tissue carboxylesterase activity of rainbow trout
TRANSCRIPT

2506
Environmental Toxicology and Chemistry, Vol. 18, No. 11, pp. 2506–2511, 1999q 1999 SETAC
Printed in the USA0730-7268/99 $9.00 1 .00
TISSUE CARBOXYLESTERASE ACTIVITY OF RAINBOW TROUT
MACE G. BARRON,*† KEM A. CHARRON,‡ WILLIAM T. STOTT,‡ and STEPHANIE E. DUVALL§†Stratus Consulting, 1881 9th Street, Suite 201, Boulder, Colorado 80302, USA
‡Health and Environmental Research Laboratories, Dow Chemical, 1803 Building, Midland, Michigan 48674, USA§University of Colorado, Environmental Studies Program, Boulder, Colorado 80309, USA
(Received 18 June 1998; Accepted 18 February 1999)
Abstract—The activity of carboxylesterase (CaE), a class of nonspecific serine hydrolases, was evaluated in vitro in tissues andmicrosomes of rainbow trout and compared to esterase activity in rats, other fish species, and embryo to adult life stages of trout.Trout gill and liver microsomes exhibited substantial CaE activity and limited variation over the range of 2 to 408C, with atemperature optimum of approximately 228C. Trout sera and rat liver microsomes exhibited a temperature optimum of approximately35 to 408C. The CaE of trout liver (maximum reaction rate [Vmax] 5 672 nmol/min/mg microsomal protein) was four times lessthan in rats. Apparent Michaelis constant (Km) values ranged from 28 (trout liver) to 214 (trout sera) mM. Values of Vmax/Km
suggested that in vivo CaE activity of trout liver would be about three times higher than serum, 135 times higher than gill, andthree times lower than rat liver. The CaE activity in whole rainbow trout homogenates significantly increased 300% per gram oftissue to 1,200% per milligram of protein between the yolk-sac and juvenile stages. The CaE activity of whole fish homogenateswas not significantly different in juvenile rainbow trout, channel catfish, fathead minnows, and bluegill. The results demonstratethat rainbow trout had high esterase activity over a broad range of temperatures, that CaE activity significantly increased betweenthe yolk-sac and juvenile life stages, and that variation between the CaE activity in trout and three other families of freshwaterfish was limited. The CaE activity in fish is expected to substantially influence the accumulation and toxicity of pesticides andother esters entering the aquatic environment.
Keywords—Xenobiotic metabolism Biotransformation Enzyme kinetics Fish
INTRODUCTION
Carboxylesterases (EC 3.1.1.1) (CaEs) are a class of serinehydrolases that catalyze the cleavage of an alcohol from avariety of ester substrates. Carboxylesterases seem to belongto the B esterase class, which exhibits nearly irreversible phos-phorylation in the presence of organophosphate cholinesteraseinhibitors. Carboxylesterase activity is present in a variety oftissues in vertebrate and invertebrate species [1]. A numberof CaE isozymes have been isolated that even in purified formdisplay broad substrate specificity [2,3].
Investigation of CaE activity in mammalian species hasbeen primarily concerned with the metabolism of prodrugs andaryl ester toxicants. Investigations of biotransformation sys-tems in aquatic animals have primarily focused on oxidationand conjugation pathways, rather than dealkylation activitiesof CaEs. Characterization of CaE activity in aquatic animalsis needed to evaluate species differences in xenobiotic dis-position and to evaluate the environmental fate of ester con-taminants. Only limited investigation of CaE activity in fishhas been conducted, despite the large number of esters presentin the aquatic environment. Herbicide and phthalate esters rep-resent important classes of chemical esters that are extensivelybiotransformed by CaEs [4–7]. Research on a limited numberof ester substrates indicates that fish have high esterase activity,and ester hydrolysis can significantly reduce the bioaccumu-lation of hydrophobic contaminants [4–7]. In addition, recentwork has demonstrated that CaEs protect against the toxicityof organophosphate and organophosphorothioate insecticides(OPs) in both mammals and fish [8,9]. For example, inhibition
* To whom correspondence may be addressed([email protected]).
of CaE activity potentiated the toxicity of OPs and lower he-patic CaE activity in juvenile rats contributed to the greatersensitivity to OP toxicity than adult rats [10,11].
The objective of this study was to quantify CaE activity inrainbow trout (Oncorhynchus mykiss) because this fish speciesis important in aquatic toxicology and carcinogenicity inves-tigations, and is an important ecological receptor for estercontaminants in the environment. The enzyme kinetics (Mi-chaelis constant [Km], maximum reaction rate [Vmax]) and tem-perature dependence of CaE activity of trout gill and livermicrosomes and sera were compared to concurrently measuredCaE activity in rat hepatic microsomes. Additionally, the CaEactivity in whole trout homogenates was compared to activityin channel catfish (Ictalurus punctatus), bluegill (Lepomis ma-crochirus), and fathead minnows (Pimephales promelas). Ho-mogenates of whole trout embryos, yolk-sac fry, juveniles,and adults were assayed to provide insight into the develop-ment of CaE activity in this species. In all experiments, CaEactivity was determined using the model substrate 4-nitro-phenyl acetate because it is a substrate for multiple CaE iso-zymes [2]. The results of this study should have general ap-plicability to a variety of xenobiotic esters.
