ultrastructure of the blood-brain barrier in the dolphin (stenella coeruleoalba)
TRANSCRIPT

Brain Research, 414 (1987) 205-218 205 Elsevier
BRE 12651
Research Reports
Ultrastructure of the blood-brain barrier in the dolphin (Stenella coeruleoalba)
Ilya I. Glezer 1'2, Myron S. Jacobs 2'3 and Peter J. Morgane 2'4 1The City University of New York Medical School, New York, NY IO031 (U.S.A.), 20sborn Laboratories of Marine Sciences, New
York Aquarium, Brooklyn, NYl1224 (U.S.A.), 3New York University College of Dentistry, New York, NY IO010 (U.S.A.) and 4Worcester Foundation for Experimental Biology, Shrewsbury, MA 01545 (U.S.A.)
(Accepted 11 November 1986)
Key words: Dolphin brain; Blood-brain barrier; Glia; Tight junction; Gap junction; Brain capillary; Angioarchitectonics; Glioarchitectonics; Glio-glial junction; Astroglia-like cell
Light and electron microscopic methods were used for investigation of angioarchitectonics, glioarchitectonics and the structural ba- sis of the blood-brain barrier in the dolphin Stenella coeruleoalba. It was shown that the cortical plate of the dolphin brain is extremely rich in capillaries and small arteries that are organized into a complicated net of continuous loops surrounding neuronal groups. The density of the capillary loops is related to the cytoarchitectural density of the cortex. It was also found that the neuronal microenviron- ment in the dolphin cortex is characterized by the presence of a large number of the astroglia-like cells that make a multi-layered in- vestment surrounding capillaries and small arteries. These glial cells, unlike typical astrocytes of terrestrial mammals, have a large number of different organelles and their nuclei are similar to those of the oligocytes. The ultrastructure of the blood-brain barrier in the dolphin is characterized by the presence of extremely long tight junctions between endothelial cells and by specialized junctions between pericapillary astroglia-like cells. A belt of the glial end-feet interlocked with different types of junctions such as zonulae ad- herentes, maculae adherentes and gap junctions was found around all investigated capillaries. This system of specialized interen- dothelial and glio-glial junctions is tentatively hypothesized to be a feature of adaptation of the dolphin to the aquatic environment.
INTRODUCTION
The p rob lem of the neuronal microenvi ronment ,
i.e. neuro-gl io-vascular relat ions, has par t icular im-
por tance for compara t ive neuroana tomy and phys-
iology. A special morphophysio logica l regulat ing
mechanism defined as the b l o o d - b r a i n bar r ie r exists in all ver tebra tes 1'2'4'5, though it may have d i f ferent
features in various classes of animals 6-14. Cetacea,
the small and great whales, occupy a unique niche
among all mammal ian groups that is shared, in terms
of being total ly commit ted to the aquat ic environ-
ment , only with Sirenia. The cetaceans are air-
breathing, diving mammals and spend their ent ire
life-cycle in the aquatic environment .
One of the main acquisit ions of cetaceans along the
pathways of aquatic adapta t ion and specialization
was the deve lopment of a highly specialized central
nervous system that combines evolut ional ly progres- sive and conservative features 2°'21,25'29. Our recent
electron microscopic and Golgi findings in the dol-
phin convexity cortex have revealed an unusual com-
binat ion of both conservative and highly specialized
features 21. Thus, we demons t ra t ed an ext reme rich-
ness of dendri t ic branching in the dolphin neocor tex
accompanied by the presence of large numbers of
synaptic vesicles. Also, we found that capil lary en-
dothelial cells are jo ined by ex t remely long tight
junctions and that there are different types of junc- tional complexes compris ing the glio-vascular ana-
tomical relations. The present ul t rastructural obser-
vations suggest a highly deve loped gl io-endothel ia l
Correspondence: I.I. Glezer, The City University of New York Medical School, The Science Building, Rm. J-914, 138th St., at Con- vent Ave., New York, NY 10031, U.S.A.
0006-8993/87/$03.50 I~) 1987 Elsevier Science Publishers B.V. (Biomedical Division)

206
interaction in addition to the usual interendothelial connections seen in most vertebrates. There are no
previous reports on the structure of the cetacean blood-brain barrier or on their glio-vascular rela-
tions in the literature. It is of particular interest to assess how these mor-
phological peculiarities of the dolphin brain are a re-
sult of adaptation and specialization to the aquatic environment 33. In previous studies, we found that Ce-
tacea have specially organized features of peripheral sources of the arterial blood supply to the brain 19'28'35. In Tursiops truncatus 28'35 as well as in
Monodon monoceros and Delphinapterus leucas 36'37, the main source of arterial blood supply to the brain is via a thoracic rete mirabile fed from the aorta via the intercostal arteries and extending into the cranial cavity from which the cerebral arterial blood supply
is derived. In earlier studies we found that Odontoce-
ti (dolphins and other toothed whales) have no sys-
tems of internal carotid and vertebral arteries which
characterize the arterial blood supply to the brain in terrestrial mammals 2s. It might be expected that such
a radical change in the arterial blood supply to the brain in the course of cetacean evolution would be ac- companied by changes in microvasculature as well as
in the structure and function of the blood-brain bar- rier. The present study was designed to investigate
this issue. We have focused our investigations on the angio-
architecture and main structural elements of the
blood-brain barrier in the neocortex of the dolphin in an attempt to answer two specific questions, name-
ly: (1) what is the architectonic relationship between the blood vessels, neurons and glial cells in the neo- cortex of the dolphin; and (2) what are the special structural features of the blood-brain barrier (neu- ro-glio-vascular relations) in the dolphin and how do
they differ from those in terrestrial mammals.
