the planar cell polarity pathway drives pathogenesis of...
TRANSCRIPT

Microenvironment and Immunology
The Planar Cell Polarity Pathway Drives Pathogenesis ofChronic Lymphocytic Leukemia by the Regulation ofB-Lymphocyte Migration
Mark�eta Kauck�a1, Karla Plevov�a2,4, Š�arka Pavlov�a2,4, Pavlína Janovsk�a1, Archana Mishra1, Jan Verner2,4,Ji�rina Proch�azkov�a1, Pavel Krej�cí1,5,6, Jana Kotaškov�a2,4, Petra Ovesn�a3, Boris Tich�y2,4, Yvona Brychtov�a4,Michael Doubek2,4, Alois Kozubík1,5, Ji�rí Mayer2,4, Š�arka Pospíšilov�a2,4, and Vít�ezslav Bryja1,5
AbstractThe planar cell polarity (PCP) pathway is a conserved pathway that regulates cell migration and polarity in
various contexts. Here we show that key PCP pathway components such as Vangl2, Celsr1, Prickle1, FZD3, FZD7,Dvl2, Dvl3, and casein kinase 1 (CK1)-e are upregulated in B lymphocytes of patients with chronic lymphocyticleukemia (CLL). Elevated levels of PCP proteins accumulate in advanced stages of the disease. Here, we show thatPCP pathway is required for the migration and transendothelial invasion of CLL cells and that patients with highexpression of PCP genes, FZD3, FZD7, and PRICKLE1, have a less favorable clinical prognosis. Our findingsestablish that the PCP pathway acts as an important regulator of CLL cell migration and invasion. PCP proteinsrepresent an important class of molecules regulating pathogenic interaction of CLL cells with their microen-vironment. Cancer Res; 73(5); 1491–501. �2012 AACR.
IntroductionWnt signaling pathways are crucial for cell-to-cell commu-
nication, differentiation, and morphogenesis in embryonicdevelopment. Dysfunction or deregulation of Wnt signalingaccounts for a number of developmental defects, inheriteddiseases, and many types of cancer (1). The best-known Wntpathway—canonical or Wnt/b-catenin pathway—inducesb-catenin–dependent activation of T-cell factor/lymphoidenhancer factor (TCF/LEF)-mediated transcription. However,Wnt ligands can activate other so-called noncanonical Wntpathways, which are b-catenin–independent and biochemi-cally distinct from the canonical Wnt pathway (2). Amongthose, the pathway that regulates planar cell polarity (Wnt/PCP pathway) is the best known (3). The core components ofthe Wnt/PCP pathway (referred here as PCP or polarity
proteins) are conserved in evolution and include Vang-likeprotein 2 (Vangl2), cadherin EGF LAG 7-pass transmembraneG-type receptor (Celsr1), Frizzleds (FZD, in mammals mainlyFZD3, FZD6, and FZD7), which act as receptors, and cyto-plasmic components such as Disheveled (Dvl1-3 in mammals),Prickle-like proteins (mainly Prickle1), casein kinase 1e (CK1e),and the small GTPases Rho/Rac and downstream kinasesROCK/JNK. PCP proteins regulate cell polarity (3), convergentextension movements during gastrulation and neurulation,and polarity of the hair cells in the inner ear (3, 4).
In the present study, we investigated the role of PCP proteinsin chronic lymphocytic leukemia (CLL). CLL is characterizedby clonal expansion and apoptosis dysregulation of CD5þ Blymphocytes. Neoplastic CLL cells accumulate in blood, bonemarrow, and lymphoid tissue. It is recognized that cell migra-tion and recirculation between peripheral blood and lymphoidniches in bone marrow and lymphoid tissue are importantfactors contributing to CLL biology (5–7) and that alterationsin survival and proliferation of CLL are affected by theirinteraction with the microenvironment. Here, we show for thefirst time that the core cassette of the PCP pathway composedof Celsr1, Prickle1, Vangl2, Dvl2, Dvl3, FZD3, and FZD7 isupregulated in B cells of patients with CLLs. We show thatPCP proteins, which are conserved regulators of the cell–cellinteraction and cell migration, are importantmediators of CLLchemotactic responses and homing.
Materials and MethodsCell isolation, purification, and cell treatments
Primary B cells from previously untreated patients withCLLs or healthy volunteers were separated using gradientcentrifugation followed by non–B-cell depletion (RosetteSepB Cell Enrichment Kit and Human CD3þ Depletion Kit;
Authors' Affiliations: 1Institute of Experimental Biology, Faculty of Sci-ence, 2CEITEC - Central European Institute of Technology, 3Institute ofBiostatistics and Analyses, Masaryk University; 4Center of MolecularBiology and Gene Therapy, Department of Internal Medicine–Hematologyand Oncology, University Hospital Brno andMedical Faculty MU; 5Depart-ment of Cytokinetics, Institute of Biophysics, Academy of Sciences of theCzech Republic, Brno, Czech Republic; and 6Medical Genetics Institute,Cedars-Sinai Medical Center, Los Angeles, California
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
K. Plevov�a and Š. Pavlov�a contributed equally to this work.
Corresponding Author: Vitezslav Bryja, Institute of Biophysics v.v.i.,Academy of Sciences of the Czech Republic, Kralovopolska 135, BrnoCZ-612 65, Czech Republic. Phone: 420-549493291; Fax: 420-541211214; E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-12-1752
�2012 American Association for Cancer Research.
CancerResearch
www.aacrjournals.org 1491
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752

StemCell Technologies; orMACSB cell IsolationKit II;MiltenyiBiotec) according to the manufacturer's instructions. Flowcytometry was conducted for evaluation of CD5 and CD19expression on purified cells (tricolor panel: CD45-TRI-COLOR,MHCD45065, Invitrogen, CD5-FITC, A08932, CD19-PE, A07769,Beckman Coulter). For further analyses, samples with thepurity higher than 95% B cells were used. The cell line MEC1was obtained from DSMZ, cell line was tested by DSMZ usingmolecular geneticmethods;MEC1 cells were cultured inRPMI-1640 supplemented with 10% FBS and antibiotics at 37oC and5% CO2.
Transwell assayThe chemotaxis assay was conducted in HTS Transwell-96
well plates (Corning Incorporated) with 5.0-mm pore sizepolycarbonate membranes following the manufacturer'sinstructions. A total of 0.5 � 106 cells (primary CLL or MEC1cells) were seeded in the Transwell upper insert, which waseither nontreated or coated with a human umbilical veinendothelial cells monolayer (HUVEC). Cells in the insert weretreated with antibodies or inhibitors as described in Supple-mentary Material. Cells were incubated overnight in full medi-um (including 10% fetal calf serum) at 37oC and 5% CO2, andafter 18 hours the migration toward the chemokine wasanalyzed by a Coulter Counter (model FN, CoulterElectronics; Figs. 2A–C, 3A–D, and 4A–C; Supplementary Figs.S2B and S4E–S4H) or by C6 flow cytometer (Accuri; Figs. 2E–G, 3E–G, and 4D; Supplementary Fig. S2E). Themigration indexwas calculated as the number of cells (treated or untreated)migrating in response to the chemokine divided by the numberof cells migrating toward the control medium only. In case ofvery variable migration indexes (in primary CLL cells), cellnumbers were normalized to the chemokine-only conditionand expressed as "relative migration."
