ultrastructural studies on normal and leukaemic human haematopoietic cells
TRANSCRIPT

Europ. 07. Cancer Vol. 5, pp. 349-360. Pergamon Press 1969. Printed in Great Britain
Ultrastructural Studies on Normal and keukaemic Human Haematopoietic Cells
ALAN ROSS and DAVID HARNDEN* Medical Research Council, Clinical and Population Cytogenetics Research Unit,
Western General Hospital, Crewe Road, Edinburgh, 4, Great Britain
INTRODUCTION THE MOST important single step in the demon- stration that a virus is involved in the aetiology of a leukaemia is the transmission of the disease by means of a cell free filtrate of material from a known case. Useful supporting evidence can, however, be obtained by the demonstration of virus particles in the leukaemic cells or other ceils of the diseased animals by electron microscopy. In man transmission experiments pose obvious difficulties and therefore numerous attempts have been made to support the idea of a virus aetiology of human leukaemia by examining material from leukaemic patients in the electron microscope. Much of the work done in this field has been on negatively stained or thin sectioned plasma pellets. The difficulty in interpreting the findings in such material is considerable and frequently authors come to quite contradictory conclusions [1, 2].
Two types of particle would be of special interest. A herpes-like virus has been found to be associated with cultured cells from lym- phomas and leukaemias in man [3] and a similar particle [4] appears in chickens with Marek's disease, one of whose features is the occurrence of lymphomas. Secondly, type-C particles occur in the murine, avian and feline leukaemias. It is now recognised that it is especially important to look for budding type-C particles. Amongst the great
Submitted for publication 19 February 1969. Accepted 6 March 1969. *Present address: Department of Cancer Studies,
Medical School, University of Birmingham, Birming- ham, Great Britain.
349
many reports of the finding of 'virus' in human leukaemic material there are only rare reports of budding [5]. Newell et al. [2] stress the need for more work on thin sectioned cellular material.
The present report deals with the examina- tion of thin sectioned material from leuk- aemic patients and controls for virus-like particles. At the same time any unusual features were noted in an effort to detect any cellular features which may be specific for a particular kind of leukaemia.
MATERIAL AND METHODS Blood and bone marrow samples
Blood samples were examined from 38 patients with leukaemia and 12 subjects with a normal haematological picture. Bone marrow samples were examined from 12 patients with leukaemia and from 12 individuals with a normal haematological picture. Plasma pellets were examined from two leukaemia patients and one haematologically normal control, while one lymph node biopsy was examined from a case of chronic lymphatic leukaemia.
The different types of leukaemia included in the study were chronic myeloid leukaernia (13 bloods, 9 bone marrows, 1 plasma pellet), acute transformation of chronic myeloid leuk- aemia (7 bloods, 2 bone marrows), acute myeloid leukaemia (5 bloods, 1 bone marrow, 1 plasma pellet), acute monocytic leukaemia (4 bloods), acute lymphatic leukaemia (5 bloods) and chronic lymphatic leukaernia (4 bloods and 1 lymph node biopsy).
Full clinical and haematological studies

350 Alan Ross and David Harnden
were made on all 38 leukaemic patients. Table 1 et al. [1] have suggested that this might gives details of the age, sex, treatment and influence the frequency with which virus-like state of health at the time the samples were particles are observed. taken. The latter is included since Levine The blood samples were all obtained by
Table 1. Details of patients included in the study
Treatment No. Age Sex Diagnosis State of health prior to sample Specimen
PB BM LN PP
1 56 F CML Well Nil + 2 65 F CML Well Busulphan + + 3 72 M CML Well (Rem) Busulphan + 4 64 M CML IU Nil + 5 40 M CML Well (Rem) Busulphan + 6 69 M CML Well Nil + + 7 40 M CML Well Nil + + 8 37 F CML Ill Nil + + 9 52 F CML Well (Rem) Busulphan +
10 44 F CML Well (Rem) Busulphan + + 11 70 M CML Ill Splen. Irr. + + 12 51 M CML Well (Rem) Busulphan + 13 65 M CML Well Nil q- 14 44 M CML Well Nil + 15 58 F CML Well Nil -b 16 50 M CML Ill Nil + 17 61 F C M L - A Well 6M-P + 18 25 M C M L - A Very ill (Rel) Busulphan + 19 43 F C M L - A Very Ill (Rel) Splen. Irr. + 20 36 F C M L - A Very Ill (Rel) Splen. Irr. +
6 M.P. Prednisolone
21 49 F CML--A Ill (Rel) Splen. Irr. q- 22 54 F C M L - A Very Ill (Rel) Busulphan -t-
Splen. Irr. 23 60 F C M L - A Ill Nil -b 24 20 M CML--A Very Ill (Rel) Busulphan q- -J- 25 71 F AML Very Ill Nil + + 26 29 M AML Ill Prednisolone + 27 62 M AML Ill Prednisolone
6 M.P. + 28 66 F AML Very Ill Nil + 29 56 M AML Ill Prednisolone + 30 53 F A. Mono L. Very ill Nil + 31 53 M A. Mono L. Ill (Rel) Cyclophosphamide +
Prednisolone 32 52 M A. Mono L. Very Ill Nil + 33 52 M A. Mono L. Ill (Rel) Prednisolone +
Cyclophosphamide 34 6 M ALL Very Ill Prednisolone ÷ 35 13 M ALL Well (Rein) Vincoblastine -t-
Prednisolone 36 2 F ALL Ill Nil ÷ 37 6 M ALL Ill Methotrexate +
6 M.P. Prednisolone
38 96 M ALL Ill Nil q- 39 66 M CLL Well Nil + 40 67 M CLL Well Nil + 41 62 F CLL Ill Nil + 42 79 M CLL Well (Rein) Chlorambucil +
i
+
+
+
C M L = chronic myeloid leukaemia; CM/.,--A= chronic myeloid leukaemia in the acute phase; A M L = a c u t e myeloid leukaemla; A. Mono. L .=acu te monocytic leukaemia; ALL=acu te lymphatic leukaemia; CLL = chronic lymphatic leukaemia; R e i n = remission; Rel = relapse; PB=peripheral blood; BM=bone marrow; L N = l y m p h node; PP=plasma pellet.

Ultrastructural Studies on Normal and Leukaemic Human Haematopoietic Cells 351
venipuncture and the bone marrow samples from the leukaemic patients by aspiration from the sternum or iliac crest. The normal bone marrow samples were obtained either by aspiration from volunteers or from ribs which had been removed during the course of thoracic surgery. Most of these latter patients had a carcinoma of the lung but the bone marrow was considered to be haematologically normal. In some of the latter cases delays of up to 30 min occurred between the removal of the rib and the marrow being put into fixative.
Preparation of peripheral blood samples The procedure used to prepare the specimens
of peripheral blood leucocytes is the method of Anderson [6] with minor modifications. Briefly, 5-20 ml heparinised venous blood is spun at 3000 rev/min for 10 rain in a siliconised centrifuge tube and plasma removed without disturbing the bully layer till only about 1/16 in. of plasma is left. Gluteraldehyde (5% in 0.1 M phosphate or 0.1 M cacodylate buffer at pH 7.2) is then added slowly so as not to disturb the buffy layer. After a minimum of I hr in fixative the disc is removed gently and thin slices not more than 1 mm thick are cut with a razor blade. These can be divided into approximately four thin broad blocks which, when embedded in araldite, will orientate themselves correctly by falling broad- face down to the bottom, so that the cutting face is at right angles to the buffy layer. The blocks are then removed, washed in cacodylate or phosphate buffered 10% sucrose, post fixed in 2% unbuffered osmium tetroxide for 1 hr, dehydrated, cleared in epoxy propane and finally embedded in araldite polymerised at 60°C for 48-72 hr.
Before thin sections are cut it is necessary to examine a thick section of the block under the light microscope, to ensure that the final t r immed block face includes the full thickness of the buffy layer. I f the buffy layer is too thick to be included in a single section, one block is tr immed to include the cells at the top of the buffy layer and another block tr immed to include ceils at the bottom of the layer. Thick sections for light microscopy are adequately stained after 5 rain in a mixture of 10% Giemsa in distilled water pre-heated to 60°C. After final trimming of the block ultra thin sections are cut using the LKB ultratome and double stained with uranyl acetate and lead citrate using the following technique. A drop of stain is deposited on to a paraffin wax coated petri dish which is flamed just before use to clean the surface. The
grid is immersed in the drop of stain and allowed to rest on the bottom of the petri dish with the sections uppermost, and the lid is then placed on the petri dish. It is found that this procedure always gives sections free of stain precipitate even when times of up to 15 min are used for either stain. The sections were examined and photographed using an AEI EM6B Electron Microscope.
Other tissues Specimens of bone marrow and lymph node
are fixed in 5% glutaraldehyde in phosphate or cacodylate buffer, washed and post-fixed in 1% osmium tetroxide and processed as above.
Ultracentrifuged plasma pellets are prepared according to the method of Porter et al. [7], fixed in 1% osmium tetroxide or Dalton's fixative, and processed as above.
Scanning procedure Up to 10 blocks were prepared from each
buffy coat, the blocks always being taken from different parts of the disc. Approxi- mately 5 thin sections (400-500A) were cut from each of five complete buffy coats, and systematically examined. Virtually all cells on each section were examined. In addition, for the bone marrow samples, areas containing several megakaryocytes were located in thick sections in the light microscope and several serial sections of these megakaryocytes were examined carefully in the electron microscope. Records were made of the types of cell present, any unusual morphological features and any particles inside or outside the ceils which could possibly have been virus particles. A special search was made of cell membranes and of platelet membranes for any evidence of budding type-C particles. While this process was very time consuming it must be remem- bered that the material examined represents only a very small fraction of the total material available. Only particles which were relatively numerous would have been detected.
RESULTS Observations on virus-like particles
We did not record in either control material or leukaemic material any structure which we considered to be a virus. We did, however, note a number of structures which bear a superficial resemblance to virus but these were equally common in leukaemic material and control material. Some can readily be identified as well known cell organelles (Fig. 1) such as nuclear pores, micropinocytotic vesicles and small granules, while others are known to

