trophic structure and dynamical complexity in simple ecological models
TRANSCRIPT

e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2
Trophic structure and dynamical complexity in simpleecological models
Vikas Rai d, Madhur Anand a,b,*, Ranjit Kumar Upadhyay c
aDepartment of Biology, Laurentian University, Ramsey Lake Road, Sudbury, Ontario, Canada P3E 2C6bDepartment of Environmental Biology, University of Guelph, Guelph, Ontario, Canada N1G 2W1cDepartment of Applied Mathematics, Indian School of Mines University, Dhanbad, Jharkhand 826004, IndiadDepartment of Applied Mathematics, HMR Institute of Technology and Management, GT Karnal Road, Delhi 110 036, India
a r t i c l e i n f o
Article history:
Received 2 April 2007
Received in revised form
27 June 2007
Accepted 27 June 2007
Published on line 13 August 2007
Keywords:
Predator–prey interaction
Generalist and specialist predators
Competition
Population cycles
Food web
Deterministic chaos
Robust chaos
Short-term recurrent chaos
a b s t r a c t
We study the dynamical complexity of five non-linear deterministic predator–prey model
systems. These simple systems were selected to represent a diversity of trophic structures
and ecological interactions in the real world while still preserving reasonable tractability.
We find that these systems can dramatically change attractor types, and the switching
among different attractors is dependent on system parameters. While dynamical complex-
ity depends on the nature (e.g., inter-specific competition versus predation) and degree (e.g.,
number of interacting components) of trophic structure present in the system, these
systems all evolve principally on intrinsically noisy limit cycles. Our results support the
common observation of cycling and rare observation of chaos in natural populations. Our
study also allows us to speculate on the functional role of specialist versus generalist
predators in food web modeling.
# 2007 Elsevier B.V. All rights reserved.
avai lable at www.sc iencedi rec t .com
journal homepage: ht tp : / /www.e lsev ier .com/ locate /ecocom
1. Introduction
Dynamical complexity refers to the variety of behaviors that
systems can exhibit over time. It can be studied through
construction of simple models, and then examination of
patterns in temporal evolution in response to changes in
model parameters. As such one can attempt to estimate the
likelihood of observing predictable (e.g., simple equilibrium,
cycles) versus unpredictable (chaotic) dynamics in phase
space and determine what causes abrupt changes between
these two types of behavior. The more likely the occurrence
of unpredictability and abrupt changes in system dynamics,
* Corresponding author at: Department of Environmental Biology, UniTel.: +1 519 824 4120; fax: +1 519 837 0442.
E-mail address: [email protected] (M. Anand).
1476-945X/$ – see front matter # 2007 Elsevier B.V. All rights reservedoi:10.1016/j.ecocom.2007.06.010
the more dynamically complex a system is understood
to be.
Typically, dynamical complexity can be studied by finding
the critical parameter values which cause changes in attractor
types. Take the classic single population dynamic model
studied by May (1975, 1976) and May and Oster (1976). In this
model, there is only one parameter to be varied, namely r, the
intrinsic growth rate. For some range of r values, dynamics
reaches a single equilibrium. For another range, the attractor
is cyclical. Once it reaches a critical point (r = 3.5), chaos is
observed. Because there is no interruption in the display of
chaotic behavior as r is increased further, we call this type of
versity of Guelph, Guelph, Ontario, Canada N1G 2W1.
d.

e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2 213
chaotic dynamics ‘‘robust’’. This behavior illustrates a very
important property: the fact that an extremely simple model
(e.g., a single population model) can display dynamical
complexity. However, as we shall see, the dynamical complex-
ity of this model is actually relatively low, when compared to
more structurally complex ecological models. This is because
it has a critical parameter value that clearly divides the phase
space into different attractor types (fixed points, stable limit
cycles, and chaos).
In this paper, we investigate the dynamical complexity of
predator–prey models. The simplest predator–prey model is the
Lotka–Volterra model (Lotka, 1920; Volterra, 1931). Because of
the existence of two interacting populations, one can argue that
this model is structurally more complex than the one studied by
May (1976). However, perhaps surprisingly, it does not display
greater dynamical complexity. In fact, one can argue that it
displays less dynamical complexity than the single population
dynamic model because there is no chaotic behavior. Several
studies, most of which build upon the work of Lotka, Volterra
and May, have however now reported robust deterministic
chaos in an appreciable range of a single critical parameter of
model predator–prey systems with three or more interacting
populations (Gilpin, 1979; Schaffer, 1985; Hastings and Powell,
1991; Rai and Sreenivasan, 1993; Rinaldi and Muratori, 1993;
Rinaldi et al., 1993; Vandermeer, 1993; Klebanoff and Hastings,
1994; King et al., 1996, 2004; Upadhyay and Rai, 1997; Edwards
and Bees, 2001; Vandermeer et al., 2001; Aziz-Alaoui, 2002). Very
few studies on predator-prey systems attempt to examine if the
chaotic dynamics is robust with respect to changes in system
parameters in a two-dimensional (2D) space. The study of
Samardzija and Greller (1988) is an exception. These authors
found that their model could exhibit chaotic dynamics in
surprising regions of parameter space due to a ‘‘fractal torus
structure’’, hence introducing the term ‘‘explosive chaos’’.
While their model included only three interacting populations,
they noted that the use of two predators and one prey was
distinctly different from previous models which included one
predator and two prey (e.g., Arneodo et al., 1980; Gilpin, 1979;
Schaffer, 1985), because these previous models all showed
robust chaos. Thus, we should expect that the structure of
models (not just the number of interacting populations) should
also affect dynamical complexity.
Generally speaking, we can expect that when there are
more parameters to be investigated, the dynamical complex-
ity of a model should increase. Indeed, while performing
simulation studies of a new class of predator–prey models,
Upadhyay et al. (1998) discovered that the presence of chaotic
dynamics is not robust to changes in paired system para-
meters. Namely, there were no large smooth regions in
parameter space with which one could identify a chaotic
phase. Similarly, Turchin and Ellner (2000) discovered an
interesting variety of chaos which they termed short-term
recurrent chaos (called strc hereafter). In this type of dynamical
complexity, chaotic behavior is interrupted by other kinds of
dynamics at irregular and unpredictable intervals. This
phenomenon also occurs in several other predator–prey
models (Upadhyay and Rai, 2001; Rai, 2004; Rai and Anand,
2004; Rai and Upadhyay, 2004).
In this paper, we examine a suite of predator–prey models
to attempt to shed light on when we should expect to see strc
versus robust chaos. We hypothesize that the reason why
chaos is rarely observed in natural populations might lie with
the structure of ecological systems (e.g., one versus two
predators) and the nature of ecological interactions (e.g.,
competition, mutualism and predation). We use some
published models, but also create new models by manipulat-
ing the trophic structure in simple coupled oscillators. As
such, we attempt to explore the diversity of interaction effects
on simple ecological systems.