In vivo experimentsNonirradiated mice (8- to 16-week-old) nonobese diabetic/
severe combined immunodeficient (NOD/SCID) IL2Rg-null(NSG)micewere used for transplantation experiments. Freshlyisolated human CLL lymphocytes (25 � 106 per condition)were pretreated as indicated for 20 hours and stained withCalcein AM. Mice underwent transplantation by intraperito-neal injection of 20� 106 of pretreated and calcein AM–stainedhuman CLL lymphocytes in 120 mL of sterile PBS. Recipientmice were analyzed 24 hours after the injection by fluores-cence-activated cell-sorting (FACS) analysis (for details, seeSupplementary Material).
Further methods including information on patient samples,quantitative real-time (qRT)-PCR, Western blotting, flow cyto-metry, nucleofection, and statistics are provided in the Sup-plementary Material.
ResultsCore PCP pathway components are upregulated in CLLpatients
Our earliermicroarray expression analysis (8) suggested thatcomponents of Wnt/PCP signaling are frequently upregulatedin CLLs and expressed differently in prognostically distinct
subsets of patients with CLLs. To confirm that the levels ofPCP components are indeed altered in CLLs, we collectedRNA samples from purified peripheral blood B cells (CD19þcell population purity � 95%) of control individuals (n ¼ 6)and patients with CLL (n ¼ 93). qRT-PCR analysis showedstatistically increased mRNA levels of 3 PCP genes, CELSR1,PRICKLE1, and FZD7, in CLL samples and identified that 2other genes VANGL2 and FZD3 have high levels in a subsetof patients with CLLs [Fig. 1A—normalized to the expressionof b2-microglobulin; Supplementary Fig. S1A—normalized tob-actin].
Subsequent analysis of the protein level byWestern blottingconfirmed increased protein expression of the PCP proteinsVangl2, Dvl2, Dvl3, Prickle1, and CK1e (Fig. 1B, quantifiedin Fig. 1C) in patients with CLLs. The level of b-catenin didnot differ between CLLs and healthy cells (Fig. 1B), and wefailed to detect activated (dephosphorylated) b-catenin (notshown), which suggest that the Wnt/b-catenin pathway isnot active in CLL cells. All CLL samples tested had high ex-pression of LEF1 and Ror1 (Fig. 1B), which were previouslyidentified as abundantly and consistently expressed in CLLcells (9, 10). Flow cytometry detection (Fig. 1D) further con-firmed increased cell surface levels of Ror1, FZD3, Celsr1, andVangl2 in comparison to healthy B cells.
Next, we asked whether the levels of PCP proteins arestable or they change during the disease course. When wecompared samples of 6 patients with CLLs collected at 2different time points (t1 and t2; Fig. 1E), we were able todetect changes in the levels of PCP proteins in CLL cells fromindividual time points. Specifically, we observed a clearincrease in Dvl2, Dvl3, and Prickle1 levels in patients fol-lowed in time, which is quantified in Supplementary Fig.S1B. The most prominent changes were observed mainly inpatients, who progressed from Rai stages (11) 0/I/II to thestage III or IV (see patients C, H, P). The levels of LEF1 andb-catenin do not change (Fig. 1E).
In summary, using several methods, we provide evidencethat a host of PCP proteins are overexpressed in B cells ofpatientswithCLLs, with some accumulating to a greater extentin advanced stages of the disease.
Wnt5a, a PCP ligand, increasesmigrationof primary CLLcells in a chemokine gradient via a Wnt/PCP pathway
