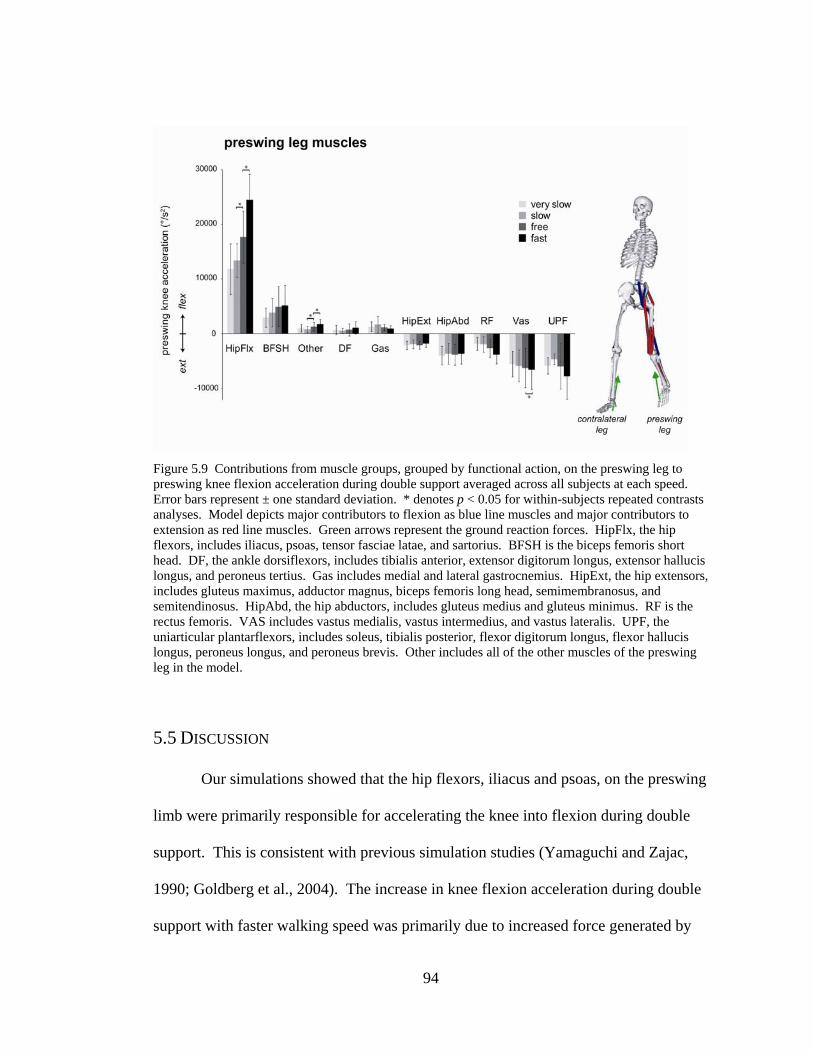
94
Figure 5.9 Contributions from muscle groups, grouped by functional action, on the preswing leg to preswing knee flexion acceleration during double support averaged across all subjects at each speed. Error bars represent ± one standard deviation. * denotes p < 0.05 for within-subjects repeated contrasts analyses. Model depicts major contributors to flexion as blue line muscles and major contributors to extension as red line muscles. Green arrows represent the ground reaction forces. HipFlx, the hip flexors, includes iliacus, psoas, tensor fasciae latae, and sartorius. BFSH is the biceps femoris short head. DF, the ankle dorsiflexors, includes tibialis anterior, extensor digitorum longus, extensor hallucis longus, and peroneus tertius. Gas includes medial and lateral gastrocnemius. HipExt, the hip extensors, includes gluteus maximus, adductor magnus, biceps femoris long head, semimembranosus, and semitendinosus. HipAbd, the hip abductors, includes gluteus medius and gluteus minimus. RF is the rectus femoris. VAS includes vastus medialis, vastus intermedius, and vastus lateralis. UPF, the uniarticular plantarflexors, includes soleus, tibialis posterior, flexor digitorum longus, flexor hallucis longus, peroneus longus, and peroneus brevis. Other includes all of the other muscles of the preswing leg in the model.
5.5 DISCUSSION
Our simulations showed that the hip flexors, iliacus and psoas, on the preswing
limb were primarily responsible for accelerating the knee into flexion during double
support. This is consistent with previous simulation studies (Yamaguchi and Zajac,
1990; Goldberg et al., 2004). The increase in knee flexion acceleration during double
support with faster walking speed was primarily due to increased force generated by

95
the hip flexors. Neptune et al. (2008) also found a dramatic increase in iliopsoas
muscle work to accelerate the preswing leg at faster walking speeds. In our subjects,
peak hip flexion moment during double support increased three-fold between very
slow and fast speeds, a larger increase than either the knee or ankle moments. Large
increases in hip moment during double support with increasing speed were also
observed in other studies (Stansfield et al., 2001b; van der Linden et al., 2002;
Schwartz et al., 2008). Additionally, we found the vasti, rectus femoris, and hip
abductors and extensors decelerate knee flexion during double support, consistent with
other studies (Neptune et al., 2001, Fig. 8; Goldberg et al., 2004).
Our results showed the two major plantarflexors, soleus and gastrocnemius,
had opposite effects on preswing knee acceleration. The knee extension acceleration
of soleus is consistent with other studies (Goldberg et al., 2004; Neptune et al., 2001,
Fig. 8). The mild knee flexion acceleration of gastrocnemius that we found contrasts
with reports that the gastrocnemius has a large flexion effect (Yamaguchi and Zajac,
1990; Goldberg et al., 2004) or an extension effect (Neptune et al., 2001, Fig. 8) on the
preswing knee. The gastrocnemius generates a plantarflexion moment that induces
knee extension acceleration and a knee flexion moment that induces knee flexion
acceleration; thus, its action is sensitive to the muscle’s ankle and knee moment arms,
body position, and foot contact model, which varied among the studies. Although the
net effect of the plantarflexors may be to extend the knee, studies have suggested they
play a role in swing initiation since it has been observed that hip flexors compensate
during preswing when plantarflexors are weak (Nadeau et al., 1999) or absent

96
(Zmitrewicz, 2007). Neptune et al. (2008) suggested that gastrocnemius contributes to
swing initiation by delivering energy to the preswing leg.
The roles of hip flexion and ankle plantarflexion moments in swing initiation
have been demonstrated in dynamic walking models. Kuo et al (2002) reported that
an increased hip torque produces increased step frequency, while an increased toe-off
impulse produces longer steps at an approximately constant step frequency; they
suggested that a combination of hip work and toe-off impulse may improve walking
energetics. Other studies have demonstrated a torque applied at the hip and/or a push-
off impulse applied to the foot can produce stable gait on level ground (Collins et al.,
2005).
In our simulations, the contralateral leg muscles contributed an extension
acceleration to the preswing knee at all speeds through their action on the pelvis.
Muscles on the contralateral leg including hip extensors, posterior hip abductors
(which have hip extension moment arms), and vasti extended the knee and hip of the
contralateral leg. In the body configuration of double support, this caused the pelvis to
tilt posteriorly, list upward on the side of preswing leg, and rise slightly. As a result,
the hip joint on the preswing leg was pushed upward (superiorly) and forward
(anteriorly). Reaction forces at the preswing hip joint extended the preswing knee
since the foot of the preswing leg remained on the ground during double support
(Movie 2). At faster walking speeds, the contralateral leg muscles had a stronger
deceleration effect on preswing knee flexion (Fig. 5.3).
Classic texts and studies of passive dynamic walkers have suggested that
muscle activity during preswing sets the initial conditions for passive knee motion

