the importance of primary producers for … importance of primary producers for benthic nitrogen and...
TRANSCRIPT

KAREN J. MCGLATHERY, KRISTINA SUNDBACK, IRIS C. ANDERSON
THE IMPORTANCE OF PRIMARY PRODUCERS FOR BENTHIC NITROGEN AND PHOSPHORUS CYCLING
I. INTRODUCTION
In shallow coastal systems where most of the seafloor lies within the photic zone, benthic photoautotrophy plays a key role in regulating nutrient cycling. In these systems, production of submerged vascular plants (seagrasses), macroalgae and benthic microalgae is high and typically exceeds that of phytoplankton (Borum and Sand Jensen 1996). Changes in the patterns of primary production as well as in habitat structure and trophic dynamics (Valiela et al. 1992; Nixon et al. 1996) have been directly related to nutrient over-enrichment; this is also recognized as one of the greatest threats to maintaining marine biodiversity in coastal regions (NRC 2000). The widely-accepted scenario following nutrient enrichment is a shift in the dominance of primary producers, from seagrasses and perennial macroalgae to fast- growing green macroalgae and phytoplankton (Sand-Jensen and Borum 199 1 ; Duarte 1995; Valiela et al. 1997; see Chapter 3). Interestingly, only one of these models (Sand-Jensen and Borum 199 1) has included explicitly the potential importance of benthic microalgae. Ultimately, one might expect a shift to phytoplankton dominance in the most heavily eutrophied shallow estuaries (Duarte 1995; Valiela et al. 1997), even though total production might not change (Borum and Sand-Jensen 1996). Given this transition, the question then becomes, how might such a shift in primary producer dominance influence nutrient cycling in shallow coastal waters? The influence of primary producers on nutrient transformations has important consequences for the role of shallow estuaries as a buffer zone between land and sea (Fig. 1). Uptake and temporary retention of nutrients in plant biomass, burial of recalcitrant organic matter, and the direct effects of autotrophic metaboiism on bacterially- and chemically-mediated processes all influence nutrient cycling rates and pathways. Variations in the rates and dominance of these processes as primary producer communities change, will ultimately determine the fate and retention of watershed nutrients on their trajectory to the open ocean.
S. Nielsen, G. Bantu und M. Pedersen (ed.~.), Estuarine Nutrient Cyclit~g: The Influence of Primary Procilrcers, 23 1-261. O 2004 Kl~rwer Academic Puhlisher.~. Printed in the Netherlands.

232 K.J. McGlathery, K. Sundback and I.C. Anderson
Fig. I . Role of primary producers on the fate and retention of nzitrients at the land-sea interface.
In this chapter we address the influence of primary producer groups (seagrasses, benthic microalgae, macroalgae) on nutrient cycling processes. We consider the effects of plant metabolism on biogeochemical transformations within the sediments and nutrient exchanges between the sediments and water column (see Chapter 10 fol discussion of denitrification). Our focus is on the retention and loss processes thti influence both the responses to nutrient enrichment and the transport of nutrients through these shallow systems. We consider these processes in ecosystems that var) with respect to tidal amplitude, water residence times, seasonality and sedimen type. Nutrient assimilation and turnover by benthic primary producers appear tc play a dominant role in regulating the magnitude and timing of nutrient fluxes ir these systems. This being the case, we pay particular attention to the competitive interactions between primary producer communities and to the fate of nutrients bound in plant tissue.
2. BENTHIC NUTRIENT CYCLING PROCESSES
The role of benthic primary producer communities as sources or sinks of nutrients reflects the net influences of assimilatory nutrient uptake, nitrogen fixation, leakage by live and grazed autotrophs, oxygen release, nutrient production and losses due mineralization, and nitrification - denitrification (Fig. 2). Nutrients can be transferred between the sediment and water column as dissolved inorganic or organic forms or as gases. Exchange rates are also influenced by benthic faunal communities (e.g., Pelegri et a!. 1994; Hansen and Kristensen 1997) and by physical factors such as sediment resuspension and porewater advection. Often there are die1

Benthic N and P cycling 233
patterns driven by the primary producers, with an efflux of nutrients during the dark and uptake in the light (e.g., Eyre and Ferguson 2002). The balance between nutrient losses and gains in benthic communities also is influenced by nutrient transfer via mobile foraging animals (see Chapter 6), and export or import of particulate organic matter (see Chapter 4). Nutrient cycling in benthic primary producer communities appears to be relatively conservative, with much of the nutrient demand met by recycling within the sediments. For example, Ottosen et al. (unpublished data) found by field 15~-labelling of sandy sediment that 90% of labeled " PON disappeared from the top 3 cm of sediment after 12 weeks. This rather long retention could be explained if remineralized N was efficiently recycled within the microbial mat (Lomstein et al. 1998), and could be responsible for sustaining high rates of productivity by benthic microalgae during summer in low- nitrogen sandy sediments (Sundback et al. 2000). A similar closed recycling of both N and P also appears to apply to benthic microalgal communities in oligotrophic carbonate sediments of coral reefs (Miyajima et al. 2001; Suzumura et al. 2002) and for both temperate and tropical seagrass systems (Alcoverro et al. 2000; McGlathery et al. 2001b; Miyajima et al. 2001; Risgaard-Petersen and Ottosen 2001). Conservative recycling of sediment N to support autotrophic communities is also the common pattern observed in Spartina alternzjlora vegetated intertidal sediments (Anderson et al. 1997). Although the nutrient demands of seagrasses appear to be met in large part by internal recycling of organic-bound nutrients in the sediment (Hemminga et al. 1991; Alcoverro et al. 2000; McGlathery et al. 2001b), seagrasses and other primary producers in the community also obtain external nutrients that are transported by water flow. Resorption of nutrients from senescing leaves reduces the demand for either recycled or external nutrients in seagrass (e.g., Pedersen and Borum 1993; Stapel and Hemminga 1997; Alcoverro et al. 2000) and in salt marsh systems (Anderson et al. 1997).
2.1 Seagrasses
Seagrasses have a major impact on sediment nutrient cycling by leakage of oxygen and dissolved organic matter (DOM) from the roots. Both processes are photosynthetically-driven. Oxygen produced by photosynthesis in the leaves is transported to the roots to support aerobic respiration via a well-developed lacuna1 system, a series of air channels comprising up to 60% of the total plant volume. The oxygen that is not respired by the roots is released into the rhizosphere, creating oxidized zones in an otherwise reducing environment. At the same time, seagrasses release DOM from the roots, typically as simple organic carbon compounds that are the recent products of photosynthesis (Koepfler et al. 1993; Ziegler and Benner 1999). Bacterial productivity in the rhizosphere has been linked to these organic exudates. Moriarty et al. (1986) showed that 11% of recent photosynthate was released within 6 hours into the rhizosphere of the tropical seagrass Halodule wrightii. Seasonal and light-induced variation in bacterial activity and the typical sub-surface peak in bacterial processes in the sediments where root biomass is highest, also clearly indicate that root exudates stimulate bacterial activity (e.g., Welsh et al. 1996; McGlathery et al. 1998; Blaabjerg et al. 1998).
i
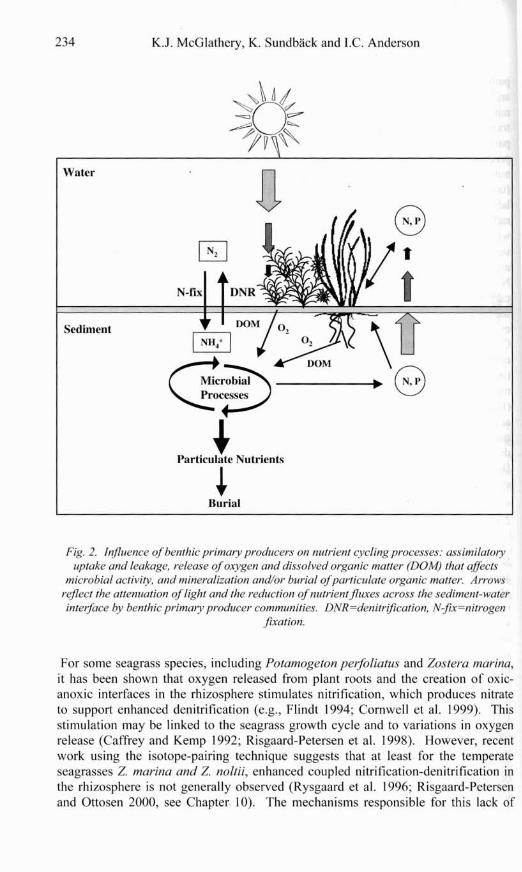
K.J. McGlathery, K. Sundback and I.C. Anderson
Particul;~te Nutrients
4 Burial
Fig. 2. Influence of benthic primary producers on nutrient cycling processes: assimilatory uptake and leakage, release of oxygen and dissolved organic matter (DOM) that affects
microbial activity, and mineralization and/or burial of particulate organic matter. Arrows reflect the attenuation of light and the reduction of nutrient fluxes across the sediment-water interface by benthic primary producer communities. DNR=denitrification, N-fix=nitrogen
furation.
For some seagrass species, including Potamogeton perfoliatus and Zostera marina, it has been shown that oxygen released from plant roots and the creation of oxic- anoxic interfaces in the rhizosphere stimulates nitrification, which produces nitrate to support enhanced denitrification (e.g., Flindt 1994; Comwell et al. 1999). This stimulation may be linked to the seagrass growth cycle and to variations in oxygen release (Caffrey and Kemp 1992; Risgaard-Petersen et al. 1998). However, recent work using the isotope-pairing technique suggests that at least for the temperate seagrasses Z. marina and Z. noltii, enhanced coupled nitrification-denitrification in the rhizosphere is not generally observed (Rysgaard et al. 1996; Risgaard-Petersen and Ottosen 2000, see Chapter. 10). The mechanisms responsible for this lack of

Benthic N and P cycling 235
stimulation are thought to be low rates of rhizosphere nitrification and competition for ammonium between nitrifying bacteria and benthic microalgae (Ottosen et al. 1999; Risgaard-Petersen and Ottosen 2000). This does not preclude the possibility that enhanced denitrification may occur in sediments vegetated by other seagrass species, in particular in tropical species such as Thalassia testudinum, which allocate more biomass to roots and rhizomes than temperate species, and which tend to be strongly P-limited in carbonate sediments so that competition between bacteria and plants for N is unlikely to occur. Organic matter concentrations and bacterial oxygen demand tend to be lower in tropical carbonates sediments, which may make oxygenation of the rhizosphere more efficient in these systems. There are relatively few measurements of denitrification in seagrass-vegetated carbonate sediments in tropical/subtropical environments, but rates tend to be high (Blackburn et al. 1994; Miyajima et al. 2001; Kemp and Cornwell 2001). The available data suggest that both denitrification rates and nitrogen fixation rates are higher in tropical/subtropical seagrass meadows than in temperate seagrass meadows (Table 1). Nitrogen fixation activity is enhanced in seagrass meadows relative to unvegetated sediments, both in temperate and tropical environments (e.g., O'Donohue et al. 1991; Welsh et al. 1996; McGlathery et al. 1998; Hansen et al. 2000). The availability of organic substrate is a major factor controlling N fixation rates in seagrass sediments. Many studies have shown increases of N fixation by addition of organic compounds, and seasonal and die1 variations that are consistent with the role of photosynthetic exudates stimulating N-fixing bacteria (e.g., McGlathery et al. 1998, Hansen et al. 2000; Welsh 2000). Much of this activity (25 - 95%) has been associated with sulfate reducers (e.g., Welsh et al. 1996; McGlathery et al. 1998), which also are believed to be stimulated by root exudates (Blaabjerg et al. 1998; Hansen et al. 2000). Depth profiles of N fixation activity in 2. marina sediments showed a close association with rootlrhizome biomass, with seasonal shifts in the magnitude and depth distribution of N fixation matching the shifts in belowground biomass (McGlathery et al. 1998). Overall, N fixation rates in sediments of seagrass meadows are higher in tropical/subtropical systems than in temperate systems (see references in McGlathery et al. 1998 and Welsh 2000), and this may in part be due to increased DOM release from the greater rootlrhizome biomass of many tropical seagrass species. Autotrophic, heterocystous cyanobacterial epiphytes on seagrass leaves also contribute to the total N fixation rates, especially in warm tropical environments, although on an area basis the activity is lower than that of heterotrophic bacteria in the sediments (e.g., Capone and Taylor 1977; O'Donohue et al. 1991). Few studies have measured N fixation and denitrification simultaneously in seagrass beds to determine if N gains by fixation can compensate for N losses via denitrification, and the results are equivocal (Table 1). In a tropical seagrass meadow, Blackburn et al. (1994) observed that N fixation rates accounted for less than half of the N loss by denitrification, whereas in a temperate seagrass meadow, Risgaard-Petersen et al. (1998) found that N fixation more than compensated for the N loss by denitrification. Both oxygen and DOM release by seagrass roots influence the forms and availability of sediment P. Oxidation in the rhizosphere results in the formation of iron oxides that effectively bind P in the solid phase. The redox conditions in the sediment and the formation of autogenic P minerals influence the release of P to overlying waters (Rozan et al. 2002). In carbonate sediments that typically dominate tropical seagrass

