tree species mediated effects on leaf litter dynamics in pure and mixed stands of oak and beech
TRANSCRIPT

Tree species mediated effects on leaf litterdynamics in pure and mixed stands of oak andbeech
Mathieu Jonard, Frederic Andre, and Quentin Ponette
Abstract: This study aimed to evaluate the relative importance of the factors whereby tree species composition can influ-ence leaf litter dynamics. Leaf litter production and chemical composition were measured in pure and mixed stands of oak(Quercus petraea Liebl.) and beech (Fagus sylvatica L.). Pure and mixed leaf litter of both species were incubated in eachstand type to assess separately the environmental, litter quality, and litter mixture effects on decomposition. To betterunderstand the environmental effects, ground climate was measured in the different stands and the effects of soil watercontent on decomposition were evaluated using roofs to simulate drought conditions. Although total leaf litter amountswere not affected by stand composition, leaf decomposition varied with litter quality and with the environmental condi-tions. In the same environment, oak leaf litter disappeared on average 1.7 times faster than beech leaf litter. Decomposi-tion of oak leaves increased significantly in the mixed-species litterbags. In contrast, the overall mass loss of the mixedlitter tallied with the mass loss estimated by examining the decomposition of the component litter separately (additive-ef-fect hypothesis). The effects of stand type appeared in the third year of incubation: leaf mass loss of both species wasgreater in the beech stand. In addition, soil water content affected leaf decomposition: the oak and beach leaf mass lossesdropped by 24% and 17%, respectively, in the dry modality.
Resume : Cette etude a pour objectif d’evaluer l’importance relative des facteurs par lesquels la composition specifiquepeut influencer la dynamique des litieres foliaires. La production et la composition chimique des litieres foliaires ont etemesurees dans des peuplements purs et melanges de chene (Quercus petraea Liebl.) et de hetre (Fagus sylvatica L.). Deslitieres pures et melangees des deux especes ont ete mises en incubation dans chaque type de peuplements de maniere aevaluer separement l’impact de l’environnement, de la qualite et du melange des litieres sur la decomposition. Afin demieux cerner les effets de l’environnement, le pedoclimat a ete suivi dans les differents peuplements et l’effet de la teneuren eau du sol sur la decomposition a ete mesure grace a un dispositif creant des conditions de secheresse. Alors que laproduction totale de litiere foliaire n’a pas ete affectee par la composition du peuplement, la decomposition a ete influen-cee par la qualite de la litiere et par les conditions environnementales. Dans un meme environnement, la litiere foliaire dechene a disparu en moyenne 1,7 fois plus vite que celle de hetre. Dans les sachets de litiere melangee, la decompositiondes litieres foliaires de chene a ete significativement acceleree. Par contre, la perte de masse globale des litieres melangeesn’etait pas significativement differente de celle predite sur base de la decomposition dans les sachets de litieres pures (hy-pothese d’additivite des effets). L’effet « peuplement » est apparu au cours de la troisieme annee d’incubation, la perte demasse des litieres foliaires des deux especes etant plus grande en hetraie. Par ailleurs, nous avons observe un effet de lateneur en eau sur la decomposition ; la perte de masse des litieres foliaires de chene et de hetre a ete reduite respective-ment de 24 % et 17 % dans la modalite seche.
IntroductionLitterfall transfers dead organic matter from the above-
ground vegetation to the forest floor, where it gradually de-composes. Decomposition results from the interactingactivities of microorganisms and soil fauna, which are con-trolled by litter quality (McClaugherty and Berg 1987; Aber etal. 1990), environmental factors such as climate (Meente-meyer 1978; Gholz et al. 2000), and soil properties (Vesterdal
1999). Without major disturbances (fire or timber harvesting),decomposition together with litterfall regulate litter accu-mulation (Olson 1963).
Tree species composition can substantially affect leaf lit-ter dynamics because litter production and chemical compo-sition vary widely among species (Bray and Gorham 1964;Berg and McClaugherty 2003). Moreover, tree-canopy char-acteristics influence decomposition conditions (Cote andFyles 1994; Prescott et al. 2000; Albers et al. 2004; Hobbieet al. 2006) through their effects on ground climate, soilproperties, and understory vegetation. In mixed forests, dif-ferent species can affect leaf litter dynamics by creating spe-cific substrate properties and decomposition conditions.Studies comparing mixed- with single-species litters havereported increased, decreased, or unchanged decompositionrates (McTiernan et al. 1997; Wardle et al. 1997). The non-additive effects are generally attributed to changes in diver-sity and (or) abundance of soil fauna, to nutrient transfers
Received 3 April 2007. Accepted 18 September 2007. Publishedon the NRC Research Press Web site at cjfr.nrc.ca on23 February 2008.
M. Jonard,1 F. Andre, and Q. Ponette. Universite catholiquede Louvain, Faculte d’Ingenierie Biologique, Agronomique etEnvironnementale, Unite des Eaux et Forets, Croix du sud 2/0091348 Louvain-la-Neuve, Belgium.
1Corresponding author (e-mail: [email protected]).
528
Can. J. For. Res. 38: 528–538 (2008) doi:10.1139/X07-183 # 2008 NRC Canada

between litter of different quality, and to compounds (suchas polyphenols) acting either as inhibitors or stimulators(Hattenschwiler 2005).
On a given site, litter quality is generally considered to bethe most important factor affecting decomposition; environ-mental conditions are secondary (Blair et al. 1990). How-ever, experiments that distinguish between the litter qualityand the environmental factors associated with species com-position are rare (Hobbie et al. 2006).
Our study aimed to evaluate the relative importance of thefactors whereby species composition can influence leaf litterdynamics in an oak (Quercus petraea Liebl.) and beech(Fagus sylvatica L.) forest. First, leaf litter production andchemical composition were measured in pure and mixedstands of oak and beech. Second, a transplantation litterbagexperiment was carried to separate litter quality from envi-ronment effects on both single- and mixed-species leaf litterdecomposition. Finally, to better understand the influence ofenvironment on decomposition, ground climate was meas-ured in the different stands and the effects of soil water con-tent on leaf litter decomposition were evaluated using roofsto simulate drought conditions and broaden the range of soilwater contents usually observed under the various stands.
Materials and methods
Study siteThe study site is located in the western part of the Belgian
Ardennes at 300 m elevation (50801’N, 4824’E). The meanannual rainfall is slightly >1000 mm, and the mean annualtemperature is 8 8C. The site consists of 60 ha oak and beechforest lying on acid brown earth soil (Dystrochrepts; US De-partment of Agriculture classification) with an AhBwC pro-file. The soil has formed on a loamy, stony solifluxion sheetin which the weathering products of the bedrock (Lower Dev-onian sandstone and schist) were mixed with added perigla-cial loess. The soil exchange complex is dominated by Al,and the base saturation is less than 20% in all soil horizons.In the Ah horizon, pH (H2O) is approximately 4 and increaseswith depth, becoming 4.5–5.0 at 1 m depth. According to theFrench classification (Brethes et al. 1995), the humus formunder beech is eumoder with well-developed OL and OFhorizons and a thin OH horizon (total forest floor thickness3.5 cm); the humus form under oak is oligomull with an OLhorizon, a thin OF horizon, and no OH horizon (total forestfloor thickness 1.5 cm). Ponge et al. (1997) and David et al.(1993) have shown that, in the Belgian Ardennes, oligomullhumus is characterized by a higher diversity of macrofaunalgroups (earthworms, mollusks, woodlice, and millipeds),whereas eumoder humus is mainly colonized by mesofauna.
