thermosensitivity of red blood cells from down's syndrome individuals
TRANSCRIPT

Ž .Bioelectrochemistry 52 2000 239–249www.elsevier.comrlocaterbioelechem
Thermosensitivity of red blood cells from Down’s syndrome individuals
Maria Przybylska a,), Maria Bryszewska b, Jozef Kedziora c´a Department of Thermobiology, Institute of Biophysics, UniÕersity of Lodz, 12r16 Banacha St., 90-237 Lodz, Poland
b Department of General Biophysics, Institute of Biophysics, UniÕersity of Lodz, 12r16 Banacha St., 90-237 Lodz, Polandc Department of Physiology, Institute of Fundamental Sciences, Military Medical Academy of Lodz, 1 Haller Sq., 90-647 Lodz, Poland
Received 4 January 2000; received in revised form 15 March 2000; accepted 19 March 2000
Abstract
Ž .Biochemical disturbances of the reactive oxygen species metabolism revealed in subjects with Down’s syndrome DS , and thefindings indicating that heat-induced cell alterations have been, at least, partly mediated by reactive oxygen species, made the elucidationof the response of trisomic cells to elevated temperatures of special interest. Kinetic analysis of cell-survival curves, accompanied by the
Ž .flow cytometry and the scanning electron microscopy SEM examinations, and their relationship with the cell membrane fluidity, wereŽ .undertaken. At each temperature 48–548C , D parameters, representing the ability to accumulate sublethal damages, were similar forqŽ .both cell groups. D parameters inverse leakage rates; D s1rk were greater for DS cells at each temperature below 548C. The0 0
Ž .haemolysis sensitivity ratio HSR showed that DS erythrocytes were, in average, 1.60 times more resistant to heat injury than those fromnormal subjects. Activation energies of haemolysis, calculated according to the Arrhenius equation, were similar both for normalŽ w x. Ž w x.290.8"6.5 kJrmol and DS erythrocytes 288.0"5.5 kJrmol . Flow cytometry studies showed that the scattering properties of
Ž .intact DS erythrocytes reflecting size, volume, shape and cell membrane surface morphology were different than those of normal cells.Ž .Scanning electron micrographs and scattering diagrams obtained for cells submitted to heat stress 518C confirmed that DS erythrocytes
were more resistant, to a certain extent, to heat-induced disruption than normal cells. The steady-state fluorescence anisotropy ofŽ Ž . .TMA-DPH 1- 4-trimethyl-ammoniumphenyl -6-phenyl-1,3,5-hexatriene showed that untreated DS erythrocytes had substantially lower
Ž . Ž .fluidity rs0.356"0.008 of the outer monolayer of cell membranes as compared to normal cells rs0.324"0.011 . The increase ofthe cell membrane fluidity during exposure to heat was observed. The greatest elevation of cell membrane fluidity occurred during thepreleakage period, immediately upon the heat treatment and was considered as a rate-limiting step of heat-induced haemolysis. q 2000Elsevier Science S.A. All rights reserved.
Keywords: Down’s syndrome; Erythrocyte; Flow cytometry; Scanning electron microscopy; TMA-DPH; Cell membrane fluidity
1. Introduction
Ž .Down’s syndrome DS is one of the best-known exam-ples of genetic disorders caused by the trisomy of the 21st
w xchromosome 1 . The prevalence of this aneuploidy variesbetween 1:500 and 1:1000 live birth and the social trendsprogressively tend to further increase the frequency of thisaneuploidy, due to its strong association with the advanced
w xmaternal age 1–3 . In addition to the widely reportedmental deficiency and other serious pathological conse-quences, individuals suffering from DS have pronounced
w xrisk of development of the different kinds of cancers 4–6 .
) Corresponding author. Fax: q48-42-35-44-73.Ž .E-mail address: [email protected] M. Przybylska .
DS is especially frequently associated with the childhoodw xhaematological malignancies 7 . In affected children, the
risk of leukaemia is increased about 10- to 20-fold of thatw xobserved in the nontrisomic population 8 .
The surplus of genetic information in subjects affectedby DS, causing an augmentation of the catalytic activity of
Ž .Cu–Zn superoxide dismutase — SOD-1 EC: 1.15.1.1. ,an antioxidant enzyme, coded on the distal segment of the
w x21 chromosomes 9–12 associated with an adaptive rise inthe activity of the H O yscavenging enzymes — glu-2 2
Ž . Ž .tathione peroxidase EC: 1.11.1.9. , catalase EC: 1.11.1.6.Ž . w xand glutathione reductase EC: 1.6.4.2. 12 , makes DS
cells a convenient model system with the potentially al-tered resistance to the reactive oxygen species.
The widespread use of elevated temperatures, in combi-nation with chemotherapy or radiotherapy, in clinical treat-
0302-4598r00r$ - see front matter q 2000 Elsevier Science S.A. All rights reserved.Ž .PII: S0302-4598 00 00106-9

( )M. Przybylska et al.rBioelectrochemistry 52 2000 239–249240
ment of broad spectrum of cancers, has increased attentionfor the molecular mechanisms of the heat-induced distur-
w xbance of normal as well as tumour tissues 13–20 . Takinginto account the fact that the heat-induced cell injury is, atleast, partly mediated by the reactive oxygen speciesw x21,22 , it may be expected that thermosensitivity of cellsderived from individuals suffering from DS will be altered,as compared to normal cells.
Despite their particular structural and biochemical prop-erties and the high thermal stability in comparison withother mammalian cells, erythrocytes have very often beenused to study the mechanism of cell sensitivity to heatw x w x23–28 . According to Lepock et al. 26 , membrane pro-teins were involved in the heat-induced red blood cellsdamage due to the relatively high value of activation
Ž .energy of this process 300 kJrmol . Contrary to this,w xresults obtained by Ivanov 29,30 revealed the mechanism
of haemolysis of red blood cells submitted to heat stress asbeing of the colloid-osmotic type. He assumed that thethermally induced haemolysis was depended on the mem-brane sphingomyelinrphosphatidylocholine ratio and the
w xfatty acids acyl-chain order 31 .The fundamental alterations occurring in red blood cells
from healthy subjects submitted to the elevated tempera-w xture have previously been described 23–26 . Studies of
thermosensitivity of red blood cells from individuals suf-fering from DS have not yet been published; thus, the
Ž .elucidation of the specific response of trisomic q21 cellsto heat is of special interest.
The aim of this study was to determine DS red bloodcell thermosensitivity and its relationship with the cellmembrane fluidity. For this purpose, kinetic analysis ofthermally induced disturbances of normal and DS erythro-cytes, accompanied by the application of flow cytometry
Ž .and scanning electron microscopy SEM examinations,were carried out. Additionally, the steady-state fluores-cence anisotropy measurements of the fluorescent probe
Ž Ž .TMA-DPH 1- 4-trimethyl-ammoniumphenyl -6-phenyl-.1,3,5-hexatriene as an assay of plasma membrane fluidity
were undertaken.
2. Materials and methods
2.1. Materials
Fluorescent probe TMA-DPH was purchased fromŽ .Molecular Probes Eugene, OR, USA . All other chemicals
were of the best quality commercially available.
2.2. Subject
ŽHuman peripheral blood from adult healthy donors 11.males and nine females aged in average 32.4"5.4 were
supplied by the Blood Bank of Lodz and blood samplesŽderived from DS patients six males and seven females
.aged in average 31.3"4.7 were obtained from the Mili-tary Medical Academy of Lodz.
