the language of the cerebellum - hse.ru manto, marien_2016... · cerebellum in a broad spectrum of...
TRANSCRIPT

Full Terms & Conditions of access and use can be found athttp://www.tandfonline.com/action/journalInformation?journalCode=paph20
Download by: [University of Groningen] Date: 17 October 2016, At: 02:04
Aphasiology
ISSN: 0268-7038 (Print) 1464-5041 (Online) Journal homepage: http://www.tandfonline.com/loi/paph20
The language of the cerebellum
Kim van Dun, Mario Manto & Peter Mariën
To cite this article: Kim van Dun, Mario Manto & Peter Mariën (2016) The language of thecerebellum, Aphasiology, 30:12, 1378-1398, DOI: 10.1080/02687038.2015.1132297
To link to this article: http://dx.doi.org/10.1080/02687038.2015.1132297
Published online: 12 Jan 2016.
Submit your article to this journal
Article views: 218
View related articles
View Crossmark data

The language of the cerebellumKim van Duna, Mario Mantob and Peter Mariëna,c
aClinical and Experimental Neurolinguistics, Vrije Universiteit Brussel, Brussels, Belgium; bUnité d’Étude duMouvement, FNRS Neurologie, ULB Erasme, Brussels, Belgium; cDepartment of Neurology and MemoryClinic, ZNA Middelheim, Antwerp, Belgium
ABSTRACTBackground: During the past three decades neuroanatomical, neu-roimaging, and clinical studies have substantially altered the viewon the role of the cerebellum as a sole coordinator of sensorimotorfunction. Currently, the cerebellum is believed to be also cruciallyinvolved in cognitive, affective, and behavioural functioning.Aims: This paper aims to summarise a number of critical insightsfrom different research areas (anatomy, functional imaging, clinicalpractice) that provide evidence for a role of the cerebellum inmotor speech and nonmotor language processing.Main contribution: By means of identifying a dense network ofcrossed reciprocal connections between the cerebellum and thesupratentorial association areas, neuroanatomical studies pro-vided a robust basis for the development of new insights in themodulatory role of the cerebellum in neurocognition, includingnonmotor language processing. A topological distinction wasestablished between the “motor” cerebellum, projecting to thecortical motor areas, and the “cognitive/affective” cerebellum,connected with the cortical and limbic association areas.Neuroimaging studies demonstrated cerebellar involvement inseveral different language tasks, even after controlling formotor aspects. In addition, several clinical studies identified avariety of nonmotor linguistic deficits after cerebellar damage,implying a prominent role for the cerebellum in these linguisticprocesses. Functional neuroimaging confirmed the functionalimpact of cerebellar lesions on remote, structurally intact corticalregions via crossed cerebello-cerebral diaschisis.Conclusion: Evidence from neuroanatomical, neuroimaging, andclinical studies suggests a strongly lateralised involvement of thecerebellum in a broad spectrum of nonmotor language functionsthrough crossed cerebello-cerebral connectivity. It is argued thatthe cerebellum is involved in language in a similar manner as it isinvolved in motor functions: through monitoring/coordinatingcortical functions.
ARTICLE HISTORYReceived 2 April 2015Accepted 12 December 2015
KEYWORDSCerebellum; language;diaschisis; Schmahmann’ssyndrome; timinghypothesis; sequencinghypothesis
Introduction
For centuries, the role of the cerebellum was considered to consist of pure sensorimotorcontrol (Manto & Mariën, 2015). In the last few decades, however, neuroanatomical,
CONTACT Peter Mariën [email protected]
APHASIOLOGY, 2016VOL. 30, NO. 12, 1378–1398http://dx.doi.org/10.1080/02687038.2015.1132297
© 2016 Taylor & Francis

neuroimaging, and clinical studies have changed this view substantially. The cerebellumis now believed to play a prominent role in cognitive, affective, and behavioural func-tions, including language (De Smet, Paquier, Verhoeven, & Mariën, 2013; Mariën et al.,2014). In this paper, the rapidly growing evidence derived from a number of neuroana-tomical, neuroimaging, and clinical studies is summarised to demonstrate crucial invol-vement of the cerebellum in a wide variety of nonmotor linguistic processes. In addition,current hypotheses about the possible underlying pathophysiological and cognitivemechanisms will be briefly discussed.
Evidence for a role for the cerebellum in language
Anatomical
Neuroanatomically, the cerebellum is not only densely interconnected with the supra-tentorial motor areas, but also with the paralimbic and association cortices throughreciprocal crossed cerebello-cerebral pathways. The cerebellum receives input fromthe cerebral cortical regions via cortico-ponto-cerebellar projections, while feedback isprovided via cerebello-thalamo-cortical projections (Mariën et al., 2014). Histologicaltract tracing studies in monkeys have shown that these feedforward and feedbackprojections form closed loops: a cerebellar area projects back to the cortical areas fromwhich it receives input (Strick, Dum, & Fiez, 2009) (Figure 1).
In addition, neuroanatomical and neurophysiological studies have identified atopological distinction between a “motor” and a “cognitive/affective” cerebellum.Motor-related cortices project primarily to the contralateral anterior cerebellar lobe(lobules I–V) –extending into medial lobule VI, and to lobule VIII via the caudal halfof the pons. Prefrontal, parietal, and temporal association areas project to theposterior cerebellar lobe (Crus I, Crus II, lobule VI/VII/IX) via the mediorostral(prefrontal areas) and dorso- and ventrolateral pons (posterior cerebral hemi-spheres) (Habas et al., 2009; Keren-Happuch, Chen, Ho, & Desmond, 2014; Mariënet al., 2014; Stoodley & Schmahmann, 2010). The affective processes are believed tobe primarily situated in the cerebellar posterior vermis (De Smet et al., 2013;Stoodley & Schmahmann, 2009, 2010) (Figure 2). This topographical division ofthe human cerebellum is also supported by separate olivo-cerebellar connections.While cutaneo-kinesthetic information is provided to the cerebellum via spinocer-ebellar and trigeminocerebellar tracts (with the medial and dorsal accessory olivarynuclei projecting on the sensorimotor cerebellum (lobules I–V, VI, VIII)), the princi-pal olivary nucleus (with little or no spinal cord input) is connected with lobule VIIand the dentate nucleus (DN), and to a smaller degree with lobule VI (Stoodley &Schmahmann, 2010). A similar topographic division between motor and nonmotorareas can also be found in the dentate nuclei, the thalamus, and the pons, theareas involved in the reciprocal cerebello-cerebral loops (see Figure 1). Since mostof the output and input of the cerebellum is regulated by the DN, a divisionbetween a motor and a nonmotor area is also found in the DN (Habas, van Dun,& Mariën, 2015). Projections to and from the motor areas pass through the dorsalpart of the DN, while the projections from the associative cortices pass through the
APHASIOLOGY 1379

Figure 1. (A) Diagram depicting the cerebello-cerebral connectivity network underlying cognitiveand affective processes. The feedback or efferent loop originates from the deep nuclei of thecerebellum that project to the motor (grey arrows) and nonmotor (blue arrows) nuclei of thethalamus. In turn, the motor nuclei of the thalamus project to motor and premotor cortices (greyarrows) but also to nonmotor association cortices (blue arrows). The nonmotor nuclei of the thalamusproject only to association cortices (blue arrows). After Schmahmann and Pandya (1997). Adaptedfrom Mariën et al. (2013). (B) Topographic distribution of motor-related cortices and associationcortex feedforward or afferent projections to the cerebellum. Both motor corticopontine projectionsand association cortex projections are somatotopically organised in the pons. See also Stoodley andSchmahmann (2010). Adapted from Grimaldi and Manto (2012).
Figure 2. Simplified representation of the dichotomy of the cerebellum in a motor (anterior) and acognitive/affective (posterior/vermis) cerebellum. Adapted from Manto and Mariën (2015).
1380 K. VAN DUN ET AL.

