two bestrophins cloned from xenopus laevis oocytes express ... file3 introduction ca-activated cl...
TRANSCRIPT

1
Two Bestrophins Cloned from Xenopus laevis Oocytes
Express Ca-activated Cl Currents.
Zhiqiang Qu, Raymond W. Wei, Wesley Mann, and H. Criss Hartzell†
Department of Cell Biology
Emory University School of Medicine
535 Whitehead Biomedical Research Building
615 Michael St.
Atlanta, GA 30322
† Corresponding author.
telephone: 404-727-0444
fax: 404-727-6256
email: [email protected]
Running title. Xenopus bestrophins are Ca-dependent Cl Channels
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on August 25, 2003 as Manuscript M308414200 by guest on D
ecember 31, 2019
http://ww
w.jbc.org/
Dow
nloaded from

2
Summary
Ca-activated Cl channels play important diverse roles from fast block to polyspermy
to olfactory transduction, but their molecular identity has not been firmly established. By
searching sequence databases with the M2 pore domain of ligand-gated anion channels, we
identified potential Ca-activated Cl channels which included members of the bestrophin
family. We cloned two bestrophins from Xenopus oocytes, which express high levels of Ca-
activated Cl channels. The Xenopus bestrophins were expressed in a variety of tissues. We
predict that bestrophin has 6 transmembrane domains with the conserved RFP domain
playing an integral part in ionic selectivity. When Xenopus bestrophins were heterologously
expressed in HEK-293 cells, large Ca-activated Cl currents were observed. The currents are
voltage-and time-independent, do not rectify, have a Kd for Ca of ~210 nM, and exhibit a
permeability ratio of I > Br > Cl >> aspartate. The W93C and G299E mutations produce non-
functional channels that exert a dominant negative effect on wild type channels. We conclude
that bestrophins are the first molecularly identified Cl channels that are dependent on
intracellular Ca in a physiological range.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

3
Introduction
Ca-activated Cl currents play a variety of important physiological roles which include
functions as diverse as the fast block to polyspermy and olfactory transduction (1). Despite
their physiological importance, the molecular identity of these channels remains in question.
The CLCA family, initially cloned from bovine trachea by Cunningham et al. (2), has been
proposed to code for Ca-activated Cl channels. Heterologous expression of hClCA1,
hCLCA2, mCLCA1, mCLCA2, bCLCA1, and pCLCA1 results in an increased Cl current that
is stimulated by intracellular Ca (3-7). However, CLCA proteins have not been universally
accepted as Ca-activated Cl channels for several reasons (8;9). The properties of CLCA-
induced currents differ depending on the cell type in which they are expressed. Furthermore,
some CLCAs are cell adhesion proteins or secreted proteins with poorly conserved trans-
membrane architecture. Furthermore, the current is activated by very high, non-physiological
Ca concentrations and native expression of CLCAs does not correlate with Ca-activated Cl
currents.
Several years ago, we began an in silico approach to identify novel Cl channel genes.
Our previous studies on the endogenous Ca-activated Cl channels of Xenopus oocytes
suggested that these channels might have a pore architecture similar to ligand gated anion
channels (LGACs) of the GABA(A), GABA(C), and glycine receptor family (10;11).
LGACs and Ca-activated Cl channels have similar anion selectivity sequences and similar
pore dimensions. Furthermore, both are blocked by anion channel blockers that enter the pore
selectively from the extracellular side, unlike CFTR and ClC channels that seem to have the
wide end of their pores oriented cytoplasmically (12).
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

4
The determinants of ligand-gated channel anion selectivity have been well-studied.
Mutagenesis of both anion- and cation-selective ligand-gated channels have established that
3 rings of charged residues formed by the M2 transmembrane domains of the channel
subunits play a major role in ion selectivity (13;14). In LGACs, both the intermediate and
extracellular rings are positively charged, whereas in cation-selective channels these residues
are negatively charged. Although both the extracellular (15) and intermediate rings (16)
contribute to ion selectivity, the intermediate ring is critical because the pore tapers to a
constriction at this point to bring the charged residues of the intermediate ring close to the
permeant ion. This geometric constraint is apparently conferred on the channel by a proline
residue at position -2’ (17;18). Mutations affecting the proline and the positively charged
intermediate ring can convert selectivity from anionic to cationic (17;19).
We used degenerate M2 domains of LGACs to search for potential unique Cl
channels. Among the hits we obtained was a family of proteins called bestrophins. Human
bestrophin-1 (VMD2) was positionally cloned in 1998 from a Swedish family with an
inherited form of early-onset macular degeneration called Best vitelliform macular dystrophy
(20). Bestrophin was initially thought to be involved in fatty acid transport (20), but more
recently, it has been shown that human bestrophin-1 expressed in heterologous systems
induces a chloride current (21). The possibility that bestrophin is a Cl channel is particularly
exciting because the hallmark diagnostic feature of Best disease is an abnormal electro-
oculogram (22;23). The slow light peak of the electro-oculogram, which is reduced in Best
disease, is thought to reflect an increase in Cl conductance across the basolateral membrane
of the retinal pigment epithelium (RPE) (24). Trans-RPE transport plays an important role in
maintaining the fluid and ionic composition of the fluid surrounding the photoreceptors.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

5
Recently, it has been demonstrated that human bestrophin-1 is localized to the basolateral
membrane of RPE (25), making it a candidate for the basolateral Cl channel of RPE.
In this paper, we report the cloning and expression of two bestrophins from Xenopus
oocyte mRNA. These proteins have high similarity to human bestrophins and induce Cl
currents when expressed in heterologous systems.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

6
Experimental Procedures
Cloning.
ESTs for Xenopus bestrophins were identified by BLAST searches using conserved
domains of mammalian bestrophins. PCR primers were constructed and full-length Xenopus
bestrophins were obtained by RT-PCR using total RNA isolated from Xenopus oocytes. The
resulting PCR products were subcloned into TOPO-PCRII and sequenced in both forward
and reverse directions. Each clone was sequenced at least three times. xBest-2b was
subcloned into pIRES2-EGFP (Invitrogen) by PCR. The forward PCR primer contained a
Bgl II restriction site and the reverse primer contained a Sac II restriction site for insertion
into the pIRES2-EGFP vector.
Site-specific mutation of xBest-2a:
Site-specific mutations were made using a PCR-based site-directed mutagenesis kit
(Quickchange, Stratagene). Specific mutations were introduced into primers. The template,
xBest-2a in pCMV-Sport6, was amplified with the primers and Pfu DNA polymerase by the
polymerase chain reaction. The methylated original templates were digested with Dpn 1 and
the PCR products were transformed into XL-1 Blue E. coli. Mutations were confirmed by
DNA sequencing.
Heterologous expession of bestrophins in HEK-293 cells
XBest-2a subcloned in pCMV-Sport6 was transfected into HEK-293 cells with
pIRES2-EGFP vector at a ratio of 10 : 1. XBest-2b was subcloned in pIRES2-EGFP vector.
Plasmid transfection was carried out with Fugene 6 kit (Roche) or Ca-PO4 precipitation
(Clontech). Transfected cells were dissociated and replated one day after transfection and
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

7
spread on glass coverslips. Fluorescent cells were used for patch clamp experiments within
three days.
Antibody production.
Antibodies were raised against a peptide of xBest-2a. The amino acid sequence was
EFQSQEPIQDPPYN, which corresponds to amino acids 451-464 of xBest-2a. xBest-2b
differs in 6 of these 14 residues (VFQFPETVQDPPNN). Blast hits with an E value <10
included only xBest-2a. Peptides were synthesized by Research
Genetics (Invitrogen) using MAP resin technology to enhance antigenic response. The
peptides (with a cysteine added to the C-terminus) were conjugated to KLH, emulsified with
an equal volume of Freud’s incomplete adjuvant and injected into two New Zealand white
rabbits. The animals received a boost after two, six, and eight weeks. Sera was assayed by
ELISA and western blot analysis with GST-fusion proteins of bestrophins. The sera were
then affinity purified.
GST fusion proteins.
GST fusion proteins of several bestrophins were engineered by sub-cloning the
bestrophin C-terminal tail into pGEX-4T expression vectors (Amersham) in frame with GST.
The C-terminal fragments used for xBest-2a and xBest-2b were residues 291 – 512
(GEQLIN….LSVAT). The resulting GST fusion protein consisted of N-terminal GST
followed by the C-terminal fragments of the bestrophin. The fusion proteins were grown in
BL21-Gold(DE3) competent E. coli (Stratagene), and induced by IPTG at final concentration
of 1mM for 3-4 hours at 37oC. Bacterial pellets were resuspended in 50mM Tris, pH 8.0,
40mM EDTA, 2.5% sucrose, 0.02% NaN3, 1mM PMSF, 2.5mg/ml lysozyme, and protease
inhibitor cocktail set II (Calbiochem, 5 µl/ 20 mg bacteria). The lysate was subjected to
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

