tnfr2 is not associated with rheumatoid arthritis susceptibility in a caucasian population
TRANSCRIPT

Jose Luis Andreu, MD, PhDHospital Universitario Clinica Puerta de HierroMadrid, SpainDomingo Ly-Pen, MD, PhDPrimary Care Unit GandhiMadrid, Spain
DOI 10.1002/art.21200
TNFR2 is not associated with rheumatoid arthritissusceptibility in a Caucasian population
To the Editor:Linkage and association analysis provides evidence of a
role for the TNFR2 gene in the pathology of several inflam-matory diseases and pathologic reactions. Our group originallyreported an association between the TNFR2 exon 6(�196M/R) gene polymorphism and susceptibility to familial,but not sporadic, rheumatoid arthritis (RA) (1). However, itwas not clear whether this polymorphism was functional. Thus,although it was associated with interleukin-6 production insystemic lupus erythematosus patients, it had no effect on theexpression, shedding, or ligand binding of TNFR2 (2). Alter-natively, this association may have been a result of linkagedisequilibrium (LD) between the �196M/R polymorphism andthe true, but as-yet-unidentified, susceptibility variant. Theassociation between the TNFR2 gene and RA susceptibility,therefore, required further investigation. This report providesevidence that the original observation was spurious and re-sulted from errors in genotyping that occurred with the use ofthe earlier and less robust genotyping method.
We performed high-density single-nucleotide polymor-phism (SNP) marker analysis in a Caucasian case–controlcohort. We compared 192 RA probands from 96 affectedsibling pairs with 183 unrelated controls. Details on thesesubjects are reported elsewhere (3). Twelve SNPs that arelocated within, or in close proximity to, the TNFR2 gene (Table1) were selected and genotyped by either RealSNP assay(Sequenom, Cambridge, UK) or 5� allelic discrimination assay(TaqMan; Applied Biosystems, Warrington, UK). In addition,the �196M/R polymorphism (SNP ID: rs1061622) was geno-typed by Pyrosequencing (Biotage, Manchester, UK). Geno-typing assays were performed according to the manufacturer’sguidelines, or modified as described elsewhere (4). The primerand probe details are available by request from the authors. Nodeviation from Hardy-Weinberg expectations was observed ineither cases or controls (P � 0.01). No significant associationwith RA susceptibility was detected for any of the polymor-phisms (P � 0.2 in all comparisons) (Table 1). Extensive LDwas exhibited across the centromeric subregion, but not the 5�or 3� region of the TNFR2 gene (Table 1).
The absence of an association between the �196M/Rpolymorphism and RA susceptibility was unexpected becausethe subjects in the present cohort overlapped with thoseinvestigated in our original study (1). Comparisons betweenthe raw data of both studies highlighted discrepancies in thegenotypes obtained from 28% of samples common to bothstudies. Repeat genotyping of the present cohort by an alter-
native method, namely SNaPshot (Applied Biosystems), dem-onstrated 100% concordance with the Pyrosequencing data(data not shown). In contrast, our earlier study was based ongenotyping by restriction fragment length polymorphism(RFLP), which was widely used at the time. There are 2possible explanations for the erroneous genotyping. These are1) incomplete digestion by the Nla III enzyme, and 2) thepresence of a second, as-yet-unidentified SNP within theenzyme binding site. We investigated the latter hypothesis bysequencing the surrounding region in 8 samples wronglygenotyped as R/R by RFLP (Lark Technologies, Essex, UK).However, no unexpected polymorphisms were identified.
Hence, it is likely that incomplete enzyme digestionwas responsible for the incorrect genotyping. Although incom-plete enzyme digestion has long been recognized as a potentialproblem with RFLP-based genotyping methods, several qual-ity control measures were included in the original study. First,no deviation from Hardy-Weinberg equilibrium was observed.Second, allele frequencies were compared in the controlpopulation and found to be similar to those described inprevious and subsequent reports. Finally, and most impor-tantly, the association with familial RA was replicated in asecond set of RA patients with a family history of RA. Indeed,a subsequent independent European study demonstrated asimilar association with familial RA (5). It remains unclear asto why a systematic bias occurred in genotyping the familialRA cases, but not in sporadic RA cases or controls. Thesefindings serve to indicate the importance of robust genotypingplatforms. Having tested the association of SNPs spanning theentire TNFR2 gene, we now conclude that the gene is notassociated with RA.