By the end of the 19th century, the forest was probably anoak coppice with a few standards. Taking advantage of themassive oak regeneration in the 1880s, the forest devel-oped progressively into a high forest and was then invadedby beech. In 2001, the area was covered by even-aged oaktrees and heterogeneously sized beech trees. In addition, anunderstory of hornbeam (Carpinus betulus L.) occurred inthe areas dominated by oak.
Experimental designFour plots were established on the site: two plots in stands
dominated either by oak (0.65 ha) or by beech (0.65 ha) andtwo plots with a mixture of both species; one had a basalarea equivalent to that of the pure plots (mixture; 0.51 ha),and the other had a higher basal area (dense mixture;1.03 ha). These plots are all situated on the same tableland(305–312 m elevation) and were selected in such a way thatspecies composition was the main varying factor. The beechand mixed plots are located side by side, the oak plot is600 m away from them, and the dense mixed plot is locatedhalfway between the oak and the two other plots. Soil ho-mogeneity was evaluated on the basis of a detailed charac-terization of the Ah and Bw horizons of the plots (Table 1).In each plot, all the trees were measured (stem circumfer-ence at a height of 1.3 m and total tree height) and posi-tioned (coordinates) in 2001 (Table 2).
Each experimental plot was subdivided into triangularzones delimited by its own trees. This subdivision was per-formed with ArcView1 GIS software on the basis of thetree coordinates using the triangular irregular network (TIN)function. The basal area and species composition of eachzone were then computed using the trunk circumference ofthe trees.
Six zones (10–55 m2) were selected in each plot to ensureregular repartition within the plot and in such a way thattheir species composition matched that of the plot. In twozones of each of the pure plots and of the mixture, dry sub-zones (3 m � 2.5 m) were created using transparent polycar-bonate roofs (mean height 1.5 m, slope: 20%). Throughfallintercepted by the roofs was directed to gutters connected totubes collecting some of the water in plastic barrels andevacuating the remainder 5 m away from the subzones.Transparent panels were used to make walls that minimizedrainfall blowing under the roof; however, a space of about1 m between the ground and the edge of the panels was leftuncovered to ensure air circulation. The dry subzones werewatered weekly with 56 L of throughfall water, exceptwhen the water barrels were empty because of prolongeddrought periods; they received a mean of 320 mm�year–1
(50%–60% of the throughfall volume). We used only thecentral part (1 m � 1.5 m) of the subzones for the litterbagincubation to avoid edge effects.
Leaf litter production and chemical compositionIn August 2001, litter traps (0.7 m � 0.7 m) were placed
in the four stands (21, 17, 40, and 23 traps in the oak,mixed, dense mixed, and beech plot, respectively, for a totalof 101). Litterfall was collected once a year after leaf shed-ding in 2001, 2002, and 2003. The collected samples wereair-dried and sorted into leaves and other litter materials.All components were weighed and subsamples were oven-dried at 65 8C for 48 h to convert fresh mass into dry mass.More detailed information on leaf litter collection can be ob-tained in Jonard et al. (2006).
Chemical analyses were performed only on the leaf littercollected in 2001. Elemental analyses (K, Mg, Ca, Na, Mn,Al, Fe, and P) were carried out by inductively coupled plasmaspectrometry after combustion in a muffle furnace at450 8C for 12 h and acid digestion with a mixture ofHNO3–HClO4–HF (Pinta 1971). Total C and N contentswere determined using a CN analyzer (FlashEA 1112 Series).The proportion of lignin, cellulose, hemicellulose, and solu-
Jonard et al. 529
# 2008 NRC Canada

ble compounds was determined by using a slightly modi-fied version of the Van Soest (1963) method, in which thesoluble compounds are extracted with an ethanol–benzenesolution.
Leaf litter decomposition
Preparation of litterbagsThe leaves collected in all plots in December 2001 were
pooled by species, homogenized by hand mixing, and usedfor filling the litterbags. Litterbags consisted of two nets ofdifferent mesh size pasted together (12 cm � 12 cm). A finemesh (2 mm) was used for the sheet lying on the ground tolimit small litter losses and a coarse mesh (7 mm) was usedfor the top sheet to allow soil fauna entrance. The bags werepacked with 4.3 g (oven-dried mass) of either pure beech,pure oak, or 1:1 (w/w) mixed leaf litter; this amount wasequivalent to the leaf fall in 2001 on an areal basis. Anotherset of bags was filled with 3.86 g (oven-dried mass) of cel-lulose (Whatman No. 40 filter).
Incubation and retrieval of litterbagsIn early November 2002, the single-species litterbags
were incubated in the six zones of each plot, whereas themixed-species and cellulose litterbags were incubated inonly three of the six zones in all plots except in the densemixture. In the dry subzones, only single-species litterbagswere incubated. The litterbags were placed in the upper hol-organic layer and covered with freshly fallen litter. After 3,6, 9, 12, 24, and 36 months of exposure, litterbags were col-lected in plastic bags to avoid losses. On each samplingdate, three litterbags per litter type were collected in allzones and in the dry subzones, which resulted in a total of1404 litterbags collected over the 3 year period. After re-moval of external debris and separation of oak and beechleaves in the mixed-species litterbags (except for the lastcollection due to the high degree of decomposition), littersamples were oven-dried at 65 8C and weighed. The per-centage of organic matter in the litter samples was deter-mined by loss on ignition (450 8C in a muffle furnace for24 h).
Climate measurementIn all zones of each plot and in the dry subzones, soil
temperature and volumetric soil water content were meas-ured hourly, using probes (thermocouples for soil tempera-ture and TDR for soil water content, model CS615;Campbell Scientific Ltd., Courtaboeuf, France) inserted inthe hemiorganic horizon at about 2 cm below the base ofthe holorganic layers. The monitoring of ground climatewas carried out from November 2002 to November 2005.Air temperature and precipitation were monitored duringthe same period in an automatic meteorological station lo-cated in an open field 200 m away from the forest site.
Leaf litter accumulationFrom 25 February to 5 March 2002, the forest floor sam-
pling was carried out in the six zones of each plot, using atemplate of 30 cm � 30 cm. The forest floor was collectedin four separate layers according to Brethes et al. (1995):OLn, recently fallen litter; OLv, discoloured litter; OF, frag-T
able
1.Se
lect
edso
ilpr
oper
ties
ofth
efo
urex
peri
men
tal
plot
saf
ter
Jona
rd(2
005)
.
Exc
hang
eabl
eca
tions
(cm
olc�k
g–1)
Tot
alpo
ol(m
g�g–1
)
Plot
and
soil
hori
zon
Dep
th(c
m)
pH(H
2O)
Sand
(%)
Silt
(%)
Cla
y(%
)K
Mg
Ca
Na
EC
EC
aC
NK
Mg
Ca
Na
Oak A
h0–
63.
