thermal shock induces host proteostasis disruption...
TRANSCRIPT

Thermal Shock Induces Host Proteostasis Disruption andEndoplasmic Reticulum Stress in the Model Symbiotic CnidarianAiptasiaClinton A. Oakley,*,† Elysanne Durand,‡ Shaun P. Wilkinson,† Lifeng Peng,† Virginia M. Weis,§
Arthur R. Grossman,∥ and Simon K. Davy†
†School of Biological Sciences, Victoria University of Wellington, Wellington 6012, New Zealand‡Department of Ecology and Environmental Sciences, Universite Pierre et Marie Curie, Paris 75005, France§Department of Integrative Biology, Oregon State University, Corvallis, Oregon 97331, United States∥Department of Plant Biology, The Carnegie Institution for Science, Stanford, California 94305, United States
*S Supporting Information
ABSTRACT: Coral bleaching has devastating effects on coralsurvival and reef ecosystem function, but many of thefundamental cellular effects of thermal stress on cnidarianphysiology are unclear. We used label-free liquid chromatog-raphy−tandem mass spectrometry to compare the effects ofrapidly (33.5 °C, 24 h) and gradually (30 and 33.5 °C, 12days) elevated temperatures on the proteome of the modelsymbiotic anemone Aiptasia. We identified 2133 proteins inAiptasia, 136 of which were differentially abundant betweentreatments. Thermal shock, but not acclimation, resulted insignificant abundance changes in 104 proteins, including thoseinvolved in protein folding and synthesis, redox homeostasis,and central metabolism. Nineteen abundant structural proteinsshowed particularly reduced abundance, demonstrating proteostasis disruption and potential protein synthesis inhibition. Heatshock induced antioxidant mechanisms and proteins involved in stabilizing nascent proteins, preventing protein aggregation anddegrading damaged proteins, which is indicative of endoplasmic reticulum stress. Host proteostasis disruption occurred beforeeither bleaching or symbiont photoinhibition was detected, suggesting host-derived reactive oxygen species production as theproximate cause of thermal damage. The pronounced abundance changes in endoplasmic reticulum proteins associated withproteostasis and protein turnover indicate that these processes are essential in the cellular response of symbiotic cnidarians tosevere thermal stress.
KEYWORDS: Aiptasia, cnidaria, coral reefs, endoplasmic reticulum, label-free proteomics, symbiosis, thermal stress,unfolded protein response
■ INTRODUCTION
Corals are the engineers of highly diverse coral reef ecosystems,building the physical structure of the reef and contributing toreef nutrient cycling through their production of photo-synthates and labile mucus.1 Reef-building corals are amutualism between the coral animal host and intracellulardinoflagellate symbionts of the genus Symbiodinium, in whichalgal photosynthates are exchanged for nitrogenous compoundsand inorganic carbon from the host.2 Existing in a highlyenergetic environment in shallow tropical seas and exposed towarm temperatures and high ultraviolet radiation, coral reefsrequire a narrow temperature range and are sensitive toextreme thermal events.3 These abiotic stresses elicit damageDNA, membranes, and proteins and induce universal cellularstress responses (CSRs).4,5 Corals are incapable of thermalhomeostasis and, as sessile organisms, must acclimate in situ at
the cellular level to changes in water temperature. The coral−dinoflagellate symbiosis and reefs worldwide are increasinglythreatened by anthropogenic climate change, ocean acid-ification, and coral bleaching, in which periods of high seasurface temperatures result in the loss of algal symbionts fromthe host, potentially resulting in regional coral mortality.6,7
Coral tissues possess ideal conditions for the generation ofreactive oxygen species (ROS) owing to their high metabolicrates and the abundant oxygen produced by photosynthesis oftheir endosymbiotic algal partners.8 There have been extensiveefforts to investigate the roles of light, ROS, and oxidativedamage during thermal stress in symbiotic cnidarians,3,9−12 andalgal symbiont-derived ROS have been proposed as instigators
Received: August 31, 2016Published: May 5, 2017
Article
pubs.acs.org/jpr
© XXXX American Chemical Society A DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX

of the bleaching response.8,12 Recently, increased attention hasbeen paid to the role of host endogenous ROS and nitric oxide(NO) generation during cnidarian thermal stress.10,13 Theendoplasmic reticulum (ER) is a potent source of up to 25% oftotal ROS generation in eukaryotic cells due to the transfer ofelectrons to O2 during peptide disulfide bond formation.14
Maintaining proteome stability, or proteostasis, during thermalstress depends on the rapid initiation of the highly conservedheat shock response, primarily the induction of heat shockproteins (Hsp’s), which serve as chaperones to maintain correctprotein folding.15 Misfolded proteins are potentially toxic andprone to aggregation and must be repaired or destroyed.15 Theunfolded protein response (UPR) is induced by the aggregationand import of misfolded proteins beyond the processingcapacity of the ER protein-folding machinery, and it mitigatesER stress by refolding misfolded or damaged proteins.16,17 ERstress provokes a number of cellular responses in addition tothe UPR, including ER expansion, increased phospholipidsynthesis, chaperone upregulation, reduced global proteintranslation, and enhanced protein export from the ER.16
Collectively, these mechanisms serve to increase protein-foldingcapacity and purge misfolded or aggregated proteins from theER.18 If these mechanisms are insufficient, the terminal step ofthe UPR is apoptosis. Host apoptosis has been demonstrated inresponse to thermal stress in symbiotic cnidarians,19 but its rolein bleaching is disputed.20
Determining the mechanisms used by the cnidarian host toacclimate to varying thermal conditions and severe thermalstress is important to better understand the resilience of thecoral−dinoflagellate mutualism in future climate-changescenarios. Corals are differentially susceptible to bleachingdue to host and symbiont identities, local physical ocean-ography (e.g., depth and water flow), and frequency ofanomalous thermal events.21 Thermal coral bleaching can beinduced by sea surface temperature fluctuations of as little as 1to 2 °C greater than the average maximum sustainedtemperature.22 Therefore, the host physiological mechanismsof acclimation to high temperatures as well as the CSRsinduced by rapid elevation in temperature (“heat shock”) arecrucial. The circumtropical anemone Aiptasia is a modelorganism for studies of the coral−dinoflagellate symbiosis,23,24
and genetic and gene expression databases of this system arerapidly expanding.25−27 To compare the effects on thecnidarian host of gradual high-temperature acclimation tothose of a rapid thermal assault, Aiptasia were either acclimatedto higher temperatures over a period of 2 weeks or wereexposed to short-term (24 h) thermal shock withoutacclimation. We then used label-free liquid chromatography−electrospray ionization mass spectrometry (LC−ESI−MS/MS)“shotgun” proteomics to characterize the Aiptasia proteomefollowing each of these treatments. We performed a detailedanalysis of the specific proteins and cell mechanisms induced byacclimation and heat shock, providing additional insight intothe cellular biology of the initial phases of coral thermal stressand the interplay between the host and symbiont prior todetectable bleaching.
■ MATERIALS AND METHODS
Aiptasia Culture and Physiology
Aiptasia sp. (= Exaiptasia pallida28) were isolated from aPacific-sourced aquarium, and a clonal population, strain NZ1,was maintained in the laboratory for several years at 25 °C and
∼40 μmol quanta m−2 s−1. We have maintained using theAiptasia nomenclature here for agreement with publishedgenomic resources.26 This host strain associates withSymbiodinium minutum (clade B).24,29 Pairs of anemoneswere maintained in 500 mL glass beakers at 25 °C and 100μmol quanta m−2 s−1 with a 12 h/12 h light/dark cycle.Anemones were allowed to acclimate to these conditions for 2weeks. Anemones were fed Artemia sp. nauplii, and the waterwas changed with 0.22 μm filtered seawater twice per week. Tominimize contamination from Artemia protein, anemones werenot fed for 1 week prior to sampling. The dark-adaptedquantum yield of chlorophyll fluorescence of three to sixanemones per treatment was measured daily using a divingpulse-amplitude modulated (PAM) fluorometer (Heinz Walz,Effeltrich, Germany) to monitor the maximum efficiency ofphotosystem II, beginning 4 days before the start of theexperiment. Sampling was conducted by homogenizing thepooled anemones, followed by separating the algal symbiontsfrom the host homogenate by centrifugation. Symbionts werefixed with formalin and counted using a hemocytometer toassess symbiont density. Sample protein content wasdetermined by the Bradford assay. All reagents were acquiredfrom Sigma-Aldrich (Auckland, New Zealand) unless noted.
Experimental Design and Statistical Rationale
Anemones were exposed to four thermal treatments: control(25 °C), 30 °C with acclimation, 33.5 °C with acclimation, anda 33.5 °C “heat shock” treatment without thermal acclimation.These temperatures were chosen to represent the range knownto induce sublethal thermal stress and bleaching in Aiptasia.13,30
Six biological samples (n = 6), each consisting of two pooledanemones, were analyzed per treatment, and each biologicalsample (24 total) was analyzed twice, forming technicalreplicates. After 2 weeks of acclimation under controlconditions, the temperature of the anemones in the 30 and33.5 °C-acclimated treatments was increased by 1 °C per day to30 °C, and the anemones were maintained at this temperaturefor 5 days. For the 33.5 °C-acclimated treatment, thetemperature was further increased to 33.5 °C over 7 days,followed by sampling. Control, 30 °C-acclimated, and 33.5 °C-acclimated anemones were sampled simultaneously. Anemonesexposed to heat shock were acclimated to the 25 °C controltank for 2 weeks, after which the temperature was increased to33.5 °C over 1 h, and the anemones were sampled 24 h later.All samples were analyzed in a stratified random order tominimize the effects of instrument drift.