2. Model systems
There are three ways to generate chaos in a mathematical
model: (1) by coupling two non-linear oscillators (Vandermeer,
1993, 2004; Rai, 2004), (2) by incorporating seasonal variation in
a model parameter (Rinaldi et al., 1993; Schaffer, 1988;
Upadhyay et al., 1998), and (3) by incorporating effect of space
in an oscillatory predator–prey system (Pascual, 1993; Med-
vinsky et al., 2002; Morozov et al., 2004; Petrovski et al., 2004).
In this paper, we examine scenario 1 only. This scenario is
appropriate for large interacting populations, which generate
well-mixed conditions.
We assume that ecologists generally accept a loose
classification of predators into specialists and generalists.
Two ecological principles that form the skeleton of the model
systems studied in the present paper are: (1) a specialist
predator population decays exponentially fast in the absence
of its prey and (2) per capita growth of a generalist predator is
limited by dependence on its favorite prey and severity of this
limitation is inversely proportional to per capita availability of
prey at any instant of time.
The Rosenzweig–MacArthur (RM) model (Rosenzweig and
MacArthur, 1963) describes the dynamics of a specialist
predator and its prey. The model is represented by following
coupled ordinary differential equations:
dXdt¼ a1X 1� X
K
� �� wYX
Xþ D(1a)
dY
dt¼ �a2Y þ w1YX
Xþ D1; (1b)
where X is prey for specialist predator Y with Holling type II
functional response, a1 is the rate of self-reproduction for the
prey, K measures the carrying capacity of the environment for
prey X, w is the maximum rate of per capita removal of prey
species X due to predation by its predator Y, D is that value of
population density of X at which per capita removal rate is half
of w, the parameter a2 measures how fast the predator Y will
die when there is no prey to capture, kill and eat; w1X=Xþ D1is
the per capita rate of gain in predator Y, w1 is its maximum
value and D1 denotes the population density of the prey at
which per capita gain per unit time in Y is half its maximum
value (w1). The assumptions underlying this formulation of
predator–prey interaction are as follows: the life histories of
each population involve continuous growth and overlapping
generations. The predator Y dies out exponentially in the
absence of its most favorite food X. The predator’s feeding
rate saturates at high prey densities. The age structure of both
the populations is ignored.

e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2214
A generalist predator differs from a specialist in that it does
not die out when its favorite food is absent or is in short
supply, but rather its population continues to grow because
individuals switch over to alternative food options when their
most preferred food is in short supply. A model known as
Holling–Tanner model (Holling, 1965; Tanner, 1975; Pielou,
1977; May, 2001) describes the dynamics of a generalist
predator and its prey:
dZdt¼ AZ 1� Z
K1
� �� w3UZ
Zþ D3(2a)
dUdt¼ cU�w4U2
Z; (2b)
where Z is the most favorite food for the generalist predator U,
A the per capita rate of self-reproduction of the predator, and
K1 is the carrying capacity of its environment. The parameter
w3 measures the maximum of the per capita rate of removal of
the prey population by its predator U. D3 is the half-saturation
constant for prey Z. c represents the per capita rate of self-
reproduction of the predatorU and w4 measures severity of the
limitation put to growth of predator population by per capita
availability of its prey. It can be seen that the generalist
predator U does not die out exponentially in the absence of
its favorite prey Z, instead, grows logistically to aZ, where
að¼ c=w4Þ is the proportionality constant. The underlying
assumption of this formulation is that the predator’s carrying
capacity is proportional to the population density of its
most favorite prey. This assumption is phenomenological
(P. Abrams, pers. commun., 2004). The positive aspect of this
formulation of predator–prey interaction is that it takes care of
our inability to write down growth equations for all the species
a generalist predator feeds on. This is simply because it is
practically impossible to identify all of them. Although this
formulation is too simplistic, yet it has positive features which
are not found in other models of predator–prey interaction
involving a generalist predator.
Graphical representation of the dynamical structure of the
Holling–Tanner model in a 2D parameter space was presented
by May (2001). There exists considerably large area in the
parameter space where stable limit cycle solutions occur (see
May, 2001, p. 192, Fig. A.1). Graphical representation of the
dynamics in a 2D parameter space of the Rosenzweig–
MacArthur model was presented in the recent work of Rai
(2004). It can be noted that the stable limit cycle solutions are
confined to a narrow area of the biologically relevant
parameter space (a2, K) (Fig. 1 in Rai, 2004; a1=b1 ¼ K) of RM
model. As is well known, a minimum of three degrees of
freedom are needed to generate chaos, thus the RM model
does not display chaotic dynamics. Rinaldi and colleagues
(Rinaldi and Muratori, 1993; Rinaldi et al., 1993) considered
sinusoidal variations in model parameters, which drove it into
chaos. Recently, Vandermeer et al. (2001) studied phase
locking in a seasonally forced RM model. Another ecologically
sound way of generating chaos would be to couple two
systems dynamically. When parameters are chosen in such a
way that both RM and HT systems display regular persistent
periodic oscillations, these systems behave as oscillators.
When suitably coupled, these systems force each other to
generate chaos (Rai, 2004; Rai and Anand, 2004; Rai and
Upadhyay, 2004). We have studied some of the models that
can be obtained by coupling two subsystems. We found that
the nature of ecological interactions affect the observability of
chaos, however, the models that we have examined so far
have not led to generalizations about these interactions. This
is what we study in the present paper.
2.1. Model 1: one generalist, one specialist, two prey
Consider an ecological community with both generalist and
specialist predators. This leads to perhaps the most
straightforward coupling of RM and HT models. In this,
the only effect of the introduction of the generalist predator
(and a second prey species) is that the specialist predator
population is reduced because of the presence of the
generalist predator. This model is given by the following
equations:
dXdt¼ a1X 1� X
K
� �� wYX
Xþ D(3a)
dY
dt¼ �a2Y þ w1YX
Xþ D1� w2Y2U
Y2 þ D22 þ Z
(3b)
dZ
dt¼ AZ 1� Z
K1
� �� w3UZ
Zþ D3(3c)
dUdt¼ cU� w4U2
Y þ Z; (3d)
where parameter a1 is the rate of self-reproduction andK is the
carrying capacity of prey X. w=ðXþ DÞ is the per capita rate of
removal of prey X by specialist predator Y. D is a measure of
protection provided by the environment to prey X. Y is a
specialist predator, i.e. X is the only food for it. Therefore, Y
dies out exponentially in the absence of X. w is the maximum
value that function wX=ðXþ DÞ can attain. a2 is the rate at
which Y dies out exponentially in the absence of its prey X.
w1X=ðXþ D1Þ denotes the per capita gain in the specialist
predator population due to proportionate loss in its prey. w1
is the maximum value that this function can take. D1 is the
half-saturation constant for specialist predator Y. A and K1 are
respectively the rate of self-reproduction and carrying capa-
city for prey Z. The last term in Eq. (3b) represents the
functional response of the generalist predator U. It switches
its prey whenever its favorite prey Z is in short supply. The
last term in Eq. (3d) describes how loss in abundance of
generalist predator U depends on per capita availability of
its prey (Z and Y).
