the molecular cell biology of anterior pituitary cellsbertram/papers/hypothalamus/cell_endo… ·...
TRANSCRIPT

C H A P T E R
2c0002
The Molecular Cell Biology of AnteriorPituitary Cells
Arturo E. Gonzalez-Iglesias and Richard BertramFlorida State University, Tallahassee, FL, USA
s0010INTRODUCTION
“Do notAU:1 stop to question whether these ideas arenew or old, but ask, more properly, whether theyharmonize with nature.” Marcello Malpighi
p0015 The dawn of neuroendocrinology is coinci-dent with the beginning of the tale of aremarkable gland: the pituitary gland. AroundAD 170 Galen postulated that the “pituita”(from the Greek ptuo, “to spit,” and the Latinpituita, “mucus”) secreted waste products(phlegm, one of the four humors of the body)from the brain into the nasal cavities. Nineteencenturies later, we have found not only thatGalen’s concept was far from the truth but alsothat a fascinating � yet still primitive � under-standing of the gland has emerged.
p0020 The pituitary is one of the two elements thatmake up the hypothalamo�hypophysial unit,the joint anatomical structure comprisinghormone-producing neurons and cells by whichthe brain regulates the vital functions of thebody. Indeed, a key regulator of body homeosta-sis during development, stress, and other
physiological processes, the pituitary gland actsas a double interpreter, mediating the talkbetween the brain and the peripheral organs,and integrating their respective cues as well asthose of its own (local autocrine and paracrinefactors). Being functionally and anatomicallyconnected to the hypothalamus by the medianeminence via the infundibular stalk,1 the pitui-tary has two embryologically and functionallydistinct divisions: the neurohypophysis (or neu-ral lobe) and the adenohypophysis (anteriorpituitary and intermediate lobes). The remark-able molecular and cellular aspects of the biol-ogy of the cells that constitute the anteriorpituitary is the focus of this chapter.
s0015ANTERIOR PITUITARY:ORGANIZATION, CELL TYPES,HORMONES AND FUNCTIONS
p0025The anterior pituitary is an endocrine glandresponsible for secreting hormones that regu-late a wide range of functions. These hormones
19Cellular Endocrinology in Health and Disease.
DOI: http://dx.doi.org/10.1016/B978-0-12-408134-5.00002-0 © 2014 Elsevier Inc. All rights reserved.
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

are synthesized and released by distinct groupsof polygonal endocrine cells that are organizedas interlacing cords and lined up on an anasto-mizing web of capillary vessels (the secondarycapillary plexus) derived from the hypophysialartery. The cytoplasm of these cells containsgranules of stored hormone that are released byexocytosis. The endothelial cells of the capillar-ies are fenestrated to facilitate the exchange ofmolecules between the endocrine cells and theblood, which not only bring in the hypotha-lamic and peripheral factors (through the longportal vessels and hypophysial arteries, respec-tively) that regulate the activity of the gland butalso carry the released pituitary hormonesaway into the general circulation.1 In addition,an extensive web of interconnected folliculos-tellate (FS) cells surround the endocrine cells.These cells regulate both the interaction ofneighboring endocrine cells and the exchangeof molecules between them and the capillaries.FS cells represent about 5�10% of the anteriorpituitary cell population.
p0030 The traditional view of the pituitary holdsthat there are five endocrine cell types that areresponsible for synthesizing six anterior pitui-tary hormones (Table 2.1). For each cell type,several immortalized cell lines have been devel-oped, characterized and used extensively.2
p0035 Somatotrophs, which synthesize and releasegrowth hormone (GH), are the major endo-crine cell type in the anterior pituitary andconstitute 40�50% of its cell population. Theyare localized predominantly to the lateral por-tions of the anterior lobe. Somatotroph func-tion is primarily regulated by hypothalamicfactors: growth hormone-releasing hormone(GHRH) produced by neurons in the arcuatenucleus is stimulatory, whereas somatostatin(STT) produced by neurons in the periventri-cular nucleus is inhibitory. STT suppressesboth basal and GHRH-induced GH release,having no effect on GH synthesis. GH produc-tion and secretion also receives inhibitory feed-back from the major target of GH, insulin-like
growth factor-I from the liver. Somatotrophsexpress receptors for many other regulators ofGH synthesis and release, including ghrelin,pituitary adenylate cyclase-activating peptide,thyroid hormone, glucocorticoids, insulin andendothelins. GH is secreted from the somato-trophs at a pulse frequency of about 1�2 hwith a half-life that ranges between 6�20 min,and the pattern exhibits gender differences. Inthe case of males, the pulses are much largerearly at night, whereas in females the patternis more irregular and the pulses tend to bemore uniform throughout the day. The patternof GH release appears to be driven by the rateat which GHRH is released from the arcuatenucleus neurons.
p0040GH is also called somatotropin (soma,“body”) because of its profound and wide-spread anabolic effects throughout the body.In its absence, growth is stunted. Although vir-tually every tissue responds to some degree,skeletal muscle cells, liver, and chondrocytes(cartilage cells) are particularly sensitive to GHlevels. Though the metabolic effects are directactions of GH, it is now apparent that most, ifnot all, of the anabolic effects of GH are medi-ated by the production of a family of peptidehormone intermediaries known as insulin-likegrowth factors (IGFs) which are secreted bythe liver, cartilage, muscle, and other tissueswhere they can act locally in a paracrine orautocrine fashion. GH, acting through theIGFs, stimulates protein synthesis, cell growthand a positive nitrogen balance, leading to anincrease in lean body mass and a decrease inbody fat. Many hours must elapse after admin-istration of GH before its anabolic, growth-promoting effects become evident.
p0045Thyrotrophs comprise approximately 5% ofthe anterior endocrine cell population and aretypically spread over the anteriomedial andlateral portions of the gland. Thyrotrophs syn-thesize and secrete thyroid-stimulating hor-mone (TSH), also known as thyrotropin, whichis controlled by central and peripheral
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
20 2. BIOLOGY OF ANTERIOR PITUITARY CELLS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

regulators. The dominant stimulatory controlof thyrotroph function and TSH secretion isexerted by the hypothalamic neurohormonethyrotropin-releasing hormone (TRH) releasedby neurons of the paraventricular nucleus.TSH is secreted in pulses lasting for 2�3 hwith a nocturnal surge before sleep. Once thesleep phase begins, TSH release is curtailed.The half-life of the hormone in blood isapproximately of 1 h.
p0050 TSH is a glycoprotein with a molecularweight of 28,000 and consists of a heterodimerof two subunits (α and β) that are tightly associ-ated by noncovalent forces and encoded byseparate genes. Although both subunits arerequired for receptor binding and hormoneaction, the β subunit confers biological specific-ity to the TSH molecule, as the α subunit is alsoa component of the anterior pituitary gonado-tropin hormones luteinizing hormone (LH) andfollicle-stimulating hormone (FSH). The essen-tial actions of TSH are those exerted on the thy-roid gland, where it promotes growth anddifferentiation of the gland and stimulates allsteps in the secretion of the thyroid hormones
thyroxine (T4) and triiodothyronine (T3). Thesesteps include glandular uptake of iodide, itsorganification, the completion of thyroid hor-mone synthesis, and the subsequent release ofthyroid gland products. T3 and T4 act at thehypothalamic and pituitary levels to block thesecretion of TSH via feedback inhibition.Fasting decreases thyrotroph responsiveness toTRH, while exposure to cold increases it.
p0055Corticotrophs constitute about 15% of theadenohypophysial endocrine cell populationand are scattered throughout the anterior lobein adult animals, but are primarily found in theanteromedial part of the gland. They synthesizeproopiomelanocortin (POMC) and release itsproteolytic derivatives, adrenocorticotropinhormone (ACTH), α-melanocyte-stimulatinghormone (α-MSH), lipotropic hormone (LPH)and endorphins. The main releasing factors forare the hypothalamic corticotropin-releasinghormone (CRH) and arginine-vasopressin( AU:2AVP), which acts in synergy with CRH topotentiate hormone release. Glucocorticoids,secreted by the adrenal cortex, are the majorphysiological inhibitor. ACTH levels in plasma
t0010 TABLE 2.1 Hypothalamic Neurohormones and Anterior Pituitary Endocrine Cell Types and Hormones
Hypothalamic Releasing
or InhibitingNeurohormone
Target Cell in
Anterior PituitaryGland
Anterior Pituitary Hormone
Release Stimulated orInhibited Target Anterior Pituitary Hormone
Growth hormone releasinghormone (GHRH)
Somatotroph Growth hormone (GH) Multiple somatic tissues
Somatostatin (STT) GH Multiple somatic tissues
Thyrotropin releasinghormone (TRH)
Thyrotroph Thyroid stimulating hormone(TSH)
Stimulates synthesis and release ofthyroid hormone by thyroid gland
Corticotropin releasinghormone (CRH)
Corticotroph Adrenocorticotrophic hormone(ACTH)
Stimulates synthesis and release ofsteroids by adrenal cortex
Gonadotropin releasinghormone (GnRH)
Gonadotroph Follicle stimulating hormone(FSH) and luteinizing hormone(LH)
Stimulates synthesis and release ofgonadal steroids, gametogenesis,ovulation
Dopamine Lactotroph Prolactin Milk synthesis and secretion by themammary gland
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
21ANTERIOR PITUITARY: ORGANIZATION, CELL TYPES, HORMONES AND FUNCTIONS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