2.3. Preparation of erythrocyte suspension
Blood samples, anticoagulated with 3% water solutionof trisodium citrate, were centrifuged at 600=g for 10min to remove plasma and the white buffy coat. The redblood cell pellet was then washed three times with anequal volume of an isotonic solution of sodium phos-
Ž y3phate-buffered saline PBS: 0.15 M NaCl; 1.9=10 M,y3 .NaH PO ; 8.1=10 M Na HPO ; pH 7.4 . After wash-2 4 2 4
ing, the packed erythrocytes were resuspended in PBS at ahaematocrit of 50%. All experiments were performed on
Žfreshly drawn blood because the red blood cells storage at.48C or at room temperature increased the rate of haemoly-
sis.
2.4. SEM
To carry out the SEM examinations, the erythrocytesuspensions, treated as described above, were heated in awater bath at 518C for 15 min or 1 h. Additionally, controlsamples without heating were prepared. After desired timetubes were cooled down under the tap water for 5 min tostop further heat damage to the cells and were centrifugedat 600=g for 10 min. The supernatants were discardedand erythrocyte pellets were washed twice with an equal
Žvolume of the sodium–potassium phosphate buffer 0.164.M Na HPO , 0.036 M KH PO ; pH 7.4 and finally2 4 2 4
resuspended in 1% glutaraldehyde in the same buffer at ahaematocrit of 50%. Fixed cells were allowed to settle onstandard microscopic cover glasses. After 1 h, the coverglasses were washed twice with the phosphate buffer.Subsequently samples were dehydrated with successive
Žwashes in an ascending ethanol series from 30% to 95%;.vrv , in 100% acetone and finally dried with CO at the2
triple point. Next, cells were coated with gold–palladiumand examined in a Stereoscan 600 scanning electron mi-
Ž .croscope Cambridge Instruments, Great Britain at theElectron Microscopy Laboratory of the Medical Universityin Lodz.
2.5. Flow cytometry
For quantitative evaluation of the heat-induced distur-Ž .bance of red blood cells, a Scatron Argus Norway flow
cytometer supplied with a 100 W HBO mercury arc lamp,Ž .with simultaneous separate detection at a low angle LS1
Ž .and a right angle LS2 was used. Scattering diagrams ofŽ 3.erythrocyte suspensions 0.5 cm , containing approxi-
mately 106 cellsrcm3, subjected to heating at 518C from15 min to 1 h, as described above, were registered. The
Ž .light scattered near the forward direction low angle isŽ .expected to be proportional to the size volume of the
particle and is independent of the cell refractive index and
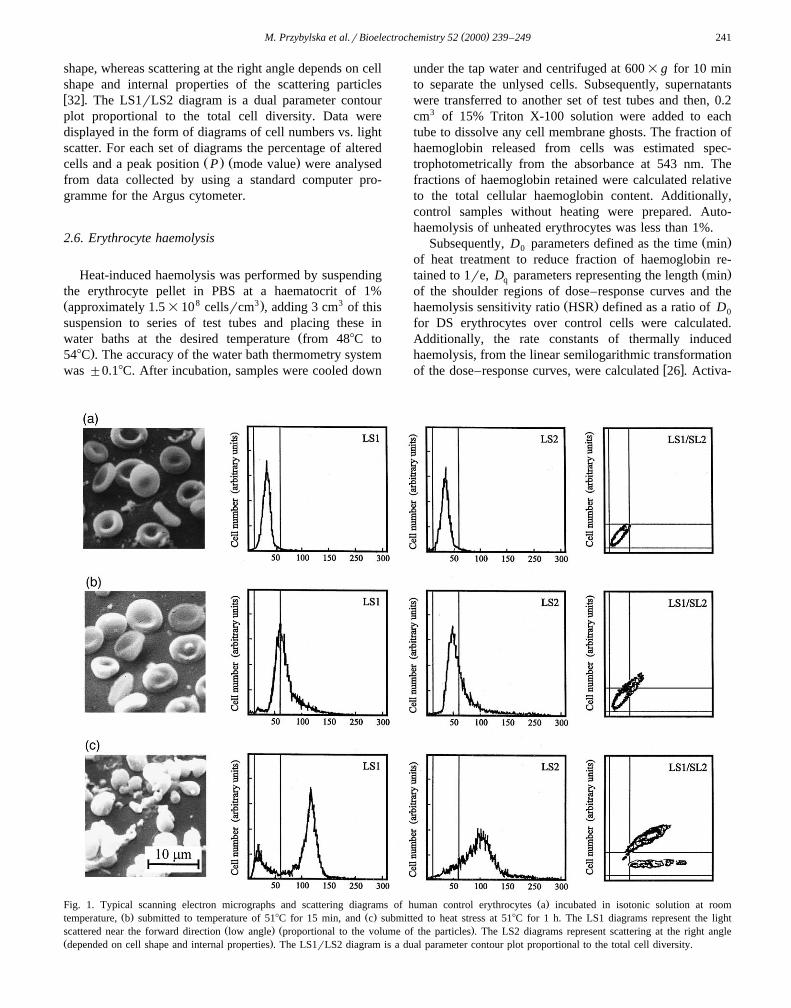
( )M. Przybylska et al.rBioelectrochemistry 52 2000 239–249 241
shape, whereas scattering at the right angle depends on cellshape and internal properties of the scattering particlesw x32 . The LS1rLS2 diagram is a dual parameter contourplot proportional to the total cell diversity. Data weredisplayed in the form of diagrams of cell numbers vs. lightscatter. For each set of diagrams the percentage of altered
Ž . Ž .cells and a peak position P mode value were analysedfrom data collected by using a standard computer pro-gramme for the Argus cytometer.
2.6. Erythrocyte haemolysis
Heat-induced haemolysis was performed by suspendingthe erythrocyte pellet in PBS at a haematocrit of 1%Ž 8 3. 3approximately 1.5=10 cellsrcm , adding 3 cm of thissuspension to series of test tubes and placing these in
Žwater baths at the desired temperature from 488C to.548C . The accuracy of the water bath thermometry system
was "0.18C. After incubation, samples were cooled down
under the tap water and centrifuged at 600=g for 10 minto separate the unlysed cells. Subsequently, supernatantswere transferred to another set of test tubes and then, 0.2cm3 of 15% Triton X-100 solution were added to eachtube to dissolve any cell membrane ghosts. The fraction ofhaemoglobin released from cells was estimated spec-trophotometrically from the absorbance at 543 nm. Thefractions of haemoglobin retained were calculated relativeto the total cellular haemoglobin content. Additionally,control samples without heating were prepared. Auto-haemolysis of unheated erythrocytes was less than 1%.
Ž .Subsequently, D parameters defined as the time min0
of heat treatment to reduce fraction of haemoglobin re-Ž .tained to 1re, D parameters representing the length minq
of the shoulder regions of dose–response curves and theŽ .haemolysis sensitivity ratio HSR defined as a ratio of D0
for DS erythrocytes over control cells were calculated.Additionally, the rate constants of thermally inducedhaemolysis, from the linear semilogarithmic transformation
w xof the dose–response curves, were calculated 26 . Activa-
Ž .Fig. 1. Typical scanning electron micrographs and scattering diagrams of human control erythrocytes a incubated in isotonic solution at roomŽ . Ž .temperature, b submitted to temperature of 518C for 15 min, and c submitted to heat stress at 518C for 1 h. The LS1 diagrams represent the light
Ž . Ž .scattered near the forward direction low angle proportional to the volume of the particles . The LS2 diagrams represent scattering at the right angleŽ .depended on cell shape and internal properties . The LS1rLS2 diagram is a dual parameter contour plot proportional to the total cell diversity.