352 Alan Ross and David Harnden
be constituents of at least some types of normal cell although their precise function is unknown, e.g. perichromatin bodies which are discussed in detail below. In some cases, however, isolated particles were noted in the cytoplasm which fit neither of the above categories (Fig. 2). Such particles are very rare and do not seem to occur in any particular type of cell, either normal or leukaemic, but in every instance it was considered that the particle differed in some important respect from any known virus. In particular the location of the particle usually did not tally with the expected location of such a particle if it were virus; Fig. 2b shows an intracytoplasmic particle which resembles a herpes virus as it would appear in the nucleus. Alternatively the internal structure of the particle was not clear enough to enable one to say, in the absence of other evidence, that this was indeed virus.
One structure which can at first sight be con- fused with a vacuole containing virus particles is the compound vacuole which has a definite sub-structure composed of spherical sub-units (Fig. 3a). Similarly some polymorph granules may also have substructure of spherical units (Fig. 3b). Both entities can however easily be distinguished from known viruses.
We examined only three plasma pellets, one from an acute myeloid leukaemia, one from a chronic myeloid leukaemia and one from a control. The acute myeloid leukaemia and the control gave amorphous structureless pellets. The pellet from the chronic myeloid leukaemia case, however, contained an enormous number of membraneous structures of many shapes and sizes. Many of these were approximately
circular in section and it was found possible to select areas which contained a number of particles similar to those described by Levine et al. [1] as virus-like particles (Fig. 4). Since these were only a few amongst a great mass of material of various shapes and sizes which was obviously not virus we believe that in the absence of supporting evidence it is more reasonable to consider these to be subcellular constituents of disrupted cells.
A comparison of our material with thin sections of platelets from the peripheral blood of a kitten inoculated with material from a case of feline lymphosarcoma* was made. In this material particles were seen budding from the cytoplasmic membrane of platelets while mature type-C particles were seen extra-cellularly and in the intra-cellular vacu- oles of the platelets (Fig. lb). This comparison leaves us convinced that there were no type-C particles in the material examined by us. In particular a special search of material from 8 cases of leukaemia failed to reveal any virus particles in 500 sections taken from approxi- mately 75 megakaryocytes. This does not enable us to say that no virus was present in the peripheral blood or bone marrow of the patients whom we studied, but we can say that, if such particles were present in these tissues they were very much less numerous than in similar material from leukaemias of other species.
Observations on morphology It is not the purpose of the present paper
*We are grateful to Dr. Helen Laird, Animal Leukaemia Research Unit, Glasgow, for giving us this material.
Key to abbreviations C Centriole GO Golgi apparatus N Nucleolus F Fibril GL Glycogen NB Nuclear body F R Free ribosomes M Mitochondrion PG Perichromatin granule G Granule MV Micropinoeytotic PV Pinocytosis vacuole
vesicle V Vacuole
F i g . la.
Fig. 16.
Fig. lc.
Fig. ld.
Fig. le.
Fig. lf.
A micropinocytosis vesicle invaginating into the cytoplasm of the cell on the right. These vary in size but the one shown is approximately 110 m~t in diameter ( x 80,000).
C type particles budding from a platelet of a kitten injected with tissue extract from a case of feline lymphosarcoma. They measure approximately 100 mB in diameter ( X 80,000).
Section of a platelet showing an area of microtubules cut in cross section. They range in size from 30-70 m~t in diameter ( X 80,000).
Nuclear pores in cross section. They are approximately 100 m~t in diameter (× 80,000).
Micropinocytosis vesicles in the cytoplasm of a red cell precursor. They are approximately 75 m~t in diameter (X 80,000).
Perichromatin bodies in the nucleus of a blood cell. These are electron dense particles approxi- mately 50 mix in diameter surrounded by a halo of less electron dense material which separates the dense
core from the nuclear chromatin. The overall diameter is approximately 85 m B (×80,000).

See facCng page for captions f
(to face p. 352)
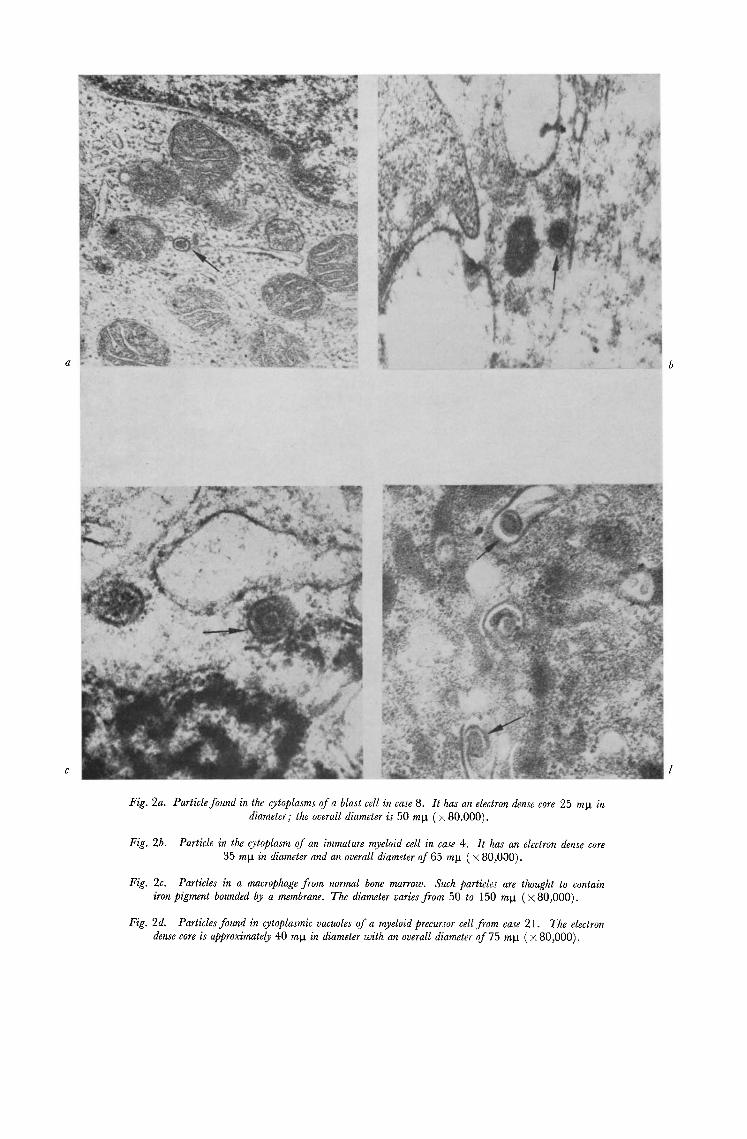
Fig. 2a. Particle found in the cytoplasms of a blast cell in case 8. It has an electron dense core 25 mg in diameter; the overall diameter is 50 mg ( × 80.000).
Fig. 2b. Particle in the cytoplasm of an immature myeloid cell in case 4. It has an electron dense core 35 mg in diameter and an overall diameter of 65 ml, t ( × 80,000).
Fig. 2c. Particles in a macrophage from normal bone marrow. Such particles are thought to contain #on pigment bounded by a membrane. The diameter varies from 50 to 150 mg (× 80,000).
Fig. 2d. Particles found in cytoplasmic vacuoles of a myeloid precursor cell from case 21. The electron dense core is approximately 40 mg in diameter with an overall diameter of 75 mla ( x 80,000).