Supported by the Arthritis Research Campaign. Dr. Barton’swork is supported by a Wellcome Advanced Fellowship.
Catherine Potter, BScJane Worthington, PhDAlan Silman, FRCPAnne Barton, MRCPUniversity of ManchesterManchester, UK
1. Barton A, John S, Ollier WE, Silman A, Worthington J. Associationbetween rheumatoid arthritis and polymorphism of tumor necrosisfactor receptor II, but not tumor necrosis factor receptor I, inCaucasians. Arthritis Rheum 2001;44:61–5.
2. Morita C, Horiuchi T, Tsukamoto H, Hatta N, Kikuchi Y, ArinobuY, et al. Association of tumor necrosis factor receptor type IIpolymorphism 196R with systemic lupus erythematosus in theJapanese: molecular and functional analysis. Arthritis Rheum 2001;44:2819–27.
3. Barton A, Jury F, Eyre S, Bowes J, Hinks A, Ward D, et al.Haplotype analysis in simplex families and novel analytic ap-proaches in a case–control cohort reveal no evidence of associationof the CTLA-4 gene with rheumatoid arthritis. Arthritis Rheum2004;50:748–52.
4. Barton A, Bowes J, Eyre S, Spreckley K, Hinks A, John S, et al. Afunctional haplotype of the PADI4 gene associated with rheuma-toid arthritis in a Japanese population is not associated in a UnitedKingdom population. Arthritis Rheum 2004;50:1117–21.
5. Dieude P, Petit E, Cailleau-Moindrault S, Osorio J, Pierlot C,
LETTERS 2579

Tab
le1.
Com
pari
son
ofge
noty
pefr
eque
ncie
sbe
twee
nrh
eum
atoi
dar
thri
tisaf
fect
edsi
blin
gpa
irca
ses
and
unre
late
dco
ntro
ls,d
eter
min
edus
ing
am
etho
dth
atac
coun
tsfo
rth
eus
eof
rela
ted
case
s*
SNP
ID
Posi
tion
(1p3
6)(b
uild
34)
Dis
tanc
efr
ompr
eced
ing
SNP,
bp
Gen
otyp
efr
eque
ncy,
no.(
%)
Tre
ndte
stM
ean
LD
acro
ssth
esu
breg
ion
Con
trol
sC
ases
n1/
11/
22/
2n
1/1
1/2
2/2
�2
PR
2(S
D)
D�
(SD
)
5�re
gion
0.34
(0.1
)0.
83(0
.2)
rs59
0368
1193
2997
–13
152
(48)
63(4
0)16
(12)
164
68(4
1)70
(43)
26(1
6)0.
0342
0.85
32rs
9768
8111
9433
0010
,303
163
88(5
4)54
(33)
21(1
3)14
676
(52)
53(3
6)17
(12)
0.00
620.
9373
rs37
6673
011
9502
226,
922
176
123
(70)
49(2
8)4
(2)
172
119
(69)
47(2
7)6
(4)
0.05
080.
8217
rs47
4247
1195
5721
1,14
615
410
6(6
9)45
(29)
3(2
)94
69(7
3)21
(22)
4(4
)0.
0875
0.76
74C
entr
omer
icsu
breg
ion
0.79
(0.1
)0.
93(0
.06)
rs60
3151
1195
7837
2,11
615
266
(43)
77(5
1)9
(6)
139
65(4
7)67
(48)
7(5
)0.
2006
0.65
43rs
6536
6711
9613
543,
517
159
62(3
9)80
(50)
17(1
1)18
375
(41)
81(4
4)27
(15)
0.04
780.
8269
rs10
6162
211
9625
011,
147
130
71(5
5)54
(42)
5(4
)18
010
1(5
6)71
(40)
8(4
)0.
0093
0.92
33rs
5909
7711
9649
062,
405
153
99(6
5)51
(33)
3(2
)17
711
3(6
4)57
(32)
7(4
)0.
1891
0.66
37rs
2352
4911
9677
772,
871
178
95(5
3)74
(42)
9(5
)17
410
0(5
8)65
(37)
9(5
)0.
3229
0.56
99rs
5746
053
1197
1844
4,06
717
510
3(5
9)66
(38)
6(3
)16
610
2(6
1)59
(36)
5(3
)0.