9610
6228
0.50
0.55
1.38
0.21
12.5
179
.01
7.50
13.1
1.1
0.4
1.8
Bw
6–27
4.28
1154
350.
090.
090.
430.
154.
9017
.36
0.87
17.7
2.0
0.4
3.1
Den
sem
ixtu
reA
h0–
63.
8215
7411
0.37
0.39
1.68
0.22
12.4
972
.20
5.06
13.6
1.4
0.9
2.9
Bw
6–20
4.37
1461
250.
090.
080.
370.
144.
2723
.17
1.01
16.6
1.8
0.8
3.5
Mix
ture
and
beec
hA
h0–
84.
1514
6224
0.28
0.23
1.33
0.21
10.4
112
1.49
3.67
15.0
1.3
0.6
2.4
Bw
8–20
4.27
1560
250.
130.
080.
550.
225.
9828
.57
1.04
17.0
1.4
0.5
2.6
aE
CE
C,
effe
ctiv
eca
tion-
exch
ange
capa
city
:K
+ +M
g2++
Ca2+
+N
a++
Mn2+
+H
+ +A
l3+.
530 Can. J. For. Res. Vol. 38, 2008
# 2008 NRC Canada

mented litter mixed with faecal pellets; OH, well-decom-posed and humified organic matter. The samples were air-dried, sorted to remove twigs, fruits and coarse roots(>2 mm), and finally weighed. Subsamples were oven-driedat 65 8C during 48 h to convert fresh mass into dry mass. Itis important to note that the forest floor was sampled 2months after the peak of leaf litterfall and that its mass wasclose to the annual maximum.
In addition to the litterbag technique, we used the massbalance method to evaluate leaf litter decomposition. Whenthe steady state is reached, the mean annual decompositionrate (kmb) of the forest floor can be estimated from the an-nual litterfall (L in kg OM�ha–1�year–1) and the forest floormass (FF in kg OM�ha–1) according to the mass balanceequation (Jenny et al. 1949):
½1� kmb ¼L
FF
This method was applied to estimate the annual leaf decom-position rate using the annual leaf litterfall and to make arough estimate of the accumulated amount of decomposingleaves (forest floor samples from which coarse debris wasexcluded).
These estimates were compared with the mean annual de-composition rates obtained by the litterbag technique (klb).The latter was calculated from the current annual decompo-sition rates computed for each incubation year by dividingthe mass loss during the considered year by the remainingmass at the beginning of that year.
Statistical analysesMixed linear models were used to test (i) for the effects
of stand composition on leaf litterfall, leaf litter accumula-tion, and ground climate and (ii) for the effects of littertype, litter mixture, stand composition, and drought on theremaining litterbag mass expressed as the percentage of ini-tial organic matter content. Concerning the litterbag experi-ment, each sampling date was analyzed separately to avoidproblems of heteroscedasticity (Wider and Lang 1982). Inall cases, the zone was considered to be a random effect.Contrasts or multiple comparison tests (Tukey) were used to
compare factors levels. For example, we used contrasts totest whether the mass loss of the mixed litter differed fromthe mean mass loss of the corresponding pure litters. Allanalyses were carried out with the MIXED procedure of theSAS software (version 8.20; SAS Institute Inc., Cary, N.C.).
Because there is only one plot per species composition(n = 1), the stand composition effect cannot be evaluatedstatistically; however, the plot effect can be tested becausemeasurements were repeated within the plots (n = 2, 3, or6 zones). In the following sections, we discuss the plot ef-fect as a stand composition effect, assuming that it is themain varying factor between plots.
Results
Leaf litter production and chemical compositionTotal annual amounts of leaf litter were similar in the
oak, mixed, and beech stands and somewhat higher in thedense mixture (P < 0.001). However, the species composi-tion of leaf litter varied according to stand composition(Fig. 1a).
Compared with beech, oak leaves had significantly lowerP and K concentrations and higher Ca, Mg, and Mn concen-trations; N and C concentrations were similar for both spe-cies. Oak and beech leaves contained approximately thesame proportions of lignin. The cellulose proportion washigher in beech leaves, whereas oak leaves contained morehemicellulose and soluble compounds (Table 3).
Leaf litter decomposition
Litter quality effectsIn the single-species litterbags, oak leaf litter disappeared
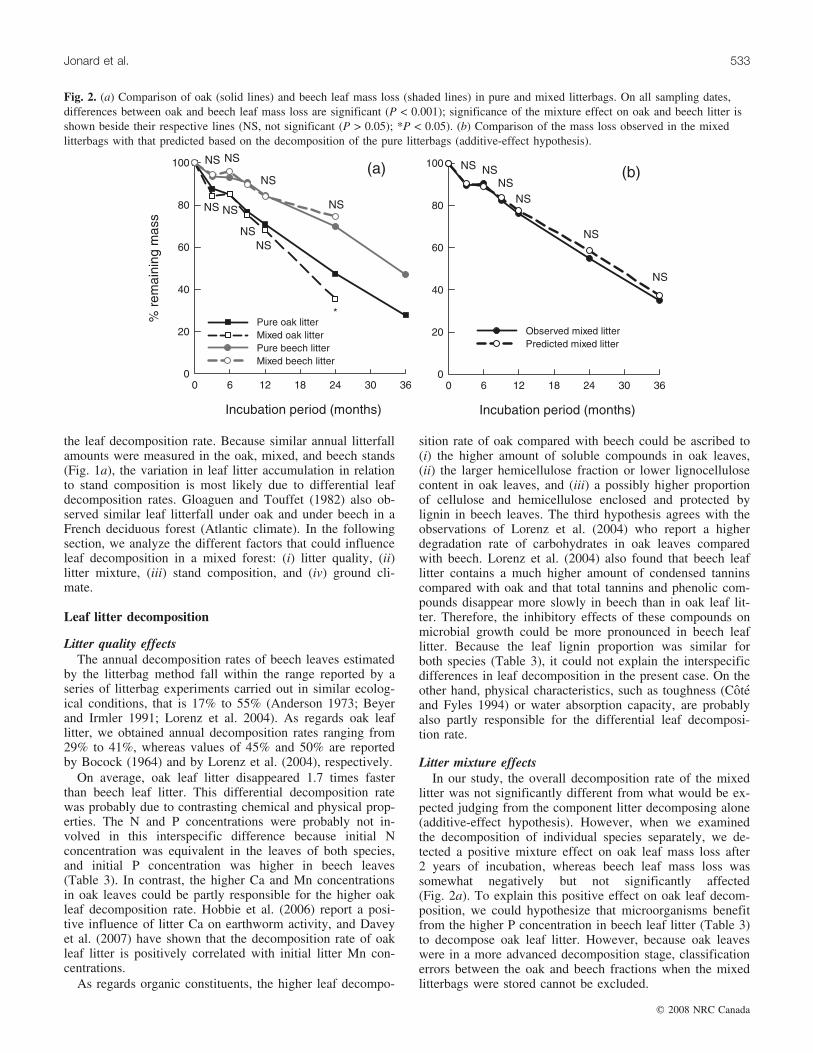
at mean rates of 29%, 33%, and 41%�year–1 during the first,second, and third incubation years, whereas the decomposi-tion rate of beech leaf litter amounted to 16%, 17%, and33%�year–1, respectively. Over the whole incubation period,oak leaf litter disappeared on average 1.7 times faster thanbeech leaf litter (Fig. 2a). Leaf mixture significantly increasedoak leaf mass loss after 24 months of exposure (P = 0.002),whereas the mixture effect on beech leaf was slightly neg-ative but not significant (P = 0.219) (Fig. 2a). On all sam-
Table 2. Mean stand characteristics (SD in parentheses) of the four experimental plots in 2001.