ventral part of the DN (Schlerf, Wiestler, Verstynen, & Diedrichsen, 2014; Stricket al., 2009).
From an evolutionary point of view, the larger size of the DN in humans than inprimates is considered the result of expansion of the ventrolateral “nonmotor” part ofthe human DN (Matano, 2001). Enlargement of the DN might reflect the increasedimportance of nonmotor functions in humans (Strick et al., 2009), especially since itsdevelopment during evolution coincided with the expansion of the prefrontal cortex(Middleton & Strick, 1994, 2001). Leiner, Leiner, and Dow (1986) suggested that theparallel expansion of the ventral DN and the frontal cortical areas, which are intrinsicallyimplicated in cognitive functioning, implies involvement of the cerebellum in cognitiveand linguistic processes as well (Leiner et al., 1986). This hypothesis is firmly endorsed bycurrent insights in the neuroanatomical projections from the ventral DN to the prefron-tal cortex.
Functional imaging (see Table 1 for a summary of the cited studies)
Positron emission tomography (PET)In 1988 and 1989, Petersen et al. identified right cerebellar activation in language taskexperiments with positron emission tomography (PET), a technique to measure brainactivity. In their study, the authors used three conditions: first, single words werepresented both aurally and visually to 17 right-handed healthy subjects without anyaction required (sensory task), then they asked these subjects to repeat the presentedword (output task), and finally they had to perform a noun–verb association task(association task) in which an auditory presented noun had to be silently associatedwith a semantically related verb. Activation of the right lateral inferior cerebellum wasfound, specifically in the noun–verb association task. Referring to the anatomical paperof Leiner et al. (1986), they speculated that this activation rather had a “cognitive” rolethan a motor or a sensory one (Petersen, Fox, Posner, Mintun, & Raichle, 1989). Since thisstudy, a large number of experimental and clinical neuroimaging studies have confirmedconsistent involvement of the right posterior cerebellum in silent word generation tasks(e.g., Grabowski et al., 1996; Martin, Haxby, Lalonde, Wiggs, & Ungerleider, 1995;Papathanassiou et al., 2000).
Functional magnetic resonance imaging (FMRI)An activation-likelihood estimate (ALE) meta-analysis performed by Stoodley andSchmahmann (2009) included 11 functional magnetic resonance imaging (fMRI) studies(performed between 1998 and 2006) investigating language-specific activations, con-trolled for motor activation. Several different language tasks were used and contrastedwith various motor tasks to isolate specific language functions such as phonological andsemantic processing, and word generation. For all language tasks, the strongest peaks inthe analysis were lateralised in lobule VI, Crus I/II, and midline lobule VII of the rightcerebellum. Only a small lateral cluster was found in left-hemisphere lobule VI (Stoodley& Schmahmann, 2009). A more recent meta-analysis performed by Keren-Happuch et al.(2014) confirmed the activation of the right lateral posterior cerebellum during languagetasks. This study specifically targeted cerebellar activation clusters. Four studies wereincluded in addition to the meta-analysis performed by Stoodley and Schmahmann
APHASIOLOGY 1381

Table1.
Overview
ofcitedimagingstud
ies.
Functio
nalimaging
Type
ofimaging
Stud
yParticipants
Tasks
Cerebellaractivations
PET
Petersen
etal.(1989)
17R(11F,6
M)
Sensorytask
(fixatio
npo
int)
Outpu
ttask
(passive
words)
Associationtask
(repeatwords)
Non
eSuperio
rAn
terio
rCB
(L+R)
Inferio
rLateralC
B(R)/PosteriorCB
(L+R)
Martin
etal.(1995)
12R(3F,9M
)Co
lour
wordgeneratio
n(reading
alou
d)Verb
generatio
n(reading
alou
d)Non
eLateralC
B(R)
Grabowskie
tal.(1996)
18R
Outpu
ttask
(passive
words)
Associationtask
(repeatwords)
Inferio
rLateralC
B(R
+L)
Papathanassio
uet
al.(2000)
8R(M
)Passivestorylistening
(rest)
Covertverb
generatio
n(rest)
PosteriorCB
(R>L)
Anterio
rCB
(R+L)/Posterio
rCB
(R)
fMRI
Stoodley
andSchm
ahmann
(2009)
Meta-analysisof
11articles
(ALE-m
etho
d)Severald
ifferentlang
uage
tasks
Lobu
leVI—Crus
I(R)/CrusI-II—
lobu
leVIIAt(R)/Lobu
leVI
(L)
Keren-Happuch
etal.(2014)
Meta-analysisof
15stud
ies
(ALE-m
etho
d)Expressive
lang
uage
tasks
Receptivelang
uage
tasks
Lobu
leVI—Midlobu
leVIII—
Crus
I(R)/CrusI(R+L)/Crus
II(R
+L)
Crus
I(R)/Lob
uleVI
(L)
Hubrich-Ungureanu
etal.
(2002)
1R(F),1L
(F)
Covertph
onolog
ical
verbalfluency(rest)
R:CB
(R)
L:CB
(L)
SPECT
Stud
yPatients
Deficit(s)
Lesion
site
Hypop
erfusion
Crossedcerebellardiaschisis
Abeet
al.(1997)
15(6F,9M
)Broca’sAp
hasia
Spared
base
offron
talg
yrus
Base
offron
talg
yrus
Non
eCB
R<L
Mariën,
Pickut,
Engelborghs,Martin,&
DeDeyn(2001)
83yRM
AoS
Phon
olog
icalagraph
iaLanterio
rinsula/adjacent
fron
talo
percular
cortex
CBR<L
Crossedcerebello-cerebral
diaschisis
Mariënet
al.(1996)
73yRM
Luria’sdynamicaphasia
RSCA
Parieto-occipitalregion(R
+L)/
Fron
to-parietalregion(L)
Zettin
etal.(1997)
46yRM
Expressive
agrammatism
RCB
haem
atom
aFron
to-tem
poralcortex(L)/Basal
gang
lia(L)/Thalam
us(L)
Mariënet
al.(2009)
58yRM
Mild
transcorticalsensoryaphasia/Visual
dyslexia/surface
dysgraph
iaRSCAischem
icinfarctio
nMedialfrontallobe
(L)
Baillieux
etal.(2010)
18R(6F,12M)
Attention(72%
)Executive(50%
)Mem
ory(50%
)Behaviou
ral/affective(50%
)
RCB
(vascular:3PICA
,3SCA;
tumou
r:2)
LCB
(vascular:4PICA
,1SCA;
tumou
r:1)
BilateralC
B(vascular:1SCA)
Verm
is(3)
Fron
tal(8/13)
Parietal(2/13)
DeSm
etet
al.(2011)
74yRM
Ataxicdysarthria/dynam
icaphasia/
anom
ia/apraxicagraph
iaRCB
haem
orrhage
Posteriorparietal(L)
Motor/premotor
areas(L)
86yRF
Ataxicdysarthria/apraxicagraph
ia/m
ildattentionprob
lems
R>LCB
ischem
iclesion
sOld
Rtempo
ro-parietal
ischem
icinfarctio
n
Parietallob
e(R)
Motor/premotor
areas(R)
76yRM
Apraxicagraph
ia/frontalprob
lem
solving/mentalfl
exibility
RPICA
infarctio
nOld
Rpo
steriorparietal
ischem
icinfarctio
n
Lateralp
refron
tallob
e(L
+R)
Inferio
rlateralfrontallobe
(L+R)
Inferiormedialfrontallobe
(L+R)
PATIEN
TS:R
:right-handed;
L:left-handed;
F:female;
M:m
ale;
y:years;OTH
ERCO
LUMNS:
R:rig
ht;L:left;CB
:cerebellum;S
CA:sup
eriorcerebellarartery;P
ICA:
posteriorinferio
rcerebellar
artery;SCA
6:SpinocerebellarAtaxia6;
ALE:activation-likelihoodestim
ation.
1382 K. VAN DUN ET AL.

(2009), expanding the range from 2006 to 2010 (Keren-Happuch et al., 2014). Thus,involvement of the right posterior lateral cerebellar hemisphere in language via crossedanatomical connections with the language-dominant left frontal, parietal, and temporalassociation areas was confirmed by functional neuroimaging.
The fMRI study performed by Hubrich-Ungureanu, Kaemmerer, Henn, and Braus(2002) also demonstrated crossed cerebello-cerebral activations subserved by crossedanatomical tracts between the supratentorial association cortices and the cerebellum. Aright-handed subject with typical left hemisphere language dominance and a left-handed subject with atypical right hemisphere language dominance were asked tosilently perform a verbal fluency task in the scanner. In addition to activation of theleft fronto-parietal cortex involvement, the right cerebellar hemisphere was found in theright-handed subject, while in the left-handed subject, the left cerebellar hemispherewas co-activated with the right fronto-parieto-temporal cortex (Hubrich-Ungureanuet al., 2002). This study not only confirmed involvement of the cerebellum in languagetasks, but also showed a clear lateralisation of cerebellar involvement contralateral to thelanguage-dominant left or right cerebral hemisphere.
Single photon emission computerised tomography (SPECT)Crossed (cerebro-)cerebellar diaschisis. Evidence for a pattern of crossed activationshas also been provided by SPECT studies which measure cerebral blood perfusion. Baron(1980) described the phenomenon of crossed cerebellar diaschisis, denoting the distantfunctional impact of a supratentorial lesion on the contralateral cerebellar hemisphere.Crossed cerebellar diaschisis associated with language deficits was investigated by Abe,Ukita, Yorifuji, and Yanagihara (1997). They performed a group study with MRI andSPECT in 30 patients with chronic Broca’s aphasia after cerebral damage. It was shownthat infarcts affecting the lower part of the left frontal gyrus were associated withcrossed cerebellar diaschisis, implying functional and/or anatomical connections of thissupratentorial region with the right cerebellar hemisphere. Interestingly, patients withcrossed cerebellar diaschisis presented with classical Broca’s aphasia, while patientswithout damage to the lower left frontal gyrus mainly presented with word-findingdifficulties without additional more complex language impairments (Abe et al., 1997). Inaddition, several case studies using SPECT imaging in patients with Apraxia of Speech(AoS) after left insular infarctions, provided support for a close interconnection betweenthe language-dominant left frontal regions, and the right cerebellum (e.g., Mariën,Pickut, et al., 2001). After an isolated infarction in the precentral gyrus of the lan-guage-dominant insula, directly anterior to the central insular sulcus, an 83-year-oldright-handed patient presented with severe AoS and phonological agraphia. A Tc-99m-ECD SPECT scan nine days after admission revealed a hypoperfusion in the left inferiorfrontal gyrus and precentral gyrus, and in the right cerebellar hemisphere (Mariën,Pickut, et al., 2001).
Crossed cerebello-cerebral diaschisis. The reversed phenomenon in which (focal)cerebellar damage exerts a distant functional effect at the supratentorial level is calledcrossed cerebello-cerebral diaschisis. Decreased cerebral blood flow in the cerebralhemisphere contralateral to the lesioned cerebellar hemisphere is an indirect measureof function often found in the anatomoclinically suspected supratentorial regions
APHASIOLOGY 1383