8
French press twice at 1000 psi. The lysate was centrifuged at 10,000 rpm for 15min, and
then passed over a glutathione Sepharose 4B columns (Amersham). The fusion proteins
were eluted from the columns with 10mM reduced Glutathione, 50mM Tris-HCl, pH 8.0.
Total RNA isolation and purification:
Total RNA from Xenopus RPE, neural retina, brain, spleen, gut, lung, liver, blood,
heart, and oocyte were isolated using Trizol Reagent (Invitrogen), a mono-phasic solution of
phenol and guanidine isothiocyanate. Tissue (1 g/ 10 ml Trizol) was homogenized using a
Polytron (Brinkman) homogenizer. The Trizol was extracted two times with chloroform, and
the total RNA was recovered by precipitation with isopropyl alcohol. DNA was removed by
DNase treatment and – DNA-freeTM (Ambion).
Semi-Quantitative PCR:
cDNA was synthesized from total RNA using ThermoScriptTM reverse transcriptase
(Invitrogen) primed with oligo(dT). The reaction mixture contained 4 µg of total RNA, 2.5
µM oligo-dT, 1 mM dNTP mix, 0.1 mM DTT, 2U/µl RNaseOUT, and 0.75 U/µl
ThermoScriptTMRT. The cDNA was amplified by using XBest-2a specific primers and
SYBR Green JumpStartTM Taq ReadyMixTM (Sigma). The concentration of primers was 40
nM. Data were analyzed with iCycler software, which determines the threshold cycle for
each sample. The house keeping gene GAPDH of the same cDNA sample was also
amplified. The quantification of the xBest-2a gene is defined by the ratio of the threshold
cycle values of xBest-2a against the GAPDH in the same sample. The thermocycler used for
quantitative detection was iCycler (BioRad), and the protocol was: one cycle at 95oc for 2
min, 40 cycles at 95oc for 30 sec, 60oc for 30 sec, and 68oc for 30 sec. The primers for xBest-
2a were 5’ - TTG GCT GAA GGT GGG TGA ACA – 3’ and 5’ – GGG CGC GGG TCT
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

9
GAG TGA TT – 3’. The primers for GAPDH were 5’ – GAC CTG CCG CCT GCA GAA G
– 3’ and 5’ – GAC TAG CAG GAT GGG CGA C – 3’.
SDS-PAGE and Western blots.
Xenopus laevis were acutely decapitated and organs were quickly collected and
weighed. The organs were homogenized (50mg tissue per ml) in the LSB buffer (60mM
NaCl, 25mM Na-PIPES pH 6.9, 1mM EDTA, 2mM NaN3, 0.3 mM β-mecaptoethanol, 0.1
mM PMSF and 100-fold diluted protease inhibitor cocktail III (Calbiochem)). SDS was
added to a final concentration of 1%. DNA was sheared by passing the solution through a 20
gauge needle. Protein concentration was measured by the Bradford assay (Bio-Rad) and
checked with Coomassie blue staining in gels. Protein samples were run on 4-15% gradient
polyacrylamide gels in 25mM Tris-HCl pH 8.3, 200mM Glycine, 0.5% SDS with ~10 µg of
protein per well. The proteins were electrophoretically transferred to Hybond nitrocelluose
membranes in 25 mM Tris-HCl pH 8.3, 200mM Glycine, 20% methanol.. The membranes
were blocked with 5% dry milk in PBS with 0.1% Tween 20 (PBS-T) overnight at 4oC or 1
hr at room temperature. The blot was incubated with primary antibody (1/1000 dilution) and
horseradish peroxidase conjugated goat anti-rabbit IgG (1/7000) (Jackson Immuno Research)
in PBS-T with 1% dry milk. Immunoreactivity was visualized by enhanced
chemiluminescence (ECL kit, Amersham Biosciences).
Immunocytochemistry.
HEK-293 cells were fixed in 4% paraformaldehyde in 0.1 M sodium phosphate buffer
pH 7.3 for 2 hr. The cells were washed 3 times with PBS (136 mM NaCl, 2.7 mM KCl, 10
mM Na2HPO4, 6 mM NaH2PO4, pH 7.3) and then blocked with PBS containing 0.1% Triton-
X100, 3% bovine serum albumin and 10% normal goat serum for 30 min. The cells were
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

10
then incubated at 4oC overnight with 180 ng/ml affinity-purified A5925 in PBS containing
2% BSA overnight. The cells were then washed extensively with PBS and incubated with
Alexa-488 (Molecular Probes) conjugated goat-anti-rabbit IgG (Molecular Probes) diluted
1:200. The cells were visualized using a Zeiss 510 confocal microscope.
Electrophysiological Methods
Recordings were performed using the whole-cell recording configuration of the patch
clamp technique. Patch pipettes were made of borosilicate glass (Sutter Instrument Co.,
Novato CA), pulled by a Sutter P-2000 puller (Sutter Instrument Co.), and fire-polished.
Patch pipets had resistances of 3 - 5 MΩ filled with the standard intracellular solution (see
below). The bath was grounded via a 3M KCl-agar bridge connected to a Ag-AgCl-
reference electrode. Solution changes were performed by gravity-feed of the 1 ml chamber at
a speed of ~ 4 ml/min. To measure the steady-state current-voltage relationship, the cells
were voltage clamped from a holding potential of 0 mV with 700-msec duration pulses from
-100 mV to +100 mV. To measure the instantaneous current-voltage relationship, a pre-pulse
was first applied to +100mV for 200ms and then 500 msec duration voltage steps were
applied between -100 mV and +100 mV. Data were acquired by an Axopatch 200A amplifier
controlled by Clampex 8.1 via a Digidata 1322A data acquisition system (Axon Instruments,
Foster City CA). Experiments were conducted at room temperature (~ 24 °C).
Recording Solutions.
The standard pipette solution contained (mM): 146 CsCl, 2 MgCl2, 5 (Ca)-EGTA, 8
HEPES, 10 sucrose, pH 7.3. The “zero” free Ca2+ pipette solution contained 5 mM EGTA
without added Ca2+ while high free Ca2+ pipette solution contained 5mM Ca-EGTA made by
the pH-metric method described by (26). Working solutions having different free Ca2+ were
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

11
prepared by mixing the zero-Ca2+ solution with the high Ca2+ solution in various ratios. The
free [Ca2+] was calculated from the equation
[Ca2+] = K d x [Ca-EGTA] / [EGTA]
where Kd is the Kd of EGTA (Kd=1.0 x 10-7 M at 24°C, pH 7.3, ionic strength 0.16 M). The
calculated Ca2+ concentrations were confirmed in each solution by fura-2 (Molecular Probes)
measurements using an LS-50B luminescence spectrophotometer (Perkin Elmer, Norwalk
CT). In some experiments, pipette solution contained (mM) 148 CsCl, 2 MgCl2, 0.5 CaCl2, 5
EGTA, 8 HEPES, pH 7.3 with free [Ca] = 164 nM.
The standard extracellular solution contained (mM) 140 NaCl, 5 KCl, 2 CaCl2, 1
MgCl2, 15 glucose, 10 HEPES, pH 7.4. When Cl was substituted with another anion, NaCl
was replaced with NaX, where X is the substitute anion.
Solution osmolarity was 300 mOsm for both intra- and extracellular solutions (Micro
Osmometer, Model 3300, Advanced Instrument, INC Norword, Massachusetts, USA). Slight
differences in osmolarity were adjusted by addition of sucrose.
Analysis of Data
For the calculations and graphical presentation, we used Origin 6.0 software
(Microcal, Northampton MA). Relative halide permeability of the channels was determined
by measuring the shift in Erev upon changing the bath solution from one containing 151 mM
Cl to another with 140 mM X and 11 mM Cl, where X is the substitute anion. The
permeability ratio was estimated using the Goldman-Hodgkin-Katz equation (27).
Px / PCl = [Cl]i / ([X]o exp(∆ErevF/RT))-[Cl] o / [X] o
where ∆Erev is the difference between the reversal potential with the test anion X and that
observed with symmetrical Cl, and F, R, and T have their normal thermodynamic meanings.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