Height (m)
Plot (plot area) and species No.�ha–1 C130 (cm)a Basal area (m2�ha–1) Top Meanb
Oak (0.65 ha)Oak 197 101 (27) 16.9 22.5 21 (2.3)Beech 125 46 (36) 3.4 14 (4.9)
Mixture (0.51 ha)Oak 101 108 (19) 9.7 24 23 (1.5)Beech 135 101 (35) 12.3 25.5 22 (5.6)
Dense mixture (1.03 ha)Oak 122 116 (21) 13.3 26 24 (1.7)Beech 351 66 (39) 16.5 27.5 18 (6.9)
Beech (0.65 ha)Oak 49 94 (13) 3.5 23 (1.2)Beech 286 82 (41) 19.5 25.5 19 (6.3)
aStem circumference at a height of 130 cm.bMean height of the 100 highest trees per hectare.
Jonard et al. 531
# 2008 NRC Canada

pling dates, the overall mass loss of the mixed litter wasslightly higher than might be expected based on the addi-tive effects of the component litter decomposing alone;however, the difference was insignificant (Fig. 2b).
Stand composition effectsStand composition had no significant effects on oak and
beech leaf decomposition after 1 year (Figs. 3a and 3b). Afterthe second incubation year, we noticed a lower mass lossof oak leaves in the oak stand compared with the otherstands (Fig. 3a) and a lower mass loss of beech leavesunder the oak and mixed stands compared with the beechstand and the dense mixture (Fig. 3b); these differenceswere not significant. After the third incubation year, the re-maining mass of oak leaf litter was lower under beech thanunder oak (P = 0.027) (Fig. 3a), whereas the increasingdifferences between stands were still not significant wherebeech leaves were concerned (Fig. 3b). The disappearanceof cellulose filters started after 6 months of incubation andwas nearly completed after 2 years. There were no signifi-cant differences among incubation stands (Fig. 3c).
Ground climate effectsDuring the growing period (14 May to 12 November), the
mean soil temperature was similar in the oak and mixedstands, whereas it was 0.3–0.4 8C and 0.6–0.8 8C lower inthe beech stand and the dense mixture, respectively(Table 4). During the dormant period (13 November to 13May), the differences between stands were generally not sig-nificant, except in the dormant period 2002–2003 duringwhich the mean soil temperature of the dense mixed standwas 0.2–0.3 8C higher than in the other stands (Table 4).
The differences in soil water content between the oak,mixed, and beech stands were not significant for both peri-ods; however, the dense mixture was somewhat moisterthan the mixture during the growing period (Table 4).
The roofs reduced the mean soil water content by 18%and 14% during the dormant and growing periods, respec-tively (P < 0.05). However, roof impact varied with yearand stand composition; it was never significant under oak,except during the 2002–2003 dormant period, and was morepronounced under mixture than under beech (Table 5).
The leaf mass loss of both species was progressively re-duced by drought (Fig. 4). After 6 months of incubation,the mass loss of oak leaves declined significantly in the drymodality of all stand types; however, under oak, the differ-ence in leaf mass loss in the two moisture modalities disap-peared after 2 years. As regards beech leaves, the depressiveeffect of drought appeared, respectively, after 1 and 3 yearsunder mixture and beech but was never significant underoak.
Leaf litter accumulationThe organic matter stored in the forest floor was 2.6 times
greater under beech than under oak (Fig. 1b). The store inthe mixed stand was not different from what would be ex-pected judging from the amounts observed in pure standsand considering an equal proportion of the two species (P =0.241) (Fig. 1b). The quantity of organic matter stored in thedense mixture was slightly lower than in the mixed stand,but the difference was not significant (P = 0.175) (Fig. 1b).
Based on the mean annual leaf litter inputs and on thestore of accumulated leaf litter, the mean annual leaf decom-position rates calculated using the mass balance equationwere 67%, 41%, 54%, and 25% under oak, mixture, densemixture and beech, respectively.
Discussion
On our site as well as in the other oak and beech forestsof the western Ardennes, leaf litter accumulation increasesin proportion to the local beech abundance for a given grow-ing stock level (Fig. 1b). This pattern could be explained ei-ther by an increase in leaf litter inputs or by a decrease in
Fig. 1. (a) Mean leaf litterfall and (b) forest floor mass in the fourstands. The error bars are the 95% confidence intervals.
Table 3. Initial element concentrations (mg�g–1) and or-ganic constituents (%) of oak and beech leaf litter.
Oak Beech PC 520.0 523.0 0.238N 11.8 11.4 0.090P 0.4 0.5 <0.001K 1.2 1.4 0.003Ca 8.8 7.7 <0.001Mg 1.1 0.7 <0.001Mn 5.2 4.6 0.027Lignin 33.8 37.2 0.136Cellulose 20.7 32.8 <0.001Hemicellulose 33.2 20.9 <0.001Soluble 8.4 5.0 <0.001
Note: The P values give the significance of the interspecificdifferences evaluated with a one-way ANOVA (n = 17 for theelement concentrations and n = 3 for the organic constituents).
532 Can. J. For. Res. Vol. 38, 2008
# 2008 NRC Canada
the leaf decomposition rate. Because similar annual litterfallamounts were measured in the oak, mixed, and beech stands(Fig. 1a), the variation in leaf litter accumulation in relationto stand composition is most likely due to differential leafdecomposition rates. Gloaguen and Touffet (1982) also ob-served similar leaf litterfall under oak and under beech in aFrench deciduous forest (Atlantic climate). In the followingsection, we analyze the different factors that could influenceleaf decomposition in a mixed forest: (i) litter quality, (ii)litter mixture, (iii) stand composition, and (iv) ground cli-mate.
Leaf litter decomposition
Litter quality effectsThe annual decomposition rates of beech leaves estimated
by the litterbag method fall within the range reported by aseries of litterbag experiments carried out in similar ecolog-ical conditions, that is 17% to 55% (Anderson 1973; Beyerand Irmler 1991; Lorenz et al. 2004). As regards oak leaflitter, we obtained annual decomposition rates ranging from29% to 41%, whereas values of 45% and 50% are reportedby Bocock (1964) and by Lorenz et al. (2004), respectively.
On average, oak leaf litter disappeared 1.7 times fasterthan beech leaf litter. This differential decomposition ratewas probably due to contrasting chemical and physical prop-erties. The N and P concentrations were probably not in-volved in this interspecific difference because initial Nconcentration was equivalent in the leaves of both species,and initial P concentration was higher in beech leaves(Table 3). In contrast, the higher Ca and Mn concentrationsin oak leaves could be partly responsible for the higher oakleaf decomposition rate. Hobbie et al. (2006) report a posi-tive influence of litter Ca on earthworm activity, and Daveyet al. (2007) have shown that the decomposition rate of oakleaf litter is positively correlated with initial litter Mn con-centrations.