LC−ESI−MS/MS Methods
Samples (20 μg final protein mass) were prepared by a sodiumdeoxycholate in-solution digestion method with 1 μg ofsequencing-grade trypsin using the methods and instrumentsettings described in Oakley et al.27 Samples were analyzed byLC−ESI−MS/MS with a 300 min gradient (buffer A: 0.1%formic acid; buffer B: 80% acetonitrile, 0.1% formic acid) at 0.3μL min−1 on an Acclaim PepMap C18, 3 μm, 100 Å column(Thermo Scientific, Auckland, New Zealand) and Ultimate3000 (Dionex, Sunnyvale, CA), with the column oven at 35 °C.An LTQ Orbitrap XL (Thermo Scientific) was used to analyzepeptides that were injected at a 1.9 kV spray voltage with aresolution of 30 000. The ion trap analyzed MS/MS spectrafrom the top eight MS peaks, rejecting +1 charge states withdynamic exclusion enabled (180 s). The LC−ESI−MS/MSinstruments were operated with Chromeleon Xpress(v2.11.0.2914, Dionex), Thermo Xcalibur (v2.1), and Thermo-
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
B

Tune Plus (v2.5.5, Thermo Scientific). Spectra were searchedusing a custom sequence database previously used for Aiptasiaprotein identification.27 This database consists of the Aiptasiagenome,26 all UniProt cnidarian sequences, all open readingframes from an Aiptasia strain CC7 transcriptome,25 and adatabase of common contaminants31 (320 798 total sequences)and was processed by MScDb to reduce peptide-levelredundancy.32 Peak lists were generated by ProteomeDiscoverer (v1.4.1.14, Thermo Scientific) via SEQUEST HTsearches,33 followed by Percolator scoring. SEQUEST HTsearches assumed trypsin digestion with a maximum of twomissed cleavages, one minimum matching peptide per protein,a minimum peptide length of six amino acids, a maximum deltaCn of 0, a parent ion tolerance of 10.0 ppm, and a fragment ionmass tolerance of 0.60 Da. Carbamylation of the N-terminusand oxidation of methionine were specified as variablemodifications, and carbamidomethylation of cysteine wasspecified as a fixed modification.Proteins were quantified by total ion current (TIC) in
Scaffold (Table S1), and statistical analyses were carried out inR.34 The TIC values for all protein clusters per sample weresummed to determine if TIC normalization between sampleswas necessary. Variation in grand total TIC values was muchgreater between biological replicates than between technicalreplicates, indicating differences in total protein loadingbetween samples (Figure S1). To account for differences inprotein loading between samples, a scaling factor approach wasused based on the assumption that the majority of proteins arenot differentially expressed in response to changes intemperature or by indirect thermal effects (e.g., differentgrowth rates). A pairwise matrix of scaling factors for eachsample (x) against each other sample (y) was calculated as themedian of the vector
=xy
i m1, 2, 3, ...,i
i
where m is the total number of proteins that were detected inall 48 samples (345 proteins). The sample with the lowestprotein loading depth (the reference sample) was identified asthe matrix column with the lowest average scaling factor. Allother samples were then normalized to this sample using thescale factors from the reference sample column.To aid in parametric model-fitting, TIC values were then
assigned for low-abundance protein clusters whose concen-trations were below the detection limit of the instrument(“non-detects”). Protein clusters with nondetect rates exceed-ing 33% were removed from the data set, and the nondetects inthe remaining clusters were imputed by robust regression onorder statistics (assuming a log-normal distribution of proteinconcentrations within samples) using the R package “NADA”.35
Following imputation, the technical replicates were averaged,and protein clusters that differed in concentration betweentreatments were identified. For multivariate analysis oftreatment effects, the data set was log-transformed, andBray−Curtis dissimilarities were calculated for each pairwisesample comparison. Analysis of variance by permutation ofdissimilarities was carried out using the “adonis” function in theR package “vegan” (v2.2-1).36 To test for differences inindividual protein clusters between treatments, a generalizedlinear model with treatment effects was fitted and comparedagainst a null model (intercept only) using an analysis ofdeviance X2 test with α = 0.05. Protein clusters showingdifferences in concentration between treatments were identified
with the false discovery rate (FDR) correction procedure ofBenjamini and Hochberg,37 using a q-threshold of 0.1 (TableS2). To ensure that the regression on order statisticsimputation procedure did not have an undue impact on theresults, the procedure was repeated 100 times with randomreallocations of imputed values.Scaffold (v4.3.4, Proteome Software) was used for further
validation and for quantification via label-free TIC using theSEQUEST HT and X!Tandem algorithms (The GPM, thegpm.org; version Cyclone 2010.12.01.1). Protein probabilities wereassigned by Protein Prophet.38 FDRs were calculated using adecoy database.39 The FDR threshold for reported peptidesand proteins was set to 0.1 and 1%, respectively, with aminimum of two peptides per protein. Proteins that sharedsignificant peptide evidence were grouped into clusters usingScaffold’s Protein Cluster Analysis algorithm. Following furtherstatistical analysis (see above), protein clusters that were foundto be significantly different between treatments were annotatedby manual BLAST searches of each database sequence againstthe UniProtKB database. Each protein cluster was given theannotation of the UniProtKB entry with the highest BLASTscore that had an E value of <1.0 × 10−10. Functions wereinferred from these annotations, and fold changes (FCs) werederived by performing log2 transformation of proteinabundance ratios. To further verify the functions of significantlydifferent proteins, UniProtKB entries were assigned biologicalprocess gene ontology (GO) terms using QuickGO.40,41 Massspectrometry data were deposited to the ProteomeXchangeConsortium42 via the PRIDE partner repository with the dataset identifier PXD004257.
■ RESULTS AND DISCUSSION
Symbiont density and Fv/Fm are widely used physiologicalmetrics of coral bleaching,43 and our data indicate that changesin the host proteome preceded the breakdown of the symbiosis.Clear signs of thermal stress were observed in the hostproteome; however, symbiont density was not significantlydifferent at the time of sampling among any of the treatments(Figure 1A, p > 0.05, single-factor ANOVA). Furthermore,none of the treatments resulted in a significant change in thedark-adapted quantum yield of photosystem II (Fv/Fm), ameasure of the photophysiological health of the dinoflagellatesymbionts44 (Figure 1B).The levels of many highly abundant host proteins were
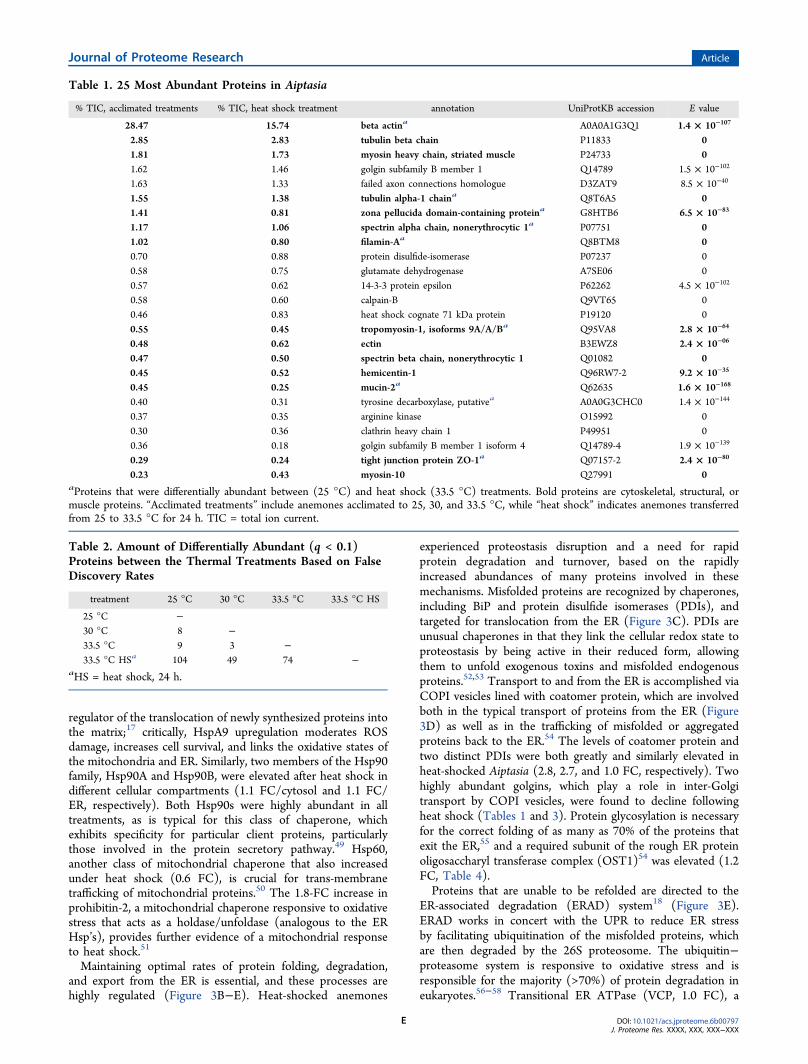
greatly reduced following heat shock conditions (33.5 °C, 24 h)when compared with those corresponding proteins in control(25 °C) or acclimated (30 and 33.5 °C) animals (Table 1).These proteins include several components involved in motorfunction, cellular matrix structure, and protein trafficking. Weidentified a total of 2187 protein clusters (from here on referredto as “proteins”) across all treatment groups using X!Tandem(Table S2). The 25 proteins with the greatest average TICvalues are listed in Table 1 in order of their average abundanceacross all samples, as determined by the percentage of theindividual protein TIC values relative to that of all proteins inthe given sample. We discarded 45 decoy sequences and ninecontaminant proteins from the data set, while 1253 proteinswere not analyzed further due to having >33% nondetects. Ofthe remaining 934 proteins, 593 had at least one value imputed.For all 100 iterations, the multivariate p values for alldifferentially expressed protein clusters identified in thegeneralized linear model analysis were <0.05. Significant
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
C

differences existed between treatments (multivariate p value =0.002).After anemones were acclimated to 30 or 33.5 °C, there were
only eight and nine proteins, respectively, that were differ-entially abundant (q < 0.1) between the control and acclimatedanimals (Tables 2 and 3). In contrast, there were many moredifferentially abundant proteins between the heat shocktreatment and the control, 30 °C acclimation, and 33.5 °Cacclimation treatments (104, 49, and 74 proteins, respectively;Table 2). The proteins that had significantly differentabundances between the control and heat shock treatmentsare given in Table 4, where they are organized by biologicalfunction. Differentially abundant proteins between the controland heat shock treatments were also assigned biological process