MATERIALS AND METHODS
The investigation was carried out on the brain of a young spotted dolphin, Stenella coeruleoalba, esti- mated to be approximately 3 months old. Cortical blocks were taken from the lateral gyrus and were processed for rapid Golgi and EM studies. This gyrus has been shown in electrophysiological studies 26'34 to be the site of visual representation in the dolphin
brain.
For rapid Golgi impregnation, cortical samples
were chromated (2.5% potassium dichromate) for 4
days, transferred to a chrom-osmium mixture (1:1) for an additional 4 days, and then were treated with
0.75% silver nitrate for one week. These steps were
carried out in the dark at a thermostatically main- tained temperature of 27 °C. After alcohol dehydra-
tion and embedding in celloidin, the cortical samples were sectioned at 100-150/~m, mounted on glass
slides and used for analysis of angioarchitectonics
and glioarchitectonics. For transmission electron microscopic (EM) stud-
ies, the brain samples were prefixed in a cold 2% glu- taraldehyde-4% paraformaldehyde mixture in caco- dylate buffer (pH 7.5, 0.1 M) for one hour and then
transferred to 1% OsO4 in cacodylate buffer (pH 7.5) for another hour. The osmicated samples were then
dehydrated in alcohols and embedded in a mixture of Epon and Araldite. After polymerization, blocks
were cut with a diamond knife on an LKB-3 ultra-
tome, mounted on copper grids, double-stained with uranylacetate and lead citrate 24'3° and examined
using a Philips-301 transmission electron microscope.
The region of the brain in which the angioarchitec-
tonic, glioarchitectonic and ultrastructural organiza- tion of the blood-brain barrier was studied was lim-
ited to the mid-anteroposterior level of the lateral gy- rus, including cortical sites in the depths and both
banks of the enterolateral suicus. A computerized Image Analysis SMI-MICRO-
COMP System was used for measurement of the den-
sity of the capillaries in one cubic millimeter of the ce- rebral cortex of the dolphin and for estimation of the
diffusion path length. Both parameters were mea- sured with the help of the PM (Planimetry) program. The Golgi and Nissl sections were projected with the help of the Videcon tube on computer monitors (IBM XT) and measurements were made with the cursor and digitizing tablet. The program then auto- matically computed and statistically evaluated the produced measurements. The following parameters were measured: integral length of the capillary net as defined on the screen within the area of the section, diameters of the capillary loops, and diameters of the cell perikarya. Cells and capillaries were measured in randomly chosen areas of different cortical layers. In each layer 10 areas were measured in 5 sequential

sections. Based on diameters of the capillary loops and
diameters of the perikarya, a diffusion pathway dis- tance was calculated based on the formula:
[(X + Y)-(x + y)]/4,
where X -- large diameter of the capillary loop; Y = small diameter of the capillary loop; x = large diame- ter of the perikaryon; y -- small diameter of the peri-
karyon. This formula is based on the assumption that
each cell or cell group is located in the geometrical center of the loop, a fact that was established by our
observations in Golgi sections.
RESULTS
Angioarchitectonics
Angioarchitectonic analysis of the lateral gyrus of
the dolphin reveals many interesting features. The general relationships between arteries, arterioles
I III III III
I' V V
207
• and capillaries at the light microscopic level are simi-
lar to those in terrestrial mammals. The radial arte-
ries and veins originate from the pial vasculature and
pierce the layers of the cortex perpendicular to the pial surface (Fig. 1). The diameter of these radial ar-
teries is about 10-11/~m. At the level of the cortical plate itself, these small arteries form a dense mesh-
like net of capillaries, having diameters ranging from
2.9 to 3.4/~m. These capillary nets connect neighbor- ing radial arteries and veins. The neurons and glial
cells are located within the loops of the capillary net
(Fig. 2). The 3-dimensional net of capillaries exhibits
variations that appear to depend on the distribution of neurons in the various cortical layers. Thus, in lay-
er II where the neurons are relatively small and ex- tremely densely packed, the capillary net consists of
narrow and tightly packed loops (Fig. 2A). On the other hand, in layer III , as well as in layer V, the loops are less tightly packed, more rounded, and larg-
er in diameter (Fig. 2B). The mean diameter of the
loops in these layers is about 24-30/~m. At the level
Fig. 1. Angioarchitectonics of the lateral gyrus (visual cortex) in the dolphin Stenella coeruleoalba. Note the radial vessels (Rar) and network of capillaries (Cap) connecting them. This figure also shows the distribution of the veins, arteries and capillaries in the differ- ent architectonic layers. Note different orientation of the capillary network in upper and lower layers of the cortical plate. At the level of the layer II the capillaries are arranged in clusters (C), whereas in the layers III and V capillaries have no special orientation of their loops. At the level of layer VI both large vessels and capillary loops have a tangential direction, parallel to the pia. Rapid Golgi im- pregnation.