Fig. 3a. Compound vacuole in the ctyoplasm of a lymphocyte from case 28. It contains a number of small particles approximately 50 m~t in diameter ( × 80,000).
Fig. 3b. Neutrophil granule from a normal polymorph showing a double membrane which contains particles ranging from 20 to 40 mg in diamoter ( × 80,000).
Fig. 4. Plasma pallet prepared from case 13 and sectioned at two different levels. (a) Small particles ranging from 25 to 50 mg in diameter.
(b) Particles of a larger size three of which resemble a mature C type particle. They contain a core approximately 50 mg in diameter with an overall diameter of 90 mla (× 80,000).

Fig. 5. Normal neutrophil showing perichromatin granules in the nucleus ( × 19,500).
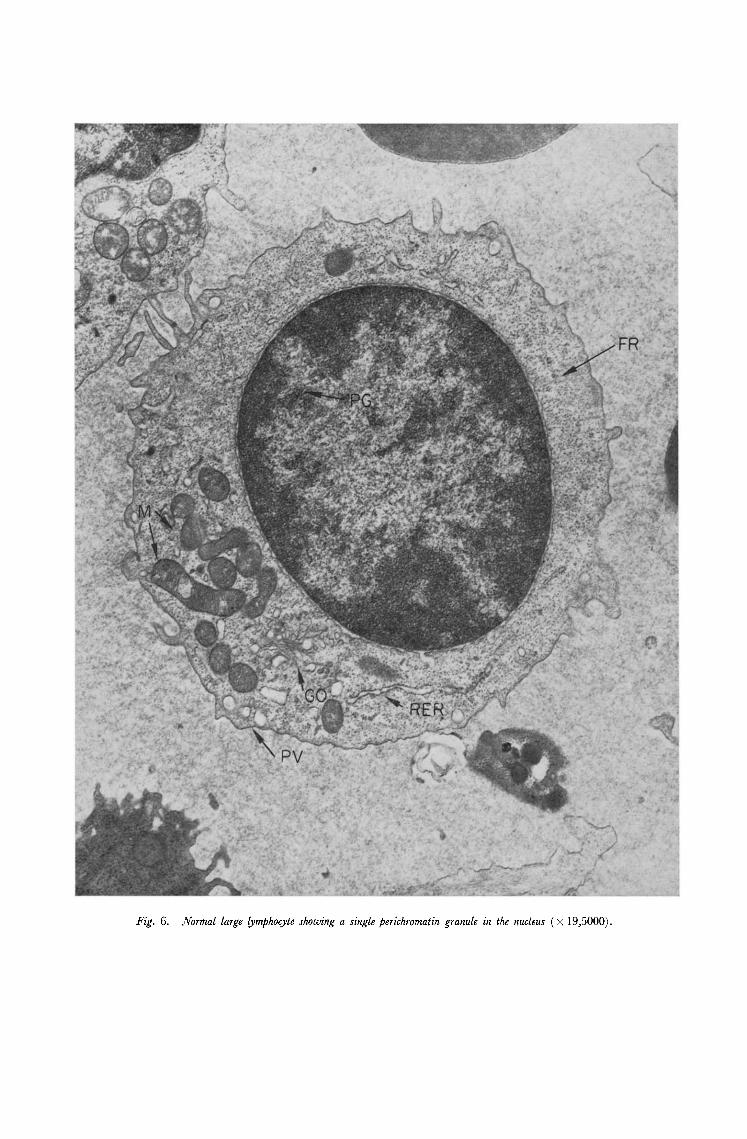
Fig. 6. Normal large lymphocyte showing a single perichromatin granule in the nucleus ( × 19,5000).
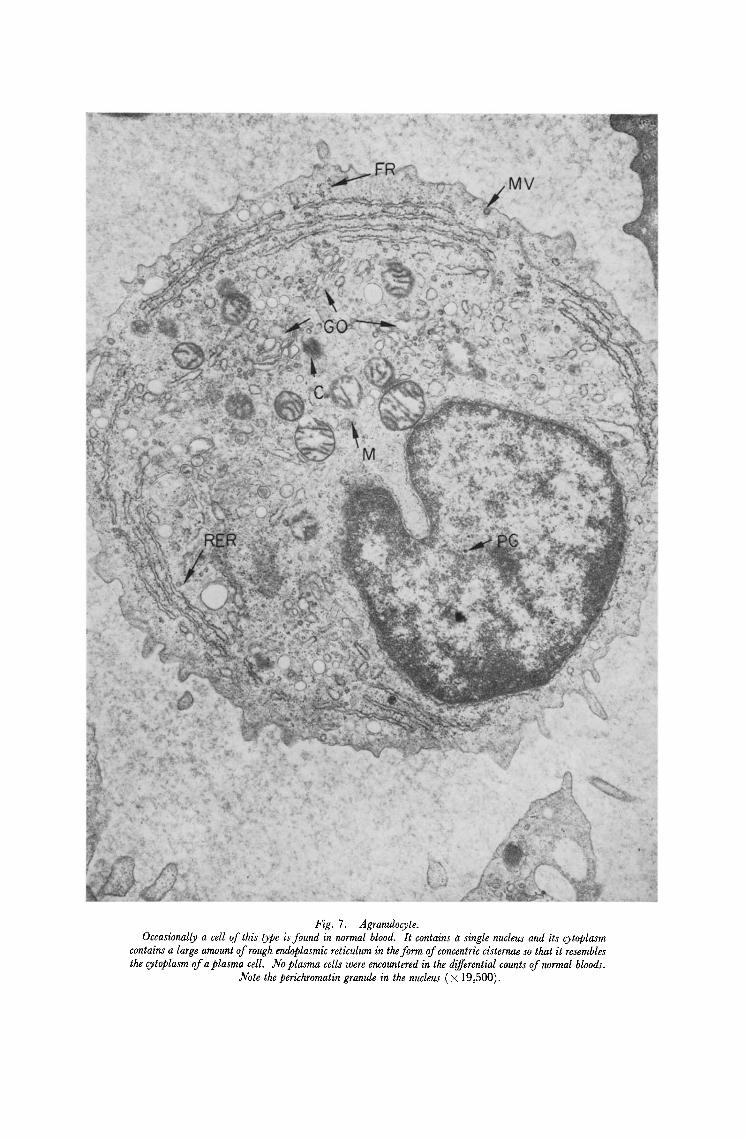
Fig, 7. Agranulocyte. Occasionally a cell of this type is found in normal blood. It contains a single nucleus and its cytoplasm
contains a large amount of rough endoplasmic reticulum in the form of concentric cisternae so that it resembles the cytoplasm of a plasma cell. No plasma cells were encountered in the differential counts of normal bloods.
Note the perichromatin granule in the nucleus ( × 19,500).
1
i
! ~ ~ i I ~ !i !
1 2
RER
PV
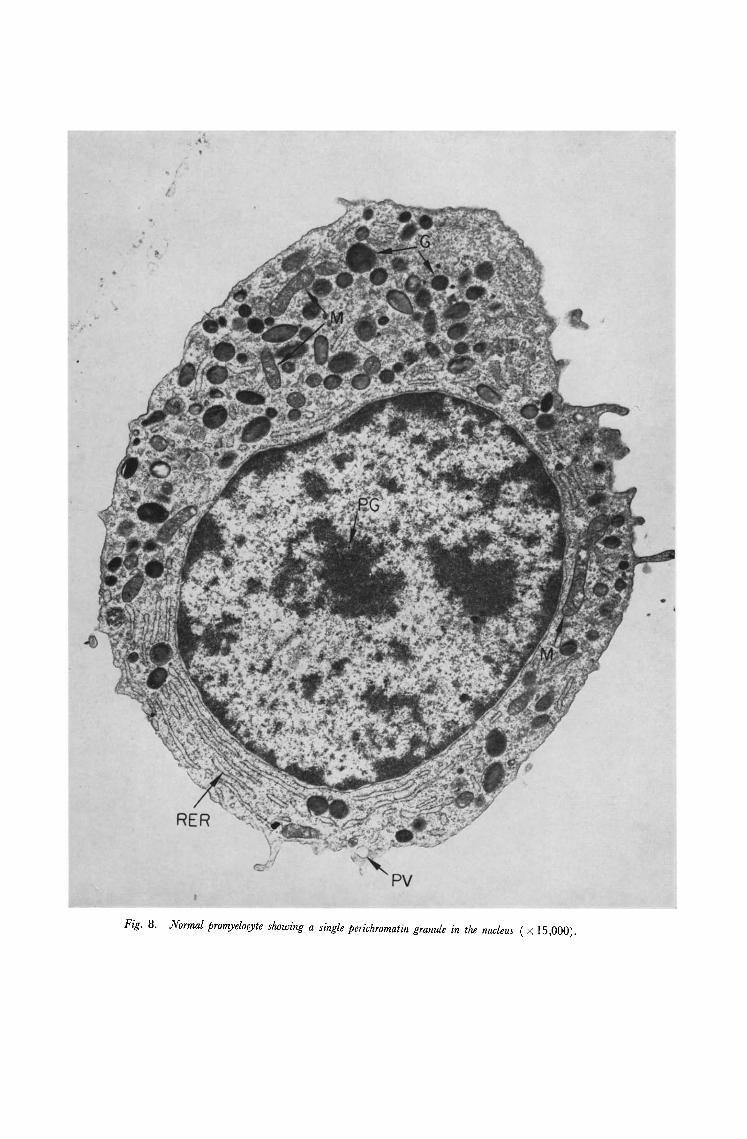
Fig. 8. Normal promyelocyte showing a single perichromatin granule in the nucleus ( × 15,000).
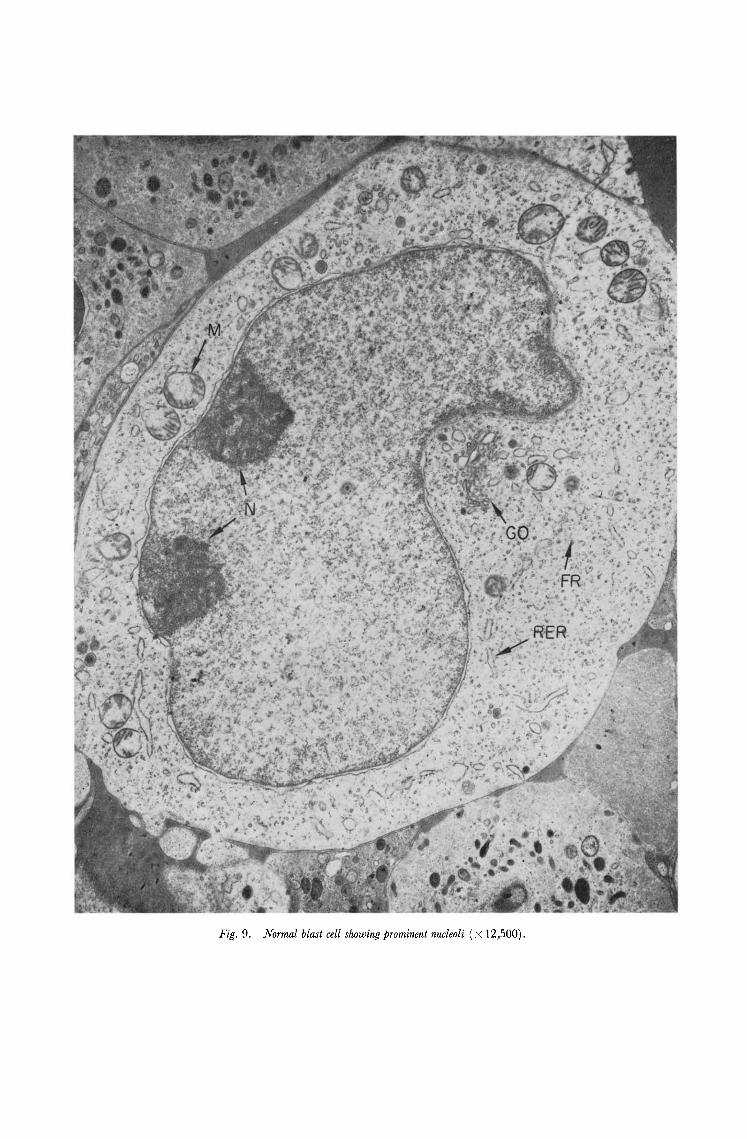
Fig. 9. Normal blast cell showing prominent nucleoli (x 12,500).