subserving linguistic and cognitive functions (Baillieux et al., 2010; De Smet,Engelborghs, Paquier, De Deyn, & Mariën, 2011; Mariën et al., 1996, 2007; Zettin et al.,1997).
Clinical evidence (see Table 2 for a summary of the cited studies)
Clinically, several different types of motor and nonmotor language deficits have beenreported after cerebellar damage, ranging from pure motor speech disorders to high-level metalinguistic disturbances.
Motor speech productionAtaxic dysarthria is a typical cerebellar motor speech disorder, mainly characterisedby distorted articulation and prosody (Spencer & Slocomb, 2007). Traditionally, ataxicdysarthria is considered a disorder of motor execution. However, recent studies havemodified this view to also include disruption of motor speech programming inaddition to a dysfunctional execution of speech (Mariën & Verhoeven, 2007;Spencer & Slocomb, 2007). This was based on some typical characteristics of ataxicdysarthria which are more compatible with a programming/planning disorder thanwith an execution one (Spencer & Slocomb, 2007). Moreover, ataxic dysarthria seemsto share some overt semiological characteristics with AoS, a typical planning andprogramming disorder, which has led to the hypothesis that both disorders aresubserved by related pathophysiological mechanisms (Mariën et al., 2006). Mariënet al. (2006) and Mariën and Verhoeven (2007) observed a significant correlationbetween the perfusional deficits in the right cerebellum and the remission of AoSsymptoms in two patients with a subtype of AoS, suggesting an involvement of thecerebellum in the planning and programming of speech.
Verbal fluency and language dynamicsDisruption of verbal fluency and language dynamics has repeatedly been observed inpatients with focal and degenerative cerebellar lesions (Leggio, Silveri, Petrosini, &Molinari, 2000; Peterburs, Bellebaum, Koch, Schwarz, & Daum, 2010; Schweizer,Alexander, Susan Gillingham, Cusimano, & Stuss, 2010), with phonological fluencymore prominently affected than semantic fluency. Although Hubrich-Ungureanu et al.(2002) confirmed lateralisation of cerebellar involvement in a (silent) phonologicalfluency task and Schweizer et al. (2010) observed a clear lateralisation effect, anumber of other studies contradict unilateral involvement of the cerebellum in verbalfluency tasks (Cook, Murdoch, Cahill, & Whelan, 2004; Leggio et al., 2000). However,despite some conflicting results with respect to laterality, a consensus exists withregard to the role for the cerebellum in verbal fluency and lexical retrieval (Mariënet al., 2014).
Mariën et al. (1996) described a patient who, after an ischemic infarction in thevascular territory of the right superior cerebellar artery (SCA), presented with aphasicsymptoms corresponding to Luria’s dynamic aphasia (characterised by severelyreduced, adynamic, fragmented self-generated speech despite normal confrontationalnaming and phonological skills). A review by Mariën, Engelborghs, Fabbro, and DeDeyn (2001) disclosed several additional cases in which language dynamics were
1384 K. VAN DUN ET AL.

Table2.
Overview
ofcitedcase
repo
rts.
Clinicalevidence
Lang
uage
deficit
Stud
yPatient
Deficits
Lesion
site
Imaging
Motor
speech
prod
uctio
nMariënand
Verhoeven(2007)
53yRF
FAS
Fron
to-parietal
stroke
SPECT(33days
poststroke):
hypo
perfusionin
CB(R)
61yRM
Cond
uctio
n-likeaphasia/FA
SLbasalg
anglia
haem
orrhageinto
Lparietotempo
ral
region
SPECT(1
mon
thpo
ststroke):
hypo
perfusionin
CB(R)
Spencerand
Slocom
b(2007)
20stud
ies(health
yparticipants)19
stud
ies
(dysarthric
patients)
Non
eAtaxicdysarthria
Non
eCerebellar
lesion
sfM
RI/PET/ERP
fMRI/PET
•Bilateralsup
eriorCB
:speechmotor
control
•RCB
:plann
ingandprocessing
ofspeech
Verbalfluency
Legg
ioet
al.(2000)
25patients(cerebellar
disease)
14healthycontrols
Phon
emicfluency:CB
patients<
controls
Semantic
fluency:CB
patients~
controls
Focalo
rdegenerativeCB
disease(L:n
=13;R
:n=6;
idiopathic:
n=6)
Schw
eizeret
al.
(2010)
22patients(chron
iccerebellar
lesion
s)30
healthycontrols
Phon
emicfluency:RCB
patients<L
CBpatients~controls
Semantic
fluency:RCB
patients<
controls
LCB
:n=12
RCB
:n=10
Cook
etal.(2004)*
5patients(vascularcerebellar
lesion
s)Ph
onem
icfluency/sentence
constructio
n/worddefinitio
ns/
multip
ledefinitio
ns/figu
rative
lang
uage/w
ordassociations/sem
antic
absurdities/synon
yms/
antonyms
LCB
(ischem
icor
haem
orrhagic)
Mariënet
al.
(1996)*
73yRM
Luria’sdynamicaphasia
RSCA
SPECT(30days
poststroke):
hypo
perfusionin
Parieto-occipitalregion
(R+L)
Fron
to-parietalregion(L)
Riva
(1998)
4y2m
FMutism–>
redu
cedspon
taneou
sspeech/aph
onic/aprosod
ic/sequential
mem
ory
Viralcerebellitis
Gasparin
ietal.
(1999)*
51yRM
Agrammatism/dysprosod
yRCB
infarctio
nSPECT:hypo
perfusionin
RCB (C
ontin
ued)
APHASIOLOGY 1385

Table2.
(Con
tinued).
Clinicalevidence
Lang
uage
deficit
Stud
yPatient
Deficits
Lesion
site
Imaging
Fabb
roet
al.
(2000)*
21yRM
Syntax/synon
yms/attributegeneratio
nArachn
oidalcystin
PF28yRF
Syno
nym/arithm
etics/writingto
dictation
Hem
angiob
lastom
ain
PF48yRM
Dysarthria/fluency/
prop
osition
ing/
lexicalaccess/morph
olog
y/syntax/
semantics/behaviou
ral/w
ritingto
dictation
CBastrocytom
ain
verm
is
59yF
Grammar/arithm
etics/
reading/syntax/
writingto
dictation
LCB
astrocytom
a
Riva
andGiorgi
(2000)
26patients
Mutism
=>dysarthriaORAff
ective/
social
behaviou
rald
isturbances
RCB
astrocytom
a(n
=7)
LCB
astrocytom
a(n
=8)
CBverm
ismedulloblastoma
(n=11)
Grammatical/Syntactical
processing
Silverie
tal.(1994)
67yRM
Expressive
agrammatism
RCB
infarct
SPECT(5
weeks
poststroke):
hypo
perfusionin:Cerebralh
emisph
ere(L)
Zettin
etal.(1997)
46yRM
Expressive
agrammatism
RCB
haem
atom
aSPECT(4
weeks
poststroke):
hypo
perfusionin
Fron
to-tem
poralcortex
(L)Basalg
anglia
(L)Thalam
us(L)
Ackerm
annet
al.
(1999)
9(3F,6M
)Ataxia/receptiveagrammatism
Degenerative
cerebellaratroph
y
Mariënet
al.
(1996)*
73yRM
Luria’sdynamicaphasia
RSCA
SPECT(30days
poststroke):
hypo
perfusionin
Parieto-occipitalregion
(R+L)
Fron
to-parietalregion(L)
Gasparin
ietal.
(1999)*
51yRM
Agrammatism/dysprosod
yRCB
infarctio
nSPECT:hypo
perfusionin
RCB (C
ontin
ued)
1386 K. VAN DUN ET AL.