12
Accession numbers.
XBest-2a has been enetered in Genbank as accession AY273825 and XBest-2b has
been entered as AY273826. hBest1 (VMD2): NM_004183, hBest2 (VMD2L1):
NM_017682, hBest3 (VMD2L2): NM_017682, hBest4 (VMD2L3): NM_152439.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

13
Results
In silico cloning of Ca-activated Cl Channels.
Blast searching (28) of protein databases with degenerate M2 domains of LGACs
found a number of proteins of the bestrophin family. Fig. 1 shows an alignment of the M2
domain of the human GABA(A) α1 subunit with four human bestrophins. The residues of the
GABA(A) receptor are labeled in accordance with Keramidas (29) with the arginines of the
intermediate and extracellular rings located at position 0’ and 19’, respectively. The
homology between the bestrophins and the GABA(A) receptor in this region is rather weak
(19% identity, 39% strong similarity). However, the critical residues for GABA(A) receptor
ion selectivity are conserved: the 19’ R of the extracellular ring, the 0’ R of the intermediate
ring, the -5’ negative charge of the cytoplasmic ring, and the -2’ proline at the pore
constriction. The physical properties of the M2 domain of the hGABA(A)R and the RFP
domain of xBest-2a are quite similar (Table 1). Interestingly, the RFP sequence (positions 0
to -2’) of the bestrophins that corresponds to the intermediate ring and associated proline of
the GABA receptor is invariant among all 40+ members of the bestrophin family for which
sequences are available, from C. elegans to human. This strongly suggests that the RFP
sequence is crucial to channel function. We propose it is involved in ion selectivity.
Because Ca-activated Cl channels are expressed at high levels in Xenopus oocytes
(12), we used the sequences obtained from mammalian bestrophins to find ESTs for Xenopus
bestrophins. One Xenopus EST (BE669309, Image clone 3401219) was obtained and
sequenced and found to be a full-length bestrophin. The sequence was confirmed by
sequencing full-length PCR products obtained by RT-PCR of mRNA isolated from Xenopus
oocyte mRNA. At least 3 overlapping sequencing runs were performed. The PCR products
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

14
from different clones differed reproducibly in sequence from one another, so new primers
were designed to amplify a second, closely related transcript (xBest-2b). Fig. 2 compares the
sequences of the two Xenopus bestrophins to four human bestrophins. Evolutionary
relationships between 15 vertebrate bestrophins for which complete sequence information is
available was determined by the neighbor joining method of Saitou and Nei (1987) (Fig. 3A).
The bestrophins from Zebrafish, Fugu, Xenopus, mouse, rat, and human fall into 4 groups
with the both Xenopus bestrophins being in group 2. The two Xenopus bestrophins are 89%
identical. Because Xenopus laevis is pseudo-tetraploid and xBest-2a and xBest-2b are more
closely related to one another than any of the four human bestrophins are to one another, we
believe that the Xenopus bestrophins are orthologs.
All bestrophins are highly conserved in the first ~300 residues, but differ significantly
in the C-terminal third of the protein. xBest-2a is 95% identical to xBest-2b in the first 307
amino acids, but only 85% identical in the remaining 205 amino acids. xBest-2a is 81%
identical to hBest-2 in the first 307 amino acids, but only 50% identical from residue 307 to
the end.
The Xenopus bestrophins contain multiple PKA, casein kinase, and PKC
phosphorylation consensus (PROSITE) sequences, but only three were conserved in the
human proteins (Fig. 2). T164 is a predicted casein kinase site that is conserved in all six
bestrophins shown here and is conserved in a majority of the other bestrophins we have
examined. T6 and T216 are predicted PKC phosphorylation sites found in 4 and 3,
respectively, of the bestrophins shown here. The other phosphorylation sites were not
conserved across species.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

15
Kyte-Doolittle hydropathy analysis of xBest-2a with a window of 17 found 6
hydrophobic domains (Fig. 3B). The SOSUI algorithm predicted 6 transmembrane domains
(shown as red lines over the sequence in Fig. 2). The (150)RFP sequence is at the C-terminal
end of the 4th transmembrane domain and thus would be predicted to be located at the
cytoplasmic mouth of the channel. Fig. 3C shows our model for xBest-2a bestrophin
topology.
Bestrophin Expression.
Semi-quantitative PCR. Real-time PCR was used to evaluate the tissue-specific
expression of XBest-2. The primers that were chosen were identical to sequences in both
XBest-2a and XBest-2b. Thus, the quantification represents the total of both orthologs (Fig.
4). XBest-2 mRNA is expressed at high levels in RPE, liver, spleen and to a lesser extent in
neural retina and lung. Relatively little message was found in brain, heart, and gut. Even
though we were able to RT-PCR full-length xBest-2a and xBest-2b from RNA isolated from
oocyte, we were unable to quantify the level of xBest-2 message by real-time PCR.
Western Blot Analysis. Polyclonal antibodies were developed to peptide sequences of
the Xenopus bestrophins. Affinity-purified antibody A5925 was raised against residues 441-
454 of xBest-2a (EFQSQEPIQDPPYN). This antibody specifically recognized GST-fusion
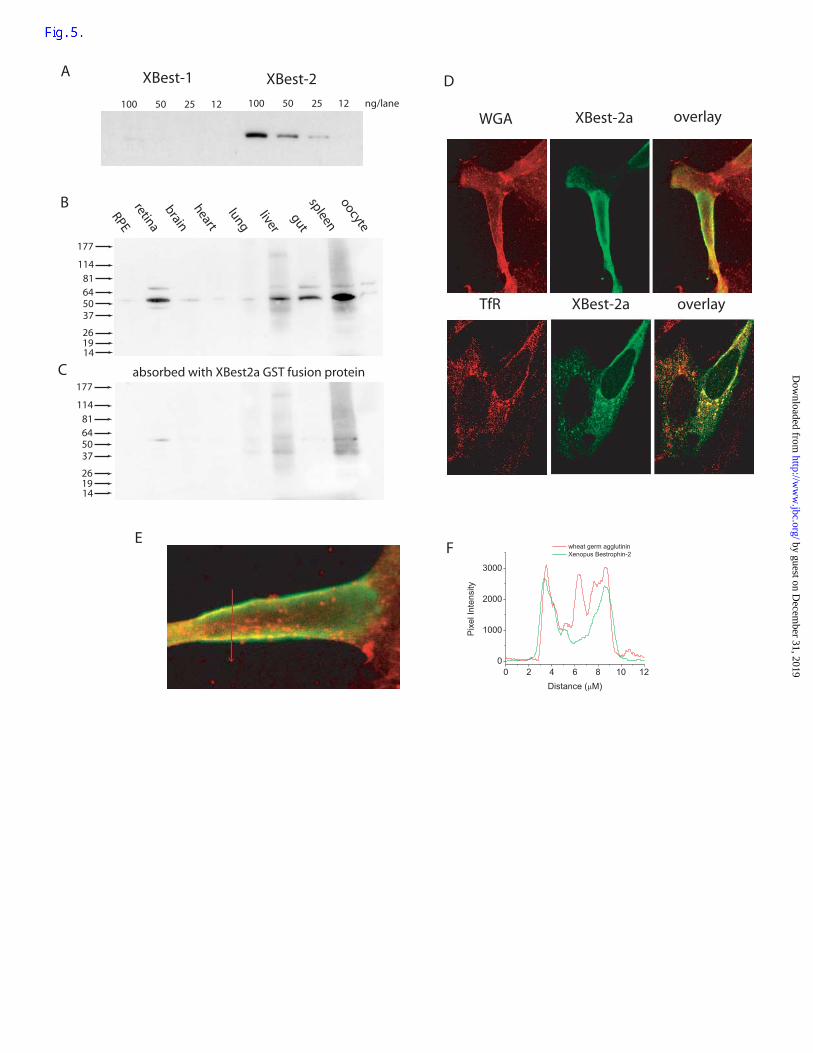
proteins of the C-terminus of xBest-2a but not xBest-2b (Fig. 5A). Western blots with
various Xenopus tissues showed that the A5925 antibody labeled a strong 54kD band in
neural retina, liver, gut, and spleen (Fig. 5B). A faint 54kD band was also observed in brain,
heart, lung, and oocyte. No band was observed in RPE. The 54kD band was absent when the
antibody was pre-absorbed with the GST-xBest-2a fusion protein (Fig. 5C). The mass of
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