Fig. lO. Normal megakaryocyte. Note the perichromatin granules in the nucleus ( × 9,000).
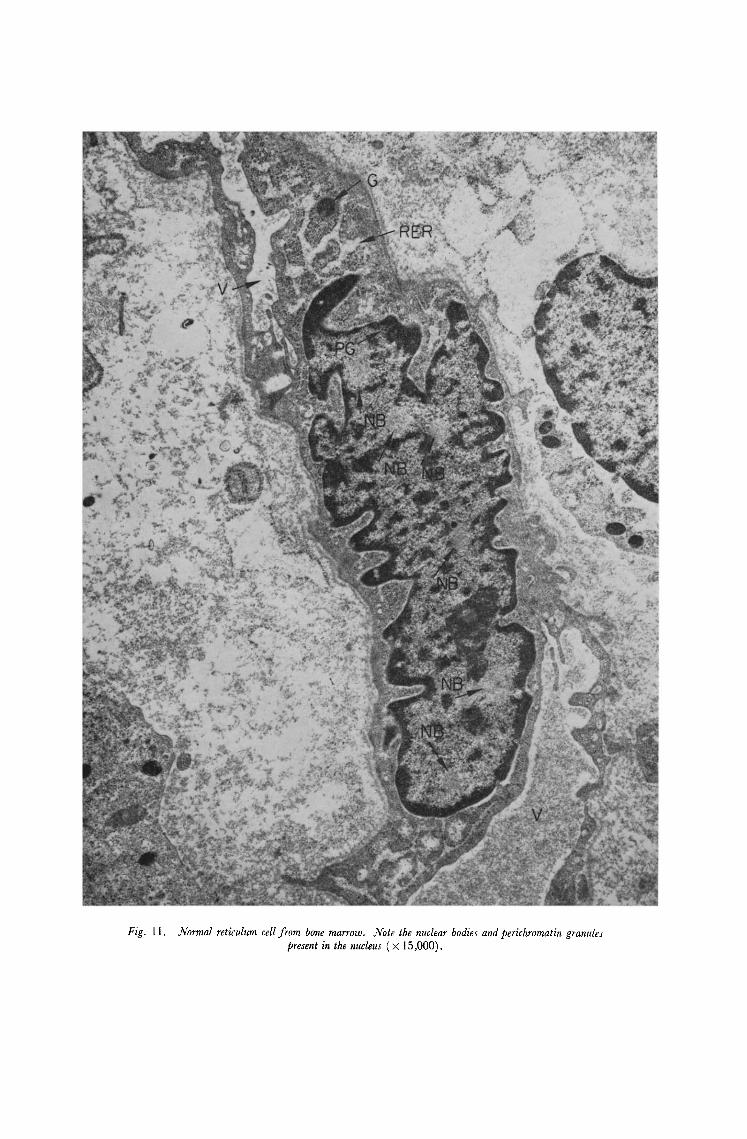
Fig. 11. Normal reticulum ceil from bone marrow. Note the nuclear bodies and perichromatin granules present in the nucleus ( × 15,000).