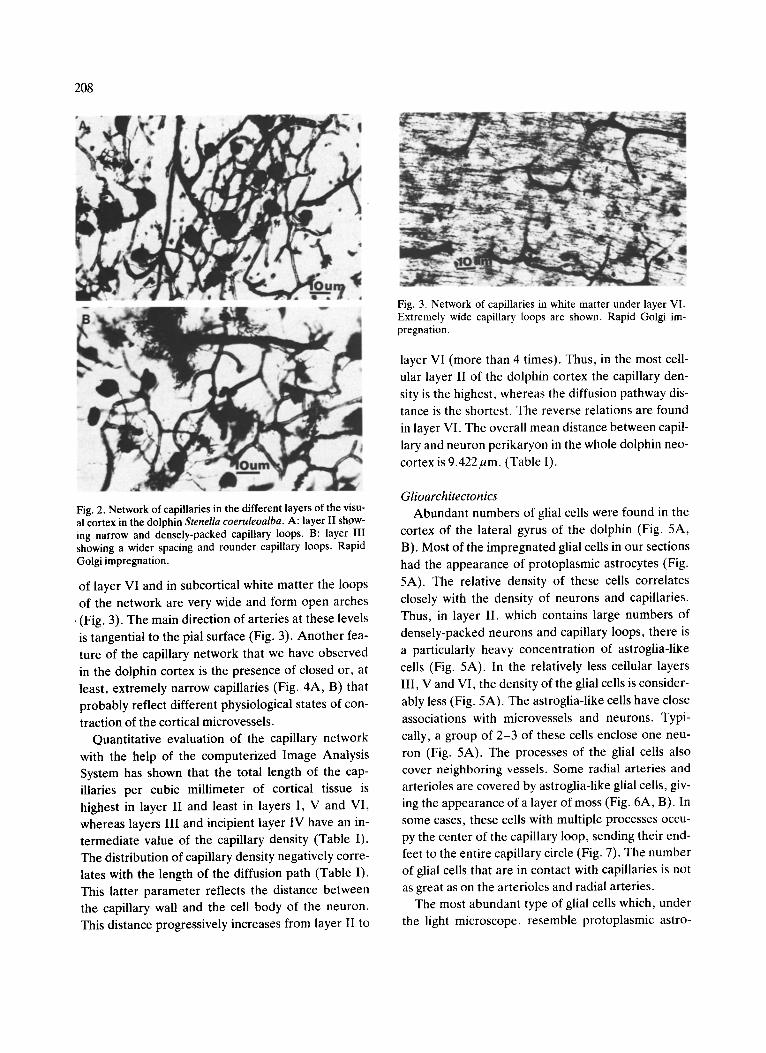
208
Fig. 2. Network of capillaries in the different layers of the visu- al cortex in the dolphin Stenella coeruleoalba. A: layer II show- ing narrow and densely-packed capillary loops. B: layer III showing a wider spacing and rounder capillary loops. Rapid Golgi impregnation.
of layer VI and in subcortical white matter the loops of the network are very wide and form open arches
• (Fig. 3). The main direction of arteries at these levels
is tangential to the pial surface (Fig. 3). Another fea- ture of the capillary network that we have observed in the dolphin cortex is the presence of closed or, at least, extremely narrow capillaries (Fig. 4A, B) that probably reflect different physiological states of con-
traction of the cortical microvessels. Quantitative evaluation of the capillary network
with the help of the computerized Image Analysis System has shown that the total length of the cap- illaries per cubic millimeter of cortical tissue is highest in layer II and least in layers I, V and VI, whereas layers III and incipient layer IV have an in- termediate value of the capillary density (Table I). The distribution of capillary density negatively corre- lates with the length of the diffusion path (Table I). This latter parameter reflects the distance between the capillary wall and the cell body of the neuron. This distance progressively increases from layer 11 to
Fig. 3. Network of capillaries in white matter under layer VI. Extremely wide capillary loops are shown. Rapid Golgi im- pregnation.
layer VI (more than 4 times). Thus, in the most cell- ular layer II of the dolphin cortex the capillary den- sity is the highest, whereas the diffusion pathway dis- tance is the shortest. The reverse relations are found in layer VI. The overall mean distance between capil- lary and neuron perikaryon in the whole dolphin neo- cortex is 9.422/zm. (Table 1).
Glioarchitectonics
Abundant numbers of glial cells were found in the cortex of the lateral gyrus of the dolphin (Fig. 5A, B). Most of the impregnated glial cells in our sections had the appearance of protoplasmic astrocytes (Fig. 5A). The relative density of these cells correlates closely with the density of neurons and capillaries. Thus, in layer II, which contains large numbers of densely-packed neurons and capillary loops, there is a particularly heavy concentration of astroglia-like cells (Fig. 5A). In the relatively less cellular layers III, V and VI, the density of the glial ceils is consider- ably less (Fig. 5A). The astroglia-like cells have close
associations with microvessels and neurons. Typi- cally, a group of 2-3 of these cells enclose one neu- ron (Fig. 5A). The processes of the glial cells also cover neighboring vessels• Some radial arteries and arterioles are covered by astroglia-like glial cells, giv- ing the appearance of a layer of moss (Fig. 6A, B). In some cases, these cells with multiple processes occu- py the center of the capillary loop, sending their end- feet to the entire capillary circle (Fig. 7). The number of gliai cells that are in contact with capillaries is not as great as on the arterioles and radial arteries•
The most abundant type of glial cells which, under the light microscope, resemble protoplasmic astro-

Fig. 4. Different functional states of capillaries in dolphin visual cortex. A: a capillary loop with entirely open lumen. B: a capil- lary loop with partially closed lumen (arrow head). Fragment of astrocyte (As) is also shown. Rapid Golgi impregnation.
cytes, are found in the electron microscope to be of
an intermediate type combining features of both as-
trocytes and oligocytes (Fig. 8). These astrocyte/oli-
gocyte cells contain numerous intermediate fila-
ments, a feature restricted to fibrous astrocytes, but
their nuclei and abundan t endoplasmic reticulum are
209
similar to features of oligocytes in terrestrial mam-
mals.
Ultrastructure o f the b l o o d - b r a i n barrier
Features of the cerebral vasculature in the dolphin
brain as in most other mammal ian species include ex-
tensive specialization of the morphological elements
comprising the b l ood - b r a i n barrier. Thus, as in ter-
restrial mammals, in most microvessels along the
basal membrane which covers the outer surface of
the endothelial cells, there is a thick uninter rupted
layer of cytoplasm of the glial end-feet. In arterioles
and small arteries, there are even several layers of
glial end-feet. As a rule, these glial end-feet contain a
network of intermediate filaments, large oval mito-
chondria and a small amount of the endoplasmic reti-
culum and polyribosomes (Fig. 9A).