GO terms (Figure 2). A complete list of differentially abundantproteins is given in Table S3.Only nine proteins were differentially abundant between
control anemones kept at 25 °C and those that were allowed toslowly acclimate to 33.5 °C (Table 3). Of this limited set,metabolic enzymes include an increase in 75 kDa NADH-ubiquinone oxidoreductase subunit 1 (NDUFS1) and decreasesin pyruvate carboxylase and phosphoinositide phospholipaseCη2. This minimal number of changes, in contrast with thelarger differences between control and heat-shocked anemones,reflects the ability of the animal to maintain homeostasisthrough means that do not involve changes in proteinabundance. Two elongation factors, EF1γ and EF2, exhibitedreduced accumulation in anemones acclimated to 33.5 °C,which may indicate the arrest of cell division and animal growthunder sustained elevated temperature.45 Compared with theirlevels in control anemones, proteins that play roles in multiplethermal stress response mechanisms were upregulated in heat-shocked Aiptasia (Figure 2). From here on, unless otherwisestated, “differentially abundant proteins” refer to changesbetween control (25 °C) and 33.5 °C heat-shocked treatments.Our proposed model (Figure 3) interprets these changes asevidence of broad oxidative damage to proteins and disruptionof proteostasis through synergistic oxidative and ER stress,resulting in an increase in proteins associated with redoxcontrol, protein synthesis, and protein degradation.
Structural and Cytoskeletal Proteins
The effects of heat shock were most pronounced on the highlyabundant cytoskeletal, actin-interacting, cell−cell binding, andmuscle fiber proteins, which together comprised 45% of thetotal identified protein biomass (Table 1, Figure 3A). Nineteenproteins with structural, muscular, or cytoskeletal rolesexhibited significantly lower abundances under heat-shockconditions compared with controls (Table 4), including 14 ofthe 25 most abundant proteins. Interestingly, this groupincluded the most abundant detected protein, β-actin, whichexhibited a −1.1 log2 FC. Actin is the most abundant protein inthe eukaryotic cytoskeleton and the principle component of themicrofilament network; it is also a muscle fiber constituent andsensitive to oxidative damage.46 Oxidative stress has beendemonstrated to have broad effects on cytoskeleton organ-ization and structure as well as on the polymerization of itsconstituent proteins.47 Collectively, cytoskeletal and structuralprotein abundances were reduced by 51% in heat-shockedanemones compared with control organisms. The large,concerted reductions in these proteins signify reorganizationof the cell and disruption of multiple proteostasis mechanisms.
Induction of the Heat Shock and Unfolded ProteinResponses
The heat shock response and UPR are highly conservedmechanisms of responding to thermal disruption. Out of the 44conservatively defined “minimal stress proteome” (MSP)proteins that are universal among all three domains of life,5
we detected 10 with an increased abundance in heat-shockedAiptasia compared with controls (Table 4). This group includesmultiple classes of Hsp’s (Figure 3B). Two Hsp70s wereincreased in heat-shocked anemones: the binding immunoglo-bulin protein (BiP; 1.2 FC) and stress-70 protein (HspA9; 2.8FC). BiP is a principal ATP-dependent protein-folding andrefolding chaperone of the heat shock response localized in theER.48 HspA9 is an Hsp70-family chaperone located in themitochondrial matrix that serves as both a chaperone and a
Figure 1. Density and quantum yield of photosystem II chlorophyllfluorescence of algal symbionts in Aiptasia. Anemones were kept at 25°C (25 °C), acclimated to 30 °C (30 °C), acclimated to 33.5 °C (33.5°C), or heat-shocked at 33.5 °C for 24 h without thermal acclimation(33.5 °C HS). (A) Algal cell density at the time of proteome sampling,n = 6. (B) Dark-adapted quantum yield of photosystem II chlorophyllfluorescence of Aiptasia symbionts prior to and during the thermaltreatment, n = 3−6 per time point. Day 0 (dashed line) indicates theend of the acclimation period and the beginning of the thermaltreatments.
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
D
regulator of the translocation of newly synthesized proteins intothe matrix;17 critically, HspA9 upregulation moderates ROSdamage, increases cell survival, and links the oxidative states ofthe mitochondria and ER. Similarly, two members of the Hsp90family, Hsp90A and Hsp90B, were elevated after heat shock indifferent cellular compartments (1.1 FC/cytosol and 1.1 FC/ER, respectively). Both Hsp90s were highly abundant in alltreatments, as is typical for this class of chaperone, whichexhibits specificity for particular client proteins, particularlythose involved in the protein secretory pathway.49 Hsp60,another class of mitochondrial chaperone that also increasedunder heat shock (0.6 FC), is crucial for trans-membranetrafficking of mitochondrial proteins.50 The 1.8-FC increase inprohibitin-2, a mitochondrial chaperone responsive to oxidativestress that acts as a holdase/unfoldase (analogous to the ERHsp’s), provides further evidence of a mitochondrial responseto heat shock.51
Maintaining optimal rates of protein folding, degradation,and export from the ER is essential, and these processes arehighly regulated (Figure 3B−E). Heat-shocked anemones
experienced proteostasis disruption and a need for rapidprotein degradation and turnover, based on the rapidlyincreased abundances of many proteins involved in thesemechanisms. Misfolded proteins are recognized by chaperones,including BiP and protein disulfide isomerases (PDIs), andtargeted for translocation from the ER (Figure 3C). PDIs areunusual chaperones in that they link the cellular redox state toproteostasis by being active in their reduced form, allowingthem to unfold exogenous toxins and misfolded endogenousproteins.52,53 Transport to and from the ER is accomplished viaCOPI vesicles lined with coatomer protein, which are involvedboth in the typical transport of proteins from the ER (Figure3D) as well as in the trafficking of misfolded or aggregatedproteins back to the ER.54 The levels of coatomer protein andtwo distinct PDIs were both greatly and similarly elevated inheat-shocked Aiptasia (2.8, 2.7, and 1.0 FC, respectively). Twohighly abundant golgins, which play a role in inter-Golgitransport by COPI vesicles, were found to decline followingheat shock (Tables 1 and 3). Protein glycosylation is necessaryfor the correct folding of as many as 70% of the proteins thatexit the ER,55 and a required subunit of the rough ER proteinoligosaccharyl transferase complex (OST1)54 was elevated (1.2FC, Table 4).Proteins that are unable to be refolded are directed to the
ER-associated degradation (ERAD) system18 (Figure 3E).ERAD works in concert with the UPR to reduce ER stressby facilitating ubiquitination of the misfolded proteins, whichare then degraded by the 26S proteosome. The ubiquitin−proteasome system is responsive to oxidative stress and isresponsible for the majority (>70%) of protein degradation ineukaryotes.56−58 Transitional ER ATPase (VCP, 1.0 FC), a
Table 1. 25 Most Abundant Proteins in Aiptasia
% TIC, acclimated treatments % TIC, heat shock treatment annotation UniProtKB accession E value
28.47 15.74 beta actina A0A0A1G3Q1 1.4 × 10−107
2.85 2.83 tubulin beta chain P11833 01.81 1.73 myosin heavy chain, striated muscle P24733 01.62 1.46 golgin subfamily B member 1 Q14789 1.5 × 10−102
1.63 1.33 failed axon connections homologue D3ZAT9 8.5 × 10−40
1.55 1.38 tubulin alpha-1 chaina Q8T6A5 01.41 0.81 zona pellucida domain-containing proteina G8HTB6 6.5 × 10−83
1.17 1.06 spectrin alpha chain, nonerythrocytic 1a P07751 01.02 0.80 filamin-Aa Q8BTM8 00.70 0.88 protein disulfide-isomerase P07237 00.58 0.75 glutamate dehydrogenase A7SE06 00.57 0.62 14-3-3 protein epsilon P62262 4.5 × 10−102
0.58 0.60 calpain-B Q9VT65 00.46 0.83 heat shock cognate 71 kDa protein P19120 00.55 0.45 tropomyosin-1, isoforms 9A/A/Ba Q95VA8 2.8 × 10−64
0.48 0.62 ectin B3EWZ8 2.4 × 10−06
0.47 0.50 spectrin beta chain, nonerythrocytic 1 Q01082 00.45 0.52 hemicentin-1 Q96RW7-2 9.2 × 10−35
0.45 0.25 mucin-2a Q62635 1.6 × 10−168
0.40 0.31 tyrosine decarboxylase, putativea A0A0G3CHC0 1.4 × 10−144
0.37 0.35 arginine kinase O15992 00.30 0.36 clathrin heavy chain 1 P49951 00.36 0.18 golgin subfamily B member 1 isoform 4 Q14789-4 1.9 × 10−139
0.29 0.24 tight junction protein ZO-1a Q07157-2 2.4 × 10−80
0.23 0.43 myosin-10 Q27991 0aProteins that were differentially abundant between (25 °C) and heat shock (33.5 °C) treatments. Bold proteins are cytoskeletal, structural, ormuscle proteins. “Acclimated treatments” include anemones acclimated to 25, 30, and 33.5 °C, while “heat shock” indicates anemones transferredfrom 25 to 33.5 °C for 24 h. TIC = total ion current.