16
XBest-2a predicted from the cDNA sequence is 59kD. We presume that the 54kD band
corresponds to the predicted 59kD protein.
Expression in HEK cells. xBest2a was expressed in HEK293 cells by transient
transfection. We observed two patterns of localization by immunocytochemistry. In some
cells, xBest-2a was concentrated at and near the cell surface (Fig. 5D-F). Figure 5D (top)
compares the distribution of XBest2a (green) with the distribution of wheat germ agglutinin
(red). Wheat germ agglutinin (WGA) is seen as a sharp band at the cell surface and in
intracellular compartments. XBest2a is also located at the cell surface and relatively little
intracellular staining is observed. A plot of pixel intensity across the cell shows that WGA
and xBest2a staining coincide very closely at the cell surface (Fig. 5E-F). These results
suggest that xBest2a can be expressed as a plasma membrane protein. In contrast, in other
cells, immunoflourescent staining was concentrated in intracellular organelles that were
identified as early endosomes by co-staining with an antibody to transferrin receptor (Figure
5D, bottom). These cells usually had patches of fluorescence at or near the plasma membrane
as well, but the staining was often not uniform or light. We do not understand why the
distribution appears to be different in different cells. Usually cells with pronounced
endosomal staining were flat, large cells.
Ionic currents of heterologously expressed XBest2a currents.
xBest-2a was transfected into cells along with a vector encoding for GFP expression.
Cells expressing xBest-2a were identified by GFP fluorescence. Whole-cell patch clamp of
xBest2a-expressing cells revealed that these cells had a Ca-dependent Cl current that was not
present in cells expressing GFP alone. Cells were patch-clamped in the whole cell
configuration with solutions having the same [Cl] inside and outside. The major intracellular
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

17
cation was Cs and the major extracellular cation was Na. Under these conditions with
intracellular free [Ca] buffered to <10 nM with 5 mM EGTA and 1 mM BAPTA, the currents
were small. In contrast, when free [Ca] was raised to 164 nM, currents were ~5-fold larger.
Fig. 6A shows the averages for one of several experiments. For both xBest-2a and xBest-2b
expressing cells, whole-cell currents with Ca <10 nM were <400 pA. However, when
intracellular free [Ca] was raised to 164 nM, currents ~1.5 nA were observed. In cells
expressing GFP alone, the currents were <200 pA.
We were concerned that expression of xBest-2 might up-regulate endogenous
swelling-activated currents in the HEK-293 cells. One set of experiments was performed in
cells bathed in 20 µM tamoxifen to block endogenous swelling-activated Cl currents (30).
Although the currents were smaller in the presence of tamoxifen, Ca stimulated the currents
~6-fold (Fig. 6B).
Figure 6 C-E shows current traces and current-voltage relationships from xBest-2a
expressing cells patched with different intracellular free [Ca]. With [Ca] < 10 nM, currents
were very small (Fig. 6C). In contrast, with 400 nM free Ca, large time- and voltage-
independent currents were observed (Fig. 6D). The currents were smaller at intermediate
[Ca], but the waveforms of the currents were virtually the same. The current-voltage
relationships for currents recorded with different intracellular free [Ca] are shown in Fig. 6E.
The current-voltage relationships were linear for all [Ca] and reverse near 0 mV as expected
for a Cl-selective current. A plot of current amplitudes vs. [Ca] shows that the EC50 for Ca is
210 nM (Fig. 6F).
The currents recorded in cells expressing xBest-2b were very similar (Fig. 7). The
calculated EC50 for xBest-2b was 228 nM.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

18
Because the xBest currents exhibited a reversal potential near 0 mV, which was the
same as the calculated ECl, but was very different from the calculated equilibrium potentials
of the major cations, Cs and Na, we concluded that the Ca-activated current was a Cl current.
To confirm that the current was Cl-selective, extracellular Cl was replaced with I, Br, or
aspartate and the shift in Erev was measured. Fig. 8 shows that the reversal potential shifted to
the left for I and Br, but shifted to the right for aspartate. These results suggest that the
channel has a relative permeability ratio of I > Br > Cl >> Aspartate.
W93C and G229E are dominant negative mutations.
To test whether the Cl currents that appeared when we over-expressed XBest-2a were
in fact mediated by this protein and were not caused by up-regulation of an endogenous
current, we expressed two XBest-2a mutants. Both mutations (W93C and G299E) were
selected because these mutations in human Best-1 produce dominantly-inherited Best
vitelliform macular dystrophy (20). The W93C construct expressed alone produced
negligible Ca-activated current (Fig. 9A, triangles). Furthermore, co-expression of W93C and
wild-type XBest-2a produced Ca-dependent currents that were 50% reduced in amplitude
compared to those with wild-type XBest-2a alone (Fig. 9A, circles). These data suggest that
the W93C mutation is able to reduce the function of wild type protein. A more dramatic
result was obtained with the G299E mutation. Expression of G299E alone produced no Ca-
dependent current (Fig. 9B, circles). Expression of G299E with wild-type suppressed the
wild-type current more than 90% (Fig. 9B, triangles). These data strongly suggest that xBest-
2a is a subunit of a Ca-activated Cl channel.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

19
Discussion
We have shown that Xenopus bestrophins produce Ca-dependent Cl currents when
they are expressed heterologously in HEK-293 cells. The bestrophin proteins are trafficked to
the plasma membrane and produce currents that are anion-selective with a permeability
sequence of I > Br > Cl >> aspartate. The currents are activated by physiological
concentrations of cytosolic Ca (EC50 = ~210 nM). Two bestrophin mutations, W93C and
G299E, produce non-functional channels that exert a dominant negative effect on the
expression of wild type channels. On this basis, we conclude that Xenopus bestrophins are
good candidates for a molecular component of Ca-dependent Cl channels. In contrast,
currents induced by the CLCA family are stimulated only by millimolar concentrations of Ca,
and to date there have been no mutagenesis studies demonstrating an effect of point
mutations on CLCA induced currents.
The currents induced by heterologous expression of xBest-2a are similar to the native
Ca-activated Cl currents in Xenopus oocytes (10;31). Both currents have EC50’s for Ca in the
range of 0.2 to 1 µM and exhibit similar anion permeability sequences (permeability ratios
for I : Br : Cl : aspartate are 4 : 2 : 1 : 0.7 for oocyte and 2.6 : 1.7 : 1 : 0.15 for xBest-2a). At
saturating [Ca], both currents are time-and voltage independent. However, there is a notable
difference between the native current and the bestrophin current at [Ca] near the EC50. The
native current has a voltage-dependent Ca affinity such that at low [Ca], the current exhibits
voltage-dependent gating behavior (31). This is not seen with the bestrophin current.
However, Ca-activated Cl currents reported in the literature differ significantly in their
voltage dependent characteristics. This suggests the possibility that, like other channels,
bestrophin channel behavior might be modulated by other subunits.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

20
Resemblance to GABA(A) receptor.
We identified bestrophin as a potential Ca-activated Cl channel by its similarity to the
pore domain of LGACs. The rationale was based on our previous studies that the native Ca-
activated Cl channel in Xenopus oocytes had very similar permeation properties to GABA(A)
and glycine receptors in terms of anionic selectivity and estimated pore dimensions and
orientation (10;12). The M2 domain of LGACs is related to what we believe is the 4th
transmembrane domain of bestrophin. We call this region the RFP domain.
Although the RFP and M2 domains have important structural similarities, this does
not imply that there is any evolutionary relationship between LGACs and bestrophins. There
is no significant homology between these proteins in other regions and, according to our
model, there are 6 transmembrane domains in bestrophins compared to 4 in LGACs.
Furthermore, the homology between the RFP domain and the M2 domain are seen only when
the sequences are compared in reverse orientation (the N-to-C terminal orientation of
bestrophins is compared to the C-to-N orientation of the GABA(A) receptor in Fig. 1).
Our model of the topology of bestrophin differs from models suggested by others.
Bakall et al. (32) and Sun et al. (21) suggested 4 transmembrane domains that correspond to
transmembrane domains 1, 3, 5 and 6 in our model. The protein is proposed to have
cytoplasmic N- and C-termini. White et al. (33) proposed 5 transmembrane domains that
correspond to our transmembrane domains 2, 3, 5, and 6 plus a domain that we believe is in
the cytoplasmic loop between transmembrane domains 4 and 5. They propose that the N-
terminus is cytoplasmic and the C-terminus is extracellular. The models by Bakall (34), Sun
et al. (21), and White et al. (33) differ from our model in that their models place the RFP
domain (our 4th transmembrane domain) in a cytoplasmic loop. However, the high degree of
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