236 K.J. McGlathery, K. Sundback and I.C. Anderson
systems, P turnover rates are generally high in the porewaters relative to N, indicating that P is preferentially removed (Hines and Lyons 1982; McGlathery et al. 2001b). The mechanisms responsible for this include surface adsorption onto carbonate grains, direct precipitation of Ca-P mineral phases, and uptake by P- limited primary producers. In vegetated carbonate sediments in Bermuda, Jensen et al. (1998) found that only 2% of the total sediment P was in the loosely-adsorbed pool that could be readily released to the porewater; 15 to 20% was more strongly adsorbed to the surface of the sediment, and the remainder (80%) was bound in the mineral matrix of the sediments, probably in the form of carbonate fluorapatite and other carbonates. It was previously thought that carbonate sediments were a permanent sink for P in these tropical environments and that mineral P was not available to plants. However, recent studies suggest that seagrass metabolism facilitates the dissolution of carbonate minerals in the rhizosphere releasing bound P (Jensen et al. 1998; Burdige and Zimmerman 2003). One mechanism for this is the decrease in pH resulting from root respiration or the stimulation of bacterial respiration by root DOM release (Burdige and Zimmerman 2003). The oxidation of sulfides in the rhizosphere may also decrease pH, as has been shown for saltmarsh grasses (Giblin and Howarth 1984). Another mechanism is the release of organic acids from the roots that also acidify the rhizosphere and cause the dissolution of the carbonate minerals, as is the case for terrestrial plants (Hoffland 1992; Knight et al. 1992). Iron associated with carbonate minerals can serve as a sorption site for inorganic P (Jensen et al. 1998), and also can react with sediment sulfides to precipitate iron- sulfide minerals that reduce sulfide toxicity to primary producers (Carlson et al. 1994; Erskine and Koch 2000). Chambers et al. (2001) found that iron additions increased the total sediment P pool and reduced exposure of the seagrass T. testudinum to free sulfide, but found no growth response probably because most of the sediment P was bound in unavailable forms. One might expect a more significant effect of iron additions on seagrasses in carbonate sediments where P is not as strongly limiting (i.e., where P is in more readily exchangeable forms). On an ecosystem scale, seagrass systems can be important temporary sinks for nutrients, although there is less information than for benthic micro- and macro-algal communities. The source-sink role will vary seasonally, depending on the growth requirements of seagrasses. Temperate Z. marina meadows may act as strong sinks for N from the beginning of the growing season in the spring to the time of maximum productivity in late summer; in the fall when decomposition rates are typically relatively high, but plant N demand low, the seagrass community may become a source of nutrients (Risgaard-Petersen et al. 1998; Risgaard-Petersen and Ottosen 2000). Denitrification was an important sink relative to plant uptake in these seagrass meadows only in winter when both plant demand and decomposition were low (Risgaard-Petersen and Otteson 2000). Since N fixation may only support a small percentage of seagrass N demand in these systems (Welsh 2000 and references therein), most of the nutrient influx is likely uptake of water column nutrients by seagrass leaves and associated primary producers (benthic microalgae, epiphytes, macroalgae) and accumulation of imported organic matter sedimented into the meadow.

Benthic N and P cycling 237
Table I . Comparison of nitrogen fucation and denitrification rates in temperate and tropical/suhtropical seagrass meadows.
Seagrass Location N Fixation Denitrification Reference u
species mmol N m-2 &' mmol N m-2 d-'
Temperate Zostera marina
Zostera marina
Zostera marina
Zostera noltii
Zostera noltii
Zostera capricorni
Vaucluse Shores, Virginia Limfjord, Denmark Limfjord, Denmark
Arcachon Bay, France Arcachon Bay and Etang du Prevost, France Edmunds Bay, Australia
0.36 Capone (1988)
0.07 - 0.43 McGlathery et al. (1998)
0.4 Risgaard- Petersen et al. ( 1998)
0.10-0.41 0.05 - 0.14 Welsh et al. (2000)
0.33 Rysgaard et al. (1995)
0.19 Eyre & Ferguson (2002)
Tropicall Subtropical Thalassia Oyster Bay, 1 2 - 4 Blackburn et testudinum Jamaica al. (1994) Thalassia Florida Bay, 0.7 - 4 1.6 - 3 Kemp & testudinum Florida Cornwell
(2001) Thalassia Gulf of 1.1 Moriarty & hemprechii Carpentaria, 0' Donohue
Australia (1993) Syringodium Gulf of 1.1 -3.4 Moriarty & isoetifolium Carpentaria, 0' Donohue
Australia ( 1993) Enhalus Gulf of 1.8 Moriarty & acoroides Carpentaria, 0' Donohue
Australia (1 993) Zostera Moreton Bay, 0.7 - 2.9 O'Donohue et capricorni Australia al. (1991)

238 K.J. McGlathery, K. Sundback and I.C. Anderson
Seagrasses enhance nutrient concentrations in the sediments, both in particulate and dissolved forms, by an increased input of organic matter from plant biomass and imported material. Roots and rhizomes typically decay in situ, and some aboveground material may remain in the system rather than be exported by water flow. However, as with salt marsh ecosystems, it appears that much, if not most, of the organic matter buried and decomposed in seagrass sediments is imported (Pedersen et al. 1997; Boschker et al. 2000; Gacia et al. 2002; see Chapter 8). Fine, organic-rich particles settle out as the water flow is slowed by the seagrass canopy, and the roots and rhizomes also help to stabilize the sediments and prevent resuspension. As a result, organic matter and nutrient concentrations are typically higher in vegetated than in unvegetated sediments. Nutrient accumulation may occur in seagrass meadows over the time scale of years, but this source-sink role may in part depend on the trophic status of the system. Pedersen et al. (1997) found that colonization and stand development of the seagrass Cymodocea nodosa. in Mediterranean carbonate sediments resulted in a net accumulation of N, but not P, over a 5 year period, with about half of the accumulated N accounted for by living and dead plant material. In C. nodosa meadows where production was nutrient limited, as is the case during stand development, Perez et al. (1997) observed a net accumulation of N and P in slowly-degrading belowground detritus, yet in eutrophic waters where belowground production was reduced and decay rates of leaves were high, the seagrass meadow becomes a source of nutrients.
2.2 Benthic microalgae
The importance of microphytobenthic production in shallow coastal areas is well documented (Underwood and Kromkamp 1999), as are the effects of microalgal production on N cycling. Photosynthesis and respiration by benthic microalgae in the top few mm of the sediments cause large die1 variations in oxygen concentrations and penetration depth, dissolved inorganic carbon (DIC) concentrations and pH (see Chapter 10). In addition, assimilation of porewater ammonium and nitrate influences the concentrations and depth distributions of porewater N. The net effect of these changes may be die1 variations in nitrification and denitrification activity (Risgaard-Petersen et al 1994, Lorenzen et al. 1998, An and Joye 2001, see Chapter lo), with higher rates of coupled nitrification- denitrification in light due to stimulated nitrification activity from photosynthetic oxygen release. However, this is not always the case (Risgaard-Petersen 2003 and references therein), and light stimulation may only occur when there is sufficient N to prevent N limitation (Rysgaard et al. 1995; An and Joye 2001). Recent field data from I8 European estuaries showed that net autotrophic sediments colonized by microalgae had lower rates of coupled nitrification-denitrification than net heterotrophic sediments (Risgaard-Petersen 2003). This was also the case for shallow bays in Sweden, where denitrification rates were higher (20% of remineralized N) in net heterotrophic than in net autotrophic sediments (10% of remineralized N) (Sundback and Miles 2002). Benthic microalgal communities in both tropical and temperate sediments are generally dominated by diatoms and cyanobacteria. Nitrogen fixation rates in

Benthic N and P cycling 239
surface sediments, and on seagrass leaves, tend to be high relative to those in the water column, and to be stimulated by light (e.g., Capone and Taylor 1977; Capone et al. 1992). Significant autotrophic N fixation occurs even in seagrass-vegetated sediments where the leaf canopy shades the sediment surface, although area- integrated rates generally are lower than heterotrophic N fixation (e.g., O'Neil and Capone 1989). Rates are generally higher in tropical carbonate sands than in temperate siliclastic sediments (e.g., Howarth et al. 1988; Capone et al. 1992). There is currently little evidence of diurnal variations in N fixation activity in benthic microalgal communities in submersed sediments, however, Currin et al. (1996) found a night-time peak in N fixation in filamentous cyanobacterial mats and a day-time peak in coccoid cyanobacterial mats in intertidal sediments. Among the primary producers of shallow-water areas, benthic microalgae appear to be the component whose quantitative role in nutrient cycling is least understood at the ecosystem level. Integrating the effect of benthic microalgae at the ecosystem scale is difficult. Although benthic microalgae are present and active throughout the year (Rizzo et al. 1996 and references therein), their relative quantitative importance for nutrient turnover depends on the presence, seasonal pattern and area coverage of other primary producers, as well as physical characteristics of the embayment. For example, McGlathery et al. (2001a) found that the contribution of benthic microalgae to carbon production varied from 4-99% during the year in a shallow lagoon. The influence of benthic microalgae on net autotrophy and N cycling in the sediments appears to be particularly important in cool microtidal areas (Sundback et al. 2000), where the growth of seagrasses and ephemeral floating macroalgae is confined to the warm season (May-September). In warmer systems, macroscopic primary producers may occur during most of the year, implying more continuous shading and nutrient competition, although this has been observed primarily in eutrophic systems (e.g., Sfiiso et al. 1992; Viaroli et al. 1996). Studies performed in different types of shallow-water ecosystems, ranging from cool temperate (e.g. Rizzo et al. 1992, 1996; Cerco and Seitzinger 1997, Sundback and Miles 2000; Thornton et al. 2002) to warm temperate (Eyre and Ferguson 2002) to subtropical and tropical areas (Miyajima et al. 2001; Suzumura et al. 2002), suggest that benthic microalgae may be an important temporary sink for nutrients. Studies from temperate, relatively N poor areas, suggest that benthic microalgae may turn sediments into sinks of N for the majority of the year, but particularly in winter and spring (e.g., Sundback et al. 2000). In such sandy areas, N may be limiting for benthic microalgae (Nilsson et al. 1991). Also in P-limited oligotrophic subtropical carbonate sediments, microalgal activity has been found to enhance accumulation of combined N (Miyajima et al. 2001).
2.3 Macroalgae
Bloom-forming macroalgae have become dominant in nutrient-enriched waters over at least the last two decades throughout temperate and tropical regions (e.g., Cambridge & McComb 1984; Sfriso et al. 1992; Viaroli et al. 1996; Valiela et al. 1997; Pihl et al. 1999). These "nuisance" algae are typically filamentous or sheet- like forms -- mainly chlorophytes (e.g., Ulva, Cladophora, Chaetomorpha) -- that can accumulate in extensive thick mats over the sediment surface or in the water