Table 2. Amount of Differentially Abundant (q < 0.1)Proteins between the Thermal Treatments Based on FalseDiscovery Rates
treatment 25 °C 30 °C 33.5 °C 33.5 °C HS
25 °C −30 °C 8 −33.5 °C 9 3 −33.5 °C HSa 104 49 74 −
aHS = heat shock, 24 h.
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
E

protein of the ERAD and ubiquitin systems, binds topolyubiquinated proteins and uses ATP to unfold or remodeltargeted proteins, often facilitating their degradation by theproteasome.59 We also detected a 2.3-FC increase in theproteasome 19S cap protein subunit Rpn1, which serves toshuttle bound ubiquinated proteins into the proteasomecomplex for degradation.60 Together, the changes in theabundance of these proteins in heat-shocked anemones supporta role for ERAD in maintaining proteostasis after heat shock incnidarians.Given the dramatic reductions in cytoskeletal protein
abundance after heat shock, large-scale protein degradationand turnover would demand simultaneous increases inchaperones to assist in replacing the lost proteins. Chaperonins,a subset class of chaperones, are ring-shaped protein complexesthat assist in protein folding by providing isolation andprotection from the cytosolic milieu.61 The cytosolicchaperonin containing TCP-1 (CCT) is a complex composedof two eight-subunit rings62,63 (Figure 3A), and we observed1.0- to 1.6-FC increases in four CCT subunits in response toheat shock (Table 4). Up to 15% of all cytosolic proteins havebeen proposed to pass through CCT during folding andassembly.64 The most important role of CCT is in the foldingof cytoskeletal proteins like actin and tubulin,65 which foldparticularly slowly, on the order of several minutes, and aretherefore susceptible to aggregation.66 The induction of CCTand Hsp’s indicates a need for the resynthesis of compromisedproteins, particularly the highly abundant cytoskeletal compo-nents, after proteome disruption.
Calcium-Binding Proteins
Calcium is an important secondary messenger ion, and changesin Ca2+ concentration induce many CSRs.67 The ERsubstantially envelops mitochondria, and Ca2+ communicationbetween the two occurs through multiple mechanisms,including direct exchange through tight junctions stabilizedby the cytosolic isoform of HspA917,67 (Figure 3F). Ca2+
transfer to the mitochondria stimulates respiratory and TCAcycle activity, eventually inducing apoptosis,17 and the role ofCa2+ homeostasis in cnidarian bleaching has been previouslyexplored.68−70 Our results detail large abundance shifts incytoskeletal proteins, many of which have Ca2+-binding activity,as well as in specific Ca2+ regulators. Calumenin is a Ca2+-binding EF-hand-containing protein that is localized to the ER.It has been demonstrated to be upregulated in response to ERstressors, including oxidative stress, and plays a chaperone-likerole in alleviating ER stress and subsequent apoptosis.10,71
Calumenin was found to be greatly upregulated in heat-shockedAiptasia (4.5 FC). Ganot et al. suggested a role of calumenin inthe cnidarian−dinoflagellate symbiosis, demonstrating multiplecnidarian-specific gene duplications and strong upregulation ofcalumenin in the symbiotic sea anemone Anemonia viridisrelative to aposymbiotic animals.72 Bellantuono et al. foundcalumenin mRNA to decline in response to heat stress.73
Calbindins (Cbp53, −2.5 FC) are a family of proteins thatsense Ca2+ concentration and serve as buffers by sequesteringintracellular Ca2+.74 Given the upregulation of multiple ERstress mechanisms and the role of Ca2+ as a direct messengerbetween the ER and mitochondria, our data are consistent witha role for Ca2+ homeostasis in the control of cnidarian thermalstress.
Oxidative Stress and Apoptosis
Redox and protein homeostasis mechanisms overlap andinteract with one another, and oxidative stress stimulates amajor signaling pathway that may integrate with other stresssignals, such as Hsp’s, through redox control.5,17,75 Commoncellular reactions involved in regulating oxidative stress involvesuperoxide dismutases, catalases, glutathione S-transferases(GSTs), and thioredoxins. The levels of two GSTs increasedin heat-shocked Aiptasia (1.5 and 2.4 FC), as did twothioredoxin-domain containing PDIs, PDIA4, and ERP44 (2.7and 1.0 FC, respectively; Table 4). Notably, ROS are generatedas a result of PDI protein folding activity (Figure 3C), which
Table 3. Differentially Abundant Proteins between Aiptasia Acclimated to Control (25 °C) and High Temperature (30 and 33.5°C) Conditions for 2 Weeks
a“Fold change” represents the log2 ratio of proteins from anemones maintained at 25 °C versus those from anemones subjected to elevatedtemperatures.
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
F

Table 4. Selected Differentially Abundant (q < 0.1) Proteins between Aiptasia Acclimated to Control (25 °C) and Heat-Shocked(33.5 °C, 24 h) Conditions
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
G

catalyzes the formation and disruption of cysteine-cysteinedisulfide bonds in the ER.76 Additionally, several Hsp’s,particularly Hsp60, have been demonstrated to mediate cellstress signaling, activate the innate immune response, andinduce NO production.77 Although caspase activity has beendemonstrated to increase in response to thermal shock inAiptasia,13 and multiple caspases were identified in the Aiptasiaproteome here, none exhibited altered abundance betweentreatments. This may reflect the nature of caspase regulation;caspases are constitutively synthesized and only activated whennecessary to alleviate ROS stress.3 Superoxide dismutase wasalso detected but showed no differential abundance.Mitochondria are a major site of ROS generation as a
consequence of their oxidative phosphorylation activity3
(Figure 3G), and our data suggest changes in response toheat shock that may alter ROS generation. NDUFS1 is thelargest subunit of complex I, the first complex of themitochondrial oxidative phosphorylation electron transportchain,78 and NDUFS1 abundance was 1.5 FC greater in heat-shocked anemones (Table 4). Furthermore, complex I can alsotransfer electrons from NADH/FADH to ubiquinone, and theubiquinone redox state of the host tissue (not the algal cells) ofthe coral Acropora millepora has been demonstrated to behighly responsive to hyperthermal stress.79 Greater NDUFS1abundance following heat shock may support an increased rateof respiratory electron flow and therefore the production ofROS (Figure 3G). The NDUFS1 complex is involved inapoptosis due to its cleavage by “executioner” caspases, such ascaspase-3, which disrupt mitochondrial electron flow andpromote ROS generation, triggering apoptosis.80 Furthermore,mitochondrial 3-mercaptopyruvate sulfurtransferase (3-MST)abundance increased in heat-shocked Aiptasia (1.0 FC). 3-MSTproduces hydrogen sulfide, which in low concentrationsprovides electrons that support mitochondrial electron flowcoupled to aerobic respiration.81 It also enhances glutathione(GSH) production and increases mitochondrial resilience to
oxidative stress.82 The increased abundances of NDUFS1 and3-MST indicate potential increases in both ROS generation andATP production to protect against ROS toxicity.