It is becoming increasingly evident that the role of micro-environment in the pathogenesis of CLLs is of a crucialimportance (7, 12). Chemokines and their receptors, mainlyCXCL12-CXCR4, CXCL9/10/11-CXCR3, and CCL19/21-CCR7,have been shown tomediate invasiveness and transendothelialmigration of CLL cells (13–16). In that line, it has been recentlyshown that Wnt-5a, which is the major ligand for Ror1/2receptors and a crucial component of the Wnt/PCP pathway(17–19), promotes cell polarization and directional movementof melanoma cells and T cells in a chemokine (CXCL12)gradient (20, 21).
We thus hypothesized thatWnt5a, which is also expressed inCLL cells (Supplementary Fig. S2A) together with receptor PCPprotein complexes,may contribute to the pathogenesis of CLLsby the effects on the polarized migration of CLL cells toward
Kauck�a et al.
Cancer Res; 73(5) March 1, 2013 Cancer Research1492
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752
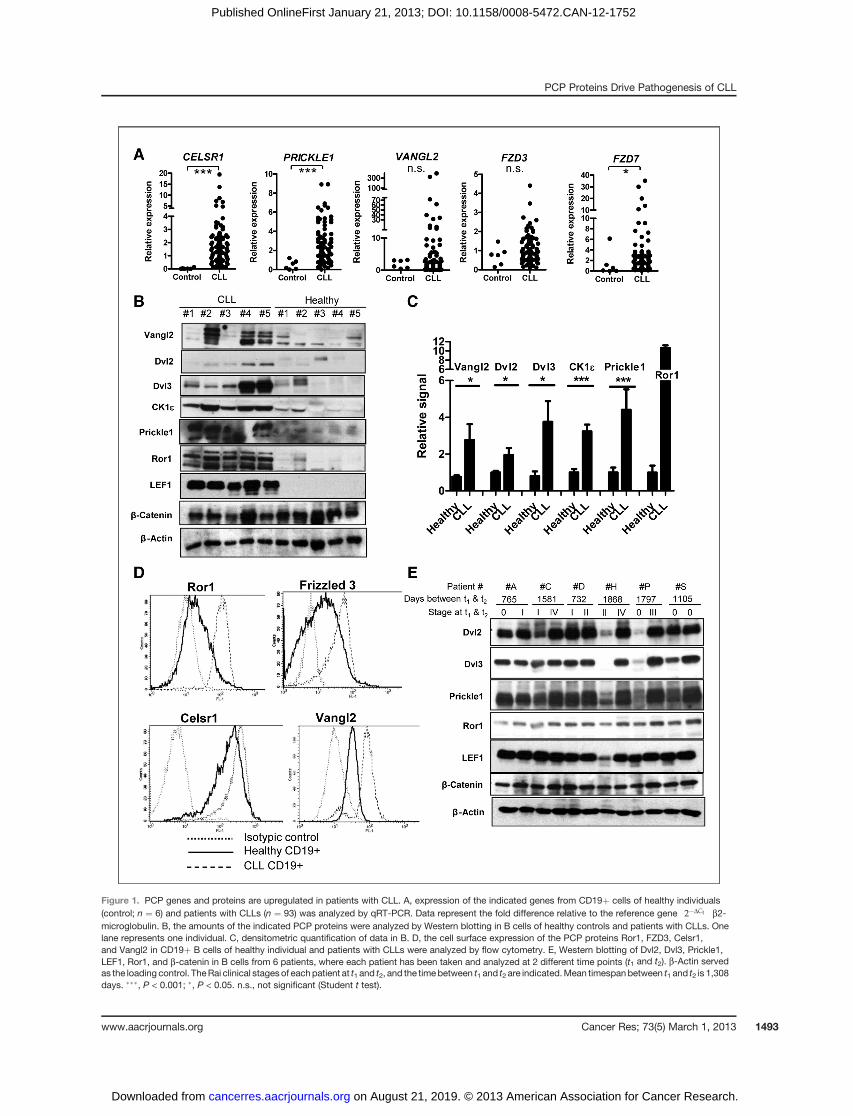
Figure 1. PCP genes and proteins are upregulated in patients with CLL. A, expression of the indicated genes from CD19þ cells of healthy individuals(control; n ¼ 6) and patients with CLLs (n ¼ 93) was analyzed by qRT-PCR. Data represent the fold difference relative to the reference gene 2�DCt
� �b2-
microglobulin. B, the amounts of the indicated PCP proteins were analyzed by Western blotting in B cells of healthy controls and patients with CLLs. Onelane represents one individual. C, densitometric quantification of data in B. D, the cell surface expression of the PCP proteins Ror1, FZD3, Celsr1,and Vangl2 in CD19þ B cells of healthy individual and patients with CLLs were analyzed by flow cytometry. E, Western blotting of Dvl2, Dvl3, Prickle1,LEF1, Ror1, and b-catenin in B cells from 6 patients, where each patient has been taken and analyzed at 2 different time points (t1 and t2). b-Actin servedas the loading control. TheRai clinical stagesof eachpatient at t1 and t2, and the timebetween t1 and t2 are indicated.Mean timespanbetween t1 and t2 is 1,308days. ���, P < 0.001; �, P < 0.05. n.s., not significant (Student t test).
PCP Proteins Drive Pathogenesis of CLL
www.aacrjournals.org Cancer Res; 73(5) March 1, 2013 1493
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752
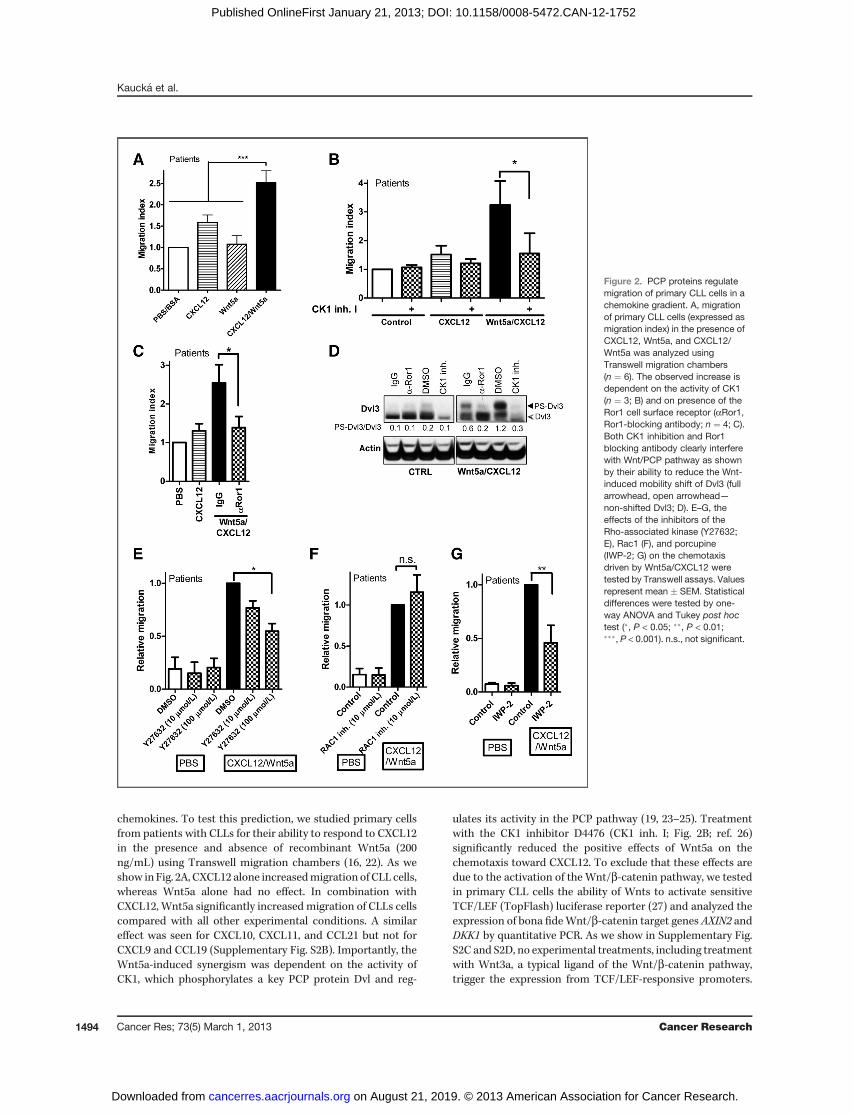
chemokines. To test this prediction, we studied primary cellsfrom patients with CLLs for their ability to respond to CXCL12in the presence and absence of recombinant Wnt5a (200ng/mL) using Transwell migration chambers (16, 22). As weshow in Fig. 2A, CXCL12 alone increasedmigration of CLL cells,whereas Wnt5a alone had no effect. In combination withCXCL12, Wnt5a significantly increased migration of CLLs cellscompared with all other experimental conditions. A similareffect was seen for CXCL10, CXCL11, and CCL21 but not forCXCL9 and CCL19 (Supplementary Fig. S2B). Importantly, theWnt5a-induced synergism was dependent on the activity ofCK1, which phosphorylates a key PCP protein Dvl and reg-
ulates its activity in the PCP pathway (19, 23–25). Treatmentwith the CK1 inhibitor D4476 (CK1 inh. I; Fig. 2B; ref. 26)significantly reduced the positive effects of Wnt5a on thechemotaxis toward CXCL12. To exclude that these effects aredue to the activation of the Wnt/b-catenin pathway, we testedin primary CLL cells the ability of Wnts to activate sensitiveTCF/LEF (TopFlash) luciferase reporter (27) and analyzed theexpression of bona fideWnt/b-catenin target genes AXIN2 andDKK1 by quantitative PCR. As we show in Supplementary Fig.S2C and S2D, no experimental treatments, including treatmentwith Wnt3a, a typical ligand of the Wnt/b-catenin pathway,trigger the expression from TCF/LEF-responsive promoters.