Table2.
(Con
tinued).
Clinicalevidence
Lang
uage
deficit
Stud
yPatient
Deficits
Lesion
site
Imaging
Justus
(2004)
16patients16
controls
Grammaticalmorph
olog
yRCB
(n=3)
LCB
(n=3)
MidlineCB
(n=1)
BilateralC
B(n
=9)
Adam
aszeket
al.
(2012)
8Rpatients(1F,7M
)8R
controls(2F,6M
)Syntactic
processing
RCB
(n=2)
LCB
(n=6)
Reading
Morettiet
al.
(2002)
10Rpatients(4F,6M
)10Rcontrols(5F,5M
)Reading
Verm
ian/
paraverm
iantumou
r
Mariënet
al.
(2009)*
58yRM
Mild
transcorticalsensoryaphasia/
Visualdyslexia/surface
dysgraph
iaRSCAischem
icinfarctio
nSPECT(5
weeks
poststroke):
hypo
perfusionin
CB(R)Medialfrontal
lobe
(L)
Writing
Silverie
tal.(1997)
67yRM
Spatiald
ysgraphia
Cerebellaratroph
ySilveri,Misciagna,
Legg
io,and
Molinari(1999)
26yRF
Spatiald
ysgraphia
CBhaem
orrhage
(vermis+R>L
hemisph
ere)
Mariënet
al.
(2007)
72yRM
Apraxicagraph
iaRCB
haem
orrhage
SPECT(1
and6mon
thspo
ststroke):
hypo
perfusionin
CB(R)Medial/lateral
prefrontalarea
(L)
DeSm
etet
al.
(2011)
74yRM
Ataxicdysarthria/dynam
icaphasia/
anom
ia/apraxicagraph
iaRCB
haem
orrhage
SPECT(1
and6mon
thspo
ststroke):
hypo
perfusionin
Posteriorparietal(L)
Motor/premotor
areas(L)
86yRF
Ataxicdysarthria/apraxicagraph
ia/
mild
attentionprob
lems
R>LCB
ischem
iclesion
sOld
Rtempo
ro-parietal
ischem
icinfarctio
n
SPECT(1
and6mon
thspo
ststroke):
hypo
perfusionin
Parietallob
e(R)Motor/
prem
otor
areas(R)
76yRM
Apraxicagraph
ia/frontalprob
lem
solving/mentalfl
exibility
RPICA
infarctio
nOld
Rpo
steriorparietal
ischem
icinfarctio
n
SPECT(2
weeks
poststroke):
hypo
perfusionin
Lateralp
refron
tallob
e(L
+R)
Inferio
rlateralfrontallobe
(L+R)
Inferio
rmedialfrontallobe
(L+R)
Mariënet
al.
(2009)*
58yRM
Mild
transcorticalsensoryaphasia/
Visualdyslexia/surface
dysgraph
iaRSCAischem
icinfarctio
nSPECT(5
weeks
poststroke):
hypo
perfusionin
CB(R)Medialfrontal
lobe
(L)
(Con
tinued)
APHASIOLOGY 1387
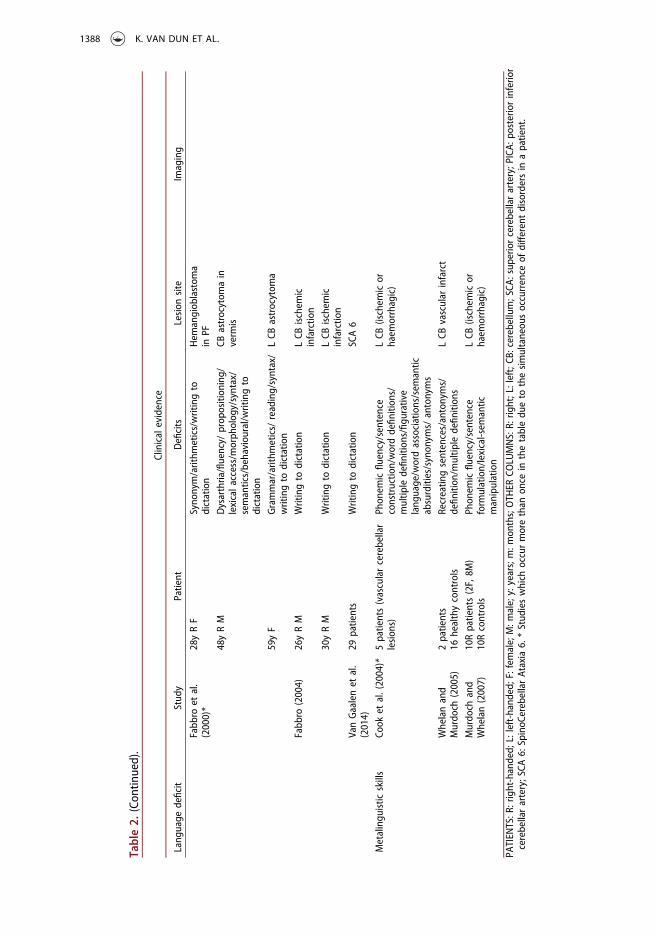
Table2.
(Con
tinued).
Clinicalevidence
Lang
uage
deficit
Stud
yPatient
Deficits
Lesion
site
Imaging
Fabb
roet
al.
(2000)*
28yRF
Syno
nym/arithm
etics/writingto
dictation
Hem
angiob
lastom
ain
PF
48yRM
Dysarthria/fluency/
prop
osition
ing/
lexicalaccess/morph
olog
y/syntax/
semantics/behaviou
ral/w
ritingto
dictation
CBastrocytom
ain
verm
is
59yF
Grammar/arithm
etics/
reading/syntax/
writingto
dictation
LCB
astrocytom
a
Fabb
ro(2004)
26yRM
Writingto
dictation
LCB
ischem
icinfarctio
n
30yRM
Writingto
dictation
LCB
ischem
icinfarctio
n
VanGaalenet
al.
(2014)
29patients
Writingto
dictation
SCA6
Metalingu
istic
skills
Cook
etal.(2004)*
5patients(vascularcerebellar
lesion
s)Ph
onem
icfluency/sentence
constructio
n/worddefinitio
ns/
multip
ledefinitio
ns/figu
rative
lang
uage/w
ordassociations/sem
antic
absurdities/synon
yms/
antonyms
LCB
(ischem
icor
haem
orrhagic)
Whelanand
Murdo
ch(2005)
2patients
16healthycontrols
Recreatin
gsentences/antonyms/
definitio
n/multip
ledefinitio
nsLCB
vascular
infarct
Murdo
chand
Whelan(2007)
10Rpatients(2F,8M
)10Rcontrols
Phon
emicfluency/sentence
form
ulation/lexical-sem
antic
manipulation
LCB
(ischem
icor
haem
orrhagic)
PATIEN
TS:R
:right-handed;
L:left-handed;
F:female;M:m
ale;y:years;m:m
onths;OTH
ERCO
LUMNS:R:
right;L:left;CB
:cerebellum;SCA
:sup
eriorcerebellarartery;P
ICA:
posteriorinferio
rcerebellarartery;SCA
6:SpinoC
erebellarAtaxia6.
*Stud
ieswhich
occurmorethan
once
inthetabledu
eto
thesimultaneou
soccurrence
ofdifferentdisordersin
apatient.
1388 K. VAN DUN ET AL.

