thérapie cellulaire de l'insuffisance cardiaque dans...
TRANSCRIPT

UNIVERSITE DE GENEVE Faculté de MédecineSection de Médecine CliniqueDépartement de GériatrieLaboratoire de Biologie du Vieillissement
Thèse préparée sous la direction du Prof. Karl-Heinz Krause
Travail effectué sous la supervision du Dr. Marisa Jaconi
THERAPIE CELLULAIRE DEL'INSUFFISANCE CARDIAQUE
DANS UN MODELE D'INFARCTUS DUMYOCARDE CHEZ LE RAT
THÈSE
présentée à la Faculté de Médecinede l´Université de Genève
pour obtenir le grade de Docteuren médecine
par
Qing HE
De Shangai, Chine
Thèse n° 10358
Genève, 2004

2
To my husband To my husband Ke-zhengKe-zhengand my daughter and my daughter Yun-YunYun-Yunwith all my lovewith all my love

3
TABLE OF CONTENTS
A. List of abbreviations 4
Résumé et introduction (en français) i-ivB. Summary 5
C. Introduction 71. Seeking stem cells desperately 72. Embryonic stem cells (ESC) 83. No stem cells in the heart ? 114. Cell therapy of the heart 11
4.1 ESC into the heart 124.2. Human ESC 134.3. Skeletal muscle cells into the heart 144.4. Bone marrow stem cells 15
D. Material and methods 251. ESC culture and differentiation 252. Isolation of ESC-derived and neonatal mouse cardiomyocytes 273. Plasmid constructions, stable transfections and isolation of ESC clones 274. Rat model of myocardial infarction 285. Experimental animal protocol 306. Transplantation procedure 317. Immunosuppression treatment 328. Evaluation of the left ventricular function by echocardiography 339. Histological and immunohistochemistry studies 33
9.1. Antibodies 349.2. Immunohistochemistry 34
10. Statistical analysis 35
E. Results 361. Grafting of undifferentiated ESC in normal (non-infarcted) heart 36
2. Rat model of myocardial infarction (MI) 41
3. Grafting of undifferentiated ESC in acute phase of MI (1 week) 43
4. Grafting of undifferentiated ESC in chronic phase of MI (4 weeks) 45
5. Grafting of ESC-derived CMC in MI rats 496. Effect of time and immunosuppression on teratoma formation 50
7. The evaluation of left ventricular function by echocardiogram 51
F. Discussion 54
G. Conclusions 58
H. Implementation and future experiments 59
I. Reprints 60
J. Acknowledgments 61
K. References 62

4
A. LIST OF ABBREVIATIONS
BSA Bovine serum albumin
CCM Cellular cardiomyoplasty
CHF Congestive heart failure
CGR8 Center Genome Research 8
CsA Cyclosporin A
Cx43 Connexin 43, gap-junction protein
EB Embryoid Body
EBFP Enhanced Blue Fluorescent Protein
EG Embryonic germ
EGFP Enhanced Cyan Fluorescent Protein
ESC Embryonic stem cells
FACS Fluorescence –assisted cell sorting
FS Fractional Shorting
GFP Green Fluorescent Protein
H&E Hematoxylin and eosin
LIF Leukemia Inhibitory Factor
LVEF Left Ventricular Ejection Fraction
LVEDD Left Ventricular End-Diastolic Diameter
LVESD Left Ventricular End-Systolic Diameter
MEF Murine Embryonic Fibroblast
MHC Myosin Heavy Chain
MI Myocardial infarction
MSC Mesenchymal stem cells
NIH National Institute of Health
PCNA Proliferating Cell Nuclear Antigen
TGF-ββ Transforming Growth Factor β-1

5
B. SUMMARY
Congestive heart failure (CHF) is a major cause of cardiovascular morbidity and mortality in
developed countries. Treatment consists basically of long-standing symptomatic drug therapy,
effective percutaneous and surgical revascularization. The only causal treatment at the end-stage
of CHF is heart transplantation. However, the shortage of donor hearts the complications of
immunosuppression, the failure of grafted organs and, not at least, the advanced age of patients
suffering from CHF limit the utility of cardiac transplantation significantly. In recent years the
idea emerged that cells rather than whole organs can be used to replace damaged tissues.
Two different cell types displaying these qualities have been identified so far: 1) embryonic
stem cells (ESC), and 2) adult somatic stem cells. ESC are pluripotent cells derived from the
inner cell mass of the blastocyst. They have the ability to differentiate into virtually all kinds of
cell types, a capacity that becomes progressively restricted with development.
Can ESC be used as the source of cells for the therapy of CHF? Could a failing heart instruct
the engrafted cells to become cardiac cells in situ, or is it first necessary to differentiate them in
culture into cardiomyocytes?
To address these fundamental questions, our research focused on the intracardiac
implantation of undifferentiated murine ESC into a rat model of myocardial infarction caused by
coronary artery ligature. We studied the possibility of cell survival and differentiation in situ. In
particular we address the necessity of using immunosuppression when implanting non-
autologous cells. Specifically, we investigated:
1) The influence of cell implantation as a function of time after MI (acute (1 week) versus
chronic phase (4 weeks));
2) Different period after cell implantation (1 versus 4 weeks of engraftment);
3) Different types of ESC, feeder cell dependent and LIF (Leukemia Inhibitory Factor)
dependent;
4) The effect of immunosuppression treatment using CsA (Cyclosporin A) on cell fate.
The results show that under the circumstance of immunosuppression by CsA, the implantation
of different types of undifferentiated ESC inevitably leads to teratoma formation. The occurrence
increased over time from 20% at 1 week to 100% 4 weeks after implantation. The teratoma
formation was not influenced by cardiac injury (MI). In the absence of immunosuppression
(CsA), the injected ESC could not be retrieved. We conclude that it is necessary to use
immunosuppression when grafting undifferentiated murine ESC to avoid cell rejection. Even

6
though the use ESC as a source of cell therapy for CHF remain a promising idea, there is still a
long way to go from basic research to the clinic application. Careful attention must be paid to
numerous aspects of stem cell therapy to avoid deleterious effects.

7
C. INTRODUCTION
Chronic congestive heart failure represents a major cause of cardiovascular morbidity and
mortality in developed countries. It is caused by the loss of functional heart muscle, which is due
either to ischemic heart disease or the presence of dysfunctional muscle resulting from a variety
of causes, including hypertension, viruses, and idiopathic factors. Following myocardial
infarction for example, functional contracting cardiomyocytes are replaced with nonfunctional
scar tissue. This ventricular remodeling leads to ventricle dilatation and progressive heart failure
which constitute a major clinical problem (Grounds et al., 2002). The remodeling process is
characterized by the removal of necrotic cardiomyocytes accompanied by granulation tissue
formation with the simultaneous induction of neovascularization in the peri-infarcted bed. The
latter is a prerequisite for the survival of surrounding hypertrophied but viable cardiomyocytes
and the prevention of further cardiomyocyte loss by apoptosis.
Several treatments for coronary artery disease are available, which reduce the symptoms and
improve the quality of life of CHF patients. They include medical treatments (anticoagulants, β-
blocker, angiotensin-converting-enzyme inhibitors, etc), effective percutaneous and surgical
revascularization and cardiac pacing systems. Cardiac transplantation remains, however, the
ultimate solution for end-stage heart failure. However, the shortage of donor hearts, the
complications of immunosuppression, the failure of grafted organs and, not at last, the advanced
age of patients suffering from CHF limit the utility of cardiac transplantation significantly.
Cell therapy as a mean to repair damaged tissues unable to heal is an increasingly attractive
concept in modern transplantation medicine. For many clinical situations, i.e. congestive heart
failure, Parkinson's disease, diabetes, traumatic injuries (spinal cord) and iatrogenic destruction
of the cell (chemotherapy), replacement of lost cells would be the ideal treatment. In many cases,
however, the development of cell treatment approaches is hampered by an increasing lack of
donors or by the lack of cells suitable for transplantation.
C. 1. Seeking stem cells desperately
Which cells could be used to repair a failing heart and restore its contractile power?
Two major types of stem cells would eventually fulfill suitable characteristics: 1) embryonic
stem cells (ESC), derived from the inner cell mass of the blastocyst, and 2) adult somatic stem
cells isolated form several organs (bone marrow, skeletal muscle, brain, etc).

8
What is a stem cell? Stem cells are commonly defined as undifferentiated cells. They have the
ability to differentiate into virtually all kinds of cell types, a capacity that becomes progressively
restricted with development. As shown in Figure I, they have two important characteristics that
distinguish them from other types of cells. First, as unspecialized cells, they can proliferate and
renew themselves for long periods through cell division. The second is that under certain
physiologic or experimental conditions, they can be induced to become cells with special
functions. As the matter of fact, they provide a theoretically inexhaustible supply of cells that,
depending on type can give rise to some or all body tissues.
Figure I. Definition of a stem cell.
Self renewal Commitment Differentiation
Stem cells are typically found in the embryo and fetus. In the adult body, they have been
identified in various tissue niches, including bone marrow, brain, liver, and skin, as well as in the
circulation. They have been termed “adult stem cells”. An extremely attractive concept is that
adult stem cells could be harvested from a patient, induced to specialize in culture, then
incorporated into a tissue construct, and put back into the same individual when repair become
necessary, bypassing the need for immunosuppression.
The experimental donor cells for cellular cardiomyoplasty (CCM), as an alternative
treatment of cardiovascular disease, include skeletal myoblasts, bone marrow-derived
Mesenchymal stem cells, purified (enriched) haematopoietic stem cell populations and blood and
bone marrow-derived endothelial progenitor cells.
C. 2. Embryonic stem cells (ESC)
Pluripotent murine stem cells are derived from two main embryonic sources: ESC from the
blastocyst and EG cells from the gonadal ridge of the embryo after gastrulation (Figure II). The
successful derivation of murine ESC from the inner cell mass of mouse blastocytes was achieved
in 1981 (Martin, 1981), while embryonic germ (EG) cells have been isolated and cultured from
primordial germ cells (Stewart et al., 1994).

9
Figure II. Origin and establishmentof pluripotent embryonic stem (ES)and embryonic germ (EG) cell linesfrom the inner cell mass (ICM) ofmouse blastocysts and fromprimordial germ cells, respectively.
These cells were shown to be pluripotent, i.e., capable of forming all mature cell phenotypes
derived from the three embryonic layers: endoderm, ectoderm, and mesoderm, as shown in
Figure III.
Figure III. Tissue derivative of the three embryonic layers.
ICM
ES cells
Primordialgerm cells
EG cells
Pluripotent Stem Cell Lines
3.5d Blastocyst 8.5d Embryo
Endoderm
Yolk sac
Primitive gut
Liver Pancreas Digestive tubes
Mesoderm
Lung
Axial Paraaxial Intermediate Lateral Head
Notochord
Sclerotomes
Myotomes
Dermatomes
AxialSkeleton
SkeletalMuscles
Connectivetissue of skin
Mullerianducts
OviductsUterus
Mesonephros
KidneyOvaryTestis
Somatic
mesoderm
Visceralmesoder
m
Connectivetissue of bodywall & limbs
MesenteriesHeartBlood vessels
Ectoderm
Epidermis
Epidermalplacodes
Neural tubes
Neuralcrest
SkinHairMammaryglandsLensinner ear
BrainSpinal cord
PNS
BlastocystZygote

10
In vitro, murine ESC remain undifferentiated when grown in the presence of leukemia
inhibitory factor (LIF) and for some cell lines, cultured on murine embryonic fibroblast (MEF)
as feeder cells. The mechanism to maintain stem cell as a undifferentiated state is linked to the
LIF/Stat3 signaling pathway and the transcription factor Oct-3/4, but it is still unclear how these
components works together (Niwa, 2001). When LIF or feeder cells are withdrawn, most types
of ESC differentiate spontaneously to form aggregates called embryoid bodies (EBs). Embryoid
bodies are comprised of the heterogeneous cells that derived from all three germ layers. These
tri-dimensional cell-cell contacts allow the formation of heterogeneous cultures of differentiated
cell types including cardiomyocytes, hematopoietic cells, endothelial cells, neurons, skeletal
muscle chondrocytes, adipocytes, liver, and pancreatic islets.
Figure IV. The derived ES or EG cells are differentiated in vitro by culturing them via embryo-likeaggregates, the embryoid bodies (EBs). After plating of EBs onto adhesive substrates, differentiated cellsgrow out from the EBs (from Wobus, 2001).
When murine ESC were cultured into embryoid bodies, some differentiated into a diverse
range of cardiomyocyte phenotypes, including ventricular, atrial, sinus nodal, Purkinje, and
pacemaker-like cells. These different cardiac phenotypes exhibit developmentally controlled
expression of cardiac-specific genes, structural proteins, ion channels, and receptors, and can be
distinguished on the basis of their action potentials. A few cell areas on the embryoid bodies
display spontaneous contractile activity under light microscope, and are identifiable as ESC-
derived cardiomyocytes. Since ESC are pluripotent, considering their use raises the potential risk
of teratoma formation. Thus, the purification of differentiated ESC-derived cardiomyocytes from
cultures is a key issue. Unfortunately, so far, none of the approaches used on murine ESC can
give 100% yield of cells with the required phenotype.

11
C 3. No stem cells in the heart?
In the case of the heart, no local stem cells have been so far identified. Cardiomyocytes
undergo terminal differentiation soon after birth and are generally considered to irreversibly
withdrawn from the cell cycle. Cardiomyocyte DNA synthesis occurs primarily in uteri, with
proliferating cells decreasing from 33% at mid-gestation to 2% at birth (MacLellan and
Schneider, 2000). Therefore, upon injury, the adult heart results incapable to regenerate the
damaged tissue which instead become fibrotic.
On note is the fact that the group of Piero Anversa was the only one to report that, in
humans, some ventricular cardiomyocytes may have the capacity to proliferate or at least to
undergo nuclear replication in response to ischemic injury. The dividing myocytes have been
identified on the basis of immunohistochemical staining of proliferating nuclear structures such
as Ki67 and cell surface expression of specific surface markers CD117. (Itescu et al., 2003).
However, this is still a matter of debate and convincing proofs of cell division as a general event
are still pending. It remains to be determined where does the dividing cells which homed to the
damaged myocardium come from? Are those a resident source of cardiac stem cells, or do they
come from a renewable source of circulating bone marrow-derived stem cells? It is not very clear
so far (Anversa et al., 2003; Beltrami et al., 2001).
C 4. Cell therapy of the heart
The cell-based myocardial repair technology “cellular cardiomyoplasty” (CCM), attempt to
regenerate functioning muscle in previously infarcted, scarred or dysfunctional myocardial tissue
after transplantation of myogenic cells. The use of such a cell therapy approach to replace lost
cardiomyocytes with new engraftable ones would represent an invaluable, low-invasiveness
technique for treatment of heart failure as an alternative to whole heart transplantation.
Replacement and regeneration of functional cardiac muscle after ischemia could be achieved
either by stimulating proliferation of endogenous mature cardiomyocytes or by implanting
exogenous donor-derived or allogeneic cardiomyocytes. The newly formed cardiomyocytes must
integrate precisely into the existing myocardial wall to augment contractile function of the
residual myocardium in a synchronized manner and avoid alteration in the electrical condition
and syncytial contraction of the heart (Itescu et al., 2003).
To date, many types of cells have been tested as a source of cell therapy for the
augmentation of myocardial performance in different experimental models of heart failure.
Those include fetal cardiomyocytes (Leor et al., 1996; Li et al., 1996; Reinecke et al., 1999),