It can be easily seen that this model has been obtained by
coupling the RM model with HT model via the term
ð�w2Y2U=ðY2 þ D2 þ ZÞÞ. This is the Holling type III functional
response term with Z added to the denominator to signify the
fact that the generalist predator switches to Y whenever it is
difficult to find the favorite food (Z). Rai (2004) has studied the
model with no Z in the denominator of the coupling term. At
this point, it should be noted that the older version of the
model is justified if the generalist predator switches its prey
only in extreme situations, for example, if there exist
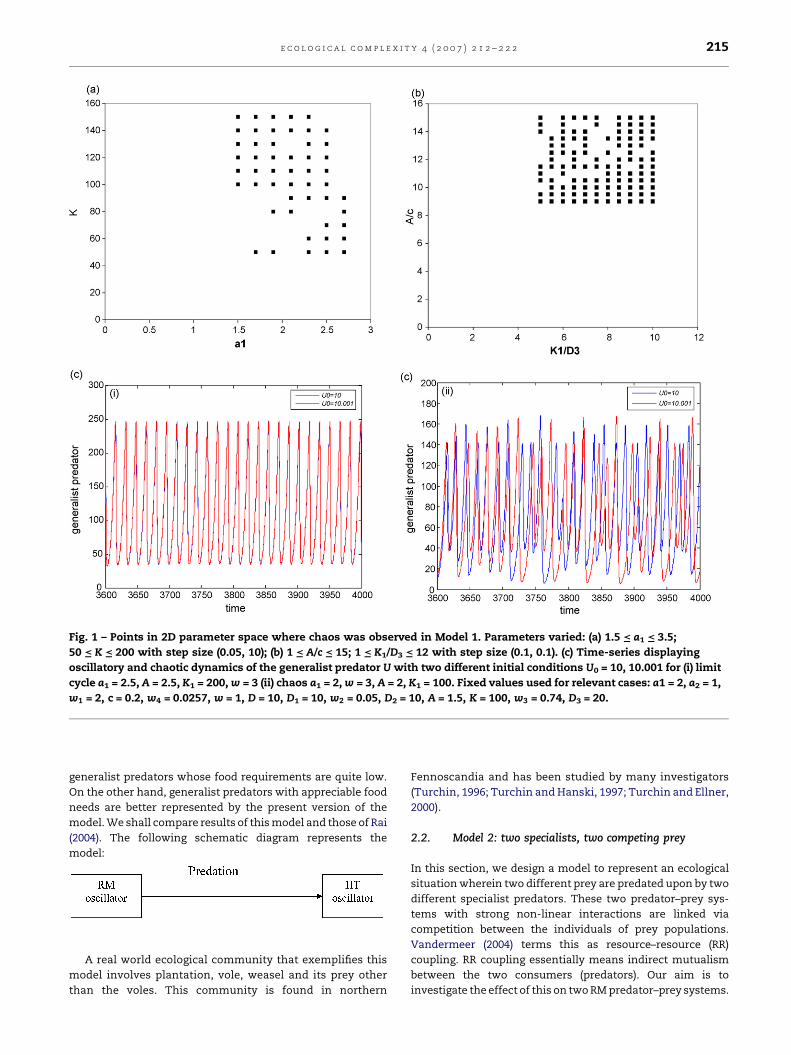
Fig. 1 – Points in 2D parameter space where chaos was observed in Model 1. Parameters varied: (a) 1.5 = a1 = 3.5;
50 = K = 200 with step size (0.05, 10); (b) 1 = A/c = 15; 1 = K1/D3 = 12 with step size (0.1, 0.1). (c) Time-series displaying
oscillatory and chaotic dynamics of the generalist predator U with two different initial conditions U0 = 10, 10.001 for (i) limit
cycle a1 = 2.5, A = 2.5, K1 = 200, w = 3 (ii) chaos a1 = 2, w = 3, A = 2, K1 = 100. Fixed values used for relevant cases: a1 = 2, a2 = 1,
w1 = 2, c = 0.2, w4 = 0.0257, w = 1, D = 10, D1 = 10, w2 = 0.05, D2 = 10, A = 1.5, K = 100, w3 = 0.74, D3 = 20.
e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2 215
generalist predators whose food requirements are quite low.
On the other hand, generalist predators with appreciable food
needs are better represented by the present version of the
model. We shall compare results of this model and those of Rai
(2004). The following schematic diagram represents the
model:
A real world ecological community that exemplifies this
model involves plantation, vole, weasel and its prey other
than the voles. This community is found in northern
Fennoscandia and has been studied by many investigators
(Turchin, 1996; Turchin and Hanski, 1997; Turchin and Ellner,
2000).
2.2. Model 2: two specialists, two competing prey
In this section, we design a model to represent an ecological
situation wherein two different prey are predated upon by two
different specialist predators. These two predator–prey sys-
tems with strong non-linear interactions are linked via
competition between the individuals of prey populations.
Vandermeer (2004) terms this as resource–resource (RR)
coupling. RR coupling essentially means indirect mutualism
between the two consumers (predators). Our aim is to
investigate the effect of this on two RM predator–prey systems.

e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2216
The model is represented by following set of non-linear
coupled ordinary differential equations:
dX1
dt¼ a1X1 1� X1
K� X2
K1
� ��wY1X1
X1 þ D(4a)
dY1
dt¼ �a2Y1 þ
w1Y1X1
X1 þ D1(4b)
dX2
dt¼ a3X2 1� X2
K2� X1
K1
� ��w2Y2X2
X2 þ D2(4c)
dY2
dt¼ �a4Y2 þ
w3Y2X2
X2 þ D3; (4d)
The meaning of the symbols is the same as in Eqs. (1a) and (1b).
A population’s growth is limited either by the severity of
competition among its own individuals or/and by that of
competition among individuals of other species, which
depend on the same resource. K is the carrying capacity for
the prey species X1 and K2 for that of X2. K1 measures the
strength of interference competition between the two prey
species. The unit of K1 is number of individuals per unit
volume. The meaning of different symbols will be more clear
if one keeps in view the fact that this system is obtained by
coupling the two RM oscillators as shown in the following
schematic:
A real world community exemplifying this ecological
situation is Epirrita autumnata (a lepidopteran forest insect
pest) feeding on the plants in northern Fennoscandia and the
voles of the genera Microtus and Clethrionomys feedings on
another set of plants. The first herbivore system has 9-year
population cycle and the second system has a 5-year cycle
(Klemola et al., 2002).