97
during swing (Mochon and McMahon, 1980; Mcgeer, 1990; Perry and Newsam, 1992;
Gage and Schwartz, 2004; Boakes, 2006). However, muscles are active during swing,
and studies muscle-actuated models have found that this muscle activity contributes
during both preswing and early swing to achieve appropriate knee motion before and
during swing. Although this study focused on double support, muscular or passive
forces during early swing phase (the period of swing before peak knee flexion) may
also affect peak knee flexion during swing. The knee undergoes a large flexion
acceleration during double support to prepare for toe-off (Fig. 5.2C). After toe-off the
knee undergoes an extension acceleration throughout early swing (Arnold et al.,
2007b). During double support or early swing, forces causing inadequate knee flexion
acceleration or excessive knee extension acceleration may limit peak knee flexion
during swing, resulting in stiff-knee gait. In this study we found that during double
support the preswing knee is accelerated into flexion mainly by swing leg muscles
(primarily the hip flexors and biceps femoris) and gravity at all speeds. Arnold et al.
(2007a; 2007b) found that during early swing the swing knee is extended by stance leg
muscles (mainly vasti and uniarticular plantarflexors) and velocity-related forces at all
speeds. It is necessary to analyze muscle contributions during both preswing and early
swing to investigate possible causes of stiff-knee gait.
Our results should be interpreted in light of several limitations of this study.
First, our estimates of muscle contributions to knee flexion acceleration were
dependent on the force produced by each muscle during the simulation. Although
experimental joint moments and EMG were generally consistent with simulated
values, it was not feasible to compare simulated muscle forces to experimentally

98
measured forces. Secondly, residual actuators in the model, which applied external
forces and torques to the pelvis, contributed to knee flexion acceleration with a
magnitude comparable to net muscle contribution in some trials. These residual
actuators do not represent real physical forces, but instead characterize errors in
kinematic and kinetic measurements and deficiencies in the model, such as the lack of
arms and joint simplifications. We chose a method to reduce rather than eliminate
these residual forces, as the latter can result in implausible motions of the back. In
these trials, net muscle contributions were affected, but the relative magnitude of
individual muscle contributions was consistent with trials in which residuals did not
contribute substantially to knee flexion acceleration. Thirdly, although care was taken
to validate simulated muscle activations with experimental EMG, rectus femoris and
gastrocnemius activations for some subjects during double support did not increase
with speed as expected (Hof et al., 2002; den Otter et al., 2004; Cappellini et al., 2006;
Schwartz et al., 2008). If activations and forces in the gastrocnemius and rectus
femoris were greater, we would have observed slightly larger contributions to knee
acceleration.
This study identifies the factors that contribute to knee flexion acceleration
during double support and provides a framework for future studies to investigate the
muscular and passive contributions to knee flexion acceleration in subjects with stiff-
knee gait. Further studies may further elucidate the role of the ankle plantarflexors in
other aspects of swing initiation, such as forward propulsion of the swing leg.

99
6 PRELIMINARY WORK: RECTUS FEMORIS VELOCITIES
BEFORE AND AFTER RECTUS FEMORIS LENGTHENING SURGERY
6.1 INTRODUCTION
There is a need for evidence-based clinical indicators for rectus femoris
lengthening surgery. Rectus femoris intramuscular lengthening surgery intends to
increase or maintain peak knee flexion in swing when simultaneous hamstrings
lengthening is performed. A proposed mechanism by which this surgery affects knee
motion is a reduction of rectus femoris spasticity that limits rectus femoris lengthening
velocity and knee flexion prior to surgery. However, it is unclear whether rectus
femoris lengthening velocity increases after surgery or whether greater rectus femoris
velocity leads to improved knee range of motion during gait. We have conducted some
preliminary investigation to determine whether peak rectus femoris lengthening
velocity increases after rectus femoris lengthening surgery and whether preoperative
peak rectus femoris lengthening velocity is predictive of increase in knee range of
motion following surgery.
6.2 METHODS
We analyzed 42 subjects (68 limbs), with cerebral palsy who received rectus
femoris intramuscular lengthening. These patients were treated at Connecticut
Children’s Medical Center between 1991 and 2008, and many of them received other

100
surgeries in addition to rectus femoris intramuscular lengthening (e.g., 53 limbs also
received medial hamstring lengthenings). Subjects underwent both preoperative and
postoperative kinematic and kinetic data collection as part of the standard of care (Fig.
6.1). The average time of postoperative gait analysis after surgery was 18.7 months.
The average age at surgery was 9 ± 3 years.
Figure 6. 1 Average and standard deviation of knee flexion angle over gait cycle for rectus femoris lengthening limbs (red) and unimpaired control limbs (grey).
We positioned a three-dimensional musculoskeletal model (Delp et al., 1990)
with subjects’ hip and knee kinematics to estimate rectus femoris musculotendon
lengths (i.e., the origin-to-insertion lengths) over the gait cycle (Fig. 6.2). The
trajectory of musculotendon lengths over the gait cycle were low-pass filtered using a
second-order Butterworth filter with a cut-off frequency of 3 Hz. Rectus femoris
musculotendon velocities were then calculated by differentiating rectus femoris

101
lengths with respect to time (Fig. 6.3). For each limb, the peak rectus femoris
lengthening velocity, which occurs before toe-off, was identified. Peak rectus femoris
velocities were also calculated for 14 typically developing subjects. Musculotendon
velocities were normalized by peak velocity of the typical subjects during gait. Rectus
femoris peak velocities were considered “slow” if they were slower than two standard
deviations below the average of the typical group.

102
Figure 6. 2 Average and standard deviation of rectus femoris musculotendon lengths (upper) and velocities (lower) for unimpaired limbs (grey) normalized by peak length and velocity of the unimpaired group. Preoperative rectus femoris musculotendon length and velocity for a representative subject (red).

103
Univariate linear regression was used to test whether a significant correlation
existed between (1) preoperative peak rectus femoris lengthening velocity and change
in velocity after surgery and (2) preoperative peak rectus femoris velocity and
improvement in knee range of motion (defined as maximum minus minimum knee
flexion during gait) after surgery. Multivariate linear regression was used to determine
whether preoperative peak rectus femoris lengthening velocity was predictive of
change in knee range of motion during gait when accounting for confounding
variables (e.g. concomitant surgeries, walking speed, and age) which may influence
both the outcome variable and the predictor of interest. A subset of the limbs (n = 57)
were included in the multivariate model whose preoperative knee range of motion
during gait was less than two standard deviations below the average for the group of
typical subjects.
6.3 RESULTS
We found that 34 out of 68 limbs had faster peak RF lengthening velocities
after surgery. Rectus femoris muscles that were slower preoperatively tended to have
greater increases in lengthening velocity after rectus femoris intramuscular
lengthening (Fig. 6.4; p < 0.0001). Only 3 of the 21 rectus femoris muscles that were
not slow preoperatively lengthened faster after surgery.

104
Figure 6. 3 Normalized preoperative peak rectus femoris musculotendon lengthening velocities for each rectus femoris lengthening subject limb plotted against his or her change in normalized rectus femoris peak musculotendon lengthening velocity after surgery.
Preoperative rectus femoris peak velocity had a significant (p < 0.01;
determined by t-test) but weak (r2 = 0.10) correlation with improvement in knee range
of motion following surgery that included rectus femoris lengthening (Fig. 6.5).
Subjects with slower preoperative rectus femoris velocities showed a greater increase
in knee range of motion during gait.
Figure 6. 4 Normalized preoperative peak rectus femoris musculotendon lengthening velocities for each rectus femoris lengthening subject limb plotted against his or her change in knee range of motion during gait after surgery.