240 K.J. McGlathery, K. Sundback and I.C. Anderson
column. Variations in light availability within the mats, and in nutrient, oxygen, and pH conditions linked to algal photosynthesis, respiration, and decomposition, influence nutrient cycling at the sediment-water interface, and perhaps even deep into the sediments. The presence of dense macroalgal mats can move the location of the oxic-anoxic interface up from the sediments into the macroalgal mat since only the upper few cm of the mat may be within the photic zone (Krause-Jensen et al. 1999; Astill and Lavery 2001). Sediment nutrient cycling is enhanced by the presence of macroalgae, presumably due to the input of organic matter (Trimmer et al. 2000; Tyler et al. 2003). Increased mineralization rates cause a build up of ammonium deep within the mat where light does not penetrate (McGlathery et al. 1997; Astill and Lavery 2001), and may affect porewater ammonium as deep as 13 cm in the sediment (Burton et al, unpublished data). This recycling of organic-bound nitrogen may provide an important nitrogen source to sustain macroalgal production, particularly in areas where tidal exchange is limited (McGlathery et al. 1997; Trimmer et al. 2000). Increased ammonium concentrations at depth in sediments may also be partly related to local extinction of bioturbating infauna caused by anoxia (Hansen and Kristensen 1997). Where bioturbating infauna is present, increased oxygen diffusion into the sediments may stimulate nitrification rates (Rysgaard et al. 1995; Hansen and Kristensen 1997). Compared to benthic microalgae and seagrasses, there is little information on the influence of macroalgae on denitrification rates. In studies performed in harbors on the south coast of England, Trimmer et al. (2000) found that both direct denitrification, using nitrate supplied from the water column and denitrification couple to sedimentary nitrification were low in sediments underlying macroalgal mats. High free sulfide concentrations occur in organic-rich sediments underlying macroalgal accumulations (Viaroli et al. 1996), and these conditions may inhibit nitrification (Henriksen and Kemp 1998; Sloth et al. 1995) and partially account for the low denitrification rates. In a laboratory experiment, Krause-Jensen et al. (1999) showed that the net effect of dense macroalgal mats was to move the zone of denitrification from the sediments up into the mat, but not to influence the rates significantly. Denitrification rates peaked at the oxic-anoxic interface in the middle of the mat. Presumably the same regulating mechanisms would apply over the larger spatial scale in a macroalgal mat as have been determined in benthic microalgal communities in the sediment. Macroalgal photosynthesis and respiration induce diurnal variations in oxygen penetration depth within the upper productive layers of the mat, which in turn, may cause diurnal variations in nitrification activity and in coupled nitrification-denitrification. Mineralization of algal-bound nutrients in the lower layers of the mat where there is insufficient light for photosynthesis creates a diffusion gradient of ammonium up into the oxidized layers. Like benthic microalgae, it is expected that macroalgae will compete with bacteria for ammonium and nitrate. It appears that macroalgae do not stimulate denitrification, however, more work is needed to address the controls on denitrification in sediments underlying benthic macroalgal communities and within the mats themselves. At the ecosystem scale, macroalgae can store significant quantities of nutrients. In highly enriched waters, it is not unusual for macroalgal populations to attain peak biomass of over 0.5 kg dry wt m-2 and for canopy heights to exceed 0.5 m. In Waquoit Bay, Massachusetts, nitrogen stored in peak macroalgal biomass was of the

Benthic N and P cycling 24 1
same magnitude as the annual nitrogen load from the watershed (Valiela et al. 1997). Many studies have shown sediments overlain with macroalgae to be a net sink for inorganic nutrients (e.g., Krause-Jensen et al. 1996; Tyler et al. 2001). In Hog Island Bay, Virginia, macroalgae were a net sink for DIN throughout the year, and at the same time a net source for organic N (Tyler et al. 2003); urea and dissolved free amino acids (DFAA) were sequestered by macroalgae, but up to 22% of the total nitrogen uptake was released as dissolved combined amino acids (DCAA). On a seasonal time scale, it is common for macroalgae in eutrophic waters to switch from being a net sink early in the growing season, to a net source of nutrients in late summer when productivity declines due to self-shading within the mat and high temperatures increase respiration. High rates of macroalgal metabolism can lead to periods of oxygen supersaturation or, following senescence, oxygen depletion throughout the entire water column (e.g., Sfriso et al. 1992, Valiela et al. 1992, Boynton et al. 1996). Anoxic events in macroalgal-dominated systems tend to be episodic and to occur during the warm summer months when macroalgal production declines. Such "boom and bust' cycles of high production and senescence are typical of dense macroalgal populations in eutrophic waters (e.g., McComb and Humphries 1992, Sfriso et al. 1992, Valiela et al. 1997). In Hog Island Bay, Virginia, Tyler et al. (2001) reported dissolved inorganic and organic N release rates following the collapse of a macroalgal bloom (primarily composed of Gracilaria tikvahiae and Ulva lactuca) that were sufficient to result in complete mineralization of the macroalgal biomass (up to 650 gdw m-2) within approximately 13 days. Release of plant-bound nutrients following these "dystrophic" events may stimulate phytoplankton and bacterial metabolism in the water column (Valiela et al. 1997; McGlathery et al. 2001a; Lunsford 2002).
3. SEDIMENT - WATER COLUMN FLUXES
Nutrient assimilation by benthic primary producers can reduce the efflux of remineralized nutrients from the sediments to the overlying water, effectively decoupling nutrient turnover within the sediments from water column processes. During times of the year when primary producer populations are most active, this "filter" effect completely intercepts nutrient fluxes across the sediment-water interface and thereby reduces nutrient availability for phytoplankton and bacteria, as well as for floating macroalgal mats, in the overlying water. If the sediment nutrient sources are insufficient to meet growth demands, there also may be a downward flux of nutrients from the water column to the benthic community. It is generally assumed that in littoral zone ecosystems biogeochemical processes occurring in the benthos are closely coupled with those in the pelagic zone. For example, primary production in the water column supports heterotrophic metabolism in underlying sediments. In turn, organic matter remineralization in sediments releases nutrients that support primary production in overlying water. In situations where benthic - pelagic coupling is strong it should be possible to calculate ammonium fluxes out of sediments based on measured rates of net ecosystem metabolism (NEM; dissolved oxygen uptake or DIC release) (Hopkinson et al.

242 K.J. McGlathery, K. Sundback and I.C. Anderson
2001). In oxic sediments where nitrification is likely to play an important role in transformation of ammonium, it may be the total dissolved inorganic nitrogen (DIN) flux that is more closely related to the stoichiometry of NEM, whereas in anoxic sediments where denitrification is important the estimated DIN flux will not equal that predicted by NEM stoichiometry. Calculation of denitrification has often been based upon this "missing" DIN. Using ' 5 ~ isotope dilution techniques, it is possible to measure both gross N mineralization and consumption of ammonium in sediments amended with 1 5 ~ ~ 4 ' , thereby allowing determination of net mineralization rates (Anderson et al, 2003). Net mineralization may also be measured during incubation of unamended sedimentlwater cores. Where gross and net mineralization and sediment-water column exchanges of DIN have been measured simultaneously in autotrophic sediments, gross rates of ammonium production generally exceed net rates by a large margin and net production of ammonium in the sediments is often not reflected in fluxes to the water column (e.g., Kristensen et al. 2000, Anderson et al. 2003). Benthic microalgal assimilation is likely to be largely responsible for this (Anderson et al. 2003), although sediment bacteria may also play a role in N immobilization depending on the C/N of the sediment organic matter undergoing decomposition. In sediments with large amounts of macrophyte detritus with C/N as high as 45, and with the short turnover times typically observed for the ammonium pool ( < I d; Kristensen et al. 2000; Anderson et al. 2003), bacterial immobilization of N would be expected. In general, benthic microalgal uptake probably represents a major fate of mineralized N compared to coupled nitrification - denitrification in net autotrophic sediments. This has been observed in Hog Island Bay, Virginia, sediments (Anderson et al. unpublished), in southern England sediments dominated by macroalgae (Trimmer et al. 2000), in the Tagus Estuary, Portugal (Cabrita and Brotas 2000), and at sites along the west coast of Sweden (Sundback et al. 2000). The DIN fluxes observed in these littoral zone systems differ markedly from those observed in sublittoral zone systems, which typically exhibit release of ammonium or ammonium plus nitrate, closer to values predicted by NEM (e.g., Giblin et al. 1997; Burdige and Zheng 1998; Hopkinson et al. 1999). The idea that benthic microalgae may significantly modify the flux of nutrients between sediment and the overlying water column was put forward more than 20 years ago (Henriksen et al. 1980). As the density of benthic microalgae - and other microbiota- in the top few mm of sediment is orders of magnitude higher than in the water column, such a filter function should be expected, but had been previously overlooked because flux measurements were traditionally made only in the dark. More recently, several studies have shown that the presence of benthic microalgae can indeed significantly decrease the net efflux of N, P and silica, or even change the direction from efflux to influx (e.g., Reay et a1.1995, Cerco and Seitzinger 1997; Sundback et al. 2000; Suzumura et al. 2002; Anderson et al. 2003; Tyler et al. 2003; see Table 2). This filter effect is mediated by both microalgal assimilation and photosynthetic oxygenation of the sediment surface, which in turn creates dynamic chemical and biological gradients. High-resolution microsensors have made it possible to study the influence of photoautotrophic activity on N turnover in more detail (see Chapter 10).

Benthic N and P cycling 243
The majority of studies on the regulatory effect of benthic microalgae on sediment- water nutrient fluxes are from lighddark incubations of flooded sediment, mimicking subtidal conditions. Such incubations do not account for the effect of emersion on fluxes in intertidal sediments that occur in many shallow estuaries (e.g., Falcao and Vale 1995). However, Thornton et al. (1999) showed that illumination during emersion reduced fluxes of ammonium to the overlying water after immersion and concluded that the effect of the illumination and emersion periods should be considered when calculating nutrient budgets for intertidal, cohesive sediments. Like benthic microalgae, accumulations of benthic macroalgae can function temporarily as a filter by intercepting dissolved nutrient fluxes from the sediments to the overlying water (Thybo-Christensen et al. 1993; McGlathery et al. 1997; Tyler et al. 2001). The effectiveness of this interception depends on the biomass and productivity of the macroalgae as well as on the stability of macroalgal accumulations. Unattached macroalgal mats tend to be patchy and unstable; however, Astill and Lavery (2001) showed that nutrient and oxygen gradients developed quickly as macroalgae accumulated (within 24 hr), suggesting that this filter function occurs even in dynamic environments.
Table 2. Ammonium and nitrate,jluxes from sediments in two temperate locations, illustrating the eflect o f benthic microalgae on the magnitude and direction of nutrientJluxes. Values represent annual means (* se) from monthly - bimonthly measurements. Fluxes from Hog
Island Bay sediments show the injluence of net autotrophy (data from Tyler et al. 2003). while those from the Swedish coastal bays show the influence of sediment type (data from Sundback
et al. 2000).
Ammonium Flux Nitrate Flux pmol N m-* d-' pmol N m'* d-'
Hog Island Bay, Virginia Autotrophic sediments -327 * 104 -78 * 29
-115 *36 -80 * 27 Heterotrophic sediments 418 * 90 9 47
384 * 126 6 % 13
Swedish west coast Sandy sediments - Vallda 108 * 1 13 -87 * 64
145 * 72 -396 * 325 Silty sediments - 337 124 -46 * 130
RotAngen 306* 110 -161 + 149
Most studies have focused on macroalgal interception of dissolved inorganic nutrient fluxes (e.g., Thybo-Christesen et al. 1993; McGlathery et al. 1997); however, macroalgae also may be important in regulating the flux of dissolved organic nutrients across the sediment-water interface. Tyler et al. (in press) showed that the chlorophyte Ulva lactuca prevented the flux of urea and DFAA from the