Figure 2. PCP proteins regulatemigration of primary CLL cells in achemokine gradient. A, migrationof primary CLL cells (expressed asmigration index) in the presence ofCXCL12, Wnt5a, and CXCL12/Wnt5a was analyzed usingTranswell migration chambers(n ¼ 6). The observed increase isdependent on the activity of CK1(n ¼ 3; B) and on presence of theRor1 cell surface receptor (aRor1,Ror1-blocking antibody; n ¼ 4; C).Both CK1 inhibition and Ror1blocking antibody clearly interferewith Wnt/PCP pathway as shownby their ability to reduce the Wnt-induced mobility shift of Dvl3 (fullarrowhead, open arrowhead—non-shifted Dvl3; D). E–G, theeffects of the inhibitors of theRho-associated kinase (Y27632;E), Rac1 (F), and porcupine(IWP-2; G) on the chemotaxisdriven by Wnt5a/CXCL12 weretested by Transwell assays. Valuesrepresent mean � SEM. Statisticaldifferences were tested by one-way ANOVA and Tukey post hoctest (�, P < 0.05; ��, P < 0.01;���, P < 0.001). n.s., not significant.
Kauck�a et al.
Cancer Res; 73(5) March 1, 2013 Cancer Research1494
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752

This strongly suggests that Wnt/b-catenin pathway is notinvolved in the observed phenotypes.The effects similar to the inhibition of CK1 have been
observed, when we blocked the putative Wnt5a receptor inthe PCP complex, Ror1, with the goat Ror1 blocking antibody(aRor1) but not with the control goat IgG (Fig. 2C). Theefficiency of this antibody, which binds Ror1 and promotesits internalization, has been shown earlier (28). Both D4476(CK1 inh. I) and aRor1 antibody clearly inhibited activation ofPCP pathway in primary CLL cells as shown by their ability toblock phosphorylation-dependent shift of endogenous Dvl3(PS-Dvl3), which is a hallmark of Dvl3 activation in the non-canonical Wnt pathway (Fig. 2D; ref. 19).In the Wnt/PCP pathway, Dvl mediates the activation of
small GTPases RhoA and Rac1, which trigger subsequent
cytoskeletal rearrangements and cell migration. As we showin Fig. 2E, the inhibition of RhoA-driven signaling by Y27632,an inhibitor of Rho-associated ROCK protein kinase (29) butnot the inhibition of Rac1 by a Rac1 inhibitor NSC23766(Fig. 2F; ref. 30), blocked the migration of primary CLL cells.No inhibitors used in this section affect the viability of prim-ary CLL cells, as assessed by WST1 test, and the effects onchemotaxis thus do not reflect differences in the condition ofCLL cells (Supplementary Fig. S3).
The experiments above suggest that PCP pathway iscapable to promote and is required for the efficient chemo-tactic migration of primary CLL cells. To further elaborateon the mechanism of the interaction between Wnt/PCP andthe chemokine signaling, we tested functionally the role ofthe CXCR4 (the receptor for CXCL12) and the role of
Figure 3. Regulation ofMEC1 cellmigration byPCPproteins. A,migration ofMEC1 cellswas analyzed in presence ofCXCL12, CCL19, andCCL21 (200 ng/mL)with or without Wnt5a using Transwell migration assays. Wnt5a does not further promote migration driven by CXCL12, CCL19, and CCL21. B–G,MEC1 migration triggered by CCL21 and/or CCL19 chemokines is blocked by CK1 inhibitor D4476 (CK1 inh. I; 100 mmol/L; B), PF670462 (CK1 inh. II;50 mmol/L; C), knockdown of Dvl2, the efficiency of knockdown was determined by Western blot analysis and quantified by densitometry (Dvl2/actin; D),inhibition of the Rho-associated kinase (Y27632; E), and the inhibitor of endogenous Wnt production (inhibitor of porcupine IWP-2; G). The inhibitor ofRac1 (F) does not have an effect. Values represent mean� SEM. Statistical differences were tested by one-way ANOVA and Tukey post hoc test (�, P < 0.05;��, P < 0.01; ���, P < 0.001; n.s., not significant). Each experiment has been carried out in at least 3 independent replicates.
PCP Proteins Drive Pathogenesis of CLL
www.aacrjournals.org Cancer Res; 73(5) March 1, 2013 1495
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752

endogenous Wnts in CLL chemotaxis. Interestingly, CXCR4-blocking antibody was unable to inhibit CLL migration(Supplementary Fig. S2E), whereas the disruption of theendogenous Wnt production by the porcupine inhibitorIWP-2 (31) reduced CLL migration (Fig. 2G). Given the factthat CXCL12-CXCR4 signaling does not regulate Wnt-5aexpression and Wnts do not regulate expression of CXCR4in CLL cells (see Supplementary Fig. S2F), we conclude that(i) the effects do not take place at the level of transcription,(ii) that the autocrine stimulation by Wnts contributes to theCLL chemotaxis. and (iii) that the chemotactic response isnot critically mediated by CXCR4.
PCP protein Dvl and its kinase CK1« are required for themigration in the chemokine gradient
To study the role of Wnt/PCP pathway in CLL migration inmore detail and to overcome the natural variability amongpatients and limitations in the experimental manipulationwith primary CLL cells, we carried out further experimentsinMEC1 cells (32).MEC1 is awell-defined cell line derived froma patients with CLLs, which recapitulates many aspects of theCLL biology and is used as the transplantation model of CLL(33). Quantitative analysis of mRNA (WNT5A, CELSR1, PRICK-LE1, FZD3, FZD7; Supplementary Fig. S4A) and protein levels(Ror1, Vangl2, Dvl2, Dvl3, CK1e; Supplementary Fig. S4B)showed that MEC1 cells express most PCP genes in high levels,although the levels of Ror1 were relatively low (SupplementaryFig. S4B).
MEC1 cells were unable to respond to CXCL12 butresponded clearly to CCL19 and CCL21 in a dose-dependentmanner (Fig. 3A; Supplementary Fig. S4C). Interestingly,
treatment with Wnt5a did not further promote the promi-gratory effect of chemokines (Fig. 3A) and Wnt5a-blockingantibody did not block the migration (Supplementary Fig.S4E) despite the high endogenous expression of Wnt5a (seeSupplementary Fig. S4A and S4D) Inhibition of CK1e, akinase, which is required for PCP signaling due to its rolein the phosphorylation of Dvl, with 2 unrelated inhibitorsD4476 (CK1 inh. I) and PF670462 (CK1 inh. II; 50 mmol/L;ref. 34) blocked the chemotactic response of MEC1 cellstoward CCL21 and CCL19 (Fig. 3B and C). It can be expectedthat CK1 blocks CLL migration via phosphorylation of thekey PCP protein Dvl. Indeed, even partial Dvl2 knockdownefficiently reduced the motility of MEC1 cells (Fig. 3D).Similarly to primary CLL cells, even in MEC1, the chemotaxiswas blocked by the inhibitors of Rho-associated kinaseROCK (Fig. 3E) and by the inhibitor of Wnt secretionIWP-2 (Fig. 3F) but not by the inhibitor of Rac1 (Fig. 3G).These data suggest that endogenous Wnts (but not uniquelyWnt5a), Dvl2, and likely the whole PCP machinery activatingRho and ROCK kinase is required for the polarized migrationof CLL cells.