105
Multivariate linear regression showed that when confounding variables
(walking speed in particular) were accounted for, preoperative rectus femoris peak
lengthening velocity was not a significant predictor (p > 0.05) of change in knee range
of motion during gait following surgery (Table 6.1). Concomitant femoral derotation
osteotomy, however, was strongly (p < 0.001) predictive of a decrease in knee range
of motion during gait.
Table 6. 1 p-values and coefficients for variables in the multivariate model predicting change in knee range of motion after surgery.

106
6.4 DISCUSSION
Our results showed that subjects with slower rectus femoris lengthening
velocities preoperatively tended to show greater increases in rectus femoris
lengthening velocity after surgery. However, it is unclear whether increase in peak
rectus lengthening velocity is due to rectus femoris lengthening or other concomitant
surgeries, such as hamstrings lengthening. Most of the limbs analyzed received
concomitant hamstrings lengthening which may increases rectus femoris velocity by
increasing knee flexion velocity with improvement in knee range of motion during
gait. Despite the surgical cause, we observed that rectus femoris muscles that were not
slow preoperatively did not tend to get faster after surgery.
Preoperative rectus femoris lengthening velocity was not significantly
predictive of improvement in knee range of motion after surgery when accounting for
confounding variables. There may be several explanations for this finding. One
possibility is that rectus femoris lengthening may reduce rectus femoris spasticity, but
other factors, such as concomitant surgeries may have a greater effect on knee range of
motion. This finding seems to be suggested by the large impact other surgeries, such
as femoral derotation osteotomy and gastrocnemius lengthening, had on range of
motion in the multivariate model. Another possibility is that slow rectus femoris
lengthening velocity may not be caused by rectus femoris spasticity. For example,
hamstrings spasticity may directly limit knee extension velocity in swing while
indirectly limiting knee flexion velocity, and therefore rectus femoris velocity, to
maintain balance. A final possible explanation is that rectus femoris musculotendon
lengthening velocity was not sufficiently representative of rectus femoris sapsaticity.

107
There are several potential criticisms and limitations of this study. It is unclear
to what extent overall musculotendon velocity is representative of muscle fiber
velocity in the rectus femoris. Although muscles with more compliant tendons are
more susceptible to inconsistencies between musculotendon length changes and fiber
length changes, it is unclear whether the assumption is valid for rectus femoris.
Additionally, we have not modeled the effects of bony deformities which may be
present in the limbs of our subjects. In particular, we have not accounted for the
effects of patella alta in estimating change in musculotendon lengths. If patella alta
affects the moment arm of the rectus femoris, this could impact the velocity of the
muscle over the gait cycle. However, it is difficult to account for the effect of patella
alta since there is currently no consensus in the literature on whether patella alta
increases or does not affect the moment arm of the knee extensors (Ward et al., 2005;
Luyckx 2009; Sheehan 2008). Additionally, it is possible that femoral anteversion may
affect rectus femoris lengths and velocities. We have addressed this possibility by
performing a sensitivity analysis of calculated rectus femoris musculotendon lengths
to varying degrees of femoral anteversion over the range of -80 to 90 degrees of
femoral anteversion. We found that peak rectus femoris musculotendon length over
the gait cycle varied by less than 2 % over this large range of femoral anteversion
angles, meaning that degree of femoral anteversion has an insignificant effect on
estimated muscultotendon lengths and velocities.
We used a multivariate model to assess the significance of one variable,
preoperative peak rectus femoris lengthening velocity, in predicting change in knee

108
range of motion while accounting for confounding factors such as age, involvement,
and concomitant surgeries. However, this data set and limited number of subjects is
not ideally suited for determining the best multivariate model, with the most powerful
combination of preoperative variables, for predicting change in knee range of motion
following surgery. An important next step would be to create a multivariate model,
with more subjects and a larger selection of preoperative predictive variables, to
determine whether some combination of preoperative data can improve predictions of
postoperative outcome.
Our multivariate model showed that concomitant femoral derotation osteotomy
had a negative effect on knee range of motion during gait after surgery. Another
important step would be to determine the cause of this effect. One would expect a
derotational osteotomy, as we observe for the tibial derotation osteotomy, to have a
restorative effect on muscle function and limb alignment, thereby resulting in
improved knee range of motion. It is possible that a non-included factor correlated
with the incidence of femoral derotation osteotomy is contributing to this effect.
Though our multivariate model accounted for overall involvement, the addition of a
preoperative severity of crouch variable may help shed some light on this
unanticipated result.
One final follow-up step would be to improve the accuracy of both the
musculoskeletal model and the statistical model. It is recommended to test the
accuracy of the rectus femoris musculotendon length and velocity measurements in the
presence of patella alta. If patella alta greatly affects these measurements it would be
necessary to obtain some clinical measure of patella height in order to accurately

109
represent it in the musculoskeletal model. Regarding the statistical model, much of the
data was taken from two limbs of a single subject, making these non-independent data.
A statistical correction is recommended to account for this effect since it breaks the
assumption of independent data in the multivariate model.

110
7 CONCLUSION 7.1 SUMMARY
The goal of this work was to clarify biomechanical causes of stiff-knee gait
and the functional mechanism of surgical treatment. The motivation for this work was
to contribute to the understanding of stiff-knee gait as a step towards improving
indications for treatment and reducing the variability of surgical outcomes. We used
computer simulation of gait to quantify muscle function during inappropriate timing,
after surgical transfer, and in unimpaired gait. We also implemented an addition to
perturbation analysis to allow comprehensive quantification of contributions to knee
flexion acceleration.
The findings of this work clarify the indications for rectus femoris transfer and
illuminate muscle coordination of normal gait. The main conclusions of this research
are:
• Preswing rectus femoris activity can contribute to stiff-knee gait
In our comparison of the simulated effects of preswing to early swing phase
activity of rectus femoris in stiff-knee gait, we identified preswing rectus femoris
activity as a potential contributor to stiff-knee gait. In many subjects, preswing
rectus femoris activity had a greater limiting effect on peak knee flexion in swing
than early swing activity. Our results suggest that preswing rectus femoris EMG
may be a useful indicator for surgical treatment. Additionally, comparison of
patient EMG to speed-matched unimpaired EMG may help to diagnose improper

111
preswing rectus femoris activity, since unimpaired subjects exhibit preswing rectus
femoris activity at free and fast speeds while many patients with stiff-knee gait
walk at slower speeds. Care should be taken in interpreting surface EMG of the
rectus femoris, as it may be subject to cross-talk from the vasti (Barr et al., 2010).
• Rectus femoris transfer improves knee flexion primarily by reduction of the
muscle’s knee extension moment
By comparing the simulated knee flexion resulting from models of rectus femoris
transfer surgery, we determined that reduction of the muscle’s knee extension
moment to values comparable to experimental measures (Asakawa et al., 2002)
was sufficient to achieve improved peak knee flexion. Simulated peak knee flexion
improvement using the scarred transfer model was most similar to average
clinically reported improvement in peak knee flexion following isolated transfer
(Hemo et al., 2007).
• Hip flexor muscles are the largest contributors to knee flexion acceleration
during double support in unimpaired gait
Comparing contributions averaged over double support to knee flexion
acceleration from muscles and passive dynamics, we determined hip flexors to be
the primary contributors at all speeds. Hip flexor muscles were also primarily
responsible for increasing knee flexion acceleration in double support with
increasing walking speed.