Our material reveals that the capillary endotheli-
um (Fig. 9A, B) contains cells rich in different cyto-
plasmic organelles such as endoplasmic reticulum,
mitochondria and different types of vesicular pro-
files. In some capillaries there is also an abundance of
vesicles or tangentially-cut channels of smooth endo-
plasmic reticulum (Fig. 9B). As a rule, two or three
endothelial cells constitute the wall of each capillary.
The nuclei of the endothelial cells have the appear-
ance of a typical non-neuronal nucleus with a chro-
matin belt along the inner nuclear membrane (Fig.
9A). The basal membrane in most of the cortical ves-
sels is represented by a layer of fibrillar substance
that is located between the outer endothelial mem-
brane and the glial end-foot (Fig. 9A). The basal
membrane often encloses pericytes and insinuates it-
TABLE I
Length of the capillaries per mm 3 of the cortical tissue and diffusion pathway length in dolphin and some terrestrial mammals
Layers Length of capillaries in mm per mm s Diffusion pathway length in l~m
Dolphin Cat* Rat* Dolphin Man* Rat* Horse*
I 1181 ± 169 845 ± 51 942 II 1749 ± 163 921 ± 52 -
III 1231 ± 103 1031 ± 57 1086 IV 1278 ± 244 1076 ± 62 1208 V 1063 ± 125 881 ± 50 1029
VI 792 ± 69 863 ± 50 789
Mean 1215 ± 117 936 ± 54 1011
4.046 + 0.71 - - - 5.048 + 0.61 - - - 7.055 + 0.99 - - -
11.560 + 0.99 - - - 17.030 + 2.09 - - -
9.422 + 1.11 30.00 25.00 37.00
* +Data from 'The Human Brain in Tables and Figures' by Blinkov and Glezer 3.
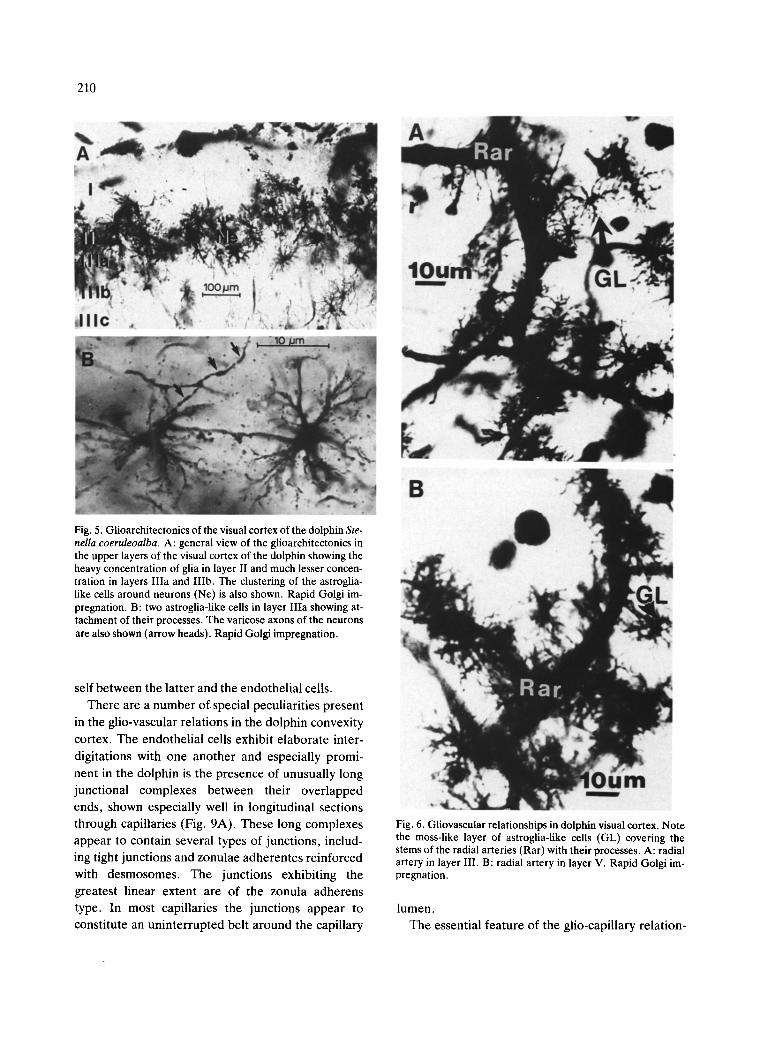
210
Fig. 5. Giioarchitectonics of the visual cortex of the dolphin Ste- nella coeruleoalba. A: general view of the glioarchitectonics in the upper layers of the visual cortex of the dolphin showing the heavy concentration of glia in layer II and much lesser concen- tration in layers Ilia and IIIb. The clustering of the astroglia- like cells around neurons (Ne) is also shown. Rapid Golgi im- pregnation. B: two astroglia-like cells in layer IIIa showing at- tachment of their processes. The varicose axons of the neurons are also shown (arrow heads). Rapid Golgi impregnation.
self be tween the la t ter and the endothel ia l cells.
There are a number of special peculiari t ies present
in the glio-vascular relat ions in the dolphin convexity
cortex. The endothel ia l cells exhibit e labora te inter-
digitations with one another and especially promi-
nent in the dolphin is the presence of unusually long
junct ional complexes between their over lapped
ends, shown especially well in longitudinal sections
through capillaries (Fig. 9A). These long complexes
appear to contain several types of junctions, includ-
ing tight junctions and zonulae adherentes reinforced
with desmosomes. The junct ions exhibit ing the
greatest l inear extent are of the zonula adherens
type. In most capil laries the junct ions appear to
constitute an unin ter rupted belt a round the capil lary
Fig. 6. Gliovascular relationships in dolphin visual cortex. Note the moss-like layer of astroglia-like cells (GL) covering the stems of the radial arteries (Rar) with their processes. A: radial artery in layer III. B: radial artery in layer V. Rapid Golgi im- pregnation.
lumen.