As regards organic constituents, the higher leaf decompo-
sition rate of oak compared with beech could be ascribed to(i) the higher amount of soluble compounds in oak leaves,(ii) the larger hemicellulose fraction or lower lignocellulosecontent in oak leaves, and (iii) a possibly higher proportionof cellulose and hemicellulose enclosed and protected bylignin in beech leaves. The third hypothesis agrees with theobservations of Lorenz et al. (2004) who report a higherdegradation rate of carbohydrates in oak leaves comparedwith beech. Lorenz et al. (2004) also found that beech leaflitter contains a much higher amount of condensed tanninscompared with oak and that total tannins and phenolic com-pounds disappear more slowly in beech than in oak leaf lit-ter. Therefore, the inhibitory effects of these compounds onmicrobial growth could be more pronounced in beech leaflitter. Because the leaf lignin proportion was similar forboth species (Table 3), it could not explain the interspecificdifferences in leaf decomposition in the present case. On theother hand, physical characteristics, such as toughness (Coteand Fyles 1994) or water absorption capacity, are probablyalso partly responsible for the differential leaf decomposi-tion rate.
Litter mixture effectsIn our study, the overall decomposition rate of the mixed
litter was not significantly different from what would be ex-pected judging from the component litter decomposing alone(additive-effect hypothesis). However, when we examinedthe decomposition of individual species separately, we de-tected a positive mixture effect on oak leaf mass loss after2 years of incubation, whereas beech leaf mass loss wassomewhat negatively but not significantly affected(Fig. 2a). To explain this positive effect on oak leaf decom-position, we could hypothesize that microorganisms benefitfrom the higher P concentration in beech leaf litter (Table 3)to decompose oak leaf litter. However, because oak leaveswere in a more advanced decomposition stage, classificationerrors between the oak and beech fractions when the mixedlitterbags were stored cannot be excluded.
(a)
Incubation period (months)
0 6 12 18 24 30 36
%re
mai
ning
mas
s
0
20
40
60
80
100
Pure oak litterMixed oak litterPure beech litterMixed beech litter
(b)
Incubation period (months)
0 6 12 18 24 30 360
20
40
60
80
100
Observed mixed litterPredicted mixed litter
NS NS
NSNS
NS
NS
NS
NS
*
NS
NSNS
NS
NS
NS
Fig. 2. (a) Comparison of oak (solid lines) and beech leaf mass loss (shaded lines) in pure and mixed litterbags. On all sampling dates,differences between oak and beech leaf mass loss are significant (P < 0.001); significance of the mixture effect on oak and beech litter isshown beside their respective lines (NS, not significant (P > 0.05); *P < 0.05). (b) Comparison of the mass loss observed in the mixedlitterbags with that predicted based on the decomposition of the pure litterbags (additive-effect hypothesis).
Jonard et al. 533
# 2008 NRC Canada

Stand composition effectsDuring the first 2 years of incubation, the environmental
effects associated with the incubation stand were not signifi-cant (Fig. 3), probably because of limited differences in
ground climate (Table 4). However, the stand compositioneffects appeared gradually with time and became significantfor oak leaves after an incubation period of 36 months(Fig. 3); this may have occurred because the litterbags wereperiodically covered with fresh leaf litter and sank progres-sively into the forest floor, where decomposition conditionswere more favorable. Indeed, accumulated decomposing or-ganic matter has a greater water absorption capacity thanfreshly fallen litter and maintains higher water contents dur-ing drought periods (Virzo De Santo et al. 1993). In addi-tion, release of nutrients and easily utilizable C from theoverlying organic layers could also favor leaf decomposition(priming effect). We would expect these effects to be morepronounced on thick forest floors, which could explain thehigher leaf decomposition rate observed in the beech stand(Fig. 3). Similarly, Binkley (1984), and Piene and VanCleve (1978) observed a higher cellulose mass loss in bagsincubated in the deeper layers of the forest floor. The differ-ence in decay patterns between stand types could also be re-lated to the temporal changes in leaf litter chemistry; themicroflora of the beech forest floor is probably more effi-cient at decomposing the recalcitrant compounds accumulat-ing in litter during the late decay stages. Finally, the effectsof stand composition on leaf decomposition might be parti-ally confounded with other factors such as soil properties,because there is just one plot per stand type (pseudorepli-cates).
Ground climate effectsThe ground climate differences between stands (Table 4)
could partly be attributed to their contrasting canopy cover.In the same stands, Jonard (2005) observe a higher leaf areaindex under beech (6.1) than under oak and mixture (5.2 and5.1, respectively), which can be related to the lower meansoil temperature measured under beech during the growingperiod (Table 4). Concerning the mixed stands, the higherleaf area index in the dense mixture (6.3) was associatedwith a lower mean soil temperature and an higher mean soilwater content during the growing period (Table 4).
In turn, these differences in ground climate could affectleaf litter decomposition; Jonard et al. (2007) show that theheterotrophic respiration of the forest floor increased expo-nentially with soil temperature, and we observed that themass losses in oak and beech leaves after 3 years of incuba-tion were, respectively, 24% and 17% lower in the dry mo-dality (Fig. 4). Taylor and Wolters (2005) report an annualmass loss of 22.3% and 4.4%, respectively, for beech leavesincubated in normal conditions and for those under a roof(no watering). Compared with our study, this drought effectwas much more acute, probably because of the marked dif-ferences in moisture conditions. Salamanca et al. (2003) car-ried out a rainfall manipulation experiment using threetreatments: fully covered, partially covered (50% of rain-fall), and control. In the partially covered and control treat-ments, litter moisture and litter decomposition werecomparable, whereas they fell by 50% and 23%, respec-tively, in the fully covered modality. The dry modality ofour study could be halfway between the fully and partiallycovered modalities of Salamanca et al. (2003). Indeed, thewater inputs to the dry modality were equivalent to 50%–60% of throughfall; however, part of this water volume was
0 6 12 18 24 30 36
%re
ma
inin
gm
ass
0
20
40
60
80
100
0 6 12 18 24 30 36
%re
ma
inin
gm
ass
0
20
40
60
80
100
Incubation period (months)
0 6 12 18 24 30 36
%re
ma
inin
gm
ass
0
20
40
60
80
100
NSNS
NS
NS
NS
*
* NSNS
NS
NS
NS
NS NS
NS
NS
NS
NS
Oak leaves
Beech leaves
Cellulose filters
Oak stand
Mixture
Dense mixture
Beech stand
(a)
(b)
(c)
Fig. 3. Stand effects on the mass loss of (a) oak and (b) beechleaves and (c) cellulose filters. Significance of the stand effect isindicated as follows: NS, not significant; *P < 0.05.
534 Can. J. For. Res. Vol. 38, 2008
# 2008 NRC Canada
probably rapidly leached below the forest floor, because theintensity associated with the weekly watering (90 mm�h–1)was higher than the rainfall intensities usually occurringunder the canopy (1–10 mm�h–1).