21
sequence conservation in the RFP domain, its hydrophobicity, and its similarity to the M2
domain of LGACs suggests that this region is involved in forming the pore. The importance
of the RFP sequence is underlined by the fact that alanine mutations of the RFP sequence
result in non-functional channels (unpublished). Like the signature PAR sequence of LGACs,
the RFP sequence is located at the cytoplasmic end of a transmembrane helix by several
topology prediction algorithms.
In addition to the RFP – M2 similarity, another feature of bestrophins that attracted
our attention to thinking that they might be Ca-activated Cl channels was the highly
conserved acidic domain immediately following the last transmembrane domain. Although
this domain is only weakly related to known Ca binding domains, its acidic nature suggested
to us that it might be involved in Ca binding.
Tissue expression.
By semi-quantitative PCR, it is very difficult to distinguish between xBest-2a and
xBest-2b because of their high homology. We were unable to design primers for real-time
PCR that were specific for the different orthologs. Message for xBest-2a and.or xBest-2b is
widely expressed. The tissues that expressed message most robustly were spleen, liver, RPE,
neural retina, and lung. The A5925 antibody which is specific for xBest-2a, shows that
xBest-2a protein was expressed in spleen, gut, liver, and neural retina. Faint bands were
observed in brain, heart, and lung. Thus, spleen, liver, and neural retina have high levels of
both mRNA and protein, whereas brain and heart have much less. In contrast, in RPE the
level of mRNA is high but the level of protein is low. This could be explained if the mRNA
in RPE was primarily xBest-2b. Because human bestrophin-1 is expressed almost exclusively
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

22
in RPE, one might hypothesize that xBest-2b is playing the functional role in Xenopus RPE
that bestrophin-1 plays in human.
Bestrophin function.
Best vitelliform macular dystrophy is an early onset form of macular degeneration
that has been linked to over 80 different mutations in the human VMD2 gene that codes for
bestrophin-1 (33;35). No other diseases have been reported to be linked to VMD2 mutations.
These mutations tend to cluster, according to our model, in the 1st and 3rd transmembrane
domains, near the acidic cluster following the last transmembrane domain, and in the second
cytoplasmic loop. No disease-associated mutations have been reported in the C-terminal third
of the protein or in the RFP domain. Perhaps mutations in the RFP domain produce a more
serious defect that has not yet been identified. Many of the amino acids that when mutated
cause Best disease are conserved in in the Xenopus bestrophins. Because Best disease is an
autosomal dominant disease, we tested the effect of two Best disease mutations on
homologous residues in xBest-2a. Both the W93C and G299E mutations produce non-
functional channels and reduce the currents produced by wild type xBest-2a. Not only do
these results bolster our conclusion that bestrophins are channels, they also suggest that the
channels form multimeric complexes and that the mutant channels have a dominant negative
effect on channel function. How dysfunction of bestrophin channels may produce macular
degeneration remains to be seen. The finding that human bestrophin-1 is located in retinal
pigment epithelial cells has lead to the suggestion that bestrophin may be the molecular
counterpart of the basolateral Cl channel in retinal pigment epithelial cells. Although we have
tried to find a bestrophin in Xenopus that is more closely related to human bestrophin-1 than
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

23
xBest-2a or xBest-2b, we have not succeeded. Our semi-quanititative PCR and western blot
results suggest that in Xenopus xBest-2b may play the same role as human bestrophin-1.
The high level of xBest-2a expression in neural retina suggests that this bestrophin may be a
component of Ca-activated Cl channels in photoreceptors or retinal neurons. In many species,
both rods and cones have prominent Ca-activated Cl currents (36-39). These currents have
properties that are similar to the bestrophin currents we have described here. It has been
suggested that the Ca-activated Cl current in cones plays a role in modulating the voltage
gain of the photoresponse and in mediating the effect of horizontal cell feedback (40). In
rods, Ca activated Cl currents play a role in shaping and modulating the light response.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

24
Acknowledgements
Supported by NIH grant GM-60448 to HCH.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

25
References
1. Fuller, C. M. (2002) Calcium-activated Chloride Channels, Academic Press, San Diego
2. Cunningham, S. A., Awayda, M. S., Bubien, J. K, Ismailov, I. I., Arrate, M. P., Berdiev, B. K., Benos, D. J., and Fuller, C. M. (1995) J.Biol.Chem. 270, 31016-31026.
3. Gruber, A. D., Elble, R. C., Ji, H.-L., Schreur, K. D., Fuller, C. M., and Pauli, B. U.
(1998) Genomics 54, 200-214
4. Gruber, A. D., Schreur, K. D., Ji, H-L., Fuller, C. M., and Pauli, B. U. (1999) Am.J.Physiol 276, C1261-C1270.
5. Loewen, M. E., Bekar, L. K., Gabriel, S. E., Walz, W., and Forsyth, G. W. (2002)
Biochemical & Biophysical Research Communications 298, 531-536
6. Loewen, M. E., Gabriel, S. E., and Forsyth, G. W. (2002) Am.J.Physiol.Cell Physiol. 283, C412-C421
7. Romio, L., Musante, L., Cinti, R., Seri, M., Moran, O., Zegarra-Moran, O., and Galietta, L. J. (1999) Gene 228, 181-188
8. Jentsch, T. J., Stein, V., Weinreich, F., and Zdebik, A. A. (2002) Physiol.Rev. 82, 503-568
9. Papassotiriou, J., Eggermont, J., Droogmans, G., and Nilius, B. (2001) Pflugers Archiv. 442, 273-279
10. Qu, Z. and Hartzell, H. C. (2000) J.Gen.Physiol. 116, 825-844
11. Qu, Z. and Hartzell, H. C. (2001) J.Biol.Chem. 276, 18423-18429
12. Machaca, K., Qu, Z., Kuruma, A., Hartzell, H. C., and McCarty, N. (2002) In Fuller, C. M., editor. Calcium-activated Chloride Channels, Academic Press, San Diego
13. Lester, H. A. (1992) Annual Review of Biophysics & Biomolecular Structure 21, 267-292
14. Karlin, A. and Akabas, M. H. (1995) Neuron 15, 1231-1244
15. Langosch, D., Laube, B., Rundstrom, N., Schmieden, V., Bormann, J., and Betz, H. (1994) EMBO J. 13, 4223-4228
16. Xu, M. and Akabas, M. H. (1996) J.Gen.Physiol. 107, 195-205
17. Keramidas, A., Moorhouse, A. J., French, C. R., Schofield, P. R., and Barry, P. H.. (2000) Biophys. J. 78, 247-259.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

26
18. Keramidas, A., Moorhouse, A. J., Pierce, K. D., Schofield, P. R., and Barry, P. H. (2002) J.Gen.Physiol. 119, 393-410
19. Keramidas, A., Moorhouse, A. J., Pierce, K. D., Schofield, P. R., and Barry, P. H. (2002) J.Gen.Physiol. 119, 393-410
20. Petrukhin, K., Koisti, M. J., Bakall, B., Li, W., Xie, G., Marknell, T., Sandgren, O., Forsman, K., Holmgren, G., Andreasson, S., Vujic, M., Bergen, A. A. B., McGarty-Dugan, V., Figueroa, D., Austin, C. P., Metzker, M. L., Caskey, C. T., and Wadelius, C. (1998) Nature Genetics 19, 241-247
21. Sun, H., Tsunenari, T., Yau, K.-W., and Nathans, J. (2002) Proceedings of the National Academy of Sciences 99, 4008-4013
22. Francois, J., De Rouck, A., and Fernandez-Sasso, D. (1967) Archives of Ophthalmology 77, 726-733
23. Deutman, A. F. (1969) Archives of Ophthalmology 81, 305-316
24. Gallemore, R. P., Hughes, B. A., and Miller, S. S. (1998) In Marmor, M. F. and Wolfensberger, T. J., editors. The Retinal Pigment Epithelium, Oxford University Press, Oxford
25. Marmorstein, A. D., Mormorstein, L. Y., Rayborn, M., Wang, X., Hollyfield, J. G., and Petrukhin, K. (2000) Proceedings of the National Academy of Sciences 97, 12758-12763
26. Tsien, R. Y. and Pozzan, T. (1989) Meth.Enzymol. 172, 230-262
27. Hille, B. (1992) Ion Channels of Excitable Membranes, 2nd Ed., Sinauer Associates, Inc., Sunderland, MA
28. Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990) J.Mol.Biol. 215, 403-410
29. Keramidas, A., Moorhouse, A. J., Pierce, K. D., Schofield, P. R., and Barry, P. H. (2002) J.Gen.Physiol. 119, 393-410
30. Nilius, B., Eggermont, J., Voets, T., Buyse, G., Manolopoulos, V., and Droogmans, G. (1997) Progress in Biophysics.& Molecular Biology 68, 69-119
31. Kuruma, A. and Hartzell, H. C. (2000). J.Gen.Physiol. 115, 59-80.. 32. Bakall, B., Marknell, T., Ingvast, S., Koisti, M. J., Sandgren, O., Li, W., Bergen, A. A.,
Andreasson, S., Rosenberg, T., Petrukhin, K., and Wadelius, C. (1999) Hum.Genet. 104, 383-389
33. White, K., Marquardt, A., and Weber, B. H. F. (2000) Hum.Mutat. 15, 301-308.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