244 K.J. McGlathery, K. Sundback and I.C. Anderson
sediments to the overlying water. Sediments are considered to be a significant source of nutrients to supporting production of macroalgae, since nutrient concentrations are typically one to several orders of magnitude higher in sediment porewaters than in the water column, even in eutrophic waters. This has been shown in Hog Island Bay, Virginia where the efflux of DIN and urea from sediments underlying macroalgal mats was sufficient to meet 27-75% of the macroalgal N demand (Tyler et al. 2003). Likewise, Stimson and Lamed (2000) determined that the N efflux from the sediments of a subtropical bay was essential to meet the growth demand of the dominant macroalgae and was at least in part responsible for the persistence of macroalgae in an otherwise oligotrophic environment. Sediment nutrient efflux is typically consumed in the bottom layers of dense macroalgal mats, and diffusion from the overlying water provides a necessary nitrogen supply for the light-saturated algae in the upper layers (McGlathery et al. 1997). Factors that reduce macroalgal productivity, such as decreased water clarity from suspended sediments or high phytoplankton biomass, decreased insolation, or self- shading within the lower layers of dense mats during summer also decrease macroalgal uptake and reduce the filter effect (Sfriso et al. 1992; D'Avanzo and Kremer 1994; McGlathery et al. 1997). Often one, or a combination of these factors, leads to the late-summer collapse of the macroalgal population, and the release of plant-bound nutrients to the water column. Macroalgal mats that are light- limited may show a diurnal pattern of nutrient release, with nutrients diffusing from the sediments through the mat to the water column in the dark (McGlathery et al. 1997). This occurs because algae growing in low-light conditions are typically N- saturated and lack carbon reserves; as such, they are dependent on recent photosynthate to build amino acids. This close coupling between nutrient uptake and photosynthesis is also characteristic of N-replete phytoplankton (Turpin 1991 and references therein). There have been fewer studies of mechanisms by which seagrass communities influence benthic dissolved nutrient fluxes. Like benthic micro- and macro-algae, one would expect assimilatory nutrient uptake by seagrasses to reduce the flux of nutrients across the sediment-water interface. Risgaard-Petersen & Ottosen (2000) in a study performed in Denmark showed a seasonal pattern of DIN exchange between eelgrass (2. marina) communities and the water column similar to that which has been shown for sediments with benthic microalgal and macroalgal communities. There was an influx of N to eelgrass-vegetated sediments in the spring and summer when plant metabolism was highest, and a smaller influx in the colder winter months. In the fall when plant N demand was low, but decomposition rates were relatively high, nutrients were released to the water column. In studies performed in Virginia, where seagrass growth is high in spring and fall and decomposition rates high in July - August, seasonal patterns of nutrient exchange were slightly different from those in Denmark in that there was a large efflux of DIN (primarily ammonium) to the water column in June and August and a negligible efflux in April and October (Anderson and Moore, unpublished data). Both studies illustrate the important influence of assimilatory nutrient uptake by seagrasses on the direction of nutrient fluxes at the sediment-water interface. In tropical carbonate sediments, P fluxes are typically low (nM), in part because the P is bound in the solid phase (Jensen et al. 1998) and in part because turnover rates in sediment porewaters are high to meet seagrass demand (McGlathery et al. 2001 b).

Benthic N and P cycling 245
Other primary producers associated with seagrass communities also can play a key role in regulating the magnitude and direction of the nutrient flux. Miyajima et al. (2001) found an influx of N in both bare and seagrass-vegetated carbonate sediments, and attributed this at least in part to high benthic microalgal activity. The presence of epiphytes on seagrass blades also may be important, as Eyre and Ferguson (2002) found a net influx of N in warm-temperate Australian seagrass communities during light incubations only when epiphytes were included. Likewise, it has been shown that epiphytes can remove more ammonium and nitrate from the water column than the seagrass Ruppia megacarpa even though their biomass was significantly less (Dudley et al. 2001).
4. COMPETITION BETWEEN PRIMARY PRODUCERS
There is ample evidence to support the general hypothesis that phytoplankton and benthic primary producers compete for resources (nutrients, light) in shallow coastal waters (e.g., Sand-Jensen and Borum 1991; Fong et al. 1993), and that the outcome of this competition is unstable. Likewise, competition between benthic primary producers occurs, especially in systems that receive high external nutrient loads (Valiela et al. 1997 and references therein). Shifts in primary producer dominance that result from this competition have an impact on the rates and pathways of nutrient transformations within these systems. Most studies indicate that at certain times of the year the reduction, or elimination, of the internal supply of nutrients to the water column from sediment regeneration that is attributed to benthic microalgal uptake decreases nutrient availability for phytoplankton (e.g., Sundback et al. 2000; Suzumura et al. 2002). However, Cerco and Seitzinger (1997) have suggested that benthic microalgae actually enhance phytoplankton production on an annual time scale. By sequestering mineralized nutrients in the winter and spring, these authors suggest that benthic microalgae may function as a temporary storage pool for regenerated nutrients; nutrients are released in the summer when the benthic microalgae become light-limited by increased phytoplankton biomass in the water column. The net effect is to extend the summer phytoplankton bloom beyond the time when it would normally become nutrient- limited. It has been suggested that uptake of mineralized nutrients in the sediment by benthic microalgae also may inhibit, or at least defer, the initiation of benthic macroalgal blooms in the spring. A study in shallow sheltered embayments in Sweden suggested that benthic microalgae could successfully compete with ephemeral green algae for the sediment pool of regenerated nutrients during the period that is critical for the onset of the macroalgal growth (Sundback and Miles 2002). On a daily basis, the benthic microalgae decreased the efflux of inorganic N by 30 - loo%, P by 70 - 100% and silica by 1&95%. Dense macroalgal mats, in turn, decrease light availability at the sediment surface (90-100% for macroalgal biomass 100-500 gdw m-2; Krause-Jensen et al. 1996; Astill and Lavery 2001), and sediments underlying stable algal accumulations tend to be heterotrophic (Tyler et al. 2003). Surprisingly, there have been few direct studies of the effect of mat-forming macroalgae on benthic microalgal productivity (Sundback et al. 1990, 1996a). Persistent, dense macroalgal mats would be expected

246 K.J. McGlathery, K. Sundback and I.C. Anderson
to decrease production of sediment microalgal communities by reducing light levels and by creating anoxic conditions at the sediment surface. In Hog Island Bay, Virginia, benthic microalgal production increased in importance following a mid- summer macroalgal decline, suggesting that competition for light limited benthic microalgal production when macroalgal densities were high (McGlathery et al. 2001a). A more recent study in this lagoon found an inverse relationship between macroalgal biomass and both sediment chlorophyll a and sediment dissolved oxygen production over an annual cycle (Tyler et al. 2003), indicating decreased benthic microalgal activity in sediments underlying macroalgal accumulations. At lower densities, or when macroalgae are floating in the water column and thereby do not create intense shading and/or bottom water anoxia, sediment microalgal production would not be as strongly influenced (Sundback et al. 1996a). By sequestering and storing sediment-derived nutrients, dense macroalgal mats also may outcompete phytoplankton during the spring and early summer when macroalgal growth rates are highest (Viaroli et al. 1992; McGlathery et al. 1997; Valiela et al. 1997). This probably accounts for the high water clarity in many shallow macroalgal-dominated systems, despite high external nutrient loads (Valiela et al. 1997). Macroalgae also can sustain maximum growth rates for longer periods of time than phytoplankton because their nutrient storage capacity obviates the need for a constant nutrient supply (Fong et al. 1993; Pedersen and Borum 1996). Stored nutrients in phytoplankton can only sustain maximum growth for a day or less, whereas growth of bloom-forming macroalgae such as Ulva, Cladophora and Chaetomorpha can be sustained by stored nutrients for several days, or more, depending on growth conditions (light and temperature) (Borum 1996). However, the outcome of competition also can be reversed when phytoplankton blooms begin to shade macroalgal communities. This usually happens when macroalgal metabolism slows down in late summer and the macroalgae become a less effective filter for the effluxed sediment nutrients. As a consequence, nutrients diffuse into the water column and stimulate phytoplankton growth, which, in turn, further reduces macroalgal productivity through shading. Valiela et al. (1997) observed interannual variation in Waquoit Bay, Massachusetts, that was consistent with this model. In the most N-enriched subestuary, phytoplankton abundance and activity increased at the same time that macroalgal biomass declined. Many studies have shown that low macroalgal biomass coincides with peak phytoplankton biomass (e.g., Sfriso et al. 1992; Viaroli et al. 1992). Typically, these moderately-enriched systems are unstable, and are characterized by shifts in dominance between benthic macroalgae and phytoplankton. The same scenario has been suggested for systems in which microalgae dominate the benthic community and where shading by phytoplankton blooms can cause a temporary collapse of benthic microalgal productivity (Blanchard and Montagna 1995; Cerco and Seitzinger 1997). The competition between rooted macrophytes and benthic microalgae has not been studied specifically. One might expect low benthic microalgal biomass and production in densely-vegetated seagrass systems due to shading of the sediment by the leaf canopy, yet many studies have shown that benthic microalgae can contribute significantly to the total productivity of seagrass beds (e.g., Murray and Wetzel 1987; Moncrieff et al. 1992). The same has been observed in Spartina alternzyora vegetated intertidal sediments (Anderson et al. 1997). In some seagrass systems, benthic microalgal biomass and primary production has been found to be as high as

Benthic N and P cycling 247
in bare sediments (e.g., Moncreiff et al. 1992, Pollard and Kogure 1993, Kemp and Comwell 2001), or even higher (Miyajima et al. 2001). The contribution of this production to nutrient retention and turnover in seagrass beds is unclear. Hansen et al. (2000) found that uptake by benthic microalgae in a temperate seagrass bed was the major sink for DIN in the spring. Likewise, Kemp and Comwell (2001) found that benthic microalgal N uptake was as important as that by seagrasses in Florida Bay sediments in August. It is likely that the interaction between benthic microalgae and seagrass varies seasonally with changes in light and nutrient availability as the seagrass canopy develops. In nutrient-enriched systems, macroalgal blooms can outcompete seagrasses, although the outcome of this competition is reversible if macroalgal biomass decreases. Hauxwell et al. (2000) showed that dense macroalgal canopies caused a decrease in recruitment of new shoots and rates of leaf appearance in existing shoots of the eelgrass Z. marina in an enriched subestuary of Waquoit Bay, Massachusetts. Using model calculations to estimate light attenuation by the different autotrophs (phytoplankton, epiphytes, macroalgae), they confirmed that the primary cause of eelgrass loss was light reduction by the macroalgal canopy, in particular for newly recruiting shoots. One might expect that some tropical seagrasses, such as Thalassia testudinurn, that have a proportionately greater allocation to belowground biomass (and carbon reserves) may be more resilient to ephemeral shading by macroalgal canopies. Increased ammonium concentrations within macroalgal mats (>25 pM) also may be toxic to seagrass (van Katwijk et al. 1997), and again, this effect is likely to be most important for newly recruiting shoots that exist entirely within the macroalgal canopy. A decrease in redox and an increase in sediment sulfide concentrations resulting from decomposition in the anoxic, organic-rich sediments and decaying macroalgal layer also may reduce seagrass photosynthesis (Goodman et al. 1995; but see Terrados et al. 1999). Decreased photosynthetic oxygen production at all light levels also decreases the potential for oxygen translocation and release to the rhizosphere, and creates a positive feedback that reduces sulfide oxidation around the roots and hrther elevates sediment sulfide levels, which decrease nutrient uptake and plant energy status (Pregnall et al. 1984).
5. THE ROLE OF PRIMARY PRODUCERS IN THE ESTUARINE 'FILTER' - FATE AND RETENTION OF ASSIMILATED NUTRIENTS
Nutrient assimilation and temporary retention is probably the most important process regulating the source-sink role of benthic communities. In the short term (days-months), uptake and temporary assimilation influence the fluxes of nutrients between the sediment and water column. Carbon and nitrogen that are fixed may be rapidly re-released to the water column as DOC or DON. The DOM released by benthic autotrophs may be processed by the microbial loop, mineralized, or transferred to higher trophic levels. Abiotic processes such as humification are also operative in sediments. In the long term, while most of the biomass of benthic primary producers is decomposed, grazed, or exported from the system, some is retained for possible burial (Cebrian 2002; see Chapter 8). Seagrass-dominated systems have higher rates of permanent burial in the sediments because of the