exhibit a circadian rhythm, with a peak in theearly morning followed by a gradual declineduring the night, and is controlled by clockneurons in the suprachiasmatic nucleus thatsynapse with CRH-producing neurons in thehypothalamic paraventricular nucleus. ACTHalso exhibits faster, ultradian and hourlyrhythms due to feedback between corticotrophsand adrenal cells.3
p0060 As its name implies, one of the majoractions of ACTH is the promotion of growth ofadrenal cortex cells. ACTH effects on thesecells are also necessary for both basal andstress-induced secretion of glucocorticoids andaldosterone. Its half-life of 10 min allows forrapid adjustments of circulating levels ofglucocorticoids. ACTH, CRH, and glucocorti-coids are the main effector molecules of thehypothalamo�pituitary�adrenal (HPA) axis,the system that activates to help the organismadapt and cope with different stress cues(hypoglycemia, anesthesia, surgery, trauma,hemorrhage, infection, pyrogens) and psychiat-ric disorders like anxiety or depression.
p0065 Gonadotrophs constitute about 10�15% ofthe anterior pituitary endocrine cells and arelocalized throughout the pars distalis andmost of the pars tuberalis of the anterior lobe.They form intimate contacts with lactotrophcells, with which they have extensive cell-to-cell (paracrine) interactions. Gonadotrophssynthesize and release two hormones essentialto the growth and function of the gonads inboth genders (hence their common designationas gonadotropins): luteinizing hormone (LH)and follicle-stimulating hormone (FSH).Synthesis and release of both hormones arestimulated by gonadotropin-releasing hormone(GnRH), which is secreted in a pulsatile man-ner by neurons that are dispersed within themediobasal hypothalamus and preoptic areas.In addition, other hypothalamic factors such asgonadotropin-inhibiting hormone, vasopressin,substance P, as well as feedback of gonadalfactors (estrogens, progestogens, androgens
and inhibin) contribute to the regulation ofgonadotropins. The release of LH and FSHoccurs in phase with pulses of GnRH, withintervals that range from 30 to 60 min depend-ing on the species and, for females, the stageof the ovarian cycle. During the ovarian cycle,the levels of LH and FSH in plasma correlatestrongly with the gonadotropin content of thepituitary, and both are highest just beforeovulation.
p0070LH, with a molecular weight of 28,000, andFSH, with a molecular weight of 33,000, areglycoproteins with similar structures. Each iscomposed of the common pituitary hormone αsubunit, also found in TSH, and a unique βsubunit that confers them hormonal specificity.The carbohydrate moiety of the latter subunitalso enhances their half-life (1 h for LH andabout 3 h for FSH) and is critical for receptorbinding and biological responses, and its mod-ification allows considerable variation of thebioactivity of secreted LH and FSH moleculesin different physiological conditions. LH sti-mulates the interstitial cell lines of male andfemale gonads (Leydig and thecal cells) mainlyto secrete androgens (particularly testoster-one), whereas FSH stimulates testicular Sertoliand ovarian granulosa cells to secrete estro-gens (particularly estradiol) and a variety ofprotein products essential to spermatogenesisand oogenesis, respectively. During the initia-tion of the reproductive cycle of females, FSHacts on primary follicles to stimulate growth ofthe granulosa cells. Once the follicular phase isadvanced and draws near to ovulation, LHalso acts in female granulosa cells to promoteprogesterone production.
p0075Lactotrophs make up about 15�25% of theadenohypophysial endocrine cell populationand are a particularly non-homogeneousgroup of endocrine cells scattered throughoutthe anterior pituitary. A significant number arealso found in the posterior medial portion ofthe gland. They synthesize and release prolac-tin ( AU:3PRL), a 198 amino-acid protein that owes
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
22 2. BIOLOGY OF ANTERIOR PITUITARY CELLS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

its name to its role in milk production duringlactation in mammals. PRL has structural simi-larity and a comparable half-life (20 min) toGH. Like somatotrophs, lactotrophs inherentlyhave high secretory activity due to their spon-taneous electrical activity, though the formercells are not as active as lactotrophs in terms oftheir secretory activity. Thus, unlike all otheranterior pituitary hormones, the physiologicalcontrol of PRL is predominantly via tonichypothalamic inhibition mediated by dopami-nergic neurons located primarily in the arcuatenucleus. Therefore, a drop in the levels ofdopamine (AU:4 DA) in portal plasma often trans-lates into an increase of the PRL-releasingactivity of these cells. Hormone release by lac-totrophs can also be stimulated by an exten-sive range of factors such as TRH, oxytocin,vasoactive intestinal peptide (VIP), angiotensinII, endothelin-1, serotonin and estrogens, butnone of them have been established as a physi-ologically relevant PRL-releasing hormone.Like other trophic hormones, PRL secretionrises at night, and release occurs in episodicpulses.
p0080 In addition to its roles in the growth of alve-olar breast cells and milk production, prolactininhibits GnRH-induced release of gonadotro-pins in the anterior pituitary as well as theactions of gonadotropins on the gonads. Thismay be the mechanism by which it preventsovulation in lactating women and normalsperm production in males. Indeed, PRL facili-tates the release of DA from theAU:5 ME and thusacts in a negative feedback loop to inhibit itsown secretion. Many other roles have beendescribed for prolactin, including control ofsexual behavior, induction of maternal behav-ior in pregnancy, and support of the corporalutea during pregnancy and pseudopregnancy(in rodents). Similarly to ACTH, PRL release isstimulated by different stressor cues, includinginsulin-induced hypoglycemia, infection, sur-gery, anesthesia, and fear. Exercise and stimu-lation of the nipples both stimulate prolactin
secretion, an effect that is thought to be medi-ated by oxytocin.
s0020DISEASES OF THE ANTERIORPITUITARY
p0085Diseases of the pituitary can be divided intotwo major categories of pituitary disturbances,namely, hyperactivity (termed hyperpituita-rism) and hypoactivity (hypopituitarism).4 Wefocus on those that arise from disorders of theanterior pituitary. Because the function of theanterior pituitary is based on the specializedaction (and interaction) of its different endo-crine cells, the type of clinical response to eachpituitary condition varies with the type of cellthat is affected.
s0025Hypopituitarism
p0090With an annual estimated incidence (num-ber of new cases per population in a giventime period) of 4.2 per 100,000 and prevalence(proportion of the total number of cases to thetotal population) of 45.5 per 100,000, hypopitu-itarism might be caused by either an inabilityof the gland itself to produce hormones or aninsufficient supply of hypothalamic-releasinghormones. It is causally associated with pitui-tary tumors (61%), non-pituitary lesions (9%),and non-cancerous causes (30%), includingperinatal insults, genetic causes, trauma andidiopathic cases. Often, mutations in genesencoding single hormones (or the receptor fortheir cell-specific hypothalamic releasing fac-tor) result in single pituitary hormone defi-ciency. Unless successfully treated,hypopituitarism is chronic and lifelong, and incases of shortage of ACTH or TSH it can causelife-threatening events and lead to increasedmortality. If there is decreased secretion ofmost pituitary hormones, the term panhypopi-tuitarism is used.
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
23DISEASES OF THE ANTERIOR PITUITARY
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

s0040 ACTH DEFICIENCYp0095 This causes adrenal atro-phy and ACTH-receptor downregulation, lowblood pressure, low blood sugar level, fatigue,weight loss, and low tolerance for stress.
s0045 TSH DEFICIENCYp0100 Features of TSH deficitinclude underactive thyroid, fatigue, coldintolerance, weight gain, constipation, hairloss, dry skin and cognitive slowing.
s0050 LH, FSH DEFICIENCIESp0105 In premenopausalwomen, absent or infrequent menstrual cycles,infertility, vaginal dryness, dyspareunia (pain-ful sexual intercourse), loss of libido, and lossof some female characteristics occur. In men,these deficiencies are associated with impo-tence, shriveling of testes, decreased spermproduction, infertility, loss of libido, and lossof some male characteristics. In childhood,deficiencies result in delayed or missing onsetof puberty.
s0055 GH DEFICIENCYp0110 The hallmarks of insuffi-cient GH release are decreased muscle massand strength, visceral obesity, fatigue,decreased quality of life, and impairment ofattention and memory; there is stunted growthand dwarfism in children.
s0060 PRL DEFICIENCYp0115 Inability to produce breastmilk after childbirth occurs in some women.However, elevated prolactin concentrationssometimes occur in hypopituitarism because ofdisruption of inhibitory signals by the hypothal-amus, causing lactation, tenderness of the breast,and suppression of gonadotropins, leading tosymptoms of hypogonadism.
s0065 Hyperpituitarism
p0120 Hyperpituitarism is the primary hypersecre-tion of pituitary hormones. It typically resultsfrom a pituitary tumor or adenoma, whichrepresent from 10�25% of all intracranial
neoplasms. Most pituitary adenomas arebenign, functional, and secrete a hormone thatproduces clear symptoms characteristic of theircondition. The four most common types ofadenoma-related hyperpituitarism are prolacti-noma, corticotropinoma (Cushing’s disease),somatotropinoma (gigantism), and null celladenomas that do not secrete hormones. Sincethe enlargement of the anterior pituitary glandcan damage the optic nerves and compress thehypothalamus, some of its common symptomsare headache, loss of side or peripheral vision,and hyposecretion of neighboring anteriorpituitary hormones. Clinically active pituitaryadenomas affect approximately one in 1000 ofthe general population but are rare in children.
s0080PROLACTINOMA p0125This is an adenoma oflactotroph cells and the most common type ofpituitary tumor (30% of pituitary adenomas). Inwomen, high blood levels of prolactin oftencause changes in menstruation (periods maybecome irregular or disappear) and those whoare not pregnant or nursing may begin produc-ing breast milk (galactorrhea). Because of thehyperprolactinemia-induced inhibition of thegonadotropin axis, women with hyperprolacti-nemia also exhibit symptoms of gonadotropindeficit, such as loss of libido, dyspareunia, vagi-nal dryness and hypogonadism. In males, themost common symptom of prolactinoma isimpotence, loss of libido, hypogonadism, oli-gospermia, and diminished ejaculate volume.
s0085CORTICOTROPINOMA p0130Also known asCushing’s disease, this is a tumor of cortico-troph cells that accounts for 20% of pituitaryadenomas. The symptoms reflect the presenceof excess cortisol or ACTH and include weightgain, high blood pressure, poor short-termmemory, irritability, extra fat around the neck,a round and ruddy face, fatigue, and poor con-centration. Women also exhibit menstrualirregularity and hirsutism (abnormal hairgrowth).
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
24 2. BIOLOGY OF ANTERIOR PITUITARY CELLS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