12
ESC-derived cardiomyocytes (Min et al., 2002), skeletal myoblasts (Murry et al., 1996; Taylor
et al., 1998), immortalized myoblasts (Robinson et al., 1996), fibroblasts (Hutcheson et al.,
2000), smooth muscle cells (Li et al., 1999), fibroblasts (Murry et al., 1996), adult cardiac-
derived cells (Li et al., 2000), and bone marrow-derived stem cells (Orlic et al., 2001). Different
heart models have been used to study the effect of cell engraftment in the heart, e.g. normal heart
tissue, cryoinjuried heart tissue, ischemic heart, infarcted scar tissue, dilated cardiomyopathy
heart.
C 4.1. ESC into the heart
Several groups have demonstrated the in vivo feasibility of the intra-cardiac implantation
of ESC. Klug et al. were the first to engraft genetically-modified and differentiated
murine ESC-derived cardiomyocytes into the left ventricular free wall of mdx mice (Klug
et al., 1996). Pure cardiomyocyte cultures were obtained by stable transfecting ESC with
a transgene comprised of the α-cardiac myosin heavy chain (MHC) promoter driving a
neomycin resistance gene. This construct being expressed only in cardiac cells, in the
presence of neomycin (G418) it allowed the only survival of ESC-derived
cardiomyocytes. The successful engraftment of donor ESC was confirmed by
immunopositivity for dystrophin, and engrafted cells were found to be aligned with the
host cardiomyocytes. More recently, Min et al. implanted cardiomyocytes derived from
the D3-ESC line into a rat model of ischemic heart. Cardiomyocytes were selected from
embryoid bodies by dissecting the spontaneously beating clusters via a sterile
micropipette. After transfection with a green fluorescent protein (GFP) marker to identify
survival of engrafted ESC, transplantation was performed within 30 minutes after
induction of MI, created by ligation of the left anterior descending coronary artery. Under
these conditions, the cardiac function was significantly improved 6 weeks after cell
transplantation in MI animals compared with the MI control group (Min et al., 2002).
However, some areas with undifferentiated cells were still observed, indicating that their
extraction from the EBs was poor. Recently, Behfar et al reported that undifferentiated
ESC directly implanted into an infarcted heart seems able to commit to the cardiac
phenotype. Their study suggested that ESC differentiation require a paracrine pathway in
the heart. In vitro, pretreatment of embryonic stem cells with TGFβ growth factor
members results in embryoid bodies with greater areas of cardiac differentiation
(increased beating areas) and normal sarcomeric organization (as revealed by
immunostaining). 5 weeks after the In vivo transplantation of undifferentiated ESC
carrying the ECFP (cyan fluorescent protein) marker under the control of the cardiac α-

13
actin promoter, they identified ESC-derived blue cardiomyocytes by immunostaining.
The left ventricular ejection fraction (measured by echocardiography) was improved in a
small number animals (n=3), as compared with control group (Behfar et al., 2002). More
recently, Johkura et al. transplanted ESC-derived cardiomyocytes into the
retroperitoneum of the adult nude mice. These myocytes proliferated, differentiated and
remained viable and contractile for up to 30 days in the ectopic site around large blood
vessels. However, contamination of the donor cells with the residual ESC committed to
other lineages was likely to occur, even if the embryoid body outgrowths were
transplanted after beating cardiomyocytes had appeared. This leaded to the formation of
teratoma in the host retroperitoneum (Johkura et al., 2003).
In summary, the main issues which limit the research and the use of ESC for CCM
include difficulties in obtaining pure and sufficient number of ES-derived
cardiomyocytes, especially of ventricular- like cardiomyocytes, high risk of teratoma
formation and immune rejection.
C 4.2. Human ESC
Presently, several human ESC lines are available since their first isolation by Thomson
in 1998 (Thomson et al., 1998). A registry of them is published by the NIH web site
(http://stemcells.nih.gov/registry/). The usage of human ESC as a resource for cell
therapy is presently an intensive field of research (Mummery et al., 2003; Nir et al.,
2003). At a basic research level, many biological differences should exist between the
murine and human ESC and further fundamental studies are required before to investigate
the suitability and feasibility of using human ES-derived cardiomyocytes for cell
transplantation in humans. From a legal and ethical point of view, research involving
human embryonic cells is highly controversial and many countries are reviewing their
legislation. Importantly, main ethical issues are raised concerning the derivation of
human ESC from human in vitro fertilized embryos, the moral status of the embryo, and
the acceptability of using such derived cells for therapeutic purposes. The application of
human ESC therapy for the treatment of cardiac diseases in humans is far from being a
reality.
C 4. 3. Skeletal muscle cells into the heart
The growth and repair of skeletal muscle is usually initiated by the activation of a
population of muscle precursors, called satellite cells. Satellite cells normally lie near the
basal lamina of the skeletal muscle fibers and can differentiated into myofibers.

14
Following tissue injury, they are rapidly mobilized, proliferate and fuse, thereby effecting
repair and regeneration of the damaged fibers.
Cardiac and skeletal muscles have a similar sophisticated organization of contractile
proteins into sarcomeres and are collectively referred to as striated muscles. Growth of
the heart is generally characterized by division of muscle cells during the embryonic
stages of life, followed after birth by into a post-mitotic state. The postnatal capacity for
cell replication during growth and regeneration of cardiac and skeletal muscle cells is
markedly different. Skeletal muscle cells are multinucleated and can readily regenerate
from precursor cells (satellite cells / myoblasts), whereas post-natal cardiac muscle is
incapable of tissue repair, since cardiac cells exit the cell cycle soon after birth and
become post-mitotic (either in vivo or in vitro).
Autologous skeletal myoblasts appear to be the most well studied and best first
generation cells for cardiac repair. This process was pioneered in the 1980s (Sola et al.,
1985), and has been applied clinically with varied success. The whole procedure includes
extraction of the myoblasts from skeletal muscle, expansion in tissue culture, and
injection into the heart muscle. For the success of cell transplantation, the introduced
donor cells must be able to survive in their host environment. Intramuscular injection of
cultured isolated myoblasts in classical myoblast transfer therapy shows that there is a
massive and rapid necrosis of donor myoblasts, with 90% dead within the first hour after
injection (Beauchamp et al., 1999). Host natural killer cells appear to play a particularly
important role in this rapid death of cultured donor myoblasts.
In vivo, in various species, such as rabbits and rats, engraftment of skeletal myoblasts
were shown colonized injured cardiac tissue (Murry et al., 1996; Pouzet et al., 2000;
Taylor et al., 1998). A survival up to 12 weeks after transplantation was observed in
normal heart (Reinecke et al., 2002), and up to 18 weeks in cryoinjured myocardium
(Reffelmann and Kloner, 2003). An important factor is that skeletal myoblasts are
relatively resistant to ischemia and, in contrast to cardiomyocytes (witch injure rapidly
within 20 min), they can withstand several hours of severe ischemia without becoming
irreversibly injured. Several studies reported improvement of left ventricular performance
after myoblast transplantation, e.g. reducing left ventricular dilatation, increasing ex vivo
systolic pressures and improving in vivo exercise capacity (Atkins et al., 1999; Jain et al.,
2001).
However, the outcome of engraftment of skeletal myoblasts still remains highly
controversial. Some reports claim that transplanted skeletal myoblasts could differentiate
and develop into striated cells within the damaged myocardium, thus preventing the

15
progressive ventricular dilatation by improving heart function (Taylor, 2001). On the
other hand, some investigators have reported negative results and adverse effects. The
systematic investigations by Reinecke et al. did not support the concept of a true
transdifferentiation into cardiomyocytes as demonstrated by the lack of α-myosin-heavy-
chain, cardiac troponin I, or atrial natriuretic peptide- expression in the grafts. Murry et
al. also concluded that skeletal satellite muscle cells differentiate into mature skeletal
muscle and do not express cardiac-specific genes after grafting into the heart, lacking any
transdifferentiation potential (Reinecke et al., 2002).
Recently, attempts to use autologous skeletal satellite cells to repair damaged hearts
have received considerable attention for the possibility of autotransplantation into human
myocardium. The autologous origin would clearly overcomes all problems related to
availability, ethics and immunogenicity, key factors for large-scale clinical applicability
(Menasche, 2003). Menasche et al. first reported a three-steps protocol in a CHF patient
in 2001 (Hagege et al., 2003; Menasche et al., 2003). A muscular biopsy was retrieved
from the thigh under local anesthesia and, after enzymatic and mechanical dissociation,
the cells were grown for 2-3 weeks in culture, providing at least 400x106cells, 50% of
which were myoblasts. These cells are then reimplanted across the post-infarcted scar at
the time of coronary bypass graft surgery (Menasche, 2003).
Several clinical trials are presently running in several countries, with different
preliminary results. Some trials reported the presence of arrhythmia and sudden death,
which leading to the suggestion to implant AICD´s on the time of procedure. The need of
randomized trials is claimed in many conferences and meetings.
There are many important questions remain to be answered for the clinical use of
myoblasts. They are related to cell survival, integration, differentiation, and functional
effect. E.g. for the long-term benefit of skeletal myoblasts transplantation myocardial
injury, cells must be able to survive for years in the heart. The long-term fate of
myoblasts is unclear. The effect of transplanted myoblasts on electrical stability of the
heart, as they do not form normal electrical junctions with the host, it will increase the
risk of arrhythmia. In a phase I human clinical trial, 4 out of 10 cases showed sustained
ventricular tachycardia during the early (3 weeks) postoperative period (Menasche et al.,
2003). The mechanisms of these arrhythmias might be inhomogeneity in action potential
conduction creating reentry pathways. So, how myoblasts electrically integrate into the
surrounding myocardium is still a big issue from the clinical point of view. Can
myoblasts function as cardiac-like myocytes and thus improve contraction, including
adaptation to chronic workload and integration into the host, or can myoblasts just

16
prevent further deterioration of the injured heart? How do we control continuing
proliferation after transplantation to avoid undesirable disturbance of local left ventricular
geometry? All those questions still need to be answered.
C 4. 4. Bone Marrow stem cells
Bone marrow is a mesodermal-derived tissue and contains haematopoietic stem cells
(HSCs) and mesenchymal stem cells (MSCs), which may both, be derived from a
common primitive blast-like cell. The HSC are archetypal stem cells. They have the
ability to balance self-renewal against differentiation cell fate decisions They are
multipotent, a single stem cell producing at least eight to ten distinct lineages of mature
cells. HSC have an extensive proliferative capacity that yields a large number of mature
progeny. These cells are rare, comprising only 1/10'000 to 100'000 of total blood cells,
and can be obtained from the bone marrow, peripheral blood umbilical cord and fetal
liver (Bonnet, 2002; Preston et al., 2003). Krause et al., demonstrated that a single HSC
was not only able to repopulate the haematopoietic system in irradiated mice, but also
differentiated into lung epithelium, skin, liver and the gastrointestinal tract (Krause et al.,
2001).
Figure V illustrates the transdifferentiation potential observed either in culture or after
in vivo injection of cells.
Figure V. Possible pathways of differentiation in adult stem cells. (from Science (Holden andVogel, 2002)
Too good to be true? Studies in mice have yielded evidence, now being
reassessed, that stem cells from the bone marrow compartment can produce progeny in

17
different organs. Bone marrow, which has several types of stem cells, seems particularly
versatile.
MSCs have the main capacity to differentiate, both in vivo and in vitro, into osteoblast,
chondroblasts, and adipocytes when exposed to the appropriate stimuli. Approximately
30% of human marrow aspirate cells adhering to plastic are considered to be MSCs. They
are in general more difficult to characterized than the HSC populations. MSCs have been
functionally identified in adult murine and human bone marrow by their ability to
differentiate to lineages of diverse mesenchymal tissues. Those include bone, cartilage,
fat, tendon, and both skeletal and cardiac muscle, which express specific surface markers
but lack of haematopoietic lineage markers such as CD34 or CD 45. Adult mouse MSCs
in culture were reported to generate spontaneously beating cardiomyocytes (Makino et
al., 1999). When MSCs are pre-exposed to 5-azacytidine they are capable of
differentiating into skeletal and cardiac muscle phenotypes (Makino et al., 1999).
However the reproducibility of this treatment is highly questioned.
Recently, a number of studies have shown that bone marrow stem cells transplantation
is beneficial for myocardial repair/regeneration in different animals and human being.
Kamihata et al. injected autologous bone marrow-derived mononuclear cells into
myocardial infarcted zone of swine (Kamihata et al., 2001). Three weeks later, regional
blood flow and capillary densities were significantly higher, and cardiac function was
improved. They concluded that bone marrow implantation may achieve optimal
therapeutic angiogenesis through potent angiogenic ligands and cytokines secreted by
those cells incorporated into foci of neovascularization. Orlic et al. reported that the
injection into female mice of a subgroup of Lin ---c-kit+ bone marrow cells from EGFP
transgenic male donors at the borderline of an ischemic area resulted in the colonization
of the infarcted area. Within 9 days, male EGFP-positive cells had proliferated in situ and
expressed protein characteristic of cardiac tissue, including connexin43, thus suggesting
intercellular communication (Orlic et al., 2001).
In human, several trials are ongoing. Perin and Dohmann recently reported that
transendocardial injections of autologous mononuclear bone marrow cells in patients with
end-stage ischemic heart disease could safely promote neovascularization and improve
perfusion and myocardial contractility. 21 patients, all of them with previous myocardial
infarction and documented with multivessel disease, were enrolled sequentially, with the
first 14 patients assigned to the treatment group and the last 7 patients to the control
group. Approximately 4 hours before the cell injection procedure, bone marrow (50ml)
was aspirated; mononuclear cells were isolated by density gradient. The

18
electromechanical map was used to identify viable myocardium for treatment; the cells
were injected by NOGA catheter in the treatment group. Both of group patients
underwent 2-month noninvasive follow-up, and treated patients along underwent a 4-
month invasive follow-up. At 2-months, there was a significant reduction in total
reversible defect and improvement in global left ventricular function within the two
groups on quantitative SPECT analysis. At 4 months, the ejection fraction improved from
a baseline of 20% to 29% (p=0.003), concomitantly to a reduction in end-systolic volume
(p=0.03) in the treated patients. The limitation of the study includes the small number of
patients, the short period of follow-up, and no placebo as control (Perin et al., 2003). Tse
et al demonstrated somewhat similar results (Tse et al., 2003). They studied percutaneous
delivery (via the Biosense Electromechanical NOGA mapping catheter) of autologous
bone-marrow-derived mononuclear cells in eight patients with stable angina. After 3
months of follow-up, there was improvement in myocardial perfusion, regional
myocardial wall motion and thickening, but LVEF remained unchanged (Tse et al.,
2003). Hamano and Stamm reported two studies with 5 and 6 patients study respectively
(Hamano et al., 2001; Stamm et al., 2003), in which both of groups received autologous
bone marrow cells at the time of coronary artery bypass grafting, and were followed up
for 3 to 12 months. Their results showed an improvement of perfusion of the infarcted
myocardium possibly due to neoangiogenesis.
Presently, bone marrow stem cells provide an interesting and promising option to
restore myocardial viability. The transplantation in human study appears feasible,
relatively safe and effective, no tumor formation has been scored. However, a major
criticism to these preliminary trials is the fact that the number of patients is too small to
derive a meaningful efficacy and definitive safety data. Also, no data are available about
cell survival following intra-myocardial needle injection, or whether implanted cells did
survive and differentiate along the cardiac myocyte and/or endothelial lineage. In
addition, some studies in big animals reported that injected cells are very low in number
and tend to disappear with time. Further studies are required to better characterize the
phenotype and the fate of injected cells.
Moreover, the in vitro proliferation potential of MSCs is not yet reproducible in order to
obtain a suitable number of differentiated and characterized cells. Clearly, cell
duplication and reproduction is the first condition to fully recolonize a diseased
myocardium and thus improve ventricular function. Another factor is if undifferentiated
stem cells are used, a risk is still present of developing other type of tissue (Laham and
Oettgen, 2003; Scorsin and Souza, 2001).