Fig. 12. Blast cell from case 20 showing a nuclear body of the Bouteille type 3 in the nucleus ( × 20,000).

Fig. 13. (a)Blast cell showing a nuclear bleb from case 18 ( × 37,500).
(b) BasophiI showing a nuclear ring from case 8 ( × 37,500).

Fig. 14. Blast cell from case 33 showing a large amount of fibrillar material in the cytoplasm arranged in a whorl ( X 20,000).

Fig. 15. Tubular inclusion found in an occasional lymphocyte from normal peripheral blood ( × 108,000).

Ultrastructural Studies on Normal and Leukaemic Human Haematopoietic Cells 353
to describe the morphology of normal or leukaemic blood and bone marrow in detail. The general morphology of normal human blood has already been described [8, 9] and our observations are in good accord with these previous reports. A neutrophil, lymphocyte and an agranulocyte are shown in Fig. 5-7. There are several previous reports which deal with normal bone marrow [e.g. 10, 11] and many deal with various aspects of leukaemic cell structure. These latter observations are reviewed by Anderson [8] and Kakefuda [12]. In normal marrow the various cell lineages could be readily indentified and a promyelocyte, blast cell and megakaryocyte are illustrated in Figs. 8-10. The uniformity of cell type in the leukaemic marrow as compared with normal marrow was a striking feature. Two cell types, monoblasts and lymphoblasts which are not seen in normal peripheral blood or bone marrow were observed in material from patients with acute monocytic and acute lymphatic leukaemia respectively.
A number of features have already been described in the light microscope which are specific for certain types of leukaemic cells and the ultrastructure of these features is already well documented, e.g. Freeman [13, 14] and White [15] have described Auer rods in detail and are in general agreement on their structure and probable origin. There are, however, a number of features which have only been noted since the introduction of electron microscopic techniques and some of these have been claimed as specific for leukaemic cells or to be the result of treatment. There are also a number of features which have been claimed to be more numerous in malignant cells than in normal cells. We present here our observations on these features and record some other unusual features of normal and leukaemic haematopoietic cells.
1. Perichromatin granules. These are electron dense circular bodies approximately 25 ml~ in diamater which are found in the nucleus separated from the chromatin by a clear halo 75m1~ in diameter (Fig. lf). These were originally described [16, 17] in cells of several different tissues of the rat and the mouse including eosinophil polymorphs but they have not previously been reported in human haematopoiefic cells. In the present study they were observed in all types of cell in normal and leukaemic peripheral blood and normal and leukaemic bone marrow. The frequency, which was roughly the same in all cases, averaged five granules per section of nucleus. Megakaryocytes however frequently had more
than this and it is suggested that the number of bodies may be related to nuclear size. I t has been suggested by Lynn et al. [18], who examined 102 tumour specimens and 60 non- neoplastic specimens that these granules are increased in number in malignant cells, but our observations on leukaemic cells do not support this view.
The nature of these bodies is obscure. Some authors have suggested that they may bear some relationship to virus particles [17, 18]. Because of their ubiquitous nature it seems much more probable that they are normal nuclear organelles and that if their number is related to nuclear size, this could suggest some direct relationship with the number of chromo- somes in the nucleus.
2. dVuclear bodies. These are intranuclear fibrillar structures sometimes enclosing groups of granules, first reported by de Th6 et al. [19] and described in detail by Bouteille et al. [20] and by Krishan et al. [21]. The bodies are of several types which have been classified by Bouteille et al. [20] and by Popoff and Stewart [22] who also summarise the present state of observations. It is agreed that these bodies occur in both normal and malignant tissue of a variety of different species but that they are commoner in malignant tissue and also in other tissues which are hyperactive. They are also said to be common in cells of patients with diseases of proved or supposed viral etiology [20]. It appears further that types 3 and 4 of Bouteille are commoner in malignant cells and in virus infected tissues.
In the present study nuclear bodies were not observed in normal peripheral blood, but they were found in all other tissues examined. In normal bone marrow they were only rarely observed and were normally of the type 1 of Bouteille. They were observed in both rib marrow samples and in aspirated marrows. Although they do not appear to be specific for one cell type they occur more commonly in reticulum cells, which may have several bodies in a single nucleus (Fig. 11). In leukaemic marrow they occur at roughly the same frequency as in normal marrow but in leukaemic peripheral blood, of all cell types, they appear to be relatively common. Further, in these cells the bodies are more often of types 2 and 3 (Fig. 12). Since these are more easily observed this may have contributed something to the impression of increased incidence but they are nevertheless still considered more common in leukaemic blood than in any other tissue examined.
Two points emerge from these observations.

354 Alan Ross and David Harnden
Firstly, we are not aware of nuclear bodies being described previously in normal human haematopoietic cells. Secondly, these observ- ations tend to support the view that nuclear bodies are more common in the cells of leukaemic patients than in normal individuals.
The nature of these bodies is not known. There has been much speculation but little fact has emerged. In particular there is little at present to support the view that they are virus associated inclusion bodies [23] but rather that they are associated with specific types of normal (and possibly abnormal) cellular hyper- activity [20].
,Nuclear blebs are small protrusions of the nucleus enclosing an area of cytoplasm (Fig. 13a). In some instances the outfolded portion of the nucleus appears to be separated from the main portion of the nucleus so that it forms a ring-like structure (Fig. 13b). These struc- tures were first described [24] in the immature granulocytes of patients with neoplastic disease and also in bone marrow cells of cases of pernicious anaemia treated with fluorouracil. They have since been described in normal and malignant cells of several different species. They have been found in cultured ceils of Burkitt's lymphoma [25], cells from patients with leukaemia [8, 26, 27] and in normal human blood cells [28-30]. Ahearn et al. [31] suggested an association with treatment with cytosine arabinoside. Davies and Small [32] have described the ultrastructure of rings and blebs in detail and show that they appear to be composed of two layers of nuclear membrane enclosing a single layer of the ordered structure which is found immediately below the nuclear membrane of the main part of the nucleus and are therefore closely related to normal nuclear structure.
In the present study blebs and rings were noted in a small number of normal blood and bone marrow cells. In leukaemic material blebs and rings were again found in both blood and bone marrow. In some leukaemic cases they were present in large numbers in peri- pheral blood ceils but there appeared to be no correlation with any type of leukaemia, but they do occur more commonly on polymorphs than on lymphocytes.
They were observed more frequently in leukaemic peripheral blood than in any other material. They were observed in both treated and untreated cases and we have found no correlation with any particular type of treat- ment. Our observations therefore tend to support the view that blebs are more common in malignant tissue than in normal tissue. A
further study of the factors contributing to the formation of blebs could yield useful infor- mation on normal nuclear structure and on the behaviour of malignant cells.
Micro fibrils have been reported [8] to be present in some cells of both normal and leuk- aemic patients. This finding was confirmed in the present study and in general the quantity of fibrillar material was no greater in leukaemic patients than in controls. In one case of acute myeloid leukaemia however many blast cells were found to contain large amounts of fibrous material. This material was always arranged in whorls (Fig. 14) enclosing cyto- plasmic material which appeared in some cases to be degenerating endoplasmic reti- culum. It may be relevant that this was the only case in which Auer rods were observed.
Tubular inclusions. In a small number of cases occasional lymphocytes in normal blood were found to contain in the cytoplasm aggregates of tubules which were cut at a variety of different angles (Fig. 15). The aggregates were irregular in shape, variable in size and were not enclosed in a membrane. Individual tubules were approximately 25 m~t in diameter. Somewhat similar structures have been seen in lymphocytes of a patient with chronic rheumatoid arthritis and leukopaenia [33] but in that case the tubules were sometimes contained in a vacuole and the aggregates were much larger in size. The nature of these structures is not known and they were only noted in normal blood cells.
DISCUSSION There have been many reports of virus-like
particles being found in electron microscopic studies of material from human leukaemias. The situation has, however, been complicated by the changing standards of technology in electron microscopy during the period when these studies have been made. Haemopoietic cells, particularly those of the peripheral blood, have in the past proved to be difficult to fix and section to give good preservation of the ultra structure and it is only within the last few years that thin sections of good quality could be produced with regularity from peripheral blood cells. For this reason many of the early reports which claimed to demon- strate the presence of virus particles in human leukaemic material should be considered with the greatest caution. This problem was recog- nised by the W H O subcommittee [34] which, in 1966, laid down criteria which should be fulfilled before particles observed in leukaemic material are claimed to be virus particles.