s0090 SOMATOTROPINOMAp0135 This is a tumor ofsomatotroph cells that represents 15% of pitui-tary adenomas. The resulting hypersecretion ofGH in adults causes acromegaly (an over-growth of the terminal parts of the skeletonsuch as the nose, mandible, hands and feet).When it presents in children and adolescents itcauses gigantism; disruption of sexual matura-tion is common, either because of hormonehypersecretion or because of manifestationscaused by compression of hypothalamic con-nections by the adenoma.
s0095 NULL CELL ADENOMASp0140 These are dys-functional tumors that account for 25% of pitu-itary adenomas. Because they do not secretehormones, symptoms are restricted to head-ache, increased intracranial pressure andvisual field defects.
s0100 GONADOTROPHIC ADENOMASp0145 These aretumors of gonadotroph cells that account for10% of pituitary adenomas. Although func-tional, they are usually clinically silent.
s0105 THYROTROPHIC ADENOMASp0150 These arerare (less than 1%) but have the particularcharacteristic of being plurihormonal, sincethey produce the common glycoproteinα-subunit, prolactin, and the specific β-subu-nits of LH and FSH.
s0110 PHENOTYPIC PROFILE OFANTERIOR PITUITARY CELL
TYPES: DIFFERENT, YETSOMEWHAT SIMILAR
p0155 The anterior pituitary endocrine cell typesare classically defined on the basis of theexpression of a specific hormone and its corre-sponding mRNA, and this is determined byvarious lineage-specific transcription factors.5
Although two-thirds of the population of eachanterior pituitary cell type exclusively express
their respective hormone at both mRNA andprotein levels, the remainder coexpress themRNAs of two to four different hormones.6
For example, a GH cell can also express themRNAs of PRL, TSH-β, LH-β and POMC. Inaddition, a significant fraction of the non-hormone producing cells contain multiple hor-mone mRNAs that fail to be translated into themature protein. Thus, at the mRNA level, thevarious anterior pituitary hormones are sharedby a fraction of cells that express multilineagephenotypes. These multiple mRNA-containing(or polyhormonal) cells are thought to be pro-genitor cells or “reserve” cells that uponappropriate signals will terminally differenti-ate depending on developmental and/or phys-iological needs. The origin and biologicalrelevance of these cells are currently a matterof intense investigation and debate.
p0160The current paradigm for hypothalamiccontrol of anterior pituitary hormone secretionholds that each hypothalamic-releasing hor-mone modulates the release of a single pitui-tary hormone by acting on a single cell type.However, when attempts were made to char-acterize cell phenotypes on the basis ofwhether their intracellular calcium concentra-tion ([Ca21]i) responded to a specific hypotha-lamic releasing hormone, many cells werefound to respond to two or more of theseagents. Furthermore, hormone release assaysin single living cells revealed that multi-responsiveness also exists with respect to hor-mone release. Such “paradoxical” hormonerelease by a noncorrresponding hypothalamicreleasing hormone had, in fact, been observedin a number of in vitro and in vivo studiesusing normal and pathological cells. Indeed, agrowing body of evidence suggests that30�40% of normal pituitary cells remain in amultipotential state, able to respond to asmany as four different hypothalamic secreta-gogues.7 In some pituitary adenomas, thesemulti-responsive cells can constitute up to 80%of the endocrine cells. Moreover, much like the
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
25PHENOTYPIC PROFILE OF ANTERIOR PITUITARY CELL TYPES: DIFFERENT, YET SOMEWHAT SIMILAR
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

polyhormonal cells described above, multi-responsive cells are not restricted to any partic-ular cell type. Thus, different cell types canalso share receptors to allegedly cell-specifichypothalamic neurohormones. In fact, themRNA of the receptor for the classic hypotha-lamic GH inhibitory hormone, somatostatin, isfound among all five major cell types of theanterior pituitary. The type 1 CRH receptor(CRH-R1) is not exclusively expressed by corti-cotrophs; subsets of lactotrophs, gonadotrophsand thyrotrophs also express it in differentdegrees. It has also been shown that about 35%of somatotrophs bind biotinylated GnRH, thestimulating hormone for gonadotrophs. It hasbeen hypothesized that estrogens and possiblyactivin stimulate the expression of GnRHreceptors in pre-existing GH cells at midcycle,rendering them capable of fully responding asa gonadotroph to help support the GnRH-mediated proestrus surge of LH and FSHrelease. Additionally, retention of GHRHreceptors would allow supporting the GHneeds of the reproductive system at this time.8
p0165 Whether these multi-responsive cells derivefrom an independent multipotential subset orfrom pre-existing differentiated cells (termedtransdifferentiation), they provide a cellularbasis for anterior pituitary plasticity. Twoobservations are consistent with this. The firstone is that multi-responsive cells are maxi-mally abundant at puberty, a time signed byhuge endocrine and physiological transforma-tion. Secondly, multifunctional cells seem to bemore abundant in females than in males, sug-gesting not only that the female pituitary maybe more plastic than the male pituitary, butalso that the hormonal changes associated withthe female reproductive cycle may promotetransdifferentiation.
p0170 Unstimulated anterior pituitary cells canalso be classified by their biophysical and bio-chemical properties, more specifically, on theirpatterns of electrical activity and intracellular
Ca21 dynamics.9,10 A general feature of cul-tured anterior pituitary cells is that their mem-brane potential (Vm) oscillates betweenpotentials of 260 to 250 mV. When Vm
reaches the threshold level, pituitary cells fireaction potentials (APs), a feature called sponta-neous electrical activity and is observed in15�80% of the cells, depending on the celltype and of cultural and/or recording condi-tions. The firing of APs causes transients ofintracellular Ca21 concentration ([Ca21]i) thatin turn reflect the pattern of electrical activity.Two patterns of electrical activity are typicallyobserved in anterior pituitary cells. The first,termed axonal-type AP spiking, is typicallyfound in gonadotrophs,11 thyrotrophs,12 andlargely in corticotrophs,13 and is characterizedby sharp single APs that are short in duration(less than 100 ms), with spike frequencies ofabout 0.7 Hz and amplitudes of more than60 mV. Axonal-type AP spiking is associatedwith small-amplitude [Ca21]i transients thatrange from 20 nM to 70 nM and low basal hor-monal release in these cells. The second pat-tern, termed pseudo-plateau bursting andcharacteristic of cultured somatotrophs14 andlactotrophs,15 exhibits broader Vm oscillationsin the form of a depolarizing plateau withsuperimposed bursts of small amplitude APsthat usually do not reach 0 mV. Such burstshave a longer duration (several seconds) thangonadotroph APs, so that the burst frequencyis significantly lower (about 0.3 Hz), and resultin an oscillatory increase in [Ca21]i of highamplitude that ranges from 0.3 to 1.2 μM andis sufficient to maintain high and steady hor-monal (GH and prolactin) release. Althoughthe typical patterns of spontaneous electricalactivity are useful for the identification of thedifferent cell populations of the anterior pitui-tary, they should not be used as the sole crite-ria, since the electrical activity of these cellsoften experience spontaneous reversible transi-tions between the two modes.
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
26 2. BIOLOGY OF ANTERIOR PITUITARY CELLS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