248 K.J. McGlathery, K. Sundback and I.C. Anderson
inherently greater refractory content of seagrass tissue and the presence of significant quantities of belowground material (roots and rhizomes) that are less likely to be exported (Buchsbaum et al. 1991; Enriquez et al. 1993; Duarte et al. 1996; Klap et al. 2000; see Chapter 8). However, decay rates of seagrass leaves can be high, and similar in magnitude as those for macroalgae (Buchsbaum et al. 1991). Risgaard-Petersen and Ottosen (2000) found for Z. marina that the pool of tissue- bound N did not accumulate in the leaves on an annual time scale. Nutrients that were temporarily retained in leaf tissue were lost when the leaves were detached and transported from the systems, or if retained, were nearly completely mineralized. (95-98%) within a year. Similarly, most of the fixed C and N in macroalgae is released during senescence over a period of several months (Buchsbaum et al. 1991). Benthic microalgae also generally have high growth rates and low CM contents; thus a rapid turnover of N in benthic microalgae would be expected. Calculating assimilation is difficult because benthic primary producers fix more carbon and nutrients than are needed to meet the metabolic needs for growth and some of the fixed C and N (and probably P) is leaked from living tissue. For example, the release of up to 22% of DlN uptake by macroalgae as DON compounds, means that calculations of macroalgal N assimilation based on growth rates and tissue C:N content are severe underestimates (Tyler et al. 2003). Similarly, calculations of benthic microalgal nitrogen demand are subject to potential errors of either over- or underestimation. Calculations of N demand may be based either on measurements of net primary production (NPP) (Sundback and Miles 2000) or gross primary production (GPP) (Anderson et al. 2003). If NPP estimates are based on DO or DIC fluxes measured in sediment cores, sediment community respiration will result in an underestimation of microalgal NPP and, therefore, N demand. On the other hand, estimation of N demand based on GPP may result in overestimation since GPP does not take into account exudation of DOM, which may be a large component (up to 75%, see below) of the carbon fixed. Use of I3c and "N tracers would provide more realistic measures of benthic autotroph N demand; however, a complete understanding would require both inorganic and organic tracers since benthic primary producers are known to take up DON (e.g., Nilsson and Sundback 1996; Tyler et al. 2003). There have been some attempts to compare the importance of benthic microalgal and macroalgal assimilation of nutrients in coastal embayments, in which the assimilation rate of N by benthic microalgae has been found to be similar to, or even exceed, that of macroalgae (e.g., McGlathery et al. 2001 a; Sundback and Miles 2002). Carbon and nitrogen that are not used for growth may be stored in tissue, exuded as attached extracellular polymeric substances (EPS), or released to the surrounding water as colloidal organic matter (COM). For example, it has been observed that up to 75% of carbon fixed by benthic diatoms may be exuded as EPS or COM (Goto et al. 1999; Smith and Underwood 2000; Wolfstein et al. 2002). Wolfstein et al (2002) found that the percentage of fixed C excreted appeared to vary with irradiance (decreased excretion at higher irradiance) and with bacterial activity. In an in situ pulse - chase I3c labeljng experiment Middelburg et al. (2000) noted rapid transfer of I3c from benthic microalgal into bacterial biomass. They suggest that exudation of EPS may be a medium for this exchange. There are also reports of DOC leakage from macroalgal fronds which may be as high as 39% of gross production (Velimirov 1986). In contrast, there appears to be more limited release of organic

Benthic N and P cycling
compounds from living seagrass tissues. Of the carbon that is fixed during photosynthesis, only 1-2% is released by the leaves (e.g., Velimirov 1986; Moriarty et al. 1986) and up to 11% by the roots and rhizomes (Moriarty et al. 1986). Nitrogen retention in benthic primary producer communities depends in part on the leakage of DON from live tissues, yet there is considerably less known about this process than for DOC. To our knowledge no studies have demonstrated exudation of DON by benthic microalgae, although DON release has been observed for pelagic cyanobacteria such as Synechococcus spp. (Bronk, 1999) and for the nitrogen fixing Trichodesmium spp. (Glibert and Bronk, 1994). Rates of DON release have been observed to vary with light availability and nutrient status, depth of the water column, DIN species composition and concentration, and season (Ward and Bronk, 2001). The composition of DON released includes DFAA, DCAA, dissolved primary amines, and urea (Berman and Bronk, in press). DON may be'released by passive release, excretion, sloppy feeding during grazing by copepods or direct release by microzooplankton, and viral lysis. Macroalgae also may release significant quantities of DON as DCAA; Tyler et al. (in press) found benthic fluxes of DCAA that were nearly &fold higher in the presence of macroalgae, and were higher in the light, suggesting a photosynthetically-driven process (Tyler et al. 2003). We are unaware of any studies on DON release from living seagrasses.
In phytoplankton-based systems, release of DOC or DON can stimulate water column bacteria and higher trophic levels (Azam et al. 1983). It is likely that this also occurs in benthic primary producer communities, although it is more difficult to show this link. In Hog Island Bay, Virginia, highest rates of water column metabolism of DOC and DON were observed in August, the period when the macroalgal bloom declines, suggesting that the labile DOM was derived from macroalgal detritus (Lunsford, 2002). Rates of DOM metabolism in a small Massachusetts phytoplankton-dominated estuary bounded by both fresh and saltwater marshes were only 25% of those observed in the Virginia lagoon. The DOM in the Massachusetts estuary was primarily allochthonous in origin, whereas that in the Virginia coastal lagoon was mainly autochthonous. Studies such as this suggest that bacterial activity in the water column can be an important link between sediment detrital material and higher trophic levels, and may play an important role in nutrient transformations in shallow coastal systems.
The retention time of tissue-bound nutrients also depends on the transformations once assimilated, which can take three pathways in the benthic food web: 1) through the macrofaunal grazingldeposit feeding chain (Asmus and Asmus 1985; Duarte and Cebrian 1996; Herman et al. 2000), 2) through the 'small food chain' consisting of micro- and meiofauna (Kuipers et al. 1981; Sundback et al. 1996b and references therein), or 3) burial or export (Admiraal et al. 1984; Boschker et al. 1999; Middelburg et al. 2000). Since much of the organic matter is released as DOM by live and dead material, processing of detritus by grazers and bacteria only accounts for a portion of the plant organic matter. For benthic microalgae, an in situ '3~-labelling experiment showed that assimilated C was transferred within a few days through the heterotrophic components in an order bacteria > macrofauna > meiofauna (Middelburg et al. 2000). Sundback et al. (1996b) found by in situ dual- labeling ( I 4 c - ~ c o 3 , 3H-thymidine; Montagna 1984) of sediment, that meiofauna grazed between 2 and 12% of the microalgal biomass per day and had a significant impact on microalgal biomass in spring and autumn, but not in summer.

K.J. McGlathery, K. Sundback and I.C. Anderson
6. CHANGES IN NUTRIENT CYCLING WITH SHIFTS IN PRIMARY PRODUCER DOMINANCE
The generally accepted conceptual models of eutrophication (Sand-Jensen and Borum 199 1 ; Duarte 1995; Valiela et al. 1997) predict that with increasing nutrient loading, benthic primary production will decrease; sediments will become increasingly heterotrophic, resulting in increased efflux of nutrients from the sediment to the water column. Ultimately, this will cause a positive feedback accelerating eutrophication through internal loading of nutrients. Some empirical evidence supporting this hypothesiyed series of events has been found (e.g., Rizzo et al. 1992; Meyercordt and Meyer-Reil 1999; Eyre and Ferguson 2002). Initially, both seagrasses and benthic microalgae will be stimulated by increased nutrient levels, particularly in sandy sediments (Nilsson et al. 1991; Flothman and Werner 1992). However, at the same time the growth of filamentous macroalgae and epiphytes will be stimulated (e.g., Nilsson et al. 1991; Havens et al. 2001), with increased shading of the benthos. Systems dominated by fast-growing macroalgae and phytoplankton are characterized by short-lived (days-months) bloom events and rapid turnover of nutrients bound in plant biomass. Whether and when planktonic or macroalgal communities outcompete benthic microalgae and seagrasses by shading will depend on the duration and extent of bloom events. We hypothesise that, with proceeding eutrophication and light attenuation from high phytoplankton biomass, the negative impact on the microphytobenthic community will be more gradual and slower than for benthic macroscopic primary producers, including seagrasses and macroalgae. Thus, a partial beneficial "buffering" effect of benthic microalgae on shallow sediment systems may persist even in more heavily eutrophied systems, as has been shown by Krom et al. (1991) for a hypertrophic fish pond. This scenario is based on the assumption that benthic microalgal communities possess, due to high diversity and functional redundancy, a certain degree of plasticity, increasing the overall resilience of shallow-water sediment systems after pelagic bloom events. Benthic diatoms can survive periods of only a few % of incident light, or even darkness, and high sulphide levels, and can rapidly resume photosynthesis when exposed to light or after an anoxic event (Admiraal et al. 1984; Kennett and Hargraves 1985; Sundback and GranCli 1988; Sundback et al. 1990). Similarly, upward migrating benthic diatoms were found to rapidly restore the oxygenation of the sediment surface after a simulated sedimentation event (Wulff et al. 1997; Underwood and Paterson 1993). This scenario, with benthic microalgae surviving despite deteriorating conditions, may apply particularly to cool microtidal waters, where macroalgal bloom events last only a few months (e.g. Pihl et al. 1999; Dalsgaard in press; McGlathery et al. 2001a), leaving the rest of the year open to benthic microalgal primary production. In warm, eutrophic microtidal systems, benthic microalgae can be outcompeted by shading and also by dystrophic events when the macroalgal blooms eventually collapse (Viaroli et al. 1996). The efflux of N and P from the sediments will increase as eutrophication proceeds due to prolonged shading of the benthos, as suggcsted by experiments at varying light intensities (Rizzo et al. 1992; Sundback and GranCli 1988; Cerco & Setizinger 1997). This will further stimulate pelagic primary productivity, while the sediment system will become more heterotrophic. Several studies have specifically pointed