112
• Muscles other than rectus femoris may contribute to stiff-knee gait in
children with cerebral palsy
Our study of muscle contributions in chapter 5, revealed that several muscles
contribute to double support knee flexion acceleration. Diminished force in the
contributors, such as hip flexors and biceps femoris short head, may contribute to
stiff-knee gait. Additionally, over-activity in the muscles that contribute to knee
extension acceleration, such as vasti and soleus, may cause stiff-knee gait.
7.2 FUTURE WORK
Though this dissertation work has important implications for the diagnosis and
treatment of stiff-knee gait, much additional work is needed to improve treatment
outcomes. In order to correctly diagnosis the cause of stiff-knee gait, more research is
required to identify contributors to stiff-knee gait, aside from rectus femoris over-
activity, in children with cerebral palsy. Though our study in chapter 5 investigated
potential muscle contributors to stiff-knee gait, it is necessary to evaluate the
prevalence of these contributors in a clinical population and to establish protocols for
diagnosing these causes. Additionally, clinical studies evaluating the efficacy of
interventions targeting these additional muscle contributors would be a valuable step
in improving treatment planning. There may be additional contributors to stiff-knee
gait that are not muscular in origin. For example, it is unknown how bony deformities,
such as femoral anteversion or patella alta, may affect the ability of muscles to effect
knee motion. Another potential cause of stiff-knee gait that deserves more research
effort is the relationship between stiff-knee and crouch gait. Although some

113
preliminary work has investigated the relationship (van der Krogt et al., 2010), it
remains unclear whether crouch gait can lead to stiff-knee gait, or what the appropriate
treatment to address this cause should be.
More thorough investigation into the biomechanical effects of surgical
treatment could also lead to improved surgical technique and patient selection. In
chapter 4, we modeled the effects of scarring of the rectus femoris by altering the
muscle’s capacity to generate a knee moment, but more detailed models representing
force transmission between muscles through connective tissue could provide
additional insight into the transferred muscle’s postoperative function. Furthermore,
muscle remodeling and neural adaptation following musculoskeletal surgery is not
well understood, and could have important implications for surgical technique and
postoperative rehabilitation procedures.
A related area of future research that has the potential for immediate clinical
impact is identifying better indications for rectus femoris surgeries. To reduce
variability in outcomes, and prevent subjects who would have poor outcomes from
having the surgery, it is necessary to refine the current indications for rectus femoris
transfer and lengthening surgeries. Some preliminary work has identified outcome-
based quantitative metrics for recommending rectus femoris transfer surgery (Reinbolt
et al., 2009). Additional work may identify a model with stronger, more clinically
relevant predictors and may test the model across multiple institutions.

114
8 REFERENCES Abdulhadi, H.M., Kerrigan, D.C. and LaRaia, P.J., 1996. Contralateral shoe-lift: effect on oxygen cost of walking with an immobilized knee. Arch Phys Med Rehabil 77, 670-2. Adolfsen, S.E., Ounpuu, S., Bell, K.J. and DeLuca, P.A., 2007. Kinematic and kinetic outcomes after identical multilevel soft tissue surgery in children with cerebral palsy. J Pediatr Orthop 27, 658-67. Albert, T.A., Yelnik, A., Bonan, I., Lebreton, F. and Bussel, B., 2002. Effectiveness of femoral nerve selective block in patients with spasticity: preliminary results. Arch Phys Med Rehabil 83, 692-6. Anderson, F.C., Goldberg, S.R., Pandy, M.G. and Delp, S.L., 2004. Contributions of muscle forces and toe-off kinematics to peak knee flexion during the swing phase of normal gait: an induced position analysis. J Biomech 37, 731-7. Anderson, F.C. and Pandy, M.G., 1999. A Dynamic Optimization Solution for Vertical Jumping in Three Dimensions. Comput Methods Biomech Biomed Engin 2, 201-231. Anderson, F.C. and Pandy, M.G., 2003. Individual muscle contributions to support in normal walking. Gait Posture 17, 159-69. Andriacchi, T.P., Ogle, J.A. and Galante, J.O., 1977. Walking speed as a basis for normal and abnormal gait measurements. J Biomech 10, 261-8. Arnold, A.S., Anderson, F.C., Pandy, M.G. and Delp, S.L., 2005. Muscular contributions to hip and knee extension during the single limb stance phase of normal gait: a framework for investigating the causes of crouch gait. J Biomech 38, 2181-9. Arnold, A.S., Schwartz, M.H., Thelen, D.G. and Delp, S.L., 2007a. Contributions of muscles to terminal-swing knee motions vary with walking speed. J Biomech 40, 3660-71. Arnold, A.S., Thelen, D.G., Schwartz, M.H., Anderson, F.C. and Delp, S.L., 2007b. Muscular coordination of knee motion during the terminal-swing phase of normal gait. J Biomech 40, 3314-24. Arsenault, A.B., Winter, D.A. and Marteniuk, R.G., 1986. Is there a 'normal' profile of EMG activity in gait? Med Biol Eng Comput 24, 337-43. Asakawa, D.S., Blemker, S.S., Gold, G.E. and Delp, S.L., 2002. In vivo motion of the rectus femoris muscle after tendon transfer surgery. J Biomech 35, 1029-37. Asakawa, D.S., Blemker, S.S., Rab, G.T., Bagley, A. and Delp, S.L., 2004. Three-dimensional muscle-tendon geometry after rectus femoris tendon transfer. J Bone Joint Surg Am 86-A, 348-54.

115
Barr, K.M., Miller, A.L. and Chapin, K.B., 2010. Surface electromyography does not accurately reflect rectus femoris activity during gait: Impact of speed and crouch on vasti-to-rectus crosstalk. Gait Posture Bleck, E.E., 1987. Orthopaedic Management in Cerebral Palsy. MacKeith Press, Oxford. Boakes, J.L., Rab, G.T. , 2006. Muscle activity during walking. In: Rose (Eds.), Human Walking. Lippincott Williams and Wilkins, Baltimore, pp. Burgen, A.S., Dickens, F. and Zatman, L.J., 1949. The action of botulinum toxin on the neuro-muscular junction. J Physiol 109, 10-24. Cappellini, G., Ivanenko, Y.P., Poppele, R.E. and Lacquaniti, F., 2006. Motor patterns in human walking and running. J Neurophysiol 95, 3426-37. Carney, B.T. and Oeffinger, D., 2003. Sagittal knee kinematics following combined hamstring lengthening and rectus femoris transfer. J South Orthop Assoc 12, 149-53. Carney, B.T., Oeffinger, D. and Gove, N.K., 2006. Sagittal knee kinematics after rectus femoris transfer without hamstring lengthening. J Pediatr Orthop 26, 265-7. Caty, G.D., Detrembleur, C., Bleyenheuft, C., Deltombe, T. and Lejeune, T.M., 2008. Effect of simultaneous botulinum toxin injections into several muscles on impairment, activity, participation, and quality of life among stroke patients presenting with a stiff knee gait. Stroke 39, 2803-8. Cavanagh, P.R. and Komi, P.V., 1979. Electromechanical delay in human skeletal muscle under concentric and eccentric contractions. Eur J Appl Physiol Occup Physiol 42, 159-63. CDC, 2004. Centers for Disease Control and Prevention: Economic costs associated wtih mental retardation, cerebral palsy, hearing loss, and vision impairment. Morbidity and Mortality Weekly Report, 53, 57-59. Chambers, H., Lauer, A., Kaufman, K., Cardelia, J.M. and Sutherland, D., 1998. Prediction of outcome after rectus femoris surgery in cerebral palsy: the role of cocontraction of the rectus femoris and vastus lateralis. J Pediatr Orthop 18, 703-11. Chambers, H.G., 2001. Treatment of functional limitations at the knee in ambulatory children with cerebral palsy. Eur J Neurol 8 Suppl 5, 59-74. Chantraine, F., Detrembleur, C. and Lejeune, T.M., 2005. Effect of the rectus femoris motor branch block on post-stroke stiff-legged gait. Acta Neurol Belg 105, 171-7. Cheung, F., Tuzson, A., Abel, M.F. and Granata, K.P., 2003. Dynamic constraint of spasticity on muscle lengthening velocity during walking. Gait & Posture 18, S7. Chung, C.Y., Stout, J. and Gage, J.R., 1997. Rectus femoris transfer - Gracilis versus Sartorius. Gait & Posture 6, 137-146.