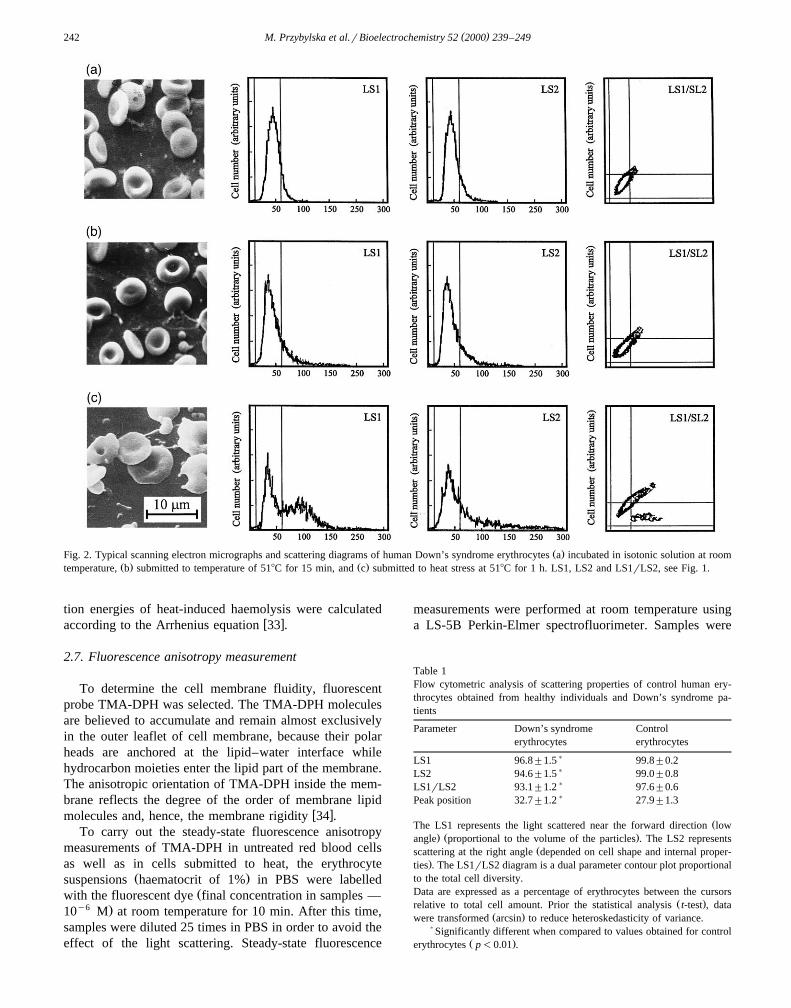
( )M. Przybylska et al.rBioelectrochemistry 52 2000 239–249242
Ž .Fig. 2. Typical scanning electron micrographs and scattering diagrams of human Down’s syndrome erythrocytes a incubated in isotonic solution at roomŽ . Ž .temperature, b submitted to temperature of 518C for 15 min, and c submitted to heat stress at 518C for 1 h. LS1, LS2 and LS1rLS2, see Fig. 1.
tion energies of heat-induced haemolysis were calculatedw xaccording to the Arrhenius equation 33 .
2.7. Fluorescence anisotropy measurement
To determine the cell membrane fluidity, fluorescentprobe TMA-DPH was selected. The TMA-DPH moleculesare believed to accumulate and remain almost exclusivelyin the outer leaflet of cell membrane, because their polarheads are anchored at the lipid–water interface whilehydrocarbon moieties enter the lipid part of the membrane.The anisotropic orientation of TMA-DPH inside the mem-brane reflects the degree of the order of membrane lipid
w xmolecules and, hence, the membrane rigidity 34 .To carry out the steady-state fluorescence anisotropy
measurements of TMA-DPH in untreated red blood cellsas well as in cells submitted to heat, the erythrocyte
Ž .suspensions haematocrit of 1% in PBS were labelledŽwith the fluorescent dye final concentration in samples —
y6 .10 M at room temperature for 10 min. After this time,samples were diluted 25 times in PBS in order to avoid theeffect of the light scattering. Steady-state fluorescence
measurements were performed at room temperature usinga LS-5B Perkin-Elmer spectrofluorimeter. Samples were
Table 1Flow cytometric analysis of scattering properties of control human ery-throcytes obtained from healthy individuals and Down’s syndrome pa-tients
Parameter Down’s syndrome Controlerythrocytes erythrocytes
)LS1 96.8"1.5 99.8"0.2)LS2 94.6"1.5 99.0"0.8)LS1rLS2 93.1"1.2 97.6"0.6)Peak position 32.7"1.2 27.9"1.3
ŽThe LS1 represents the light scattered near the forward direction low. Ž .angle proportional to the volume of the particles . The LS2 represents
Žscattering at the right angle depended on cell shape and internal proper-.ties . The LS1rLS2 diagram is a dual parameter contour plot proportional
to the total cell diversity.Data are expressed as a percentage of erythrocytes between the cursors
Ž .relative to total cell amount. Prior the statistical analysis t-test , dataŽ .were transformed arcsin to reduce heteroskedasticity of variance.
)Significantly different when compared to values obtained for controlŽ .erythrocytes p-0.01 .
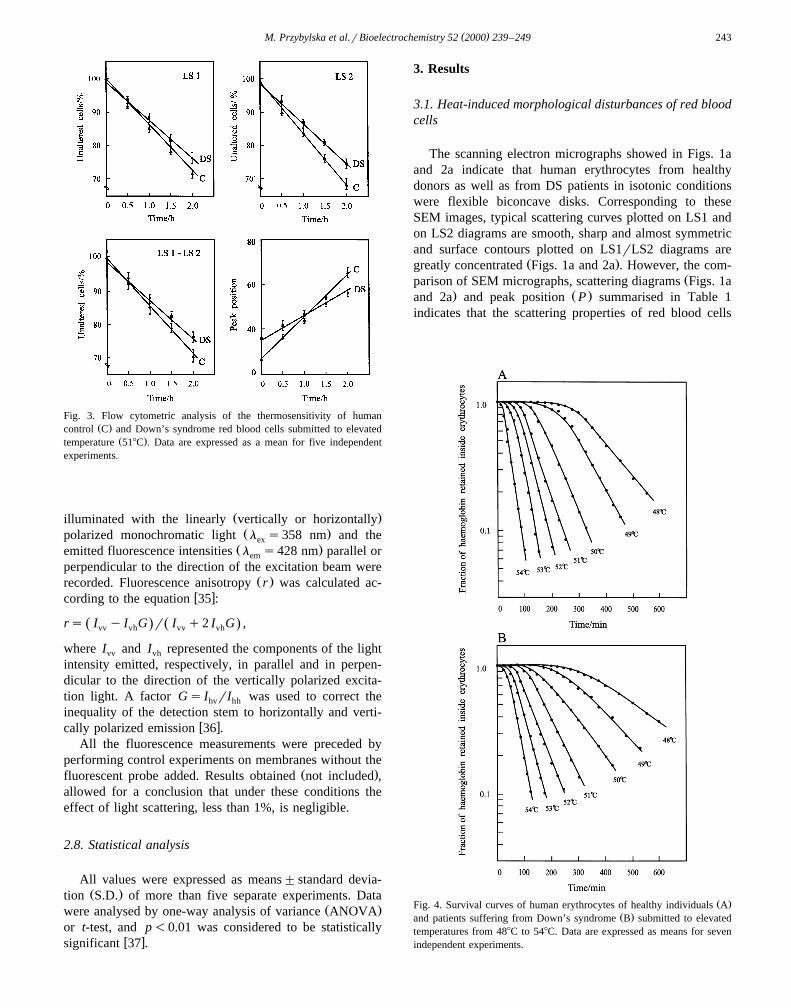
( )M. Przybylska et al.rBioelectrochemistry 52 2000 239–249 243
Fig. 3. Flow cytometric analysis of the thermosensitivity of humanŽ .control C and Down’s syndrome red blood cells submitted to elevated
Ž .temperature 518C . Data are expressed as a mean for five independentexperiments.