s0115 HETEROGENEITY WITHINANTERIOR PITUITARY CELL
TYPES: SIMILAR, YET SOMEWHATDIFFERENT
p0175 Although different cell types have similari-ties, it is also apparent that similar cells (that is,cells within a cell type population) may presentstriking differences. It has been long recognizedthat each of the different types of anterior pitui-tary cells are functionally heterogeneous, whichis thought to be related to structural heteroge-neity. One of the best known examples is foundamong gonadotrophs,16,17 which are heteroge-neous in size (cell area ranging from30�170 μm2), morphology (large rounded,small oval and angular stellate cells), physicaldensity (allowing their separation using sedi-mentation techniques at unit gravity), ultra-structural characteristics (type I gonadotrophscharacterized by dilated rough ER (RER) cister-nae and secretory granules of 200 nm and300�700 nm in diameter, type II gonadotrophsby flattened RER cisternae and 200�250 nmsecretory granules, type III gonadotrophs by astellate shape and secretory granules of220�250 nm), hormone content (small gonado-trophs seem to store only one of the gonadotro-pins, whereas most of the larger cells eitherstore both LH and FSH, or FSH alone) andresponsiveness (variable capacity to bind andrespond to GnRH). These gonadotroph subpo-pulations may account for the differential regu-lation of LH and FSH secretion in vivo.
p0180 The other anterior pituitary cell types exhibitheterogeneity of these cellular features as well.In the porcine pituitary, somatotrophs comprisetwo morphologically distinct subpopulations oflow- (LD) and high-density (HD) cells, separa-ble by Percoll gradient, that respond differentlyto hypothalamic regulators. In LD somato-trophs, somatostatin inhibits GHRH-inducedGH secretion, whereas somatostatin alone sti-mulates GH release from HD somatotrophs.
Functional heterogeneity of corticotrophs dis-plays striking sexual dimorphism. In males, thecorticotrophs are of the orthodox phenotype,that is, monohormonal (storing only ACTH)and monoreceptorial (responding only toCRH). Their female counterparts are made ofabout equal parts of orthodox and multifunc-tional cells. Sexual dimorphism and functionalheterogeneity are even more striking in thyro-trophs, which are mostly polyhormonal in bothgenders, but only female thyrotrophs co-storeGH and/or ACTH in addition to prolactinand/or LH. Among lactotrophs, three morpho-logical types in rodents have been defined byelectron microscopy. Type I “classical” lacto-trophs contain large irregular-shaped electron-dense secretory granules (diameter300�700 nm), type II cells contain numerousmedium sized spherical electron-dense gran-ules (diameter 100�250 nm) and type III cellscontain small (,100 nm) spherical granules.Though thought to represent different stages ofcell maturity, these morphologically-definedlactotroph subpopulations have functional dif-ferences, as these subtypes are differentiallysensitive to hypothalamic and local regulatorsof prolactin secretion.18
p0185Several substates of a particular pattern ofelectrical activity can be found within a givenanterior pituitary cell type. In somatotrophs,the burst period of somatotrophs ranges from2 to 10 s, with longer active phase durationassociated with the slower bursting, whichresults in increased Ca21 influx throughvoltage-gated Ca21 channels (VGCC AU:6) andhigher amplitude Ca21 oscillations. An expla-nation of the heterogeneity of the active phaseduration has been recently provided.14,19
p0190Lactotrophs also display considerable vari-ability in their spontaneous and receptor-controlled patterns of electrical activity and Ca21
dynamics. The authors have recently addressedthe heterogeneity in the [Ca21]i responses of lac-totrophs to TRH.20 Responses were evaluated in
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
27HETEROGENEITY WITHIN ANTERIOR PITUITARY CELL TYPES: SIMILAR, YET SOMEWHAT DIFFERENT
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.
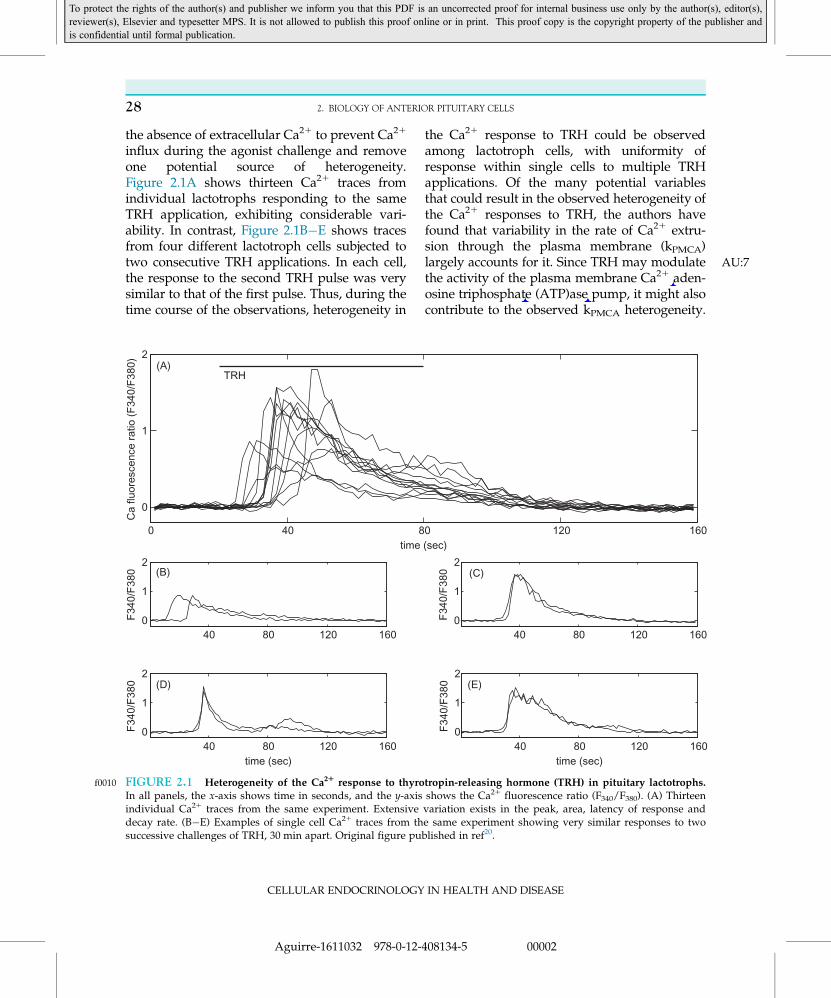
the absence of extracellular Ca21 to prevent Ca21
influx during the agonist challenge and removeone potential source of heterogeneity.Figure 2.1A shows thirteen Ca21 traces fromindividual lactotrophs responding to the sameTRH application, exhibiting considerable vari-ability. In contrast, Figure 2.1B�E shows tracesfrom four different lactotroph cells subjected totwo consecutive TRH applications. In each cell,the response to the second TRH pulse was verysimilar to that of the first pulse. Thus, during thetime course of the observations, heterogeneity in
the Ca21 response to TRH could be observedamong lactotroph cells, with uniformity ofresponse within single cells to multiple TRHapplications. Of the many potential variablesthat could result in the observed heterogeneity ofthe Ca21 responses to TRH, the authors havefound that variability in the rate of Ca21 extru-sion through the plasma membrane (kPMCA)largely accounts for it AU:7. Since TRH may modulatethe activity of the plasma membrane Ca21 aden-osine triphosphate (ATP)ase pump, it might alsocontribute to the observed kPMCA heterogeneity.
0 40 80 120 160
0
1
2(A)
time (sec)
Ca
fluor
esce
nce
ratio
(F
340/
F38
0)
40 80 120 1600
1
2(B)
F34
0/F
380
40 80 120 1600
1
2(C)
F34
0/F
380
40 80 120 1600
1
2(D)
time (sec)
F34
0/F
380
40 80 120 1600
1
2(E)
time (sec)
F34
0/F
380
TRH
f0010 FIGURE 2.1 Heterogeneity of the Ca21 response to thyrotropin-releasing hormone (TRH) in pituitary lactotrophs.In all panels, the x-axis shows time in seconds, and the y-axis shows the Ca21 fluorescence ratio (F340/F380). (A) Thirteenindividual Ca21 traces from the same experiment. Extensive variation exists in the peak, area, latency of response anddecay rate. (B�E) Examples of single cell Ca21 traces from the same experiment showing very similar responses to twosuccessive challenges of TRH, 30 min apart. Original figure published in ref20.
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
28 2. BIOLOGY OF ANTERIOR PITUITARY CELLS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

We also found that variability in the [Ca21] inthe endoplasmic reticulum ([Ca21]ER) is likely akey element of heterogeneity. In addition, ourresults did not show a positive correlationbetween the peak and decay rate of the intracel-lular Ca21 transients, suggesting that variationsin the Gq/IP3 signaling pathway are not themain source of heterogeneity.
s0120 PATTERNS OF ANTERIORPITUITARY CELL SIGNALING AND
SECRETION
s0125 Molecular Mechanisms UnderlyingExcitability and Basal Secretion
p0195 In excitable anterior pituitary cells, the restingmembrane potential is controlled by classicalinward rectifier K1 (Kir), ether-a-go-go-relatedgene (ERG) and TWIK-related (TREK-1) K1-conducting channels. Because the Vm of anteriorpituitary cells ranges between 250 to 260 mV,other depolarizing conductances must contributeto maintaining the resting potential. Recent find-ings indicate that tetrodotoxin (TTX)-insensitiveNa1-conducting channels, along with low-voltage-activated transient (T)-type voltage-gatedCa21 channels, not only participate in the controlof the resting potential but also act as pacemak-ing currents underlying the spontaneous activityfrequently observed in isolated cells.10 In physio-logical (in vivo) conditions, TTX-sensitive voltage-gated Na1 channels may play a role in theproduction of action potentials in hyperpolarizedcells. However, high voltage-activated,dihydropyridine-sensitive long lasting (L)-typeCa21 channels account for the bulk of the con-ductance during spike depolarization as removalof extracellular Ca21 and addition of VGCCblockers abolish electrical activity in the majorityof the endocrine pituitary cells without affectingtheir resting membrane potential. Finally, theconcerted activity of delayed-rectifying K1 chan-nels with both BK (large conductance) and SK
(small conductance) types of Ca21-activated K1
channels repolarize the Vm during the down-stroke of an AP. In bursting cells, BK channelsare also functionally important.
p0200The distinctive patterns of electrical activityand Ca21 dynamics exhibited by differenttypes of pituitary cells are thought to arisefrom differences in the expression levels ofion channels such as those involved in burst-ing and repolarization. For example, BK chan-nels are expressed at higher levels insomatotrophs and lactotrophs (which exhibitpseudo-plateau bursting activity) than ingonadotrophs (which display continuous,axonal-type AP spiking).21 This correlationbetween BK channel expression levels andpattern of electrical activity suggests thatlarger BK conductance favors the generationof bursting activity and global Ca21 signals.Indeed, blockade of BK channels in pituitarysomatotrophs can switch the activity patternof these cells from bursting to spiking, greatlyreducing the amplitude of [Ca21]i oscilla-tions.22 In other words, it is possible to con-vert the firing phenotype of somatotrophs tothat of gonadotrophs by reducing their BKconductance. The reciprocal conversion is alsotrue: addition of an artificial BK conductanceto cultured gonadotrophs through thedynamic clamp technique changes their activ-ity pattern from spiking to bursting.19
Mathematical modeling predicted that theactivation time constant of the BK conduc-tance is important. BK activation must be fastto promote bursting; if too slow, then the BKcurrent does not promote bursting andinstead takes on its traditional inhibitory roleon electrical activity.19 Faster BK activationkinetics limit spike amplitude, preventing fullactivation of the repolarizing delayed rectifierK1 current, so membrane potential weaklyoscillates around a depolarized level14,22,23
before falling back toward rest, which resultsin bursting, increased Ca21 influx and globalCa21 signals.
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
29PATTERNS OF ANTERIOR PITUITARY CELL SIGNALING AND SECRETION
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