27
34. Bakall, B., Marknell, T., Ingvast, S., Koisti, M. J., Sandgren, O., Li, W., Bergen, A. A., Andreasson, S., Rosenberg, T., Petrukhin, K., and Wadelius, C. (1999) Hum.Genet. 104, 383-389
35. Weber, B. H. F. and Krämer, F. (2002) VMD2 Mutation Database. http://www.uni-wuerzburg.de/humangenetics/vmd2.html .
36. Bader, C. R., Bertrand, D., and Schwartz, E. A. (1982) J.Physiol. 331, 253-284
37. Barnes, S. and Hille, B. (1989) J.Gen.Physiol. 94, 719-743
38. Yagi, T. and Macleish, P. R. (1994) J.Neurophysiol. 71, 656-665
39. Morgans, C. W., El Far, O., Berntson, A., Wassle, H., and Taylor, W. R. (1998) J.Neurosci. 18, 2467-2474
40. Barnes, S. and Hille, B. (1989) J.Gen.Physiol. 94, 719-743
41. Hirokawa, T., Seah, B. C., and Mitaku, S. (1998) Bioinformatics 14, 378-379. 42. Kyte, J. and Doolittle, R. F. (1982) J.Mol.Biol. 157, 105-132 by guest on D
ecember 31, 2019
http://ww
w.jbc.org/
Dow
nloaded from

28
Footnotes
Abbreviations: LGAC: ligand gated anion channels, CFTR:, cystic fibrosis
transmembrane conductance regulator, RPE: retinal pigment epithelium, PCR: polymerase
chain reaction, GST: glutathione S-transferase, XBest-2a: Xenpous bestrophin-2a
(GENBANK AY273825), XBest-2b: Xenbopus bestrophin-2b (GENBANK AY273826).
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

29
Figure Legends
Figure 1. Comparison of human GABA(A) receptor α1 subunit M2 domain with the
bestrophin RFP domain. The M2 domain of the human GABA(A)α1 subunit is shown from
residue 301 – 277. The intermediate positively charged ring at R282 is labeled 0’. (17). The
outer positively charged ring is R301 at position 19’ and the negatively charged inner ring is
E277 at position -5’. The bestrophins are shown from residue 130 – 155. Identical residues
are highlighted in red, similar residues in orange, and weakly similar residues in blue.
Figure 2. Alignment of human and Xenopus bestrophins. Identical residues are shaded.
Transmembrane domains predicted by the SOSUI algorithm (41) are indicated by horizontal
lines. Consensus sequences for phosphorylation are highlighted in green for PKA, red for
PKC, and blue for casein kinase II. The signature RFP residues are cyan. The last 44 residues
of hBest-1 are not shown. Genbank accession numbers are xBest-2a: AY273825, xBest-2b:
AY2273826, hBest-1: NM_004183, hBest-2: NM_017682, hBest-3: NM_017682, hBest-4:
NM_152439.
Figure 3. Properties of bestrophins. (A) Cladogram of vertebrate bestrophins. The
relationships between Xenopus (x), human (h), mouse (m), Fugu (f), rat (r), and zebrafish (z)
bestrophins were determined using the method of Saitou and Nei (1987). Accession numbers
as in Figure 2, except that Fugu sequences were obtained from the Fugu Genomics project
assembly (fBest1 is SINFRUP00000141703, fBest2 is SINFRUP00000151123, and fBest-3
is SINFRUP00000134584). The Zebrafish bestrophins were cloned in our lab (unpublished).
(B). Hydropathy analysis of xBest-2a by the Kyte-Doolittle algorithm (42) with a window of
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

30
17 residues shows 6 distinct hydrophobic domains. This finding, coupled with the results of
the SOSUI transmembrane prediction shown in Fig. 2, suggests 6 transmembrane domains.
(C). Proposed topology of xBest-2a. Transmembrane domains are shaded in yellow.
Positively charged residues are red, negatively charged residues are blue, and proline is
orange. Note the location of the RFP sequence at the cytoplasmic end of transmembrane
helix 4.
Figure 4. Quantitative PCR. The amount of bestrophin-2 message in different Xenopus
tissues was quantified by real time PCR. The amount of bestrophin-2 message was compared
to the amount of GAPDH message as described in Methods.
Figure 5. Expression of bestrophin proteins in Xenopus tissues. (A). Specificity of antibody
A5925. Different amounts of xBest-2b and xBest-2a GST fusion proteins (12-100 ng per
well) were run on SDS-PAGE, blotted onto nitrocellulose, probed with affinity-purified
A5925 antibody (90 ng/ml), visualized with peroxidase-conjugated goat anti-rabbit
secondary antibody by chemi-luminescence. (B). Various Xenopus tissues were
homogenized, dissolved in SDS buffer (methods), run on SDS-PAGE, blotted onto
nitrocellulose and probed with affinity-purified A5925 antibody. (C). A companion blot was
probed with primary antibody (A5925) that had been incubated 2 hours with
electrophoretically-purified xBest-2a GST fusion protein. The blots in B and C were treated
in parallel identically with the exception of the primary antibody absorption step. (D).
Localization of xBest-2a in HEK-293 cells. xBest-2a in pCMV-SPORT6 was transfected into
HEK-293 cells. 1-2 days after transfection, the cells were fixed permeabilized, and stained
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

31
with antibody A5925 against xBest-2a visualized with Alexa-488 conjugated second
antibody (green). In the top row, cells were also stained with tetramethylrhodamine-
conjugated wheat-germ agglutinin (WGA, red left column), xBest-2 staining (green, middle
column), and an overlay of the two stains (right column). In the bottom row, the cells were
stained with A5925 against XBest-2a and a monoclonal mouse antibody against human
transferrin receptor (TfR) followed by rhodamine-goat-anti-mouse secondary antibody. E.
The distribution of xBest-2a at the membrane was determined by comparing red and green
fluorescence along a line drawn across a cell. F. Distribution of WGA and xBest-2a
associated fluorescence along the line across the cell in E.
Figure 6. Currents in xBest-2 expressing HEK-293 cells. xBest-2a or xBest-2b was
transfected into HEK-293 cells as described in methods. 1-2 days after transfection, the cells
were whole-cell patch clamped. (A). Amplitude of steady-state currents in transfected cells.
The steady-state currents during a 1-sec voltage pulse to +100 mV was measured in cells
either transfected with (a) a GFP expression vector (pIRES2-EGFP), (b) xBest-2a in pCMV-
SPORT6 plus the GFP-expression vector at a ratio of 10 : 1, or (c) the bi-cistonic construct
pIRES2-EGFP encoding for xBest-2b and GFP. The cells were patched with a pipet solution
containing either <10 nM free Ca (open bars), or 164 nM free Ca (hatched bars). (B).
Steady-state currents in the presence of 20 µM tamoxifen to block swelling-activated
currents. Transfections with GFP or xBest-2a plus GFP were performed and cells were
patched in the presence of 20 µM tamoxifen. Internal solution contained either <10 nM free
Ca or 400 nM free Ca as indicated. (C) Current traces with <10 nM free Ca. Voltage
protrocol is shown above current traces. (D). Current traces with 400 nM free Ca. (E)
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

32
Current-voltage relationships obtained with different free Ca concentrations. Steady state
current at the beginning of the 350 msec pulse is plotted vs. membrane potential. squares:
zero Ca, circles: 130 nM Ca, up triangles: 180 nM Ca, down triangles: 250 nM Ca,
diamonds: 400 nM Ca, side triangles: 4.5 µM Ca. (F) Dependence of the current on
intracellular [Ca]. The amplitude of the average current at +100 mV was plotted vs. [Ca]. n=
5-10 for each [Ca].
Figure 7. Currents in xBest-2b expressing cells. The conditions were the same as in Fig. 8,
except that the cells were transfected with pIRES-GFP vector encoding xBest-2b. A. Current
traces with <10 nM free Ca. (B) Current traces with 400 nM free Ca. (C) Current-voltage
relationships of selected [Ca]. Square: zero Ca, circle: 180 nM Ca, up triangle: 250 nM Ca,
down triangle: 4 µM Ca. (D). Dependence of the current on intracellular [Ca]. n= 3-10 for
each [Ca].
Figure 8. Ionic selectivity of the current induced by xBest-2a. HEK-293 cells expressing
xBest-2a were patched in whole cell configuration. Extracellular Cl was replaced with either
I, Br, or aspartate. (A) Current-voltage relationships. The current-voltage relationships were
obtained as shown in Fig. 7 with I (up triangles), Br (down triangles), Cl (squares), or
asparate (circles). The reversal potentials shifted as expected for a Cl-selective channel with
pI>pBr>pCl>>pAspartate. B. The relative permeabilities were calculated from the GHK
equation:
Px / PCl = [Cl]i / ([X]o exp (∆ErevF/RT))-[Cl] o / [X] o.
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