Metabolic Responses to Thermal Shock
Maintaining homeostasis in response to thermal changerequires shifts in metabolism to accommodate changes in theorganism’s energy requirements (e.g., ATP, NADH) or of theflux of different metabolite classes (e.g., amino acids, lipids),and our study reveals changes in multiple metabolic pathwaysin response to thermal shock. Enzymes that catalyze rate-limiting reactions in central metabolism are likely to be tightlyregulated, and an increase in tricarboxylic acid (TCA) cycleactivity is a highly conserved response to cellular stress.5 Threeenzymes of the TCA cycle were found to increase in heat-shocked Aiptasia: citrate synthase (2.9 FC), isocitratedehydrogenase (IDH, 1.2 FC), and isocitrate lyase (0.9 FC).Citrate synthase is thermally sensitive and stabilized byHsp90,83 while IDH, also a member of the conserved MSP,catalyzes the rate-limiting step of the TCA cycle and is one ofthe few sources of NADPH in animal cells.5 Isocitrate lyase ispart of the glyoxylate cycle, which short-circuits the TCA cycleto enable enhanced utilization of 2-carbon compounds, such asacetyl-CoA from fatty acid catabolism.84 Because antioxidantand macromolecule repair mechanisms require significant ATPand NADPH, the increased ATP availability from elevatedTCA cycle activity and oxidative phosphorylation may supportHsp functions.Increased NADPH production confers resistance to ROS
and NO stress by regenerating reduced antioxidants, and theprimary source of NADPH in animals is the pentose phosphatepathway (Figure 3H), two enzymes of which, transaldolase andtransketolase, increased in abundance after heat shock (0.7 and1.0 FC, respectively).85−87 Furthermore, two oxidoreductasesthat catalyze NADPH regeneration, L-xylulose reductase andhydroxysteroid dehydrogenase-like protein 2, were elevated(2.3 and 1.0 FC, respectively).88,89 Much of the NADPH
Table 4. continued
a“Fold change” represents the log2 ratio of proteins from heat shocked anemones to those of control anemones. bFunctional categories are notexclusive; proteins may appear in multiple categories. Red and blue indicate increased and decreased abundance in heat-shocked anemones,respectively.
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
H

produced by these mechanisms is used to regenerate theantioxidant GSH86 (Figure 3H), and several enzymes of theGSH-producing folate and methionine cycles increased inabundance in response to heat shock. GSH is synthesized viaserine,90 which is generated from glycolysis by phosphogly-cerate dehydrogenase (PHGDH, 2.8 FC) and serine hydrox-ymethyltransferase (SHMT2, 2.0 FC). SHMT2 and PHGDHare also essential for maintaining mitochondrial redox homeo-stasis through the production of NADPH.91 Methionineadenyltransferase (MAT, 2.7 FC) catalyzes the rate-limitingstep of the methionine cycle, producing S-adenosyl methionine(SAMe), the primary methyl donor in eukaryotic cells and anintermediate in GSH biosynthesis. In contrast, betainehydroxymethyltransferase (−4.6 FC) synthesizes the alternativeproduct of the methionine cycle and was reduced followingheat shock. SAMe concentrations are maintained by adenosinekinase,92 which was also increased following heat shock (1.7FC). Phosphoethanolamine N-methyltransferase uses SAMe asa substrate and plays a role in glycerophospholipid synthesis,90
and its increased abundance (3.6 FC) may reflect the need toregenerate host membranes damaged by ROS. Elevation ofthese enzymes suggests increases in NADPH supply forantioxidant mechanisms following heat shock.
Proteome Reorganization of Heat-Shocked Aiptasia
Changes in the protein abundances of heat-shocked Aiptasiasignify the induction of multiple stress-response pathways,particularly those involved in protein repair, degradation, andresynthesis. The concerted upregulation of multiple chaperonesand protein folding/refolding mechanisms demonstrates thatthermal stress is felt acutely at the ER. The marked reduction inthe levels of many otherwise highly abundant cytoskeletalproteins after 24 h of heat shock, combined with thesimultaneous increase in several antioxidant mechanisms (e.g.,GSTs, PDIs) and enzymes that supply them with reductant(ATP and NADPH), indicates that generalized oxidativedamage occurs in response to heat shock. Whether theseROS originated from the algal symbiont or were endogenouslyproduced cannot be definitively determined, but endogenousROS stress would most likely be generated from mitochondrialactivity and the observed changes in Ca2+ homeostasis proteinssupport this view. Excess oxidative damage to mitochondriainduces apoptosis, and while this may have occurred in a subsetof host cells, mitochondrial proteins were either unchanged orupregulated overall, eliminating the possibility of wholesalemitochondrial degradation. We propose that thermal shockresulted in elevated ROS production by the host, causingwidespread protein damage, which was followed by the
Figure 2. Counts of proteins that were differentially abundant (q < 0.1) between control (25 °C) and heat-shocked (33.5 °C, 24 h) Aiptasia,grouped by biological process gene ontology terms. (A) More abundant proteins in heat-shocked Aiptasia (minimum four shown) or (B) lessabundant proteins in heat-shocked anemones (minimum three shown).
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
I

induction of CSRs to repair, degrade, or refold damagedproteins (Figure 3), an interpretation supported by GO termanalysis (Figure 2). The magnitude of the insult, likely taxing tothe ER, required increased levels of multiple chaperones tofacilitate correct protein folding as well as the induction ofERAD and the UPR to degrade misfolded proteins.The large decline in the abundances of actin and other
structural proteins over a relatively short time scale (24 h),combined with the lack of an obvious biological function forsuch a response, is indicative of elevated protein degradation,impaired protein synthesis, or a combination of the two. Wepropose that the dramatic loss of cytoskeletal proteins is a resultof widespread inhibition of protein synthesis and folding due toER stress, enhanced by accelerated protein degradation. The∼50% reduction in cytoskeletal proteins after 24 h of heatshock is in line with a previously reported proteolysis capacityof the 26S proteasome of 3 to 4% degradation of total cellprotein per hour.56 This experiment was not designed to assessprotein turnover per se; therefore, we cannot be definitive inour interpretation of the mechanism of protein degradation.
Reductions in epidermal and gastrodermal layer thickness of∼25% have previously been described in the coral Acroporaaspera after exposure to temperature increases in a smallermagnitude than those imposed here.19 This is in accord withour observation that heat-shocked anemones exhibiteddiminution of four protein toxins (Table 4), presumablylocalized to the cnidocytes of the epidermal layer, which do notharbor algal symbionts and therefore would not be directlyexposed to algal-derived ROS. Hanes and Kempf illustrated theappearance of autophagic structures in Aiptasia tissue after 48 hof heat shock, indicating that in addition to ERAD andproteasome activity autophagy may play a role in restructuringthe host cytoskeleton if the thermal stress is sustained.30
Furthermore, Sawyer and Muscatine demonstrated host-celldetachment during thermal and Ca2+-disruption stress.70
Detachment of whole cells may play a role in the bleachingresponse at the organismal level,93 but because it would have auniversal effect on all cell proteins, it seems unlikely to explainthe cytoskeletal modifications detailed here.
Figure 3. Cellular processes proposed to be induced by thermal shock in Aiptasia. (A) Cytoskeletal proteins are damaged by reactive oxygen species(ROS) exposure (red regions). The damaged proteins are ubiquinated (green circles) and targeted to the proteasome for degradation. The folding ofnew cytoskeleton proteins is stabilized by chaperonins (CCT). (B) Nascent proteins either fold correctly (blue) or are misfolded (red). Misfoldingrates increase under high temperatures, resulting in an accumulation of misfolded proteins and the induction of the unfolded protein response. (C)Misfolded proteins are identified by heat shock proteins (Hsp’s), such as Hsp90, and are either repaired or unfolded for retrotranslocation from theendoplasmic reticulum (ER), which is then followed by ubiquination and subsequent degradation by the proteasome (ER-associated degradation(ERAD)). Protein unfolding by protein disulfide isomerase results in ROS generation. (D) Increases in coatomer (COPI) protein reflect anincreased amount of COPI vesicle cycling between the ER and Golgi apparatus, which increases the influx of damaged or misfolded proteins into theER. (E) Proteins that are unable to be refolded are exported from the ER, ubiquinated, and transported to the proteasome for degradation via ERAD.(F) The secondary messenger Ca2+ is exchanged between the ER and mitochondria through tight junctions stabilized by HspA9. Transfer of Ca2+
from the ER to the mitochondria increases during ER stress, stimulating mitochondrial respiration and potentially inducing apoptosis. (G)Upregulation of both the tricarboxylic acid (TCA) cycle and oxidative phosphorylation increases the reductant available for antioxidant mechanismsas well as the generation by NADH-ubiquinone oxidoreductase (NDUFS1) of ROS, some of which may leak from the mitochondria. (H) Theantioxidant glutathione (GSH) repairs oxidized proteins, and its oxidized form (GSSG) is reduced by NADPH generated by the pentose phosphatepathway (PPP). GSH is synthesized as a result of the methionine/S-adenosyl methionine (SAMe) cycle, enzymes of which are upregulated byincreased temperatures.
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
J

The CSRs and corresponding changes in protein abundancedescribed here are congruent with previous studies in symbioticcnidarians. Larvae of Acropora millepora exhibited upregulationof Hsp90 under short-term (4 h) thermal stress but not underlong-term stress.94 Additionally, an analysis of the A. milleporatranscriptome revealed greater changes in the gene expressionof heat-shocked corals compared with those that had beenallowed to thermally acclimate, but the transcript profiles of thethermally shocked corals showed little overlap with our dataset.73 Proteomic analysis of thermally stressed A. microphthalmaidentified 11 upregulated proteins, including multiple Ca2+-sensing proteins.95 Furthermore, Desalvo et al. detectedchanges in transcripts encoding Ca2+-binding proteins inbleached corals,69 suggesting that disruption of Ca2+ homeo-stasis, potentially resulting from ROS or NO production,induces organelle damage and cytoskeletal rearrangement. Ourdata show that these and other CSR mechanisms are not simplya result of elevated temperature but rather are indicative of arapid disruption of cellular homeostasis prior to algalphotoinhibition.