2.3. Model 3: two competing specialists, two prey
Here we again design a model to examine the nature and
extent to which dynamics of an ecological system is
determined by coupling of two RM oscillators. In this case,
the predators can be thought of as specialized on a particular
prey, but consume an alternative prey on which another
predator specializes (MacArthur, 1970). Vandermeer (2004)
refers to this type of coupling as consumer–resource (CR)
coupling. CR coupling implies competition between two
predators. The model is represented by following set of
non-linear coupled differential equations:
dX1
dt¼ a1X1 1� X1
K
� ��wX1
Y1
1þ bðX1 þ bX2Þþ bY2
1þ bðX2 þ bX1Þ
� �
(5a)
dY1
dt¼ wðX1 þ bX2ÞY1
1þ bðX1 þ bX2Þ� a2Y1 (5b)
dX2
dt¼ a3X2 1� X2
K
� ��wX2
Y2
1þ bðX2 þ bX1Þþ bY1
1þ bðX1 þ bX2Þ
� �
(5c)
dY2
dt¼ wðX2 þ bX1ÞY2
1þ bðX2 þ bX1Þ� a4Y2; (5d)
where X1, X2 are the two prey and Y1, Y2 are corresponding
predators. These predators prey on each other’s prey. a1 and a3
are per capita reproductive growth rates for the prey X1 and X3.
K is the carrying capacity for the prey and b is the parameter
for the functional response of predators. Parameter b mea-
sures the strength of CR coupling. a2, a4 are mortality rates for
the two predators. The parameter w represents the consump-
tion rates of predators. The model is parameterized in such a
way that variables take any value between 0 and 1.
2.4. Model 4: one specialist, two competing prey
In this model, two prey species compete for the same resource
and each one is predated by the same specialist predator. We
thus consider switching behavior in the specialist predator, as
no predator is a specialist predator in the mathematical sense
encrypted in Eqs. (1a) and (1b). Some specialist predators do
switch to alternative prey when their most favorite food is in
short supply; e.g. lynx switches to red squirrel when snowshoe
hare becomes scarce (O’Donoghue et al., 1998), and the
Eurasian badger switches to rabbits when its preferred prey
(plants, worms and insects) becomes scarce (Fedriani et al.,
1998). The model is described by following equations:
dX1
dt¼ r1X1 1� X1
K1� c12X2
K1
� �� f1F1ðX1;X2ÞY (6a)
dX2
dt¼ r2X2 1� X2
K2� c21X1
K2
� �� f2F2ðX1;X2ÞY (6b)
dYdt¼ e1 f1F1ðX1;X2ÞY þ e2 f2F2ðX1;X2ÞY � dY; (6c)
where
f1 ¼pX1
pX1 þ ð1� pÞX2and f2 ¼
ð1� pÞX2
pX1 þ ð1� pÞX2;
where p is the prey preference and takes a value between 0
and 1.
FiðX1;X2Þ ¼AiXi
ð1þ B1X1 þ B2X2Þ;

Fig. 2 – Points in 2D parameter space where chaos was
observed in Model 2. (a) 0.5 = a1 = 1.9; 30 = K1 = 65 with
step size (0.1, 5). (b) 0.2 = a2 = 0.8; 30 = K1 = 60 with step
size (0.05, 5) (c) 0.5 = a3 = 1.9; 30 = K1 = 60 with step size
e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2 217
where Ai is the maximum harvest rate of predator Y for prey Xi
and 1/Bi is proportional to the half-saturation constants. The
constants e1 and e2 are conversion rates of preyXi to predator Y.
Setting f1 and f2 equal to 1 in Eqs. (6a)–(6c), one gets the
system without switching. This system can be decoupled into
two RM oscillators (Rai, 2004). The two oscillators are
connected through inter-specific competition.
Gakkhar and Naji (2003) have discovered chaos in the
model without switching. However, scans of parameter space
were not presented in their publication. The assessment of
dynamical complexity is not possible without studying the
model in more detail. We performed simulation experiments
to investigate dynamical chaos in both versions of the model
(with and without switching).
2.5. Model 5: two generalists, two prey
This model is obtained by linking the two HT oscillators. It is
assumed that two generalist predators feed on both prey
species although they have their own favorite food items (X for
Y and Z for U). The model is represented by the following set of
differential equations:
dXdt¼ r1X 1� X
K1
� �� X
wY1þ Xþ aZ
þ aU1þ Xþ aZ
� �(7a)
dYdt¼ c1Y1 �
w2Y2
Xþ aZ(7b)
dZdt¼ r2Z 1� Z
K2
� �� Z
bY1þ Zþ bX
þ w1U1þ Zþ bX
� �(7c)
dUdt¼ c2U� w3U2
Zþ bX; (7d)
where parameters b and a are measures of intensity with
which two subsystems are connected to one another. Van-
dermeer (1993) called them parameters of connection asym-
metry (when b 6¼ a). r1, K1 and r2, K2 are per capita growth rates
and carrying capacities for the two prey. He found chaos for
a = 0.005 and b = 0.008. Here we present results of detailed
simulation experiments, which were performed in the spirit
of the methodology in Upadhyay et al. (2001).
Model 5 contains two types of parameters: (1) connection
asymmetry (a and b) and (2) driving asymmetry (r1, r2; K1, K2). The
(0.05, 5). Fixed values used for relevant cases: a2 = 1, a4 = 1,
a3 = 1.2, K = 35, K2 = 30, D = 10, D1 = 10, D2 = 15, D3 = 15,
w = 1, w1 = 2, w2 = 1, w3 = 2.
e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2218
connection asymmetry parameters are measures of predation
intensity between the generalist predator and the other prey
(the one which is not its most favorite prey). On the other
hand, parameters of driving asymmetry are the per capita
reproductive rates and carrying capacities for the two prey. We
chose both kinds of parameters as control parameters to
perform simulation experiments.