19
The following three tables (Table I, II, III) are taken from a review by L. Field
(Dowell et al., 2003) and represent to date an extensive list of all the cell therapy studies
published so far.
Table I. Cardiomyocyte transplantation
Donor cell: F, fetal cardiomyocyte; N, neonatal cardiomyocyte; A, adult cardiomyocyte; C, atrial tumor cell line.Species, donor species/host species: M, mouse; R, rat; H, human; D, dog; P, pig; HM, hamster; C, cell line; °,autologous transplant.Tracking: D, dye; H, histology; I, immunostain; T, transgenic; CH, Y chromosome; G, donor specific gene(mdx; or sry); V, viral transfection; Ca, calcium phosphate transfection; E, electroporation; Li, liposome genedelivery; M, metabolic label.Heart injury: Normal, Cryoinjury, Reperfusion injury, Coronary occlusion, Genetic (i.e., mdx), Chemical,Cardiotoxic agent.Function improvement/assay: +, improved; L-B, isolated perfused Langendorff with intraventricular balloon; L-F, Langendorff with fluorescence microscopy; M, closed-chest intraventricular micromanometer; M-B,micromanometer with intraventricular balloon; M&S, micromanometer and sonomicrometer; E,echocardiography; F, in vitro force measurements; SPECT, (99mTc) MIBI single photon emission computedtomography; ND, not determined.Angiogenesis: +, angiogenesis; ND, not determined.Cell transplant survival intervention: Y, yes; N, no; Casp Inh, caspase inhibitor; Matrix, support matrix (i.e.,scaffold).

20
Table II. Myoblast transplantation
Donor cell: C, myoblast cell line; NM, neonatal myoblast; AM, adult myoblast; F, single muscle fiber.Species, donor species/host species: M, mouse; R, rat; H, human; D, dog; P, pig; RB, rabbit; HM, hamster; S,sheep; C, cell line; °, autologous transplant.Tracking: D, dye; H, histology; I, immunostain; T, transgenic; CH, Y chromosome; G, donor specific gene(mdx, or sry); V, viral transfection; Ca, calcium phosphate transfection; E, electroporation; Li, liposome genedelivery; M, metabolic label.Heart injury: Normal; Cryoinjury, Reperfusion injury, Permanent coronary occlusion, Genetic (i.e., mdx),Chemical, Cardiotoxic agent.Function improvement/assay: +, improved; L-B, isolated perfused Langendorff with intraventricular balloon; M,closed-chest with intraventricular micromanometer; M-B, micromanometer with balloon; M&S,micromanometer and sonomicrometer; E, echocardiography; F, in vitro force measurements; Ex, exerciseregimens; ND, not determined.Angiogenesis: +, angiogenesis; ND, not determined.Cell transplant survival intervention: Y, yes; N, no; HS, heat shock

21
Table III. Stem cell and stem cell-derived cardiomyocyte transplantation
a No cardiomyogenesis observed.Cell type: ESC, embryonic stem cell; BMSC, bone marrow stem cell; NSC, neuronal stem cell; EnSC,endothelial stem cell; HpSC, hepatocyte stem cell; CSC, cardiac stem cell; UNK, unknown cell type.Species, donor species/host species: M, mouse; R, rat; H, human; D, dog; P, pig; HM, hamster; C, cell line; °,autologous transplant.Cell status at transplant: SC, stem cell; CM, cardiomyocyte; Mobile, mobilized cell population.Delivery: CI, cardiac injection; IU, intrauterine injection; IV, venous injection; AI, arterial injection; BMT, bonemarrow transplant; SCI, subcutaneous injection; BI, blastocyst injection; HHT, human heart transplant; -, notapplicable.Tracking: D, dye; H, histology; I, immunostain; T, transgenic; CH, Y chromosome; G, gene (mdx, or sry); V,viral transfection; Ca, calcium phosphate transfection; E, electroporation; Li, liposome gene delivery; M,metabolic label; PCR, PCR.Heart injury: Normal, Cryoinjury, Reperfusion injury, Permanent coronary occlusion, Genetic (i.e., mdx).Function improvement/assay: +, improved; L-B, isolated perfused Langendorff with intraventricular balloon; M,closed-chest intraventricular micromanometer; M-B, micromanometer with intraventricular balloon; M&S,micromanometer and sonomicrometer; E, echocardiography; Ex, exercise regimens; TSPECT, Thallium-201single photon emission computed tomography; SPECT, (99mTc) MIBI SPECT; V, ventriculography; F, in vitroforce measurements; ND, not determined.Angiogenesis: +, angiogenesis; ND, not determined.

22
References in tables I-III:1. R. SoRelle, Myoblast transplant to heart attempted. Circulation
102 (2000), pp.E9030-9031.
2. B.E. Strauer, M. Brehm, T. Zeus, M. Kostering, A. Hernandez,R.V. Sorg et al., Repair of infarcted myocardium by autologousintracoronary mononuclear bone marrow cell transplantation inhumans. Circulation 106 (2002), pp. 1913-1918.
3. M.H. Soonpaa, G.Y. Koh, M.G. Klug and L.J. Field, Formationof nascent intercalated disks between grafted fetal cardiomyocytesand host myocardium. Science 264 (1994), pp. 98-101.
4. G.Y. Koh, M.H. Soonpaa, M.G. Klug, H.P. Pride, B.J. Cooper,D.P. Zipes et al., Stable fetal cardiomyocyte grafts in the hearts ofdystrophic mice and dogs. J Clin Invest 96 (1995), pp. 2034-2042.
5. C.H. Van Meter, Jr., W.C. Claycomb, J.B. Delcarpio, D.M.Smith, H. deGruiter, F. Smart et al., Myoblast transplantation inthe porcine model: a potential technique for myocardial repair. JThorac Cardiovasc Surg 110 (1995), pp. 1442-1448.
6. J. Leor, M. Patterson, M.J. Quinones, L.H. Kedes and R.A.Kloner, Transplantation of fetal myocardial tissue into theinfarcted myocardium of rat. A potential method for repair ofinfarcted myocardium?. Circulation 94 (1996), pp. II332-336.
7. R.K. Li, Z.Q. Jia, R.D. Weisel, D.A. Mickle, J. Zhang, M.K.Mohabeer et al., Cardiomyocyte transplantation improves heartfunction. Ann Thorac Surg 62 (1996), pp. 654-660 discussion pp.660-661 .
8. A.L. Connold, R. Frischknecht and G. Vrbova, A simple methodfor cardiac surgery in rats. Cell Transplant 5 (1996), pp. 405-409.
9. S. Gojo, S. Kitamura, W.T. Germeraad, Y. Yoshida, K. Niwayaand K. Kawachi, Ex vivo gene transfer into myocardium usingreplication-defective retrovirus. Cell Transplant 5 (1996), pp. S81-84.
10. A.L. Connold, R. Frischknecht, M. Dimitrakos and G. Vrbova,The survival of embryonic cardiomyocytes transplanted intodamaged host rat myocardium. J Muscle Res Cell Motil 18 (1997),pp. 63-70.
11. M. Aoki, R. Morishita, J. Higaki, A. Moriguchi, S. Hayashi, H.Matsushita et al., Survival of grafts of genetically modified cardiacmyocytes transfected with FITC-labeled oligodeoxynucleotidesand the -galactosidase gene in the noninfarcted area, but not themyocardial infarcted area. Gene Ther 4 (1997), pp. 120-127.
12. S. Gojo, S. Kitamura, O. Hatano, A. Takakusu, K. Hashimoto,Y. Kanegae et al., Transplantation of genetically marked cardiacmuscle cells. J Thorac Cardiovasc Surg 113 (1997), pp. 10-18.
13. M. Scorsin, A.A. Hagege, F. Marotte, N. Mirochnik, H. Copin,M. Barnoux et al., Does transplantation of cardiomyocytesimprove function of infarcted myocardium?. Circulation 96 (1997),pp. II188-193.
14. R.K. Li, D.A. Mickle, R.D. Weisel, M.K. Mohabeer, J. Zhang,V. Rao et al., Natural history of fetal rat cardiomyocytestransplanted into adult rat myocardial scar tissue. Circulation 96(1997), pp. II179-186 discussion 186-187 .
15. Z.Q. Jia, D.A. Mickle, R.D. Weisel, M.K. Mohabeer, F.Merante, V. Rao et al., Transplanted cardiomyocytes survive inscar tissue and improve heart function. Transplant Proc 29 (1997),pp. 2093-2094.
16. M. Scorsin, A.A. Hagege, I. Dolizy, F. Marotte, N. Mirochnik,H. Copin et al., Can cellular transplantation improve function indoxorubicin-induced heart failure?. Circulation 98 (1998), pp.II151-155 discussion II155-156.
17. E. Watanabe, D.M. Smith, Jr., J.B. Delcarpio, J. Sun, F.W.Smart, C.H. Van Meter, Jr. et al., Cardiomyocyte transplantation ina porcine myocardial infarction model. Cell Transplant 7 (1998),pp. 239-246.
18. T. Sakai, R.K. Li, R.D. Weisel, D.A. Mickle, Z.Q. Jia, S.Tomita et al., Fetal cell transplantation: a comparison of three celltypes. J Thorac Cardiovasc Surg 118 (1999), pp. 715-724.
19. H. Reinecke, M. Zhang, T. Bartosek and C.E. Murry, Survival,integration, and differentiation of cardiomyocyte grafts: a study innormal and injured rat hearts. Circulation 100 (1999), pp. 193-202.
20. R.K. Li, Z.Q. Jia, R.D. Weisel, D.A. Mickle, A. Choi and T.M.Yau, Survival and function of bioengineered cardiac grafts.Circulation 100 (1999), pp. II63-69.
21. J. Leor, S. Aboulafia-Etzion, A. Dar, L. Shapiro, I.M. Barbash,A. Battler et al., Bioengineered cardiac grafts: a new approach torepair the infarcted myocardium?. Circulation 102 (2000), pp.III56-61.
22. M. Scorsin, A. Hagege, J.T. Vilquin, M. Fiszman, F. Marotte,J.L. Samuel et al., Comparison of the effects of fetalcardiomyocyte and skeletal myoblast transplantation onpostinfarction left ventricular function. J Thorac Cardiovasc Surg119 (2000), pp. 1169-1175.
23. T.S. Li, K. Hamano, K. Kajiwara, M. Nishida, N. Zempo andK. Esato, Prolonged survival of xenograft fetal cardiomyocytes byadenovirus-mediated CTLA4-Ig expression. Transplantation 72(2001), pp. 1983-1985.
24. S. Etzion, A. Battler, I.M. Barbash, E. Cagnano, P. Zarin, Y.Granot et al., Influence of embryonic cardiomyocytetransplantation on the progression of heart failure in a rat model ofextensive myocardial infarction. J Mol Cell Cardiol 33 (2001), pp.1321-1330.
25. H. Yokomuro, R.K. Li, D.A. Mickle, R.D. Weisel, S. Vermaand T.M. Yau, Transplantation of cryopreserved cardiomyocytes. JThorac Cardiovasc Surg 121 (2001), pp. 98-107.
26. Y. Sakakibara, K. Tambara, F. Lu, T. Nishina, G. Sakaguchi,N. Nagaya et al., Combined procedure of surgical repair and celltransplantation for left ventricular aneurysm: an experimentalstudy. Circulation 106 (2002), pp. I193-197.
27. Y. Sakakibara, K. Tambara, F. Lu, T. Nishina, N. Nagaya, K.Nishimura et al., Cardiomyocyte transplantation does not reversecardiac remodeling in rats with chronic myocardial infarction. AnnThorac Surg 74 (2002), pp. 25-30.
28. Y. Sakakibara, K. Nishimura, K. Tambara, M. Yamamoto, F.Lu, Y. Tabata et al., Prevascularization with gelatin microspherescontaining basic fibroblast growth factor enhances the benefits ofcardiomyocyte transplantation. J Thorac Cardiovasc Surg 124(2002), pp. 50-56.
29. A. Ruhparwar, J. Tebbenjohanns, M. Niehaus, M. Mengel, T.Irtel, T. Kofidis et al., Transplanted fetal cardiomyocytes ascardiac pacemaker. Eur J Cardiothorac Surg 21 (2002), pp. 853-857.
30. W. Roell, Z.J. Lu, W. Bloch, S. Siedner, K. Tiemann, Y. Xia etal., Cellular cardiomyoplasty improves survival after myocardialinjury. Circulation 105 (2002), pp. 2435-2441.
31. Rubart M, Soonpaa MH, Nakajima H, Nakajima H, PasumarthiK, Field LJ. In-situ 2-photon microscopy reveals functionalintegration of grafted fetal cardiomyocytes in the adult hostmyocardium. American Heart Association 2002 ScientificSessions, Abstract #104755.
32. M. Scorsin, F. Marotte, A. Sabri, O. Le Dref, M. Demirag, J.L.Samuel et al., Can grafted cardiomyocytes colonize peri-infarctmyocardial areas?. Circulation 94 (1996), pp. II337-340.
33. T. Matsushita, M. Oyamada, H. Kurata, S. Masuda, A.Takahashi, T. Emmoto et al., Formation of cell junctions betweengrafted and host cardiomyocytes at the border zone of ratmyocardial infarction. Circulation 100 (1999), pp. II262-268.
34. T. Imanishi, C.E. Murry, H. Reinecke, T. Hano, I. Nishio,W.C. Liles et al., Cellular FLIP is expressed in cardiomyocytesand down-regulated in TUNEL-positive grafted cardiac tissues.Cardiovasc Res 48 (2000), pp. 101-110.
35. M. Zhang, D. Methot, V. Poppa, Y. Fujio, K. Walsh and C.E.Murry, Cardiomyocyte grafting for cardiac repair: graft cell deathand anti-death strategies. J Mol Cell Cardiol 33 (2001), pp. 907-921.
36. S. Miyagawa, Y. Sawa, S. Taketani, N. Kawaguchi, T.Nakamura, N. Matsuura et al., Myocardial regeneration therapy forheart failure: hepatocyte growth factor enhances the effect ofcellular cardiomyoplasty. Circulation 105 (2002), pp. 2556-2561.