PCP proteins regulate chemokine-driventransendothelial invasion of CLL cells
In the human body, cells behave in a complex environmentand have to pass via extracellular matrix or endothelial bar-riers, which is a process known as cell invasion. We have testedthe role of PCP proteins in the invasion of CLL cells by theanalysis of their invasion through a layer of HUVEC. In thisexperimental system, Wnt5a significantly promoted transen-dothelialmigration of primary patient CLL cells in a gradient of
Figure 4. Regulation of transendothelial migration of CLL cells by PCPpathway. A–D, transendothelial invasion of primary humanCLL lymphocytes andMEC1cells was tested in the Transwell assays coated with the HUVEC endothelial cell monolayer and is indicated as the migration index. The level ofinvasion of human primary CLL lymphocytes (A) was analyzed in the presence of chemokine CCL19, Wnt5a, or a combination of CCL19 and Wnt5a. Wnt5astrongly promotes CCL19-induced invasion of CLL cells through the endothelial monolayer. B–D, the positive effect of CCL19 on transendothelialmigration of MEC1 cells is fully blocked by the addition of the D4476 (CK1 inh. I; B). siRNA-mediated knockdown of Dvl2 (C) and the inhibition ofRho-associated kinase (Y27632) and endogenous Wnt production (IWP-2; D). Statistical differences were tested by one-way ANOVA and Tukey post hoctest (�, P < 0.05; ��, P < 0.01; ���, P < 0.001). Each experiment has been carried out in at least 3 independent replicates.
Kauck�a et al.
Cancer Res; 73(5) March 1, 2013 Cancer Research1496
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752

CCL19 (Fig. 4A), to a lesser extent in the gradient of CXCL12(Supplementary Fig. S4F) but showed no effect in the gradientof CCL21 (Supplementary Fig. S4G). This suggests that coop-eration between Wnt/PCP and chemokine signaling is notlimited to chemotaxis but takes place also in the process oftransendothelial invasion.To check whether invasion is controlled by PCP signaling,
specifically via CK1e-Dvl2-Rho pathway (as shown in Fig. 3 forchemotaxis), we have carried out another set of experiments inMEC1 cells. O the 3 tested chemokines, only CCL19 was able toinduce transendothelial migration of MEC1 cells (Supplemen-tary Fig. S4H). This response has not been promoted byWnt5abut has been almost completely abolished by CK1 inhibition(Fig. 4B). Similar to the basal chemotaxis, the downregulationof the key PCP protein Dvl2 by siRNA (Fig. 4C) as well asinhibition of ROCK kinase (by Y27632) and porcupine (by IWP-2; Fig. 4D) efficiently reduced also transendothelial invasion ofMEC1 cells. In summary, these results show that PCP proteinsmediate both migration/chemotaxis and chemokine-driveninvasion of CLL cells.
PCP proteins regulate CLL homing in vivoTo test the role of PCP proteins in CLL cells in vivo, we
transplanted primary CLL cells into NSG mice (Fig. 5A),which lack mature T cells, B cells, and functional naturalkiller cells (35). Primary CLL cells efficiently home in NSGmice and can be detected mainly in spleen, liver, and bonemarrow. Interestingly, pretreatment with the casein kinase 1inhibitor D4476 decreased homing to spleen, liver, and bonemarrow without any effect on the apoptosis of the cells (Fig.5B and C; for typical dot plots for apoptosis assay, seeSupplementary Fig. S5A). This experiment suggests thatinhibition of CK1 diminishes the ability of CLL cells tomigrate and colonize murine tissues. Interference with theRor1- and FZD7-driven signaling by treatment with anti-bodies directed against Ror1 and FZD7 (for validation ofanti-FZD7 antibody, see Supplementary Fig. S5B) did notaffect the survival of CLL cells either (Fig. 5D). Importantly,treatment with the blocking anti-Ror1 antibody (28) effi-ciently eliminated homing of CLL cells from most patients tothe spleen (Fig. 5E). The homing to bone marrow could notbe determined because treatment with any antibody (evencontrol IgG) for unknown reasons completely abolishedmigration to bone marrow in vivo (data not shown). Liverinfiltration was diminished only for some patients (Fig. 5F).Treatment with the anti-FZD7 antibody did not show anyeffect on the CLL cells in liver (Fig. 5F); however, decreasedspleen infiltration by CLL cells occurred in a subset ofpatients (Fig. 5E). Interestingly, the patient subset respond-ing to anti-FZD7 antibody had mutated p53, a strong markerof aggressive disease, whereas the patients not responding toanti-FZD7 antibody treatment had wild-type (wt) p53.In summary, these results show that CK1 activity and the
cell surface receptors Ror1 and FZD7 regulate biologicproperties of CLL cells in vivo. These observations areconsistent with the in vitro results and further strengthenthe conclusion that PCP proteins act as regulators of CLLbiology and pathogenesis.
PCP-high patient cohorts show worse clinicalcharacteristics
The expression aswell as functional data suggested that PCPproteins are strongly associated with CLL cell behavior. In thenext step, we analyze the clinical significance of PCP proteinexpression by analysis of samples from 93 previously untreatedpatients (identical to patients analyzed in Fig. 1A). The nor-malized expression of FZD3, FZD7, CELSR1, VANGL2, andPRICKLE1 assessed by qRT-PCR (see also Fig. 1A) was corre-lated to the clinical parameters.
Importantly, the increased expression of FZD3, FZD7,VANGL2, and PRICKLE1 correlated positively with unmutatedimmunoglobulin heavy chain (IGHV; Table 1), which is strong-ly associated with worse prognosis. In contrast, patients withunfavorable chromosomal aberrations del11q22-23 (ATM) ordel17p13 (TP53) did not show significantly higher expressionof PCP genes. However, it should be noted that the numberof patients with del11q22-23 (n ¼ 17; 18%) or del17p13 (n ¼ 7;7.5%) was rather low and insufficient for robust statisticalanalysis.