3. Methodology
The model systems presented above are multi-parameter
systems. A few hundred parameter combinations (choosing
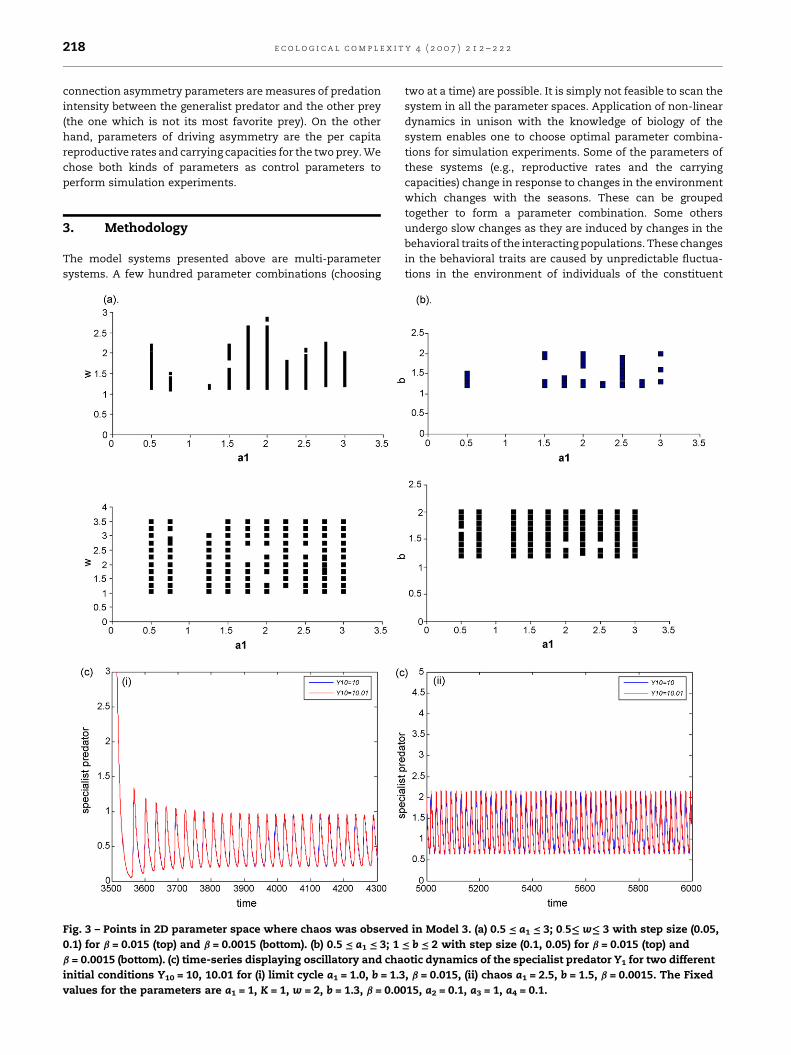
Fig. 3 – Points in 2D parameter space where chaos was observe
0.1) for b = 0.015 (top) and b = 0.0015 (bottom). (b) 0.5 = a1 = 3; 1
b = 0.0015 (bottom). (c) time-series displaying oscillatory and cha
initial conditions Y10 = 10, 10.01 for (i) limit cycle a1 = 1.0, b = 1.3
values for the parameters are a1 = 1, K = 1, w = 2, b = 1.3, b = 0.00
two at a time) are possible. It is simply not feasible to scan the
system in all the parameter spaces. Application of non-linear
dynamics in unison with the knowledge of biology of the
system enables one to choose optimal parameter combina-
tions for simulation experiments. Some of the parameters of
these systems (e.g., reproductive rates and the carrying
capacities) change in response to changes in the environment
which changes with the seasons. These can be grouped
together to form a parameter combination. Some others
undergo slow changes as they are induced by changes in the
behavioral traits of the interacting populations. These changes
in the behavioral traits are caused by unpredictable fluctua-
tions in the environment of individuals of the constituent
d in Model 3. (a) 0.5 = a1 = 3; 0:5��� w��� 3 with step size (0.05,
= b = 2 with step size (0.1, 0.05) for b = 0.015 (top) and
otic dynamics of the specialist predator Y1 for two different
, b = 0.015, (ii) chaos a1 = 2.5, b = 1.5, b = 0.0015. The Fixed
15, a2 = 0.1, a3 = 1, a4 = 0.1.

e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2 219
populations. To fix the ranges of parameters appearing in
parameter combinations for simulation experiments, we use
the method suggested by Upadhyay et al. (1998). The system of
equations is integrated using standard numerical procedures
to generate time-histories for a given set of parameter values.
The distinguishing property of chaos is sensitive dependence
on initial conditions. Two initially close trajectories in the state
space (or in the phase space) diverge exponentially fast to the
extent that after elapse of a certain amount of time, the two
appear to originate from two different dynamical systems. We
choose this basic feature of chaotic dynamics to detect the
presence as well as absence of chaos in these model systems.
The system parameters are chosen in such a way that
asymptotic dynamics of both the subsystems confine to a
stable limit cycle. If the two time-histories (generated by
integrating the system of differential equations and recorded
after transients are died out) corresponding to two different,
but very close, initial conditions overlap each other comple-
tely, the absence of dynamical chaos at that particular set of
Fig. 4 – Points in 2D parameter space where chaos was observed
switching behavior. For (a): (i) 0.5 = r1 = 4.5; 50 = K2 = 350 with s
(0.2, 10). The parameter values for limit cycle solutions are: r1 =
c12 = 0.204, C21 = 0.208, d = 1.21, B1 = 0.54, B2 = 0.55, e1 = 2.5, e2 = 0
25). (ii) 1.5 = r2 = 4; 50 = K1 = 250 with step size (0.25, 25). The pa
K1 = 150, K2 = 155, A1 = 1.1, A2 = 0.26, c12 = 1.1, c21 = 0.2, d = 1.2, B
parameter values is inferred. In the case when two histories
appear to be quite different, it is understood that chaos exists
at the chosen set of parameter values. There is one more
important aspect of these simulation experiments: choosing
the step size for the variation of a system parameter from a
parameter combination within the chosen range. This
depends on the nature of the parameter concerned (whether
it is a slow varying or fast varying one).
We chose the range of the two critical parameters based on
the available information about the parameters (Jorgensen,
1979), which are intrinsic attributes of the system. The critical
parameters are varied one at a time in the chosen range. ODE
Workbench from AIP (IBM PC Version 1.5; Aguirregabiria, 1994)
was used to integrate the system of differential equations
numerically. Dormance and Prince 5 (Hairer et al., 1993) was
the integration algorithm chosen for the purpose. Comparing
two time histories is the most simple and efficient way of
detecting the presence as well as absence of chaos in model
dynamical systems. This enables one to study two forms of
in Model 4 without switching behavior ( f1, f2 = 1) and with
tep size (0.1, 10). (ii) 1 = r2 = 4; 50 = K1 = 250 with step size
2.6, r2 = 2.65, K1 = 150, K2 = 155, A1 = 1.06, A2 = 0.35,
.52. For (b) (i) 1 = r1 = 2.8; 50 = K2 = 350 with step size (0.1,
rameter values for limit cycle solutions are: r1 = 2.6, r2 = 2.1,
1 = 0.54, B2 = 0.01, e1 = 2.5, e2 = 0.52, p = 0.5.
e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2220
chaos in dissipative dynamical systems (dynamical chaos as
well as intermittent chaos) (Berge et al., 1986; Vandermeer,
1993; Letellier and Aziz-Alaoui, 2002). Of course, the method is
not helpful to distinguish between long-lived chaotic tran-
sients and chaotic dynamics on a bonafide chaotic attractor.
The phenomenon of chaotic transients (Grebogi et al., 1983;
Rai and Schaffer, 2001) is more prevalent in spatially extended
systems, which we do not study in the present paper. We also
mention that we did not use the maximum Lyapunov
exponent to detect chaos in the models we studied because
the method we chose was time saving. Moreover, a positive
maximum Lyapunov exponent does not guarantee dynamical
chaos even in a deterministic system (Alligood et al., 1997). Of
course, one has to take precautions to detect weak chaos using
this methodology. A large number of data points are thrown
out as transients so that there is sufficient time for chaos to
develop.