Ultrastructural Studies on Normal and Leukaemic Human Haematopoietic Cells 355
Even in recent work there is a wide divergence of opinion on whether or not virus particles can be seen. There are many reports which described particles which may be virus but which in the absence of supporting evidence cannot categorically be stated to be virus. A very small number of reports have clearly demonstrated structures which, even in the absence of infectivity studies, would be accepted by most workers as virus particles [35]. But even in these cases the question remains as to what is the relationship between the virus and the leukaemia.
There are two particular areas of interest, the herpes-fike viruses and the type-C particles. Herpes-like viruses, first described by Epstein et al. [3] in cultured cells from Burkitt's lymphoma have now been detected in many different lines of these cells by many different authors. Similar particles have been demon- strated in cultured blood cells from patients with leukaemia of several different cell types [36, 37, 39] from patients with other malig- nancies [37, 39], from patients with infectious mononucleosis [38, 39] and also from normal subjects [39]. It has been shown that contact with virus-containing nonproliferating cells can confer on normal lymphocytes the property of indefinite proliferation in suspension culture [40]. This change is associated with the transfer of the herpes-like virus. There is serological evidence that the virus is common in many different areas of the world [41, 42] and that it may be the causal agent of infectious mononucleosis [38]. Its relationship to Bur- kitt's lymphoma and to human leukaemia is still obscure but the view which Dameshek [43] has held for some time, that infectious mono- nucleosis is a leukemia which undergoes spontaneous regression, takes on a new signi- ficance in the light of the present observations. Reports of herpes-like virus in uncultured human leukaemic material are rare but Carper et al. [44] has recently reported a herpes virus in peripheral blood cells of a leukaemic child and its unaffected mother. Interest in viruses of this type as possible causal agents in human leukaemia has been sharpened by the demonstration that a herpes-virus may be the causal agent of Marek's disease in chickens where one important feature is the occurrence of lymphosarcoma [4].
Most attention has, however, been directed to attempts to demonstrate type-C particles in human leukaemia, since such particles are known to be associated with many different types of leukaemia in the mouse, with rat leukaemia, guinea pig leukaemia, feline lyre-
phosarcoma and the avian leukoses. Experience with these animal leukaemias suggests that certain sources of material harbour larger numbers of virus particles. In both avain and murine leukaemia, ultraspun pellets of peripheral blood plasma are a particularly rich source of virus particles. In the mouse leukaemias particles are also commonly seen in megakaryocytes and platelets [5, 45, 46]. Particles have, however, been observed in other blood cells with a low frequency, e.g. lympho- blasts, normoblasts and eosinophils [5].
The search for virus-like particles in man is to some extent governed by the nature of the material available. The majority of studies that have been made so far have been carried out on plasma pellets, using negative staining techniques or thin sections [1, 2, 5, 7, 47-51]. Studies on thin sectioned cellular material are less common but positive findings have been reported [5, 52, 53] while de Harven [54] and Arnoult and Hagenau [51] make negative reports. All the reports are agreed that virus-like particles are present but the contra- dictions arise at the level of interpretation. The only type of evidence which it is generally agreed would be conclusive proof of the presence of virus particles of the murine leukaemia type is the demonstration of budding particles. The few reports of budding in human material come almost exclusively from Dmochowski. His recent pictures are of great interest but they do not yet show the same detail as his own pictures of budding murine leuk- aemia particles. Further work in this field will be watched with great interest. The recent work of Laird et al. [55, 56] on the presence of budding particles in platelets of peripheral blood from kittens infected with a tissue extract from a case of feline lympho- sarcoma strengthens the conviction that only the demonstration of budding in human leuk- aemia will be acceptable as evidence for the presence of type-C particles in these cases.
In the present study a detailed search has been made for virus, virus-like particles or budding particles in thin sectioned blood and bone marrow material from a variety of different types of leukaemia and from normal individuals. Most of the virus-like particles which were observed can be identified as specific cell structures while a small number remain unidentified. None of these particles, however, fits any category of virus particle known to us and in particular they do not resemble either herpes virus or the type-C particles of the murine, avain and feline leukaemias.

356 Alan Ross and David Harnden
This result is therefore at variance with the previous observations on thin sectioned leuk- aemic ceils [5, 52, 53, 57] all reporting the finding of virus particles in a low proportion of the cases examined. On the other hand it agrees with the work of de Harven [54] and of Arnoult and Hageneau [51] who considered that none of the particles which they observed in thin sectioned material was virus.
Clearly there is still much work to be done in this field but already a number of facts are beginning to emerge. Although we found no virus particles we would not claim that this rules out their presence in human leukaemic material. Similarly the vast majority of authors who claim to find virus in human leukaemic material find it only with difficulty in a low proportion of the cases examined. It follows therefore either that virus is present in human teukaemic material in very much smaller quantities than in the animal leuk- aemias or it is in a state which renders it not easily demonstrable by the present techniques or it is not there at all and the positive reports are either artefactual or else represent the accidental presence of viruses not relevant to the aetiology of the disease.
Much of the work that has been done in this field has been influenced by the earlier studies on animal leukaemias. It now seems clear that the hypothesis of a direct parallel is no longer tenable. This is hardly surprising since work with mouse leukaemias has been done with selected stocks of virus in highly inbred animals. Similarly avain leukosis has been studied with virus which has been passed many times in experimental animals and selected domestic stocks of birds. In human leukaemia, one is dealing with an outbred population and any virus, if it is present, will not have been subjected to artificial selection. The situation may, therefore, appear to be quite different when using the approaches that have been developed for the study of the animal leukaemias. Consideration is now being given to factors which might interplay with virus in the initiation of the disease, e.g. it is quite clear that genetic factors can influence the susceptibility of animals to leukaemogenic viruses [58, 59]. A study of physiological host factors which influence the development
of virus leukaemias, whether these are environ- mentally or genetically determined, may show that the difference between human and animal leukaemias is more apparent than real.
One other possible approach is the develop- ment of new techniques for looking for evidence of virus infection. One could try to develop systems of pretreatment which might render a virus more readily detectable, e.g. Henle and Henle [60] have shown that the percent- age of cells from cell lines cultured from Burkitt's lymphoma in which virus can be readily detected may be increased dramatically by growing the cells in deficient media.
From the ultrastructural point of view one could also look for morphological features of leukaemic cells which might be indicative of virus infections although virus particles them- selves are not observed. Several of the unusual morphological features described in this paper have been claimed to be associated with the presence of virus in the cell, e.g. perichromatin bodies [18], nuclear bodies [23] or nuclear blebs [25]. While there is at present no convincing evidence that such structures are indicative of a virus infection the evidence does warrant further investigation of the conditions which control the appearance of these morphological features, e.g. the resem- blance between nuclear blebs and some of the abnormal structures which appear during infection with herpes simplex are too striking to be discarded lightly.
Ae. lmowledgm~t t~ - - -The authors are indebted to Dr. N. C. Allan, Dr. W. R. C. Altchison, Dr. S. H. Davies, Dr. C. H. Davidson, Professor K. W. Donald, Professor A. S. Douglas, Dr. M. Douglas, Professor J. O. Forfar, Dr. E. B. French, Dr. E. Innes, Dr. J . Innes, Dr. A . J . Keay, Mr. A. Logan, F.R.C.S., Dr. J . G. i a c l e o d , Professor McWhirter, Dr. M. B. Matthews, Dr. R. M. Murray Lyon, Dr. A. Muir, Dr. E. G. Oastler, Dr. J. A. Orr, Dr. J. Richmond, Dr. G. Ritchie, Dr. J . S. Robson, Dr. W. I-I. S. Schutt, Dr. J. Syme and Mr. J. D. Wade, F.R.C.S., for providing samples of peripheral blood and bone marrow from their patients; to Dr. N. C. Allan and Dr. S. FI. Davies for valuable advice and discussion; to Dr. H. Laird for the sample of cat material; to Dr. N. C. Allan and Dr. H. Laird for their critical review of this manuscript; to Miss C. W. Burns for her help with specimen collection; to Mr. N. Daviclson for assistance with the photography; and to Mr. Alexander Sutherland and Mr. Andrew Ross for their technical assistance.
RESUME La prgsence de particules virales fur recherctde, au microscope glectronique, dans des coupes fines de cellules sanguines et de ceUules mgduUaires provenant de malades atteints de leudraie et de sujets normaux. On ne trouva aucune particule qui pat ~tre considgrge comme gtant un virus. Les particules ayant l'aspect de virus furent identifiges soit comme