Other factors influenced by the roofs could have affectedthe differences in leaf litter decomposition observed betweenthe two moisture modalities. For example, roofs could havecreated a greenhouse effect and increased soil temperature.However, air circulation under the roofs probably limitedthis potential effect. Unfortunately, we have no data to ver-ify this hypothesis.
Comparison of the litterbag and the mass balanceestimates
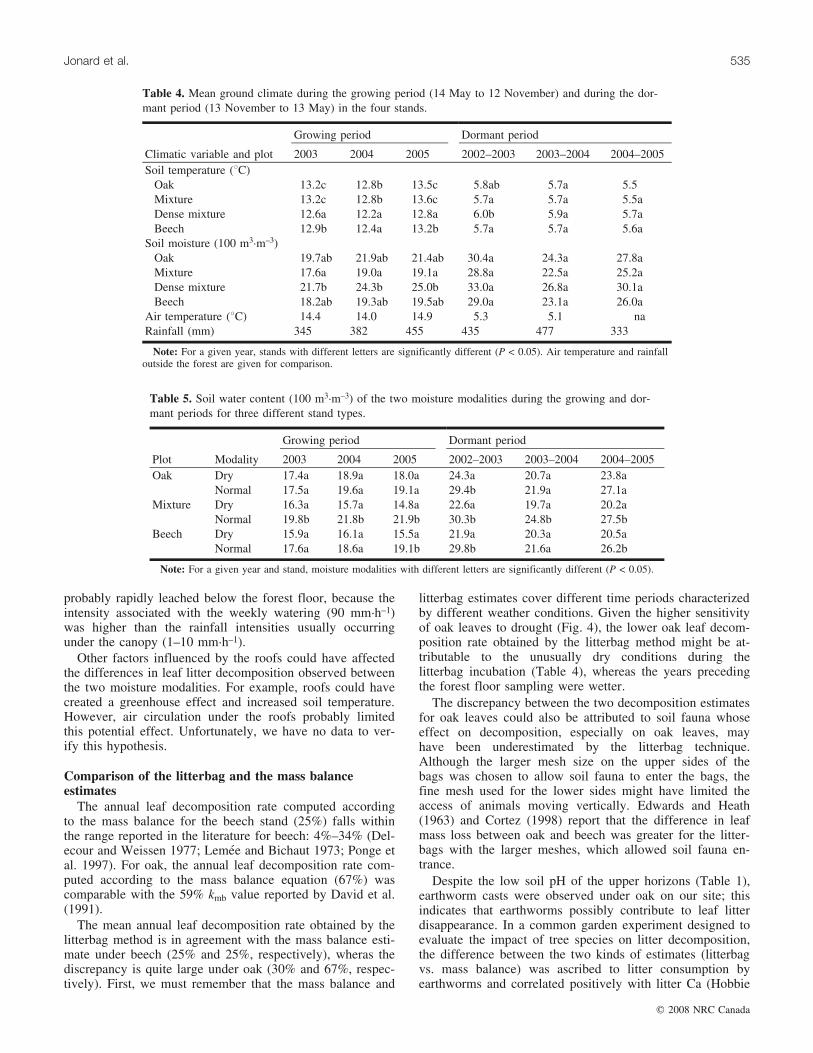
The annual leaf decomposition rate computed accordingto the mass balance for the beech stand (25%) falls withinthe range reported in the literature for beech: 4%–34% (Del-ecour and Weissen 1977; Lemee and Bichaut 1973; Ponge etal. 1997). For oak, the annual leaf decomposition rate com-puted according to the mass balance equation (67%) wascomparable with the 59% kmb value reported by David et al.(1991).
The mean annual leaf decomposition rate obtained by thelitterbag method is in agreement with the mass balance esti-mate under beech (25% and 25%, respectively), wheras thediscrepancy is quite large under oak (30% and 67%, respec-tively). First, we must remember that the mass balance and
litterbag estimates cover different time periods characterizedby different weather conditions. Given the higher sensitivityof oak leaves to drought (Fig. 4), the lower oak leaf decom-position rate obtained by the litterbag method might be at-tributable to the unusually dry conditions during thelitterbag incubation (Table 4), whereas the years precedingthe forest floor sampling were wetter.
The discrepancy between the two decomposition estimatesfor oak leaves could also be attributed to soil fauna whoseeffect on decomposition, especially on oak leaves, mayhave been underestimated by the litterbag technique.Although the larger mesh size on the upper sides of thebags was chosen to allow soil fauna to enter the bags, thefine mesh used for the lower sides might have limited theaccess of animals moving vertically. Edwards and Heath(1963) and Cortez (1998) report that the difference in leafmass loss between oak and beech was greater for the litter-bags with the larger meshes, which allowed soil fauna en-trance.
Despite the low soil pH of the upper horizons (Table 1),earthworm casts were observed under oak on our site; thisindicates that earthworms possibly contribute to leaf litterdisappearance. In a common garden experiment designed toevaluate the impact of tree species on litter decomposition,the difference between the two kinds of estimates (litterbagvs. mass balance) was ascribed to litter consumption byearthworms and correlated positively with litter Ca (Hobbie
Table 4. Mean ground climate during the growing period (14 May to 12 November) and during the dor-mant period (13 November to 13 May) in the four stands.
Growing period Dormant period
Climatic variable and plot 2003 2004 2005 2002–2003 2003–2004 2004–2005Soil temperature (8C)
Oak 13.2c 12.8b 13.5c 5.8ab 5.7a 5.5Mixture 13.2c 12.8b 13.6c 5.7a 5.7a 5.5aDense mixture 12.6a 12.2a 12.8a 6.0b 5.9a 5.7aBeech 12.9b 12.4a 13.2b 5.7a 5.7a 5.6a
Soil moisture (100 m3�m–3)Oak 19.7ab 21.9ab 21.4ab 30.4a 24.3a 27.8aMixture 17.6a 19.0a 19.1a 28.8a 22.5a 25.2aDense mixture 21.7b 24.3b 25.0b 33.0a 26.8a 30.1aBeech 18.2ab 19.3ab 19.5ab 29.0a 23.1a 26.0a
Air temperature (8C) 14.4 14.0 14.9 5.3 5.1 naRainfall (mm) 345 382 455 435 477 333
Note: For a given year, stands with different letters are significantly different (P < 0.05). Air temperature and rainfalloutside the forest are given for comparison.
Table 5. Soil water content (100 m3�m–3) of the two moisture modalities during the growing and dor-mant periods for three different stand types.
Growing period Dormant period
Plot Modality 2003 2004 2005 2002–2003 2003–2004 2004–2005Oak Dry 17.4a 18.9a 18.0a 24.3a 20.7a 23.8a
Normal 17.5a 19.6a 19.1a 29.4b 21.9a 27.1aMixture Dry 16.3a 15.7a 14.8a 22.6a 19.7a 20.2a
Normal 19.8b 21.8b 21.9b 30.3b 24.8b 27.5bBeech Dry 15.9a 16.1a 15.5a 21.9a 20.3a 20.5a
Normal 17.6a 18.6a 19.1b 29.8b 21.6a 26.2b
Note: For a given year and stand, moisture modalities with different letters are significantly different (P < 0.05).