Benthic N and P cycling 25 1
out that the autotrophyheterotrophy balance is a good indicator of nutrient flux from sediments (e.g., McGlathery et al. 2001a; Eyre and Fergusson 2002; Laursen and Seitzinger 2002; Sundback et al. 2003; Tyler et al. 2003), with net autotrophic systems functioning as sinks and net heterotrophic systems as sources. As systems become increasingly enriched with organic matter, net heterotrophy will increase and the systems will become self-generating and demonstrate hysteresis; e.g., internal nutrient loading from the sediments will continue to sustain the growth of ephemeral macroalgal mats for some period of time after nutrient loads begin to decrease (Pihl et al. 1999; Sundback et al. 2003). However, measurements from some Danish shallow fjords suggest that the release rate of the sediment N pool can be significantly lower only a few years after the nutrient load to the water column has been decreased (Dalsgaard et al. 1999). When nutrient loads are decreased, a reversed scenario has been predicted, where benthic primary producers are favored because of better light conditions, thus accelerating the process of recovery (Christensen et al. 1998) by reoxygenation of the sediment, interception of the nutrient flux, and by increased retention of nutrients in the sediment. Seagrass-dominated systems will have both a longer retention time of nutrients temporarily bound in plant material and a greater permanent removal of nutrients by burial.
7. CONCLUSION
In this chapter we have reviewed the effects of seagrass, macroalgal, and benthic microalgal communities on specific nitrogen and phosphorus cycling processes and on the ecosystem-level role of these communities as sources or sinks of nutrients. Photosynthetic and respiratory activities of benthic primary producers alter chemical conditions in the sediments, causing spatial and die1 variations in DO, DIC, pH and nutrients that influence the rates and pathways of nutrient transformations. Uptake and retention of nutrients by plant biomass and the effects of primary producers on N losses from denitrification are key regulators of sediment-water column fluxes. As such, benthic primary producers play an important role regulating phytoplankton production in the overlying water column. However, nutrients bound in plant biomass turnover rapidly and probably make little difference on annual times scales to the overall nutrient retention within the system. It also appears that although there are examples of both stimulation and suppression of denitrification rates by benthic primary producer communities, there is not a consistent and significant increase of N losses by denitrification on annual time scales caused by benthic primary producers. This being the case, the key drivers to total estuarine nutrient retention of external inputs are likely dependent on: 1) the amount of burial of refractory compounds (including particulates imported to system and trapped as well as in situ production) and 2) the water residence time (i.e., to what extent recycled nutrients support primary and secondary production within the system before being exported). Both of these processes will be dependent on the character of the primary producer community. Coupling of ecological and physical processes is central to our understanding of how shallow coastal ecosystems function as a filter of watershed nutrient inputs, yet we know relatively little about the fate and turnover of

252 K.J. McGlathery, K. Sundback and I.C. Anderson
nutrients bound in plant biomass or the complex hydrodynamics of many shallow coastal estuaries.
8. REFERENCES
Admiraal W. (1984). The ecology of estuarine sediment-inhabiting diatoms. In: Round, F. E., Chapman, D. J. (ed.) Progress in phycological research. Biopress Ltd, Bristol, p. 269-314
Alcoverro T., M. Manzanera and J. Rornero. (2000). Nutrient mass balance of the seagrass Posidonia oceanica: the importance of nutrient retranslocation. Marine Ecology Progress Series 194: 13-2 1
An S., S.B. Joye. (2001). Enhancement of coupled nitrification-denitrification by benthic photosynthesis in shallow estuarine sediments. Limnology and Oceanography 46: 62-74
Anderson I.C., K.J. McGlathery and A.C. Tyler. (2003). Microbial mediation of 'reactive' nitrogen transformations in a temperate lagoon. Marine Ecology Progress Series 246:73-84.
Anderson I.C., C.R. Tobias, B.B. Neikirk and R.L. Wetzel. (1997). Development of a process-based mass balance model for a Virginia Sparlina altern~JIora salt marsh: Implications for net DIN flux. Marine Ecology Progress Series 159, 13-27.
Asmus H. and R. Asmus. (1985). The importance of grazing food chain for energy flow and production in three intertidal sand bottom communities of the northern Wadden Sea. Helgolander Meeresuntersuchungen 39: 273-301
Astill H. and P. Lavery. (2001). The dynamics of unattached benthic tnacroalgal accumulations in the Swan-Canning Estuary. Hydrological Processes 15: 2387-2399
Azam R., T. Fenchel, J.G. Field, J.S. Gray, L.A. Meyerriel and F. Thingstad. (1983). The ecological role of water column microbes in the sea. Marine Ecology Progress Series 10: 257-263.
Berman T. and D.A. Bronk. (in press). Dissolved organic nitrogen: a dynamic participant in aquatic ecosystems. Aqzratic Microbial Ecology
Blaaberg V.K., K.M. Mouritsen and K. Finster. (1998). Diel cycles of sulphate reduction rates in sediments of a Zostera marina bed, Denmark. Aquatic Microbial Ecology 15:97-102
Blackbuni T.H., D.B. Nedwell and W.J. Wiebe. (1994). Active mineral cycling in a Jamaica seagrass sediment Marine Ecology Progress Series 1 10: 233-239
Blanchard G.F. and P.A. Montagna. (1995). Assessment of a brown tide impact on microalgal benthic communities in Baffin Bay (Texas) in 1990 using a primary production simulation model. Oceanologia Acra 18: 37 1-377
Bomm J. (1996). Shallow waters and LandISea Boundaries. In Eutrophication in Coastal Marine Ecosystem, Jsrgensen, B. B. and K. Richardson, eds. American Geophysical Union, Washington, D. C., pp. 179-204.
Borurn, J. and K. Sand-Jensen. (1996). 1s total primary production in shallow coastal matine waters stimulated by nitrogen loading? Oikos 76:406-410
Boschker H.T.S., A. Wielemaker, B.E.M. Schaub, and M. Holmer. (2000). Limited coupling of macrophyte production and bacterial carbon cycling in the sediments of Zostera spp. meadows. Marine Ecology Progress Series 203: 18 1 - 189.
Boynton W.R., J.D. Hagy, L. Murray, C. Stokes, and W.M. Kemp. (1996). A comparative analysis of eutrophication patterns in a temperate coastal lagoon. Estuaries 19: 408-42 1.

Benthic N and P cycling 253
Buchsbaum R., I. Valiela, T. Swain, M. Dzierzeski and S. Allen. (1991). Available and refractory nitrogen in detritus of coastal vascular plants. Marine Ecology Progress Series 72: 131-143.
Burdige D.J. and R.C. Zimmerman. (2002). Impact of seagrass density on carbonate dissolution in Bahamian sediments. Limnology and Oceanography 47L 1761-1 763.
Bronk D.A. (1999). Rates of NH4' uptake, intracellular transformation and dissolved organic nitrogen release in two clones of marine Synechococcus spp. Journal ofPlankton Research 2 I: 1337-1353.
Burdige D.J., S. Zheng. (1998). The biogeochemical cycling of dissolved organic nitrogen in estuarine sediments. Limnology and Oceanography 43: 1796- 18 13
Cabrita M.T. and V. Brotas. (2000). Seasonal variation in denitrification and dissolved nitrogen fluxes in intertidal sediments of the Tagus Estuary. Marine Ecology Progress Series 202: 5 1-65
Caffrey J.M. and W.M. Kemp. (1992). Influence of the submersed plant Potamogeton perfuliatus, on nitrogen cycling in estuarine sediments. Limnology and Oceanography 37: 1483-1495
Cambridge M.L. and A.J. McComb. (1984). The loss of seagrasses in Cockbum Sound, Western Australia. I. The time course and magnitude of seagrass decline in relation to industrial development. Aquatic Botany 20: 229-243
Capone D.G. (1988). Benthic nitrogen fixation: microbiology, physiology and ecology. In: Blackburn TH, J Sorensen and T Roswall (eds). Nitrogen cycling in marine coastal environments. SCOPE Series. J Wiley & Sons, New York, p 85-123
Capone D.G. and B.F. Taylor. (1977). Nitrogen fixation (acetylene reduction) in the phyllosphere of Thalassia testudinum. Marine Biology 40: 19-28.
Capone D.G., S.E. Dunham, S.G. Horrigan and L.E. Duguay. (1992). Microbial nitrogen transformations in unconsolidated coral reef sediments. Marine Ecology Progress Series 80: 75-88
Carlson P.R., L. Yarbro and T. Barber. (1994). Relationship of sediment sulfide to mortality of Thalassia testudinum in Florida Bay. Bulletin ofMarine Science 54: 733-746
Cebrian J. (2002). Variability and control of carbon consumption, export and accumulation in marine communities. Limnology and Oceanography 47: 1 1-22
Cerco C.F. and S.P. Seitzinger. (1997). Measured and modeled effects of benthic algae on eutrophication in Indian River-Rehoboth Bay, Delaware. Estuaries 20: 23 1-248
Chambers R.M., J.W. Fourqurean, S.A. Macko and R. Hoppenot. (2001). Biogeochemical effects of iron availability on primary producers in a shallow marine carbonate environment. Limnology and Oceanography 46: 1278- 1286
Christensen P.B. et al. (1998). The Danish marine environment: Has action improved its state. Danish Environmental Protection Agency. Report 62:
Comwell J.C., W.M. Kemp and T.M. Kana. (1999). Denitrification in coastal ecosystems: Environmental controls and aspects of spatial and temporal scaling. Journal of Aquatic Ecology 33: 41-54
Currin C.A., S.B. Joye and H.W. Paerl. (1996). Diel rates of N2-fixation and denitrification in a transplanted Spartina alterniflora marsh: implications for N-flux-dynamics. Estuarine Coastal and ShelfScience 42: 597-61 6
Dalsgaard T., P.B. Christensen, S. Rysgaard and N. Risgaard-Petersen. (1999). Nitrogen removal in Danish waters: importance and regulation (In Danish). In: A,, L. B. (ed.) Havmilj0et ved hs indsk i f te t ( Marine Environments into the Millenium). Vol: Olsen & Olsen, Fredensborg, p. 101- 118

254 K.J. McGlathery, K. Sundback and I.C. Anderson
D'Avanzo C. and J.N. Kremer. (1994). Diel oxygen dynamics and anoxic events in an eutrophic estuary of Waquoit Bay, Massachusetts. Estuaries 17: 13 1-39
Duarte C.M. (1995). Submerged aquatic vegetation in relation to different nutrient regimes. Ophelia 41: 87- 1 12.
Duarte C.M. and J. Cebrian. (1996). The fate of marine autotrophic production. Limnology and Oceanography 4 1 : 1758- 1766
Dudley B.J., A.M.E. Gahnstrom and D.I. Walker. (2001). The role of benthic vegetation as a sink for elevated inputs of ammonium and nitrate in a mesotrophic estuary. Morine Ecology Progress Series 2 19: 99- 107.
Enriquez S., C.M. Duarte and K. Sand-Jensen. (1993). Patterns in decomposition rates among photosynthetic organisms: The importance of detritus C:N: P content. Oecologia 94: 457-471.
Erskine J.M. and M.S. Koch. (2000). Sulfide effects on Thalassia testudinum carbon balance and adenylate energy charge. Aquatic Botany 67: 275-285
Eyre B.D. and A.J.P. Ferguson. (2002). Comparison of carbon production and decompositopn, benthic nutrient fluxes and denitrification i seagrass, phytoplankton, benthic microalgae- and macroalgae- dominated warm-temperate Australian lagoons. Marine Ecology Progress Series 229: 43-59
Falcao M. and C. Vale. (1995). Tidal flushing of ammonium from intertidal sediments of Ria Formosa, Portugal. Netherlands Journal of Aquatic Ecology 29: 239-244
Flindt M.R. (1994). Measurements of nutrient fluxes and mass balances by on-line in situ dialysis in a Zostera marina bed culture. Verhandlungen Internationale Vereinigung Limnologie 25:2259-2264
Flothmann S. and I . Werner. (1992). Experimental eutrophication on an intertidal sandflat: effects on microphytobenthos, meio- and macrofauna. In: Colombo G, I Ferrari, V Ceccherelli, and R Rossi (ed.) Marine eutrophication and population dynamics. Vol: Olsen & Olsen, Fredensborg, p. 93-100
Fong P., R.M. Donohoe and J.B. Zedler. (1993). Competition with macroalgae and benthic cyanobacterial mats limits phytoplankton abundance in experimental microcosms. Marine Ecology Progress Series 100: 97- 102
Gacia E., C.M. Duarte and J.J. Middelburg. (2002). Carbon and nutrient deposition in a Mediterranean seagrass (Posidonia oceanica) meadow. Limnology and Oceanography 47,: 23-32.
Giblin A. and R.W. Howarfh. (1984). Pore water evidence for a dynamic sedimentary iron cycle in salt marshes. Limnology and Oceanography 29: 47-63.
Giblin A.E., C.S. Hopkinson Jr and J. Tucker. (1997). Benthic metabolism and nutrient cycling in Boston Harbor, Massachusetts. Estuaries 20:346-364
Glibert P.M. and D.A. Bronk. (1994). Release of dissolved organic nitrogen by marine diazotrophic cyanobacteria, Trichodesmium spp. Applied and Environmental Microbiology 60: 3996-4000.
Goodman J.L., K.A. Moore and W.C. Dennison. (1995). Photosynthetic responses of eelgrass (Zostero marina L.) to light and sediment sulfide in a shallow barrier island lagoon. Aquatic Botany 50: 37-47
Goto N., T. Kawamura, 0. Mitamura and H. Terai. (1999). Importance of extracellular organic carbon production in the total primary production by tidal-flat diatoms in comparison to phytoplankton. Marine Ecology Progress Series 190:289-295.
Hansen K, and E. Kristensen. (1997). Impact of macrofaunal recolonization on benthic metabolism and nutrient fluxes in a shallow marine sediment previously overgrown with macroalgal mats. Estuarine Coastal and ShelfScience 45:6 13-628.