Ž .illuminated with the linearly vertically or horizontallyŽ .polarized monochromatic light l s358 nm and theex
Ž .emitted fluorescence intensities l s428 nm parallel orem
perpendicular to the direction of the excitation beam wereŽ .recorded. Fluorescence anisotropy r was calculated ac-
w xcording to the equation 35 :
rs I y I G r I q2 I G ,Ž . Ž .vv vh vv vh
where I and I represented the components of the lightvv vh
intensity emitted, respectively, in parallel and in perpen-dicular to the direction of the vertically polarized excita-tion light. A factor Gs I rI was used to correct thehv hh
inequality of the detection stem to horizontally and verti-w xcally polarized emission 36 .
All the fluorescence measurements were preceded byperforming control experiments on membranes without the
Ž .fluorescent probe added. Results obtained not included ,allowed for a conclusion that under these conditions theeffect of light scattering, less than 1%, is negligible.
2.8. Statistical analysis
All values were expressed as means"standard devia-Ž .tion S.D. of more than five separate experiments. Data
Ž .were analysed by one-way analysis of variance ANOVAor t-test, and p-0.01 was considered to be statistically
w xsignificant 37 .
3. Results
3.1. Heat-induced morphological disturbances of red bloodcells
The scanning electron micrographs showed in Figs. 1aand 2a indicate that human erythrocytes from healthydonors as well as from DS patients in isotonic conditionswere flexible biconcave disks. Corresponding to theseSEM images, typical scattering curves plotted on LS1 andon LS2 diagrams are smooth, sharp and almost symmetricand surface contours plotted on LS1rLS2 diagrams are
Ž .greatly concentrated Figs. 1a and 2a . However, the com-Žparison of SEM micrographs, scattering diagrams Figs. 1a
. Ž .and 2a and peak position P summarised in Table 1indicates that the scattering properties of red blood cells
Ž .Fig. 4. Survival curves of human erythrocytes of healthy individuals AŽ .and patients suffering from Down’s syndrome B submitted to elevated
temperatures from 488C to 548C. Data are expressed as means for sevenindependent experiments.

( )M. Przybylska et al.rBioelectrochemistry 52 2000 239–249244
Table 2Kinetic parameters of the heat-induced haemolysis of human red blood cells
w xt 8C Down’s syndrome erythrocytes Control erythrocytes HSR4 y1 4 y1w x w x w x w x w x w xD min D min k=10 s D min D min k=10 sq 0 q 0
) )48 301.2"17.3 292.4"15.4 0.57"0.03 284.3"19.2 189.2"10.8 0.88"0.05 1.56"0.02) )49 210.8"13.1 208.3"10.4 0.80"0.04 215.0"16.4 134.3"8.7 1.24"0.08 1.55"0.04
) )50 140.2"9.8 157.2"8.9 1.06"0.06 130.0"10.2 84.9"5.2 1.96"0.12 1.85"0.03) )51 103.1"7.2 98.0"6.2 1.70"0.11 105.0"8.2 65.9"3.2 2.54"0.12 1.49"0.04) )52 63.2"3.9 78.6"4.4 2.12"0.12 62.0"4.2 54.2"2.5 3.04"0.16 1.45"0.03) )53 50.0"2.8 55.9"3.4 2.98"0.18 51.4"3.6 34.0"2.0 4.84"0.29 1.64"0.02) )54 38.3"2.5 40.7"2.1 4.10"0.21 35.2"2.0 24.4"1.9 6.84"0.33 1.67"0.02
D — the length of the shoulder regions of dose–response curves.q
D — the time of heat treatment reducing fraction of haemoglobin retained to 1re on the linear region of dose–response curves.0
k — rate constant of heat-induced haemolysis.HSR — the haemolysis sensitivity ratio, defined as a ratio of D over control cells for DS.0
Data are expressed as a mean"S.D. for seven independent experiments.) Ž .Significantly different when compared to values obtained for control erythrocytes p-0.01 .
Žreflecting erythrocyte size, shape and cell membrane sur-.face morphology derived from DS patients are different
Ž .p-0.001 than those of erythrocytes obtained fromhealthy individuals.
ŽA typical experiment conducted with heating 15 min or.1 h, 518C of red blood cells suspended in isotonic solution
is illustrated in Figs. 1b–c and 2b–c. Scanning electronimages and scattering diagrams show that the elevatedtemperatures transform red blood cells gradually fromdiscocytes into more spherical shapes, with loss of theirconcavity and progressively lead to their disruption, espe-cially for longer incubation time. The curves plotted onLS1 and on LS2 diagrams become more irregular, asym-metric, flattened and eventually double-peaked as com-pared to those for unheated cells. Also the peak positionŽ .P is significantly shifted. The distinctly split surfacecontour plotted on LS1rLS2 diagram suggests the exis-tence of two subpopulations of particles of altered sizewhich corroborates the results obtained by SEM showingthe formation of numerous vesicles and intercellular links.The comparison of scanning electron micrographs, both
Žscattering diagrams, the dual parameter contour plot Figs.. Ž .1b–c and 2b–c , peak position P and curves represent-
Ž .ing the percentage of unaltered cells Fig. 3 indicated thatthermosensitivity of DS red blood cells is substantiallyreduced compared to normal cells. The greatest variety oferythrocyte shapes and sizes was especially visible for the
Ž .longer incubation time )1 h .
3.2. Kinetic analysis of cell-surÕiÕal curÕes
The effect of elevated temperatures on mammalian cellsassayed in terms of cell damage leads to survival curves,similar to those described for ionising radiation treatment.The fractions of haemoglobin retained by human erythro-cytes obtained from DS patients and healthy individualsplotted vs. time of exposure to temperatures from 488C to
548C are presented in Fig. 4. For each temperature, Dq
parameters are inversely proportional to temperature andŽ .almost the same p)0.1 for both groups of cells. A
comparison of temperature dependence of D parameters0Ž .Table 2, Fig. 4 shows that D parameters, for the0
temperature range from 488C to 548C, are greater for DScells, as compared to normal ones and the differences areinversely proportional to the temperature. At the tempera-ture above 54.58C, similar values of D parameters were0
Ž .obtained for both groups of cells Fig. 5 . The HSR showsthat DS red blood cells are in average 1.60 times moreresistant to heat-induced haemolysis than those from nor-
Ž .mal subjects Table 2 .Arrhenius plots of rate of haemoglobin leakage both
from control as well as DS erythrocytes are linear over theŽ .temperature range from 488C to 548C Fig. 6 . Activation
energies of haemolysis calculated according to the Arrhe-nius equation are similar both for normal and DS erythro-
Fig. 5. Temperature dependence of D parameters derived from survival0
curves of human erythrocytes of healthy individuals and Down’s syn-drome patients submitted to temperatures from 488C to 548C. Data areexpressed as means for seven independent experiments.