The essential feature of the glio-capil lary re la t ion-
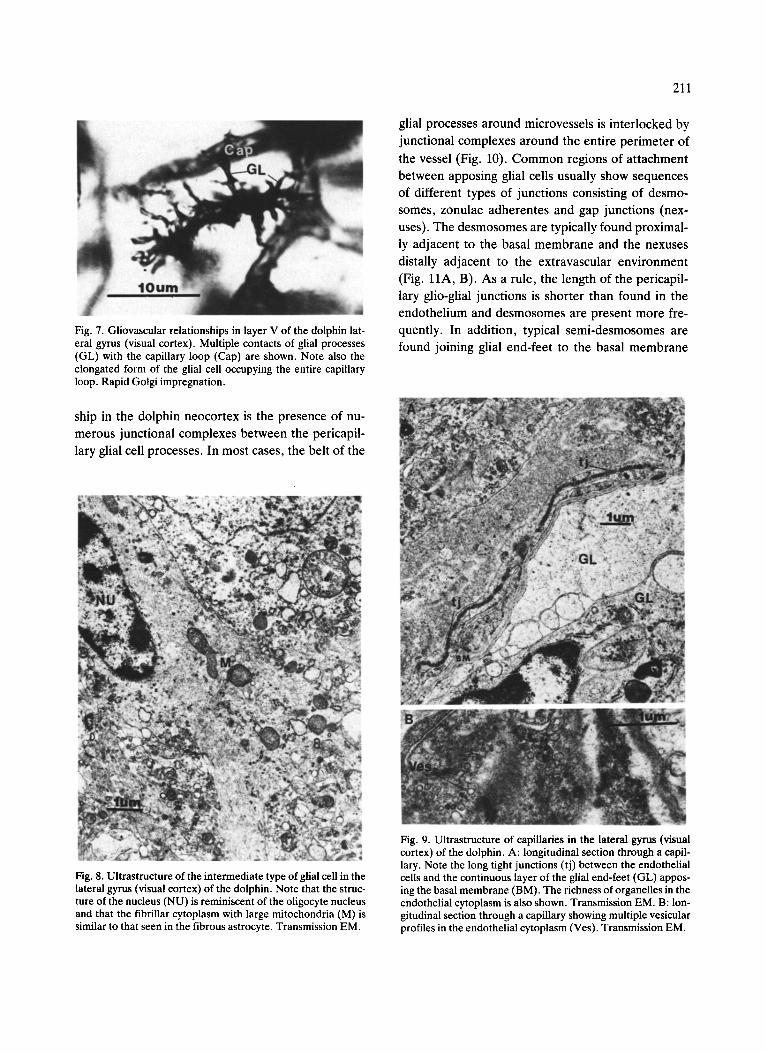
211
Fig. 7. Gliovascular relationships in layer V of the dolphin lat- eral gyrus (visual cortex). Multiple contacts of glial processes (GL) with the capillary loop (Cap) are shown. Note also the elongated form of the glial cell occupying the entire capillary loop. Rapid Golgi impregnation.
glial processes around microvessels is interlocked by
junctional complexes around the entire perimeter of
the vessel (Fig. 10). Common regions of at tachment
between apposing glial cells usually show sequences of different types of junctions consisting of desmo-
somes, zonulae adherentes and gap junctions (nex-
uses). The desmosomes are typically found proximal- ly adjacent to the basal membrane and the nexuses
distally adjacent to the extravascular environment
(Fig. 11A, B). As a rule, the length of the pericapil-
lary glio-glial junctions is shorter than found in the
endothelium and desmosomes are present more fre- quently. In addition, typical semi-desmosomes are
found joining glial end-feet to the basal membrane
ship in the dolphin neocortex is the presence of nu-
merous junctional complexes between the pericapil- lary glial cell processes. In most cases, the belt of the
Fig. 8. Ultrastructure of the intermediate type of glial cell in the lateral gyrus (visual cortex) of the dolphin. Note that the struc- ture of the nucleus (NU) is reminiscent of the oligocyte nucleus and that the fibrillar cytoplasm with large mitochondria (M) is similar to that seen in the fibrous astrocyte. Transmission EM.
Fig. 9. Ultrastructure of capillaries in the lateral gyrus (visual cortex) of the dolphin. A: longitudinal section through a capil- lary. Note the long tight junctions (tj) between the endothelial cells and the continuous layer of the glial end-feet (GL) appos- ing the basal membrane (BM). The richness of organelles in the endothelial cytoplasm is also shown. Transmission EM. B: lon- gitudinal section through a capillary showing multiple vesicular profiles in the endothelial cytoplasm (Ves). Transmission EM.

212
(Figs. 12A, B; 13A, B). Conspicuous long-gap junc- tions between apposed pericapillary and periarterial cells are also found around some vessels (Fig. l l A , B). In some cases, we found small areas of apposing glio-glial contacts that are reminiscent of tight junc- tions (Fig. 12A, B).