Benthic N and P cycling 255
Hansen J.W., J.W. Udy, C.J. Perry, W.C. Dennison and B.A. Lomstein. (2000). Effect of the seagrass Zosfera capricorni on sediment microbial processes. Marine Ecology Progress Series 199: 83-96
Hauxwell J., J. Cebrian, C. Furlong and I. Valiela. (2000). Macroalgal canopies contribute to eelgrass (Zosfera marina) decline in temperate estuarine ecosystems. Ecology 82: 1007- I022
Havens K.E. , J. Hauxwell, A.C. Tyler, S. Thomas, K.J. McGlathery, J. Cebrian, I . Valiela, A.D. Steinman and S.-J. Hwang. (2001). Complex interactions between autotrophs in shallow marine and freshwater ecosystems: implications for community responses to nutrient stress. Environmental PoNution 1 13: 95-107
Hemminga, M.A., P.G. Hamson and F. van Lent. (1991). The balance of nutrient losses and gains in seagrass meadows. Marine Ecology Progress Series 7 1 :85-96
Henriksen K., J. Hansen and T.H. Blackbum. (1980). The influence of benthic infauna on exchange rates of inorganic nitrogen betwcen sediment and water. Ophelia Suppl. I: 249-256 .
Henriksen K. and M.W. Kemp. (1988). Nitrification in estuarine and coastal marine sediments 201-255. In: Blackburn TH and J Ssrensen [eds.], Nitrogen cycling in coastal marine environments, SCOPE. John Wiley & Sons Ltd
Herman P.M.J., M.J. Widdows, C.H. Licas and C.H.R. Heip. (2000). Stable isotopes as trophic tracers: combining field sampling and manipulative labelling of food resources for macrobenthos. Marine Ecology Progress Series 204: 79-92
Hines M. E. and W.B. Lyons. (1982). Biogeochemistry of nearshore Bermuda sediments. I. Sulfate reduction rates and nutrient generation. Marine Ecology Progress Series 8: 87-94
IHoffland E. (1992). Quantitative evaluation of the role of organic acid exudation in the mobilization of rock phosphate by rape. Plant Soil 140: 279-289.
Hopkinson C.S., A.E. Giblin and J. Tucker. (2001). Benthic metabolism and nutrient regeneration on the continental shelf of Eastern Massachusetts, USA. Marine Ecology Progress Series 224: 1- 19.
Hopkinson C.S., A.E. Giblin, J. Tucker and R.H. Garritt. (1999). Benthic metabolism and nutrient cycling along an estuarine salinity gradient. Estuaries 222363-881
Howarth R.W., R. Marino, J. Lane and J.J. Cole. (1988). Nitrogen fixation in freshwater, estuarine, and marine ecosystems. I . Rates and importance. Limnology and Oceanography 33: 669-687
Jensen H.S., K.J. McGlathery, R. Marino and R.W. Howarth. (1998). Forms and availability of sediment phosphorus in carbonate sand of Bermuda seagrass beds. Limnology and Oceanography 43: 799-8 10
Kemp W.M. and J.C. Cornwell. (2001). Role of benthic communities in the cycling and balance of nitrogen in Florida Bay. Final report to the U.S. Environmental Protection Agency, Region 4, Atlanta, GA. 53 pp.
Kennett D.M. and P.E. Hargraves. (1985). Benthic diatoms and sulfide fluctuations: upper basin of Pettaquamscutt River, Rhode Island. Estuarine Con~tal and ShevSciencc 2 1 : 577-586
Klap V.A. M.A. Hemminga and J.J. Boon. (2000). Retention of lignin in seagrasses: angiosperms that returned to the sea. Marine Ecology Progress Series 194: 1 - 1 1.
Knight W.G., L.M. Dudley and J.J. Jurinak. (1992). Oxalate effects on solution phosphorus in a calcareous soil. Arid Soil Research and Rehabilitation 6: 11-20.
Koepfler E.T., R. Benner, and P.A. Montagna. (1993). Variability of dissolved organic carbon in sediments of a seagrass bed and an unvegetated area within an estuary in southem Texas. Esluaries 16: 391404

256 K.J. McGlathery, K. Sundback and I.C. Anderson
Krause-Jensen D., K.J. McGlathery, S. Rysgaard and P.B. Christensen. (1996). Production within dense mats of the filamentous macroalga Chaetomorpha lrnum in relation to light and nutrient availability. Marine Ecology Progress Series 134: 207-2 16
Krause-Jensen D., P.B. Christensen and S. Rysgaard. (1999). Oxygen and nutrient dynamics within mats of the filamentous macroalga Chaeotomorpha linum. Estuaries 22: 3 1-38.
Kristensen E., F.O. Andersen, N. Holmboe, M. Holmer and N. Thongtham. (2000). Carbon and nitrogen mineralization in sediments of the Rangrong mangrove area, Phuket, Thailand. Aquatic Microbial Ecology 22: 199-2 13
Krom M.D. (1991). Importance of benthic productivity in controlling the flux of dissolved inorganic nitrogen through the sediment-water interface in a hypertrophic marine ecosystem. Marine Ecology Progress Series 78: 163- 172
Kuipers B.R., P.A.W.J. de Wilde and F. Creutzberg. (1981). Energy flow in a tidal flat ecosystem. Marine Ecology Progress Series 5: 2 15-22 1
Laursen A.E. and S.P. Seitzinger. (2002). Measurement of denitrification in rivers: an integrated, whole reach approach. Hydrobiologia 485: 67-8 1.
Lomstein B.A., A.-G.U. Jensen, J.W. Hansen, I.B. Andreasen, L.S. Hansen, J. Bemtsen and H. Kunzendorf. (1998). Budgets of sediment nitrogen and carbon cycling in the shallow water of Knebel Vig, Denmark. Aquatic Microbial Ecology 14: 69-80
Lorenzen J., L.H. Larsen, T. Kjaer and N.P. Revsbech. (1998). Biosensor determination of the microscale distribution of nitrate, nitrate assimilation, nitrification, and denitrification in a diatom-inhabited freshwater sediment. Applied and Environmental Microbiology 64: 3264-3269
Lunsford T. (2002). A comparison of the fate of dissolved organic matter in two coastal systems: Hog Island Bay, VA (USA) and Plum Island Sound, MA (USA). Ph.D. Thesis, Virginia institute of Marine Science, College of William and Mary.
McComb A.J. and R. Humphries. (1992). Loss of nutrients from catchments and their ecological impacts on the Peel-harvey estuarine system, Western Australia. Estuaries 15: 529-537.
McGlathery K.J., D. Kraose-Jensen. S. Rysgaard and P.B. Christensen. (1997). Patterns of ammonium uptake within dense mats of the filamentous macroalga Chaetomorpha linum. Aquatic Botany 59: 99-115
McGlathery K.J., N. Risgaard-Petersen and P.B. Christensen. (1998). Temporal and spatial variation in nitrogen fixation activity in the eelgrass (Zosreru marina L.) rhizosphere. Marine Ecology Progress Series 168: 245-258
McGlathery K.J., I.C. Anderson and A.C. Tyler. (2001a). Magnitude and variability of benthic and pelagic metabolism in a temperate coastal lagoon. Marine Ecology Progress Series 216: 1-15
McGlathery K.J., P. Berg and R Marino. (2001 b). Using porewater profiles to assess nutrient availability in seagrass-vegetated carbonate sediments. Biogeochemistry 56: 239-263
Meyercordt J. and L.A. Meyer-Reil. (1999). Primary production of benthic microalgae to two shallow coastal lagoons of different trophic status in the southern Baltic Sea. Marine Ecology Progress Series 178: 179-191
Middelburg J.J., C. Barranguet, H.T.S. Boschker, P.M.J. Herman, T. Moens and C.H.R. Heip. (2000). The fate of intertidal microphytobenthos carbon: an in situ 13C-labelling study. Limnology and Oceanography 45: 1224- 1234

Benthic N and P cycling 257
Miyajima T., M. Suzumura, Y. Umezawa and 1. Koike. (2001). Microbial nitrogen transformation in carbonate sediments of a coral-reef lagoon and associated seagrass beds. Marine Ecology Progress Series 2 17: 273-286
Moncreiff C.A., M.J. Sullivan, and A.E. Daehnick. (1992). Primary production in seagrass beds of Mississippi Sound: the contributions of seagrass, epiphytic algae, sand microflora, and phytoplankton. Marine Ecology Progress Series 87: 16 1-17 1
Montagna P.A.. (1984). In situ measurement of meiobenthic grazing rates on sediment bacteria and edaphic diatoms. Marine Ecology Progress Series 18: 119-130
Moriarty D.W., J.R. lverson and P.C. Pollard. (1986). Exudation of organic carbon. by the seagrass Halodule wrightii and its effect on bacterial growth in the sediment. Journal of Experimerzfal Biology and Ecology 96: 1 15- 126
Moriarty D.W. and M.J. O'Donohue. (1993). Nitrogen fixation in seagrass communities during summer in the Gulf of Carpentaria, Australia. Australian Jot~rnal of Marine and Freshwafer Research 44: 117-125
Murray L. and R.L. Wetzel. (1987). Oxygen production and consumption associated with the major autotrophic components in two temperate seagrass communities. Marine Ecology Progress Series 38: 23 1-239
Nielsen L.P.. (1992). Denitrification in sediment determined from nitrogen isotope pairing. FEMS Microbiologv Ecology. 86: 357-362
Nilsson C. and K. Sundback. (1996). Amino acid uptake in natural microphytobenthic assemblages studied by microautoradiography. Hydrobiologia 332: 1 19-129.
Nilsson P., B. Jonsson, Lindstrom, I. Swanberg and K. Sundback. (1991). Response of a marine shallow- water sediment system to an increased load of inorganic nutrients. Marine Ecology Progress Series 7 1 : 275-290
Nixon S.W. et al. (1996). The fate of nitrogen and phosphorus at the land-sea margin of the North Atlantic Ocean. Biogeochernistry 35: 14 1 - 180.
National Research Council. (2000). Clean coastal waters: understanding and reducing the effects of nutrient pollution. National Academy Press, Washington, DC.
O'Donohue M.J., D.J. Moriarty and I.C. MacRae. (1991). Nitrogen fixation in sediments and the rhizosphere of the seagrass Zoslera capricorni. Microbial Ecology 22: 53-64
O'Neil J.M. and D.G. Capone. (1989). Nitrogenase activity in tropical carbonate marine sediments. Marine Ecology Progress Series 56: 145- 156
Onosen, L.D.M., N. Risgaard-Petersen and L.P. Nielsen. (1999). Direct and indirect measurements of nitrification and denitrification in the rhizosphere of aquatic macrophytes. Aqualic Microbial Ecology 19:81-91
Pedersen M.F. and J. B o N ~ . (1996). Nutrient control of algal growth in estuarine waters. Nutrient limitation and the importance of nitrogen requirements and nitrogen storage among phytoplankton and species of rnacroalgae. Marine Ecology Progress Series 142: 261-272
Pedersen M.F., C.M. Duarte and J. Cebrian. (1997). Rates of changes in organic matter and nutrient stocks during seagrass Cymodocea nodosa colonization and stand development. Marine Ecology Progress Series 159: 29-36.
Pelegri S.P., L.P. Nielsen and T.H. Blackbum. (1994). Denitrification in estuarine sediment stimulated by the irrigation activity of the amphipod Corophium volu~ator. Marine Ecology Progress Series 1051285-290.