2337. J. Muller-Ehmsen, K.L. Peterson, L. Kedes, P. Whittaker, J.S.Dow, T.I. Long et al., Rebuilding a damaged heart: long-termsurvival of transplanted neonatal rat cardiomyocytes aftermyocardial infarction and effect on cardiac function. Circulation105 (2002), pp. 1720-1726.
38. J. Muller-Ehmsen, P. Whittaker, R.A. Kloner, J.S. Dow, T.Sakoda, T.I. Long et al., Survival and development of neonatal ratcardiomyocytes transplanted into adult myocardium. J Mol CellCardiol 34 (2002), pp. 107-116.
39. T. Shimizu, M. Yamato, Y. Isoi, T. Akutsu, T. Setomaru, K.Abe et al., Fabrication of pulsatile cardiac tissue grafts using anovel 3-dimensional cell sheet manipulation technique andtemperature-responsive cell culture surfaces. Circ Res 90 (2002),p. e40.
40. T. Eschenhagen, M. Didie, J. Heubach, U. Ravens and W.H.Zimmermann, Cardiac tissue engineering. Transplant Immunol 9(2002), pp. 315-321.
41. T. Sakai, R.K. Li, R.D. Weisel, D.A. Mickle, E.J. Kim, S.Tmita et al., Autologous heart cell transplantation improvescardiac function after myocardial injury. Ann Thorac Surg 68(1999), pp. 2074-2080 discussion pp. 2080-2081 .
42. K.J. Yoo, R.K. Li, R.D. Weisel, D.A. Mickle, Z.Q. Jia, E.J.Kim et al., Heart cell transplantation improves heart function indilated cardiomyopathic hamsters. Circulation 102 (2000), pp.III204-209.
43. R.K. Li, R.D. Weisel, D.A. Mickle, Z.Q. Jia, E.J. Kim, T.Sakai et al., Autologous porcine heart cell transplantationimproved heart function after a myocardial infarction. J ThoracCardiovasc Surg 119 (2000), pp. 62-68.
44. T.M. Yau, K. Fung, R.D. Weisel, T. Fujii, D.A. Mickle andR.K. Li, Enhanced myocardial angiogenesis by gene transfer withtransplanted cells. Circulation 104 (2001), pp. I218-222.
45. G.Y. Koh, M.H. Soonpaa, M.G. Klug and L.J. Field, Long-term survival of AT-1 cardiomyocyte grafts in syngeneicmyocardium. Am J Physiol 264 (1993), pp. H1727-1733.
46. J.B. Delcarpio and W.C. Claycomb, Cardiomyocyte transferinto the mammalian heart. Cell-to-cell interactions in vivo and invitro. Ann NY Acad Sci 752 (1995), pp. 267-285.
47. G.Y. Koh, M.G. Klug, M.H. Soonpaa and L.J. Field,Differentiation and long-term survival of C2C12 myoblast grafts inheart. J Clin Invest 92 (1993), pp. 1548-1554.
48. G.Y. Koh, S.J. Kim, M.G. Klug, K. Park, M.H. Soonpaa andL.J. Field, Targeted expression of transforming growth factor-beta1 in intracardiac grafts promotes vascular endothelial cell DNAsynthesis. J Clin Invest 95 (1995), pp. 114-121.
49. S.W. Robinson, P.W. Cho, H.I. Levitsky, J.L. Olson, R.H.Hruban, M.A. Acker et al., Arterial delivery of genetically labelledskeletal myoblasts to the murine heart: long-term survival andphenotypic modification of implanted myoblasts. Cell Transplant 5(1996), pp. 77-91.
50. H. Reinecke and C.E. Murry, Transmural replacement ofmyocardium after skeletal myoblast grafting into the heart. Toomuch of a good thing?. Cardiovasc Pathol 9 (2000), pp. 337-344.
51. K. Suzuki, N.J. Brand, R.T. Smolenski, J. Jayakumar, B.Murtuza and M.H. Yacoub, Development of a novel method forcell transplantation through the coronary artery. Circulation 102(2000), pp. III359-364.
52. K. Suzuki, R.T. Smolenski, J. Jayakumar, B. Murtuza, N.J.Brand and M.H. Yacoub, Heat shock treatment enhances graft cellsurvival in skeletal myoblast transplantation to the heart.Circulation 102 (2000), pp. III216-221.
53. C.E. Murry, R.W. Wiseman, S.M. Schwartz and S.D.Hauschka, Skeletal myoblast transplantation for repair ofmyocardial necrosis. J Clin Invest 98 (1996), pp. 2512-2523.
54. H. Reinecke, G.H. MacDonald, S.D. Hauschka and C.E.Murry, Electromechanical coupling between skeletal and cardiacmuscle. Implications for infarct repair. J Cell Biol 149 (2000), pp.731-740
55. M. Jain, H. DerSimonian, D.A. Brenner, S. Ngoy, P. Teller,A.S. Edge et al., Cell therapy attenuates deleterious ventricular
remodeling and improves cardiac performance after myocardialinfarction. Circulation 103 (2001), pp. 1920-1927.
56. D. Marelli, C. Desrosiers, M. el-Alfy, R.L. Kao and R.C. Chiu,Cell transplantation for myocardial repair: an experimentalapproach. Cell Transplant 1 (1992), pp. 383-390.
57. D. Marelli, F. Ma and R.C. Chiu, Satellite cell implantation forneomyocardial regeneration. Transplant Proc 24 (1992), p. 2995.
58. A. Zibaitis, D. Greentree, F. Ma, D. Marelli, M. Duong andR.C. Chiu, Myocardial regeneration with satellite cellimplantation. Transplant Proc 26 (1994), p. 3294.
59. R.C. Chiu, A. Zibaitis and R.L. Kao, Cellular cardiomyoplasty:myocardial regeneration with satellite cell implantation. AnnThorac Surg 60 (1995), pp. 12-18.
60. P.D. Yoon, R.L. Kao and G.J. Magovern, Myocardialregeneration. Transplanting satellite cells into damagedmyocardium. Tex Heart Inst J 22 (1995), pp. 119-125.
61. D.A. Taylor, S.C. Silvestry, S.P. Bishop, B.H. Annex, R.E.Lilly, D.D. Glower et al., Delivery of primary autologous skeletalmyoblasts into rabbit heart by coronary infusion: a potentialapproach to myocardial repair. Proc Assoc Am Phys 109 (1997),pp. 245-253.
62. D.A. Taylor, B.Z. Atkins, P. Hungspreugs, T.R. Jones, M.C.Reedy, K.A. Hutcheson et al., Regenerating functionalmyocardium: improved performance after skeletal myoblasttransplantation. Nat Med 4 (1998), pp. 929-933
63. J. Dorfman, M. Duong, A. Zibaitis, M.P. Pelletier, D. Shum-Tim, C. Li et al., Myocardial tissue engineering with autologousmyoblast implantation. J Thorac Cardiovasc Surg 116 (1998), pp.744-751.
64. B.Z. Atkins, M.T. Hueman, J. Meuchel, K.A. Hutcheson, D.D.Glower and D.A. Taylor, Cellular cardiomyoplasty improvesdiastolic properties of injured heart. J Surg Res 85 (1999), pp. 234-242.
65. B.Z. Atkins, M.T. Hueman, J.M. Meuchel, M.J. Cottman, K.A.Hutcheson and D.A. Taylor, Myogenic cell transplantationimproves in vivo regional performance in infarcted rabbitmyocardium. J Heart Lung Transplant 18 (1999), pp. 1173-1180.
66. B.Z. Atkins, C.W. Lewis, W.E. Kraus, K.A. Hutcheson, D.D.Glower and D.A. Taylor, Intracardiac transplantation of skeletalmyoblasts yields two populations of striated cells in situ. AnnThorac Surg 67 (1999), pp. 124-129.
67. K.A. Hutcheson, B.Z. Atkins, M.T. Hueman, M.B. Hopkins,D.D. Glower and D.A. Taylor, Comparison of benefits onmyocardial performance of cellular cardiomyoplasty with skeletalmyoblasts and fibroblasts. Cell Transplant 9 (2000), pp. 359-368.
68. R.J. Lee, M.L. Springer, W.E. Blanco-Bose, R. Shaw, P.C.Ursell and H.M. Blau, VEGF gene delivery to myocardium:deleterious effects of unregulated expression. Circulation 102(2000), pp. 898-901.
69. K. Suzuki, B. Murtuza, R.T. Smolenski, I.A. Sammut, N.Suzuki, Y. Kaneda et al., Cell transplantation for the treatment ofacute myocardial infarction using vascular endothelial growthfactor-expressing skeletal myoblasts. Circulation 104 (2001), pp.I207-212.
70. C. Rajnoch, J.C. Chachques, A. Berrebi, P. Bruneval, M.O.Benoit and A. Carpentier, Cellular therapy reverses myocardialdysfunction. J Thorac Cardiovasc Surg 121 (2001), pp. 871-878.
71. B. Pouzet, J.T. Vilquin, A.A. Hagege, M. Scorsin, E. Messas,M. Fiszman et al., Factors affecting functional outcome afterautologous skeletal myoblast transplantation. Ann Thorac Surg 71(2001), pp. 844-850 discussion pp. 850-851 .
72. E.G. Chedrawy, J.S. Wang, D.M. Nguyen, D. Shum-Tim andR.C. Chiu, Incorporation and integration of implanted myogenicand stem cells into native myocardial fibers: anatomic basis forfunctional improvements. J Thorac Cardiovasc Surg 124 (2002),pp. 584-590.
73. N. Dib, E.B. Diethrich, A. Campbell, N. Goodwin, B.Robinson, J. Gilbert et al., Endoventricular transplantation ofallogenic skeletal myoblasts in a porcine model of myocardialinfarction. J Endovasc Ther 9 (2002), pp. 313-319

2474. S. Etzion, I.M. Barbash, M.S. Feinberg, P. Zarin, L. Miller, E.Guetta et al., Cellular cardiomyoplasty of cardiac fibroblasts byadenoviral delivery of MyoD ex vivo: an unlimited source of cellsfor myocardial repair. Circulation 106 (2002), pp. I125-130.
75. K. Suzuki, B. Murtuza, L. Heslop, J.E. Morgan, R.T.Smolenski, N. Suzuki et al., Single fibers of skeletal muscle as anovel graft for cell transplantation to the heart. J ThoracCardiovasc Surg 123 (2002), pp. 984-992.
76. S. Ghostine, C. Carrion, L.C. Souza, P. Richard, P. Bruneval,J.T. Vilquin et al., Long-term efficacy of myoblast transplantationon regional structure and function after myocardial infarction.Circulation 106 (2002), pp. I131-136.
77. H. Reinecke, V. Poppa and C.E. Murry, Skeletal muscle stemcells do not transdifferentiate into cardiomyocytes after cardiacgrafting. J Mol Cell Cardiol 34 (2002), pp. 241-249.
78. M.G. Klug, M.H. Soonpaa, G.Y. Koh and L.J. Field,Genetically selected cardiomyocytes from differentiatingembryonic stem cells form stable intracardiac grafts. J Clin Invest98 (1996), pp. 216-224.
79. A. Behfar, L.V. Zingman, D.M. Hodgson, J.M. Rauzier, G.C.Kane, A. Terzic et al., Stem cell differentiation requires a paracrinepathway in the heart. FASEB J 16 (2002), pp. 1558-1566.
80. J.Y. Min, Y. Yang, K.L. Converso, L. Liu, Q. Huang, J.P.Morgan et al., Transplantation of embryonic stem cells improvescardiac function in postinfarcted rats. J Appl Physiol 92 (2002), pp.288-296.
81. Y. Yang, J.Y. Min, J.S. Rana, Q. Ke, J. Cai, Y. Chen et al.,VEGF enhances functional improvement of postinfarcted hearts bytransplantation of ESC-differentiated cells. J Appl Physiol 93(2002), pp. 1140-1151.
82. S. Tomita, R.K. Li, R.D. Weisel, D.A. Mickle, E.J. Kim, T.Sakai et al., Autologous transplantation of bone marrow cellsimproves damaged heart function. Circulation 100 (1999), pp.II247-256.
83. J.S. Wang, D. Shum-Tim, J. Galipeau, E. Chedrawy, N.Eliopoulos and R.C. Chiu, Marrow stromal cells for cellularcardiomyoplasty: feasibility and potential clinical advantages. JThorac Cardiovasc Surg 120 (2000), pp. 999-1005.
84. K.W. Liechty, T.C. MacKenzie, A.F. Shaaban, A. Radu, A.M.Moseley, R. Deans et al., Human mesenchymal stem cells engraftand demonstrate site-specific differentiation after in uterotransplantation in sheep. Nat Med 6 (2000), pp. 1282-1286
85. K.A. Jackson, S.M. Majka, H. Wang, J. Pocius, C.J. Hartley,M.W. Majesky et al., Regeneration of ischemic cardiac muscle andvascular endothelium by adult stem cells. J Clin Invest 107 (2001),pp. 1395-1402.
86. C.E. Murry, M. Rubart, M. Soonpaa, H. Nakajima, H.Nakajima and L.J. Field, Absence of cardiac differentiation inhematopoietic stem cells transplanted into normal and injuredhearts. Circulation (abstract) 104 (2001), p. II599.
87. J.S. Wang, D. Shum-Tim, E. Chedrawy and R.C. Chiu, Thecoronary delivery of marrow stromal cells for myocardialregeneration: pathophysiologic and therapeutic implications. JThorac Cardiovasc Surg 122 (2001), pp. 699-705.
88. B.E. Strauer, M. Brehm, T. Zeus, N. Gattermann, A.Hernandez, R.V. Sorg et al., (Intracoronary, human autologousstem cell transplantation for myocardial regeneration followingmyocardial infarction). Dtsch Med Wochenschr 126 (2001), pp.932-938.
89. D. Orlic, J. Kajstura, S. Chimenti, I. Jakoniuk, S.M. Anderson,B. Li et al., Bone marrow cells regenerate infarcted myocardium.Nature 410 (2001), pp. 701-705.
90. K. Hamano, T.S. Li, T. Kobayashi, K. Hirata, M. Yano, M.Kohno et al., Therapeutic angiogenesis induced by localautologous bone marrow cell implantation. Ann Thorac Surg 73(2002), pp. 1210-1215.
91. Y. Jiang, B.N. Jahagirdar, R.L. Reinhardt, R.E. Schwartz, C.D.Keene, X.R. Ortiz-Gonzalez et al., Pluripotency of mesenchymalstem cells derived from adult marrow. Nature 418 (2002), pp. 41-49.
92. T.C. Mackenzie, A.F. Shaaban, A. Radu and A.W. Flake,Engraftment of bone marrow and fetal liver cells after in uterotransplantation in MDX mice. J Pediatr Surg 37 (2002), pp. 1058-1064.
93. T. Saito, J.Q. Kuang, B. Bittira, A. Al-Khaldi and R.C. Chiu,Xenotransplant cardiac chimera: immune tolerance of adult stemcells. Ann Thorac Surg 74 (2002), pp. 19-24 discussion p. 24 .
94. S. Tomita, D.A. Mickle, R.D. Weisel, Z.Q. Jia, L.C. Tumiati,Y. Allidina et al., Improved heart function with myogenesis andangiogenesis after autologous porcine bone marrow stromal celltransplantation. J Thorac Cardiovasc Surg 123 (2002), pp. 1132-1140. .
95. C. Toma, M.F. Pittenger, K.S. Cahill, B.J. Byrne and P.D.Kessler, Human mesenchymal stem cells differentiate to acardiomyocyte phenotype in the adult murine heart. Circulation105 (2002), pp. 93-98.
96. A.J. Wagers, R.I. Sherwood, J.L. Christensen and I.L.Weissman, Little evidence for developmental plasticity of adulthematopoietic stem cells. Science 297 (2002), pp. 2256-2259.
97. R.E. Bittner, C. Schofer, K. Weipoltshammer, S. Ivanova, B.Streubel, E. Hauser et al., Recruitment of bone-marrow-derivedcells by skeletal and cardiac muscle in adult dystrophic mdx mice.Anat Embryol (Berlin) 199 (1999), pp. 391-396.
98. D.L. Clarke, C.B. Johansson, J. Wilbertz, B. Veress, E.Nilsson, H. Karlstrom et al., Generalized potential of adult neuralstem cells. Science 288 (2000), pp. 1660-1663.
99. G. Condorelli, U. Borello, L. De Angelis, M. Latronico, D.Sirabella, M. Coletta et al., Cardiomyocytes induce endothelialcells to trans-differentiate into cardiac muscle: implications formyocardium regeneration. Proc Natl Acad Sci USA 98 (2001), pp.10733-10738.
100. N.N. Malouf, W.B. Coleman, J.W. Grisham, R.A. Lininger,V.J. Madden, M. Sproul et al., Adult-derived stem cells from theliver become myocytes in the heart in vivo. Am J Pathol 158(2001), pp. 1929-1935.
101. A.M. Hierlihy, P. Seale, C.G. Lobe, M.A. Rudnicki and L.A.Megeney, The post-natal heart contains a myocardial stem cellpopulation. FEBS Lett 530 (2002), p. 239.
102. R.H. Hruban, P.P. Long, E.J. Perlman, G.M. Hutchins, W.A.Baumgartner, K.L. Baughman et al., Fluorescence in situhybridization for the Y-chromosome can be used to detect cells ofrecipient origin in allografted hearts following cardiactransplantation. Am J Pathol 142 (1993), pp. 975-980.
103. R. Glaser, M.M. Lu, N. Narula and J.A. Epstein, Smoothmuscle cells, but not myocytes, of host origin in transplantedhuman hearts. Circulation 106 (2002), pp. 17-19.
104. M.A. Laflamme, D. Myerson, J.E. Saffitz and C.E. Murry,Evidence for cardiomyocyte repopulation by extracardiacprogenitors in transplanted human hearts. Circ Res 90 (2002), pp.634-640.
105. P. Muller, P. Pfeiffer, J. Koglin, H.J. Schafers, U. Seeland, I.Janzen et al., Cardiomyocytes of noncardiac origin in myocardialbiopsies of human transplanted hearts. Circulation 106 (2002), pp.31-35.
106. F. Quaini, K. Urbanek, A.P. Beltrami, N. Finato, C.A.Beltrami, B. Nadal-Ginard et al., Chimerism of the transplantedheart. New Engl J Med 346 (2002), pp. 5-15.
107. D. Orlic, J. Kajstura, S. Chimenti, F. Limana, I. Jakoniuk, F.Quaini et al., Mobilized bone marrow cells repair the infarctedheart, improving function and survival. Proc Natl Acad Sci USA98 (2001), pp. 10344-10349.
____________________________________________________________________________________________________________________