Jonard et al. 535
# 2008 NRC Canada

et al. 2006). On the same site, Reich et al. (2005) found apositive correlation between litter Ca and the abundance ofearthworms, particularly Lumbricus terrestris L. On oursite, the higher Ca concentration in oak leaves, comparedwith that of beech leaves, could improve their palatabilityfor macrofauna (Cortez 1998; Ponge et al. 1997). In addi-tion, the presence of hornbeam trees in the understory ofthe oak stand could increase the diversity and the abundanceof soil macrofauna.
Then again we could perhaps ask ourselves whether theforest floor had stabilized and whether harvesting practiceshad interfered with organic matter accumulation. Yet, thelast clear-cutting dated from the 1880s, and the cutting cyclewas quite long (12 years). In addition, the potentially nega-tive effects of ground-skidding are limited because light ma-chines are used and small timber volumes are harvested.
ConclusionThe purpose of our study was to evaluate the relative im-
portance of the different factors whereby species composi-tion can influence leaf litter accumulation. Although totalleaf litter production was not affected by species composi-tion, the leaf decomposition rate varied with litter qualityand with the environmental conditions associated with thevarious stand types. In the same environment, the two spe-cies featured contrasting decomposition rates. Taking themixed litter as a whole, changes in mass loss were not de-tected. However, oak leaf litter decomposition increased inmixed-species litterbags, whereas beech leaf litter decompo-sition decreased. Therefore, it would be worthwhile measur-ing the litter decomposition of each species separatelywithin mixtures, a procedure which is omitted in most ex-periments. We found that the effects of stand compositionon a given leaf litter appeared progressively and could berelated to the burial of the leaf litter in the forest floor. On
the forest floor surface, the ground climate was probablysimilar in all stand types; however, the moisture conditionscertainly differed in the forest floor itself because of the dif-ference in the forest floor thickness of the various stands.We observed that small differences in soil water contentcould bring about substantial differences in decompositionrate. In conclusion, our study showed that litter quality wasthe major factor regulating leaf litter mass loss and accumu-lation in stands with varying proportions of oak and beech.
AcknowledgementsThis study was initiated by the Forestry Department (Di-
vision de la Nature et des Forets,) of the Walloon Region(Belgium) and funded by the Regional Ministry of Agricul-ture through the project Accord-Cadre Recherches Forest-ieres. We thank S. Caja for her collaboration; F. Hardy andF. Plume for their intensive help with fieldwork; and L.Gerlache, K. Henin, and F. Piscaglia for processing thechemical analyses. The manuscript was greatly improved bythe constructive criticism of two anonymous reviewers andthe associate editor.
ReferencesAber, J.D., Melillo, J.M., and McClaugherty, C.A. 1990. Predicting
long-term patterns of mass loss, nitrogen dynamics, and soil or-ganic matter formation from initial fine litter chemistry in tem-perate forest ecosystems. Can. J. Bot. 68: 2201–2208. doi:10.1139/cjb-68-10-2201.
Albers, D., Migge, S., Schaefer, M., and Scheu, S. 2004. Decompo-sition of beech leaves (Fagus sylvatica) and spruce needles(Picea abies) in pure and mixed stands of beech and spruce.Soil Biol. Biochem. 36: 155–164. doi:10.1016/j.soilbio.2003.09.002.
Anderson, J.M. 1973. The breakdown and decomposition of sweetchestnut (Castanea sativa Mill.) and beech (Fagus sylvatica L.)
Incubation period (months)
0 6 12 18 24 30 36
%re
ma
inin
gm
ass
0
20
40
60
80
100
Dry - Under oak
Normal - Under oak
Dry - Under mixture
Normal - Under mixture
Dry - Under beech
Normal - Under beech
Incubation period (months)
0 6 12 18 24 30 360
20
40
60
80
100
NS
*
NS
*
**
**
*
**
**
*
**
*
NS
**
*
NS
**
**
NS
*
NS
NS
NS
**
NS
NS
NS
NS
**
NS
NS
*
NS
NS
*
*
Oak leaves Beech leaves
(a) (b)
Fig. 4. Effect of water treatment on the mass loss of (a) oak and (b) beech leaves in three different stand types. Significance of the watertreatment effect is given in the boxes above the figures for the oak stand (top row), the mixture (middle row), and the beech stand (bottomrow). NS, not significant (P > 0.05); *P < 0.05; **P < 0.01.
536 Can. J. For. Res. Vol. 38, 2008
# 2008 NRC Canada

leaf litter in two deciduous woodland soils I. Breakdown, leach-ing and decomposition. Oecologia, 12: 251–274.
Berg, B., and McClaugherty, C. 2003. Plant litter: decomposition,humus formation, carbon sequestration. Springer-Verlag, Berlin.
Beyer, L., and Irmler, U. 1991. The structure of humus and dy-namic of litter decomposition on a luvisol and a podzol underforest. Pedobiologia (Jena), 35: 368–380.
Binkley, D. 1984. Does forest removal increase rates of decomposi-tion and nitrogen release? For. Ecol. Manage. 8: 229–233.doi:10.1016/0378-1127(84)90055-0.
Blair, J.M., Parmelee, R.W., and Beare, M.H. 1990. Decay rates,nitrogen fluxes, and decomposer communities of single- andmixed-species foliar litter. Ecology, 71: 1976–1985. doi:10.2307/1937606.
Bocock, K.L. 1964. Changes in the amounts of dry matter, nitro-gen, carbon and energy in decomposing woodland leaf litter inrelation to the activities of the soil fauna. J. Ecol. 52: 273–284.doi:10.2307/2257595.
Bray, J.R., and Gorham, E. 1964. Litter production in forests of theworld. Adv. Ecol. Res. 2: 101–157.
Brethes, A., Brun, J.J., Jabiol, B., Ponge, J.F., and Toutain, F.1995. Classification of forest humus forms: a French proposal.Ann. Sci. For. 52: 535–546. doi:10.1051/forest:19950602.
Cortez, J. 1998. Field decomposition of leaf litters: relationshipsbetween decomposition rates and soil moisture, soil temperatureand earthworm activity. Soil Biol. Biochem. 30: 783–793.doi:10.1016/S0038-0717(97)00163-6.
Cote, B., and Fyles, J.W. 1994. Leaf litter disappearance of hard-wood species of southern Quebec: interaction between litterquality and stand type. Ecoscience, 1: 322–328.
Davey, M.P., Berg, B., Emmett, B.A., and Rowland, P. 2007. De-composition of oak leaf litter is related to initial litter Mn con-centrations. Can. J. Bot. 85: 16–24. doi:10.1139/B06-150.
David, J.-F., Ponge, J.-F., Arpin, P., and Vannier, G. 1991. Reactionsof the macrofauna of a forest mull to experimental perturbationsof litter supply. Oikos, 61: 316–326. doi:10.2307/3545239.
David, J.F., Ponge, J.F., and Delecour, F. 1993. The saprophagousmacrofauna of different types of humus in beech forests of theArdenne (Belgium). Pedobiologia (Jena), 37: 49–56.