MATERIALS AND METHODS
Animals and acclimation conditions
Rainbow trout were obtained as eyed embryos from Mt.Lassen Trout Farms (Mt. Lassen, CA, USA) and maintainedat 12 6 18C in a salmonid hatcher until they were free swim-ming. Trout were then transferred to flow-through aerated race-ways maintained at 12 6 18C. Channel catfish and bluegillwere obtained as juveniles from Ossage Catfisheries (OssageBeach, MO, USA) and acclimated to 22 6 18C for a minimum

Carboxylesterase activity in trout Environ. Toxicol. Chem. 18, 1999 2507
of 2 weeks in flow-through aerated holding tanks. Fatheadminnows were produced from laboratory cultures (Dow Chem-ical, Midland, MI, USA) and acclimated to 22 6 18C for aminimum of 2 weeks in flow-through aerated aquaria. Juvenilerainbow trout (1.64 6 0.07 g; wet weight 6 SE), channelcatfish (7.89 6 0.74 g), fathead minnows (1.02 6 0.13 g), andbluegill (1.89 6 0.25 g) were less than one year old whenused. The laboratory water was softened Lake Huron waterthat had been sand-filtered, pH adjusted with CO2, carbon-filtered, and ultraviolet irradiated. Laboratory water was mon-itored weekly for pH, alkalinity, conductivity, and hardness;and quarterly for selected inorganics, pesticides, and poly-chlorinated biphenyls. Typical water quality values were pHof 7.5, alkalinity of 43 mg/L, hardness of 70 mg/L (as CaCO3),and conductivity of 140 mhos/cm. Fish were killed by a blowto the head and placed immediately on ice before tissue prep-aration.
Preparation of microsomes
Rat (n 5 3; ;0.25 kg) and rainbow trout (n 5 4; ;2 kg)tissue microsomes (one animal per sample) were prepared ac-cording to the procedures of Guengerich [12]. Tissues wereimmediately excised, rinsed, and weighed in ice-cold Tris buff-er. Gills were trimmed from the branchial arches before weigh-ing. The Tris buffer consisted of 0.1 M Tris (pH 7.4), 0.1 mMKCl, 1 mM ethylenediaminetetraacetic acid, and 20 M of theantioxidant buterated hydroxy toluene. Blood was obtainedfrom the caudal vein of two rainbow trout, and serum wasmixed with buffer (1:5; v/v). Tissues were homogenized ice-cold in glass tubes with a motor-driven Teflont homogenizer;gills were first processed with a high-speed tissue grinder–sonicator. Tissue:buffer (w/v) dilutions were 1:5 for fish and1:10 for rats. Homogenates were filtered through cheesecloth,and the filtrate was centrifuged at 12,000 g for 60 min. Thesupernatant was removed and either assayed immediately forresidual CaE activity or centrifuged at 100,000 g for 60 minto produce a microsomal pellet [12]. Microsomes were resus-pended in buffer and stored at 2808C until assayed. Proteinwas assayed by the method of Lowry et al. [13].
Preparation of fish homogenates
Homogenates of juvenile fish (n 5 3 for each species),rainbow trout embryos (n 5 3; each sample consisted of 13–15 eggs, including the chorion), trout yolk-sac fry (n 5 3;each sample consisted of 18 or 19 fish), and adult rainbowtrout (n 5 3) were prepared using identical methods, with theexception that adult rainbow trout were initially ground tohomogeneity using a motorized tissue grinder and then sub-sampled for further analysis. Samples of all species and lifestages were rinsed and weighed in ice-cold Tris buffer, andthen homogenized on ice using a high-speed tissue grinder–sonicator. Tissue:buffer (w/v) dilutions were 1:5 for all ho-mogenates. Homogenates were filtered through cheesecloth,and then the filtrate was centrifuged at 1,000 g for 20 min.The supernatant was immediately assayed for CaE activity toprevent loss of activity with storage [14].
Assay of CaE activity
Enzyme activity was assayed by quantifying the formationof 4-nitrophenol from 4-nitrophenyl acetate using the spectro-metric method of Heymann and Mentlein [15]. Incubationswere carried out in a Tris(hydroxymethyl)aminomethane hy-drochloride (Tris-HCL) buffer (Sigma, St. Louis, MO, USA)
(0.1 mM, pH 8). Assays were run for 1.25 min and productformation was linear during all runs. The CaE activity of wholefish and embryo homogenates were assayed under saturatingconditions (440 mM 4-nitrophenyl acetate) at a constant tem-perature of 228C. The temperature optima of CaE in rat liverand trout gill and liver microsomes were determined undersaturating conditions (440 mM 4-nitrophenyl acetate) and vary-ing temperature (2–408C, depending on the tissue). The Vmax
and Km values were determined by varying the substrate con-centration (3.8–769 mM 4-nitrophenyl acetate; Heymann andMentlein [15]) at a constant temperature of 258C (trout tissues)or 378C (rat liver). The detection limit of the assay was ap-proximately 1 nmol/min/mg protein.