25
D. MATERIALS AND METHODS
1. ESC culture and differentiation
Mouse ESC CGR8 were cultured in BHK21 medium (GIBCO BRL) supplemented with
nonessential amino acids, pyruvate, mercaptoethanol, glutamine, penicillin/streptomycin, 10%
fetal calf serum and LIF-conditioned medium in a humidified 5% CO2 atmosphere at 37°C and
maintained at < 70% confluency to keep an undifferentiated phenotype (Meyer et al., 2000).
Figure A. Colonies of undifferentiated mouse ESC. a) D3-ES clone growing on mytomycin-inactivatedmouse embryonic fibroblasts (MEFs); b) CGR8-ES clone feeder-cell independent (no need ofMEFs).Culture and propagation in the presence of LIF.
The day of the injection procedure, cells were harvested by trypsinization, washed and
pelleted, then suspended at a concentration of 2x107/ml in serum-free culture medium. The
differentiation of CGR8 cell was performed by the hanging drop method (Maltsev et al., 1994).
Colonies of mouse EScells
Mitomycin-inactivatedmouse embryonic
fibroblasts
a b
10 µµm
Propagationand
self renewal
Cell dissociationby trypsin
+ LIF

26
Figure B. Cardiac cell differentiation byhanging drop method, and isolated by Percollgradient centrifugation.
As illustrated in Figure B, EBs were formed at day 2 in hanging drops (450 cells/20µl) of
differentiation medium (BHK21, as described above), containing 20% fetal calf serum and
lacking of LIF. After 6 days in suspension culture EBs were plated to gelatin-coated 24-well
plates or cover slips (Meyer et al., 2000).
The beating cardiomyocytes within EBs were counted under phase-contrast
microscope at day 8 of differentiation.
Figure C. Sequence of pictures illustrating thePassage from ESC to EBs in suspension thenadherent EBs containing beatingcardiomyocytes.
Hanging drop method for cardiac cell differentiation
and isolation of ES-derived cardiomyocytes
ES cells
PBS
Formation of Embryoid Bodies (EB)
Hanging drops with ES cells
EB culture in suspension
EB culture on gelatin-coated wells
Cell dissociation by collagenase treatment
by Percoll gradient centrifugation
Terminally-differentiated
cardiomyocytes
Days
0
2
4
6
10
12
culture medium
Embryoid bodyat day 8
Embryoid bodyat day 4
colony of mouseundifferentiated
ES cells
10 µµm
Beating area containingcontracting
cardiomyocytes
- LIF

27
2. Isolation of ES-derived and neonatal mouse cardiomyocytes
As illustrated in figure, day-8 EBs containing cardiomyocytes were detached from culture
surfaces by incubating them with 0.05% trypsin-EDTA for 1 min at 37°C. Cells were dissociated
with 1mg/ml collagenase (CLSII, Wortington), and 0.25% mg/ml pancreatin in a buffer
containing (in mmol/L) NaCl 117, HEPES 20, NaH2PO4 1.2, KCl 5.4, Mg SO4 1, glucose 5, pH
= 7.35.
As shown in Figure D, myocytes were separated by centrifugation through a discontinuous
Percoll gradient, and collected at the interface of the two layers. Neonatal cardiomyocytes were
isolated from ventricles of 2 days-old mouse neonates using a similar method as previously
described (Jaconi et al., 2000).
Figure D. Isolation of cardiomyocyte from day-8EBs by enzymatic dissociation and centrifugationon a discontinuous Percoll Gradient.
3. Plasmid construction, stable transfection and isolation of ESC clones
We designed a mouse ESC clone expressing GFP fused to CD63. This fusion protein allows
the expression of green fluorescence associated with membrane and intracellular endosomes.
CD63-GFP construct was electroporated into CGR8 cells according to the standard protocol in a
Gene Pulser (BioRad) at 240v, 500µF. Stable ESC clones were propagated in the presence of
Embryoid bodyat day 8
discontinuousPercoll gradient
centrifugation
non-muscle cells
myocytes
Dissociationwith collagenase

28
LIF and selected for 10 days using G418 (250µg/ml), and sorted by FACS. In parallel, we also
prepared a mouse ESC clone expressing the human Bcl2 gene. Human Bcl2 cDNA inserted into
pcDNA3.1(-) at EcoRI and Hind III restriction sites. After selection of stable clones, Bcl2 could
be identified by immunohistochemistry using anti-Blc2 monoclonal antibodies (Figure E).
Figure E. ESC stable clones (CGR8) containing the CD63-GFP construct and resistant to neomycin.
4. Rat model of myocardial infarction
Male Sprague-Dawley rats weighing 300-350 grams were initially anesthetized with 4-5%
isoflourane in an induction chamber. Following the shaving and weighting, the rat was intubated
with a 14-gauge catheter, tracheal ventilation was performed at 70 cycles/min with 2.5-3.0mL
tidal volume, room air supplemented with oxygen (Harvard Rodent Ventilator, Model 683,
Harvard Apparatus Co, Inc). 1.5-2% isoflourane was maintained for continuous anesthesia.
Figure F. Anestesia chamber andisofluorane delivery system.
GFPCD6320 µµm
Mouse embryonic fibroblasts
Electroporation intoundifferentiated ES cells
Selection of stableESC clones by G418
Plasmid construction Green ES cells clone

29
Three electrodes were positioned to record the electrocardiographic tracing (ECG) monitor. The
respiration curve was also recorded during all procedure, as indicated in the Figure G.
Figure G. ECG and ventilation setup (left) as installed during the procedure. ECG and ventilation tracesrecordings (right). An IV line was installed in the left femoral vein if saline infusion is needed.
A left intercostal thoracotomy was performed under aseptic technique. The forth-intercostal
space was opened carefully to avoid accidentally cutting any vessels including the internal
mammary artery. The forth and fifth ribs were separated with a small retractor (Harvard
Apparatus, France) to explore the heart. The pericardium was removed, the left anterior
descending artery and its branch was observed under surgical microscope. A 6-0 polypropylene
snare was made passing through the epicardium layer around the origin of the artery between the
left atrium and the right pulmonary outflow tract, tying the ligature permanently occluded the
artery (Ye et al., 1997).
The muscle layer and skin were closed with 3-0 suture afterwards. Before the rat woke up
completely, extubation was performed and the rat was places in a recovery cage with a supply of
oxygen for 30 to 60 minutes.

30
Figure H. Panels a,b: ligature procedure. After a left intercostal thoracotomy was performed, the fourthintercostal space was opened and the ribs were separated with a small retractor in order to expose theheart. LAD was ligated by a 6-0 polypropylene snare. Panel c: Drawing illustrating that right after LADligature, the left ventricular anterior free wall becomes hypokinetic and clearer due to the cyanosis.
5. Experimental animal protocol
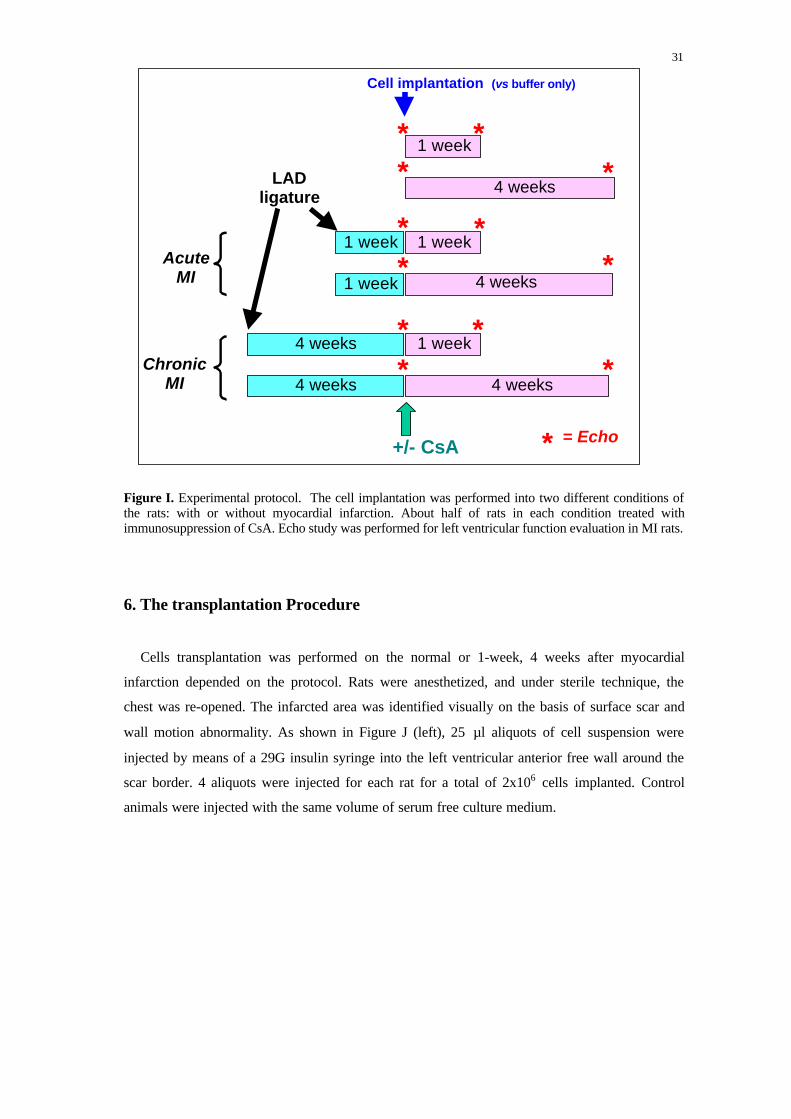
We divided the rats into two groups: normal rats (non-MI, n = 24) and MI rats (n= 47). The
MI rats were subdivided to acute phase (MI = 1week) and chronic phase (MI = 4 weeks). The
undifferentiated murine ESC was implanted to both normal and infarcted rats. Also we
implanted cardiomyocytes derived from ESC in a few rats. 1 week or 4 weeks after implantation,
the rats were sacrificed and the histologic and pathologic studies were performed. On the each
condition, about half of rats received immunosuppression treatment of cyclosporine A. To
evaluate the transformation of left ventricular function, the echocardiograph study was
performed to the rats with infarction the day before cells grafting and the day before sacrificing
respectively (Fig I).
LAD
Infarctedarea
ligature
a
b
c
31
Figure I. Experimental protocol. The cell implantation was performed into two different conditions ofthe rats: with or without myocardial infarction. About half of rats in each condition treated withimmunosuppression of CsA. Echo study was performed for left ventricular function evaluation in MI rats.
6. The transplantation Procedure
Cells transplantation was performed on the normal or 1-week, 4 weeks after myocardial
infarction depended on the protocol. Rats were anesthetized, and under sterile technique, the
chest was re-opened. The infarcted area was identified visually on the basis of surface scar and
wall motion abnormality. As shown in Figure J (left), 25 µl aliquots of cell suspension were
injected by means of a 29G insulin syringe into the left ventricular anterior free wall around the
scar border. 4 aliquots were injected for each rat for a total of 2x106 cells implanted. Control
animals were injected with the same volume of serum free culture medium.
LADligature
+/- CsA
* ** *
* **
Cell implantation (vs buffer only)
* = Echo
1 week
1 week
4 weeks
4 weeks
4 weeks
1 week
1 week
1 week
4 weeks
**
**
*
AcuteMI
ChronicMI
4 weeks

32
Figure J. Left: Cell transplantation was performed bytransmyocardial injection around the infarction borderwith 4 times 25µl for a total number of cells between1x106 and 2x106. Right: When required, osmoticminipump (ALZA Corporation) was implantedsubcutaneously to deliver CsA continuously.
7. Immunosuppression treatment
To prevent rejection and study the effect of an immunosuppression treatment on ESC fate,
groups of rats received immunosupression agent cyclosporin A delivered continuously via an
osmotic minipump (ALZA Corporation), as shown in Figure K.
Figure K. Rat with a subcutaneous implantationOf an osmotic minipump filled with CsA.
Alzet mini-osmotic pumps were filled with cyclosporin A (CsA) (Sandimmune, Novartis
50mg/1ml), and was kept overnight at 37°C in PBS before implantation. The CsA release was
adjusted at 2.5µl/hour or 10µl/hour and pump were designed for a 7- or 28-days release. The
administrated dosage of CsA was calculated as 6-9 mg/Kg/day (Sullivan et al., 2000). After hair
shaving and skin cleaning at the site for incision, a hemostat was inserted into the incision to
spread the subcutaneous tissue and create a prompt pocket for the pump. The filled pump was
implanted subcutaneously and the wound closed with suture. All procedure was performed under
sterile circumstances.
4 x 25 µµ l with 250’000-500’000cells
LADLigation
Infarctedarea
Osmotic minipump + CsA

33
8. Evaluation of the left ventricular function by echocardiography
For the evaluation of left ventricular function, transthoracic echocardiogram was performed
on the rats after myocardial infarction 1 week or 4 weeks right before implantation (baseline
echocardiogram), and 1 week or 4 weeks after implantation, before the sacrifice of the animals.
Rats were anesthetized with 4-5% isoflurane in an induction chamber. The chest was shaved, the
rats were placed in dorsal decubitus position and intubated for continuous ventilation. 1-2%
isoflurane was continuously supplied via a mask (Moises et al., 2000). 3 electrodes were adhered
to their paws to record the electrocardiographic tracing simultaneously with the cardiac image
identifying the phase of a cardiac cycle.
Echocardiograms were performed with a commercially available echocardiography system
equipped with 7.5 MHz phased–array transducer (Philips-Hewlett-Packard). The transducer was
positioned on the left anterior side of the chest (Leor, 2000 #25). At first, longitudinal images of
the heart were obtained, including the left ventricle, atrium, the mitral valve and the aorta,
followed by the cross-sectional images from the plane of the base to the left ventricular apical
region. M-mode tracings were obtained at the level below the tip of the mitral valve leaflets at
the level of the papillary muscles (Litwin et al., 1994). All of two-dimensional images, M-mode
tracings and Doppler curves were recorded on videotape for later analysis. We calculated the
fractional shorting (FS) as a measure of systolic function, according to the M-mode tracing from
the cross-sectional view: maximal LV end-diastolic diameter (at the time of maximal cavity
dimension), minimal LV end-systolic diameter (at the time of maximum anterior motion of the
posterior wall), FS (%) = {(LVEDD-LVESD) / LVEDD} x 100, (Pouzet et al., 2000). All
measurements were averaged for 3 consecutive cardiac cycles.
9. Histological and immunohistochemistry study
Either 1 or 4 weeks after cells implantation, the rats were sacrificed with intravenous injection
of potassium chloride at the level of the femoral vein, in order to stop the cardiac ventricular
contraction on the diastole. Hearts were rapidly excised, the cardiac cavities were rinsed in PBS
to removed blood and thrombus, then the hearts were fixed with 10% formalin for 24 hours, and
immediately thereafter embedded in paraffin. Blocks were sectioned into 3µm thickness slices
and stained with hematoxylin and eosin. Serial sections were immunolabeled with different
antibodies against the cell markers and structural cardiac proteins. Upon cell injection, we
retrieved the engrafted cells according to the markers carried by the ESC (GFP or human Bcl2),