116
Cimolin, V., Piccinini, L., Turconi, A.C., Crivellini, M. and Galli, M., 2010. Are knee kinematic anomalies in swing due to rectus femoris spasticity different from those due to femoral anteversion in children with cerebral palsy? A quantitative evaluation using 3D gait analysis. J Pediatr Orthop B 19, 221-5. Collins, S., Ruina, A., Tedrake, R. and Wisse, M., 2005. Efficient bipedal robots based on passive-dynamic walkers. Science 307, 1082-5. Cottrell, G.W., 1963. Role of rectus femoris in spastic children. Proceedings: American Academy of Orthopedic Surgery. J Bone Joint Surg Am 45, 1556. Cruz, A.I., Õunpuu, S., Solomito, M. and DeLuca, P., 2009. Intramuscular Rectus Femoris Lengthening for the Correction of Stiff Knee Gait in Children with Cerebral Palsy: An Evaluation using 3D Motion Analysis. Gait and Clinical Movement Analysis Society, Csongradi, J., Bleck, E. and Ford, W.F., 1979. Gait electromyography in normal and spastic children, with special reference to quadriceps femoris and hamstring muscles. Dev Med Child Neurol 21, 738-48. Damron, T.A., Breed, A.L. and Cook, T., 1993. Diminished knee flexion after hamstring surgery in cerebral palsy patients: prevalence and severity. J Pediatr Orthop 13, 188-91. Delp, S.L., Anderson, F.C., Arnold, A.S., Loan, P., Habib, A., John, C.T., Guendelman, E. and Thelen, D.G., 2007. OpenSim: open-source software to create and analyze dynamic simulations of movement. IEEE Trans Biomed Eng 54, 1940-50. Delp, S.L. and Loan, J.P., 2000. A computational framework for simulating and analyzing human and animal movement. Computing in Science & Engineering 2, 46-55. Delp, S.L., Loan, J.P., Hoy, M.G., Zajac, F.E., Topp, E.L. and Rosen, J.M., 1990. An interactive graphics-based model of the lower extremity to study orthopaedic surgical procedures. IEEE Trans Biomed Eng 37, 757-67. Delp, S.L., Ringwelski, D.A. and Carroll, N.C., 1994. Transfer of the rectus femoris: effects of transfer site on moment arms about the knee and hip. J Biomech 27, 1201-11. DeLuca, P.A., Davis, R.B., Ounpuu, S., Rose, S. and Sirkin, R., 1997. Alterations in surgical decision making in patients with cerebral palsy based on three-dimensional gait analysis. J Pediatr Orthop 17, 608-614. den Otter, A.R., Geurts, A.C., Mulder, T. and Duysens, J., 2004. Speed related changes in muscle activity from normal to very slow walking speeds. Gait Posture 19, 270-8. Downing, A.L., Ganley, K.J., Fay, D.R. and Abbas, J.J., 2009. Temporal characteristics of lower extremity moment generation in children with cerebral palsy. Muscle Nerve 39, 800-9. Duncan, W.R., 1955. Release of the rectus femoris in spastic paralysis. Proceedings: American Academy of Orthopedic Surgery J Bone Joint Surg Am 37, 634.

117
Fox, M.D. and Delp, S.L., 2010. Contributions of muscles and passive dynamics to swing initiation over a range of walking speeds. J Biomech 43, 1450-5. Fox, M.D., Reinbolt, J.A., Ounpuu, S. and Delp, S.L., 2009. Mechanisms of improved knee flexion after rectus femoris transfer surgery. J Biomech 42, 614-9. Gage, J.R., 1990. Surgical treatment of knee dysfunction in cerebral palsy. Clin Orthop Relat Res 45-54. Gage, J.R., Fabian, D., Hicks, R. and Tashman, S., 1984. Pre- and postoperative gait analysis in patients with spastic diplegia: a preliminary report. J Pediatr Orthop 4, 715-25. Gage, J.R., Perry, J., Hicks, R.R., Koop, S. and Werntz, J.R., 1987. Rectus femoris transfer to improve knee function of children with cerebral palsy. Dev Med Child Neurol 29, 159-66. Gage, J.R. and Schwartz, M.H., 2004. Pathological gait and lever-arm dysfunction. In: Gage (Eds.), The Treatment of Gait Problems in Cerebral Palsy. Mac Keith Press, London, pp. 180-204. Gold, G.E., Asakawa, D.S., Blemker, S.S. and Delp, S.L., 2004. Magnetic resonance imaging findings after rectus femoris transfer surgery. Skeletal Radiol 33, 34-40. Goldberg, S.R., Anderson, F.C., Pandy, M.G. and Delp, S.L., 2004. Muscles that influence knee flexion velocity in double support: implications for stiff-knee gait. J Biomech 37, 1189-96. Goldberg, S.R., Ounpuu, S., Arnold, A.S., Gage, J.R. and Delp, S.L., 2006. Kinematic and kinetic factors that correlate with improved knee flexion following treatment for stiff-knee gait. J Biomech 39, 689-98. Goldberg, S.R., Ounpuu, S. and Delp, S.L., 2003. The importance of swing-phase initial conditions in stiff-knee gait. J Biomech 36, 1111-6. Granata, K.P., Abel, M.F. and Damiano, D.L., 2000. Joint angular velocity in spastic gait and the influence of muscle-tendon lengthening. J Bone Joint Surg Am 82, 174-86. Guerado, E. and de la Varga, V., 2001. Proximal rectus femoris lengthening. Orthopedics 24, 649-50. Hadley, N., Chambers, C., Scarborough, N., Cain, T. and Rossi, D., 1992. Knee motion following multiple soft-tissue releases in ambulatory patients with cerebral palsy. J Pediatr Orthop 12, 324-8. Hamner, S.R., Seth, A. and Delp, S.L., 2010. Muscle contributions to propulsion and support during running. J Biomech 43, 2709-16. Hebela, N. and Keenan, M.A., 2004. Neuro-orthopedic management of the dysfunctional extremity in upper motor neuron syndromes. Eura Medicophys 40, 145-56.