DISCUSSION
The present study reveals several special features of the neuronal microenvironment in the lateral gy- rus (visual cortex) of the dolphin. Qualitative and quantitative analysis shows several important pecu- liarities of cortical angioarchitectonics in the dolphin. Thus, the presence of the densely packed loops of capillary networks in the dolphin cortex is closely correlated with the packing density of cortical neu- rons in the different layers. Each capillary loop encir- cles a small group of neurons, thus providing them
Fig. 10. Cross-section through a capillary in the lateral gyrus (visual cortex) of the dolphin brain. Endothelial and glio-glial junctional complexes (t j) are shown. Note also that a belt of the glial processes (GL) surrounds and encloses an entire perime- ter of the capillary. Transmission EM.
with metabolites and maintaining the partial pressure of oxygen at a certain level which is critical in the aquatic environment. In terrestrial mammals, the ra- dius of the cylinder of brain substance supplied by one capillary has been shown to depend on the size of the animal 23. Thus, in man it is about 30 ~tm, in a
horse about 37/~m, whereas in small mammals such as rats and mice it is about 25 ,um. Our data shows that the distance between the capillary walls and the cell body of the neurons which are located in the cen- ter of the capillary loop is not more than 9-10/~m. It is interesting that, although the packing of capillaries in the dolphin cortex is higher than in terrestrial mammals, capillary diameters are in the same range of variation (3-4t im). Comparing the densities of the capillaries per one cubic millimeter of the cortical volume in cetaceans and some terrestrial mammals, we found several important differences between these two ecologically different mammalian groups. Thus, mean density of capillaries is moderately higher in almost all cortical layers of the dolphin brain than in terrestrial mammals (see Table I). However, the most spectacular differences between these two groups are in distribution of the densities of capillaries along the vertical axis of the cortex. As it was expected, the highest density of microvessels co- incides with the highest neuronal concentration. Thus, in dolphins the highest capillary concentration is found in the accentuated layer II, whereas in pri- mates (homo), carnivora (felis, canis) and rodentia (rat) it is found at the levels of layers III and IV 3't7. We can tentatively hypothesize that the difference in cortical vasculature between terrestrial and aquatic mammals also depends on differences in afferenta- tion. In cetaceans the main afferent flow to the cortex is provided by the extremely thick layer I and accen- tuated layer II. This type of afferentation is the fea- ture of the conservative ('initial') type of cortical af- ferentation preserved in contemporary cetaceans and in some insectivores 2°'29. In most of the advanced
terrestrial mammals the main afferent flow in neo- cortex is assigned to layer IV and, to some degree, layers III and V. Evidently, these differences in func- tional activities of the cortical layers are reflected in distribution of the capillaries in the cortical layers.
The density of the capillaries is compatible with the diffusion blood pathway length. Thus, in layer II, as can be expected, the path is the shortest and in layer

213
Fig. 11. Cross-section through a capillary in the lateral gyrus (visual cortex) of the dolphin brain showing endothelial and glio-glial junctional complexes. Note the presence of the topographical sequence of glial junctions: desmosome (Des) is proximal to the basal membrane (BM) and gap junction (GP) is distal to it. A: general view of the vessel and surrounding perivascular glia. B: higher magni- fication of the area in A indicated by a rectangle. A gap junction and desmosome are shown. Transmission EM.

214
Fig. 12. Ultrastructure of the glio-glial junctional complexes in the lateral gyrus (visual cortex) of the dolphin brain. A: a general view of a capillary and glio-glial junctional complexes. Box indicates the area magnified in B. Transmission EM. B: multiple zonulae adher- entes (ZA) and maculae adherentes (Des) between apposed plasma membranes of the pericapillary glial cells. Also semidesmosomes (SM) are present on glial plasma membrane facing basal membrane (BM). Note the extreme closeness of the apposed membranes in certain locations reminiscent of tight junctions and indicated by heavy arrows. Light lines show limits of zonulae adherents (ZA). Transmission EM.

215
Fig. 13. Longitudinal section through a capillary in the lateral gyrus (visual cortex) of the dolphin brain. A: figure showing the pres- ence of morphologically similar junctions (j) in endothelium and in pericapillary glia. B: higher magnification of the area in A indi- cated by a rectangle. Note granules (GR) covering the apposed plasma membranes of the glial end-feet and multiple semi-desmo- somes (SM) facing the basal membrane (BM) and having typical tonofibrils. Transmission EM.

216
VI it is the longest. It is of special interest that the dif- fusion pathway length in the dolphin is increased more than 4 times from layer II to layer VI. It is possi- ble that in evolution the special hydrodynamics of the blood supply to the cortex has an important influence on the formation of the very thin and extremely ex- panded cetacean neocortex.
The glio-architectonic analysis has also revealed some special features of the dolphin brain. First of all, we found a great abundance of astroglia-like cells enclosing both neuronal perikarya and neigh- boring vessels. These glial cells are of intermediate character and have features common to both astro- cytes and oligocytes. The presence of intermediate type glial cells in the dolphin may be interpreted to be an indication of the ancestral radiation of cetaceans from some 'initial' group of mammals that retained non-differentiated macroglial elements. To deter- mine the actual character of these morphologically intermediate cells, it would be extremely important to apply the immunofluorescence reaction to glial fi- brillary acidic protein (GFAP) which is specific for astrocyte intermediate fibrils and highly stable in dif- ferent mammalian species 15A6.
The presence of an unusual abundance of these glial cells surrounding neurons and vessels can be in- terpreted to mean that in the dolphin brain regulation of the ionic metabolism in cortical neurons, a univer- sal feature in all vertebrates 1'2'7'11-14, has assumed
special importance. It is possible that this abundance of glial cells helps to remove accumulated K + ions from the interstitium of active neurons and thus re- duces the decrease of Na + and Ca 2+ concentrations, enhancing synaptic neuronal activity 2'1s'27. We ini-
tially interpret this abundance of glia as a special fea- ture of the structure of the blood-brain barrier in dolphins.