34
while D3-ESC, which were not genetically modified, were identified only by morphology and
the proliferative marker PCNA.
9. 1. Antibodies
All the used antibodies are summarized in the following table:
Antibody Antigen Recognized Dilution Source
Ab6556 GFP 1:400 Abcom UK
MF-20 Sarcomere myosin 1:400 DSHB, Iowa, USAPC10 PCNA 1:100 DAKO
CXN-6 Connexin-43 1:200 SIGMA
EA-53 a-actinin (sarcomeric) 1:200 SIGMA
M0887 BCL-2 1:100 DAKO
Anti-Bcl-2 antibody is a monoclonal mouse antibody specific for human Bcl-2 oncoprotein
(DAKO, M0887). Monoclonal anti- α-actinin (sarcomeric, clone EA-53, mouse ascites fluid,
Sigma) is specific for α-skeletal muscle actinin and α-cardiac muscle actinin. It stains Z lines
and dots in stress fibers of myotubes in skeletal and cardiac muscle, but not in non-sarcomeric
muscle elements (connective tissue, epithelium, nerves, and smooth muscle). Monoclonal anti-
PCNA (proliferating cell nuclear antigen) antibody (clone PC-10, DAKO) reacts with PCNA, a
useful index of cell proliferation. Anti-myosin antibody MF 20 (DSHB, Developmental Studies
Hybridoma Bank, University of Iowa, USA) is an IgG2b monoclonal antibody, which reacts
with all sarcomeric myosin. Anti-connexin-43 antibody (clone cxn-6, Sigma) is a mouse IgM
isotype monoclonal antibody, which reacts with the gap junction protein connexin 43 (Cx43).
Anti-GFP antibody (Abcam, ab6556) is a rabbit polyclonal to GFP (affinity purified) antibody.
9. 2. Immunohistochemistry
For immunohistochemical studies, tissue slices were serially rehydrated in 100%, 95%, and
70% ethanol after deparaffinization with xylol. Endogenous peroxidase in the sample was

35
blocked and the antigens were retrieved on a microwave for 15 minutes in pH=6.0, 10mM TP
citrate solution. Non-specific binding were blocked with 5% BSA/PBS solution for another 15
minutes. The slices were stained with following primary antibodies at room temperature for 1
hour: anti-Bcl-2 (1:100), anti-α-actinin (1:200), anti-PCNA (1:100), anti-GFP (1:400), anti-
sarcomere myosin (1:400), and anti-Cx43 (1:200). Then the slices were incubated for 30 minutes
at room temperature with one of the following biotinylated secondary antibodies diluted 1:100 to
1:200 in 5% BSA/PBS: Biotinylated anti-rabbit IgG (H+L), affinity purified (VECTOR, BA-
1000), Biotinylated anti-mouse IgM (mu-chain specific) affinity purified (VECTOR, BA-2020),
Biotinylated anti–mouse IgG (H+L) affinity purified and rat adsorbed. This was followed by
another 30 minutes incubation in avidin-biotin solution (Vectastin ABC Kit). All the incubations
were preceded by through rinsing with PBS. The reaction products were visualized with 3,3’-
Diaminobenzidine tetrahydrochloride Dehydrate and 3% hydrogen peroxide. Then the slices
were washed with distilled water; counter staining was performed with hemotoxyline for 1
minute. Control secondary antibodies were processed simultaneously using the same protocol,
except that the dilution solution was devoid of primary antibodies. The tissue was observed,
photographed and digitally saved on a ZEISS Axioskop 2 Plus photomicroscope.
10. Statistics Analysis
All values are presented as means ± S.E. Difference between two groups was compared by
unpaired Student’s t-test. A level of p < 0.05 was considered as significant.

36
E. RESULTS
E. 1. Grafting undifferentiated ESC in normal (non-infarcted) heart
We first established optimal conditions for the ESC implantation into the ventricle wall of
normal rat hearts (n=24). We inspected the fate of the cells 1 week or 4 weeks later. 10/24 rats
were treated with CsA. We tested different kind of ESC lines, such as the feeder-cell dependent
cell lines D3 and feeder cell-independent cell line CGR8 (wild type of containing Bcl2 or GFP
marker). As depicted in Figure 1, H&E staining revealed an consistent granulation tissue in the
control rats while 33% (1/3) of the CsA-treated animals displayed, in addition to granulation, the
growth of teratoma after 1 week. This percentage increased to 100% (6/6) after 4 weeks. At this
time point, the tumors were highly proliferative and some even bigger that the ventricle
diameter, as indicated by the positive PCNA staining (see Fig.2b and Fig. 3b). The ESC marker
Bcl-2 (as well as GFP, not shown) was also strongly positive in the teratoma area (see Figure 2c
for Bcl2-positive tumors), indicating the ESC derivation. In contrast, in the absence of
immunosuppression, no teratoma was found even after 4 weeks (0/13) (Fig. 2d and Fig. 3 c) and
no cell proliferation or granulation was still present at this later time (Fig 2e and Fig. 3d).
The tumors were identified as teratoma with characteristics of tissues originating from the
ectoderm (skin), mesoderm (cartilage, muscle) and endoderm (epithelium of bronchi and of
pancreas) (see Figure 4).

37
Figure 1

38
Figure 2

39
Figure 3

40
Figure 4

41
E. 2. Rat model of myocardial infarction (MI)
Large, reproducible lesions were induced in rat heart by coronary artery ligation, as
illustrated in Figure 5. In the left panel, the explanted heart, 4 weeks after the coronary ligature
presented a large infarcted area (dotted line) with a collapsed left ventricular wall due to the
extremely reduced thickness. Before sacrifice, the infarcted area was clearly identifiable by
echocardiography as shown in the right panel of Figure 5. Using a parasternal short axis view it
was possible to estimate the akinetic territory going from 12 to 4 o'clock, as indicated by the
dotted line. In this example, the impairment of the left ventricular ejection fraction (LVEF) was
visually estimated at 25%.
Figure 5. Model of MI in rat. The heart was explanted 4 weeks after ligation of LAD. Leftpanel: macroscopical view of the explanted heart. Right panel: echocardiographic picture of thesame heart using a parasternal short axis view.
Figure 6 illustrates heart sections produced from normal rats (panels a,b,c), acute MI rats (1
week, panels d,e,f) and chronic MI rats (4 weeks, panels g,h,i). Upon H&E staining, an acute
progressive tissue inflammation was observed after 1 week of MI (Fig 6d,e,f), as compared with
normal heart (Fig. 6a,b,c). One could observe neutrophilic infiltration, fragmentation and
smudging of the muscle fibers and coagulative necrosis (Fig.6 e, f). 4 weeks from the MI, almost
no granulation tissue was seen; instead collagen deposition and myocardial scar were present.
Infarcted heartMI=4 weeks post-ligation
Echocardiographparasternal short axis view
ligation
Infarctedanterior area LVEF ~ 25%

42
Figure 6

43
As displayed in Figure 7, we first compared normal heart with infarcted hearts 1 week after
MI. At this time point, one could see an acute and progressive tissue inflammation on left
ventricular wall. Pericarditis was present. The tissue pathology of heart sections revealed by
H&E staining or by immunohistochemistry indicated i) a strong cell proliferation in the infarcted
area (increased PCNA staining over control), ii) disorganized cardiac fibers with an abnormal α-
actinin and MHC distribution in the sarcomeres (fuzzy staining instead of z-lines and A-bands
respectively), iii) partial loss of Cx43 at the gap junctions between fibers.
The same type of abnormal pathology but exacerbated was observed after 4 weeks. At this
time the ventricular wall become very thin and completely fibrotic (Figure 5, left panel and
Figure 6 g, h, i). Only scar tissue was visible with non contracting, dead wavy fibers.
E. 3. Engraftment of undifferentiated ESC in acute phase of MI (1 week)
Dead cells or infiltrated leukocytes releases lysosome content, which result in further cellular
autolysis or heterolysis, followed by coagulated necrosis. Enzymatic digestion of cells and
protein denaturation is then removed by phagocytosis. We engrafted undifferentiated ESC at this
time point to study the influence of the inflammation on cell proliferation and differentiation. In
fact lot of cytokines are released by inflammatory cells, which are though to modify substantially
cell fate as well as tissue repair.
We compared 20 rats implanted with undifferentiated ESC and distributed in 2 groups, each
comprising animals treated and non-treated with CsA, as summarized by the following scheme:
ligature
* ** *
1 week
1 week
1 weekAcute
MI 4 weeks
+/- CsA * = Echo
Cell injection

44
Figure 7.

45
In MI rats after 1 week post-implantation the histology indicated an inflammatory reaction
still underway. Infiltrated mononuclear cells together with ingrowth fibroblasts and capillaries
formed a granulation tissue in the infarcted area, similarly to control conditions showed in Figure
6 and 7 (MI but no cells engrafted). Animals injected with buffer only displayed a tissue
morphopathology identical to non injected animals at same stages (n=4).
In contrast, 40% (2/5) of the rats treated with CsA formed a tumor after 1 week, while 100%
(8/8) presented a highly proliferating teratoma after 4 weeks (strong PCNA staining),
respectively. Figure 8 shows the heart sections corresponding to this last condition (i.e. T=4
weeks). Strikingly, in contrast to normal rats (no MI), some areas of the teratoma contained cells
positive for MHC, α-actinin and Cx43, indicating a partial differentiation of ESC into muscle
cells and fibers.
As illustrated in Figure 9, 4 weeks after implantation, only a scar tissue was visible in the
absence of CsA and no teratoma was observed. Moreover no engrafted could be identified: both
Bcl-2 and GFP staining were negative. Also, all types of the cardiomyocytes specific staining,
cardiac muscle gap protein (Cx43), sarcomere myosin (MF20), sarcomere actinin (α-actinin)
were negative on the area of MI (Fig. 9), which suggest that without CsA, no cardiomyocyte
differentiation in MI area could be identified.
E. 4. Grafting of undifferentiated ESC in chronic phase of MI (4 weeks)
Histology evaluation of the infarcted area after 4 weeks ligation confirmed the presence of
transmural fibrous tissue at the apex of left ventricle (total rats = 18). Comparing with infarcted
area after 1-week ligation, pathologic changes after 4-weeks ligation comprised a fibrotic
repairing tissue instead of necrosis. There was no more granulation tissue, but a rich vascular
network and collagen deposition. Left ventricular cavity is more dilated globular (Figure 6). The
same type of morphology was observed in CsA-immunosuppressed rats injected with
undifferentiated ESC. 20% (1/5) of these animals developed a tumoral mass 1 week later, a
percentage reaching 100% (3/3) after 4 weeks. These masses contained tissue from the three
different germ layers, typical of teratoma. They were positive for the human Bcl2 marker,
confirming their origin from injected ESC. Animals injected with culture medium or injected
with ESC but without CsA-immunosuppression did not present any teratoma or identifiable ESC
in the grafting area. Bcl-2 and PCNA staining were both negative.

46
Figure 8

47
Figure 9

48
Figure 10

49
E. 5. Grafting of ESC-derived CMC in MI rats.
CMC was derived from Bcl2-ESC after Percoll gradient isolation of beating EBs at day 8,
were injected into 1week MI rats (n=5). All the rats received CsA and were sacrificed for the
pathological examination 4 weeks later. Figure 10 shows that in 2 out of 5 animal hearts,
implanted CMC formed aggregates with positive staining for Bcl2 and PCNA. Strikingly, these
aggregates resulted positive for MHC and α-actinin Figure 11A. However, 3 out of 5 did not
show any teratoma formation, as reported in Figure 11B. In those cases, there was no positive
staining for PCNA or MHC-positive ES-derived cells.
Figure 11. Immunohistochemistry analysis of MI rat hearts implanted with ES-derived cardiomyocytes 4weeks after coronary ligature. A) Teratoma formation with extensive positive labeling for MHC and α-actinin. B) Same conditions as in A, but in 3 out of 5, no teratoma was found.
MHC αα-actinin
50X
200X
100X
400X
H&E MHC
a
b
c
d
10X 10X
A
B

50
E. 6. Effect of time and immunosuppression on teratoma formation
In our study however, independently of the type of cardiac injury (acute or chronic MI), the
injected ESC had the tendency of differentiate to teratoma in vivo, displaying different tissue
types but myocytes. The teratoma formation was exclusively dependent on the
immunosuppression treatment and the length of transplantation. Figure 12 summarizes the data
on the frequency of teratoma formation as a function of time (weeks of transplantation), normal
rats being compared to MI 1 week and 4 weeks. Clearly, in all of these three of conditions, non-
immunosuppressed rats never developed teratoma, indicating an active removal of non-
autologous cells from the immune system.
Figure 12. Effect of time and immunosuppression on the occurrence of teratoma. Normal rats werecompared to MI rats (1 week and 4 weeks). After transplantation of undifferentiated ESC, teratomaformation was dependent on the administration of CsA as well as on the duration of cell implantation, butnot the type of ESC, nor on the presence of a cardiac injury.
Normal rats MI 1 week MI 4 weeks
0
10
2030
4050
6070
8090
100
%
teratoma no teratoma
1 4 1 4 1 4 1 4 1 4 1 4
CsA - + - + - +T (# of weeks)

51
B. 7. The evaluation of left ventricular function by echocardiogram
The echocardiogram was performed before and after ESC transplantation. Cardiac parameters
were collected. Figure 16 show the two-dimensional imaging and M–mode tracing of
echocardiogram. Cardiac function of MI rat was much worse compared with non-MI rat
(control), which was identified by the echocardiograph: segmental abnormality of LV wall
motion, global decrease of LV contractility, a change in shape of LV cavity. When tumor was
formed (which may growth towards or outwards LV chamber), it could be identified from both
2-D and M-mode imaging, but it will influence on the judgment of the LV wall motion and LV
function (Figure 13).
Cardiac function was assessed according to the value measured on the M-mode tracing.
Table 1 showed the difference between the cardiac parameters such as: heart rate,
(LVEDDafterT - LVEDDbeforeT), (LVESDafter T - LVESDafter T), and (FSafter T - FSafter T).
When ESC were transplanted into acute or chronic phase of MI (1 week or 4 weeks) under
immunosuppression, comparison of delta values between those groups measured 4 weeks after
transplantation did not give any significant difference.
Ischemia-induced inflammation present at an early MI stage (up to 1 week) did not influence
heart functions.
Table 1. Comparison MI=1 week vs. MI=4weeks (delta of cardiac parameters (afterT -before T)), in the presence of CsA. Effect 4 weeks after cell transplantation (n=11).
Heart rate (bpm)
LVEED (mm)
LVESD (mm)
FS (%)
MI =1week ± S.E.
29 ± 13
2.2 ± 1.3
1.1 ± 0.92.3 ± 0.4
MI =4 weeks ± S.E.
13 ± 3.3
1.2 ± 0.6
0.3 ± 1.73.2 ± 1.6
P0.837
0.153
0.5400.683