Determination of Km and Vmax and statistical analyses
The values of Km (slope) and Vmax (intercept) were deter-mined by linear regression analysis of an Eadie–Hofstee plotof velocity/substrate concentration (i.e., v/[S] versus velocity[16]). The value Vmax is the maximum reaction rate, and Km
(Michaelis constant) is the substrate concentration at one halfof Vmax. The values of Km and Vmax were also determined usingthe Michaelis–Menton equation using nonlinear modeling withsimple weighting and the Levenberg–Marquart algorithm. Sig-nificant differences in tissue levels of CaE activity were de-termined by a two-sided Student’s t test (alpha 5 0.02).
RESULTS
CaE kinetics and temperature optima
The CaE activity exhibited Michaelis–Menton kinetics asevidenced by a curvilinear approach to Vmax on Cartesian co-ordinates (data not shown). Eadie–Hofstee linear regressionplots of CaE activity (velocity [v]) and v/[S] were used toestimate Km (slope of regression) and Vmax (intercept) (Fig. 1).Gill and sera data were linear on Eadie–Hofstee coordinates,whereas both fish and rat liver exhibited a curvilinear response(Fig. 1). The CaE activity of trout liver (Vmax 5 672 nmol/min/mg microsomal protein) was about four times less thanin rat liver (Table 1). Apparent Km values ranged from 28 (troutliver) to 214 (trout sera) mM (Table 1). Values of Vmax (nmol/min/mg microsomal protein 6 SE) determined by nonlinearmodeling were nearly identical to the values derived from theEadie–Hofstee method (Table 1): rat liver (2,683 6 200); troutliver (678 6 66); gill (16.1 6 1.7); trout sera (1,737 6 35).Values of Km (mM, 6 SE) were also nearly identical to thevalues derived from the Eadie–Hofstee method (Table 1): ratliver (38.5 6 9.5); trout liver (29.0 6 12); gill (92 6 27); troutsera (243 6 12). The CaE activity of trout and rat tissues werealso assayed at various temperatures under saturating condi-tions of substrate (Fig. 2). Trout gill and liver microsomesexhibited substantial CaE activity and limited variation overthe range of 2 to 408C, with a temperature optimum of ap-proximately 228C (Fig. 2). Trout sera and rat liver microsomesexhibited a temperature optimum of 35 to 408C (Fig. 2).
The CaE activity in tissues (per gram or whole tissue) wascalculated from the CaE activity per milligram of protein and theprotein content of the tissue (Table 1), and tissue weight (Table2). Rainbow trout sera and rat liver exhibited the highest activity(sera 5 87 mmol/min/g; rat liver 5 76 mmol/min/g). The CaEin sera and liver were estimated to constitute 57% and 10%,respectively, of the CaE in the whole rainbow trout (Table 2).