consistently disturbed after (right) cerebellar damage, resembling transcortical motoraphasia (Fabbro, Moretti, & Bava, 2000; Gasparini et al., 1999; Riva, 1998). Disruptionof language dynamics also characterises the Posterior Fossa Syndrome (PFS), a clinicalcondition consisting of transient cerebellar mutism, cognitive symptoms, and beha-vioural abnormalities that typically occurs in children after posterior fossa tumourresection (Riva & Giorgi, 2000).
Grammatical/syntactical processingGrammatical and syntactic disorders are also associated with cerebellar damage. Bothexpressive (Silveri, Leggio, & Molinari, 1994; Zettin et al., 1997) and receptive agramma-tism (Ackermann, Gräber, Hertrich, & Daum, 1999) have been observed. In most cases,the grammatical disorders are caused by right cerebellar lesions (Gasparini et al., 1999;Mariën, Engelborghs, Pickut, & De Deyn, 2000; Mariën et al., 1996; Silveri et al., 1994;Zettin et al., 1997). However, there are reports of patients with left cerebellar lesions whoalso presented with grammatical and/or syntactic deficits (Adamaszek, Strecker, &Kessler, 2012; Justus, 2004).
ReadingOnly a handful of cases are described in which acquired cerebellar damage leads toreading difficulties (Mariën et al., 2009; Moretti, Bava, Torre, Antonello, & Cazzato, 2002).Moretti et al. (2002) demonstrated that patients with cerebellar lesions made moreerrors at letter- and word level in reading words and sentences, compared to right-handed controls. More recently, Mariën et al. (2009) reported a patient with visualdyslexia after an ischemic infarction in the vascular territory of the right SCA. A quanti-fied SPECT study showed a relative hypoperfusion in the right cerebellar hemisphereand the left medial frontal lobe, which may be caused by a functional disruption of thecerebello-cerebral pathways (Mariën et al., 2009).
WritingIn the last few decades, cerebellar lesions have been frequently associated with writingdisorders. Both peripheral agraphia, a disorder of the coordination, planning, andexecution of writing movements, not attributable to motor or sensory impairments(e.g., spatial dysgraphia, apraxic agraphia) and central agraphia, involving the centralspelling processes, have been reported. Silveri, Misciagna, Leggio, and Molinari (1997,1999) described two patients with spatial dysgraphia after cerebellar damage, andMariën et al. (2007) and De Smet et al. (2011) reported several patients with acquiredapraxic agraphia associated with focal cerebellar damage. Quantified SPECT studies inthe patients with apraxic agraphia consistently showed reduced perfusion in the medialprefrontal regions of the left hemisphere, known to be involved in the planning andexecution of skilled graphomotor actions, and the right cerebellum.
Central agraphia on the other hand is only scarcely reported after cerebellardamage. Mariën et al. (2009) described a patient with surface dysgraphia (togetherwith visual dyslexia) after an ischemic infarction in the vascular territory of the rightSCA. A quantified SPECT study showed a hypoperfusion in the left medial frontal lobe.Other possible cases of central agraphia are difficult to evaluate due to limited
APHASIOLOGY 1389

information about the observed writing disorder (Fabbro, 2004; Fabbro et al., 2000;Van Gaalen et al., 2014).
Metalinguistic skillsSeveral case studies point to an involvement of the left cerebellar hemisphere in higherlevel language and metalinguistic skills. Some patients with primary left cerebellar strokeare known to have difficulties with providing multiple definitions for homophonicwords, recreating sentences, figurative and ambiguous language, word associations,antonym/synonym generation, and interpreting semantic absurdities (Cook et al.,2004; Murdoch & Whelan, 2007; Whelan & Murdoch, 2005). These symptoms are typicallyassociated with right cerebral damage (Kempler, Van Lancker, Marchman, & Bates, 1999).
Possible pathophysiological mechanisms and cognitive theories forcerebellar involvement in language
Several different hypotheses have been put forward to explain the pathophysiologicaland cognitive mechanisms by means of which the cerebellum has a role in linguisticprocesses.
Pathophysiological mechanisms
In 1987, Broich, Hartmann, Biersack, and Horn for the first time described crossedcerebello-cerebral diaschisis after a cerebellar infarction. Their SPECT study showed arelative hypoperfusion in the contralateral left hemisphere in their patient with a rightcerebellar infarction. They hypothesised that crossed perfusional deficits reflect a func-tional depression of the supratentorial areas due to a lack of excitatory stimuli throughthe cerebello-ponto-thalamo-cerebral pathways which connect the cerebellum to sev-eral cerebral association areas crucially implicated in cognitive and affective processing(Broich, Hartmann, Biersack, & Horn, 1987).
Since then, crossed cerebello-cerebral diaschisis has been frequently observed inpatients with speech and language deficits after cerebellar damage (Baillieux et al.,2010; De Smet et al., 2011; Hassid, 1995; Mariën et al., 2000, 1996, 2007; Zettin et al.,1997). For example, Mariën et al. (1996) described a 73-year-old right-handed man withan ischemic infarction in the right superior cerebellum. The patient presented withaphasic symptoms closely resembling Luria’s dynamic aphasia, complicated by receptiveand expressive agrammatism. SPECT findings revealed relative hypoperfusions in theclinically suspected prefrontal regions of the language-dominant hemisphere. Moreover,the pattern of perfusional deficits mirrored linguistic recovery: the prefrontal languageregions regained normal perfusion when remission of the aphasic symptoms wasobserved. This evidence strongly suggests a link between crossed cerebello-cerebraldiaschisis and cerebellar-induced language disturbances (Mariën et al., 2000, 1996).
Cognitive theories
In 1998, Schmahmann and Sherman introduced the concept of cerebellar cognitiveaffective syndrome (CCAS) or Schmahmann’s syndrome (Manto & Mariën, 2015). After
1390 K. VAN DUN ET AL.

screening and testing of 20 patients with isolated cerebellar lesions, they identified acluster of symptoms that could be classified in four categories: (1) executive dysfunc-tions (planning, set-shifting, abstract reasoning, and working memory), (2) visuospatialdeficits (visuospatial organisation and memory), (3) behavioural-affective disturbances(blunting of affect or disinhibited and inappropriate behaviour) and (4) language symp-toms (agrammatism and anomia), (Schmahmann & Sherman, 1998). To explain thecerebellar induced deficits, Schmahmann (1998) introduced the concept of “dysmetriaof thought”, which draws analogies with the motor system. This hypothesis suggests arole for the cerebellum as a modulator of cognitive and affective function that comparesthe intended outcome with the perceived outcome to detect and prevent or correctmismatches. Schmahmann (1998) hypothesised that the cerebellum acts as an oscilla-tion dampener, smoothing out performance of mental operations (Mariën et al., 2014;Schmahmann, 1998).
Due to its architectural homogeneity and its dense interconnections with thesupratentorial association areas, it is frequently stated that the cerebellum contri-butes to both motor and nonmotor activities in a similar manner. Schmahmannreferred to this unique computation as the “universal cerebellar transform” (UCT)(Koziol et al., 2014; Schmahmann, 2000). This phenomenon allows the cerebellum tocompare predictions of “internal models” (= “any neural representation of theexternal world” (Ito, 2008)) with the incoming feedback from the cerebrum, optimis-ing human behaviour and performance. This explanation is in accordance with theconcept of “dysmetria of thought” that Schmahmann introduced in 1998(Schmahmann, 1998) and which implies a “universal cerebellar impairment” (UCI).If the cerebellum is involved in every cortical process in the same way, cerebellardamage should result in a similar kind of disruption of these processes (Koziol et al.,2014). In this context, Schmahmann reported in a number of studies that everydeficit resulting from cerebellar damage can be interpreted as hypometric/dimin-ished or hypermetric/exaggerated behaviours (for an overview, see Koziol et al.,2014). As a result, the theories relating to motor dysfunction and cerebellar diseasewere adapted to account for the observed linguistic deficits after cerebellar damage(Mariën et al., 2014; Strick et al., 2009).
Internal models have been used to explain the ability to perform controlled andskilful movements by creating a prediction of the dynamics of a body part. Thesepredictions help to perform a complex movement in a very precise way withoutconstantly relying on sensory feedback (Ito, 2008). Ito (2008) hypothesised that theseinternal models can be used in the same manner to create and manipulate mentalrepresentations, in association with the prefrontal and the temporo-parietal cortex.These models have also been used to explain the involvement of the cerebellum inlanguage (Mariën et al., 2014). In language studies, the cerebellum is frequentlyregarded as a predictor of future states (as in the “dysmetria of thought” hypothesis(Schmahmann, 1998)) that uses these predictions to optimise cognitive and languagefunctioning by comparing them with the cortical feedback (Mariën et al., 2014).
However, which information the cerebellum uses to form these predictions remains amatter of debate (Leggio & Molinari, 2015). Below, two main hypotheses of the motorliterature are discussed in the context of cognitive and language functions. These two
APHASIOLOGY 1391