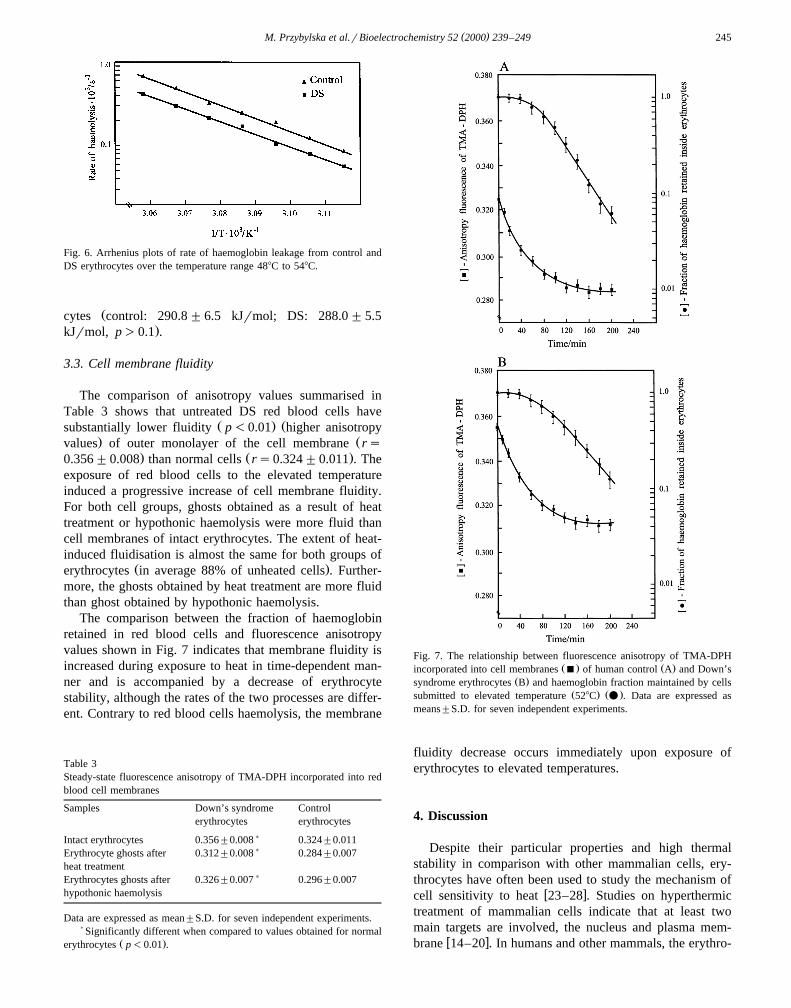
( )M. Przybylska et al.rBioelectrochemistry 52 2000 239–249 245
Fig. 6. Arrhenius plots of rate of haemoglobin leakage from control andDS erythrocytes over the temperature range 488C to 548C.
Žcytes control: 290.8 "6.5 kJrmol; DS: 288.0 "5.5.kJrmol, p)0.1 .
3.3. Cell membrane fluidity
The comparison of anisotropy values summarised inTable 3 shows that untreated DS red blood cells have
Ž . Žsubstantially lower fluidity p-0.01 higher anisotropy. Žvalues of outer monolayer of the cell membrane rs
. Ž .0.356"0.008 than normal cells rs0.324"0.011 . Theexposure of red blood cells to the elevated temperatureinduced a progressive increase of cell membrane fluidity.For both cell groups, ghosts obtained as a result of heattreatment or hypothonic haemolysis were more fluid thancell membranes of intact erythrocytes. The extent of heat-induced fluidisation is almost the same for both groups of
Ž .erythrocytes in average 88% of unheated cells . Further-more, the ghosts obtained by heat treatment are more fluidthan ghost obtained by hypothonic haemolysis.
The comparison between the fraction of haemoglobinretained in red blood cells and fluorescence anisotropyvalues shown in Fig. 7 indicates that membrane fluidity isincreased during exposure to heat in time-dependent man-ner and is accompanied by a decrease of erythrocytestability, although the rates of the two processes are differ-ent. Contrary to red blood cells haemolysis, the membrane
Table 3Steady-state fluorescence anisotropy of TMA-DPH incorporated into redblood cell membranes
Samples Down’s syndrome Controlerythrocytes erythrocytes
)Intact erythrocytes 0.356"0.008 0.324"0.011)Erythrocyte ghosts after 0.312"0.008 0.284"0.007
heat treatment)Erythrocytes ghosts after 0.326"0.007 0.296"0.007
hypothonic haemolysis
Data are expressed as mean"S.D. for seven independent experiments.)Significantly different when compared to values obtained for normal
Ž .erythrocytes p-0.01 .
Fig. 7. The relationship between fluorescence anisotropy of TMA-DPHŽ . Ž .incorporated into cell membranes B of human control A and Down’s
Ž .syndrome erythrocytes B and haemoglobin fraction maintained by cellsŽ . Ž .submitted to elevated temperature 528C v . Data are expressed as
means"S.D. for seven independent experiments.
fluidity decrease occurs immediately upon exposure oferythrocytes to elevated temperatures.
4. Discussion
Despite their particular properties and high thermalstability in comparison with other mammalian cells, ery-throcytes have often been used to study the mechanism of
w xcell sensitivity to heat 23–28 . Studies on hyperthermictreatment of mammalian cells indicate that at least twomain targets are involved, the nucleus and plasma mem-
w xbrane 14–20 . In humans and other mammals, the erythro-

( )M. Przybylska et al.rBioelectrochemistry 52 2000 239–249246
cytes were highly differentiated and have lost almost allcellular organelles. Therefore, thermosensitivity of humanerythrocytes must not have been associated with DNAinjury and the rate-limiting step of heat-induced haemoly-sis is related to different kind of molecular mechanisms.The crucial sequence of heat-induced events in cells lead-
w xing to cell death has previously been described 23–28 .Similar studies on the effects of elevated temperatures oncells from individuals suffering from DS have not yet beenpublished.
Although biochemical disturbances of the reactive oxy-gen species metabolism revealed in subjects with DS have
w xwidely been described 9–12,38,39 , the sensitivity of tri-Ž .somic q21 cells to the various physical and chemical
agents which increase a cellular oxidative stress has stillremained controversial. The occurrence of chromosomalaberrations induced by ionising radiation and different
w xchemical mutagens was elevated 40–42 or decreasedw x43,44 , depending considerably on the experimental condi-tions. Moreover, the study of gamma radiation- andacetylphenylhydrazine-induced red blood cells haemolysisshowed no statistically significant differences of the sensi-tivity of cells obtained from DS patients with respect tothose from normal donors. However, DS erythrocytes weremore resistant to the action of photoactivated riboflavinmonitored by haemolysis and haemoglobin oxidationw x45,46 . Assuming that haemolytic effect of supraoptimaltemperatures is, at least, partly mediated by highly reactive
w xoxygen species 21,22 , it may be expected that the effectof their cytotoxic action will be different in case of cellsfrom individuals suffering from Down syndrome.