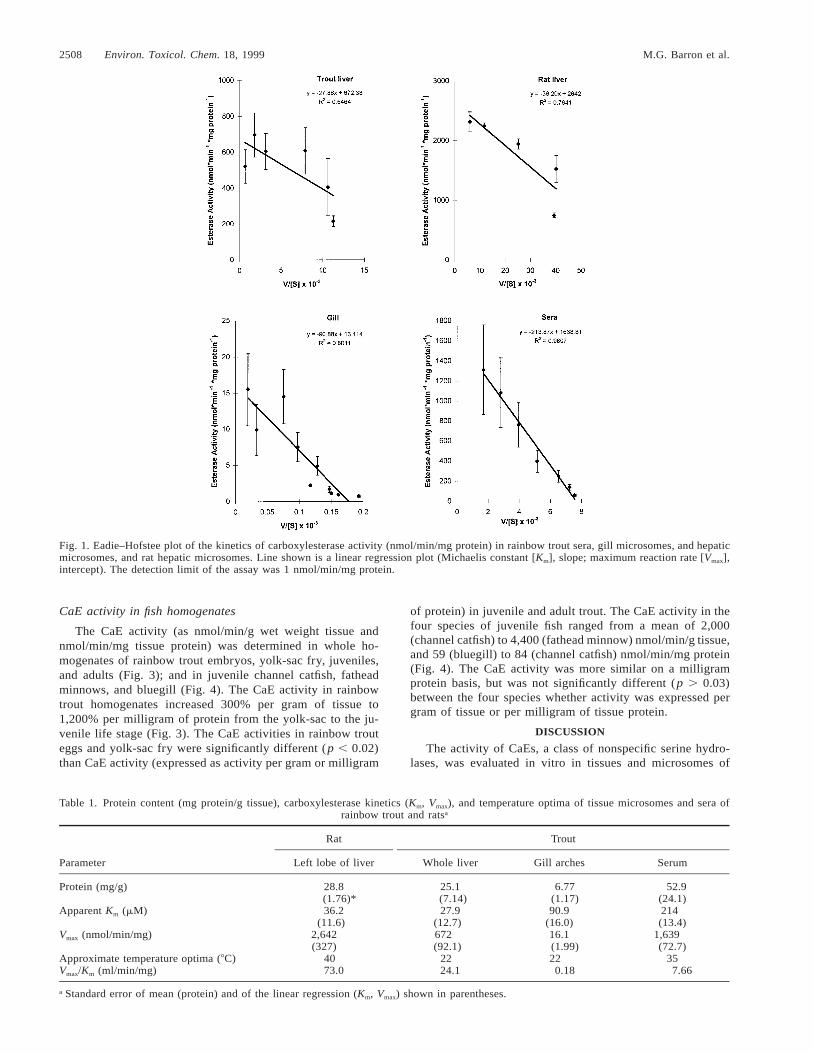
2508 Environ. Toxicol. Chem. 18, 1999 M.G. Barron et al.
Fig. 1. Eadie–Hofstee plot of the kinetics of carboxylesterase activity (nmol/min/mg protein) in rainbow trout sera, gill microsomes, and hepaticmicrosomes, and rat hepatic microsomes. Line shown is a linear regression plot (Michaelis constant [Km], slope; maximum reaction rate [Vmax],intercept). The detection limit of the assay was 1 nmol/min/mg protein.
Table 1. Protein content (mg protein/g tissue), carboxylesterase kinetics (Km, Vmax), and temperature optima of tissue microsomes and sera ofrainbow trout and ratsa
Parameter
Rat
Left lobe of liver
Trout
Whole liver Gill arches Serum
Protein (mg/g)
Apparent Km (mM)
Vmax (nmol/min/mg)
Approximate temperature optima (8C)Vmax/Km (ml/min/mg)
28.8(1.76)*36.2
(11.6)2,642(327)
4073.0
25.1(7.14)27.9
(12.7)672(92.1)
2224.1
6.77(1.17)90.9
(16.0)16.1(1.99)220.18
52.9(24.1)214(13.4)
1,639(72.7)
357.66
a Standard error of mean (protein) and of the linear regression (Km, Vmax) shown in parentheses.
CaE activity in fish homogenates
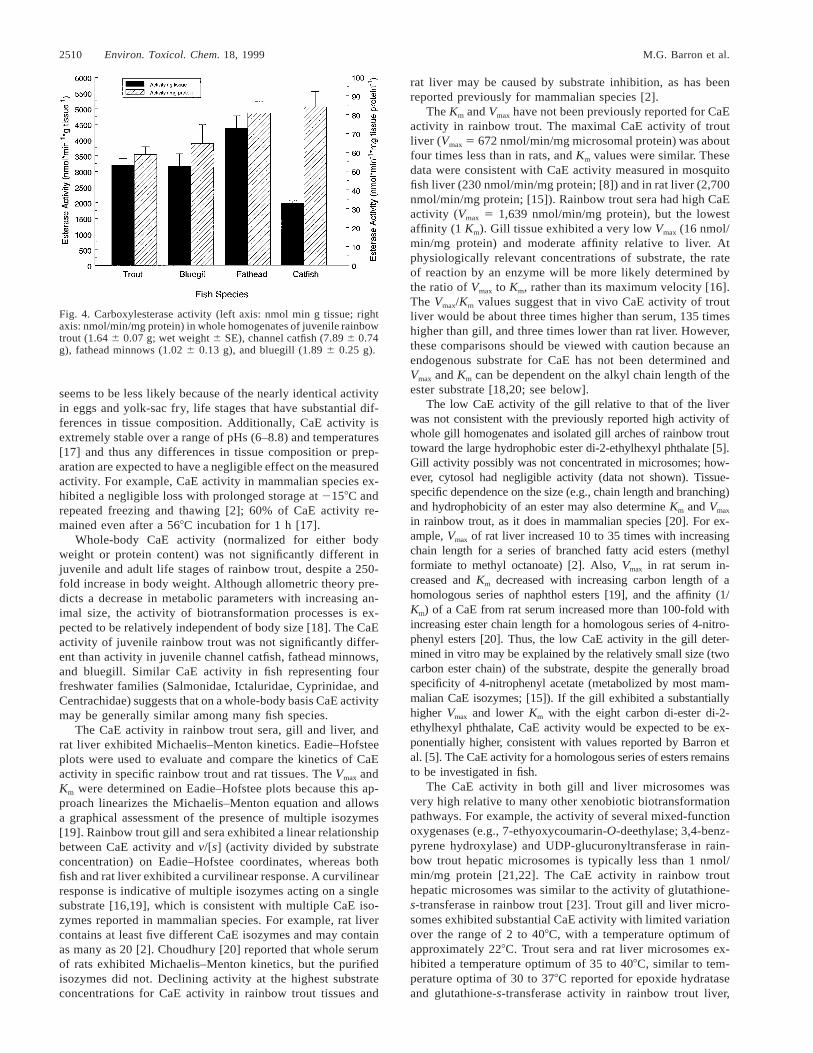
The CaE activity (as nmol/min/g wet weight tissue andnmol/min/mg tissue protein) was determined in whole ho-mogenates of rainbow trout embryos, yolk-sac fry, juveniles,and adults (Fig. 3); and in juvenile channel catfish, fatheadminnows, and bluegill (Fig. 4). The CaE activity in rainbowtrout homogenates increased 300% per gram of tissue to1,200% per milligram of protein from the yolk-sac to the ju-venile life stage (Fig. 3). The CaE activities in rainbow trouteggs and yolk-sac fry were significantly different (p , 0.02)than CaE activity (expressed as activity per gram or milligram
of protein) in juvenile and adult trout. The CaE activity in thefour species of juvenile fish ranged from a mean of 2,000(channel catfish) to 4,400 (fathead minnow) nmol/min/g tissue,and 59 (bluegill) to 84 (channel catfish) nmol/min/mg protein(Fig. 4). The CaE activity was more similar on a milligramprotein basis, but was not significantly different (p . 0.03)between the four species whether activity was expressed pergram of tissue or per milligram of tissue protein.
DISCUSSION
The activity of CaEs, a class of nonspecific serine hydro-lases, was evaluated in vitro in tissues and microsomes of

Carboxylesterase activity in trout Environ. Toxicol. Chem. 18, 1999 2509
Fig. 2. Temperature-dependence of carboxylesterase activity (nmol/min/mg protein) under saturating conditions in rainbow trout sera, gillmicrosomes, and hepatic microsomes, and rat hepatic microsomes. The detection limit of the assay was 1 nmol/min/mg protein.