Next, we analyzed the association of PCP gene expressionwith the treatment-free survival (TFS) using Kaplan–Meiersurvival curves. Patients were sorted into 2 groups (high vs.low) based on the mRNA levels (as determined by qRT-PCR)of individual PCP genes (FZD3, FZD7, CELSR1, VANGL2, andPRICKLE1). The cutoff for each gene was set up as the valuecorresponding to 75% percentile for the expression levels innormal B cells from the periphery (see Fig. 1A and below).TFS analysis was conducted in both groups of patients.Importantly, FZD3-, FZD7-, and PRICKLE1-high patientcohorts had significantly (Breslow test, P < 0.05) worseprognosis than the group of patients with low expressionof these genes (Fig. 6). Specifically, the median TFS of FZD3-low (cutoff 1.2) patients was 40 months compared with 17months in FZD3-high group, 54 versus 25 months for FZD7low versus high (cutoff 2.0), and 54 versus 31 months forPRICKLE1-low versus PRICKLE1-high (cutoff 1.0) patientcohorts. Very similar data were obtained when we con-ducted the analysis with the data normalized to b-actin(Supplementary Fig. S6).
These findings suggest that mainly more aggressive CLLcells with unmutated IGHV use PCP pathway to regulate theirchemotaxis toward chemokines. Indeed, when we looked atunmutated IGHV patient subset only, we have not observedany statistically significant difference in survival curvesbetween PCP-high and -low patient cohorts (data not shown).In summary, the expression analyses in patients with CLLsshowed that (i) the expression of key PCP genes correlates withclinically important mutational status of IGHV and (ii) on theclinical sample show the relevance of processes controlled byPCP pathway for disease pathogenesis.
DiscussionIn the present study we, for the first time, show that (i) core
PCP components such as Vangl2, Prickle1, CK1e, Dvl, andCelsr1 are upregulated in CLLs, (ii) PCP pathway componentshelp regulate chemotaxis and transendothelial migration inthe chemokine gradient and in vivo homing of CLL cells, and
PCP Proteins Drive Pathogenesis of CLL
www.aacrjournals.org Cancer Res; 73(5) March 1, 2013 1497
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752

(iii) PCP-high patient cohorts show worse clinical parameterssuch as TFS. Our findings suggest that PCP proteins affect CLLpathogenesis via regulation of chemokine-driven migrationand as such influence disease progression.
Our data suggest that PCP pathway contributes to theCLL pathogenesis mainly via regulation of chemotacticresponses to chemokines, which are the leading mediatorsof the interaction between CLL cells and their microenvi-
Figure 5. PCP proteins affect engraftment of CLL cells in vivo. A, scheme of the experiment. Typical FACS dot blot is shown (see gate G1 for transplanted cells.).B, the level of apoptosis in primary CLL cells after dimethyl sulfoxide (DMSO)/D4476 (CK1 inh. I) was determined by flow cytometric analysis of themitochondrial membrane potential using tetramethylrhodamine ethyl ester dye in combination with the green calcein staining. C, proportion of CLL cells (number ofpositive cells/number of total cells in the organ) treated with DMSO or D4476 (CK1 inh. I) that were recovered from the spleen (Ci), liver (Cii), and bone marrow(Ciii) of transplanted mice. D, treatment with the aRor1 and aFZD7 antibodies does not induce apoptosis as determined by tetramethylrhodamine ethyl esteranalysis. E and F, proportion of CLL cells, when treated with a nonspecific antibody (IgG) or antibodies directed against Ror1 and FZD7, were recovered from themouse spleen (E) or liver (F). Please note that samples responding toaFZD7come frompatientswithmutated p53 (p53MUT). B andD, graphs showmean� SEM.C and E, one symbol on the graph represents one patient. �, P < 0.05, Wilcoxon paired t test (C) or one-way ANOVA and Tukey post hoc test (E).
Kauck�a et al.
Cancer Res; 73(5) March 1, 2013 Cancer Research1498
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752

ronment (for review, see ref. 36). Components of the PCPmachinery thus become an interesting target for the noveltherapies, which interfere with the communication betweenCLL cells and their environment, similarly to the inhibitorsof chemokine receptor or B-cell receptor (BCR) signalingpathways (37–39). In agreement with these data, our obser-vations show that PCP-controlled migration is mainly rel-evant for the CLL cells with unmutated IGHV, which wereshown to be more dependent on chemokine and BCRsignaling.Our data provide to our best knowledge (i) the first
evidence for the increased levels of the conserved proteinsunique for the PCP pathway—Prickle1, Vangl2, Celsr1—in
leukemia and (ii) the first implication for the role of Celsr1 inhuman cancer. PCP signaling has been, however, function-ally implicated earlier in the metastatic process of varioussolid tumors with the best defined role in melanoma, breast,and gastric cancer (40–42). It has been convincingly shown,mainly in melanoma, that PCP pathway increases the capac-ity of tumor cells to migrate and invade into surroundingtissue (43). Our data support the possibility that PCP path-way mediates physical aspects of cell migration via its effectson cytoskeleton and that chemokine signaling can serve asthe navigator of invading cells into their final destination.Interesting prediction of this hypothesis is that PCP pathwayis dynamically and temporarily used by migrating immune
Table 1. The expression of PCP genes determined by qRT-PCR in the patient cohorts with mutated (IGHVMUT) and unmutated (IGHV UNMUT) immunoglobulin heavy chain
Gene nameIGHV MUT(n ¼ 39)
IGHV UNMUT(n ¼ 54) P (Mann–Whitney test)
FZD7 2.697 (�0.9081) 3.674 (�0.6834) 0.0075FZD3 0.7684 (�0.07373) 1.217 (�0.1130) 0.0054VANGL2 4.937 (�2.137) 11.12 (� 6.119) 0.007PRICKLE1 1.942 (�0.2322) 2.978 (�0.2950) 0.0113
NOTE: Numbers indicate the mean, �SEM; n, number of patients; P, statistical significance of the difference between both cohortsdetermined by Mann-Whitney test.
Figure 6. Expression of PCP genes defines CLL progression. Patients were sorted into 2 groups (high vs. low expression; cutoff indicated by the line inthe upper graph) based on the mRNA levels of individual PCP genes (FZD3, FZD7, and PRICKLE1), and Kaplan–Meier survival curves (TFS) wereplotted and survival analysis was conducted in both groups of patients. PRICKLE1- (A), FZD3- (B), and FZD7-high (C) patient cohorts had a significantly(Breslow test, P < 0.05) worse prognosis than the group of patients with low expression of these genes.
PCP Proteins Drive Pathogenesis of CLL
www.aacrjournals.org Cancer Res; 73(5) March 1, 2013 1499
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752

cells during the immune response. This interesting topicclearly exceeds the current study and has to be tested infuture.