Our experimental data obtained by kinetic analysis ofcell-survival curves, accompanied by flow cytometry andSEM examinations, indicated that the trisomy of 21 chro-mosomes significantly affected thermostability of red bloodcells. Erythrocytes submitted to elevated temperatures lostgradually their biconcave shape, formed vesicles and fi-nally lysed. Flow cytometry scans which reflected thesemodifications showed significant differences in erythrocyte
Ž . Žvolume LS1 diagrams , shape and surface structure LS2. Ždiagrams upon the influence of heat stress Figs. 1B–C.and 2B–C . As shown in Figs. 1B–C, 2B–C and 3, DS red
blood cells were more resistant, to a certain extent, todisruption induced by heat which was indicated by theconsiderably reduced fraction of altered cells, compared to
Ž .normal ones, especially for longer incubation time )1 h .Ž .At each temperature from 488C to 548C , the fractions
of haemoglobin retained by red blood cells from DSpatients were greater than for erythrocytes of healthyindividuals. The similar values of D parameters calcu-q
Žlated for DS cells as well as for normal erythrocytes Table.2, Fig. 4 showed that the preleakage period which indi-
cated the ability of red blood cells to accumulate sublethaldamage before a release of haemoglobin was inverselyproportional to temperature and almost the same for bothcell groups.
The comparison of temperature dependence of D pa-0Ž .rameters Table 2, Fig. 4 showed that D parameters0
were greater for DS cells as compared to normal ones andthe differences were inversely proportional to the tempera-ture. Similar values of D parameters calculated for the0
Ž .temperature of 54.58C Fig. 5 indicate that this tempera-ture seems to be critical for the thermally induced haemol-ysis. This confirms the hypothesis proposed by Lepock et
w xal. 26 , which assumed that the rate-limiting step ofheat-treated erythrocytes was the denaturation of cell mem-brane proteins. The temperature of about 548C was possi-bly responsible for the denaturation of spectrin and cyto-
Ž .plasmic domain of band 3 protein. The rate constants kof thermally induced haemolysis, calculated from linearsemilogarithmic transformation of the dose–response
Ž .curves, were greater for normal cells p-0.01 . The HSRshowed that DS red blood cells were in average 1.60 timesmore resistant to heat-induced haemolysis than those from
Ž .normal subjects Table 2 .Activation energies of haemolysis calculated according
to Arrhenius equation were relatively high and similar forŽboth cell groups control — 290.8"6.5 kJrmol; DS —
.288.0"5.5 kJrmol; p)0.1 . A straight line of ArrheniusŽ .plot Fig. 6 suggests a single mechanism for lysis. Our
w xdata corroborate results obtained by Lepock et al. 26 andconfirm the hypothesis that membrane protein denaturationis the rate-limiting step of thermally induced haemolysisw x17,18,25 .
We have also studied the relationship between ther-mosensitivity of red blood cells and cell membrane fluid-ity. The comparison of anisotropy values obtained for thefluorescent probe TMA-DPH, summarised in Table 3,shows that untreated DS red blood cells have substantially
Ž . Ž .lower fluidity p-0.01 higher anisotropy values ofŽ .outer monolayer of cell membrane rs0.356"0.008
Ž .than normal cells rs0.324"0.011 . Our results arew xsimilar to those reported by Kantar et al. 47 . They
showed that erythrocytes from DS individuals exhibit adecreased cell membrane fluidity at the lipid–water inter-face region.
The relationship between the alteration of DS erythro-cyte membrane rigidity and modification of cell membranecomposition has not been well established. Data reported
w xby Kantar et al. 47 show that the major factors influenc-ing membrane fluidity, i.e. cholesterol and a phospholipidcontent and the cholesterol to phospholipid molar ratio, arealmost the same in erythrocyte membranes from DS indi-viduals and normal donors. The analysis of the concentra-tion of polyunsaturated fatty acids in erythrocyte cell
w xmembranes 12 showed considerable changes of the rela-tive proportion of different groups of these compounds.The linoleic acid levels as well as elongation products of
Žarachidonic acid adrenic acid, docosatetrenoic acid and.docosapentenoic acid were significantly lowered in DS
patients. In contrast, the concentrations of long-chain poly-Žunsaturated fatty acids dihomo-g-linolenic acid and

( )M. Przybylska et al.rBioelectrochemistry 52 2000 239–249 247
.arachidonic acid and docosahexenoic acid were increasedin DS individuals, compared with the control group. Therelative elevation in DS patients of docosahexenoic acid
Žlevel, the most susceptible to peroxidation due to the.highest number of double bonds polyunsaturated fatty
acid, suggested that its content in cell membranes was notinfluenced substantially by the oxidative damage, probablybecause of the compensatory increase of the antioxidantenzyme cell system. The relationship between antioxidantenzyme activities and lipid peroxidation mediated by oxy-gen reactive species in DS subjects is still controversial.Most authors suggest the augmentation of oxidative stress
w xin DS individuals 11,48,49 , while others indicate that anincrease in lipid peroxidation may not be directly related to
w xan elevation of superoxide dismutase activity 50,51 . Sim-w xilar results obtained by Yatvin 52 show that the resistance
to supraoptimal temperatures is related to the degree andamount of unsaturated fatty acids incorporated into cellmembranes. A decrease of the amount of unsaturated fattyacid incorporated into cell membrane considerably de-creased cell killing.
Our results, which indicate that the DS red blood cellsmembrane fluidity is considerably diminished, are in con-
w xtrast with data obtained by Pastor et al. 12 , showing theincrease of the fatty acid unsaturation level in DS individu-als and with a current view that the higher degree of theacyl-chain unsaturation causes the cell membrane fluidisa-
w xtion 53 . In this context, we imply that the augmentationof DS red blood cells membrane fluidity is probablyrelated to a factor different than the enlargement of fattyacids unsaturation level. Our suggestion is based on theanalysis of scattering diagrams obtained for intact DS
Ž .erythrocytes Table 1 . They show that the scattering prop-Žerties of red blood cells from DS patients reflecting
erythrocyte size, shape and cell membrane surface mor-.phology are different than those of erythrocytes of healthy
individuals. Those results are confirmed by Kedziora et al.w x w x54,55 as well as Chapman et al. 56,57 . They indicatedthe existence of structural modifications of DS eryth-rocytes, suggesting the loss of membrane surface andconcluded that the protein conformational changes wereresponsible for membrane structural alterations of DS ery-throcytes revealed by SEM examinations, rather than thecell membrane lipid composition and the degree of theacyl-chain unsaturation.
A contribution of cell membrane fluidity to the develop-ment of thermostability has been reported occasionallyw x52,53,58–60 . However, the evidence for an associationbetween membrane fluidity and the red blood cells ther-mosensitivity has not been well established.