We (Rai, 2004; Rai and Upadhyay, 2004) have classified
dynamical complexity into two main categories: (1) robust
chaos and (2) short-term recurrent chaos (strc). When one finds
dynamical chaos to exist in a densely populated region defined
by two control parameters of the system, a case for robust
chaos is encountered. On the other hand, strc manifests itself
as a scatter of points in 2D parameter spaces. Extensive
simulation experiments were performed in 2D parameter
spaces spanned by per capita reproductive rate (a1) and
carrying capacity (K) of prey X; death rate (a2) and the foraging
efficiency ðw1Þ of the specialist predator; per capita reproduc-
tive rate (c) and a measure of dependence of the generalist
predator on its most favorite prey ðw4Þ to mention a few.
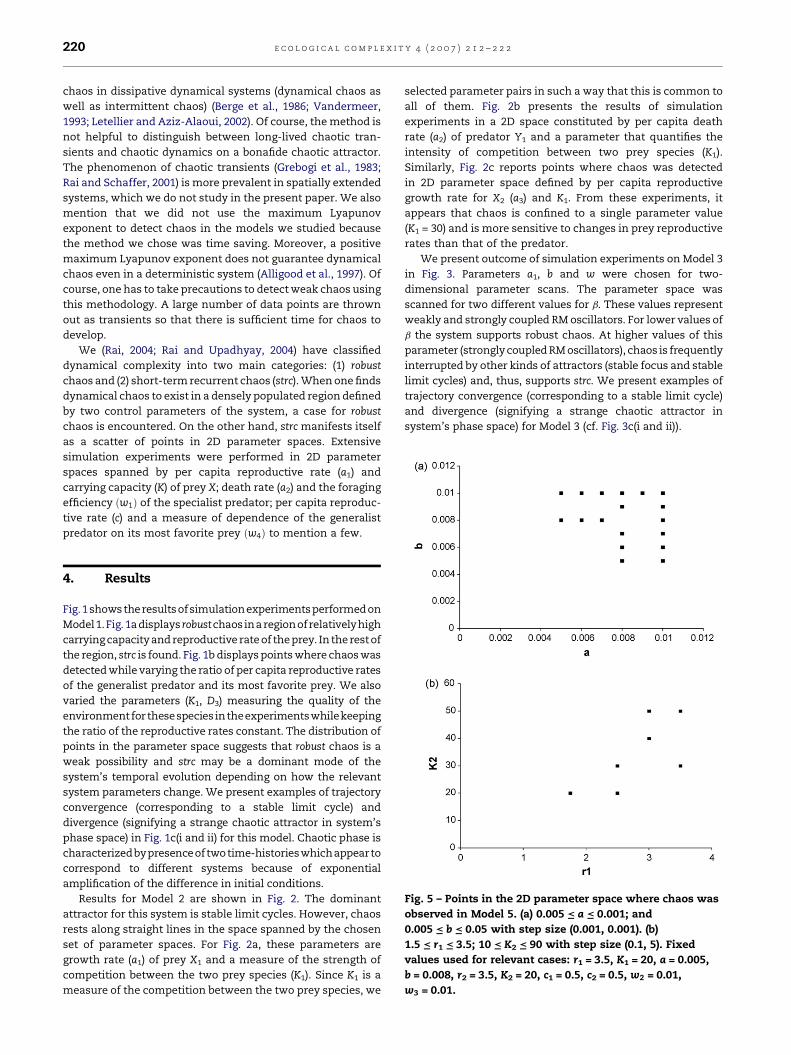
Fig. 5 – Points in the 2D parameter space where chaos was
observed in Model 5. (a) 0.005 = a = 0.001; and
0.005 = b = 0.05 with step size (0.001, 0.001). (b)
1.5 = r1 = 3.5; 10 = K2 = 90 with step size (0.1, 5). Fixed
values used for relevant cases: r1 = 3.5, K1 = 20, a = 0.005,
b = 0.008, r2 = 3.5, K2 = 20, c1 = 0.5, c2 = 0.5, w2 = 0.01,
w3 = 0.01.
4. Results
Fig.1showstheresultsofsimulationexperimentsperformedon
Model1.Fig.1adisplays robustchaos inaregionofrelativelyhigh
carrying capacity and reproductive rate of the prey. In the rest of
the region, strc is found. Fig. 1b displays points where chaos was
detected while varying the ratio of per capita reproductive rates
of the generalist predator and its most favorite prey. We also
varied the parameters (K1, D3) measuring the quality of the
environmentfor thesespecies intheexperimentswhilekeeping
the ratio of the reproductive rates constant. The distribution of
points in the parameter space suggests that robust chaos is a
weak possibility and strc may be a dominant mode of the
system’s temporal evolution depending on how the relevant
system parameters change. We present examples of trajectory
convergence (corresponding to a stable limit cycle) and
divergence (signifying a strange chaotic attractor in system’s
phase space) in Fig. 1c(i and ii) for this model. Chaotic phase is
characterizedbypresenceoftwotime-historieswhichappearto
correspond to different systems because of exponential
amplification of the difference in initial conditions.
Results for Model 2 are shown in Fig. 2. The dominant
attractor for this system is stable limit cycles. However, chaos
rests along straight lines in the space spanned by the chosen
set of parameter spaces. For Fig. 2a, these parameters are
growth rate (a1) of prey X1 and a measure of the strength of
competition between the two prey species (K1). Since K1 is a
measure of the competition between the two prey species, we
selected parameter pairs in such a way that this is common to
all of them. Fig. 2b presents the results of simulation
experiments in a 2D space constituted by per capita death
rate (a2) of predator Y1 and a parameter that quantifies the
intensity of competition between two prey species (K1).
Similarly, Fig. 2c reports points where chaos was detected
in 2D parameter space defined by per capita reproductive
growth rate for X2 (a3) and K1. From these experiments, it
appears that chaos is confined to a single parameter value
(K1 = 30) and is more sensitive to changes in prey reproductive
rates than that of the predator.
We present outcome of simulation experiments on Model 3
in Fig. 3. Parameters a1, b and w were chosen for two-
dimensional parameter scans. The parameter space was
scanned for two different values for b. These values represent
weakly and strongly coupled RM oscillators. For lower values of
b the system supports robust chaos. At higher values of this
parameter (strongly coupled RM oscillators), chaos is frequently
interrupted by other kinds of attractors (stable focus and stable
limit cycles) and, thus, supports strc. We present examples of
trajectory convergence (corresponding to a stable limit cycle)
and divergence (signifying a strange chaotic attractor in
system’s phase space) for Model 3 (cf. Fig. 3c(i and ii)).

e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2 221
Fig. 4 shows the results of our simulation experiments on
Model 4. We explored parameter spaces spanned by r1, K2 and
r2, K1. Results simulation experiments are presented in Figure
(a) without and (b) with switching in the diet of the predator.
We found few and intermittent occurrences of chaos in
general, and in the case of switching, no chaos was observed in
the parameter scan in r2 � K1 space.
Results of simulation experiments to study presence and
absence of chaos in Model 5 is presented in Fig. 5. In these
experiments, one connection parameter was varied at a time
(cf. Fig. 5a). We also study presence and absence of chaos in
this model system by varying r1 and K2 (Fig. 5b).