Fig. 3. Carboxylesterase activity (left axis: nmol/min/g tissue; rightaxis: nmol/min/mg protein) in homogenates of four developmentalstages of rainbow trout: eggs (including chorion and eyed embryo;0.078 6 0.003 g; wet weight 6 SE), yolk-sac fry (0.055 6 0.002 g),juvenile (1.64 6 0.07 g), and adult (428 6 52 g).
Table 2. Calculated mean carboxylesterase (CaE) activity (95%confidence limit; n 5 2–4) in tissues of adult rainbow trout (1.25–2
kg) and juvenile rats (approximately 0.25 kg)
Tissue
Activity perg tissue
(mmol/min/g)
Tissueweight
(g)
Activity perwhole tissue(mmol/min)
Percent ofwhole body
activitya
Trout gill
Trout sera
Trout liver
Rat liver
0.13(0.05)86.6
(27.0)16.9(7.4)76.2
(21.7)
14.1(4.4)65.7b
(4.5)20.8(4.6)—c
1.88(1.00)
2,690(1,580)
378(73)—
,0.1
57.7(37.5)10.2(2.5)—
a Calculated from CaE activity in adult rainbow trout (Fig. 3).b Serum volume calculated as 3.3% of body weight [28].c Not calculable; only left liver lobe of rat assayed.
rainbow trout and compared to esterase activity measured inrats and other fish species. Rainbow trout had substantial CaEactivity at the embryo stage of development (718 nmol/min/g egg) and exhibited a 300% increase in CaE activity betweenthe yolk-sac and juvenile life stages. The increase in activitywas even more dramatic when expressed per milligram ofprotein (1,200%). The physiologic functions of CaEs are un-certain and may include aspects of the storage and mobilizationof specific lipids, assimilation of biological and xenobioticesters, and regulatory functions such as bioinactivation of spe-
cific hormones [1]. The increase in CaE activity between theyolk-sac and juvenile life stages may be related to the fish’stransition from feeding on an endogenous to an exogenousfood source, or to differences in the tissue composition ofdifferent life stages (e.g., effects of lipid content or compo-sition on in vitro activity). However, this latter possibility
2510 Environ. Toxicol. Chem. 18, 1999 M.G. Barron et al.
Fig. 4. Carboxylesterase activity (left axis: nmol min g tissue; rightaxis: nmol/min/mg protein) in whole homogenates of juvenile rainbowtrout (1.64 6 0.07 g; wet weight 6 SE), channel catfish (7.89 6 0.74g), fathead minnows (1.02 6 0.13 g), and bluegill (1.89 6 0.25 g).
seems to be less likely because of the nearly identical activityin eggs and yolk-sac fry, life stages that have substantial dif-ferences in tissue composition. Additionally, CaE activity isextremely stable over a range of pHs (6–8.8) and temperatures[17] and thus any differences in tissue composition or prep-aration are expected to have a negligible effect on the measuredactivity. For example, CaE activity in mammalian species ex-hibited a negligible loss with prolonged storage at 2158C andrepeated freezing and thawing [2]; 60% of CaE activity re-mained even after a 568C incubation for 1 h [17].