The exposure of red blood cells to the elevated tempera-tures increased progressively the cell membrane fluidity ina time-dependent manner and this was accomplished by adecrease of red blood cell stability, although the course ofthese two changes was different. For both cell groups,ghosts obtained as a result of the heat treatment or hypo-
tonic haemolysis were more fluid than cell membranes ofŽ .intact erythrocytes Table 3 . The extent of heat-induced
fluidisation was almost the same for both groups ofŽ .erythrocytes in average 88% of unheated cells . Further-
more, the ghosts obtained by a heat treatment were morefluid than ghosts obtained by the hypotonic haemolysis.Unlike haemolysis which was characterised by an apparentdelay represented by a shoulder region of time responsecurves, a decrease of cell membrane fluidity diminishedimmediately upon exposure of cells to the heat treatmentand they might be considered as a part of damage accumu-
Ž .lated during the preleakage period of lysis Fig. 7 . Therelationship between fluidity and rate of haemolysis sug-gests that the rate-limiting step in the haemolysis involves
w xthe rupturing of the cell membrane lipid bilayer 61 .In conclusion, our experimental data suggest that tri-
somy of 21 chromosomes significantly affects thermosta-bility of red blood cells, although the mechanisms oflowered thermosensitivity of erythrocytes from personssuffering from DS still remain unclear. Better understand-ing of this phenomenon would have potential value forimproving the therapeutic effects of hyperthermia as a partof anticancer therapy.
Acknowledgements
–The authors would like to thank Dr. Milosz Faber and
Dr. Andrzej Zaborowski for their excellent technical assis-–
tance, Dr. Miroslaw Przybylski for his advice and help in–´statistical analysis of data, and Mr. Jacek Swietoslawski
from the Electron Microscopy Laboratory of the MedicalUniversity of Lodz for his kind help in electron micro-scopic measurements.
References
w x1 R.F. Mueller, I.D. Young, Chromosome disorders, Emery’s Ele-ments of Medical Genetics, Livingstone, Edinburgh, 1998, pp. 245–248.
w x2 P.E. Polani, E. Alberman, A.C. Berry, S. Blunt, J.D. Singer, Chro-Ž .mosome abnormalities and maternal age, Lancet 2 1976 516–517.
w x3 B.L. Shapiro, The environmental basis of the Down syndromeŽ .phenotype, Dev. Med. Child Neurol. 36 1994 84–90.
w x4 C.T. Fong, G.M. Brodeur, Down’s syndrome and leukemia: epi-demiology, genetics, cytogenetics and mechanism of leukemogene-
Ž .sis, Cancer Genet. Cytogenet. 28 1987 55–76.w x5 L. Iselius, P. Jacobs, N. Morton, Leukaemia and transient leukaemia
Ž .in Down syndrome, Hum. Genet. 85 1990 477–485.w x Ž .6 L.L. Robinson, Down syndrome and leukemia, Leukemia 6 1992
5–7.w x7 A.H. Ragab, A. Abdel-Mageed, J.J. Shuster, L.S. Frankel, J. Pullen,
J. van Eys, M.P. Sullivan, J. Boyett, M. Borowitz, W.M. Christ,Clinical characteristic and treatment outcome of children with acute
Ž .lymphatic leukemia and Down’s syndrome, Cancer 67 1991 1057–1063.

( )M. Przybylska et al.rBioelectrochemistry 52 2000 239–249248
w x8 F. Lampert, U. Bertram, H. Riehm, Acute lymphoblastic leukaemia,Ž .in: P.N. Plowman, C.R. Pinkerton Eds. , Paediatric Oncology.
Clinical Practice and Controversies, Chapman & Hall, London,1992, pp. 180–182.
w x Ž .9 P. Benson, Gene dosage effect in trisomy 21, Lancet 1 1975584–587.
w x10 J. Kedziora, G. Bartosz, Down’s syndrome: a pathology involvingthe lack of balance of reactive oxygen species, Free Radic. Biol.
Ž .Med. 4 1988 317.w x11 G. Gerli, L. Zenoni, G.F. Locateelli, R. Mongiat, F. Piattoni, G.B.
Orsini, A. Montagnani, M.R. Gueli, V. Gualandri, Erythrocyte an-tioxidant system in Down syndrome, Am. J. Med. Genet. Suppl. 7Ž .1990 272–330.
w x12 M.C. Pastor, C. Sierra, M. Dolade, E. Navarro, N. Brandi, E. Cabre,A. Mira, A. Seres, Antioxidant enzymes and fatty acid status in
Ž .erythrocytes of Down’s syndrome patients, Clin. Chem. 44 1998924–929.
w x13 J.R. Lepock, Involvement of membranes in cellular responses toŽ .hyperthermia, Radiat. Res. 92 1982 433–438.
w x14 A.W.T. Konings, Effect of heat and radiation on mammalian cells,Ž .Radiat. Phys. Chem. 30 1987 339–349.
w x15 A.W.T. Konings, Membranes as targets for hyperthermic cell killing,Ž .Recent Results Cancer Res. 109 1988 10–21.
w x16 E.W. Gerner, Thermal dose and time-temperature factors for biologi-Ž .cal responses to heat shock, Int. J. Hyperthermia 3 1987 319–327.
w x17 W.C. Dewey, The search for critical cellular targets damaged byŽ .heat, Radiat. Res. 120 1989 191–204.
w x18 W.C. Dewey, L.E. Hopwood, S.A. Sapareto, L.E. Gerweck, Cellularresponses to combination of hyperthermia and radiation, Radiology
Ž .123 1989 463–474.w x19 T.R. Tritton, J.A. Hickman, How to kill cancer cells: membranes
and cell signalling as targets in cancer chemotherapy, Cancer Cells 2Ž .1990 95–105.
w x20 A. Laszlo, The effects of hyperthermia on mammalian cell structureŽ .and function, Cell Proliferation 25 1992 59–87.
w x21 R.D. Issels, J.E. Biaglow, L. Epstein, L.E. Gerwick, Enhancement ofcysteamine cytotoxicity by hyperthermia and its modification bycatalase and superoxide dismutase in Chinese hamster ovary cells,
Ž .Cancer Res. 44 1984 3911–3915.w x22 E. Lengfelder, R.M. Fink, Oxidative stress during hyperthermia, in:
Ž .T. Sugahara, M. Saito Eds. , Hyperthermic Oncology 2 Taylor andFrancis, London, 1989, pp. 350–362.
w x23 T.H. Ham, S.C. Shen, E.M. Fleming, W.B. Castle, Studies on theŽ .destruction of red blood cells, Blood 3 1948 373–403.
w x24 L.A. Crum, W.T. Coakley, J.O.T. Deeley, Instability development inŽ .heated human erythrocytes, Biochim. Biophys. Acta 554 1979
76–89.w x25 N.L. Gershfeld, M. Murayama, Thermal instability of red blood cells
membrane bilayers: temperature dependence of haemolysis, J.Ž .Membr. Biol. 101 1988 67–72.
w x26 J.R. Lepock, H.E. Frey, H. Bayne, J. Markus, Relationship ofhyperthermia-induced hemolysis of human erythrocytes to the ther-mal denaturation of membrane proteins, Biochim. Biophys. Acta 980Ž .1989 190–201.
w x27 M. Koter, Erythrocyte vesiculation after single or double heat shock,Ž .Med. Sci. Res. 20 1992 633–639.
w x28 M. Koter, Changes in erythrocyte membranes after fractionatedŽ .hyperthermia, Cell Biol. Int. 17 1993 665–670.
w x29 I.T. Ivanov, Relationship between thermohemolysis and the ther-moinduced disturbance of the barrier properties of human erythro-
Ž .cyte membranes, C. R. Acad. Bulg. Sci. 45 1992 129–132.w x30 I.T. Ivanov, Investigation into the membrane alteration relevant to
Ž .the mechanism of thermohaemolysis, J. Therm. Biol. 21 1996205–212.
w x31 I.T. Ivanov, Thermohaemolysis of mammalian erythrocytes, J.Ž .Therm. Biol. 18 1993 177–183.
w x32 G.C. Salzman, S.B. Singham, R.G. Johnston, C.F. Bohren, Light
scattering and cytometry, Flow Cytometry and Sorting, Wiley-Liss,New York, 1990.
w x33 K.D. Bauer, K.J. Henle, Arrhenius analysis of heat survival curvesŽ .from normal and thermotolerant CHO cells, Radiat. Res. 78 1979
251–263.w x34 J.G. Kuhry, G. Duportail, C. Bronner, G. Laustriat, Plasma mem-
brane fluidity measurements in whole living cells by fluorescenceanisotropy of trimethylammonium-diphenylhexatriene, Biochim.