■ CONCLUSIONSThe independent thermal susceptibility of the cnidarian host,the dinoflagellate symbiont, and different host/symbiontsymbioses is an area of active research. ROS generation isconsidered to be a likely factor in the bleaching response, andthe fact that symbiont abundance and photosynthetic efficiencyremained unchanged in our experiment, even during short-termthermal shock, suggests that ROS or NOS were host-derivedand were generated before algal stress or bleaching wasdetected. Aiptasia exhibited considerable capacity to maintainhomeostasis with little change in protein abundances undergradually elevated, sustained high temperatures. Short-termheat shock, however, disrupted proteostasis and inducedmultiple mechanisms to promote protein folding and todegrade improperly folded proteins, indicating an intenseimpact on ER functions. On the basis of the observations ofreduced cytoskeletal protein abundance and elevated ERprotein folding mechanisms, the host appeared to be in themidst of large-scale protein reorganization to repair damageincurred by the heat shock. The initial mechanism of thermaldisruption cannot be identified from this study, but oxidativestress caused by increased production of ROS or NOS seemslikely. The cnidarian ER is both a generator and mediator ofROS and likely plays a central role in maintaining hosthomeostasis and symbiosis stability during thermal stress.
■ ASSOCIATED CONTENT*S Supporting Information
The Supporting Information is available free of charge on theACS Publications website at DOI: 10.1021/acs.jproteo-me.6b00797.
Figure S1. Grand sums of all total ion current values forall identified proteins per technical replicate. (PDF)Table S1. Database accession and total ion current valuefor each identified protein cluster. (XLSX)Table S2. Protein cluster report including identificationprobabilities, sequence coverage, and numbers of uniquepeptides and spectra. (XLSX)Table S3. Protein clusters that were significantlydifferentially abundant between treatments, including
BLAST statistics, UniprotKB entries, and log2 foldchange values. (XLSX)
■ AUTHOR INFORMATIONCorresponding Author
*Tel: +64 04 4639802. E-mail: [email protected]
Clinton A. Oakley: 0000-0002-4673-0645Author Contributions
The project was conceived by C.A.O. and S.K.D. andconducted with the assistance of E.D. and L.P. Technicalassistance and expertise were provided by L.P. Statisticalanalysis was provided by S.P.W. Analysis and interpretationwere contributed by C.A.O., V.M.W., A.R.G., and S.K.D. Themanuscript was written by C.A.O. with insight, editing, andreview from E.D., L.P., S.P.W., V.M.W., A.R.G., and S.K.D.Funding was acquired by S.K.D., V.M.W., and A.R.G.Notes
The authors declare no competing financial interest.Mass spectrometry data have been deposited to theProteomeXchange Consortium42 via the PRIDE partnerrepository with the data set identifier PXD004257.
■ ACKNOWLEDGMENTSThis project was funded by the Marsden Fund of the RoyalSociety of New Zealand, grant number 1202 to S.K.D., V.M.W.,and A.R.G.
■ REFERENCES(1) Wild, C.; Huettel, M.; Klueter, A.; Kremb, S. G.; Rasheed, M. Y.M.; Jørgensen, B. B. Coral mucus functions as an energy carrier andparticle trap in the reef ecosystem. Nature 2004, 428, 66−70.(2) Davy, S. K.; Allemand, D.; Weis, V. M. Cell biology of cnidarian-dinoflagellate symbiosis. Microbiol. Mol. Biol. Rev. 2012, 76 (2), 229−261.(3) Weis, V. M. Cellular mechanisms of cnidarian bleaching: stresscauses the collapse of symbiosis. J. Exp. Biol. 2008, 211, 3059−3066.(4) Kultz, D. Evolution of the cellular stress proteome: frommonophyletic origin to ubiquitous function. J. Exp. Biol. 2003, 206(18), 3119−3124.(5) Kultz, D. Molecular and evolutionary basis of the cellular stressresponse. Annu. Rev. Physiol. 2005, 67 (1), 225−257.(6) Gattuso, J. P.; Magnan, A.; Bille, R.; Cheung, W. W. L.; Howes, E.L.; Joos, F.; Allemand, D.; Bopp, L.; Cooley, S. R.; Eakin, C. M.; et al.Contrasting futures for ocean and society from different anthropogenicCO2 emissions scenarios. Science (Washington, DC, U. S.) 2015, 349(6243), aac4722−aac4722.(7) Hughes, T. P.; Kerry, J.; Alvarez-Noriega, M.; Alvarez-Romero, J.;Anderson, K.; Baird, A.; Babcock, R.; Beger, M.; Bellwood, D.;Berkelmans, R.; et al. Global warming and recurrent mass bleaching ofcorals. Nature 2017, 543, 373.(8) Lesser, M. P. Oxidative stress causes coral bleaching duringexposure to elevated temperatures. Coral Reefs 1997, 16, 187−192.(9) Lesser, M. P. Oxidative stress in marine environments:Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68,253−278.(10) Dunn, S. R.; Pernice, M.; Green, K.; Hoegh-Guldberg, O.; Dove,S. G. Thermal stress promotes host mitochondrial degradation insymbiotic cnidarians: are the batteries of the reef going to run out?PLoS One 2012, 7 (7), e39024.(11) Tolleter, D.; Seneca, O.; Denofrio, J. C.; Krediet, C. J.; Palumbi,S. R.; Pringle, J. R.; Grossman, A. R. Coral bleaching independent ofphotosynthetic activity. Curr. Biol. 2013, 23, 1782−1786.
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
K

(12) Rehman, A. U.; Szabo, M.; Deak, Z.; Sass, L.; Larkum, A.; Ralph,P.; Vass, I. Symbiodinium sp. cells produce light-induced intra- andextracellular singlet oxygen, which mediates photodamage of thephotosynthetic apparatus and has the potential to interact with theanimal host in coral symbiosis. New Phytol. 2016, 212 (2), 472−484.(13) Hawkins, T. D.; Bradley, B. J.; Davy, S. K. Nitric oxide mediatescoral bleaching through an apoptotic-like cell death pathway: Evidencefrom a model sea anemone-dinoflagellate symbiosis. FASEB J. 2013,27 (12), 4790−4798.(14) Chaudhari, N.; Talwar, P.; Parimisetty, A.; Lefebvred’Hellencourt, C.; Ravanan, P. A molecular web: endoplasmicreticulum stress, inflammation, and oxidative stress. Front. Cell.Neurosci. 2014, 8, 213.(15) Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response:life on the verge of death. Mol. Cell 2010, 40 (2), 253−266.(16) Ron, D.; Walter, P. Signal integration in the endoplasmicreticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8(7), 519−529.(17) Rainbolt, T. K.; Saunders, J. M.; Wiseman, R. L. Stress-responsive regulation of mitochondria through the ER unfoldedprotein response. Trends Endocrinol. Metab. 2014, 25 (10), 528−537.(18) Vembar, S. S.; Brodsky, J. L. One step at a time: endoplasmicreticulum-associated degradation. Nat. Rev. Mol. Cell Biol. 2008, 9(12), 944−957.(19) Ainsworth, T. D.; Hoegh-Guldberg, O.; Heron, S. F.; Skirving,W. J.; Leggat, W. Early cellular changes are indicators of pre-bleachingthermal stress in the coral host. J. Exp. Mar. Biol. Ecol. 2008, 364 (2),63−71.(20) Bieri, T.; Onishi, M.; Xiang, T.; Grossman, A. R.; Pringle, J. R.Relative contributions of various cellular mechanisms to loss of algaeduring cnidarian bleaching. PLoS One 2016, 11 (4), e0152693.(21) Donner, S. D.; Skirving, W. J.; Little, C. M.; Oppenheimer, M.;Hoegh-Guldberg, O. Global assessment of coral bleaching andrequired rates of adaptation under climate change. Glob. Chang. Biol.2005, 11 (12), 2251−2265.(22) Hoegh-Guldberg, O.; Mumby, P. J.; Hooten, A. J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C. D.; Sale, P. F.; Edwards, A. J.;Caldeira, K.; et al. Coral reefs under rapid climate change and oceanacidification. Science (Washington, DC, U. S.) 2007, 318 (5857), 1737−1742.(23) Weis, V. M.; Davy, S. K.; Hoegh-Guldberg, O.; Rodriguez-Lanetty, M.; Pringle, J. R. Cell biology in model systems as the key tounderstanding corals. Trends Ecol. Evol. 2008, 23 (7), 369−376.(24) Thornhill, D. J.; Xiang, Y.; Pettay, D. T.; Zhong, M.; Santos, S.R. Population genetic data of a model symbiotic cnidarian systemreveal remarkable symbiotic specificity and vectored introductionsacross ocean basins. Mol. Ecol. 2013, 22 (17), 4499−4515.(25) Lehnert, E. M.; Burriesci, M. S.; Pringle, J. R. Developing theanemone Aiptasia as a tractable model for cnidarian-dinoflagellatesymbiosis: the transcriptome of aposymbiotic A. pallida. BMCGenomics 2012, 13 (1), 271.(26) Baumgarten, S.; Simakov, O.; Esherick, L. Y.; Liew, Y. J.;Lehnert, E. M.; Michell, C. T.; Li, Y.; Hambleton, E. A.; Guse, A.;Oates, M. E.; et al. The genome of Aiptasia, a sea anemone model forcoral biology. Proc. Natl. Acad. Sci. U. S. A. 2015, 112 (38), 11893−11898.(27) Oakley, C. A.; Ameismeier, M. F.; Peng, L.; Weis, V. M.;Grossman, A. R.; Davy, S. K. Symbiosis induces widespread changes inthe proteome of the model cnidarian Aiptasia. Cell. Microbiol. 2016, 18,1009.(28) Grajales, A.; Rodríguez, E. Morphological revision of the genusAiptasia and the family Aiptasiidae (Cnidaria, Actiniaria, etridioidea).Zootaxa 2014, 3826 (1), 55−100.(29) LaJeunesse, T. C.; Parkinson, J. E.; Reimer, J. D. A genetics-based description of Symbiodinium minutum sp. nov. and S.psygmophilum sp. nov. (dinophyceae), two dinoflagellates symbioticwith cnidaria. J. Phycol. 2012, 48 (6), 1380−1391.