p0205 Unlike classical neuronal synapses, neuroen-docrine and endocrine cells require sustaineddepolarization to trigger the exocytotic pathway.In anterior pituitary cells, VGCC are open onlybriefly during the short time of a single AP, andthe elevated [Ca21]i is localized to the nanodo-mains that form at the inner mouths of openchannels. With longer durations and smallerburst amplitudes, VGCC stay open much longerand significant Ca21 influx ensues, resulting inthe summation of individual [Ca21]i nanodo-mains that generate a global Ca21 signal.Therefore, differences in the ability of spontane-ous firing patterns to generate global [Ca21]i sig-nals will determine their basal secretory activityFor example, the basal release of GH and pro-lactin is much higher than that of LH, which isconsistent with the pseudo-plateau burstingactivity of somatotrophs and lactotrophs thatresult in high-amplitude (global) [Ca21]i signalsand the short-duration APs of gonadotrophsthat evoke low-amplitude (local) [Ca21]i signals.
s0130 Receptor-Modulated Hormone Releaseand Voltage-Gated Ca21 InfluxAU:8 throughcAMP-Dependent Pathways
p0210 A number of hypothalamic neurohormonesand peripheral hormones control AP-drivenCa21 dynamics and Ca21-dependent hormonerelease in anterior pituitary cells through mod-ulation of cAMP-dependent pathways. Thesebind and activate a G protein-coupled receptor(GPCR) to modulate the basal activity of ade-nylyl cyclase (AC, the enzyme responsible forsynthesizing cAMP) and, in turn, Ca21 entry.Those that promote AP-driven Ca21 dynamicsand Ca21 dependent secretion bind to GPCRscoupled to the stimulatory class of G proteins(Gs) to increase AC activity through the activa-tion of the α subunit of the Gs protein, whereasthose that reduce Ca21 signaling and secretionbind to GPCRs coupled to the inhibitory classof G proteins (Gi/o) to decrease AC activity.
p0215The stimulatory Gs-signaling pathway is trig-gered by CRH receptors in corticotrophs, GHRHreceptors in somatotrophs and VIP/ AU:9pituitaryadenylate cyclase-activating polypeptide(PACAP) receptors in lactotrophs, somatotrophsand FS cells. Activation of the Gs-operatedGPCRs causes plasma membrane depolarization,increased electrical activity and Ca21 entry. Thetype of [Ca21]I response is a plateau elevation of[Ca21]I or an increase in the frequency and/oramplitude of [Ca21]I transients as elevatedcAMP levels promote, directly or indirectly, elec-trical activity and voltage-gated Ca21 influx AU:10
(VGCI). The direct pathway consists of the acti-vation of a background Na1 conductance thatresults upon binding of cAMP tohyperpolarization-activated and cyclicnucleotide-regulated (HCN) channels. HCNchannels are expressed in somatotrophs,24 lacto-trophs,25 gonadotrophs and thyrotrophs,12 andprobably corticotrophs.26 Because they are non-selective cation channels, they are likely to play arole in the initiation of the pacemaker depolari-zation. The indirect pathway consists of the acti-vation of cAMP-dependent kinases (PKAs) thatinduce phosphorylation-mediated modulation ofthe function of several plasma membrane ionchannels. In corticotrophs, CRH-induced PKAactivation inhibits Kir channels to promote slowdepolarization and enhanced excitability. Insomatotrophs, GHRH-stimulated PKA-mediated phosphorylation results in the openingof not only T-type and L-type Ca21 channels, butalso TTX-insensitive voltage-gated Na1 chan-nels, leading to the upstroke of a voltage spike.
p0220The inhibitory Gi/o-signaling pathway is trig-gered in lactotrophs by dopamine D2 andendothelin-1 ETA receptors, and in both lacto-trophs and somatotrophs by somatostatin sst1,sst2 and sst5 receptors. Other receptors linked tothis pathway and expressed by anterior pitui-tary cells include those for adenosine,γ-aminobutyric acid (GABA), serotonin, melato-nin and neuropeptide Y. Activation of this path-way opposes the actions mediated by the Gs-
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
30 2. BIOLOGY OF ANTERIOR PITUITARY CELLS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

signaling pathway resulting in membrane hyper-polarization, silencing of electrical activity andinhibition of Ca21 entry and Ca21-dependenthormone secretion. Gi/o-mediated inhibitoryactions, which can be irreversibly blocked byapplication of pertussis toxin, comprise twomajor signaling branches. The first one stemsfrom the Gi/o α-subunit-mediated inhibition ofAC activity, downregulating all cAMP-stimulated effects on electrical activity, Ca21
entry and secretion. The second is due to the acti-vation of Gi/o βγ dimers that activate Kir3 (alsoknown as G protein-gated inwardly rectifyingK1 channels, GIRK) and inhibit L-type Ca21
channels in a cAMP/PKA-independent fashion,leading to hyperpolarization and cessation of APfiring. Interestingly, three of the four mammalianKir3 channels are specifically induced by estra-diol in lactotrophs in proestrus,27 underlyingdopamine effects that are only observed in thisstage of the cycle: strong Kir conductance androbust hyperpolarization, the latter playing a crit-ical role in the prolactin secretory rebound thatfollows dopamine withdrawal.28 A novel mecha-nism has been recently described for endothelinETA and dopamine D2 receptors in lactotrophs.In addition to the Gi/o class of G proteins, thesereceptors couple to Gz proteins, a subfamily ofGi/o proteins that are insensitive to pertussistoxin. Activation of the α subunit and βγ dimersof Gz by these receptors potently block VGCI andprolactin release by inhibiting AC activity andthe exocytotic machinery responsible forsecretion.29,30
s0135 Receptor-Modulated Hormone Releasethrough Ca21-Mobilizing Pathways
p0225 All anterior pituitary cell types expressCa21-mobilizing GPCRs. Examples in gonado-trophs include the receptors for GnRH,endothelins, PACAP and substance P; in lacto-trophs such receptors include those for acetyl-choline, angiotensin II, TRH, oxytocin, ATP,
endothelin, serotonin, galanin and substance P;somatotrophs express ghrelin and endothelinreceptors; in corticotrophs, AVP and norepi-nephrine receptors; and in thyrotrophs, TRH(the main thyrotroph secretagogues) andendothelin elicit Ca21 mobilization. When acti-vated, these Ca21-mobilizing GPCRs couple tothe Gq protein class of heterotrimeric G pro-teins, eliciting the dissociation of the αq subunitthat triggers phospholipase C-mediated phos-phoinositide hydrolysis resulting in the forma-tion of inositol 1,4,5 triphosphate (IP3) anddiacylglycerol (DAG) (Figure 2.2). IP3 binds toIP3 AU:11receptors expressed in the membrane of theendoplasmic reticulum (ER), the primary store-house of Ca21 in most cells, causing a mobiliza-tion of Ca21 out of this compartment andleading to a large and fast increase of [Ca21]I.
31
p0230Two different Ca21 signaling patterns can beobserved after the initial [Ca21]I spike. The firstone, termed “biphasic,” is a non-oscillatory pat-tern found in lactotrophs, somatotrophs andthyrotrophs where the transient [Ca21]I spike isfollowed by a slow decline to a plateau level thatis above basal. However, some cells exhibit“monophasic” responses where only the spike orthe plateau is observed. A key condition of thebiphasic Ca21 response is that the IP3Rs areopened continuously throughout the time of ago-nist application. The microdomain of Ca21 thatforms near the mouth of the IP3 receptor duringthe Ca21 spike is high enough to trigger exocyto-sis. Furthermore, the transient [Ca21]I surge acti-vates the small-conductance Ca21-activated K1
channels (SK) and hyperpolarizes the plasmamembrane, terminating any previous electricalactivity.10,32 As the Ca21 is removed from thecytosol, by means of plasma membrane ATPasepumps and Na1/Ca21 exchangers as well assarco-endoplasmic reticulum Ca21 ATPase(SERCA) pumps, the SK channels close and themembrane depolarizes. Depletion of the ER Ca21
store also provides a signal for the activation oftransient receptor potential cation channels AU:12
(TRPCs) and other store-operated Ca21 channels
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
31PATTERNS OF ANTERIOR PITUITARY CELL SIGNALING AND SECRETION
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

(SOCs) important for depolarization.Diacylglycerol may directly activate TRP chan-nels or may inhibit Kir currents through proteinkinase C (PKC)-dependent phosphorylation(Figure 2.2). In addition to these events, downre-gulation of an M/ERG channel via PLC-mediated PiP2 depletion leads to a sustaineddepolarization phase that activates voltage-gatedCa21 channels, further depolarizing the cell andinitiating single spiking or bursting (Figure 2.2).The resulting Ca21 entry refills the ER Ca21 store,enabling continued Ca21 signaling.
p0235 In contrast to the biphasic Ca21 responsedescribed above, GnRH-stimulated gonado-trophs and norepinephrine-stimulated cortico-trophs engage in an oscillatory Ca21 responseafter the initial transient of [Ca21]I. Interestingly,oscillations in IP3 are not required to generateoscillatory Ca21 release in gonadotrophs. The IP3receptor itself is the source of the oscillation, sinceit is activated by cytosolic Ca21 on a fast timescale and inhibited on a longer time scale. Thedelayed inhibition of the channel produces oscil-lations.33 This is a true ER-mediated oscillation,as it can be produced even when the membrane
potential is clamped at a voltage sufficiently highto allow Ca21 influx that refills the ER betweeneach bout of Ca21 release.34 In the unclampedcell, a key feature of the oscillatory response isthe antiphasic pattern of electrical activity andCa21 release due to the SK channel-mediatedinhibitory action of each Ca21 pulse on theplasma membrane (see ref10). Once [Ca21]Ireturns to a low level following the Ca21 pulse,firing resumes. The electrical activity and secre-tion are out of phase; the former serves to refillthe ER Ca21 store that periodically releases Ca21
and evokes secretion during sustainedstimulation.
s0140PARACRINE (LOCAL)REGULATION AND
INTERCELLULAR SIGNALINGWITHIN THE ANTERIOR
PITUITARY
p0240More than 100 bioactive substances areexpressed by cells of the anterior pituitary andcan act within the gland through specific
TRPTRP KirKv HCNHCN
ergM
PP
α–90
0
0
1
80
0
β
γ
SKSK
Ca2+
IP3
K+ K+ K+ K+
PLC
DAG
PKC
PIP2
Ca2+
IP3 Sensitive Pool
ER
Pum
p
Na+/K+ Na+/K+
Vm (mV)
[Ca2+]i (μM)
[LH], (ng/ml)
(A) (B)Gq/11-coupled GPCRs
f0015 FIGURE 2.2 (A) Mechanism of receptor-induced electrical activity and Ca21 signaling by activated Gq- coupled, Ca21-
mobilizing receptors. (B) Schematic representation of a biphasic response typically observed in gonadotrophs uponGnRH-induced stimulation of electrical activity (top), cytosolic Ca21 (middle) and hormone secretion (bottom). The gener-ation of a biphasic response reflects the tonic activation of IP3 receptors (see text for details).
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
32 2. BIOLOGY OF ANTERIOR PITUITARY CELLS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