5. Discussion and conclusion
In all the models we studied chaotic dynamics is interrupted by
predictable changes in system parameters. The deterministic
change in system parameters is to be contrasted with stochastic
environmental influences that act on initial conditions.
Seasonality is one of the agencies causing these deterministic
changes in system parameters (reproductive rates and carrying
capacities). Even if one discovers a way to deal with the impact
of long-term environmental fluctuations so as to get a suitable
time-series which can be analyzed using concepts and
techniques fromnon-linear time series analysisand forecasting
(Turchin and Ellner, 2000), the likelihood that an unambiguous
case for chaos will be found is small. The on–off character of
chaos will render its capture in the wild elusive. In the presence
(and absence) of exogenous environmental perturbations,
ecological systems will display short-term recurrent chaos
(Turchin and Ellner, 2000; Rai, 2004; Rai and Upadhyay, 2004,
2006). The challenge remains the same: to distinguish between
deterministic chaos and noisy periodicity (Olsen and Schaffer,
1990; Kendall, 2001; Rai and Schaffer, 2001). We emphasize that
the source of this challenge is frequent interruptions in chaotic
dynamics caused by non-linear interactions.
We found that in most of the models, the dominant
behavior found was stable limit cycles. We hypothesize that
the coupling between the dynamics of specialist non-
competing predators (herbivorous insects and small mam-
mals, e.g., voles) and their prey (e.g., different varieties of
plants) explains pronounced population cycles observed in
many predator–prey systems, for example the herbivores in
northern Fennoscandia (Klemola et al., 2002). No population-
extrinsic condition is needed to understand the differences
between these observed cycles and ideal cycles. In harsh
environments experiencing strong seasonality, the inter-
specific competition between different prey (functional
varieties of plants) may be severe enough to put the parameter
(K1) at high values. Thus, occasional chaotic bursts can explain
the apparent departures from ideal cycles observed in real
population dynamics.
Our study allows us to make some comments about the
relationship between trophic structure and dynamical com-
plexity. It appears that the most complex models (Models 1
and 3) displayed dynamical complexity (in the form of strc). It is
interesting to note that Model 1 is the only one that
incorporates both generalist and specialist predators. This
provides a nice complement to results of Fussman and Heber
(2002) who showed that the frequency of chaotic dynamics
increases with the number of trophic levels (they did not
differentiate between generalist and specialist predators). If
we compare the results from an earlier work on a different
version of Model 1, which assumes that the switching to
alternative prey in the generalist predator’s foraging strategy
is infrequent (Rai, 2004), we find that the system’s tendency to
exhibit episodes of chaotic dynamics is considerably increased
(cf. description of Model 1 in Section 2.1). Model 2 contains two
specialist predators whose prey are in competition with each
other. In this model, we see a few points where chaotic
dynamics exists. This suggests that most likely outcome of
models involving the same kind of predators (either the
specialist or generalist) in natural systems is cycles not chaos.
This could be an explanation for the abundance of population
cycles in nature (Kendall et al., 1999). Of course, the observed
dynamical patterns will not conform to ideal cycles. These
departures are caused by existence of chaotic dynamics in
some (albeit small) regions of the parameter space. Thus, non-
linear interactions, and not exogenous environmental factors
could explain observations of these deviations from ideal
cycles. It is interesting to note that coupling two generalists
(Model 5) shows more points of observation of chaos than
coupling two specialist predators (Model 2) and the model with
one specialist predator with switching in its diet (Model 4).
This suggests that the generalist behavior may be functionally
important for dynamical complexity in these types of systems
(Tanabe and Namba, 2005).
Acknowledgements
M.A. acknowledges financial support from a Canada Research
Chair and Discovery grant (Natural Sciences and Engineering
Council of Canada) and the Ontario Ministry of Economic
Development and Trade (Science and Technology Division) to
undertake this collaborative project. V.R. gratefully acknowl-
edges the Department of Biology at Laurentian University for
hosting his 4-month visit and Bai-Lian (Larry) Li for helpful
discussions during a brief visit to his lab. R.U. acknowledges
financial support from D.S.T., Government of India.
r e f e r e n c e s
Aguirregabiria, J.M., 1994. ODE Workbench. Physics AcademicSoftware.
Alligood, K.T., Sauer, T.D., Yorke, J.A., 1997. Chaos: AnIntroduction to Dynamical Systems. Springer, Berlin,Germany.
Arneodo, A., Coullet, P., Tresser, C., 1980. Occurrence of strangeattractors in three-dimensional Volterra equation. Phys.Lett. 79A, 259–263.
Aziz-Alaoui, M.A., 2002. Study of a Leslie–Gower type tri-trophicpopulation model. Chaos Solitons Fract. 14 (8), 1275–1293.
Berge’, P., Pomeau, Y., Vidal, C., 1986. Order Within Chaos:Towards a Deterministic Approach to Turbulence. JohnWiley & Sons, NY, USA.
Edwards, A.M., Bees, M.A., 2001. Generic dynamics of a simpleplankton population model with a non-integer exponent ofclosure. Chaos Solitons Fract. 12 (2), 289–300.

e c o l o g i c a l c o m p l e x i t y 4 ( 2 0 0 7 ) 2 1 2 – 2 2 2222
Fedriani, J.M., Ferreras, P., Delibes, M., 1998. Dietary response ofthe Eurasian badger, Meles meles, to a decline of its mainprey in the Donana National Park. J. Zool. 245, 214–218.
Fussman, G.F., Heber, G., 2002. Food web complexity and chaoticpopulation dynamics. Ecol. Lett. 5, 394–401.
Gakkhar, S., Naji, R.K., 2003. Existence of chaos in two-prey, onepredator system. Chaos Solitons Fract. 17, 639–649.
Gilpin, M.E., 1979. Spiral chaos in a predator–prey model. Am.Nat. 113, 306–308.
Grebogi, C., Ott, E., Yorke, J.A., 1983. Crises, sudden changes inchaotic attractors and transient chaos. Phys. D 7, 181–200.
Hastings, A., Powell, T., 1991. Chaos in a three-species foodchain. Ecology 72, 896–903.
Hairer, E., Norsett, S.P., Wanner, G., 1993. Solving DifferentialEquations I: Nonstiff Problems, 2nd ed. Springer, Berlin.
Holling, C.S., 1965. The functional response of invertebratepredators to prey density. Mem. Entomol. Soc. Can. 45, 3–60.
Jorgensen, S.E., 1979. Handbook of Environmental Data andEcological Parameters. Pergamon Press, Amsterdam.
Kendall, B.E., 2001. Cycles, chaos and noise in predator–preydynamics. Chaos Solitons Fract. 12, 321–332.
Kendall, B.E., Briggs, C.J., Murdock, W.W., Turchin, P., Ellner,S.P., McCauley, E., Nisbet, R.M., Wood, S.N., 1999. Why dopopulations cycle? A synthesis of statistic and mechanicalmodeling approaches. Ecology 80, 1789–1805.