(30) Hanes, S. D.; Kempf, S. C. Host autophagic degradation andassociated symbiont loss in response to heat stress in the symbioticanemone. Aiptasia pallida. Invertebr. Biol. 2013, 132 (2), 95−107.(31) Mellacheruvu, D.; Wright, Z.; Couzens, A.; Lambert, J.; St-Denis, N., St; Li, T.; Miteva, Y. V.; Hauri, S.; Sardiu, M. E.; Low, T. Y.The CRAPome: a contaminant repository for affinity purification massspectrometry data. Nat. Methods 2013, 10 (8), 730−736.(32) Marx, H.; Lemeer, S.; Klaeger, S.; Rattei, T.; Kuster, B. MScDB:A mass spectrometry centric protein sequence database. J. ProteomeRes. 2013, 12 (6), 2386−2398.(33) Eng, J. K.; McCormack, A. L.; Yates, J. R. An approach tocorrelate tandem mass spectral data of peptides with amino acidsequences in a protein database. J. Am. Soc. Mass Spectrom. 1994, 5(11), 976−989.(34) R Development Core Team. R: A Language and Environment forStatistical Computing; R Foundation for Statistical Computing: Vienna,Austria, 2016.(35) Lee, L. NADA: Nondetects and Data Analysis for EnvironmentalData, 2013.(36) Oksanen, J.; Blanchet, F. G.; Kindt, R.; Legendre, P.; Minchin,P. R.; O’Hara, R. B.; Simpson, G. L.; Solymos, P.; Stevens, M. H. H.;Wagner, H. Package “vegan”, 2016.(37) Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate:a practical and powerful approach to multiple testing. J. R. Stat. B1995, 57 (1), 289−300.(38) Nesvizhskii, A. I.; Keller, A.; Kolker, E.; Aebersold, R. Astatistical model for identifying proteins by tandem mass spectrometry.Anal. Chem. 2003, 75 (17), 4646−4658.(39) Kalli, A.; Hess, S. Effect of mass spectrometric parameters onpeptide and protein identification rates for shotgun proteomicexperiments on an LTQ-Orbitrap mass analyzer. Proteomics 2012, 12(1), 21−31.(40) Binns, D.; Dimmer, E.; Huntley, R.; Barrell, D.; O’Donovan, C.;Apweiler, R. QuickGO: a web-based tool for Gene Ontologysearching. Bioinformatics 2009, 25 (22), 3045−3046.(41) Huntley, R. P.; Sawford, T.; Mutowo-Meullenet, P.; Shypitsyna,A.; Bonilla, C.; Martin, M. J.; O’Donovan, C. The GOA database:Gene Ontology annotation updates for 2015. Nucleic Acids Res. 2015,43, 1057−1063.(42) Vizcaíno, J. A.; Deutsch, E. W.; Wang, R.; Csordas, A.; Reisinger,F.; Ríos, D.; Dianes, J. A.; Sun, Z.; Farrah, T.; Bandeira, N.; et al.ProteomeXchange provides globally coordinated proteomics datasubmission and dissemination. Nat. Biotechnol. 2014, 32 (3), 223−226.(43) Warner, M. E.; Suggett, D. J. The Photobiology of Symbiodiniumspp.: Linking Physiological Diversity to the Implications of Stress. InThe Cnidaria, Past, Present and Future: The World of Medusa and HerSisters; Goffredo, S., Dubinsky, Z., Eds.; Springer Publishing, 2016; pp489−509.(44) Hennige, S. J.; Suggett, D. J.; Warner, M. E.; McDougall, K. E.;Smith, D. J. Photobiology of Symbiodinium revisited: bio-physical andbio-optical signatures. Coral Reefs 2009, 28 (1), 179−195.(45) Thornton, S.; Anand, N.; Purcell, D.; Lee, J. Not just forhousekeeping: Protein initiation and elongation factors in cell growthand tumorigenesis. J. Mol. Med. 2003, 81 (9), 536−548.(46) Dalle-Donne, I.; Rossi, R.; Milzani, A.; Di Simplicio, P.;Colombo, R. The actin cytoskeleton response to oxidants: From smallheat shock protein phosphorylation to changes in the redox state ofactin itself. Free Radical Biol. Med. 2001, 31 (12), 1624−1632.(47) Wilson, C.; Terman, J. R.; Gonzalez-Billault, C.; Ahmed, G.Actin filaments - a target for redox regulation. Cytoskeleton 2016, 73(10), 577−595.(48) Mayer, M. P.; Bukau, B. Hsp70 chaperones: Cellular functionsand molecular mechanism. Cell. Mol. Life Sci. 2005, 62 (6), 670−684.(49) McClellan, A. J.; Xia, Y.; Deutschbauer, A. M.; Davis, R. W.;Gerstein, M.; Frydman, J. Diverse cellular functions of the Hsp90molecular chaperone uncovered using systems approaches. Cell 2007,131 (1), 121−135.
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
L

(50) Yogev, O.; Pines, O. Dual targeting of mitochondrial proteins:Mechanism, regulation and function. Biochim. Biophys. Acta, Biomembr.2011, 1808 (3), 1012−1020.(51) Nijtmans, L. G.; de Jong, L.; Artal Sanz, M.; Coates, P. J.;Berden, J. A.; Back, J. W.; Muijsers, A. O.; van der Spek, H.; Grivell, L.A. Prohibitins act as a membrane-bound chaperone for thestabilization of mitochondrial proteins. EMBO J. 2000, 19 (11),2444−2451.(52) Tsai, B.; Rodighiero, C.; Lencer, W. I.; Rapoport, T. A. Proteindisulfide isomerase acts as a redox-dependent chaperone to unfoldcholera toxin. Cell 2001, 104 (6), 937−948.(53) Tsai, B.; Ye, Y.; Rapoport, T. A. Retro-translocation of proteinsfrom the endoplasmic reticulum into the cytosol. Nat. Rev. Mol. CellBiol. 2002, 3 (4), 246−255.(54) Lee, M. C. S.; Miller, E. A.; Goldberg, J.; Orci, L.; Schekman, R.Bi-directional protein transport between the ER and Golgi. Annu. Rev.Cell Dev. Biol. 2004, 20, 87−123.(55) Dempski, R. E.; Imperiali, B. Oligosaccharyl transferase:Gatekeeper to the secretory pathway. Curr. Opin. Chem. Biol. 2002,6 (6), 844−850.(56) Rock, K. L.; Gramm, C.; Rothstein, L.; Clark, K.; Stein, R.; Dick,L.; Hwang, D.; Goldberg, A. L. Inhibitors of the proteasome block thedegradation of most cell proteins and the generation of peptidespresented on MHC class I molecules. Cell 1994, 78 (5), 761−771.(57) Aiken, C. T.; Kaake, R. M.; Wang, X.; Huang, L. Oxidativestress-mediated regulation of proteasome complexes. Mol. Cell.Proteomics 2011, 10 (5), R110.006924.(58) Imai, J.; Maruya, M.; Yashiroda, H.; Yahara, I.; Tanaka, K. Themolecular chaperone Hsp90 plays a role in the assembly andmaintenance of the 26S proteasome. EMBO J. 2003, 22 (14),3557−3567.(59) Meyer, H.; Bug, M.; Bremer, S. Emerging functions of the VCP/p97 AAA-ATPase in the ubiquitin system. Nat. Cell Biol. 2012, 14 (2),117−123.(60) Shi, Y.; Chen, X.; Elsasser, S.; Stocks, B. B.; Tian, G.; Lee, B.-H.;Shi, Y.; Zhang, N.; de Poot, S. A. H.; Tuebing, F.; et al. Rpn1 providesadjacent receptor sites for substrate binding and deubiquitination bythe proteasome. Science (Washington, DC, U. S.) 2016, 351 (6275),aad9421−aad9421.(61) Cuellar, J.; Martín-Benito, J.; Scheres, S. H. W.; Sousa, R.; Moro,F.; Lopez-Vinas, E.; Gomez-Puertas, P.; Muga, A.; Carrascosa, J. L.;Valpuesta, J. M. The structure of CCT-Hsc70 NBD suggests amechanism for Hsp70 delivery of substrates to the chaperonin. Nat.Struct. Mol. Biol. 2008, 15 (8), 858−864.(62) Martín-Benito, J.; Grantham, J.; Boskovic, J.; Brackley, K. I.;Carrascosa, J. L.; Willison, K. R.; Valpuesta, J. M. The inter-ringarrangement of the cytosolic chaperonin CCT. EMBO Rep. 2007, 8(3), 252−257.(63) Noormohammadi, A.; Khodakarami, A.; Gutierrez-Garcia, R.;Lee, H. J.; Koyuncu, S.; Konig, T.; Schindler, C.; Saez, I.; Fatima, A.;Dieterich, C.; et al. Somatic increase of CCT8 mimics proteostasis ofhuman pluripotent stem cells and extends C. elegans lifespan. Nat.Commun. 2016, 7, 13649.(64) Thulasiraman, V.; Yang, C. F.; Frydman, J. In vivo newlytranslated polypeptides are sequestered in a protected foldingenvironment. EMBO J. 1999, 18 (1), 85−95.(65) Leroux, M. R.; Hartl, F. U. Protein folding: Versatility of thecytosolic chaperonin TRiC/CCT. Curr. Biol. 2000, 10 (7), 260−264.(66) Dekker, C.; Stirling, P. C.; McCormack, E. A.; Filmore, H.; Paul,A.; Brost, R. L.; Costanzo, M.; Boone, C.; Leroux, M. R.; Willison, K.R. The interaction network of the chaperonin CCT. EMBO J. 2008, 27(13), 1827−1839.(67) Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of celldeath: the calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4 (7),552−565.(68) Fang, L. S.; Huang, S. P.; Lin, K. L. High temperature inducesthe synthesis of heat-shock proteins and the elevation of intracellularcalcium in the coral Acropora grandis. Coral Reefs 1997, 16 (2), 127−131.