Whole-body CaE activity (normalized for either bodyweight or protein content) was not significantly different injuvenile and adult life stages of rainbow trout, despite a 250-fold increase in body weight. Although allometric theory pre-dicts a decrease in metabolic parameters with increasing an-imal size, the activity of biotransformation processes is ex-pected to be relatively independent of body size [18]. The CaEactivity of juvenile rainbow trout was not significantly differ-ent than activity in juvenile channel catfish, fathead minnows,and bluegill. Similar CaE activity in fish representing fourfreshwater families (Salmonidae, Ictaluridae, Cyprinidae, andCentrachidae) suggests that on a whole-body basis CaE activitymay be generally similar among many fish species.
The CaE activity in rainbow trout sera, gill and liver, andrat liver exhibited Michaelis–Menton kinetics. Eadie–Hofsteeplots were used to evaluate and compare the kinetics of CaEactivity in specific rainbow trout and rat tissues. The Vmax andKm were determined on Eadie–Hofstee plots because this ap-proach linearizes the Michaelis–Menton equation and allowsa graphical assessment of the presence of multiple isozymes[19]. Rainbow trout gill and sera exhibited a linear relationshipbetween CaE activity and v/[s] (activity divided by substrateconcentration) on Eadie–Hofstee coordinates, whereas bothfish and rat liver exhibited a curvilinear response. A curvilinearresponse is indicative of multiple isozymes acting on a singlesubstrate [16,19], which is consistent with multiple CaE iso-zymes reported in mammalian species. For example, rat livercontains at least five different CaE isozymes and may containas many as 20 [2]. Choudhury [20] reported that whole serumof rats exhibited Michaelis–Menton kinetics, but the purifiedisozymes did not. Declining activity at the highest substrateconcentrations for CaE activity in rainbow trout tissues and
rat liver may be caused by substrate inhibition, as has beenreported previously for mammalian species [2].
The Km and Vmax have not been previously reported for CaEactivity in rainbow trout. The maximal CaE activity of troutliver (Vmax 5 672 nmol/min/mg microsomal protein) was aboutfour times less than in rats, and Km values were similar. Thesedata were consistent with CaE activity measured in mosquitofish liver (230 nmol/min/mg protein; [8]) and in rat liver (2,700nmol/min/mg protein; [15]). Rainbow trout sera had high CaEactivity (Vmax 5 1,639 nmol/min/mg protein), but the lowestaffinity (1 Km). Gill tissue exhibited a very low Vmax (16 nmol/min/mg protein) and moderate affinity relative to liver. Atphysiologically relevant concentrations of substrate, the rateof reaction by an enzyme will be more likely determined bythe ratio of Vmax to Km, rather than its maximum velocity [16].The Vmax/Km values suggest that in vivo CaE activity of troutliver would be about three times higher than serum, 135 timeshigher than gill, and three times lower than rat liver. However,these comparisons should be viewed with caution because anendogenous substrate for CaE has not been determined andVmax and Km can be dependent on the alkyl chain length of theester substrate [18,20; see below].
The low CaE activity of the gill relative to that of the liverwas not consistent with the previously reported high activity ofwhole gill homogenates and isolated gill arches of rainbow trouttoward the large hydrophobic ester di-2-ethylhexyl phthalate [5].Gill activity possibly was not concentrated in microsomes; how-ever, cytosol had negligible activity (data not shown). Tissue-specific dependence on the size (e.g., chain length and branching)and hydrophobicity of an ester may also determine Km and Vmax
in rainbow trout, as it does in mammalian species [20]. For ex-ample, Vmax of rat liver increased 10 to 35 times with increasingchain length for a series of branched fatty acid esters (methylformiate to methyl octanoate) [2]. Also, Vmax in rat serum in-creased and Km decreased with increasing carbon length of ahomologous series of naphthol esters [19], and the affinity (1/Km) of a CaE from rat serum increased more than 100-fold withincreasing ester chain length for a homologous series of 4-nitro-phenyl esters [20]. Thus, the low CaE activity in the gill deter-mined in vitro may be explained by the relatively small size (twocarbon ester chain) of the substrate, despite the generally broadspecificity of 4-nitrophenyl acetate (metabolized by most mam-malian CaE isozymes; [15]). If the gill exhibited a substantiallyhigher Vmax and lower Km with the eight carbon di-ester di-2-ethylhexyl phthalate, CaE activity would be expected to be ex-ponentially higher, consistent with values reported by Barron etal. [5]. The CaE activity for a homologous series of esters remainsto be investigated in fish.