King, A.A., Schaffer, W.M., Gordon, C., Treat, J., Kot, M., 1996.Weakly dissipative predator–prey systems. Bull. Math. Biol.58, 835–859.
King, A.A., Costantino, R.F., Cushing, J.M., Henson, S.M.,Desharnais, R.A., Denis, B., 2004. Anatomy of a chaoticattractor: subtle model predicted patterns revealed inpopulation data. Proc. Natl. Acad. Sci. U.S.A. 101, 408–413.
Klebanoff, A., Hastings, A., 1994. Chaos in three species foodchains. J. Math. Biol. 32, 427–451.
Klemola, T., Tanhuanpaa, M., Korpimaki, E., Ruohomaki, K.,2002. Specialist and generalist natural enemies as anexplanation for geographical gradients in population cyclesof northern herbivores. Oikos 99, 83–94.
Letellier, C., Aziz-Alaoui, M.A., 2002. Analysis of the dynamics ofa realistic ecological model. Chaos Solitons Fract. 13, 95–107.
Lotka, A.J., 1920. Undamped oscillations derived from the law ofmass action. J. Am. Chem. Soc. 42, 1595.
MacArthur, R.H., 1970. Species packing and competitiveequilibrium for many species. Theor. Pop. Biol. 1, 1–11.
May, R.M., 1975. Biological populations obeying differenceequations: simple points, stable cycles, and chaos. J. Theor.Biol. 49, 511–524.
May, R.M., 1976. Simple mathematical models with verycomplicated dynamics. Nature 261, 459–467.
May, R.M., Oster, G.F., 1976. Bifurcation and dynamic complexityin simple ecological models. Am. Nat. 110, 578–599.
May, R.M., 2001. Stability and Complexity in Model Ecosystems.Princeton University Press, Princeton, New Jersey.
Medvinsky, A.B., Petrovskii, S.V., Tikhonova, I.A., Malchow, H.,Li, B.-L., 2002. Spatio-temporal complexity of plankton andfish dynamics. SIAM Rev. 44, 311–370.
Morozov, A., Petrovski, S., Li, B.-L., 2004. Bifurcations and chaosin a predator–prey system with Allee effect. Proc. R. Soc.Lond. B 271, 1407–1414.
O’Donoghue, M., Boutin, S., Krebs, C.J., Zuleta, G., Murray, D.L.,Hofer, E.J., 1998. Functional response of coyotes and lynx tothe snowshoe hare cycle. Ecology 79, 1193–1208.
Olsen, L.F., Schaffer, W.M., 1990. Chaos versus noisy periodicity:alternative hypotheses for childhood epidemics Science249, 499–504.
Pascual, M., 1993. Diffusion-induced chaos in a spatial predator–prey system. Proc. R. Soc. Lond. B 251, 1–7.
Pielou, E.C., 1977. Mathematical Ecology: An Introduction.Wiley, NY, USA.
Petrovski, S., Li, B.-L., Malchow, H., 2004. Transition tospatiotemporal chaos can resolve the paradox ofenrichment. Ecol. Complexity 1, 37–47.
Rai, V., Sreenivasan, R., 1993. Period doubling bifurcationsleading to chaos in a model food-chain. Ecol. Model. 69,63–77.
Rai, V., Schaffer, W.M., 2001. Chaos in ecology. Chaos SolitonsFract. 12, 197–203.
Rai, V., 2004. Chaos in natural populations: edge or wedge? Ecol.Complexity 1 (2), 127–138.
Rai, V., Anand, M., 2004. Is dynamic complexity of ecologicalsystems quantifiable? Int. J. Ecol. Env. Sci. 30, 123–130.
Rai, V., Upadhyay, R.K., 2004. Chaotic population dynamics andthe biology of the top-predator. Chaos Solitons Fract. 21,1195–1204.
Rai, V., Upadhyay, R.K., 2006. Evolving to the edge of chaos:chance or necessity? Chaos Solitons Fract. 30, 1074–1087.
Rinaldi, S., Muratori, S., 1993. Conditioned chaos in seasonallyperturbed predator–prey models. Ecol. Model. 69, 79–97.
Rinaldi, S., Muratori, S., Kuznetsov, Y., 1993. Multiple attractors,catastrophes and chaos in seasonally perturbed predator–prey communities. Bull. Math. Biol. 55, 15–35.
Rosenzweig, M.L., MacArthur, R.H., 1963. Graphicalrepresentation and stability conditions of predator–preyinteractions. Am. Nat. 97, 209–223.
Schaffer, W.M., 1985. Order and chaos in ecological systems.Ecology 66, 93–106.
Schaffer, W.M., 1988. Perceiving order in the chaos of nature. In:Boyce, M. (Ed.), Life Histories: Theory and Patterns fromMammals. Yale University Press, New Haven, pp. 313–350.
Samardzija, N., Greller, L.D., 1988. Explosive route to chaosthrough a fractal torus in a generalized Lotka–Volterramodel. Bull. Math. Biol. 50 (5), 465–491.
Tanabe, K., Namba, T., 2005. Omnivory creates chaos in simplefood web models. Ecology 86 (12), 3411–3414.
Tanner, J.T., 1975. The stability and intrinsic growth rates ofprey and predator populations. Ecology 56, 855–867.
Turchin, P., 1996. Non-linear time-series modeling of volepopulation fluctuations. Res. Pop. Ecol. 38, 121–132.
Turchin, P., Hanski, I., 1997. An empirically-based model for thelatitudinal gradient in vole population dynamics. Am. Nat.149, 842–874.
Turchin, P., Ellner, S.P., 2000. Living on the edge of chaos:population dynamics of Fennoscandian voles. Ecology 81,3099–3116.
Upadhyay, R.K., Rai, V., 1997. Why chaos is rarely observed innatural populations? Chaos Solitons Fract. 8, 1933–1939.
Upadhyay, R.K., Iyengar, S.R.K., Rai, V., 1998. Chaos: anecological reality? Int. J. Bifur. Chaos 8, 1325–1333.
Upadhyay, R.K., Rai, V., 2001. Crisis-limited chaotic dynamics inmodel ecological systems. Chaos Solitons Fract. 12,205–218.
Upadhyay, R.K., Rai, V., Iyengar, S.R.K., 2001. Species extinctionproblem: genetic vs. ecological factors. Appl. Math. Model.25, 937–951.
Vandermeer, J., 1993. Loose coupling of predator–prey cycles:entrainment, chaos and intermittency in the classicalMacArthur consumer–resource equations. Am. Nat. 141,687–716.
Vandermeer, J., Stone, L., Blasius, B., 2001. Categories of chaosand fractal basin boundaries in forced predator–preymodels. Chaos Solitons Fract. 12 (2), 265–276.
Vandermeer, J., 2004. Coupled oscillations in food webs:balancing competition and mutualism in simple ecologicalmodels. Am. Nat. 163, 857–867.
Volterra, V., 1931. Variations and fluctuations of the number ofindividuals in animal species living together. In: AnimalEcology, McGraw-Hill, Translated from 1928 edition by R.N.Chapman.