receptors, allowing these messengers to exert alocal regulatory function during specific physi-ological states. Depending on the cellular loca-tion of both the signaling molecule and itsrespective receptor, such messengers may acteither on the same cell (autocrine control), anadjacent cell (juxtacrine control), a neighboringcell (paracrine control) or even within thesame cell where the signaling molecule isproduced without being secreted (intracrinecontrol). Such interactions are highly context-dependent and constitute the cellular basis oflocally controlled anterior pituitary plasticity.In general, they either promote or inhibit hor-mone release and cell proliferation, and areswitched on/off when hormonal outputs needto be adapted to changing demands of theorganism, such as during mating, pregnancy,lactation, stress, inflammation, immuneresponses, starvation and circadian rhythms.We will briefly highlight physiologically rele-vant paracrine and autocrine interactions thathave been substantiated with reasonable evi-dence. Comprehensive reviews on this topichave been recently published.17,35
p0245 Well-established autocrine mechanisms inlactotrophs include the stimulatory loops ofVIP, galanin and TGF-α as well as the inhibi-tory loops of TGF-β1 and endothelins. In gona-dotrophs, the autocrine loop made up byactivin B (stimulatory) and follistatin (inhibi-tory), along with that of inhibin (inhibitory),constitute one of the fundamental mechanismsfor selective regulation of FSH expression andsecretion, as changes in GnRH pulse frequencycause changes of expression of these threemodulators that in turn change the FSH:LHratio.36 Nitric oxide (NO) is also an importantautocrine/paracrine modulator of gonado-trophs as both these cells and FS cells expressnitric oxide synthase (NOS). NO may playboth inhibitory and stimulatory roles, and thismight depend on the cellular source of NO, asNO from FS cells seems to inhibit GnRH-stimulated LH release whereas gonadotroph
NO stimulates basal LH and FSH secretion.PACAP also plays autocrine roles in gonado-trophs where it is specifically expressed in pro-estrus to stimulate LH release throughinteraction with the gonadotroph NO systemand GnRH signaling pathways. The purineATP has been shown to be costored with hor-mones in secretory granules and cosecreted inall endocrine cell types of the anterior pitui-tary,37 but autocrine roles have only been con-sistently shown in gonadotrophs where itpotently stimulates basal LH release. In corti-cotrophs, expression of CRH and AVP (whichhas also been detected in all endocrine celltypes except somatotrophs) and their receptorsunderlies an autocrine mechanism by whichCRH and AVP may contribute to the well-established potentiation of ACTH release byhypothalamic CRH and AVP. In somatotrophs,ghrelin has been shown to sensitize the cell toGHRH at early postnatal age (and possibly atpuberty).38 In thyrotrophs, stimulatory andinhibitory autocrine roles for leptin and neuro-medin B, respectively, have been shown to beimportant in the control of TSH secretion dur-ing adaptation to nutritional status.
p0250The majority of the paracrine interactionsreported so far are those of gonadotrophs withdifferent endocrine cell types of the anteriorpituitary, particularly lactotrophs, somato-trophs and corticotrophs. Among these, one ofthe first described is the GnRH-stimulated pro-lactin release by lactotrophs,39 an effect that ismediated by the release of a still uncharacter-ized molecule from immature postnatal gona-dotrophs.40 In immature cells, GnRH alsoelicits a biphasic hormone release response onsomatotrophs that begins with inhibition ofgrowth hormone secretion during GnRHapplication and is followed by a rapid reboundsecretion of GH that slowly returns to basallevels. These interactions might be partiallyrelated to developmental roles that GnRH mayhave on these cell types. Some of the candidateparacrine factors from gonadotrophs thought
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
33PARACRINE (LOCAL) REGULATION AND INTERCELLULAR SIGNALING WITHIN THE ANTERIOR PITUITARY
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

to be involved in the development of lacto-trophs are the glycoprotein hormone α-sub-unit, the growth factor TGF-α and prolactin-releasing peptide (PrRP). Stimulatoryparacrine interactions must be counterbalancedwith inhibitory ones for the gland to meet andmaintain homeostasis throughout differentphysiological stages; one good example of neg-ative interaction between gonadotrophs andlactotrophs is that mediated by calcitonin (andcalcitonin-like peptides). Calcitonin immunore-activity is primarily located in gonadotrophsthat are associated with cup-shaped lactotrophs(described later in this section). It inhibits basaland TRH-stimulated prolactin synthesis andrelease as well as lactotroph mitosis. This para-crine effect is likely mediated by the release ofTGF-β1 by lactotrophs, which in turn inhibitslactotroph proliferation as well as prolactinexpression and secretion.41 Consistent withthis role, estradiol, a well-known promoter oflactotroph secretory activity and proliferation,negatively regulates calcitonin expression ingonadotrophs. Positive and negative interac-tions have also been described betweengonadotrophs and corticotrophs; calcitoningene-related peptide accounts for the formerwhereas the natriuretic peptides (ANP andCNP) and adrenomedullin (a calcitonin pep-tide family member) account for the latter.Since stress responses are attenuated in preg-nancy and lactation, it is likely that the inhibi-tory tone of natriuretic peptides oncorticotroph function is upregulated duringthese states of elevated estradiol levels inplasma as estrogens are known to upregulatenatriuretic peptide expression in heart tissue.
p0255 Gonadotrophs can also be paracrine targetsthemselves. Although inhibition of ovulationduring lactation is achieved primary byendorphin-mediated inhibition of hypotha-lamic GnRH neurons triggered by the suck-ling stimulus, local inhibitory cues fromlactotrophs and corticotrophs to gonado-trophs at the anterior pituitary may contribute
to the suckling-induced negative influence onovulation. Increased release of β-endorphinand galanin by corticotrophs and lactotrophs,respectively, may contribute to inhibitpreovulatory LH secretion. In pregnancy,estrogen-induced high galanin release by lac-totrophs underlies increased lactotrophactivity and growth as well as decreasedLH release. During stress and undernutrition,stress-induced activation of thehypothalamo�pituitary�adrenal ( AU:13HPA) axisinhibits the hypothalamo�pituitary�gonadal(HPG) axis. At the pituitary level, this nega-tive influence of the HPA onto the HPG axismay be mediated by the negative paracrinesignals of corticotroph β-endorphin on GnRH-induced gonadotroph LH release.
p0260At the heart of the structure and functionof the anterior pituitary gland is the key sup-portive and dynamic role of the FS cells.These cells are also excitable and are thoughtto coordinate activity of endocrine cells. Theyform two microanatomical structures thatmay have a large impact on pituitary cellphysiology. Located in the center of the hor-monal cell cord, they are often arranged inclusters and form small follicles in rats, butare larger in humans and some other species.In the follicles, numerous microvilli protrudeand some cilia are present. Follicle-forming FScells are polarized. At the apical pole, border-ing the follicle, they form tight junctionsamong each other, although not always fullysealed, and, more laterally, junctions of the“zonula adherens” type (desmosomes). Thebasolateral side makes contact with the endo-crine cells and with other FS cells, andextends processes that end on the basal mem-brane surrounding the cell cords. The role offollicles remains unclear but the structures arethought to be involved in intercellular trans-port of metabolic products and ions. A secondgroup of FS cells extends long processesbetween the hormonal cell types within eachglandular cell cord. Although these processes
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
34 2. BIOLOGY OF ANTERIOR PITUITARY CELLS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