118
Hemo, Y., Aiona, M.D., Pierce, R.A., Dorociak, R. and Sussman, M.D., 2007. Comparison of rectus femoris transposition with traditional transfer for treatment of stiff knee gait in patients with cerebral palsy. J Child Orthop 1, 37-41. Hernandez, A., Lenz, A.L. and Thelen, D.G., 2010. Electrical stimulation of the rectus femoris during pre-swing diminishes hip and knee flexion during the swing phase of normal gait. IEEE Trans Neural Syst Rehabil Eng 18, 523-30. Hicks, J.L., Schwartz, M.H., Arnold, A.S. and Delp, S.L., 2008. Crouched postures reduce the capacity of muscles to extend the hip and knee during the single-limb stance phase of gait. J Biomech 41, 960-7. Hof, A.L., 1996. Scaling gait data to body size. Gait & Posture 4, 222-223. Hof, A.L., Elzinga, H., Grimmius, W. and Halbertsma, J.P., 2002. Speed dependence of averaged EMG profiles in walking. Gait Posture 16, 78-86. Hsu, L.C. and Li, H.S., 1990. Distal hamstring elongation in the management of spastic cerebral palsy. J Pediatr Orthop 10, 378-81. Inman, V.T., 1976. The Joints of the Ankle. Williams and Wilkins Co., Baltimore, MD. Johnson, B.A., D'Astous, J.L., Shuckra, A.L. and MacWilliams, B.A., 2011. Using Knee Kinematics in the Decision for Revision of Rectus Femoris Transfer. In Gait and Clinical Movement Analysis Society. Jonkers, I., Stewart, C., Desloovere, K., Molenaers, G. and Spaepen, A., 2006. Musculo-tendon length and lengthening velocity of rectus femoris in stiff knee gait. Gait Posture 23, 222-9. Kay, R.M., Rethlefsen, S.A., Kelly, J.P. and Wren, T.A., 2004. Predictive value of the Duncan-Ely test in distal rectus femoris transfer. J Pediatr Orthop 24, 59-62. Kerrigan, D.C., Bang, M.S. and Burke, D.T., 1999. An algorithm to assess stiff-legged gait in traumatic brain injury. J Head Trauma Rehabil 14, 136-45. Kerrigan, D.C., Burke, D.T., Nieto, T.J. and Riley, P.O., 2001a. Can toe-walking contribute to stiff-legged gait? Am J Phys Med Rehabil 80, 33-7. Kerrigan, D.C. and Glenn, M.B., 1994. An illustration of clinical gait laboratory use to improve rehabilitation management. Am J Phys Med Rehabil 73, 421-7. Kerrigan, D.C., Gronley, J. and Perry, J., 1991. Stiff-legged gait in spastic paresis. A study of quadriceps and hamstrings muscle activity. Am J Phys Med Rehabil 70, 294-300. Kerrigan, D.C., Karvosky, M.E. and Riley, P.O., 2001b. Spastic paretic stiff-legged gait: joint kinetics. Am J Phys Med Rehabil 80, 244-9.

119
Kerrigan, D.C. and Riley, P.O., 1998. The modelling of adult spastic paretic stiff-legged gait swing period based on actual kinematic data. Gait & Posture 7, 117-124. Kirtley, C., Whittle, M.W. and Jefferson, R.J., 1985. Influence of walking speed on gait parameters. J Biomed Eng 7, 282-8. Koca, K., Yildiz, C., Yurttas, Y., Bilgic, S., Ozkan, H., Kurklu, M., Balaban, B., Hazneci, B. and Basbozkurt, M., 2009. Outcomes of combined hamstring release and rectus transfer in children with crouch gait. Ortop Traumatol Rehabil 11, 333-8. Kuo, A.D., 2002. Energetics of actively powered locomotion using the simplest walking model. J Biomech Eng 124, 113-20. Lage, K.J., White, S.C. and Yack, H.J., 1995. The effects of unilateral knee immobilization on lower extremity gait mechanics. Med Sci Sports Exerc 27, 8-14. Lewek, M.D., Hornby, T.G., Dhaher, Y.Y. and Schmit, B.D., 2007. Prolonged quadriceps activity following imposed hip extension: a neurophysiological mechanism for stiff-knee gait? J Neurophysiol 98, 3153-62. Limpaphayom, N. and Prasongchin, P., 2011. Re-exploration of the rectus femoris tendon to sartorius transfer in children with spastic cerebral palsy. Asian Biomedicine 5, 143-149. Lin, C.J., Guo, L.Y., Su, F.C., Chou, Y.L. and Cherng, R.J., 2000. Common abnormal kinetic patterns of the knee in gait in spastic diplegia of cerebral palsy. Gait Posture 11, 224-32. Lin, Y.-C., Kim, H.J. and Pandy, M.G., 2011. A computationally efficient method for assessing muscle function during human locomotion. International Journal for Numerical Methods in Biomedical Engineering 27, 436-449. Liu, M.Q., Anderson, F.C., Pandy, M.G. and Delp, S.L., 2006a. Muscles that support the body also modulate forward progression during walking. Journal of biomechanics 39, 2623-2630. Liu, M.Q., Anderson, F.C., Pandy, M.G. and Delp, S.L., 2006b. Muscles that support the body also modulate forward progression during walking. J Biomech 39, 2623-30. Liu, M.Q., Anderson, F.C., Schwartz, M.H. and Delp, S.L., 2008. Muscle contributions to support and progression over a range of walking speeds. J Biomech 41, 3243-52. Luyckx, T., Didden, K., Vandenneucker, H., Labey, L., Innocenti, B. and Bellemans, J., 2009. Is there a biomechanical explanation for anterior knee pain in patients with patella alta?: influence of patellar height on patellofemoral contact force, contact area and contact pressure. J Bone Joint Surg Br 91, 344-50. Marks, M.C., Alexander, J., Sutherland, D.H. and Chambers, H.G., 2003. Clinical utility of the Duncan-Ely test for rectus femoris dysfunction during the swing phase of gait. Dev Med Child Neurol 45, 763-8.

120
Matjacic, Z. and Olensek, A., 2007. Biomechanical characterization and clinical implications of artificially induced crouch walking: Differences between pure iliopsoas, pure hamstrings and combination of iliopsoas and hamstrings contractures. J Biomech 40, 491-501. Matsuo, T., Hara, H. and Tada, S., 1987. Selective lengthening of the psoas and rectus femoris and preservation of the iliacus for flexion deformity of the hip in cerebral palsy patients. J Pediatr Orthop 7, 690-8. Mattsson, E. and Brostrom, L.A., 1990. The increase in energy cost of walking with an immobilized knee or an unstable ankle. Scand J Rehabil Med 22, 51-3. Mcgeer, T., 1990. Passive Dynamic Walking. International Journal of Robotics Research 9, 62-82. McMulkin, M.L., Baird, G.O., Barr, K.M., Caskey, P.M. and Ferguson, R.L., 2005. Proximal rectus femoris release surgery is not effective in normalizing hip and pelvic variables during gait in children with cerebral palsy. J Pediatr Orthop 25, 74-8. McNee, A.E., Shortland, A.P., Eve, L.C., Robinson, R.O. and Gough, M., 2004. Lower limb extensor moments in children with spastic diplegic cerebral palsy. Gait Posture 20, 171-6. Mena, D., Mansour, J.M. and Simon, S.R., 1981. Analysis and synthesis of human swing leg motion during gait and its clinical applications. J Biomech 14, 823-32. Miller, F., Cardoso Dias, R., Lipton, G.E., Albarracin, J.P., Dabney, K.W. and Castagno, P., 1997. The effect of rectus EMG patterns on the outcome of rectus femoris transfers. J Pediatr Orthop 17, 603-7. Mochon, S. and McMahon, T.A., 1980. Ballistic walking. J Biomech 13, 49-57. Mooney, V. and Goodman, F., 1969. Surgical approaches to lower-extremity disability secondary to strokes. Clin Orthop Relat Res 63, 142-52. Moreau, N., Tinsley, S. and Li, L., 2005. Progression of knee joint kinematics in children with cerebral palsy with and without rectus femoris transfers: a long-term follow up. Gait Posture 22, 132-7. Murray, M.P., Mollinger, L.A., Gardner, G.M. and Sepic, S.B., 1984. Kinematic and EMG patterns during slow, free, and fast walking. J Orthop Res 2, 272-80. Muthusamy, K., Seidl, A.J., Friesen, R.M., Carollo, J.J., Pan, Z. and Chang, F.M., 2008. Rectus femoris transfer in children with cerebral palsy: evaluation of transfer site and preoperative indicators. J Pediatr Orthop 28, 674-8. Muthusamy, K., Seidl, A.J., Fries, R.M., 2006. Rectus femoris transfer in children with cerebral palsy: evaluation of transfer site and preoperative indicators. Gait & Posture 24, S137-S138.