Ultrastruaural Studies on Normal and Leukaemic Human Haematopoietic Cells 357
des organelles cellulaires normales, soit comme diffgrant, pour l'une ou l'autre raison importante, de virus ¢onnus. De ces observations, et d'autres travaux r&ents, on condut que le concept d'un parallKisme direct avec les leudmies murines et avaires ne peut gtre ddfendu. La prgsence de particules virales, dans les cellules leudmiques humaines, ne peut gtre formellement &artge ; mais, sices particules existent, elles doivent se trouver en quantitd beaucoup plus faible que dans les leudmies des animaux.
Les autres caract#istiques ultrastructurelles des cellules normales et leudmiques furent dtudides. On en conclut que les granules pgrichromatiniens, les corps nuelgaires et les vdsicules nucldaires s'observant ~ la fois dans les cellules hdmatopofe'tiques normales et dans les cellules leudmiques. Cependant, alors que les granules pgrichromatiniens sont ggalement frgquents dans tousles cas dtudids, les corps et les vgsicules nucldaires sont plus abondants dans les cellules leudmiques que dans les cellules normales.
SUMMARY Thin sections of peripheral blood cells and bone marrow cells from patients with leukaemia and from normal subjects were examined in the electron microscope for the presence of virus particles. No particles were seen which were thought to be virus. Virus-like particles were either identified as normal cell organelles or else differed in some important respect from known viruses. On the basis of these and other recent observations it is concluded that the concept of a direct parallel with murine and avian leukaemias is no longer tenable. Although the presence of virus particles in human leukaemic cells is by no means ruled out, their frequency, i f present at all, must be very much lower than in the animal leukaemias so far studied.
Other unusual ultra structural features of normal and leukaemic cells were also studied. It is concluded that perichromatin granules, nuclear bodies and nuclear blebs are found in both normal and leukaemic haematopoietic cells but that while perichromatin granules are equally common in all cases studied, nuclear bodies and nuclear blebs are more common in leukaemic cells than in normal cells.
ZUSAMMENTASSUNG Diinne Schnitte peripherer Blutzellen und Knochenmarkzellen von Leuktimiepatienten und von gesunden Individuen wurden elektronenmikroskopisch auf das Vorhandensein yon Viruspartikeln untersucht. Keine Partikel wurden gefunden, die als Viren angesehen werden konnten. Virusiihnliche Partikel erwiesen sich entweder als normale Zellorganellen oder unterschieden sich in wesentlichen Punkten von bekannten Viren. Diese und andere, kiirzlich gemachten Beobachtungen fiihren zu der Schluflfolgerung daft das Konzept einer direkten Parallele zwischen der Leukiimie der Nager und der Viigel nicht liinger aufrecht erhalten werden kann. Obwohl die Gegenwart von Viruspartikeln in menschlichen Leu- kiimiezellen keineswegs ausgeschlossen werden darf, muflihre H~ufigkeit, falls iiberhaupt vorhanden, sehr vM geringer sein als in der Tierleuk~mie, soweit sie bis jetzt bekannt ist.
Es wurden auch andere ungewO'hnliche Ultrastrukturen normaler und leukiimischer Zellen untersucht. Folgende Schliisse wurden gezogen : Perichromatingranula, Kernein- schliisse und Kernausstiilpungen sind sowohl in normalen wie in leukdmischen hgmato- poetischen Zellen vorhanden. Perichromatingranula jedoch erscheinen gleichmiiflig in allen untersuchten Fgllen, w~hrend Kerneinschliisse und I(ernausstiilpungen hdufiger in leuk~mischen gellen auftreten als in normalen Zellen.
REFERENCES 1. P .H . LEVINE, J. S. HOROSZEWlCZ, J. T. GRACE, L. S. CHAI, R. ELLISON and
J. F. HOLLAND, Relationship between clinical status of leukaemia patients and virus-like particles in their plasma. Cancer, N.T. 20, 1563 (1976).
2. G. R. NEWELL, W. W. HARRIS, K. V, BOWMAN, C. W. BOONE and N. G. ANDERSON, Evaluation of "virus-like" particles in the plasmas of 255 patients with leukaemia and related diseases. New Engl. 07. Med. 278, 1185 (1968).
3. M . A . EPSTEIN, B. G. ACHONG and Y. M. BARR, Virus particles in cultured lymphoblasts from Burkitt's lymphoma. Lancet i~ 702 (1964).

358 Alan Ross and David Harnden
4. A.E. CHURCHILL and P. M. BraGs, Agent of Marek's disease in tissue culture. Nature (Lond.) 215, 528 (1967).
5. L. DMOCHOWSKI, T. YUMOTO, C. E. GREY, R. L. HARRIS, R. L. HALES, P. L. LANGFORD, H. G. TAYLOR, E. J. FREIREICH, C. C. SHULLENBERGER, J. A. SHIVELY and C. D. HOWE, Electron microscopic studies of human leukaemia and lymphoma. Cancer, N.T. 20, 760 (1967).
6. D. R. ANDERSON, A method of preparing peripheral leucocytes for electron microscopy. 07. Ultrastruct. Res. 13, 263 (1965).
7. G.M. PORTER III, A. J. DALTON, J. B. MOLONEY and E. Z. MITCHELL, Associ- ation of electron-dense particles with human acute leukaemia. 07. nat. Cancer Inst. 33, 547 (1964).
8. D .R . ANDERSON, Ultrastructure of normal and leukaemic leucocytes in human peripheral blood. 07. Ultrastruct. Res. Suppl. 9 (1966).
9. I. WATANABE, S. DONAHUE and N. HOGGATT, Methods for electron microscopic studies of circulating human leucocytes and observations on their fine structure. 07. Ultrastruct. Res. 20, 366 (1967).
10. M. BEssIs andJ. P. THmRY, Electron microscopy of white blood cells and their stem cells. Int. Rev. Cytol. 12, 199 (1961).
11. R.J . CAPONE, E. L. WEINm~L and B. CHAPMAN, Electron microscope studies on normal human myeloid elements. Blood 23, 300 (1964).
12. T. K~KEFUDA, Electron microscopy of normal and leukaemic cells. In Pathology of Leukemia (Edited by G. D. A~mOMIN) Harper & Row, New York (1968).
13. J . A . FREEMAN, The ultrastructure and genesis of Auer rods. Blood 15, 449 (1960).
14. J . A . FREEMAN, Origin of Auer rods. Blood 27, 499 (1966). 15. J. G. WHITE, Fine structural demonstration of acid phosphatase activity in
Auer rods. Blood 29, 667 (1967). 16. H. SWIFT, Nucleoprotein localization in electron micrographs: metal binding
and radioautography. Symp. int. Soc. Cell Biol. Vol. 1, 213 (1962). 17. M. WATSON, Observations on a granule associated with chromatin in the
nuclei of cells of rat and mouse, o7. Cell Biol. 13, 162 (1962). 18. J . A . LYNN, G. J. RACE, W. B. KINGSLEY, J. H. MARTIN and J. W. FINNEY,
Dense intranuclear particles, possible viral nucleoids in human cancer cells. Proc. Amer. Ass. Cancer Res. 9, 44 (1968).
19. G. DE TH~, M. RIVIERE and W. BERNHARD, Examen au microscope ~lectronique de la tumeur UX2 du lapin domesfique d~riv~e du papillome de Shope. Bull. Cancer 47, 570 (1960).
20. M. BOUTEILLE, S. R. KALIFAT and J. DELARUE, Ultrastructural variations of nuclear bodies in human diseases. 07. Ultrastruct. Res. 19, 474 (1967).
21. A. KRISHAN, B. U. UZMAN and E. T. HEDLEY-WHYTE, Nuclear bodies: a component of cell nuclei in hamster tissues and human tumours. 07. Ultrastruct. Res. 19, 563 (1967).
22. N. POPOFF and S. STEWART, The fine structure of nuclear inclusions in the brain of experimental golden hamsters. 07. Ultrastruct. Res. 23, 347 (1968).
23. N. GRANBOULON, P. TOURNIER, R. WICKER and W. BERNHARD, An electron microscope study of the development of SV40 virus. 07. Cell. Biol. 17, 423 (1963).
24. R. C. STALZER, J. M. KIELY, G. L. PEASE and A. L. BROWN, Effect of 5- Fluorouracil on human haematopoiesis. A morphologic study. Cancer, N.Y. 18, 1071 (1965).
25. M.A. EPSTEIN, Y. M. BARR and B. G. ACHONG, Preliminary observations on new lymphoblast strains (EB4, EB5) from Burkitt tumours in a British and a Ugandan patient. Brit. 07. Cancer 20, 475 (1966).
26. F. MoLto and A. STRAMIGNONI, Nuclear projections in blood and lymph node cells of human leukaemias and Hodgkin's disease and in lymphocytes cultured with phytohaemagglutinin. Brit. 07. Cancer 21, 519 (1967).
27. N.G. McDuFFIE, Nuclear blebs in human leukaemic eeUs. Nature (Lond.) 214, 1341 (1967).
28. D. HUHN, Nuclear pockets in normal monocytes. Nature (Lond.) 216, 1240 (1967).
29. G.F. SMITH and P. T. O'HARA, Nuclear pockets in normal leucocytes. Nature (Lond.) 215, 773 (1967).
30. G.F . SMITH and P. T. O'HARA, Structure of nuclear pockets in human leuco- cytes. J. Ultrastruct. Res. 21, 415 (1968).