33
Figure 9. W93C and G299E are dominant negative mutations. Open symbols: zero Ca pipet
solution. Solid symbols: High Ca (4.5 µM) pipet solution. (A) W93C mutation. HEK-293
cells were transfected with pIRES2-EGFP vector without bestrophin insert (down triangles),
wild type xBest-2a (solid squares), W93C xBest-2a (up triangles), and equal amounts of
W93C and wild type xBest-2a together (circles). (B) G299E mutation. HEK-293 cells were
transfected with wild type xBest-2a (squares), G299E xBest-2a (circles), and equal amounts
of wild type and G299E (triangles).
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

34
Table 1. Physical Properties of the M2 domain of GABA receptor and xBest-2a.
hGABA(A)R α1 xBest2a
Isoelectric point 9.6 10.9
acidic (D,E) 1 1
basic (K,R) 2 4
polar (N,C,Q,S,T,Y) 9 6
hydrophobic (A,I,L,F,W,V) 10 13
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

Fig.1 19’ 0’-2’ -5’ hGABAA (301) R-ASISLTTMTLVTTVGFVTRAPVSE hBest1 (130) RYANLGNVLILRSVSTAVYKRFPSAQ hBest2 (130) RYAGLSAVLILRSVSTAVFKRFPTID hBest3 (130) RYANLASVLVLRSVSTRVLKRFPTME hBest4 (130) RYVNLTSLLIFRSVSTAVYKRFPTMD xbest2 (130) RYCSLSGLLILRSVSTAVFKRFPTID + + P - OUTER INTERM INNER
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

Fig.2 hBest-1 1 MTITYTSQVA NARLGSFSRL LLCWRGSIYK LLYGEFLIFL LCYYIIRFIY RLALTEEQQL MFEKLTLYCD hBest-2 1 MTVTYTARVA NARFGGFSQL LLLWRGSIYK LLWRELLCFL GFYMALSAAY RFVLTEGQKR YFEKLVIYCD hBest-3 1 MTVSYTLKVA EARFGGFSGL LLRWRGSIYK LLYKEFLLFG ALYAVLSITY RLLLTQEQRY VYAQVARYCN hBest-4 1 MTVTYSSKVA NATFFGFHRL LLKWRGSIYK LLYREFIVFA VLYTAISLVY RLLLTGVQKR YFEKLSIYCD XBest-2b 1 MTVTYTARVA NARFGGFYKL LLLWRGSIYK LLYKELLAFF IMYLALSITY RFFLNEEQRL YFEKVAIYCN XBest-2a 1 MTVTYTARVA NARFGGFYKL LLLWRGSIYK LLYKEFLAFL IMYLGLSIIY RFLLNDEQRL YFEKVAIYCN hBest-1 71 SYIQLIPISF VLGFYVTLVV TRWWNQYENL PWPDRLMSLV SGFVEGKDEQ GRLLRRTLIR YANLGNVLIL hBest-2 71 QYASLIPVSF VLGFYVTLVV NRWWSQYLCM PLPDALMCVV AGTVHGRDDR GRLYRRTLMR YAGLSAVLIL hBest-3 71 RSADLIPLSF VLGFYVTLVV NRWWSQYTSI PLPDQLMCVI SASVHGVDQR GRLLRRTLIR YANLASVLVL hBest-4 71 RYAEQIPVTF VLGFYVTLVV NRWWNQFVNL PWPDRLMFLI SSSVHGSDEH GRLLRRTLMR YVNLTSLLIF XBest-2b 71 NYANLIPVSF VLGFYVTLVV SRWWNQYLSI PFPDRVMCAI SGTVHGSDEK GRLYRRTLMR YCSLSGLLIL XBest-2a 71 NYANLIPVSF VLGFYVTLVV NRWWNQYLCM PFPDRVMCAV SGTVHGSDET GRLYRRTLIR YCSLSGLLIL hBest-1 141 RSVSTAVYKR FPSAQHLVQA GFMTPAEHKQ LEKLSLPHNM FWVPWVWFAN LSMKAWLGGR IRDPILLQSL hBest-2 141 RSVSTAVFKR FPTIDHVVEA GFMTREERKK FENLNSSYNK YWVPCVWFSN LAAQARREGR IRDNSALKLL hBest-3 141 RSVSTRVLKR FPTMEHVVDA GFMSQEERKK FESLKSDFNK YWVPCVWFTN LAAQARRDGR IRDDIALCLL hBest-4 141 RSVSTAVYKR FPTMDHVVEA GFMTTDERKL FNHLKSPHLK YWVPFIWFGN LATKARNEGR IRDSVDLQSL XBest-2b 141 RSVSTAVFKR FPTIDHVVEA GFMTRLERKK FENLQSSYNK YWVPCVWFCN LAAQGRSEGR IRDDHSFKML XBest-2a 141 RSVSTAVFKR FPTIDHVVEA GFMTRLERKK FENLQSSYNK YWVPCVWFCN LAAQARSEGR IRDDHSFKML hBest-1 211 LNEMNTLRTQ CGHLYAYDWI SIPLVYTQVV TVAVYSFFLT CLVGRQFLNP AKAYPG---- ---------- hBest-2 211 LEELNVFRGK CGMLFHYDWI SVPLVYTQVV TIALYSYFLA CLIGRQFLDP AQGYKD---- ---------- hBest-3 211 LEELNKYRAK CSMLFHYDWI SIPLVYTQVV TIAVYSFFAL SLVGRQFVEP EAGAAKPQKL LKPGQEPAPA hBest-4 211 MTEMNRYRSW CSLLFGYDWV GIPLVYTQVV TLAVYTFFFA CLIGRQFLDP TKGYAG---- ---------- XBest-2b 211 MEELNTFRGN CSMLFHHDWI SVPLVYTQVV TIAVYSFFLT CLIGRQFLDP LRGYPG---- ---------- XBest-2a 211 MEELNTFRGN CGMLFHYDWI SIPLVYTQVV TIAVYSFFLT CLIGRQFLDP SRGYPG---- ---------- hBest-1 267 -HELDLVVPV FTFLQFFFYV GWLKVAEQLI NPFGEDDDDF ETNWIVDRNL QVSLLAVDEM HQDLPRMEPD hBest-2 267 -HDLDLCVPI FTLLQFFFYA GWLKVAEQLI NPFGEDDDDF ETNFLIDRNF QVSMLAVDEM YDDLAVLEKD hBest-3 282 LGDPDMYVPL TTLLQFFFYA GWLKVAEQII NPFGEDDDDF ETNQLIDRNL QVSLLSVDEM YQNLPPAEKD hBest-4 267 -HDLDLYIPI FTLLQFFFYA GWLKVAEQLI NPFGEDDDDF ETNWCIDRNL QVSLLAVDEM HMSLPKMKKD XBest-2b 267 -HELDLYIPV FTLLQFFFYA GWLKVGEQLI NPFGEDDDDF ETNFLIDRNF QVSMLAVDEM YSDVPPMEKD XBest-2a 267 -HELDLYIPV FTLLQFFFYA GWLKVGEQLI NPFGEDDDDF ETNFLIDRNF QVSMLAVDEM YSDVPPMEKD hBest-1 336 MYWNKPEPQP PYTAASAQFR -RASFMGSTF NISLNKEEME FQPNQ-E--D EEDAHAGIIG RFLGLQSHDH hBest-2 336 LYWDAAEARA PYTAATVFQL RQPSFQGSTF DITLAKEDMQ FQRLDGLDGP MGEAPGDFLQ RLLPAGAGMV hBest-3 351 QYWDEDQPQP PYTVATAAES LRPSFLGSTF NLRMSDDPEQ SLQVEAS--- -PGSGRPAPA AQTPLLGRFL hBest-4 336 IYWDDSAARP PYTLAAADYC -IPSFLGSTV QMGKQMP--- ---------K -NEWKMEDIK IPLPQPQFQC XBest-2b 336 RYWNHSDPRP PYTAATLFQK HLPSFQGSTF NMAIPKEDMQ FQPLS----D IEEMNEDTLT HTLPLFSRFP XBest-2a 336 RYWNHSDPRP PYTAATLFQK HLPSFQGSTF NMAIPKEDMQ FQPLS----D IEEMNEDTLM HPLPLLSRFL hBest-1 402 HPPRANSRTK LLWPKRESLL HEGLPKNHKA AKQNVRGQED NKAWKLKAVD AFKSAPLYQR PGYYSAPQTP hBest-2 406 AGGPLGRRLS FLLRKNSCVS EASTGASCSC AVVPEG---- ------AAPE CSCGDPLLDP --GLPEPEAP hBest-3 417 GVGAPSPAIS LRNFGRVRGT PRPPHLLRFR AEEGGD---- ------PEAA ARIEEESAES GDEALEP--- hBest-4 392 AKSDPGG--- ---------- ---------- ---------- ---------- ---------- ---------- XBest-2b 402 PGIGPSPLSS SAGLSTLFAA PGSRLTLLSH STSSFS---- ------SSSV FQFPETVQDP PNNLVDPTAP XBest-2a 402 PGVGPSPLSS SAALAAQFAA PENNFTLLRR STSAFS---- ------SSSE FQSQEPIQDP PYNLVDTKTP hBest-1 472 LSPTPMFFPL EPSAPSKLHS VTGIDTKDKS LKTVSSGAKK SFELLSESDG ALMEHPEVSQ VRRKTVEFNL +44 hBest-2 462 PPAGPEPLTL IPGPVEPFSI VTMPGPRGPA PPWLPSPIGE EEENLA---- ---------- ---------- hBest-3 ---------- ---------- ---------- ---------- ---------- ---------- ---------- hBest-4 ---------- ---------- ---------- ---------- ---------- ---------- ---------- XBest-2b 462 GLNVQEWPLE KLFSIGSQNS LFHPPKTMDG CGNVQSVMPE EAEDAASLVA T--------- ---------- XBest-2a 462 ELNVQECQIE KQFSVGSQVS LFLPPKTMDI DGNIQSVMPE EEEDTASLVA T--------- ---------- PKA PKC casein
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