The CaE activity in both gill and liver microsomes wasvery high relative to many other xenobiotic biotransformationpathways. For example, the activity of several mixed-functionoxygenases (e.g., 7-ethyoxycoumarin-O-deethylase; 3,4-benz-pyrene hydroxylase) and UDP-glucuronyltransferase in rain-bow trout hepatic microsomes is typically less than 1 nmol/min/mg protein [21,22]. The CaE activity in rainbow trouthepatic microsomes was similar to the activity of glutathione-s-transferase in rainbow trout [23]. Trout gill and liver micro-somes exhibited substantial CaE activity with limited variationover the range of 2 to 408C, with a temperature optimum ofapproximately 228C. Trout sera and rat liver microsomes ex-hibited a temperature optimum of 35 to 408C, similar to tem-perature optima of 30 to 378C reported for epoxide hydrataseand glutathione-s-transferase activity in rainbow trout liver,

Carboxylesterase activity in trout Environ. Toxicol. Chem. 18, 1999 2511
kidney, and gill tissues [23]. Appreciable CaE activity over abroad temperature range may be a favorable adaptation for fishsuch as rainbow trout that experience ranges in environmentaltemperatures of 2 to 208C.
Rainbow trout sera and rat liver exhibited the highest ac-tivity (sera 5 86 mmol/min/g; rat liver 5 76.2 mmol/min/g)on a whole-tissue basis. The CaE in sera was estimated tocomprise 57% of the activity in the whole rainbow trout (Table2). The CaE activity in tissues (per gram or whole tissue) wereextrapolated from microsomal activity (gill and liver) or fromblood protein content, and should be viewed as approxima-tions. Definitive estimates of the relative contribution of in-dividual tissues to whole-body CaE activity would requirewhole-tissue preparations and larger sample sizes.
Rapid biotransformation of a variety of xenobiotic substratesis known to reduce the bioconcentration of environmental con-taminants [24,25]. Previous work on a limited number of estersubstrates indicated that fish have high CaE activity in both seraand liver [26], and ester hydrolysis significantly reduced ac-cumulation of several hydrophobic esters [4–6]. Inhibition ofCaE activity with the selective active-site inhibitor bis-nitro-phenyl phosphate caused an estimated sevenfold increase in thebioconcentration of triclopyr butoxyethyl ester and an eightfoldincrease in di-2-ethylhexyl phthalate bioconcentration [5,27].Presystemic metabolism in the gill has also been shown to lowerthe amount of unhydrolyzed di-2-ethylhexyl phthalate enteringrainbow trout [5]. This may have favorable toxicologic con-sequences because the hydrolysis product should be less per-sistent in the fish. The CaE activity is also protective againstOP toxicity in both fish and mammals [9,11]. Atterberry et al.[11] concluded that age-related changes in CaE seemed to con-tribute to the age-dependent toxicity of the acetylcholinesterase-inhibiting insecticide chlorpyrifos in rats. Abas and Hayton [9],using computer simulations, concluded that CaE was an im-portant detoxication pathway for the acetylcholinesterase inhib-itor paroxon in rainbow trout. Carboxylesterases detoxify OPsby stoichiometrically reducing the amount of OP compoundavailable to inhibit acetylcholinesterase [10].
The results of this study demonstrated that rainbow trout hadhigh esterase activity over a broad range of temperatures, thatCaE activity significantly increased between the yolk-sac andjuvenile life stages, and that variation between the CaE activityin trout and three other families of freshwater fish was limited.The CaE activity of fish is expected to substantially influence theaccumulation and toxicity of pesticides and other esters enteringthe aquatic environment. The tissue-specific biotransformationresults reported here may be useful in understanding both thetoxicity and bioaccumulation of ester contaminants. Our resultsshould have general applicability to a variety of xenobiotic estersand may be useful in developing physiologically based phar-macokinetic models and assessing risks of ester contaminants.However, the results should be applied with the recognition thatCaE activity may be dependent on the chain length and hydro-phobicity of the ester. Additional research with fish is needed toelucidate substrate-specific biotransformation for a homologousseries of environmental esters.
Acknowledgement—We thank Elizabeth Fergeson for Michaelis–Mentonanalyses; T. Ball, P. Wilga, and S. Gorzinski for technical assistance; andM.A. Mayes, P.J. Gering, and P.G. Watanabe for advice and review.
REFERENCES1. Leinweber FJ. 1987. Possible physiological roles of carboxylic
ester hydrolases. Drug Metab Rev 18:379–439.
2. Heymann E. 1980. Carboxylesterases and amidases. In JakobyWB, ed, Enzymatic Basis of Detoxication, Vol 2. Academic, NewYork, NY, USA, pp 291–323.
3. Walker CH, Mackness ML. 1983. Esterases: Problems of iden-tification and classification. Biochem Pharmacol 32:3265–3269.