form intercellular junctions among each other,mostly of the zonula adherens-type, they arealso electrotonically coupled through gapjunctions as shown by rapid propagation ofCa21 currents over long distances in thegland.42 On this basis, it is hypothesized thatthese cells coordinate the activity of endocrinecells. In support of this is the finding of a cor-relation between the number of gap junctionsand reproductive maturation in the rat.Interestingly, estradiol seems to increase FSnetwork connectivity, as a steep rise in gapjunction number is observed at the end ofpregnancy and during lactation. Moreover, inthe estrous cycle, connectivity is highest dur-ing proestrus and estrus, and it has beenrecently suggested that this increased connec-tivity plays a significant role in the preovula-tory LH surge.43 Some FS cells make intimatefoot processes with the basal membrane of theextra-vascular spaces at the periphery of thecell cords. In some species FS cells located inthe periphery of the cell cords are juxtaposedin a way that they form sinusoid-like spaces.Intercellular lacunae are also often seenbetween endocrine cells. These lacunae, alongwith the sinusoid-like spaces surrounded byFS cells and perivascular spaces are thoughtto form a micro-channel system within thepituitary, through which hormones, local fac-tors, nutrients, ions and waste products cancirculate. The three-dimensional architectureof FS cells is under developmental control: inthe infant rat, the FS follicles are elongatedand participating FS cells have a columnarshape without cellular extensions and dis-playing very few junctions. At the onset ofpuberty, they separate into smaller follicularunits and start making extensions and junc-tions, especially tight junctions.
p0265 It is thus reasonable to hypothesize that FScells play key roles in anterior pituitary adap-tation to varying physiological conditions,including immune, nutritional and other stres-ses in which they may operate as critical
interfaces in homeostatic mechanisms.Prolonged pituitary activation during immunestress, pregnancy, lactation, starvation andother conditions, may lead to excess produc-tion of specific hormones that in turn mayresult in inhibition of essential physiologicalprocesses. In such events, it is likely that FScells provide a mechanism to circumvent thisissue through their capability to blunt manystimulated activities in the anterior pituitary.FS cells release NO that in turn may stimulateguanylyl cyclase activity and increase cyclicguanosine monophosphate (cGMP) in differentendocrine cell types to inhibit hormone release(Figure 2.3). In addition, FS cells are permis-sive for the mitogenic effect of estradiol on lac-totrophs by releasing fibroblast growth factor-2(FGF-2), and mediate the stimulatory action ofbacterial endotoxin lipopolysaccharide (LPS),tumor necrosis factor α (TNF-α), VIP, PACAPand interleukin-1 (IL-1) on ACTH secretion byreleasing IL-6 which act in corticotrophs in aparacrine fashion (Figure 2.3). FS cells maypossibly modulate responses to immune stressas they express receptors for epinephrine, ace-tylcholine, angiotensin II, calcitonin and ATPthat are known to regulate immune cell func-tions. Indeed, FS cells also express glucocorti-coid receptors and have been shown tomediate the glucocorticoid fast negative feed-back effect on ACTH (and also prolactin andGH) secretion via ATP-binding cassette (ABC)transporter-mediated externalization of annex-in 1, which in turn acts on specific bindingsites on endocrine cells to inhibit hormonerelease44 (Figure 2.3).
s0145PLASTICITY OF ANTERIORPITUITARY CELL NETWORK
ORGANIZATION AND FUNCTION
p0270The architecture of the anterior lobe is basedon interlacing strands of mostly polygonalcells separated by a tremendously developed
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
35PLASTICITY OF ANTERIOR PITUITARY CELL NETWORK ORGANIZATION AND FUNCTION
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

array of sinusoidal capillaries. Early descrip-tions of the microanatomy of the adenohy-pophysis suggested that its various cell typesare intermingled, forming cell networks(reviewed in refs35,42). Nakane later made theseminal observation that the different pituitarycell types are not distributed homogeneouslyover the various areas of the gland and withina specific cell cord.45 In fact, a recent studyusing two-photon microscopy imaging ofgenetically engineered mouse somatotrophsexpressing the enhanced green-fluorescentprotein (EGFP) showed that GH-producingcells are interconnected via adherens junctions,seeming to form a “homotypic connected 3Dcell continuum” that displays coordinatedCa21 transients.46 This cell system consists ofnumerous intercrossing strands of single GHcells, with larger clusters of GH cells
positioned at the intersection of the cords.Consistent with Nakane’s observation, thereare differences in the GH cell strand and clus-ter densities in different regions of the gland.Across the lifespan, the GH system architec-ture shows plasticity.46 In prepubertal animals,the patterning of the GH cell system is similarbetween the lateral and the median zones ofthe gland. From puberty to adulthood there isa marked increase in the proportion of GHcells in clusters in the lateral portions of thegland than in the median zone surroundingthe stalk. Interestingly, the volume-to-surfaceratios of the GH cell system returns to prepu-bertal values in the lateral zones of aged mice,indicating that the plasticity of this cell systemcontinues well into adulthood.46 More recently,it has been shown that gonadotrophs exhibitsignificantly different distributions across
GHRH
CRHABC -A1
Glucocorticoid
IL-1
TNF-α
LPS
MIF
IL-6 iNOS
NO
GC
GTP
TRH
PRL
GC
GTP
cGMP
LH
GnRH
GH
ACTH
GC
GC GTP
cGMPPOMCmRNA
cGMP
GTP
cGMP
ATP
CalcitoninVIP
PACAPAcetylcholineEpinephrine
Angiotensin II
FS cell
FGFnNOS
Annex
in-1
MITOSIS
Estradiol
f0020 FIGURE 2.3 Schematic representation of some of the paracrine interactions thought to occur between folliclostellate (FS)cells, lactotrophs, somatotrophs, gonadotrophs and corticotrophs. -, stimulatory effect; j, inhibitory effect (see text fordetails).
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
36 2. BIOLOGY OF ANTERIOR PITUITARY CELLS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

physiological states,47 so it is likely that othercell types share a similar cell system architec-ture and plasticity. In fact, Nakane hadobserved close associations between somato-trophs and corticotrophs and between gonado-trophs and lactotrophs. Some of thelactotrophs embraced the oval-shaped gonado-trophs with long cellular processes andNakane named them “cup-shaped” PRLcells.45 These cell�cell associations are thoughtto have functional consequences, not only inthe cells involved in the association, but also inthe pituitary gland as a whole. That the differ-ent cell types are both structurally and func-tionally interconnected is confirmed by the factthat genetic ablation of GH cells dramaticallyreduced the pituitary content of allhormones.48
p0275 Finally, the ample possibilities broughtabout by interconnectivity at the cellular net-work level may offer emergent properties thatmight be critical to anterior pituitary cells toadapt their physiology to suit the prevailingenvironmental conditions. As interconnectedcell populations bestowed with the role ofmounting large-scale responses to maintainhomeostasis, experience-dependent plasticitymay represent an inherent property of thesecells. This concept has been recently illustratedin female mice that underwent one or morelactations to repeatedly and selectively stimu-late activity of pituitary lactotrophs.49
Throughout the lactotroph population, eachlactation event induced alterations in func-tional connectivity through changes in struc-tural connectivity mediated by differentialhomotypic lactotroph�lactotroph gap junc-tional contacts. Following weaning, the lacto-troph population is able to maintain thepattern of functional connectivity for weeks tomonths, a hallmark of long-term experience-dependent plasticity that is reminiscent to thatobserved in neurons. Likewise, the lactotrophnetwork retains functional connectivitythrough changes in the extent and strength of
cell�cell communication, allowing repeatedlactation demands to be met with evolved net-work dynamics and improved tissue output.49
Thus, experience-dependent plasticity allowslactotrophs to adapt their hormone releasingactivity upon repeated stimulation, a featurethat is likely shared by other endocrine cellswithin the anterior pituitary and, possibly,throughout the body.
s0150CONCLUDING REMARKS
p0280Endocrine cells of the anterior pituitaryhave a remarkably varied signaling toolkit attheir disposal which, in combination withGPCR expression profiles, and gap junctionalelectrical coupling, may allow incoming sig-nals to be processed and propagated from cellto cell in a specific manner. Not only are thesefeatures subject to dynamic changes accordingto physiological needs, but also the morpho-logical arrangement and functional connectiv-ity of the different cell types of this gland canalso go through profound remodeling. Thesefundamental features allow anterior pituitarycells to adapt their responses to environmentalchallenges and demands and thereby to meettheir vital role in homeostatic control.
s0155Acknowledgements
p0285The authors wish to gratefully acknowledge Jose Arias-Cristancho for preparing Figures 2 and 3 of this chapter aswell as the support provided by National Institute ofHealth grant DK-43200.
References
1. Gonzalez-Iglesias AE, Freeman ME. Brain control overpituitary gland hormones. In: Pfaff DW, editor.Neuroscience in the 21st Century. New York: Springer-Verlag; 2012.
2. Ooi GT, Tawadros N, Escalona RM. Pituitary cell linesand their endocrine applications. Mol Cell Endocrinol2004;228:1�21.
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
37REFERENCES
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