C
A
MTV
V
V
E
EE
E
E
E
E
E EE
EE
EE
E
E
E
E
DDDD
DD
D
D
D
D
D
D
D
DD
DD D
D D
DD
EE E
E
E
E
EEE E
E
D
T
T
A
A
A
R
RR
Y
Y
R
N
F
F
GG
K
K
LL
LL
WR
GS
I Y K
LL
L
L
L
L
LL
L
L
L
L
Y
YY
Y
Y
Y
Y
II
I
V
V
F
NN
N
K
F
S
AF
F
Q C
I
AI
N
IP
SF
GF
V
VV
V
V
T
T
T
T
G
G
RN
M
WW L
L
L
LL
P
R
R
RRN
QYL
CMPF
V
VV
VV
V
SS
S
SS
S
S
S
S
S
S
P
P
P
P P
R
R
R
R
R
RR
GG
G
G
G
G
T
T
TH
MC
G
G
G
T
T
T
LY
YC
A
A
A
I
F
FFF
FF
K
KK
K
K
VV
VVED I
I
I L LL
LL
H
A
AA
A F
F
F
FF
R
N
N
NN
N Y
Y
Y
Q W
WW
P
RRC
C
Q
S
GR
H F
M
M F I
L
P S V
QF
I
L
P
ST
VQ
A
F I
L
P
S T
VQA
F
I
L
P
S T
V
Q A
F
I
L
M
N
P
S
T
V
QA
F
GI
K
L
M
N
P
S
TV
Q A
F
G
I
K
L
M
N
PS
T
V
Q
A
F
G
I
KL MN
P
R
ST
V
Q
A
F
G H
I
K
L
MNP
R
S
T
V
Y
Q
A
F
GH
I
K
L M
N
P RS
T
VY
Q
Y
A
C
D
F
G
H
I
K
L
M
N
P
R
S
T
V
Y
Q
Y
A
C
D
F
G
H
IK
L
MN
P
R
S
T
V
Y
W
Q
Y
Y
LL
LL
P
S
L
P
SF
LP
S
F
L P
S FL
P
S
F
L
N
P
SA
FL
M
N
P
S
TA
FH
L
M
N
P R
STV
Y
QC
L
T
W
L Q
LS
Q
L
S
Q
L
PS
Q
A
E
GL
MP
S
QL
AB
mBest-1rBest-1
hBest-1zBest-1
fBest-1mBest-4rBest-4
hBest-4fBest-2
zBest2xBest-2axBest-2b
hBest-2mBest-2
fBest-3hBest-3
Nucleotide Substitutions (x100)
70 60 50 40 30 20 10 0
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

RP
E
retin
a
brai
n
hear
t
lung
liver gu
t
sple
en
-4
-3
-2
Log
(xB
est2
/GA
PD
H)
Fig.4
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from
XBest-1 XBest-2
100 50 25 12 100 50 25 12 ng/lane
177
11481645037
261914
RPE
gut
liver
lung
heart
brain
retina
oocyte
spleen
177
11481645037
261914
absorbed with XBest2a GST fusion protein
A
B
C
D
WGA XBest-2a overlay
EF
TfR XBest-2a overlay
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

0 1 0 0 2 0 0 3 0 0- 3- 2- 1012
0 1 0 0 2 0 0 3 0 0- 3- 2- 1012
0 . 0 0 . 1 0 . 2 0 . 3 0 . 4 40
1
2
0 . 0
0 . 5
1 . 0
1 . 5I (n
A) 1 6 4 n M C a < 1 0 n M C a
G F P x B e s t - 2 b x B e s t - 2 a 0 . 00 . 20 . 40 . 60 . 8 T a m o x i f e n ( 2 0 µM )
X B e s t - 2 a4 0 0 n M C a
X B e s t - 2 a< 1 0 n M C a
G F P 4 0 0 n M C a
I (nA)
I (nA)
T i m e ( m s e c )
0 C a
A
FE
I (nA)
T i m e ( m s e c )
4 0 0 n M C a
B
C D
- 1 0 0 - 5 0 5 0 1 0 0
- 2
- 1
1
2 0 C a 1 3 0 n M C a 1 8 0 n M C a 2 5 0 n M C a 4 0 0 n M C a 4 . 5 µM C a
I (nA)
V m ( m V )
I (nA)
[ C a ] ( µM )
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

0 5 0 1 0 0 1 5 0 2 0 0 2 5 0 3 0 0 3 5 0- 2 . 0- 1 . 5- 1 . 0- 0 . 50 . 00 . 51 . 01 . 5
0 5 0 1 0 0 1 5 0 2 0 0 2 5 0 3 0 0 3 5 0- 2 . 0- 1 . 5- 1 . 0- 0 . 50 . 00 . 51 . 01 . 5
0 . 0 0 . 1 0 . 2 0 . 3 0 . 4 4
0 . 5
1 . 0
1 . 5
2 . 0
nA
[ C a ] ( µM )
- 1 0 0 - 5 0 5 0 1 0 0
- 1 . 5- 1 . 0- 0 . 5
0 . 51 . 01 . 5 nA
m V
0 C a 1 8 0 n M C a 2 5 0 n M C a 4 µM C a
A
B
C
D
I (nA)
T i m e ( m s e c )
4 0 0 n M C a
I (nA)
T i m e ( m s e c )
0 C a
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

-100 -50 50 100
-1
1
I Br Cl Asp0
1
2
3
B
pX /
pCl
A
Fig.8
Asp
Cl
Br
I
I (nA
)
Vm (mV)
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

-3
-2
-1
1
2
wt, 0Ca, n=5 wt, High Ca, n=8 G299E,High Ca, n=7 wt + G299E, High Ca, n=7
I (nA
)
Vm (mV)100
-100 -50
50
-3
-2
-1
1
2
Fig.9
EGFP, High Ca, n=5 wt, High Ca, n=6 W93C, High Ca, n=7 wt + W93C, High Ca, n=6 wt + W93C, 0 Ca, n=6
Vm (mV)
I (nA
)
10050
-50-100
A
B
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from

Zhiqiang Qu, Raymond W Wei, Wesley Mann and H. Criss HartzellTwo bestrophins cloned from Xenopus laevis Oocytes express Ca-activated Cl currents
published online August 25, 2003J. Biol. Chem.
10.1074/jbc.M308414200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on Decem
ber 31, 2019http://w
ww
.jbc.org/D
ownloaded from