theories are frequently cited in studies that found involvement of the cerebellum inlanguage functioning (Mariën et al., 2014).
Timing hypothesisKeele and Ivry (1990) suggested that the cerebellum is responsible for the temporalcomputation of a number of different tasks. This hypothesis was primarily based onobservations of timing deficits in cerebellar patients. Patients with cerebellar disor-ders had trouble judging the velocity of moving visual stimuli and differentiatingbetween two auditory stimuli of different lengths. As a result, Keele and Ivry (1990)proposed a role for the cerebellum in providing temporal information in motorcoordination. According to these authors, the cerebellum times different movementsbut also incorporates cortical input to adjust this timing accordingly, such as inlocomotion. The cerebellum is therefore responsible for the timing of the expectedfeedback and the actual feedback. If an expected feedback comes earlier or laterthan expected, it will be treated as an unexpected event due to the timing coordina-tion of the cerebellum (Gellman, Gibson, & Houk, 1985; Keele & Ivry, 1990). Silveriet al. (1994) and Gasparini et al. (1999) used the timing hypothesis to explain thelinguistic deficits observed after right cerebellar damage in their patients. Theyspeculated that the agrammatism they observed in patients with cerebellar lesionsmight be the result of a delay in the application of syntactic rules and linguisticprocesses (Gasparini et al., 1999). Due to this delay, caused by the damage in thecerebellum, the information held in the verbal working memory decays before all theoperations are performed, resulting in errors (Gasparini et al., 1999; Silveri et al.,1994). As such, the cerebellum times the linguistic operations performed on theinformation held in the verbal working memory.
Sequencing hypothesisAnother explanation of cerebellar involvement in language is provided by thesequencing hypothesis. The cerebellum as a monitor of sequential events has alreadybeen proven in the somatosensory system using expected and unexpected stimuli(Leggio, Chiricozzi, Clausi, Tedesco, & Molinari, 2011). With respect to language, it ishypothesised that the activity of different functional modules, necessary to performlinguistic processes, is synchronised and coordinated by the cerebellum. The cerebel-lum might act as a monitor and supervisor of the sequence in which linguisticprocesses are performed, integrating its activity with the so-called “frontal-lobesystem” (Fabbro et al., 2000). Especially in novel strategies (e.g., used in phonologicalfluency), the cerebellum would play an important role in smoothing and speeding upthe process (Leggio et al., 2011; Mariën et al., 2014).
It is also believed that the cerebellum is responsible for the temporal and sequentialorganisation of both overt and covert speech (Ackermann, 2008). Ackermann, Mathiak,and Ivry (2004) suggested that cognitive deficits observed after cerebellar damage couldbe due to an impairment of the “inner speech”, which can compromise the verbalworking memory (Koziol et al., 2014). The performance on cognitive operations invol-ving a generation of a prearticulatory speech code most likely suffers if this “innerspeech” function is compromised (Ackermann et al., 2004).
1392 K. VAN DUN ET AL.

A lateralised linguistic cerebellum
All suggested cognitive theories strongly rely on the crossed functional cerebello-cere-bral connections, via which the cerebellum exerts its modulatory influence on thecognitive functions subserved by the supratentorial areas. Each of these theories regardthe cerebellum as a coordinator/modulator of supratentorial motor, cognitive, andaffective processes, recognising the cerebellum as a non-specific high-level operationaldevice that provides active support to a variety of functions in a similar manner (Mariënet al., 2014). Since in the vast majority of right-handers (±97%) and in most left-handers(±70%) the left hemisphere is dominant for language (Knecht et al., 2000; Mariën,Paghera, De Deyn, & Vignolo, 2004), it is assumed that the involvement of the cerebel-lum in language functions is strongly lateralised to the right hemisphere, while the leftcerebellar hemisphere primarily contributes to right hemisphere functioning (Mariën,Engelborghs, et al., 2001; Stoodley & Schmahmann, 2009). Several studies have shownthat the configuration of language lateralisation at the supratentorial level is stronglyrelated to a contralateral lateralised language representation at the cerebellar level.Jansen et al. (2005) showed that atypical right-hemisphere language dominance isaccompanied by a left-lateralised cerebellar contribution to language. Additionally,Mendez Orellana et al. (2014) found a significant dependency between language later-alisation in the cerebrum and the cerebellum with an inverse correlation of cerebro-cerebellar lateralisation. This means that the stronger the language lateralisation in thecerebral hemisphere, the stronger the contralateral cerebellar hemisphere is involved inlanguage. Clinically, this is confirmed by a study of Baillieux et al. (2010). They system-atically investigated 18 patients with primary cerebellar lesions by means of an extensiveneuropsychological battery. Although disturbances of attention, executive function, andmemory were most commonly found, their analyses supported the hypothesis of afunctionally lateralised linguistic cerebellum with the right cerebellum being associatedwith logical reasoning and language processing, and the left cerebellum with attentionaland visuo-spatial skills (Baillieux et al., 2010).
Conclusion
Converging evidence from an increasing number of neuroanatomical, neuroimaging, andclinical studies unambiguously shows that the cerebellum plays a cardinal role in a varietyof linguistic processes via a dense network of cerebello-cerebral connections. Thesecrossed connections have led to the concept of a “lateralised linguistic cerebellum” inwhich the right cerebellar hemisphere is involved in left-hemispheric, and the left cere-bellar hemisphere in right-hemispheric functioning (Mariën, Engelborghs, et al., 2001).Many SPECT studies in patients with cerebellar disorders have demonstrated the functionalimpact of a cerebellar lesion on a remote, structurally intact language area, suggestingcrossed cerebello-cerebral diaschisis as the possible pathophysiological mechanism under-lying the observed linguistic deficits.
Due to the architectural homogeneity of the cerebellum, a “UCT” was proposedby Schmahmann (2000), resulting in a “UCI”. According to this hypothesis thecerebellum acts as a coordinator/modulator of both motor and nonmotor (cogni-tive/affective) functions by comparing cortical feedback with “internal models”.
APHASIOLOGY 1393

These models are internal representations/predictions of future states (Mariën et al.,2014). Which information is used to make up these “internal models”, is still amatter of debate. The “dysmetria of thought” theory indicates that the cerebellumtries to keep motor and cognitive processes around a homeostatic baseline, actinglike an oscillation dampener, without specifying which aspects of the processes aremonitored (Schmahmann, 1998). Two factors adapted from the motor literaturehave been discussed in this paper that may hypothetically account for the observedlanguage disturbances: timing and sequencing. Both mechanisms have been dealtwith in the literature to explain the observed language deficits (timing: Silveri et al.,1994; Gasparini et al., 1999; | sequencing: Fabbro et al., 2000; Ackermann et al.,2004). However, future studies are needed to unravel the specific involvement ofthe cerebellum in cognitive functioning, as it is not always clear which languagedeficits are due to a timing deficit, which to a disruption of sequencing, and whichare merely due to disturbed attention control. Hypothetically, it might be arguedthat a timing deficit leads to errors in which not all linguistic operations aresuccessfully completed, such as the use of infinitives instead of a conjugated verbin agrammatism. Due to the delay caused by the timing deficit, as hypothesised bySilveri et al. (1994) and Gasparini et al. (1999), the information temporarily stored inthe working memory system decays before all operations are completed (Gaspariniet al., 1999; Silveri et al., 1994). However, a timing deficit as described by Keele andIvry (1990) and Gellman et al. (1985) could lead to other kinds of errors due to adeficit in timing and coordinating the cortical feedback. This hypothesis merelypredicts a disruption of the incorporation of the cortical feedback due to aninaccurate timing of the expected and the actual input. This could lead to a varietyof linguistic errors. The sequencing hypothesis (Leggio & Molinari, 2015), on theother hand, predicts a deficit in the coordination and monitoring of the sequenceof application of different linguistic operations. According to this hypothesis, repe-titive patterns of temporally or spatially structured serial events are identified andsimulated by the cerebellum. Based on this, the cerebellum generates internalmodels that can be used to make predictions. Disruption of serial pattern detectionand prediction, and processing of anticipation might not only lead to the use ofinfinitives instead of a conjugated verb, but also to wrongfully conjugated verbs.
In the literature on the role of the cerebellum in motor processing, both timing andsequencing deficits have been observed after cerebellar damage (timing: Keele & Ivry,1990; | sequencing: Leggio et al., 2011). It is therefore possible that the cerebellum isinvolved in both mechanisms, resulting in complicated linguistic errors due to bothtiming and sequencing deficits.
In conclusion, it can be stated that the cerebellum contributes in a unique way to themodulation and coordination of a variety of motor and nonmotor (cognitive/affective)processing.
Disclosure statement
No potential conflict of interest was reported by the authors.
1394 K. VAN DUN ET AL.