52
Figure 13.

53
Also in our study, immunosuppression treatment did not affect the changes of cardiac
parameters (table 2). Under the same condition, i.e. 1 week after LAD ligature, ESC
transplantation for a duration of 4 weeks, there was no significant difference on the cardiac delta
value) whether CsA was administrated or not.
Table 2. Effect of CsA treatment in MI=1 week (delta of parameters), 4 weeks after
transplantation (n=12).
We could not finally conclude with the data, because of the presence of teratoma caused to
many artifacts to the measurement of the different parameters.
11 ± 45
3.7 ± 2.6
2.2 ± 1.5
3.3 ± 1.9
29 ± 13
2.2 ± 1.3
11.4 ± 0.9
2.3 ± 4.4
0.396
0.497
0.610
1.000
No CsA with CsA PHeart rate (bpm)
LVEED (mm)
LVESD (mm)
FS (%)

54
F. DISCUSSION
The major aim of this study was to investigate whether an infarcted heart could instruct
undifferentiated mouse ESC to acquire a cardiac phenotype in situ in order to improve cardiac
function in a rat model of myocardial infarction in the rat. We studied different type of murine
ESC lines engrafted into rat model of myocardial infarction. The main findings of this study
indicate that:
1. Undifferentiated murine ESC implanted into normal rat hearts developed into teratoma if the
animals were immunosuppressed with CsA. The morbidity increased with time from 20% to
100% at 4 weeks post-implantation. In contrast, in the absence of immunosuppression, an
extended granulation tissue was first observed and, thereafter, injected ESC could not be
retrieved, being most probably suppressed by the immune system.
2. The teratoma derived from the implanted ESC since those were positive for Bcl2 and GFP
ESC markers. They were not related to the type of ESC lines used, since different cell lines,
e.g., feeder cell-dependent D3 and feeder cell-independent CGR8-Bcl2 and CGR8-EFFP
gave similar results.
3. The type of cardiac injury (acute or chronic myocardial infarction) did not influence the
teratoma formation. Both infarcted and non-infarcted hearts could develop teratoma after
grafting of undifferentiated ESC when accompanied with immunosuppression.
4. When ESC-derived CMC were implanted in MI hearts we observed cells positive for cardiac
markers but not organized into mature fibers. The growth of other tissue types was also
observed.
5. Left ventricular functions were not modified by the transplantation of undifferentiated ESC
or ESC-derived CMC. Because of teratoma formation, it was impossible to observe a
reliable change (either improvement or worsening) of the cardiac functions.
That undifferentiated ESC have the ability to differentiate to all types of tissue upon
implantation it is a notion well established. However, the possibility to commit to the right
phenotype depending to which niche or organ they are exposed, this is definitely a new and
attractive concept and deserves further exploration. As first reported by Puceat et al, the
engraftment of murine ESC into chronic infarcted rat hearts leaded to their differentiation into
cardiomyocytes without the need of immunosuppression. This work proposed that the heart
might exert a TGFβ-mediated paracrine action on the implanted cells, especially under

55
conditions in which the heart undergoes a lesion such as MI. Under these conditions in fact, an
increased production of TGFβ has been reported in the literature. This could favor the mesoderm
layer development within ESC and commit them to cardiomyocytes. In addition, Puceat et al
demonstrated in vitro that ESC treated with TGFβ expressed increased amounts of cardiac
transcription factors, and more cells acquired a cardiac phenotype (Behfar et al., 2002). It is
therefore likely that cardiac injury could induce cardiac repair and thus cardiomyocyte
differentiation through TGFβ signaling.
Upon injury, several organs are able to release factors that activate repairing mechanisms and
induce resident stem cells, partially committed but not fully differentiated, to further progress
into their fate and replace damaged or dead cells with new units. Well-known examples are the
bone marrow, the skin, the liver and the skeletal muscle. This does not seem to be the case of the
brain that, despite possessing a population of neural stem cells, does not seem to be able to
activate enough brain stem cells upon a major injury or cell loss (ex. brain stroke, Parkinson's or
Alzheimer's diseases) (Cao et al., 2002). It is therefore conceivable that an healthy organ may
not be capable to locally release cues able to prime the implanted cells, while a damaged tissue
may be activated to release important molecules for both the recruitment of resident adult stem
cells when available, or distant stem cells from other compartments (i.e. the bone marrow). The
identification of those factors will therefore be fundamental to optimally initiate in vitro a suited
differentiation that could pursued in situ. One could also envisage to genetically inducing the
implanted cells to transiently secrete factors favoring a neovascularization, for example via the
insertion of VEGF (vascular endothelial growth factor) transgene under the control of an
inducible promoter. This would allow an optimal integration in the recipient organ an avoid
long-term deleterious effects of an uncontrolled long-term secretion (Springer et al., 1998;
Suzuki et al., 2001; Yang et al., 2002).
The timing of cell transplantation post-injury is like to play an important role because it
related to donor cell survival, homing and/or post transplantation proliferation. At early stages of
MI, the vascular and cellular responses of both acute and chronic inflammation reaction are
mediated by chemical factors derived from plasma or cells and triggered by inflammatory stimuli
such as cytokines. This should possibly affect donor cell survival. At later stages of MI, tissue
repairing had occurred; it was characterized by the replacement of injured/dead cells by
connective tissue, fibrosis, thus leaving a permanent scar. Hence, this situation may reduce the
positive effect of cell therapy. In our study, we compared ESC implanted at both of stages (acute
and chronic phase), and the results did not provide differences in cell survival or morphology
between different implantation timings.

56
A key point is surely the need of an immunosuppression treatment when mouse-to-rat
transplantation is envisaged. In our study, we used CsA as immunosuppressive agent,
continuously delivered by osmotic minipump, to avoid host versus donor graft rejection.
In our experimental conditions, we could not reproduce the data from Puceat et al, which did
not observe any ESC rejection by the host in the absence of immunosuppression. In contrast, we
showed that the cells did not survive if the immune system was not depressed by CsA. Soon or
later the engrafted cells turned into tumors, as compared to control conditions.
Only one study reported the occurrence of teratoma when undifferentiated murine ESC were
directly injected into the knee joint or the subcutaneous space on the back of SCID mice
(Wakitani et al., 2003) . The tumors grew and destroyed the joints.
Impede the rejection of the implanted cells by the recipient remains a barrier to be solved. In
fact, immunosuppressive drugs are associated with many highly unpleasant side effects and such
a treatment would not represent an optimally acceptable option. Interestingly, ESC seems to
express less immune-related cell surface proteins (e.g., class I products of the major
histocompatibility complex) (O'Shea, 1999). The group of Benvenisty et al. (Drukker et al.,
2002) addressed the graft rejection issue of cells derived from hESC by showing that both
undifferentiated or differentiated hESC express no MHC-II proteins or HLA-G and very low
levels of MHC class I (MHC-I) proteins on their surface. MHC-I molecules, however, may be
dramatically and rapidly induced by treating the cells with interferons. If a similar phenomenon
will be observed after transplantation, allogeneic human ESC might be rejected by cytotoxic T
lymphocytes.
It is likely that the problem of rejection of grafted human ES-derived cells could be overcome
(or at least minimized) by establishing "histocompatibility banks" of hESC with completely
HLA-typed ESC clones derived using Good Manufacture Practice (GMP) protocols. Ideally, if
large numbers of cell lines from genetically diverse populations can be maintained, this would
provide isotype-matching cells for virtually any patient. Other possibilities include means of
reducing or abolishing cell immunogenicity. ESC, unlike adult cells, can be easily modified
genetically by, for example, inserting immunosuppressive molecules such as Fas ligand, or
removing immunoactive proteins such as B7 antigens (Walker et al., 1997). Alternatively, one
could delete the foreign major histocompatibility complex (MHC) genes or insert genes coding
for the recipient's MHC (Westphal and Leder, 1997). Ultimately, neoplastic growth or
immunopathology could be suppressed by introducing into ESC before implantation suicide
genes that permit their ablation in case of misbehavior. For example, herpes thymidine kinase

57
sensitizes mouse ESC to destruction by the guanosine analog gancyclovir (Fareed and Moolten,
2002).
Total immunocompatability of tissue engineered from human ESC (Cibelli et al., 2002; Lanza
et al., 1999a; Lanza et al., 1999b; Solter and Gearhart, 1999) could be theoretically obtained by
somatic nuclear transfer (also defined as therapeutical cloning). This procedure uses the transfer
of a somatic cell nucleus from an individual into an enucleated oocyte (Wilmut et al., 1997;
Wilmut et al., 1998). Such an oocyte would then undergo embryonic development to the
blastocyst stage prior to the isolation from the inner cell mass of hESC that would be genetically
matched to the tissues of the nucleus donor. So far, one group has claimed the derivation by
nuclear transfer of a human embryo up to a six-cell stage only (Cibelli et al., 2002), but the
success of this result is still questioned. Clearly, this procedure of somatic nuclear transfer is still
highly problematic from an ethical and practical point of view.
The implantation of pure population of cells correctly committed to the suitable phenotype,
i.e. the cardiac one, seems to be a critical issue that remain to be solved.
Up to date, several studies have reported ESC-derived CMC engrafted into host myocardium.
Li et al (1996) has reported successfully transplanted cardiomyocytes isolated from fetal rat heart
into cryoinjured myocardial scar tissue, and improved heart function. Min et al (Min et al., 2002)
showed that the implantation in rats of clusters of beating mouse cardiomyocytes dissected from
EBs and labelled with GFP within 30 minutes after induction of myocardial infarction resulted in
stable intramyocardial grafts and improve cardiac function in postinfarcted failing hearts. These
authors did not discuss about immunoreactivity, rejection and teratoma formation, although some
area of undifferentiated cells were observed in the graft. We performed the same type of
experiments and we observed that engrafted CMC clusters neither terminally do not differentiate,
nor they organized into connected myofibers.

58
G. CONCLUSIONS
The clinical application of cell therapy in CHF requires a source of grafting cells providing
the following properties: i) the possibility to derive them easily; ii) the availability to propagate
to such a big mount of cell number; iii) the possibility to survive in the micro-circumstance of
the heart and to integrate with the host cell; vi) finally it should restore and improve the function
of a damaged heart.
Successful use of any stem cell-based therapy will eventually depend on our ability to isolate
specific cell types in large numbers that will differentiate to a fully functional state, as well as on
the challenging demonstration of their in vivo function. To this end, it is crucial to pursue basic
research on human ESC biology to ensure better understanding of basic principles, what genes
and proteins are essential for hES developmental progression. This comprehension does not only
specifically involve the transplantation of ESC-derived cells but will also help the development
of new therapeutic strategies to improve the transdifferentiation and expansion of adult stem
cells such as umbilical cord blood cells or bone marrow cells. Many hurdles (not only technical
but also ethical) have to be cleared before the research reaches a point where clinical trials can
begin.
Although ESC would bring several advantages for cell therapy strategies due to their great
differentiation potential, availability and expansion capacity in culture, a great attention has to be
paid to problems of growth control. The development of teratoma has to be avoided before to
envisage their use for transplantation. The use of ESC as a resource of cell therapy of CHF
remain a promising idea, but there is still a long way to go from basic research to the clinic
application, carefully attention must be paid to avoid deleterious effects.

59
H. IMPLEMENTATION AND FUTURE EXPERIMENTS
The present goals are to:
1. Improve the in vitro differentiation of ESC into cardiomyocytes using specific growth
factors.
2. Optimize the methods for the isolation of ESC-derived cardiomyocytes in order to obtain a
population as pure as possible and avoid contamination with undifferentiated cells or cells
committed to other lineages.
3. Develop new methods of cell engraftment. In particular we aim at the production of 3D-
cardiac tissue in vitro containing cardiomyocytes. Those would probably result more
appropriate in order to achieve a better engraftment.
4. To develop imaging tools such as magnetic resonance imaging (MRI) in order to monitor the
implanted cells in vivo after a labeling with nanoparticles or iron oxide. MRI will allow to
follow cell fate in vivo and measure cardiac functions in a non invasive way.

60
I. REPRINTS
He Q, Li J, Bettiol E, Jaconi ME, Embryonic Stem Cells: New Possible Therapy forDegenerative Diseases That Affect Elderly People. J Gerontol A Biol Sci Med Sci2003;58(3):M279-287.

61
J. ACKNOLEDGMENTS
I would like to thank all the persons who did help me during these last two years. Without
their support I would never had finished my research work.
First of all, I was very pleased to work with Profession Karl-Heinz Krause and Dr. Marisa
Jaconi, under the direction of whom I could finish my thesis work. I thank them for always
giving me a positive support.
I would like to thank Profession Zhang, Zhong-hui, the president of Beijing Hospital, Beijing,
for permitting my on-leave work abroad for two years, to complete the M.D. program.
I also thank the dean of the Geriatric Department, Prof. Jean-Pierre Michel, who allowed me
to do welcomed me in this Department and allowed me to perform some clinical consultations.
I thank Profession Denis Morel and his technician Manuel Costa for helping me to built up
the rat animal model of myocardial infarction and to assist me with animal experiments;
A warm thank you also to :
- Dr. Jian Li and Esther Bettiol, working with me in the laboratory and helping me with the
cell culture work,
- Dr Michel Dubois-Dauphin and Dr. Michael Stumm who unravel to me all the secrets of
immunohistochemistry,
- Dr. Pedro Trigo Trindade who performed the echocardiograms,
- Dr. François Herrmann who did the statistical study,.
- Dr. Jean-Jecques Perrenound, with whom I did cardiology consultations at the in Geriatric
Department during my first year. He was so nice to me and gave me many advice from the
clinical point of view
- Ms. Hélène Gfeller-Tillmann, our secretary; Mr. Olivier Plastre, lab technician and all of
others persons in our laboratory for their help and for being very kind to share their
experience with me.