It is well-established that the main physiological and structural mechanism of the blood-brain barrier in most vertebrates is confined to the endothelial cells interlocked with tight junctions 4-8'1°-12'31. Only in some elasmobranchs, as well as in some inverte- brates, has there been found morphological and physiological evidence of the glial blood-brain bar- rier with the endothelial barrier not being present 6'1°. The endothelium and perivascular glial cells present between the circulating blood and neuronal elements in all mammals so far studied appear to be organized
differently in Cetacea. Thus, the blood-brain inter- face in the dolphin comprises both endothelial and glial cells which are independently united by varying sequences of junctional complexes so as to form two distinct concentric rings separating the blood from the brain parenchyma. In the case of the dolphin neo- cortex we found that not only is the endothelial bar- rier represented by hyperdeveloped, long junctional complexes but also by a belt of glial end-feet inter- locked by junctions. These long endothelial junctions are similar to those found in certain fish (Chimaera monstrosa) II. The blood-brain barrier of the dolphin evidently shows a specialized structure that might be interpreted as a specific adaptation to the aquatic en- vironment. The previously mentioned abundance of glial cells around arteries and arterioles is well-corre- lated with the electron microscopic data on the unin- terrupted and sometimes multi-layered belt of glio- plasm around capillaries and with the presence of junctional complexes between glial processes. In most studies of the blood-brain barrier, it has been shown that a glial belt around brain capillaries is not part of the blood-brain barrier because of the ab- sence of tight junctions between apposed glial pro- cesses and because the injected tracers such as HRP, ferritin, etc., easily penetrate through interglial spaces 6'7'1s. However, some authors have described the presence of gap junctions between pericapillary astrocytes in terrestrial mammals s'~5'31. These gap junctions appear to be very resistant to hypoxia and are an important mechanism for molecular exchange and electrical coupling 7'22'27. The sequence of the
pericapillary glio-glial junctions which we have described in the dolphin, including desmosomes, zo- nula adherens, and gap junctions in the same region of apposed membranes, has not heretofore been described in other mammals. The gap junctions be- tween pericapillary and periarterial glial cells are ex- tremely well-developed and often have very great length. Thus, in these aquatic mammals the glial end- feet are attached to each other by sequences of ad- hering junctions (fascia adherens, desmosomes) as well as by extremely penetrable junctions (nexuses).
At this stage of our studies of the blood-brain bar- tier in dolphins, we may speculate that glio-glial in- teractions play an important regulatory role in the function of the blood-brain barrier in these animals. Based on the presence of the different kinds of junc-

217
tions between pericapillary glial processes, one might
assume that in the cetacean brain the permeability of
the b lood-bra in barrier is regulated by two contigu-
ous layers (endothelial and glial). It would be ex- pected that these layers would certainly slow down
the exchange of macromolecules between blood and
brain. For support or refutation of this hypothesis, the data on chemical composition of the CSF and, es-
pecially, on concentration of proteins in cetacean ce-
rebrospinal f luidwould be of great importance.
In our future studies we would like to determine whether, among the multiple types of glio-glial junc-
tions we have seen in the dolphin brain, there are also
present typical tight junctions besides the fasciae ad-
herentes and desmosomes. As of now, we tend to as-
cribe the presence of extremely well-developed en- dothelial junctions as well as glio-glial attachments in
cetaceans to the need for a large brain reservoir of oxygen and for prevention of damage to the nervous tissue during deep-diving and surfacing. It is well
known that most whales have the capacity to dive for periods of minutes in the case of small whales (dol- phins) and up to an hour in the case of large whales.
In this situation, prevention of the rapid exit of oxy-
gen and recycling of the high energy metabolites in
all tissues, especially in the brain, are crucial for t h e
survival of the animal. This may be an essential role for the well-developed b lood-bra in barrier in whales. Also, it has been shown 32'37 that dolphins can
dive and surface very rapidly (2 -3 meters/s for belu- ga) 32. In such conditions without these special adap-
tations of the b lood-bra in barrier, the change in sol-
ubility of the gases would result in the caisson effect
and irreversible damage of the brain tissue by the bubbling out of nitrogen. Evidently, the presence of
an extremely resistant b lood-bra in barrier would
prevent the rapid release of gases from brain tissue
to blood and would enable whales to make rapid changes of depth, thus enhancing the chances for sur-
vival while hunting or escaping from predators.
ACKNOWLEDGEMENTS
This work was supported by National Science Foundation Grants BNS 84-14523 and BNS 85- 45732.
REFERENCES
1 Abbott, N.J., Bundgaard, M. and Cserr, H.F., Fine-struc- tural evidence for a glial blood-brain barrier in the cuttle- fish, Sepia officinalis, J. Physiol. (London), 316 (1981) 52-53.
2 Abbott, N.J., The neuronal microenvironment, TINS, 9 (1986) 3-6.
3 Blinkov, S.M. and Glezer, I.I., The Human Brain in Fig- ures and Tables, Plenum, New York, 1968, pp. 434-437.
4 Bodenheimer, T.S. and Brightman, M.W., A blood-brain barrier to peroxidase in capillaries surrounded by perivas- cular space, Am. J. Anat., 122 (1968) 249-268.
5 Brightman, M.W. and Reese, T.S., Junctions between inti- mately apposed cell membranes in the vertebrate brain, J. Cell Biol., 40 (1969) 648-677.
6 Brightman, M.W., Reese, T.S., Olsson, Y. and Klatzo, I., Morphologic aspects of the blood-brain barrier to perox- idase in elasmobranchs, Prog. Neuropathol., (1971) 146-161.
7 Brightman, M.W., Zis, K. and Anders, J., Morphology of cerebral endothelium and astrocytes as determinants of the neuronal microenvironment, Acta Neuropath. (Berlin), VIII (Suppl.) (1983) 21-33.
8 Braak, E., On the fine structure of the external glial layer in the isocortex of man, Cell Tissue Res., 157 (1975) 367-390.
9 Bundgaard, M., Transport pathways in capillaries - - in search of pores, Ann. Rev. Physiol,, 42 (1980) 325-336.
10 Bundgaard, M. and Cserr, H.F., A glial blood-brain bar- rier in elasmobranchs, Brain Research, 226 (1981) 61-73.
11 Bundgaard, M., The ultrastructure of cerebral blood capil- laries in the ratfish, Chimaera monstrosa, Cell Tissue Res., 226 (1982) 145-154.