3. Walker JJ, Terry JR, Tsaneva-Atanasova K, ArmstrongSP, Mc Ardle CA, Lightman SL. Encoding and decod-ing mechanisms of pulsatile hormone secretion. JNeuroendocrinol 2010;22:1226�38.
4. Melmed S. Update in pituitary disease. J ClinEndocrinol Metab 2008;93:331�8.
5. Perez-Castro C, Renner U, Haedo MR, Stalla GK, ArztE. Cellular and molecular specificity of pituitary glandphysiology. Physiol Rev 2012;92:1�38.
6. Roudbaraki M, Lorsignol A, Langouche L,Callewaert G, Vankelecom H, Denef C. Target cellsof gamma3-melanocyte-stimulating hormonedetected through intracellular Ca21 responses ininmature rat pituitary constitute a fraction of allmain pituitary cell types, but mostly express multi-ple hormone phenotypes at the messenger ribonu-cleic acid level. Refractoriness to melanocortin-3receptor blockade in the lacto-somatotroph lineage.Endocrinology 1999;140:4874�85.
7. Villalobos C, Nunez L, Frawley S, Garcıa-Sancho J,Sanchez A. Multi-responsiveness of single anteriorpituitary cells to hypothalamic-releasing hormones: acellular basis for paradoxical secretion. Proc Natl AcadSci USA 1997;94:14132�7.
8. Childs GV. Development of gonadotropes may involvecyclic transdifferentiation of growth hormone cells.Arch Physiol Biochem 2002;110:42�9.
9. Stojilkovic SS. Molecular mechanisms of pituitaryendocrine cell calcium handling. Cell Calcium2012;51:212�21.
10. Stojilkovic SS, Tabak J, Bertram R. Ion channels andsignaling in the pituitary gland. Endocr Rev2010;31:845�915.
11. van Goor F, Zivadinovic D, Martınez-Fuentes AJ,Stojilkovic SS. Dependence of pituitary hormone secre-tion on the pattern of spontaneous voltage-gated cal-cium influx. Cell type-specific action potentialsecretion coupling. J Biol Chem 2001;276:33840�6.
12. Kretschmannova K, Kucka M, Gonzalez-Iglesias AE,Stojilkovic SS. The expression and role ofhyperpolarization-activated and cyclic nucleotide-gated channels in endocrine anterior pituitary cells.Mol Endocrinol 2012;26:153�64.
13. Liang Z, Chen L, McClafferty H, Lukowski R,MacGregor D, King JT, et al. Control of hypothalamic-pituitary-adrenal stress axis activity by the intermedi-ate conductance calcium-activated potassium channel,SK4. J Physiol 2011;589:5965�86.
14. Tsaneva-Atanasova K, Sherman A, van Goor F,Stojilkovic SS. Mechanism of spontaneous andreceptor-controlled electrical activity in pituitary soma-totrophs: experiments and theory. J Neurophysiol2007;98:131�44.
15. Gonzalez-Iglesias AE, Jiang Y, Tomic M,Kretschmannova K, Andric SA, Zemkova H, et al.Dependence of electrical activity and calcium influx-controlled prolactin release on adenylyl cyclase signal-ing pathway in pituitary lactotrophs. Mol Endocrinol2006;20:2231�46.
16. Tougard C, Tixier-Vidal A. Lactotropes and gonado-tropes. In: Knobil E, Neill JD, editors. The physiology ofreproduction. New York: Raven Press Ltd; 1994. p.1711�47.
17. Childs GV. Gonadotropes and lactotropes. In: Neill JD,editor. Knobil and neill’s physiology of reproduction. 3rded. New York: Raven Press Ltd; 2006. . p. 1483�562..
18. Christian HC, Chapman LP, Morris JF. Thyrotrophin-releasing hormone, vasoactive intestinal peptide,prolactin-releasing peptide and dopamine regulationof prolactin secretion by different lactotroph morpho-logical subtypes in the rat. J Neuroendocrinol2007;19:605�13.
19. Tabak J, Tomaiuolo M, Gonzalez-Iglesias AE, MilescuLS, Bertram R. Fast-activating voltage- and calcium-dependent potassium (BK) conductance promotesbursting in pituitary cells: a dynamic clamp study.J Neurosci 2011;31:16855�63.
20. Tomaiuolo M, Bertram R, Gonzalez-Iglesias AE, TabakJ. Investigating heterogeneity of intracellular calciumdynamics in anterior pituitary lactotrophs usign a com-bined modelling/experimental approach.J Neuroendocrinol 2010;22:1279�89.
21. van Goor F, Zivadinovic D, Stojilkovic SS. Differentialexpression of ionic channels in rat anterior pituitarycells. Mol Endocrinol 2001;15:1222�36.
22. van Goor F, Li YX, Stojilkovic SS. Paradoxical role oflarge-conductance calcium-activated K1 (BK) channelsin controlling action potential-driven Ca21 entry inanterior pituitary cells. J Neurosci 2001;21:5902�15.
23. Tabak J, Toporikova N, Freeman ME, Bertram R. Lowdose of dopamine may stimulate prolactin secretion byincreasing fast potassium currents. J Comput Neurosci2007;22:211�22.
24. Simasko SM, Sankaranarayanan S. Characterization ofa hyperpolarization-activated cation current in rat pitu-itary cells. Am J Physiol � Endocrinol Metab 1997;272:E405�14.
25. Gonzalez-Iglesias AE, Kretschmannova K, Tomic M,Stojilkovic SS. ZD7288 inhibits exocytosis in an HCN-independent manner and downstream of voltage-gatedcalcium influx in pituitary lactotrophs. Biochem BiophysRes Commun 2006;346:845�50.
26. Tian L, Shipston MJ. Characterization ofhyperpolarization-activated cation currents in mouseanterior pituitary, AtT20 D16:16 corticotropes.Endocrinology 2000;141:2930�7.
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
38 2. BIOLOGY OF ANTERIOR PITUITARY CELLS
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.

27. Christensen HR, Zeng Q, Murawsky MK, GregersonKA. Estrogen regulation of the dopamine-activatedGIRK channel in pituitary lactotrophs: implications forregulation of prolactin release during the estrous cycle.Am J Physiol � Regul Integr Comp Physiol 2011;301:R746�56.
28. Gregerson KA. Functional expression of the dopamine-activated K1 current in lactotrophs during the estrouscycle in female rats. Endocrine 2003;20:67�74.
29. Andric SA, Zivadinovic D, Gonzalez-Iglesias AE,Lachowicz A, Tomic M, Stojilkovic SS. Endothelin-induced, long lasting, and Ca21 influx-independentblockade of intrinsic secretion in pituitary cells by Gzsubunits. J Biol Chem 2005;280:26896�903.
30. Gonzalez-Iglesias AE, Murano T, Li S, Tomic M,Stojilkovic SS. Dopamine inhibits basal prolactinrelease in pituitary lactotrophs through pertussis toxin-sensitive and -insensitive signaling pathways.Endocrinology 2008;149:1470�9.
31. Berridge MJ. Neuronal signalling. Cell signaling biology.Portland Press Limited; 2012. p. 10.1�10.104
32. Li YX, Rinzel J, Keizer J, Stojilkovic SS. Calcium oscilla-tions in pituitary gonadotrophs: comparison of experi-ment and theory. Proc Natl Acad Sci USA1994;91:58�62.
33. Vergara LA, Stojilkovic SS, Rojas E. GnRH-inducedcytosolic calcium oscillations in pituitary gonado-trophs: phase resetting by membrane depolarization.Biophys J 1995;69:1606�14.
34. Kukuljan M, Vergara L, Stojilkovic SS. Modulation ofthe kinetics of inositol 1,4,5-trisphosphate-induced[Ca21 ]i oscillations by calcium entry in pituitary gona-dotrophs. Biophys J 1997;72(2 Pt 1):698�707.
35. Denef C. Paracrinicity: the story of 30 years of cellularpituitary crosstalk. J Neuroendocrinol 2008;20:1�70.
36. Kirk SE, Dalkin AC, Yasin M, Haisenleder DJ, MarshallJC. Gonadotropin-releasing hormone pulse frequencyregulates expression of pituitary follistatin in folliculos-tellate cell-enriched primate pituitary cell cultures.Endocrinology 1994;135:876�80.
37. Stojilkovic SS, He ML, Koshimizu TA, Balik A,Zemkova H. Signaling by purinergic receptors andchannels in the pituitary gland. Mol Cell Endocrinol2010;314:184�91.
38. Szabo M, Cuttler L. Differential responsiveness of thesomatotroph to growth hormone-releasing factor
during early neonatal development in the rat.Endocrinology 1986;118:69�73.
39. Denef C, Andries M. Evidence for paracrine interactionbetween gonadotrophs and lactotrophs in pituitary cellaggregates. Endocrinology 1983;112:813�22.
40. Andries M, Denef C. Gonadotropin-releasing hormoneinfluences the release of prolactin and growth hormonefrom intact rat pituitary in vitro during a limited periodin neonatal life. Peptides 1995;16:527�32.
41. Sarkar DK, Kim KH, Minami S. Transforming growthfactor-beta 1 messenger RNA and protein expressionin the pituitary gland: its action on prolactin secretionand lactotropic growth. Mol Endocrinol 1992;6:1825�33.
42. Hodson DJ, Romano N, Schaeffer M, Fontanaud P,Lafont C, Fiordelisio T, et al. Coordination of calciumsignals by pituitary endocrine cells in situ. Cell Calcium2012;51:222�30.
43. Lyles D, Tien JH, McCobb DP, Zeeman ML. Pituitarynetwork connectivity as a mechanism for the lutei-nizing hormone surge. J Neuroendocrinol2010;22:1267�78.
44. Morris JF, Christian HC, Chapman LP, Epton MJ,Buckingham JC, Ozawa H, et al. Steroid effects onsecretion from subsets of lactotrophs: role of Folliculo-Stellate cells and Annexin 1. Arch Physiol Biochem2002;110:54�61.
45. Nakane PK. Classifications of anterior pituitary celltypes with immuno-enzyme histochemistry. JHistochem Cytochem 1970;18:9�20.
46. Bonnefont X, Lacampagne A, Sanchez-Hormigo A,Fino E, Creff A, Mathieu MN, et al. Revealing thelarge-scale network organization of growth hormone-secreting cells. Proc Natl Acad Sci USA2005;102:16880�5.
47. Alim Z, Hartshorn C, Mai O, Stitt I, Clay C, Tobet S, etal. Gonadotrope plasticity at cellular and populationlevels. Endocrinology 2012;153:4729�39.
48. Waite E, Lafont C, Carmignac D, Chauvet N, CoutryN, Christian H, et al. Different degrees of somatotrophablation compromise pituitary growth hormone cellnetwork structure and other pituitary endocrine celltypes. Endocrinology 2010;151:234�43.
49. Hodson DJ, Schaeffer M, Romano N, Fontanaud P,Lafont C, Birkenstock J, et al. Existence of long-lastingexperience-dependent plasticity in endocrine cell net-works. Nat Commun 2012;3:605.
CELLULAR ENDOCRINOLOGY IN HEALTH AND DISEASE
39REFERENCES
Aguirre-1611032 978-0-12-408134-5 00002
To protect the rights of the author(s) and publisher we inform you that this PDF is an uncorrected proof for internal business use only by the author(s), editor(s),reviewer(s), Elsevier and typesetter MPS. It is not allowed to publish this proof online or in print. This proof copy is the copyright property of the publisher andis confidential until formal publication.