121
Nadeau, S., Gravel, D., Arsenault, A.B. and Bourbonnais, D., 1999. Plantarflexor weakness as a limiting factor of gait speed in stroke subjects and the compensating role of hip flexors. Clin Biomech (Bristol, Avon) 14, 125-35. Namdari, S., Pill, S.G., Makani, A. and Keenan, M.A., 2010. Rectus femoris to gracilis muscle transfer with fractional lengthening of the vastus muscles: a treatment for adults with stiff knee gait. Phys Ther 90, 261-8. Nene, A., Byrne, C. and Hermens, H., 2004. Is rectus femoris really a part of quadriceps? Assessment of rectus femoris function during gait in able-bodied adults. Gait Posture 20, 1-13. Nene, A., Mayagoitia, R. and Veltink, P., 1999. Assessment of rectus femoris function during initial swing phase. Gait Posture 9, 1-9. Nene, A.V., Evans, G.A. and Patrick, J.H., 1993. Simultaneous multiple operations for spastic diplegia. Outcome and functional assessment of walking in 18 patients. J Bone Joint Surg Br 75, 488-94. Neptune, R.R., Kautz, S.A. and Zajac, F.E., 2001. Contributions of the individual ankle plantar flexors to support, forward progression and swing initiation during walking. J Biomech 34, 1387-98. Neptune, R.R., Sasaki, K. and Kautz, S.A., 2008. The effect of walking speed on muscle function and mechanical energetics. Gait & Posture 28, 135-43. Nicholson, D.E., MacWilliams, B. A., D'Astous, J. L., 2000. Outcome prediction after distal hamstring lengthenings in children with cerebral palsy. Gait & Posture 11, 172-173. Niiler, T.A., Richards, J.G. and Miller, F., 2007. Concurrent surgeries are a factor in predicting success of rectus transfer outcomes. Gait Posture 26, 76-81. Nymark, J.R., Balmer, S.J., Melis, E.H., Lemaire, E.D. and Millar, S., 2005. Electromyographic and kinematic nondisabled gait differences at extremely slow overground and treadmill walking speeds. J Rehabil Res Dev 42, 523-34. Ounpuu, S., Muik, E., Davis, R.B., 3rd, Gage, J.R. and DeLuca, P.A., 1993a. Rectus femoris surgery in children with cerebral palsy. Part I: The effect of rectus femoris transfer location on knee motion. J Pediatr Orthop 13, 325-30. Ounpuu, S., Muik, E., Davis, R.B., 3rd, Gage, J.R. and DeLuca, P.A., 1993b. Rectus femoris surgery in children with cerebral palsy. Part II: A comparison between the effect of transfer and release of the distal rectus femoris on knee motion. J Pediatr Orthop 13, 331-5. Park, M.S., Chung, C.Y., Lee, S.H., Choi, I.H., Cho, T.J., Yoo, W.J., Park, B.S. and Lee, K.M., 2009. Effects of distal hamstring lengthening on sagittal motion in patients with diplegia: hamstring length and its clinical use. Gait Posture 30, 487-91.

122
Patikas, D., Wolf, S.I., Schuster, W., Armbrust, P., Dreher, T. and Doderlein, L., 2007. Electromyographic patterns in children with cerebral palsy: do they change after surgery? Gait Posture 26, 362-71. Patrick, J.H., 1996. Techniques of psoas tenotomy and rectus femoris transfer: "new" operations for cerebral palsy diplegia--a description. J Pediatr Orthop B 5, 242-6. Perry, J., 1987. Distal rectus femoris transfer. Dev Med Child Neurol 29, 153-8. Perry, J., 1992. Gait analysis: normal and pathological function. SLACK Inc., New Jersey. Perry, J., Hoffer, M.M., Antonelli, D., Plut, J., Lewis, G. and Greenberg, R., 1976. Electromyography before and after surgery for hip deformity in children with cerebral palsy. A comparison of clinical and electromyographic findings. J Bone Joint Surg Am 58, 201-8. Perry, J. and Newsam, C., 1992. Function of the hamstrings in cerebral palsy. In: Sussman (Eds.), The Diplegic Child: Evaluation and Management. American Academy of Orthopaedic Surgeons, Rosemont, IL, pp. 299-307. Piazza, S.J. and Delp, S.L., 1996. The influence of muscles on knee flexion during the swing phase of gait. Journal of biomechanics 29, 723-733. Piccinini, L., Cimolin, V., Turconi, A.C. and Galli, M., 2009. Relationship between kinematic knee deviations and femoral anteversion in children with cerebral palsy. Hip Int 19 Suppl 6, S63-8. Reinbolt, J.A., Fox, M.D., Arnold, A.S., Ounpuu, S. and Delp, S.L., 2008. Importance of preswing rectus femoris activity in stiff-knee gait. J Biomech 41, 2362-9. Reinbolt, J.A., Fox, M.D., Schwartz, M.H. and Delp, S.L., 2009. Predicting outcomes of rectus femoris transfer surgery. Gait & Posture 30, 100-5. Renshaw, T.S., Green, N.E., Griffin, P.P. and Root, L., 1995. Cerebral palsy: orthopaedic management. Journal of Bone and Joint Surgery 77-A, 1590-1606. Rethlefsen, S., Kay, R., Dennis, S., Forstein, M. and Tolo, V., 1999. The effects of fixed and articulated ankle-foot orthoses on gait patterns in subjects with cerebral palsy. J Pediatr Orthop 19, 470-4. Riewald, S.A. and Delp, S.L., 1997. The action of the rectus femoris muscle following distal tendon transfer: does it generate knee flexion moment? Dev Med Child Neurol 39, 99-105. Riley, P.O. and Kerrigan, D.C., 1998. Torque action of two-joint muscles in the swing period of stiff-legged gait: a forward dynamic model analysis. J Biomech 31, 835-40. Riley, P.O. and Kerrigan, D.C., 1999. Kinetics of stiff-legged gait: induced acceleration analysis. IEEE Trans Rehabil Eng 7, 420-6.