(69) Desalvo, M. K.; Voolstra, C. R.; Sunagawa, S.; Schwarz, J. A.;Stillman, J. H.; Coffroth, M. A.; Szmant, A. M.; Medina, M. Differentialgene expression during thermal stress and bleaching in the Caribbeancoral Montastraea faveolata. Mol. Ecol. 2008, 17 (17), 3952−3971.(70) Sawyer, S. J.; Muscatine, L. Cellular mechanisms underlyingtemperature-induced bleaching in the tropical sea anemone Aiptasiapulchella. J. Exp. Biol. 2001, 204 (Pt 20), 3443−3456.(71) Lee, J. H.; Kwon, E. J.; Kim, D. H. Calumenin has a role in thealleviation of ER stress in neonatal rat cardiomyocytes. Biochem.Biophys. Res. Commun. 2013, 439 (3), 327−332.(72) Ganot, P.; Moya, A.; Magnone, V.; Allemand, D.; Furla, P.;Sabourault, C. Adaptations to endosymbiosis in a cnidarian-dinoflagellate association: differential gene expression and specificgene duplications. PLoS Genet. 2011, 7 (7), e1002187.(73) Bellantuono, A. J.; Granados-Cifuentes, C.; Miller, D. J.; Hoegh-Guldberg, O.; Rodriguez-Lanetty, M. Coral thermal tolerance: tuninggene expression to resist thermal stress. PLoS One 2012, 7 (11),e50685.(74) Hagel, K. R.; Beriont, J.; Tessier, C. R. Drosophila Cbp53Eregulates axon growth at the neuromuscular junction. PLoS One 2015,10 (7), e0132636.(75) Schieber, M.; Chandel, N. S. ROS function in redox signalingand oxidative stress. Curr. Biol. 2014, 24 (10), R453−R462.(76) Tu, B. P.; Weissman, J. S. Oxidative protein folding ineukaryotes: Mechanisms and consequences. J. Cell Biol. 2004, 164 (3),341−346.(77) Colaco, C. A.; Bailey, C. R.; Walker, K. B.; Keeble, J. Heat shockproteins: stimulators of innate and acquired immunity. BioMed Res. Int.2013, 2013, 1.(78) Baradaran, R.; Berrisford, J. M.; Minhas, G. S.; Sazanov, L. A.Crystal structure of the entire respiratory complex I. Nature 2013, 494(7438), 443−448.(79) Lutz, A.; Raina, J. B.; Motti, C. A.; Miller, D. J.; Van Oppen, M.J. H. Host coenzyme Q redox state is an early biomarker of thermalstress in the coral Acropora millepora. PLoS One 2015, 10 (10),e0139290.(80) Ricci, J. E.; Munoz-Pinedo, C.; Fitzgerald, P.; Bailly-Maitre, B.;Perkins, G. A.; Yadava, N.; Scheffler, I. E.; Ellisman, M. H.; Green, D.R. Disruption of mitochondrial function during apoptosis is mediatedby caspase cleavage of the p75 subunit of complex I of the electrontransport chain. Cell 2004, 117 (6), 773−786.(81) Modis, K.; Coletta, C.; Erdelyi, K.; Papapetropoulos, A.; Szabo,C. Intramitochondrial hydrogen sulfide production by 3-mercaptopyr-uvate sulfurtransferase maintains mitochondrial electron flow andsupports cellular bioenergetics. FASEB J. 2013, 27 (2), 601−611.(82) Kimura, Y.; Goto, Y.; Kimura, H. Hydrogen sulfide increasesglutathione production and suppresses oxidative stress in mitochon-dria. Antioxid. Redox Signaling 2010, 12 (1), 1−13.(83) Jakob, U.; Lilie, H.; Meyer, I.; Buchner, J. Transient interactionof Hsp90 with early unfolding intermediates of citrate synthase:Implications for heat shock in vivo. J. Biol. Chem. 1995, 270, 7288−7294.(84) Dunn, M. F.; Ramírez-Trujillo, J. A.; Hernandez-Lucas, I. Majorroles of isocitrate lyase and malate synthase in bacterial and fungalpathogenesis. Microbiology 2009, 155 (10), 3166−3175.(85) Xu, D.; Sun, L.; Liu, S.; Zhang, L.; Yang, H. Understanding theheat shock response in the sea cucumber Apostichopus japonicus, usingiTRAQ-based proteomics. Int. J. Mol. Sci. 2016, 17 (2), 150.(86) Maeng, O.; Kim, Y. C.; Shin, H. J.; Lee, J. O.; Huh, T. L.; Kang,K.; Kim, Y. S.; Paik, S. G.; Lee, H. Cytosolic NADP+-dependentisocitrate dehydrogenase protects macrophages from LPS-inducednitric oxide and reactive oxygen species. Biochem. Biophys. Res.Commun. 2004, 317 (2), 558−564.(87) Mittler, R. Oxidative stress, antioxidants and stress tolerance.Trends Plant Sci. 2002, 7 (9), 405−410.(88) Oppermann, U. Carbonyl reductases: the complex relationshipsof mammalian carbonyl- and quinone-reducing enzymes and their rolein physiology. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 293−322.
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
M

(89) Kowalik, D.; Haller, F.; Adamski, J.; Moeller, G. In search forfunction of two human orphan SDR enzymes: Hydroxysteroiddehydrogenase like 2 (HSDL2) and short-chain dehydrogenase/reductase-orphan (SDR-O). J. Steroid Biochem. Mol. Biol. 2009, 117(4−5), 117−124.(90) Locasale, J. W. Serine, glycine and one-carbon units: cancermetabolism in full circle. Nat. Rev. Cancer 2013, 13 (8), 572−583.(91) Ye, M.; Pan, Y.; Cheng, K.; Zou, H. Protein digestion priority isindependent of protein abundances. Nat. Methods 2014, 11 (3), 220−222.(92) Bjursell, M. K.; Blom, H. J.; Cayuela, J. A.; Engvall, M. L.; Lesko,N.; Balasubramaniam, S.; Brandberg, G.; Halldin, M.; Falkenberg, M.;Jakobs, C.; et al. Adenosine kinase deficiency disrupts the methioninecycle and causes hypermethioninemia, encephalopathy, and abnormalliver function. Am. J. Hum. Genet. 2011, 89 (4), 507−515.(93) Gates, R. D.; Baghdasarian, G.; Muscatine, L. Temperaturestress causes host cell detachment in symbiotic cnidarians: implicationsfor coral bleaching. Biol. Bull. 1992, 182, 324−332.(94) Meyer, E.; Aglyamova, G. V.; Matz, M. V. Profiling geneexpression responses of coral larvae (Acropora millepora) to elevatedtemperature and settlement inducers using a novel RNA-Seqprocedure. Mol. Ecol. 2011, 20 (17), 3599−3616.(95) Weston, A. J.; Dunlap, W. C.; Beltran, V. H.; Starcevic, A.;Hranueli, D.; Ward, M.; Long, P. F. Proteomics links the redox state tocalcium signaling during bleaching of the scleractinian coral Acroporamicrophthalma on exposure to high solar irradiance and thermal stress.Mol. Cell. Proteomics 2015, 14 (3), 585−595.
Journal of Proteome Research Article
DOI: 10.1021/acs.jproteome.6b00797J. Proteome Res. XXXX, XXX, XXX−XXX
N