12 Cervos-Navarro, J., Artigas, J. and Mrsulja, B.J., Morpho- functional aspects of the normal and pathological blood- brain barrier, Acta Neuropathol. (Berlin), VIII (Suppl.) (1983) 1-19.
13 Cserr, H.F., Fenstermacher, J.D. and Rail, D.P., Compar- ative aspects of brain barrier systems for non-electrolytes, Am. J. Physiol., 234 (1978) 52-60.
14 Cserr, H.F. and Bundgaard, M., Blood-brain interfaces in vertebrates: a comparative approach, Am. J. Physiol., 246 (1984) 277-288.
15 Cuevas, P., Guttieres-Diaz, J.A., Reimers, D., Dujovny, M., Diaz, F.M. and Ausman, J.I., Aspect of interastrocytic gap junctions in the experimental penumbra area, revealed by transmission electron microscopy and freeze-fracture, Experientia, 40 (1984) 471-473.
16 Dahl, D. and Bignami, A., Immunochemical and immuno- fluorescence studies of glial fibrillary acidic protein in ver- tebrates, Brain lCesearch, 61 (1973) 279-293.
17 Duvernoy, S., Delon, S. and Vannson, J,L., Cortical blood vessels of the human brain, Brain Res. Bull., 7 (1981) 519-579.
18 Fenstermacher, J.D., Current models of blood-brain transfer, TINS, 8 (1985) 449-453.
19 Galliano, "R.E., Morgane, P.J., McFarland, W.L., Nagel, E.L. and Catherman, R.L., The anatomy of cervicothorac- ic arterial system in the bottlenose dolphin Tursiops trunca- tus with a surgical approach suitable for guided angiogra-

218
phy, Anat. Rec., 155 (1966) 325-338. 20 Giezer, I.I., Jacobs, M.S. and Morgane, P.J., The So-called
'Initial' Type of the Neocortex: Relation to Cetacean Brain Organization, Abstracts Soc. for Neurosci., 15th Annual meeting, Dallas, TX, 11 Part 2, 1985, p. 1308.
21 Glezer, I.I., Jacobs, M.S. and Morgane, P.J., Ultrastructu- ral features of the neocortex on the convexity surface (lat- eral gyrus) of the dolphin Stenella coeruleoalba, J. Neuro- sci., in press.
22 Hansen, A.J., Lund-Andersen, H. and Crone, C., K+-per - meability of the blood-brain barrier, investigated by aid of a K+-sensitlve microelectrode, Acta Physiol. Scand., 101 (1977) 438-445.
23 Horstman, E., Abstand und Durchmesser der Kapillaren im Zentralnervensystem verschiedener Wilbertierklassen. In D. Tower and J. Shade (Eds.), Structure and Function of the Cerebral Cortex, Elsevier, New York, 1960, pp. 59-63.
24 Karnovsky, M.J., Simple methods for 'staining with lead' at high pH in electron microscopy, J. Biophys. Biochem. Cy- tol., 11 (1961) 729-732.
25 Kesarev, V.S., Malofeeva, L.I. and Trikova, O.V., Struc- tural organization of the cerebral neocortex in Cetaceans, Arkhiv Anat. Gistol. Embriol., 73 (1977) 23-30.
26 Ladygina, T.F., Mass, A.M. and Supin, A.Y., Multiple sensory projections in the dolphin cerebral cortex, Zh. Vyssh. Nervn. Deyat. ira. I.P. Pavlova, 28 (1978) 1047-1050.
27 Loewenstein, W.R., Junctional intercellular communica- tion: the cell-to-cell membrane channel, Physiol. Rev., 61 (1981) 830-898.
28 McFarland, W.L., Jacobs, M.S. and Morgane, P.J., Blood
supply to the brain of the dolphin Tursiops truncatus, with comparative observations on specific aspects of the cere- brovascular supply of other vertebrates, Neurosci. Biobe- hay. Rev., Suppl. I, 3 (1979) 1-93.
29 Morgane, P.J., Jacobs, M.S. and Galaburda, A.M., Con- servative features of neocortical evolution in dolphin brain, Brain Behav. Evol., 26 (1985) 176-184.
30 Pease, D.C., Histological Technique for Electron Microsco- py, Academic, New York, 1964.
31 Rapoport, S.I., Blood-Brain Barrier in Physiology and Medicine, Raven, New York, 1976.
32 Ridgway, S.H., Bowers, C.A., Miller, D., Schultz, M.L., Jacobs, C.A. and Dooley, C.A., Diving and blood oxygen in white whale, Can. J. Zool., 62 (1984) 2349-2351.
33 Severtsov, A., Morphological Principles of Evolution, Nau- ka, Moscow, 1939.
34 Sokolov, V.Y., Ladygina, T.F. and Supin, A.Y., Localiza- tion of sensory zones in dolphin brain cortex, Dokl. Akad. Nauk SSSR, 202 (1972) 490-493.
35 Viamonte, M., Morgane, P.J., Galliano, R.E., Nagel, E.L. and McFarland, W.L., Angiography in the living dolphin and observations on blood supply to the brain, Am. J. Phys- iol., 214 (1968) 1225-1249.
36 Vogl, A.W. and Fisher, H.D., Arterial retia to supply of the central nervous system in two small-toothed whales- Narwhal ( Monodon monoceros) and Beluga ( Delphinapte- rus leucas), J. Morphol., 174 (1982) 41-56.
37 Vogl, A.W. and Fisher, H.D., Arterial circulation of the spinal cord and brain in Monodontidae (order Cetacea), J. Morphol., 170 (1981) 171-180.