Ž .Biophys. Acta 845 1985 60–67.w x35 B.W. Van der Meer, Biomembrane structure and function viewed by
Ž .fluorescence, in: H.J. Hilderson, J.R. Harris Eds. , SubcellularBiochemistry. Fluorescence Studies on Biological Membranes,Plenum, New York, 1988, pp. 1–53.
w x36 M. Shinitzky, Y. Barenholz, Fluidity parameters of lipid regionsdetermined by fluorescence polarisation, Biochim. Biophys. Acta
Ž .515 1978 367–394.w x37 J.H. Zar, Biostatistical Analysis, Prentice-Hall, London, 1984.w x38 W.W. Feaster, L.W. Kwok, C.J. Epstein, Dosage effects for superox-
ide dismutase-1 in nucleated cells aneuploid for chromosome 21,Ž .Am. J. Hum. Genet. 29 1977 563–570.
w x39 P.M. Sinet, Metabolism of oxygen derivatives in Down’s syndrome,Ž .Ann. N. Y. Acad. Sci. 396 1982 83–94.
w x40 D. Viyalaxmi, H. Evans, Bleomycin-induced chromosomal aberra-Ž .tions in Down’s syndrome lymphocytes, Mutat. Res. 105 1982
107–113.w x41 N.S. Raji, K.S. Rao, Trisomy 21 and accelerated aging: DNA-repair
parameters in peripheral lymphocytes of Down’s syndrome patients,Ž .Mech. Ageing Dev. 100 1998 85–101.
w x42 R. Preston, X-ray-induced chromosome aberrations in Down lym-phocytes: an explanation of their increased sensitivity, Environ.
Ž .Mutagen. 3 1981 85–89.w x43 R. Łukaszewicz, E. Sibinska, J. Kedziora, Down’s syndrome: de-´
creased radiation sensitivity of lymphocytes’ DNA, Hereditas 97Ž .1982 155–156.
w x44 J. Kedziora, E. Sibinska, B. Rozga, G. Bartosz, Gamma-radiation´ ´Ž .sensitivity of fibroblast DNA in trisomy 21, Hereditas 105 1986
161–162.w x45 G. Bartosz, W. Leyko, J. Kedziora, J. Jeske, Radiation sensitivity of
erythrocytes with an increased level of superoxide dismutase, Int. J.Ž .Radiat. Biol. 31 1977 19–200.
w x46 G. Bartosz, W. Leyko, J. Kedziora, J. Jeske, Superoxide dismutaseand radiation-induced haemolysis: no benefit of its increased content
Ž .in red cells, Int. J. Radiat. Biol. 38 1980 187–192.w x47 A. Kantar, P.L. Giorgi, G. Curatola, R. Fiorini, Alterations in
erythrocyte membrane fluidity in children with trisomy 21: a fluores-Ž .cence study, Biol. Cell 75 1992 135–138.
w x48 J. Kedziora, G. Bartosz, Down’s syndrome: a pathology involvingthe lack of balance of reactive oxygen species, Free Radic. Biol.
Ž .Med. 4 1988 317–330.–
w x49 J. Kedziora, G. Bartosz, J. Gromadzinska, M. Sklodowska, W.´´Wasowicz, J. Scianowski, Lipid peroxides in blood plasma and
enzymatic antioxidative defence of erythrocytes in Down’s syn-Ž .drome, Clin. Chim. Acta 154 1986 191–194.
w x50 K.G. Anneren, C.J. Epstein, Lipid peroxidation and superoxidedismutase-1 and glutathione peroxidase activities in trisomy 16 fetal
Ž .mice and human trisomy 21 fibroblasts, Pediatr. Res. 21 198788–92.
w x51 M. Hayn, K. Kremser, N. Singewald, N. Cairns, M. Nemethova, G.Lubec, Evidence against the involvement of reactive oxygen speciesin the pathogenesis of neuronal death in Down’s syndrome and
Ž .Alzheimer’s disease, Life Sci. 59 1996 537–544.w x52 M.B. Yatvin, The influence of membrane composition and procaine
Ž .on hyperthermic death of cells, Int. J. Radiat. Biol. 32 1977513–521.
w x53 T. Suda, N. Maeda, M. Sekiya, K. Matsuoka, R. Tokita, T. Shiga,Influence of membrane cholesterol on the human red cell properties,
Ž .Med. J. Osaka Univ. 29 1978 21–27.

( )M. Przybylska et al.rBioelectrochemistry 52 2000 239–249 249
w x54 J. Kedziora, M. Koter, G. Bartosz, Influence of acrocentric chromo-some G-21 on the structural changes of erythrocyte membranes,
Ž .Stud. Biophys. 90 1982b 119–120.w x55 J. Kedziora, M. Koter, H. Bartel, G. Bartosz, W. Leyko, Ultrastruc-
tural modifications of erythrocyte membrane in Down’s syndrome,Ž .Acta Biol. Med. Ger. 40 1981 423–428.
w x56 I.V. Chapman, M. Koter, J. Kedziora, M. Soszynski, G. Bartosz,´Erythrocyte membrane thiol groups and protein composition in
Ž .Down’s syndrome, IRCS Med. Sci. 14 1986 267–268.w x57 I.V. Chapman, W. Leyko, K. Gwozdzinski, M. Koter, E. Grzelinska,´ ´ ´
G. Bartosz, Hyperthermic modification of bleomycin-DNA interac-Ž .tion detected by electron spin resonance, Radiat. Res. 96 1983
518–524.
w x58 A.W.T. Konings, A.C.C. Ruifrok, Role of membrane lipids andmembrane fluidity in thermosensitivity and thermotolerance of mam-
Ž .malian cells, Radiat. Res. 102 1985 86–94.w x59 J.R. Dynlacht, M.H. Fox, Heat-induced changes in the membrane
fluidity of Chinese hamster ovary cell measured by flow cytometry,Ž .Radiat. Res. 130 1992a 48–54.
w x60 J.R. Dynlacht, M.H. Fox, The effect of 458C hyperthermia on theŽ .membrane fluidity of cells of several lines, Radiat. Res. 130 1992b
55–59.w x61 A.E. Cress, E.W. Gerner, Cholesterol levels inversely reflect the
Ž .thermal sensitivity of mammalian cells in culture, Nature 283 1980677–679.