References
Abe, K., Ukita, H., Yorifuji, S., & Yanagihara, T. (1997). Crossed cerebellar diaschisis in chronic Broca’saphasia. Neuroradiology, 39, 624–626. doi:10.1007/s002340050480
Ackermann, H. (2008). Cerebellar contributions to speech production and speech perception:Psycholinguistic and neurobiological perspectives. Trends in Neurosciences, 31, 265–272.doi:10.1016/j.tins.2008.02.011
Ackermann, H., Gräber, S., Hertrich, I., & Daum, I. (1999). Cerebellar contributions to the perceptionof temporal cues within the speech and nonspeech domain. Brain and Language, 67, 228–241.doi:10.1006/brln.1999.2056
Ackermann, H., Mathiak, K., & Ivry, R. B. (2004). Temporal organization of “internal speech” as abasis for cerebellar modulation of cognitive functions. Behavioral and Cognitive NeuroscienceReviews, 3, 14–22. doi:10.1177/1534582304263251
Adamaszek, M., Strecker, K., & Kessler, C. (2012). Impact of cerebellar lesion on syntactic processingevidenced by event-related potentials. Neuroscience Letters, 512, 78–82. doi:10.1016/j.neulet.2012.01.020
Baillieux, H., De Smet, H. J., Dobbeleir, A., Paquier, P. F., De Deyn, P. P., & Mariën, P. (2010).Cognitive and affective disturbances following focal cerebellar damage in adults: A neuropsy-chological and SPECT study. Cortex, 46, 869–879. doi:10.1016/j.cortex.2009.09.002
Baron, J. C. (1980). “Crossed cerebellar diaschisis” in human supratentorial brain infarction.Transactions of the American Neurological Association, 105, 459–461.
Broich, K., Hartmann, A., Biersack, H.-J., & Horn, R. (1987). Crossed cerebello-cerebral diaschisis in apatient with cerebellar infarction. Neuroscience Letters, 83, 7–12. doi:10.1016/0304-3940(87)90207-2
Cook, M., Murdoch, B. E., Cahill, L., & Whelan, B.-M. (2004). Higher-level language deficits resultingfrom left primary cerebellar lesions. Aphasiology, 18, 771–784. doi:10.1080/02687030444000291
De Smet, H. J., Engelborghs, S., Paquier, P. F., De Deyn, P. P., & Mariën, P. (2011). Cerebellar-inducedapraxic agraphia: A review and three new cases. Brain and Cognition, 76, 424–434. doi:10.1016/j.bandc.2010.12.006
De Smet, H. J., Paquier, P., Verhoeven, J., & Mariën, P. (2013). The cerebellum: Its role in languageand related cognitive and affective functions. Brain and Language, 127, 334–342. doi:10.1016/j.bandl.2012.11.001
Fabbro, F. (2004). Long-term neuropsychological deficits after cerebellar infarctions in two youngadult twins. Neuropsychologia, 42, 536–545. doi:10.1016/j.neuropsychologia.2003.09.006
Fabbro, F., Moretti, R., & Bava, A. (2000). Language impairments in patients with cerebellar lesions.Journal of Neurolinguistics, 13, 173–188. doi:10.1016/S0911-6044(00)00010-5
Gasparini, M., Di Piero, V., Ciccarelli, O., Cacioppo, M. M., Pantano, P., & Lenzi, G. L. (1999).Linguistic impairment after right cerebellar stroke: A case report. European Journal ofNeurology, 6, 353–356. doi:10.1046/j.1468-1331.1999.630353.x
Gellman, R., Gibson, A. R., & Houk, J. C. (1985). Inferior olivary neurons in the awake cat: Detectionof contact and passive body displacement. Journal of Neurophysiology, 54, 40–60.
Grabowski, T. J., Frank, R. J., Brown, C. K., Damasio, H., Boles Ponto, L. L., Watkins, G. L., & Hichwa,R. D. (1996). Reliability of PET activation across statistical methods, subject groups, and samplesizes. Human Brain Mapping, 4, 23–46. doi:10.1002/(SICI)1097-0193(1996)4:1<>1.0.CO;2-Y
Grimaldi, G., & Manto, M. (2012). Topography of cerebellar deficits in humans. The Cerebellum, 11(2), 336–351. doi:10.1007/s12311-011-0247-4
Habas, C., Kamdar, N., Nguyen, D., Prater, K., Beckmann, C. F., Menon, V., & Greicius, M. D. (2009).Distinct cerebellar contributions to intrinsic connectivity networks. Journal of Neuroscience, 29,8586–8594. doi:10.1523/JNEUROSCI.1868-09.2009
Habas, C., van Dun, K., & Mariën, P. (2015). Deep cerebellar nuclei and language. In M. Manto & P.Mariën (Eds.), The linguistic cerebellum. London: Elsevier/Academic Press.
Hassid, E. I. (1995). A case of language dysfunction associated with cerebellar infarction.Neurorehabilitation and Neural Repair, 9, 157–160. doi:10.1177/154596839500900304
APHASIOLOGY 1395

Hubrich-Ungureanu, P., Kaemmerer, N., Henn, F. A., & Braus, D. F. (2002). Lateralized organizationof the cerebellum in a silent verbal fluency task: A functional magnetic resonance imaging studyin healthy volunteers. Neuroscience Letters, 319, 91–94. doi:10.1016/S0304-3940(01)02566-6
Ito, M. (2008). Control of mental activities by internal models in the cerebellum. Nature ReviewsNeuroscience, 9, 304–313. doi:10.1038/nrn2332
Jansen, A., Flöel, A., Van Randenborgh, J., Konrad, C., Rotte, M., Förster, A.-F., . . . Knecht, S. (2005).Crossed cerebro-cerebellar language dominance. Human Brain Mapping, 24, 165–172.doi:10.1002/hbm.20077
Justus, T. (2004). The cerebellum and English grammatical morphology: Evidence from production,comprehension, and grammaticality judgments. Journal of Cognitive Neuroscience, 16, 1115–1130. doi:10.1162/0898929041920513
Keele, S. W., & Ivry, R. (1990). Does the cerebellum provide a common computation for diversetasks? A timing hypothesis. Annals of the New York Academy of Sciences, 608, 179–211.doi:10.1111/j.1749-6632.1990.tb48897.x
Kempler, D., Van Lancker, D., Marchman, V., & Bates, E. (1999). Idiom comprehension in childrenand adults with unilateral brain damage. Developmental Neuropsychology, 15, 327–349.doi:10.1080/87565649909540753
Keren-Happuch, E., Chen, S.-H. A., Ho, M.-H. R., & Desmond, J. E. (2014). A meta-analysis ofcerebellar contributions to higher cognition from PET and fMRI studies: A meta-analysis ofcerebellar contributions. Human Brain Mapping, 35, 593–615. doi:10.1002/hbm.22194
Knecht, S., Dräger, B., Deppe, M., Bobe, L., Lohmann, H., Floël, A., . . . Henningsen, H. (2000).Handedness and hemispheric language dominance in healthy humans. Brain, 123, 2512–2518.doi:10.1093/brain/123.12.2512
Koziol, L. F., Budding, D., Andreasen, N., D’Arrigo, S., Bulgheroni, S., Imamizu, H., . . . Yamazaki, T.(2014). Consensus paper: The cerebellum’s role in movement and cognition. The Cerebellum, 13,151–177. doi:10.1007/s12311-013-0511-x
Leggio, M. G., Chiricozzi, F. R., Clausi, S., Tedesco, A. M., & Molinari, M. (2011). The neuropsycho-logical profile of cerebellar damage: The sequencing hypothesis. Cortex, 47, 137–144.doi:10.1016/j.cortex.2009.08.011
Leggio, M. G., & Molinari, M. (2015). Cerebellar sequencing: A trick for predicting the future. TheCerebellum, 14, 35–38. doi:10.1007/s12311-014-0616-x
Leggio, M. G., Silveri, M. C., Petrosini, L., & Molinari, M. (2000). Phonological grouping is specificallyaffected in cerebellar patients: A verbal fluency study. Journal of Neurology, Neurosurgery &Psychiatry, 69, 102–106. doi:10.1136/jnnp.69.1.102
Leiner, H. C., Leiner, A. L., & Dow, R. S. (1986). Does the cerebellum contribute to mental skills?Behavioral Neuroscience, 100, 443–454. doi:10.1037/0735-7044.100.4.443
Manto, M., & Mariën, P. (2015). Schmahmann’s syndrome – identification of the third cornerstoneof clinical ataxiology. Cerebellum & Ataxias, 2. doi:10.1186/s40673-015-0023-1
Mariën, P., Ackermann, H., Adamaszek, M., Barwood, C. H. S., Beaton, A., Desmond, J., . . . Ziegler, W.(2014). Consensus paper: Language and the cerebellum: An ongoing enigma. The Cerebellum,13, 386–410. doi:10.1007/s12311-013-0540-5
Mariën, P., Baillieux, H., De Smet, H. J., Engelborghs, S., Wilssens, I., Paquier, P., & De Deyn, P. P.(2009). Cognitive, linguistic and affective disturbances following a right superior cerebellarartery infarction: A case study. Cortex, 45, 527–536. doi:10.1016/j.cortex.2007.12.010
Mariën, P., De Smet, H. J., Wijgerde, E., Verhoeven, J., Crols, R., & De Deyn, P. P. (2013). Posteriorfossa syndrome in adults: A new case and comprehensive survey of the literature. Cortex, 49(1),284–300. doi:10.1016/j.cortex.2011.06.018
Mariën, P., Engelborghs, S., Fabbro, F., & De Deyn, P. P. (2001). The lateralized linguistic cerebellum:A review and a new hypothesis. Brain and Language, 79, 580–600. doi:10.1006/brln.2001.2569
Mariën, P., Engelborghs, S., Pickut, B. A., & De Deyn, P. P. (2000). Aphasia following cerebellardamage: Fact or fallacy? Journal of Neurolinguistics, 13, 145–171. doi:10.1016/S0911-6044(00)00009-9
Mariën, P., Paghera, B., De Deyn, P. P., & Vignolo, L. A. (2004). Adult crossed aphasia in dextralsrevisited. Cortex, 40, 41–74. doi:10.1016/S0010-9452(08)70920-1
1396 K. VAN DUN ET AL.