4. Rodgers CA, Stalling DL. 1972. Dynamics of an ester of 2,4-Din organs of three fish species. Weed Sci 20:101–105.
5. Barron MG, Schulz IR, Hayton WL. 1989. Presystemic branchialmetabolism limits di-2-ethylhexyl phthalate accumulation in fish.Toxicol Appl Pharmacol 98:49–57.
6. Barron MG, Mayes MA, Murphy PG, Nolan RJ. 1990. Phar-macokinetics and metabolism of triclopyr butoxyethyl ester incoho salmon. Aquat Toxicol 16:19–32.
7. Barron MG, Albro PW, Hayton WL. 1995. Biotransformation ofdi-2-ethylhexyl phthalate by rainbow trout. Environ ToxicolChem 14:873–876.
8. Boone JS, Chambers JE. 1996. Time course of inhibition of cholin-esterase and aliesterase activities, and nonprotein sulfhydryl levelsfollowing exposure to organophosphorous insecticides in mosquitofish (Gambusia affinis). Fundam Appl Toxicol 29:202–207.
9. Abas R, Hayton WL. 1997. A physiologically based pharmaco-kinetic and pharmacodynamic model for paraoxon in rainbowtrout. Toxicol Appl Pharmacol 145:192–201.
10. Maxwell DM. 1992. The specificity of carboxylesterase protec-tion against the toxicity of organophosphorus compounds. ToxicolAppl Pharmacol 114:306–312.
11. Atterberry TT, Burnett WT, Chambers JE. 1997. Age-related dif-ferences in parathion and chlorpyrifos toxicity in male rats: targetand nontarget esterase sensitivity and cytochrome P450-mediatedmetabolism. Toxicol Appl Pharmacol 147:411–418.
12. Guengerich FP. 1982. Microsomal enzymes involved in toxicology—Analysis and separation. In Hayes AW, ed, Principles and Methodsof Toxicology. Raven, New York, NY, USA, pp 609–634.
13. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951. Proteinmeasurement with the Folin phenol reagent. J Biol Chem 193:265–275.
14. Lindstrom-Seppa P, Hanninen O. 1988. Sampling and storageconditions of rainbow trout liver affects monoxygenase and con-jugation enzymes. Comp Biochem Physiol C 89:221–224.
15. Heymann E, Mentlein R. 1981. Carboxylesterases—Amidases. InJakoby WB, ed, Methods in Enzymology, Vol 77, Detoxicationand Drug Metabolism: Conjugation and Related Systems. Aca-demic, New York, NY, USA, pp 333–341.
16. Hofstee BH. 1952. On the evaluation of the constants Vm and Km
in enzymatic reactions. Science 116:329–331.17. Hashinotsume M, Higashino K, Hada T, Yamamura Y. 1978. Pu-
rification and enzymatic properties of rat serum carboxylesterase.J Biochem 84:1325–1333.
18. Weis M, Sziegoleit W, Forster W. 1977. Dependence of phar-macokinetic parameters on the body weight. Int J Clin PharmacolRes 15:572–575.
19. Lenk W. 1976. Application and interpretation of kinetic analysesfrom the microsomal drug metabolizing oxygenases. BiochemPharmacol 25:997–1005.
20. Choudhury SR. 1974. Studies of substrate hydrolysis by an es-terase isozyme of rat serum. Biochim Biophys Acta 350:484–490.
21. Lindstrom-Seppa P, Koivusaari U, Hanninen O. 1981. Extrahe-patic xenobiotic metabolism in north-European freshwater fish.Comp Biochem Physiol C 69:259–263.
22. Forlin L, Andersson T. 1985. Storage conditions of rainbow troutliver cytochrome P-450 and conjugating enzymes. Comp BiochemPhysiol B 80:569–572.
23. Lauren DJ, Halarnkar PP, Hammock BD, Hinton DE. 1989. Mi-crosomal and cytosolic epoxide hydrolase and glutathione s-trans-ferase activities in the gill, liver, and kidney of the rainbow trout,Salmo gardneri. Baseline levels and optimization of assay con-ditions. Biochem Pharmacol 38:881–887.
24. Lech JJ, Bend JR. 1980. Relationship between biotransformationand the toxicity and fate of xenobiotic chemicals in fish. EnvironHealth Perspect 34:115–131.
25. Barron MG. 1990. Bioconcentration. Environ Sci Techol 24:1612–1618.
26. Lech J, Melancon M. 1980. Uptake, metabolism and dispositionof xenobiotic chemicals in fish. EPA-600 3-80-082. U.S. Envi-ronmental Protection Agency, Duluth, MN.
27. Karara AH, Hayton WL. 1988. Metabolic inhibition and di-2-ethylhexyl phthalate pharmacokinetics in fish. Drug Metabol Dis-pos 16:146–150.
28. Barron MG, Tarr BD, Hayton WL. 1987. Temperature dependenceof cardiac output and regional blood flow in rainbow trout (Salmogairdneri Richardson). J Fish Biol 31:735–744.