In summary, our findings provide strong evidence that corecomponents of the Wnt/PCP pathway play an important rolein CLL pathogenesis via regulation of CLL migration. We forthe first time show the biologic importance of the crosstalkbetween chemokine and PCP signaling, which might haveimplication for understanding of the biology of invasivenessin other tumors. Given the fact that Wnt/PCP pathway is anevolutionary conserved regulator of cell polarity and migra-tion, our data further emphasize the role of cellmigration in thedevelopment of CLLs.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: M. Kauck�a, P. Krej�cí, Š. Pospíšlov�a, V. BryjaDevelopment of methodology: M. Kauck�a, P. Krej�cíAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): M. Kauck�a, K. Plevov�a, S. Pavlov�a, P. Janovsk�a,
A. Mishra, J. Verner, J. Proch�azkov�a, J. Kotaškov�a, B. Tich�y, Y. Brychtov�a,M. Doubek, J. MayerAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): M. Kauck�a, S. Pavlov�a, A. Mishra, J. Proch�azkov�a,P. Krej�cí, J. Kotaškov�a, P. Ovesn�a, B. Tich�y, V. BryjaWriting, review, and/or revision of the manuscript:M. Kauck�a, K. Plevov�a,S. Pavlov�a, A. Mishra, A. Kozubík, J. Mayer, Š. Pospíšlov�a, V. BryjaAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): K. Plevov�a, J. Kotaškov�a, A. Kozubík,J. MayerStudy supervision: P. Krej�cí, Š. Pospíšlov�a, V. Bryja
Grant SupportThis work was supported by grants from the Czech Science Foundation (204/
09/H058, 301/11/0747), Ministry of Health of the Czech Republic (NT11217-5/2010, NT 13493-4/2012), Ministry of Education, Youth and Sports of the CzechRepublic (MSM0021622430), Masaryk University Rector's programme to supportMU student's creative work (MUNI/E/0128/2009, MUNI/A/0784/2011), Europe-an Regional Development Fund (CZ.1.07/2.3.00/20.0180, CZ.1.05/1.1.00/02.0068,CZ.1.07/2.3.00/20.0045), and by an EMBO Installation Grant. P. Krej�cí is sup-ported by the grants Kontakt LH12004 (Ministry of Education) and P305/11/0752(Czech Science Foundation).
The costs of publication of this article were defrayed in part by the paymentof page charges. This article must therefore be hereby marked advertisementin accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
ReceivedMay 7, 2012; revised November 29, 2012; accepted December 12, 2012;published OnlineFirst January 21, 2013.
References1. Clevers H. Wnt/beta-catenin signaling in development and disease.
Cell 2006;127:469–80.2. SemenovMV,HabasR,MacdonaldBT,HeX.SnapShot: noncanonical
Wnt signaling pathways. Cell 2007;131:1378.3. Seifert JR, Mlodzik M. Frizzled/PCP signalling: a conserved mecha-
nism regulating cell polarity and directed motility. Nat Rev Genet2007;8:126–38.
4. Torban E, Kor C, Gros P. Van Gogh-like2 (Strabismus) and its role inplanar cell polarity and convergent extension in vertebrates. TrendsGenet 2004;20:570–7.
5. Caligaris-Cappio F, Ghia P. Novel insights in chronic lymphocyticleukemia: are we getting closer to understanding the pathogenesisof the disease? J Clin Oncol 2008;26:4497–503.
6. Chiorazzi N, Rai KR, Ferrarini M. Chronic lymphocytic leukemia. N EnglJ Med 2005;352:804–15.
7. Zenz T, Mertens D, Kuppers R, Dohner H, Stilgenbauer S. Frompathogenesis to treatment of chronic lymphocytic leukaemia. Nat RevCancer 2010;10:37–50.
8. Kotaskova J, Tichy B, Trbusek M, Francova HS, Kabathova J, Mal-cikova J, et al. High expression of lymphocyte-activation gene 3(LAG3) in chronic lymphocytic leukemia cells is associated withunmutated immunoglobulin variable heavy chain region (IGHV) geneand reduced treatment-free survival. J Mol Diagn 2010;12:328–34.
9. Baskar S, Kwong KY, Hofer T, Levy JM, Kennedy MG, Lee E, et al.Unique cell surface expression of receptor tyrosine kinase ROR1 inhuman B-cell chronic lymphocytic leukemia. Clin Cancer Res2008;14:396–404.
10. Fukuda T, Chen L, Endo T, Tang L, Lu D, Castro JE, et al. Antiserainduced by infusions of autologous Ad-CD154-leukemia B cells iden-tify ROR1 as an oncofetal antigen and receptor for Wnt5a. Proc NatlAcad Sci U S A 2008;105:3047–52.
11. Rai KR, Sawitsky A, Cronkite EP, Chanana AD, Levy RN, PasternackBS. Clinical staging of chronic lymphocytic leukemia. Blood 1975;46:219–34.
12. Ghia P, Chiorazzi N, Stamatopoulos K. Microenvironmental influencesin chronic lymphocytic leukaemia: the role of antigen stimulation.J Intern Med 2008;264:549–62.
13. Vaisitti T, Aydin S, Rossi D, Cottino F, Bergui L, D'Arena G, et al. CD38increases CXCL12-mediated signals and homing of chronic lympho-cytic leukemia cells. Leukemia 2010;24:958–69.
14. HartmannTN,GrabovskyV,WangW,DeschP,RubenzerG,Wollner S,et al. Circulating B-cell chronic lymphocytic leukemia cells displayimpaired migration to lymph nodes and bone marrow. Cancer Res2009;69:3121–30.
15. Redondo-Munoz J, Jose Terol M, Garcia-Marco JA, Garcia-Pardo A.Matrix metalloproteinase-9 is up-regulated by CCL21/CCR7 interac-tion via extracellular signal-regulated kinase-1/2 signaling and isinvolved in CCL21-driven B-cell chronic lymphocytic leukemia cellinvasion and migration. Blood 2008;111:383–6.
16. Richardson SJ, Matthews C, Catherwood MA, Alexander HD, CareyBS, Farrugia J, et al. ZAP-70 expression is associated with enhancedability to respond to migratory and survival signals in B-cell chroniclymphocytic leukemia (B-CLL). Blood 2006;107:3584–92.
17. Nishita M, Itsukushima S, Nomachi A, Endo M, Wang Z, Inaba D,et al. Ror2/Frizzled complex mediates Wnt5a-induced AP-1 acti-vation by regulating Dishevelled polymerization. Mol Cell Biol2010;30:3610–9.
18. Minami Y, Oishi I, Endo M, Nishita M. Ror-family receptor tyrosinekinases in noncanonical Wnt signaling: their implications in devel-opmental morphogenesis and human diseases. Dev Dyn 2010;239:1–15.
19. Bryja V, Schulte G, Rawal N, Grahn A, Arenas E. Wnt-5a in-duces Dishevelled phosphorylation and dopaminergic differ-entiation via a CK1-dependent mechanism. J Cell Sci 2007;120:586–95.
20. Witze ES, Litman ES, Argast GM, Moon RT, Ahn NG. Wnt5a control ofcell polarity and directional movement by polarized redistribution ofadhesion receptors. Science 2008;320:365–9.