Mariën, P., Pickut, B. A., Engelborghs, S., Martin, -J.-J., & De Deyn, P. P. (2001). Phonologicalagraphia following a focal anterior insulo-opercular infarction. Neuropsychologia, 39, 845–855.doi:10.1016/S0028-3932(01)00006-9
Mariën, P., Saerens, J., Nanhoe, R., Moens, E., Nagels, G., Pickut, B. A., . . . De Deyn, P. P. (1996).Cerebellar induced aphasia: Case report of cerebellar induced prefrontal aphasic languagephenomena supported by SPECT findings. Journal of the Neurological Sciences, 144, 34–43.doi:10.1016/S0022-510X(96)00059-7
Mariën, P., & Verhoeven, J. (2007). Cerebellar involvement in motor speech planning: Some furtherevidence from foreign accent syndrome. Folia Phoniatrica Et Logopaedica, 59, 210–217.doi:10.1159/000102933
Mariën, P., Verhoeven, J., Brouns, R., De Witte, L., Dobbeleir, A., & De Deyn, P. P. (2007). Apraxicagraphia following a right cerebellar hemorrhage. Neurology, 69, 926–929. doi:10.1212/01.wnl.0000267845.05041.41
Mariën, P., Verhoeven, J., Engelborghs, S., Rooker, S., Pickut, B. A., & De Deyn, P. P. (2006). A role forthe cerebellum in motor speech planning: Evidence from foreign accent syndrome. ClinicalNeurology and Neurosurgery, 108, 518–522. doi:10.1016/j.clineuro.2005.06.006
Martin, A., Haxby, J. V., Lalonde, F. M., Wiggs, C. L., & Ungerleider, L. G. (1995). Discrete corticalregions associated with knowledge of color and knowledge of action. Science, 270, 102–105.doi:10.1126/science.270.5233.102
Matano, S. (2001). Brief communication: Proportions of the ventral half of the cerebellar dentatenucleus in humans and great apes. American Journal of Physical Anthropology, 114, 163–165.doi:10.1002/(ISSN)1096-8644
Mendez Orellana, C., Visch-Brink, E., Vernooij, M., Kalloe, S., Satoer, D., Vincent, A., . . . Smits, M.(2014). Crossed cerebrocerebellar language lateralization: An additional diagnostic feature forassessing atypical language representation in presurgical functional MR imaging. AmericanJournal of Neuroradiology. doi:10.3174/ajnr.A4147
Middleton, F. A., & Strick, P. L. (1994). Anatomical evidence for cerebellar and basal gangliainvolvement in higher cognitive function. Science, 266, 458–461. doi:10.1126/science.7939688
Middleton, F. A., & Strick, P. L. (2001). Cerebellar projections to the prefrontal cortex of the primate.The Journal of Neuroscience, 21, 700–712.
Moretti, R., Bava, A., Torre, P., Antonello, R. M., & Cazzato, G. (2002). Reading errors in patients withcerebellar vermis lesions. Journal of Neurology, 249, 461–468. doi:10.1007/s004150200040
Murdoch, B. E., & Whelan, B.-M. (2007). Language disorders subsequent to left cerebellar lesions:A case for bilateral cerebellar involvement in language? Folia Phoniatrica Et Logopaedica, 59,184–189. doi:10.1159/000102930
Papathanassiou, D., Etard, O., Mellet, E., Zago, L., Mazoyer, B., & Tzourio-Mazoyer, N. (2000). Acommon language network for comprehension and production: A contribution to the definitionof language epicenters with PET. NeuroImage, 11, 347–357. doi:10.1006/nimg.2000.0546
Peterburs, J., Bellebaum, C., Koch, B., Schwarz, M., & Daum, I. (2010). Working memory and verbalfluency deficits following cerebellar lesions: Relation to interindividual differences in patientvariables. The Cerebellum, 9, 375–383. doi:10.1007/s12311-010-0171-z
Petersen, S. E., Fox, P. T., Posner, M. I., Mintun, M., & Raichle, M. E. (1989). Positron emissiontomographic studies of the processing of singe words. Journal of Cognitive Neuroscience, 1,153–170. doi:10.1162/jocn.1989.1.2.153
Riva, D. (1998). The cerebellar contribution to language and sequential functions: Evidence from achild with cerebellitis. Cortex, 34, 279–287. doi:10.1016/S0010-9452(08)70755-X
Riva, D., & Giorgi, C. (2000). The cerebellum contributes to higher functions during developmentevidence from a series of children surgically treated for posterior fossa tumours. Brain, 123,1051–1061. doi:10.1093/brain/123.5.1051
Schlerf, J., Wiestler, T., Verstynen, T., & Diedrichsen, J. (2014). Big challenges from the little brain –Imaging the cerebellum. In T. D. Papageorgiou, G. I. Christopoulos, & S. M. Smirnakis (Eds.),Advanced brain neuroimaging topics in health and disease – Methods and applications. Rijeka:InTech. Retrieved from http://cdn.intechopen.com/pdfs-wm/46098.pdf
APHASIOLOGY 1397

Schmahmann, J. D. (1998). Dysmetria of thought: Clinical consequences of cerebellar dysfunctionon cognition and affect. Trends in Cognitive Sciences, 2, 362–371. doi:10.1016/S1364-6613(98)01218-2
Schmahmann, J. D. (2000). The role of the cerebellum in affect and psychosis. Journal ofNeurolinguistics, 13, 189–214. doi:10.1016/S0911-6044(00)00011-7
Schmahmann, J. D., & Pandya, D, D. N. (1997). The cerebrocerebellar system. In J. D. Schmahmann(Ed.), The cerebellum and cognition, International review of neurobiology (pp. 31–60). San Diego,CA: Academic Press.
Schmahmann, J. D., & Sherman, J. C. (1998). The cerebellar cognitive-affective syndrome. Brain,121, 561–579. doi:10.1093/brain/121.4.561
Schweizer, T. A., Alexander, M. P., Susan Gillingham, B. A., Cusimano, M., & Stuss, D. T. (2010).Lateralized cerebellar contributions to word generation: A phonemic and semantic fluencystudy. Behavioural Neurology, 23, 31–37. doi:10.1155/2010/102421
Silveri, M. C., Leggio, M. G., & Molinari, M. (1994). The cerebellum contributes to linguisticproduction: A case of agrammatic speech following a right cerebellar lesion. Neurology, 44,2047–2047. doi:10.1212/WNL.44.11.2047
Silveri, M. C., Misciagna, S., Leggio, M. G., & Molinari, M. (1997). Spatial dysgraphia and cerebellarlesion: A case report. Neurology, 48, 1529–1532. doi:10.1212/WNL.48.6.1529
Silveri, M. C., Misciagna, S., Leggio, M. G., & Molinari, M. (1999). Cerebellar spatial dysgraphia:Further evidence. Journal of Neurology, 246, 312–313. doi:10.1007/s004150050353
Spencer, K., & Slocomb, D. (2007). The neural basis of ataxic dysarthria. The Cerebellum, 6, 58–65.doi:10.1080/14734220601145459
Stoodley, C. J., & Schmahmann, J. D. (2009). Functional topography in the human cerebellum: Ameta-analysis of neuroimaging studies. NeuroImage, 44, 489–501. doi:10.1016/j.neuroimage.2008.08.039
Stoodley, C. J., & Schmahmann, J. D. (2010). Evidence for topographic organization in thecerebellum of motor control versus cognitive and affective processing. Cortex, 46, 831–844.doi:10.1016/j.cortex.2009.11.008
Strick, P. L., Dum, R. P., & Fiez, J. A. (2009). Cerebellum and nonmotor function. Annual Review ofNeuroscience, 32, 413–434. doi:10.1146/annurev.neuro.31.060407.125606
Van Gaalen, J., De Swart, B. J. M., Oostveen, J., Knuijt, S., Van De Warrenburg, B. P. C., & Kremer, B.H. P. (2014). Language impairment in cerebellar ataxia: LANGUAGE IMPAIRMENT IN SCA6.Movement Disorders, 29, 1307–1312. doi:10.1002/mds.25854
Whelan, B.-M., & Murdoch, B. E. (2005). Unravelling subcortical linguistic substrates: Comparisonof thalamic versus cerebellar cognitive-linguistic regulation mechanisms. Aphasiology, 19,1097–1106. doi:10.1080/02687030500174050
Zettin, M., Cappa, S. F., D’amico, A., Rago, R., Perino, C., Perani, D., & Fazio, F. (1997). Agrammaticspeech production after a right cerebellar haemorrhage. Neurocase, 3, 375–380. doi:10.1080/13554799708411976
1398 K. VAN DUN ET AL.