62
K. REFERENCES
Anversa, P., J. Kajstura, B. Nadal-Ginard, and A. Leri. 2003. Primitive Cells and Tissue Regeneration. Circ Res.92:579-582.
Atkins, B.Z., M.T. Hueman, J. Meuchel, K.A. Hutcheson, D.D. Glower, and D.A. Taylor. 1999. Cellularcardiomyoplasty improves diastolic properties of injured heart. J Surg Res. 85:234-42.
Beauchamp, J.R., J.E. Morgan, C.N. Pagel, and T.A. Partridge. 1999. Dynamics of Myoblast Transplantation Reveala Discrete Minority of Precursors with Stem Cell-like Properties as the Myogenic Source. J. Cell Biol.144:1113-1122.
Behfar, A., L.V. Zingman, D.M. Hodgson, J.M. Rauzier, G.C. Kane, A. Terzic, and M. Puceat. 2002. Stem celldifferentiation requires a paracrine pathway in the heart. Faseb J. 16:1558-66.
Beltrami, A.P., K. Urbanek, J. Kajstura, S.M. Yan, N. Finato, R. Bussani, B. Nadal-Ginard, F. Silvestri, A. Leri,C.A. Beltrami, and P. Anversa. 2001. Evidence that human cardiac myocytes divide after myocardialinfarction. N Engl J Med. 344:1750-7.
Bonnet, D. 2002. Haematopoietic stem cells. J Pathol. 197:430-40.Cao, Q., R.L. Benton, and S.R. Whittemore. 2002. Stem cell repair of central nervous system injury. J Neurosci Res.
68:501-10.Cibelli, J.B., R.P. Lanza, M.D. West, and C. Ezzell. 2002. The first human cloned embryo. Sci Am. 286:44-51.Dowell, J.D., M. Rubart, K.B. Pasumarthi, M.H. Soonpaa, and L.J. Field. 2003. Myocyte and myogenic stem cell
transplantation in the heart. Cardiovasc Res. 58:336-50.Drukker, M., G. Katz, A. Urbach, M. Schuldiner, G. Markel, J. Itskovitz-Eldor, B. Reubinoff, O. Mandelboim, and
N. Benvenisty. 2002. Characterization of the expression of MHC proteins in human embryonic stem cells.Proc Natl Acad Sci U S A. 99:9864-9.
Fareed, M.U., and F.L. Moolten. 2002. Suicide gene transduction sensitizes murine embryonic and humanmesenchymal stem cells to ablation on demand-- a fail-safe protection against cellular misbehavior. GeneTher. 9:955-62.
Grounds, M.D., J.D. White, N. Rosenthal, and M.A. Bogoyevitch. 2002. The role of stem cells in skeletal andcardiac muscle repair. J Histochem Cytochem. 50:589-610.
Hagege, A.A., C. Carrion, P. Menasche, J.T. Vilquin, D. Duboc, J.P. Marolleau, M. Desnos, and P. Bruneval. 2003.Viability and differentiation of autologous skeletal myoblast grafts in ischaemic cardiomyopathy. Lancet.2361:491-2.
Hamano, K., M. Nishida, K. Hirata, A. Mikamo, T.S. Li, M. Harada, T. Miura, M. Matsuzaki, and K. Esato. 2001.Local implantation of autologous bone marrow cells for therapeutic angiogenesis in patients with ischemicheart disease: clinical trial and preliminary results. Jpn Circ J. 65:845-7.
Holden, C., and G. Vogel. 2002. STEM CELLS: Plasticity: Time for a Reappraisal? Science. 296:2126-2129.Hutcheson, K.A., B.Z. Atkins, M.T. Hueman, M.B. Hopkins, D.D. Glower, and D.A. Taylor. 2000. Comparison of
benefits on myocardial performance of cellular cardiomyoplasty with skeletal myoblasts and fibroblasts.Cell Transplant. 9:359-68.
Itescu, S., M.D. Schuster, and A.A. Kocher. 2003. New directions in strategies using cell therapy for heart disease. JMol Med. 81:288-96.
Jaconi, M., C. Bony, S.M. Richards, A. Terzic, S. Arnaudeau, G. Vassort, and M. Puceat. 2000. Inositol 1,4,5-trisphosphate directs Ca(2+) flow between mitochondria and the Endoplasmic/Sarcoplasmic reticulum: arole in regulating cardiac autonomic Ca(2+) spiking. Mol Biol Cell. 11:1845-58.
Jain, M., H. DerSimonian, D.A. Brenner, S. Ngoy, P. Teller, A.S. Edge, A. Zawadzka, K. Wetzel, D.B. Sawyer,W.S. Colucci, C.S. Apstein, and R. Liao. 2001. Cell therapy attenuates deleterious ventricular remodelingand improves cardiac performance after myocardial infarction. Circulation. 103:1920-7.
Johkura, K., L. Cui, A. Suzuki, R. Teng, A. Kamiyoshi, S. Okamura, S. Kubota, X. Zhao, K. Asanuma, Y. Okouchi,N. Ogiwara, Y. Tagawa, and K. Sasaki. 2003. Survival and function of mouse embryonic stem cell-derivedcardiomyocytes in ectopic transplants. Cardiovasc Res. 58:435-43.
Kamihata, H., H. Matsubara, T. Nishiue, S. Fujiyama, Y. Tsutsumi, R. Ozono, H. Masaki, Y. Mori, O. Iba, E.Tateishi, A. Kosaki, S. Shintani, T. Murohara, T. Imaizumi, and T. Iwasaka. 2001. Implantation of bonemarrow mononuclear cells into ischemic myocardium enhances collateral perfusion and regional functionvia side supply of angioblasts, angiogenic ligands, and cytokines. Circulation. 104:1046-52.
Klug, M.G., M.H. Soonpaa, G.Y. Koh, and L.J. Field. 1996. Genetically Selected Cardiomyocytes fromDifferentiating Embryonic Stem Cells Form Stable Intracardiac Grafts. J. Clin. Invest. 98:216-224.
Krause, D.S., N.D. Theise, M.I. Collector, O. Henegariu, S. Hwang, R. Gardner, S. Neutzel, and S.J. Sharkis. 2001.Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell. 105:369-77.

63
Laham, R.J., and P. Oettgen. 2003. Bone marrow transplantation for the heart: fact or fiction? The Lancet. 361:11-12.
Lanza, R.P., J.B. Cibelli, and M.D. West. 1999a. Human therapeutic cloning. Nat Med. 5:975-7.Lanza, R.P., J.B. Cibelli, and M.D. West. 1999b. Prospects for the use of nuclear transfer in human transplantation.
Nat Biotechnol. 17:1171-4.Leor, J., M. Patterson, M.J. Quinones, L.H. Kedes, and R.A. Kloner. 1996. Transplantation of fetal myocardial
tissue into the infarcted myocardium of rat. A potential method for repair of infarcted myocardium?Circulation. 94:II332-6.
Li, R.K., Z.Q. Jia, R.D. Weisel, F. Merante, and D.A. Mickle. 1999. Smooth muscle cell transplantation intomyocardial scar tissue improves heart function. J Mol Cell Cardiol. 31:513-22.
Li, R.K., Z.Q. Jia, R.D. Weisel, D.A. Mickle, J. Zhang, M.K. Mohabeer, V. Rao, and J. Ivanov. 1996.Cardiomyocyte transplantation improves heart function. Ann Thorac Surg. 62:654-60; discussion 660-1.
Li, R.-K., R. Weisel, D. Mickle, Z.-Q. Jia, E.-J. Kim, T. Sakai, S. Tomita, L. Schwartz, M. Iwanochko, M. Husain,R. Cusimano, R. Burns, and T. Yau. 2000. Autologous porcine heart cell transplantation improved heartfunction after a myocardial infarction. Journal of Thoracic and Cardiovascular Surgery. 119:62-68.
Litwin, S., S. Katz, J. Morgan, and P. Douglas. 1994. Serial echocardiographic assessment of left ventriculargeometry and function after large myocardial infarction in the rat. Circulation. 89:345-354.
MacLellan, W.R., and M.D. Schneider. 2000. Genetic dissection of cardiac growth control pathways. Annu RevPhysiol. 62:289-319.
Makino, S., K. Fukuda, S. Miyoshi, F. Konishi, H. Kodama, J. Pan, M. Sano, T. Takahashi, S. Hori, H. Abe, J. Hata,A. Umezawa, and S. Ogawa. 1999. Cardiomyocytes can be generated from marrow stromal cells in vitro. JClin Invest. 103:697-705.
Maltsev, V.A., A.M. Wobus, J. Rohwedel, M. Bader, and J. Hescheler. 1994. Cardiomyocytes differentiated in vitrofrom embryonic stem cells developmentally express cardiac-specific genes and ionic currents. Circ Res.75:233-44.
Martin, G.R. 1981. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned byteratocarcinoma stem cells. Proc Natl Acad Sci U S A. 78:7634-8.
Menasche, P. 2003. Skeletal muscle satellite cell transplantation. Cardiovasc Res. 58:351-7.Menasche, P., A.A. Hagege, J.-T. Vilquin, M. Desnos, E. Abergel, B. Pouzet, A. Bel, S. Sarateanu, M. Scorsin, and
K. Schwartz. 2003. Autologous skeletal myoblast transplantation for severe postinfarction left ventriculardysfunction. Journal of the American College of Cardiology. 41:1078-1083.
Meyer, N., M. Jaconi, A. Landopoulou, P. Fort, and M. Puceat. 2000. A fluorescent reporter gene as a marker forventricular specification in ES-derived cardiac cells. FEBS Lett. 478:151-8.
Min, J.-Y., Y. Yang, K.L. Converso, L. Liu, Q. Huang, J.P. Morgan, and Y.-F. Xiao. 2002. Transplantation ofembryonic stem cells improves cardiac function in postinfarcted rats. J Appl Physiol. 92:288-296.
Moises, V.A., R.L. Ferreira, E. Nozawa, R.M. Kanashiro, O. Campos, J.L. Andrade, A.C. Carvalho, and P.J. Tucci.2000. Structural and functional characteristics of rat hearts with and without myocardial infarct. Initialexperience with Doppler echocardiography. Arq Bras Cardiol. 75:125-36.
Mummery, C., D. Ward-van Oostwaard, P. Doevendans, R. Spijker, S. van den Brink, R. Hassink, M. van derHeyden, T. Opthof, M. Pera, A.B. de la Riviere, R. Passier, and L. Tertoolen. 2003. Differentiation ofHuman Embryonic Stem Cells to Cardiomyocytes: Role of Coculture With Visceral Endoderm-Like Cells.Circulation. 107:2733-2740.
Murry, C.E., R.W. Wiseman, S.M. Schwartz, and S.D. Hauschka. 1996. Skeletal myoblast transplantation for repairof myocardial necrosis. J Clin Invest. 98:2512-23.
Nir, S.G., R. David, M. Zaruba, W.-M. Franz, and J. Itskovitz-Eldor. 2003. Human embryonic stem cells forcardiovascular repair. Cardiovascular Research. 58:313-323.
Niwa, H. 2001. Molecular mechanism to maintain stem cell renewal of ES cells. Cell Struct Funct. 26:137-48.Orlic, D., J. Kajstura, S. Chimenti, I. Jakoniuk, S.M. Anderson, B. Li, J. Pickel, R. McKay, B. Nadal-Ginard, D.M.
Bodine, A. Leri, and P. Anversa. 2001. Bone marrow cells regenerate infarcted myocardium. Nature.410:701-5.
O'Shea, K.S. 1999. Embryonic stem cell models of development. Anat Rec. 257:32-41.Perin, E.C., H.F.R. Dohmann, R. Borojevic, S.A. Silva, A.L.S. Sousa, C.T. Mesquita, M.I.D. Rossi, A.C. Carvalho,
H.S. Dutra, H.J.F. Dohmann, G.V. Silva, L. Belem, R. Vivacqua, F.O.D. Rangel, R. Esporcatte, Y.J. Geng,W.K. Vaughn, J.A.R. Assad, E.T. Mesquita, and J.T. Willerson. 2003. Transendocardial, Autologous BoneMarrow Cell Transplantation for Severe, Chronic Ischemic Heart Failure. Circulation. 107:2294-2302.
Pouzet, B., J.-T. Vilquin, A.A. Hagege, M. Scorsin, E. Messas, M. Fiszman, K. Schwartz, and P. Menasche. 2000.Intramyocardial Transplantation of Autologous Myoblasts : Can Tissue Processing Be Optimized?Circulation. 102:210III--215.
Preston, S.L., M.R. Alison, S.J. Forbes, N.C. Direkze, R. Poulsom, and N.A. Wright. 2003. The new stem cellbiology: something for everyone. Mol Pathol. 56:86-96.
Reffelmann, T., and R.A. Kloner. 2003. Cellular cardiomyoplasty-cardiomyocytes, skeletal myoblasts, or stem cellsfor regenerating myocardium and treatment of heart failure? Cardiovasc Res. 58:358-68.

64
Reinecke, H., V. Poppa, and C.E. Murry. 2002. Skeletal muscle stem cells do not transdifferentiate intocardiomyocytes after cardiac grafting. J Mol Cell Cardiol. 34:241-9.
Reinecke, H., M. Zhang, T. Bartosek, and C.E. Murry. 1999. Survival, Integration, and Differentiation ofCardiomyocyte Grafts : A Study in Normal and Injured Rat Hearts. Circulation. 100:193-202.
Robinson, S.W., P.W. Cho, H.I. Levitsky, J.L. Olson, R.H. Hruban, M.A. Acker, and P.D. Kessler. 1996. Arterialdelivery of genetically labelled skeletal myoblasts to the murine heart: long-term survival and phenotypicmodification of implanted myoblasts. Cell Transplant. 5:77-91.
Scorsin, M., and L.C. Souza. 2001. Cellular transplantation for the treatment of heart failure. State of the art. ArqBras Cardiol. 77:103-6.
Sola, O.M., D.H. Dillard, T.D. Ivey, K. Haneda, T. Itoh, and R. Thomas. 1985. Autotransplantation of skeletalmuscle into myocardium. Circulation. 71:341-8.
Solter, D., and J. Gearhart. 1999. Putting stem cells to work. Science. 283:1468-70.Springer, M.L., A.S. Chen, P.E. Kraft, M. Bednarski, and H.M. Blau. 1998. VEGF gene delivery to muscle:
potential role for vasculogenesis in adults. Mol Cell. 2:549-58.Stamm, C., B. Westphal, H.D. Kleine, M. Petzsch, C. Kittner, H. Klinge, C. Schumichen, C.A. Nienaber, M.
Freund, and G. Steinhoff. 2003. Autologous bone-marrow stem-cell transplantation for myocardialregeneration. Lancet. 361:45-6.
Stewart, C.L., I. Gadi, and H. Bhatt. 1994. Stem cells from primordial germ cells can reenter the germ line. DevBiol. 161:626-8.
Sullivan, P.G., M. Thompson, and S.W. Scheff. 2000. Continuous Infusion of Cyclosporin A PostinjurySignificantly Ameliorates Cortical Damage Following Traumatic Brain Injury. Experimental Neurology.161:631-637.
Suzuki, K., B. Murtuza, R.T. Smolenski, I.A. Sammut, N. Suzuki, Y. Kaneda, and M.H. Yacoub. 2001. Celltransplantation for the treatment of acute myocardial infarction using vascular endothelial growth factor-expressing skeletal myoblasts. Circulation. 104:I207-12.
Taylor, D.A. 2001. Cellular cardiomyoplasty with autologous skeletal myoblasts for ischemic heart disease andheart failure. Curr Control Trials Cardiovasc Med. 2:208-210.
Taylor, D.A., B.Z. Atkins, P. Hungspreugs, T.R. Jones, M.C. Reedy, K.A. Hutcheson, D.D. Glower, and W.E.Kraus. 1998. Regenerating functional myocardium: improved performance after skeletal myoblasttransplantation. Nat Med. 4:929-33.
Thomson, J.A., J. Itskovitz-Eldor, S.S. Shapiro, M.A. Waknitz, J.J. Swiergiel, V.S. Marshall, and J.M. Jones. 1998.Embryonic stem cell lines derived from human blastocysts. Science. 282:1145-7.
Tse, H.F., Y.L. Kwong, J.K. Chan, G. Lo, C.L. Ho, and C.P. Lau. 2003. Angiogenesis in ischaemic myocardium byintramyocardial autologous bone marrow mononuclear cell implantation. Lancet. 361:47-9.
Wakitani, S., K. Takaoka, T. Hattori, N. Miyazawa, T. Iwanaga, S. Takeda, T.K. Watanabe, and A. Tanigami. 2003.Embryonic stem cells injected into the mouse knee joint form teratomas and subsequently destroy the joint.Rheumatology. 42:162-165.
Walker, P.R., P. Saas, and P.Y. Dietrich. 1997. Role of Fas ligand (CD95L) in immune escape: the tumor cell strikesback. J Immunol. 158:4521-4.
Westphal, C.H., and P. Leder. 1997. Transposon-generated 'knock-out' and 'knock-in' gene-targeting constructs foruse in mice. Curr Biol. 7:530-3.
Wilmut, I., A.E. Schnieke, J. McWhir, A.J. Kind, and K.H. Campbell. 1997. Viable offspring derived from fetal andadult mammalian cells. Nature. 385:810-3.
Wilmut, I., L. Young, and K.H. Campbell. 1998. Embryonic and somatic cell cloning. Reprod Fertil Dev. 10:639-43.
Yang, Y., J.Y. Min, J.S. Rana, Q. Ke, J. Cai, Y. Chen, J.P. Morgan, and Y.F. Xiao. 2002. VEGF enhancesfunctional improvement of postinfarcted hearts by transplantation of ESC-differentiated cells. J ApplPhysiol. 93:1140-51.
Ye, J., L. Yang, R. Sethi, J. Copps, B. Ramjiawan, R. Summers, and R. Deslauriers. 1997. A new technique ofcoronary artery ligation: experimental myocardial infarction in rats in vivo with reduced mortality. MolCell Biochem. 176:227-33.