21. Ghosh MC, Collins GD, Vandanmagsar B, Patel K, Brill M, Carter A,et al. Activation of Wnt5A signaling is required for CXC chemokineligand 12-mediated T-cell migration. Blood 2009;114:1366–73.
22. Redondo-Munoz J, Escobar-Diaz E, Samaniego R, Terol MJ, Gar-cia-Marco JA, Garcia-Pardo A. MMP-9 in B-cell chronic lympho-cytic leukemia is up-regulated by alpha4beta1 integrin or CXCR4engagement via distinct signaling pathways, localizes to podo-somes, and is involved in cell invasion and migration. Blood2006;108:3143–51.
23. Witte F, Bernatik O, Kirchner K, Masek J, Mahl A, Krejci P, et al.Negative regulation of Wnt signaling mediated by CK1-phosphorylat-ed Dishevelled via Ror2. Faseb J 2010;24:2417–26.
Kauck�a et al.
Cancer Res; 73(5) March 1, 2013 Cancer Research1500
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752

24. Strutt H, Price MA, Strutt D. Planar polarity is positively regulated bycasein kinase I epsilon in Drosophila. Curr Biol 2006;16:1329–36.
25. Klein TJ, Jenny A, Djiane A, Mlodzik M. CKIepsilon/discs overgrownpromotes both Wnt-Fz/beta-catenin and Fz/PCP signaling in Dro-sophila. Curr Biol 2006;16:1337–43.
26. Rena G, Bain J, Elliott M, Cohen P. D4476, a cell-permeant inhibitor ofCK1, suppresses the site-specific phosphorylation and nuclear exclu-sion of FOXO1a. EMBO Rep 2004;5:60–5.
27. Korinek V, Barker N, Morin PJ, van Wichen D, de Weger R,Kinzler KW, et al. Constitutive transcriptional activation by abeta-catenin-Tcf complex in APC-/- colon carcinoma. Science1997;275:1784–7.
28. KauckaM, Krejci P, Plevova K, Pavlova S, Prochazkova J, Janovska P,et al. Post-translational modifications regulate signalling by Ror1. ActaPhysiol (Oxf) 2011;203:351–62.
29. Uehata M, Ishizaki T, Satoh H, Ono T, Kawahara T, Morishita T, et al.Calcium sensitization of smooth muscle mediated by a Rho-associ-ated protein kinase in hypertension. Nature 1997;389:990–4.
30. Gao Y, Dickerson JB, Guo F, Zheng J, Zheng Y. Rational design andcharacterization of a Rac GTPase-specific small molecule inhibitor.Proc Natl Acad Sci U S A 2004;101:7618–23.
31. ChenB,DodgeME, TangW, Lu J,MaZ, FanCW, et al. Smallmolecule-mediateddisruptionofWnt-dependent signaling in tissue regenerationand cancer. Nat Chem Biol 2009;5:100–7.
32. Stacchini A, Aragno M, Vallario A, Alfarano A, Circosta P, Gottardi D,et al. MEC1 and MEC2: two new cell lines derived from B-chroniclymphocytic leukaemia in prolymphocytoid transformation. Leuk Res1999;23:127–36.
33. BertilaccioMT, ScielzoC, Simonetti G, PonzoniM, Apollonio B, Fazi C,et al. A novel rag2-/-{gamma}c-/–xenograft model of human CLL.Blood 2010;115:1605–9.
34. Badura L, Swanson T, AdamowiczW, Adams J, Cianfrogna J, Fisher K,et al. An inhibitor of casein kinase I epsilon induces phase delays incircadian rhythms under free-running and entrained conditions.J Pharmacol Exp Ther 2007;322:730–8.
35. Shultz LD, Lyons BL, Burzenski LM, Gott B, Chen X, Chaleff S, et al.Human lymphoid and myeloid cell development in NOD/LtSz-scidIL2R gamma null mice engrafted with mobilized human hemopoieticstem cells. J Immunol 2005;174:6477–89.
36. Burger JA. Chemokines and chemokine receptors in chronic lympho-cytic leukemia (CLL): from understanding the basics towards thera-peutic targeting. Semin Cancer Biol 2010;20:424–30.
37. Hoellenriegel J, Meadows SA, Sivina M, Wierda WG, Kantarjian H,Keating MJ, et al. The phosphoinositide 30-kinase delta inhib-itor, CAL-101, inhibits B-cell receptor signaling and chemo-kine networks in chronic lymphocytic leukemia. Blood 2011;118:3603–12.
38. Friedberg JW, Sharman J, Sweetenham J, Johnston PB, Vose JM,Lacasce A, et al. Inhibition of Syk with fostamatinib disodium hassignificant clinical activity in non-Hodgkin lymphoma and chroniclymphocytic leukemia. Blood 2010;115:2578–85.
39. Herman SE, Gordon AL, Wagner AJ, Heerema NA, Zhao W, Flynn JM,et al. Phosphatidylinositol 3-kinase-delta inhibitor CAL-101 showspromising preclinical activity in chronic lymphocytic leukemia byantagonizing intrinsic and extrinsic cellular survival signals. Blood2010;116:2078–88.
40. Weeraratna AT, Jiang Y, Hostetter G, Rosenblatt K, Duray P, BittnerM,et al. Wnt5a signaling directly affects cell motility and invasion ofmetastatic melanoma. Cancer Cell 2002;1:279–88.
41. Pukrop T, Klemm F, Hagemann T, Gradl D, Schulz M, Siemes S,et al. Wnt 5a signaling is critical for macrophage-induced invasionof breast cancer cell lines. Proc Natl Acad Sci U S A 2006;103:5454–9.
42. KurayoshiM,OueN,YamamotoH,KishidaM, InoueA,Asahara T, et al.Expression of Wnt-5a is correlated with aggressiveness of gastriccancer by stimulating cell migration and invasion. Cancer Res2006;66:10439–48.
43. O'ConnellMP, Fiori JL, XuM,Carter AD, FrankBP,Camilli TC, et al. Theorphan tyrosine kinase receptor, ROR2, mediates Wnt5A signaling inmetastatic melanoma. Oncogene 2009;29:34–44.
PCP Proteins Drive Pathogenesis of CLL
www.aacrjournals.org Cancer Res; 73(5) March 1, 2013 1501
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752

2013;73:1491-1501. Published OnlineFirst January 21, 2013.Cancer Res Markéta Kaucká, Karla Plevová, Sárka Pavlová, et al. MigrationLymphocytic Leukemia by the Regulation of B-Lymphocyte The Planar Cell Polarity Pathway Drives Pathogenesis of Chronic
Updated version
10.1158/0008-5472.CAN-12-1752doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2013/05/06/0008-5472.CAN-12-1752.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/73/5/1491.full#ref-list-1
This article cites 43 articles, 23 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/73/5/1491.full#related-urls
This article has been cited by 10 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/73/5/1491To request permission to re-use all or part of this article, use this link
on August 21, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 21, 2013; DOI: 10.1158/0008-5472.CAN-12-1752