123
Robertson, J.V., Pradon, D., Bensmail, D., Fermanian, C., Bussel, B. and Roche, N., 2009. Relevance of botulinum toxin injection and nerve block of rectus femoris to kinematic and functional parameters of stiff knee gait in hemiplegic adults. Gait & Posture 29, 108-12. Rodda, J.M., Graham, H.K., Carson, L., Galea, M.P. and Wolfe, R., 2004. Sagittal gait patterns in spastic diplegia. J Bone Joint Surg Br 86, 251-8. Saw, A., Smith, P.A., Sirirungruangsarn, Y., Chen, S., Hassani, S., Harris, G. and Kuo, K.N., 2003. Rectus femoris transfer for children with cerebral palsy: long-term outcome. J Pediatr Orthop 23, 672-8. Schwartz, M.H., Rozumalski, A. and Trost, J.P., 2008. The effect of walking speed on the gait of typically developing children. J Biomech 41, 1639-50. Sheehan, F.T., Seisler, A.R. and Alter, K.E., 2008. Three-dimensional in vivo quantification of knee kinematics in cerebral palsy. Clin Orthop Relat Res 466, 450-8. Shiavi, R., 1985. Electromyographic patterns in adult locomotion: a comprehensive review. J Rehabil Res Dev 22, 85-98. Shiavi, R., Bugle, H.J. and Limbird, T., 1987. Electromyographic gait assessment, Part 1: Adult EMG profiles and walking speed. J Rehabil Res Dev 24, 13-23. Silfverskiold, N., 1923. Reduction of the uncrossed two joint muscles of the leg to one joint muscles in spastic conditions. Acta Chir Scand 56, 153-158. Stanley, F., Blair, E., Alberman, E., 2000. Cerebral Palsies: Epidemiology and Causal Pathways. MacKeith Press, London. Stansfield, B.W., Hillman, S.J., Hazlewood, M.E., Lawson, A.A., Mann, A.M., Loudon, I.R. and Robb, J.E., 2001a. Normalized speed, not age, characterizes ground reaction force patterns in 5-to 12-year-old children walking at self-selected speeds. J Pediatr Orthop 21, 395-402. Stansfield, B.W., Hillman, S.J., Hazlewood, M.E., Lawson, A.A., Mann, A.M., Loudon, I.R. and Robb, J.E., 2001b. Sagittal joint kinematics, moments, and powers are predominantly characterized by speed of progression, not age, in normal children. J Pediatr Orthop 21, 403-11. Stansfield, B.W., Hillman, S.J., Hazlewood, M.E. and Robb, J.E., 2006. Regression analysis of gait parameters with speed in normal children walking at self-selected speeds. Gait Posture 23, 288-94. Stewart, C., Postans, N., Schwartz, M.H., Rozumalski, A. and Roberts, A., 2007. An exploration of the function of the triceps surae during normal gait using functional electrical stimulation. Gait Posture 26, 482-8.

124
Stoquart, G.G., Detrembleur, C., Palumbo, S., Deltombe, T. and Lejeune, T.M., 2008. Effect of botulinum toxin injection in the rectus femoris on stiff-knee gait in people with stroke: a prospective observational study. Arch Phys Med Rehabil 89, 56-61. Studenski, S., Perera, S., Patel, K., Rosano, C., Faulkner, K., Inzitari, M., Brach, J., Chandler, J., Cawthon, P., Connor, E.B.et al., 2011. Gait speed and survival in older adults. JAMA 305, 50-8. Sung, D.H. and Bang, H.J., 2000. Motor branch block of the rectus femoris: its effectiveness in stiff-legged gait in spastic paresis. Arch Phys Med Rehabil 81, 910-5. Sutherland, D.H. and Davids, J.R., 1993. Common gait abnormalities of the knee in cerebral palsy. Clin Orthop Relat Res 139-47. Sutherland, D.H., Larsen, L.J. and Mann, R., 1975. Rectus femoris release in selected patients with cerebral palsy: a preliminary report. Dev Med Child Neurol 17, 26-34. Sutherland, D.H., Santi, M. and Abel, M.F., 1990. Treatment of stiff-knee gait in cerebral palsy: a comparison by gait analysis of distal rectus femoris transfer versus proximal rectus release. J Pediatr Orthop 10, 433-41. Tammik, K., Matlep, M., Ereline, J., Gapeyeva, H. and Paasuke, M., 2008. Quadriceps femoris muscle voluntary force and relaxation capacity in children with spastic diplegic cerebral palsy. Pediatr Exerc Sci 20, 18-28. Thelen, D.G. and Anderson, F.C., 2006. Using computed muscle control to generate forward dynamic simulations of human walking from experimental data. J Biomech 39, 1107-15. Thelen, D.G., Anderson, F.C. and Delp, S.L., 2003. Generating dynamic simulations of movement using computed muscle control. J Biomech 36, 321-8. Thometz, J., Simon, S. and Rosenthal, R., 1989. The effect on gait of lengthening of the medial hamstrings in cerebral palsy. J Bone Joint Surg Am 71, 345-53. Treanor, W.J., 1969. The role of physical medicine treatment in stroke rehabilitation. Clin Orthop Relat Res 63, 14-22. Tuzson, A.E., Granata, K.P. and Abel, M.F., 2003. Spastic velocity threshold constrains functional performance in cerebral palsy. Arch Phys Med Rehabil 84, 1363-8. van der Krogt, M.M., Bregman, D.J., Wisse, M., Doorenbosch, C.A., Harlaar, J. and Collins, S.H., 2010. How crouch gait can dynamically induce stiff-knee gait. Ann Biomed Eng 38, 1593-606. van der Krogt, M.M., Doorenbosch, C.A. and Harlaar, J., 2007. Muscle length and lengthening velocity in voluntary crouch gait. Gait Posture 26, 532-8.

125
van der Linden, M.L., Aitchison, A.M., Hazlewood, M.E., Hillman, S.J. and Robb, J.E., 2003. Effects of surgical lengthening of the hamstrings without a concomitant distal rectus femoris transfer in ambulant patients with cerebral palsy. J Pediatr Orthop 23, 308-13. van der Linden, M.L., Kerr, A.M., Hazlewood, M.E., Hillman, S.J. and Robb, J.E., 2002. Kinematic and kinetic gait characteristics of normal children walking at a range of clinically relevant speeds. J Pediatr Orthop 22, 800-6. Ward, S.R., Terk, M.R. and Powers, C.M., 2005. Influence of patella alta on knee extensor mechanics. J Biomech 38, 2415-22. Waters, R.L., Garland, D.E., Perry, J., Habig, T. and Slabaugh, P., 1979. Stiff-legged gait in hemiplegia: surgical correction. J Bone Joint Surg Am 61, 927-33. Wren, T.A., Rethlefsen, S. and Kay, R.M., 2005a. Prevalence of specific gait abnormalities in children with cerebral palsy: influence of cerebral palsy subtype, age, and previous surgery. J Pediatr Orthop 25, 79-83. Wren, T.A., Rethlefsen, S.A., Healy, B.S., Do, K.P., Dennis, S.W. and Kay, R.M., 2005b. Reliability and validity of visual assessments of gait using a modified physician rating scale for crouch and foot contact. J Pediatr Orthop 25, 646-50. Yamaguchi, G.T. and Zajac, F.E., 1990. Restoring unassisted natural gait to paraplegics via functional neuromuscular stimulation: a computer simulation study. IEEE Trans Biomed Eng 37, 886-902. Yngve, D.A., Scarborough, N., Goode, B. and Haynes, R., 2002. Rectus and hamstring surgery in cerebral palsy: a gait analysis study of results by functional ambulation level. J Pediatr Orthop 22, 672-6. Zajac, F.E. and Gordon, M.E., 1989. Determining muscle's force and action in multi-articular movement. Exerc Sport Sci Rev 17, 187-230. Zmitrewicz, R.J., Sachin, R.R., Kotaro, S., 2007. Mechanical energetic contributions from individual muscles and elastic prosthetic feet during symmetric unilateral transtibial amputee walking: a theoretical study. Journal of Biomechanics 40, 1824-1831.