Ultrastructural Studies on Normal and Leukaemic Human Haematopoietic Cells 359
31. M. J . AHEARN, C. W. LEWIS, L. A. CAMPBELL and J. K. LucE, Nuclear bleb formation in human bone marrow cells during cytosine arabinoside therapy. Nature (Lond.) 215, 196 (1967).
32. H .G. DAVIES and J. V. SMALL, Structural units in chromatin and their orient- ation on membranes. Nature (Lond.) 217, 1122 (1968).
33. T. Hovlo, M. JmaEMIC and P. STAVEM, A new type of inclusion body in lym- phocytes. 07. Ultrastruct. Res. 20, 298 (1967).
34. W. BERNHARD, M. BENYEsH-MELINCK and J. MELNICK, Statement on the detection of viruses by electron microscopy in human leukaemic tissues and extracts and in similar material from domestic animals. Bull. Wld Hlth Org. 34, 953 (1966).
35. M. V. VIOLA, A. J. DALTON, E. MITCHELL and J. B. MOLONEY, Virus-like particles in a patient with chronic lymphatic leukaemia. New Enfl. 07. Med. 277, 503 (1967).
36. V . H . ZEVE, L. S. LucAs and R. A. MANAKER, Continuous cell culture from a patient with chronic myelogenous leukaemia. II. Detection of a herpes-like virus by electron microscopy. 07. nat. Cancer Inst. 37, 761 (1966).
37. E .M. JENSEN, W. KOROL, S. L. DUTTMAR and T. J. MEDREK, Virus containing lymphocyte cultures from cancer patients. 07. nat. Cancer Inst. 39, 745 (1967).
38. G. HENLE, W. HENLE and V. DIEHL, Relationship of Burkitt's tumor-associa- ted herpes type virus to infectious mononucleosis. Proc. Nat. Acad. Sci., Wash. 59, 94 (1968).
39. G.E. MooaE, H. KITAMURA and R. E. GERNER, Incidence of "leukovirus" in human hematopoietic cell lines. Proc. Amer. Ass. Cancer Res. 9, 52 (1968).
40. W. HENLE, V. DIEHL, G. KOHN, H. ZUR HAUSEN and G. HENLE, Herpes-type virus-chromosome marker in normal leucocytes after growth with irradiated Burkitt cells. Science, N.Y. 157, 1064 (1967).
41. L . J . OLD, E. A. BOYSE, H. F. OETTGEN, E. DE HARVEN, G. GEERINO, B. WILLIAMSON and P. CLIFFORD, Precipitating antibody in human serum to an antigen present in cultured Burkitt's lymphoma cells. Proc. Nat. Acad. Sci., Wash. 56, 1699 (1966).
42. G. HENLE and W. HENLE, Immunofluorescence, interference and complement fixation technics in the detection of the herpes-type virus in Burkitt tumor cell lines. Cancer Res. 27, 2442 (1967).
43. W. D,~,'~ESHEK, Glandular fever cells. Lancet i, 333 (1967). 44. J. M. CARPER, W. M. O'DoNNELL and J. H. MONROE, Neonatal leukaemia.
Detection of herpes virus in the mother and child. Amer. 07. Dis. Child. 115, 61 (1968).
45. A.J . DALTON, F. HAOUENAU and J. B. MOLONEY, Further electron microscopic studies on the morphology of the Moloney agent. 07. nat. Cancer Inst. 33, 255 (1964).
46. A.J . DALTON, L. W. LAW, J. B. MOLONEY and R. A. MANAKER, An electron microscopic study of a series of murine lymphoid neoplasms. 07. nat. Cancer Inst. 27, 747 (1962).
47. C.L. BURGER, W. W. HARRIS, N. G. ANDERSON, T. W. BARTLETT and R. M. KNISELEY, Virus-like particles in human leukemic plasma. Proc. Soc. exp. Biol. Med. 115, 151 (1964).
48. K . O . SMITH, M. BENYESH-MELNICK and D. J. FERNBACI-I, Studies on human leukaemia. II. Structure and quantitation of myxovirus-like particles associated with human leukemia..7- nat. Cancer Inst. 33, 557 (1964).
49. M. BENYESH-MELNICK, K. O. SMITH and D. J. FERNBACI-I, Studies on human leukaemia. III. Electron microscopic findings in children with acute leukaemia and in children with infectious mononucleosis. 07. nat. Cancer Inst. 33, 571 (1964).
50. A. M. PRINCE and W. R. ADAMS, Virus-like particles in human plasma and serum role of platelet lysosomes. 07. nat. Cancer Inst. 37, 153 (1966).
51. J. ARNOULT and F. HAOUENAU, Problems raised by the search for virus particles in human leukaemia. A study with the electron microscope of blood plasma, cerebrospinal fluid, and megakaryocytes from the bone marrow. 07. nat. Cancer Inst. 36, 1089 (1966).
52. J . D . ALMEIDA, R. C. HASSELBACK and A. W. HAM, Virus-like particles in two leukemic patients. Science, N.Y. 142, 1487 (1963).
53. M. BRAUNSTEINER, K. FELLINGER and E. PAKESCH, On the occurrence of virus- like bodies in human leukemia. Blood 15, 476 (1960).

360 Alan Ross and David Harnden
54. E. DE HARVEN, B. CLARKSON and A. STRIFE, Electron microscopic study of human leukaemic cells in tissue culture. Cancer, N.Y. 20, 911 (1967).
55. H.M. LAIRD, W. F. H. JARRETT, 0. JARRETT and G. W. CRIGHTON, Virus-like particles in three field cases of feline lymphosarcoma. Vet. Rec. 80, 606 (1967).
56. H . M . LAIRD, 0 . JARRETT, G. W. CRIGHTON, W. F. H. JARRETT and D. HAY, Replication of leukemogenic-type virus in cats inoculated with feline lympho- sarcoma extracts. 07. nat. Cancer Inst. 41, 879 (1968).
57. L. DMOCHOWSKI, C. E. GREY, J. A. SYKES, C. C. SHULLENBERGER and C. D. HOWE, Studies on human leukaemia. Proc. Soc. exp. Biol. Med. 101, 686 (1959).
58. R . A . STEEVES, M. BENNETT and E. A. MIRAND, Genetic control by the W. locus of susceptibility to (Friend) spleen focus forming virus. Nature (Lond.) 218 372 (1968).
59. L. M. PAYNE and P. M. Bmos, Differences between highly inbred lines of chickens in the response to rous sarcoma virus of the chorioallantoic membrane and of embryonic cells in tissue culture. Virology 24, 610 (1964).
60. W. HENLE and G. HENLE, Effect of Argine-deficient media on the herpes-type virus associated with cultured Burkitt tumor cells. J. Virology 2, 182 (1968).