2052
0.65
053�
regi
on0.
1(0
.1)
0.3
(0.2
)rs
2352
1911
9739
012,
057
175
140
(80)
31(1
8)4
(2)
170
140
(82)
30(1
8)0
(0)
0.88
080.
348
rs33
9711
9768
382,
937
155
70(4
5)66
(43)
19(1
2)18
468
(37)
90(4
9)26
(14)
1.09
680.
295
rs10
6163
111
9780
451,
207
158
94(5
9)57
(36)
7(4
)18
610
7(5
8)69
(37)
10(5
)0.
1026
0.74
87
*C
alcu
latio
nsba
sed
onpu
blis
hed
alle
lefr
eque
ncie
sin
dica
ted
that
each
test
ofas
soci
atio
nha
d80
%po
wer
tode
tect
anef
fect
size
of2
atth
e5%
sign
ifica
nce
leve
l.T
hem
etho
dus
edto
com
pare
geno
type
freq
uenc
ies
betw
een
case
san
dco
ntro
lsis
desc
ribe
din
refe
renc
e6.
Lin
kage
dise
quili
briu
m(L
D)
anal
ysis
was
impl
emen
ted
with
Hel
ixT
ree
soft
war
e(G
olde
nH
elix
,).S
NP
�si
ngle
-nuc
leot
ide
poly
mor
phis
m.
2580 LETTERS
Martinez M, et al. Association between tumor necrosis factor receptorII and familial, but not sporadic, rheumatoid arthritis: evidence forgenetic heterogeneity. Arthritis Rheum 2002;46:2039–44.
6. Slager SL, Schaid DJ. Evaluation of candidate genes in case-controlstudies: a statistical method to account for related subjects. Am JHum Genet 2001;68:1457–62.
DOI 10.1002/art.21281
A cartilage-derived self peptide presented by HLA–B27molecules? Comment on the article by Atagunduz et al
To the Editor:Atagunduz et al (1) suggest that self peptides derived
from cartilage proteins can be bound by HLA–B27 molecules,serve as targets for autoimmune cytotoxic T cells, and play arole in the pathogenesis of ankylosing spondylitis (AS). Theyidentified a single peptide (C34, DRASFIKNL) which wasstimulatory for HLA–B27–restricted CD8� T cells from someAS patients. However, as the authors point out, an N-terminalacidic residue has so far not been observed in peptides elutedfrom HLA–B27 complexes (2), making it crucial to demon-strate the unequivocal existence of the HLA–B27/C34 complexin order to establish C34 as an immunodominant candidatearthritogenic peptide (1).
Atagunduz et al attempt to provide the above-described evidence using a refolding protocol for HLA–B27complexes (Figure 2 in their report). This protocol differs fromthe commonly employed procedure (3) in several importantaspects, however, most notably the absence of a redox system.
With the standard procedure, crystallization of a wide range ofHLA/peptide complexes, including various HLA–B27 mole-cules (ref. 4 and references therein), has been achieved. This isnot the case with the system used by Atagunduz et al (1).Chromatographic separation of the protein mixture yielded aminute peak at 13.7 ml, which the authors regarded as therefolded, trimeric HLA–B27/C34 complex. According to thecolumn manufacturer, this position corresponds to a MW of�30,000 and may therefore not represent an intact HLA heavychain/�2-microglobulin/peptide complex (expected MW�45,000). Insufficient description of the experimental detailsalso does not allow for an assessment of its composition as judgedby sodium dodecyl sulfate–polyacrylamide gel electrophoresis (1).Therefore, the authors’ assumption that the product peak repre-sents an HLA–B27/C34 complex remains unsubstantiated (3).