Delecour, F., and Weissen, F. 1977. Les sols bruns acides desforets de l’Ardenne: definition et amelioration. Ann. Gembloux,83: 27–42.
Edwards, C.A., and Heath, G.W. 1963. The role of soil animals inbreakdown of leaf material. In Soil organisms. Edited by J.Doeksen and J. Van der Drift. North-Holland Publishers, Am-sterdam. pp. 76–84.
Gholz, H.L., Wedin, D.A., Smitherman, S.M., Harmon, M.E., andParton, W.J. 2000. Long-term dynamics of pine and hardwoodlitter in contrasting environments: toward a global model of de-composition. Glob. Change Biol. 6: 751–765. doi:10.1046/j.1365-2486.2000.00349.x.
Gloaguen, J.C., and Touffet, J. 1982. Production de litiere dans unechenaie–hetraie atlantique. Relations avec les caracteres clima-tiques. Rev. For. Fr. 34: 108–118.
Hattenschwiler, S. 2005. Effects of tree species diversity on litterquality and decomposition. In Forest diversity and function:temperate and boreal systems. Edited by M. Scherer-Lorenzen,C. Koner, and E.D. Schulze. Springer-Verlag, Berlin. Ecol.Stud. 176. pp. 149–164.
Hobbie, S.E., Reich, P.B., Oleksyn, J., Ogdahl, M., Zytkowiak, R.,Hale, C., and Karolewski, P. 2006. Tree species effects on de-composition and forest floor dynamics in a common garden.Ecology, 87: 2288–2297. doi:10.1890/0012-9658(2006)87[2288:TSEODA]2.0.CO;2. PMID:16995629.
Jenny, H., Gessel, S.P., and Bingham, F.T. 1949. Comparativestudy of decomposition rates of organic matter in temperate andtropical regions. Soil Sci. 68: 419–432. doi:10.1097/00010694-194912000-00001.
Jonard, M. 2005. Dynamique des litieres foliaires en peuplementspurs et melanges de chene et de hetre. Retombees foliaires et pre-mieres etapes de la decomposition. Ph.D. thesis, Universite cath-olique de Louvain, Faculte d’ingenierie biologique, agronomiqueet environnementale, CIACO, Louvain-la-Neuve, Belgium.
Jonard, M., Andre, F., and Ponette, Q. 2006. Modeling leaf dispersalin mixed hardwood forests using a ballistic approach. Ecology, 87:2306–2318. doi:10.1890/0012-9658(2006)87[2306:MLDIMH]2.0.CO;2. PMID:16995631.
Jonard, M., Andre, F., Jonard, F., Mouton, N., Proces, P., and Pon-ette, Q. 2007. Soil carbon dioxide efflux in pure and mixedstands of oak and beech. Ann. For. Sci. 64: 141–150. doi:10.1051/forest:2006098.
Lemee, G., and Bichaut, N. 1973. Recherches sur les ecosystemesdes reserves biologiques de la foret de Fontainebleau II. Decom-position de la litiere de feuilles des arbres et liberation des bioe-lements. Oecol. Plant. 8: 153–174.
Lorenz, K., Preston, C.M., Kumrei, S., and Feger, K.H. 2004. De-composition of needle/leaf litter from Scots pine, black cherry,common oak and European beech at a conurbation forest site.Eur. J. For. Res. 123: 177–188.
McClaugherty, C., and Berg, B. 1987. Cellulose, lignin and nitro-gen concentrations as rate regulating factors in late stages of for-est litter decomposition. Pedobiologia (Jena), 30: 101–112.
McTiernan, K.B., Ineson, P., and Coward, P.A. 1997. Respirationand nutrient release from tree leaf litter mixtures. Oikos, 78:527–538. doi:10.2307/3545614.
Meentemeyer, V. 1978. Macroclimate and lignin control of litterdecomposition rates. Ecology, 59: 465–472. doi:10.2307/1936576.
Olson, J.S. 1963. Energy storage and the balance of producers anddecomposers in ecological systems. Ecology, 44: 322–331.doi:10.2307/1932179.
Piene, H., and Van Cleve, K. 1978. Weight loss of litter and cellu-lose bags in a thinned white spruce forest in interior Alaska.Can. J. For. Res. 8: 42–46. doi:10.1139/cjfr-8-1-42.
Pinta, M. 1971. Spectrometrie d’absorption atomique. Application al’analyse chimique. Tome II. Masson et Cie, Paris.
Ponge, J.F., Arpin, P., Sondag, F., and Delecour, F. 1997. Soilfauna and site assessment in beech stands of the Belgian Ar-dennes. Can. J. For. Res. 27: 2053–2064. doi:10.1139/cjfr-27-12-2053.
Prescott, C.E., Zabek, L.M., Staley, C.L., and Kabzems, R. 2000.Decomposition of broadleaf and needle litter in forests of BritishColumbia: influences of litter type, forest type, and litter mixtures.Can. J. For. Res. 30: 1742–1750. doi:10.1139/cjfr-30-11-1742.
Reich, P.B., Oleksyn, J., Modrzynski, J., Hobbie, S.E., Eissenstat,D.M., Chorover, J., Chadwick, O.A., Hale, C.M., and Tjoelker,M.G. 2005. Linking litter calcium, earthworms and soil proper-ties: a common garden test with 14 tree species. Ecol. Lett. 8:811–818. doi:10.1111/j.1461-0248.2005.00779.x.
Salamanca, E.F., Kaneko, N., and Katagiri, S. 2003. Rainfall ma-nipulation effects on litter decomposition and the microbial bio-mass of the forest floor. Appl. Soil Ecol. 22: 271–281. doi:10.1016/S0929-1393(02)00153-1.
Taylor, A.R., and Wolters, V. 2005. Responses of oribatid mitecommunities to summer drought: the influence of litter type andquality. Soil Biol. Biochem. 37: 2117–2130. doi:10.1016/j.soilbio.2005.03.015.
Van Soest, P.J. 1963. Use of detergents in the analysis of fibrous
Jonard et al. 537
# 2008 NRC Canada

feeds II. A rapid method for the determination of fiber and lig-nin. J. Assoc. Off. Anal. Chem. 46: 829–835.
Vesterdal, L. 1999. Influence of soil type on mass loss and nutrientrelease from decomposing foliage litter of beech and Norwayspruce. Can. J. For. Res. 29: 95–105. doi:10.1139/cjfr-29-1-95.
Virzo De Santo, A., Berg, B., Rutigliano, F.A., Aflani, A., andFioretto, A. 1993. Factors regulating early-stage decompositionof needle litters in five different coniferous forests. Soil Biol.Biochem. 25: 1423–1433. doi:10.1016/0038-0717(93)90057-I.
Wardle, D.A., Bonner, K.I., and Nicholson, K.S. 1997. Biodiversityand plant litter: experimental evidence which does not supportthe view that enhanced species richness improves ecosystemfunction. Oikos, 79: 247–258. doi:10.2307/3546010.
Wider, R.K., and Lang, G.E. 1982. A critique of the analyticalmethods used in examining decomposition data obtained fromlitter bags. Ecology, 63: 1636–1642. doi:10.2307/1940104.
538 Can. J. For. Res. Vol. 38, 2008
# 2008 NRC Canada