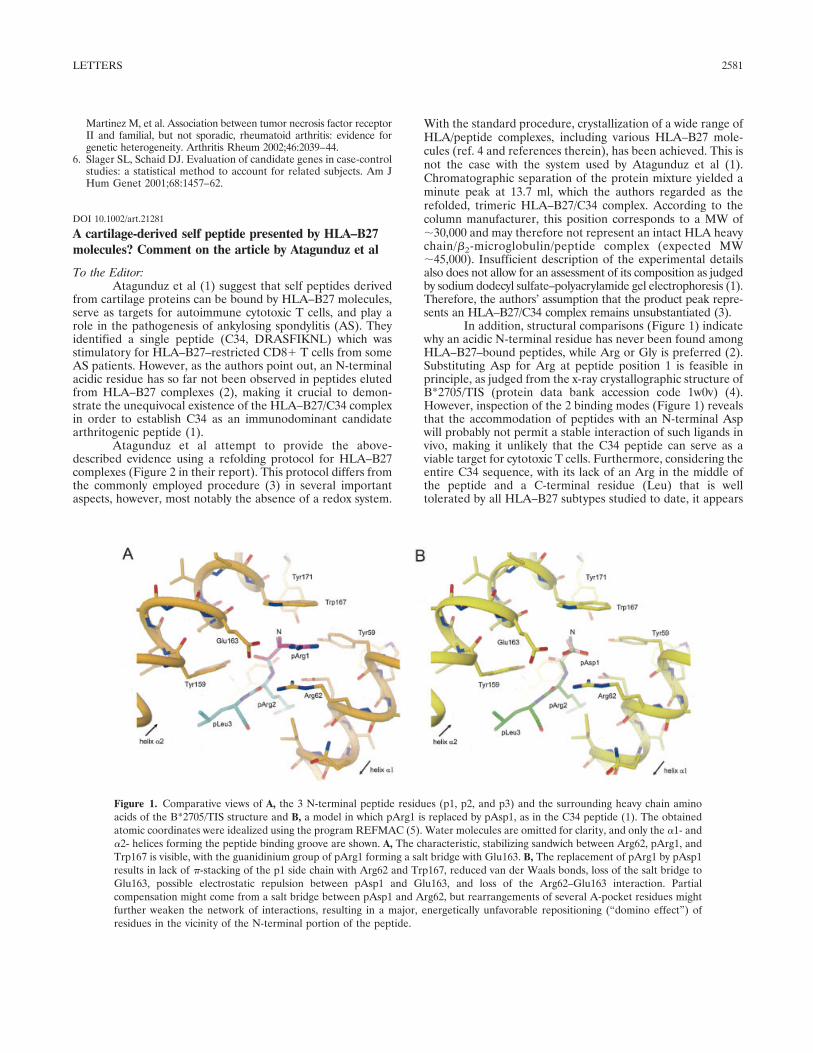
In addition, structural comparisons (Figure 1) indicatewhy an acidic N-terminal residue has never been found amongHLA–B27–bound peptides, while Arg or Gly is preferred (2).Substituting Asp for Arg at peptide position 1 is feasible inprinciple, as judged from the x-ray crystallographic structure ofB*2705/TIS (protein data bank accession code 1w0v) (4).However, inspection of the 2 binding modes (Figure 1) revealsthat the accommodation of peptides with an N-terminal Aspwill probably not permit a stable interaction of such ligands invivo, making it unlikely that the C34 peptide can serve as aviable target for cytotoxic T cells. Furthermore, considering theentire C34 sequence, with its lack of an Arg in the middle ofthe peptide and a C-terminal residue (Leu) that is welltolerated by all HLA–B27 subtypes studied to date, it appears
Figure 1. Comparative views of A, the 3 N-terminal peptide residues (p1, p2, and p3) and the surrounding heavy chain aminoacids of the B*2705/TIS structure and B, a model in which pArg1 is replaced by pAsp1, as in the C34 peptide (1). The obtainedatomic coordinates were idealized using the program REFMAC (5). Water molecules are omitted for clarity, and only the �1- and�2- helices forming the peptide binding groove are shown. A, The characteristic, stabilizing sandwich between Arg62, pArg1, andTrp167 is visible, with the guanidinium group of pArg1 forming a salt bridge with Glu163. B, The replacement of pArg1 by pAsp1results in lack of �-stacking of the p1 side chain with Arg62 and Trp167, reduced van der Waals bonds, loss of the salt bridge toGlu163, possible electrostatic repulsion between pAsp1 and Glu163, and loss of the Arg62–Glu163 interaction. Partialcompensation might come from a salt bridge between pAsp1 and Arg62, but rearrangements of several A-pocket residues mightfurther weaken the network of interactions, resulting in a major, energetically unfavorable repositioning (“domino effect”) ofresidues in the vicinity of the N-terminal portion of the peptide.
LETTERS 2581