258 K.J. McGlathery, K. Sundback and I.C. Anderson
Perez M., M.A. Mateo, T. Alcoverro and J. Romero. (1997). Variability in detritus stocks in beds of the seagrass Cymodocea nodosa. Botanica Marina 44: 523-53 I .
Pihl L., A. Svenson, P.O. Moksnes and H. Wennhage. (1999). Distribution ofgreen algal mats throughout shallow soft bonoms of the Swedish Skagerrak archipelago in relation to nutrient sources and wave exposure. Journal of Sea Research 4 1 : 28 1-294
Pollard P.C. and K. Kogure. (1993). The role of epiphytic and epibenthic algal productivity in a tropical seagrass, Syringodium isoetifolium (Aschers) Dandy, community. Australian Journal of Marine and Freshwater Research 44: 14 1 - 154
Pregnall A.M., R.D. Smith, T.A. Kursar and R.S. Alberte. (1984). Metabolic adaptations of Zostera marina (eelgrass) to diurnal periods of root anoxia. Marine Biology 83: 14 1- 147
Reay W.G., D.L. Gallagher and G.M.J. Simmons. (1995). Sediment-water column oxygen and nutrient fluxes in nearshore environments of the lower Delmawa Peninsula, USA. Marine Ecology Progress Series 1 18: 2 15-227
Risgaard-Petersen N. (2003). Coupled nitrification-denitrification in autotrophic and heterotrophic estuarine sediments: On the influence of benthic microalgae. Limnology and Oceanography 48: 93- 105
Risgaard-Petersen N., S. Rysgaard, L.P. Nielsen and N.P. Revsbech. (1994). Diurnal variation of denitrification and nitrification in sediments colonized by benthic microphytes. Limnology and Oceanography 39: 573-579.
Risgaard-Petersen N., T. Dalsgaard, S. Rysgaard, P.B. Christensen, J. Borum, K.J. McGlathery and L.P. Nielsen. (1998). Nitrogen balance of a temperate eelgrass Zostera marina bed. Marine Ecology Progress Series 174: 28 1-29 1
Risgaard-Petersen N. and L.D.M. Ottosen. (2000). Nitrogen cycling in two temperate Zostera marina beds: seasonal variation. Marine Ecology-Progress Series 198:93-107
Rizzo W.M., S.K. Dailey, G.J. Lackey, R.R. Christian, B.B. Beny, and R.L. Wetzel. (1996). A metabolism-based trophic index for comparing the ecological values of shallow-water sediment habitats. Estuaries. 19: 247-256
Rizzo W.M., G.L. Lackey, and R.R. Christian. (1992). Significance of euphotic, subtidal sediments to oxygen and nutrient cycling in a temperate estuary. Marine Ecology Progress Series 86: 5 1-61
Rozan T.F., M. Taillefert, R.E. Trouwborst, B.T. Glazer, S. Ma, J. Herszage, L.M. Valdes, K.S. Price and G.W. Luther 111 (2002). Iron-sulhr-phosphorus cycling in the sediments of a shallow coastal bay: Implications for sediment nutrient release and benthic macroalgal blooms. Limnology and Oceanography 47: 1346- 1354.
Rysgaard S., P.B. Christensen and L.P. Nielsen. (1995). Seasonal variation in nitrification and denitrification in estuarine sediment colonized by benthic microalgae and bioturbating infauna. Marine Ecology Progress Series 126: 1 1 1 - 1 2 1
Rysgaard S., N. Risgaard-Petersen and N.P. Sloth. (1996). Nitrification, denitrification, and nitrate ammonification in sediments of two coastal lagoons in Southern France. Hydrobiologia 329: 133-141
Sand-Jensen, K. and Bomm J. (1991). Interactions among phytoplankton, periphyton, and macrophytes in temperate freshwaters and estuaries. Aquatic Botany 41: 137-175.
Sfriso A,, B. Pavoni, A. Marcomini and A.A. Orio. (1992). Macroalgae, nutrient cycles, and pollutants in the lagoon of Venice. Estuaries 15: 5 17-528

Benthic N and P cycling
Sloth N.P., H. Blackbum, L.S. Hansen, N. Risgaard-Petersen and B.A. Lomstein. (1995). Nitrogen cycling in sediments with different organic loading. Marine Ecology Progress Series 116: 163-170
Smith D.J. and G.J.C. Underwood. (2000). Exopolymer production by intertidal epipelic diatoms. Limnology and Oceanography 43: 1578- 159 1.
Stapel J. and M.A. Hemminga. (1997). Nutrient resorption from seagrass leaves. Marine Biology 128: 197-206
Stimson J. and S.T. Lamed. (2000). Nitrogen efflux from the sediments of a subtropical bay and the potential contribution to macroalgal nutrient requirements. Journal of Experimental Marine Biology and Ecology 252: 159-1 80
Sundback K. and W. GranCli. (1988). Influence of microphytobenthos on the nutrient flux between sediment and water: a laboratory study. Marine Ecology Progress Series 43: 63-69
Sundback K., B. Jonsson, P. Nilsson and I. Lindstrom. (1990). Impact of accumulating drilling macroalgae on a shallow-water sediment system: an experimental study. Marine Ecology Progress Series 58: 261 -274
Sundback K., L. Carlson, C. Nilsson, B. Jonsson, A. Wulff and S. Odmark. (1996a). Response of benthic microbial mats to drifting green algal mats. Aquatic Microbial Ecology 10: 195-208
Sundback K., P. Nilsson, C. Nilsson and B. Jonsson (1996b). Balance between autotrophic and heterotrophic components and processes in the sandy sediments: a field study. Estuarine Coastal and ShelfScience 43: 689-706
Sundback K. and A. Miles. (2000). Balance between denitrification and microalgal incorporation of nitrogen in microbial sediments, NE Kattegat. Aquaict Microbial Ecology 22: 291-300
Sundback K. and A. Miles. (2002). Role of microphytobenthos and denitrification for nutrient turnover in embayments with floating macroalgal mats: a spring situation. Aquat Mar Ecol. 30: 91-101
Sundback K., A. Miles and E. Goransson. (2000). Nihogen fluxes, denitrification and the role of microphytobenthos in microtidal shallow-water sediments: an annual study. Marine Ecology Progress Series 200: 59-76
Sundback K., A. Miles, A. Hulth, L. Pihl, P. Engstrom, E. Selander and A. Svenson. (2003). Importance of benthic nuhient regeneration during initiation of macroalgal blooms in shallow bays. Marine Ecology Progress Series 246: 11 5-126
Suzumura M., T. Miyajima, H. Hata, Y. Umezawa, H. Kayanne and I. Koike. (2002). Cycling of phosphorus maintains the production of microphytobenthic communities in carbonate sediments of a coral reef. Limnology and Oceanography 47: 77 1-781
Terrados J., C.M. Duarte, L. Kamp-Nielsen, N.S.R. Agawin, E. Gacia, D. Lancap, M.D. Fortes, J. Borum, M. Lubanski and T. Greve. (1999). Are seagrass growth and survival constrained by reducing conditions of the sediment? Aquatic Botany 65: 175-107
Thomton D.C.O., L.F. Dong, G.J.C. Underwood and D.B. Nedwell. (2002). Factors affecting microphytobenthic biomass, species composition and production in the Colne Estuary (UK). Aquatic Microbial Ecology 27: 285-300
Thomton D.C.O., G.J.C. Underwood and D.B. Nedwell. (1999). Effect of illumination and emersion period on the exchange of ammonium across the estuarine sediment-water interface. Marine Ecology Progress Series 28: 1 1-20
Thybo-Christesen M., M.B. Rasmussen and T.H. Blackburn. (1993). Nutrient fluxes and growth of Cladophora sericea in a shallow Danish bay. Marine Ecology Progress Series 100: 273-8 1

260 K.J. McClathery, K. Sundback and I.C. Anderson
Trimmer M., D.B. Nedwell, D.B. Sivyer and S.J. Malcolm. (2000). Seasonal benthic organic matter mineralisation measured by oxygen uptake and denitrification along a transect of the inner and outer River Thames estuary, UK. Marine Ecology Progress Series 197: 103- 1 19
Turpin D.H.. (1991). Effects of inorganic N availability on algal photosynthesis and carbon metabolism. Journal of Phycology 27: 14-20
Tyler A.C., K.J. McGlathery and I.C. Anderson. (2001). Macroalgal mediation of dissolved organic nitrogen fluxes in a temperate coastal lagoon. Estuarine Coastal andSheljScience 53: 155-168
Tyler A.C., K.J. McGlathery and 1.C. Anderson. (2003). Benthic algae control sediment-water column fluxes of nitrogen in a temperate lagoon. Limnology and Oceanography In press.
Underwood G.J.C. and J. Kromkamp. (1999). Primary -production by phytoplankton and microphytobenthos in estuaries. Advances in Ecological Research 29: 93-153
Underwood G.J.C. and D.M. Paterson. (1993). Recovery of intertidal benthic diatoms after,biocide treatment and associated sediment dynamics. Journal of the Marine Biology Association of the United Kingdom 73: 25-45
Valiela I., K. Foreman, M. LaMontagne, D. Hersh, J. Costa, P. Peckol, B. DeMeo-Andreson, C. D'Avanzo, M. Babione, S. Sham, J. Brawley and K. Lajtha. (1992). Couplings of watersheds and coastal waters: sources and consequences of nutrient enrichment in Waquoit Bay, Massachusetts. Estuaries 15: 443-457
Valiela I., J. McClelland, J. Hauxwell, P.J. Behr, D. Hcrsh and K. Foreman. (1997). Macroalgal blooms in shallow estuaries: Controls and ecophysiological and ecosystem consequences. Limnology and Oceanography 42: 1 105- 1 1 18
Velimerov B. (1986). DOC dynamics in a Mediterranean seagrass system. Marine Ecology Progress Series28: 21-41.
van Kahvijk M.M., L.H.T. Vergeer, G.H.W. Schrnitz and J.G.M. Roelofs. (1997). Ammonium toxicity in eelgrass Zostera marina. Marine Ecology Progress Series 157: 159- 173
Viaroli P, et al. (1992). Ulva rigida growth and decomposition processes and related effects on nitrogen and phosphorus cycles in a coastal lagoon (Sacca di Goro, Po River Delta), In: G. Colombo, et al. (ed.), Marine eutrophication and population dynamics, 77-84. Olsen and Olsen, Fredensborg.
Viaroli P., M. Bartoli, C. Bondavalli and R.R. Christian. (1996). Macrophyte communities and their impact on benthic fluxes of oxygen, sulphide and nutrients in shallow eutrophic environments. Hvdrobiologia. 329: 105-1 19
Ward B.B. and D.A. Bronk. (2001). Net nitrogen u ~ t a k e and DON rclease in surface waters: imvortance . , ., . of trophic interactions implied from size fractionation experiments. Marine Ecology Progress Series 219:l 1-24
Welsh D.T., S. Bourgues, R. de Wit and R.A. Herbert. (1996). Seasonal variations in nitrogen-fixation (acetylene reduction) and sulphate-reduction rates in the rhizosphere of Zosrera noltii: nitrogen fixation by sulfate-reducing bacteria. Marine Riology 125: 619-628
Welsh D.T. (2000). Nitrogen fixation in seagrass meadows: Regulation, plant-bacteria interactions and significance to primary productivity. Ecology Letters 3:58-71
Welsh D.T., M. Bartoli, D. Nizzoli, G. Castaldelli, S.A. Riou and P. Viaroli. (2000). Denitrification, nitrogen fixation, community productivity and inorganic-N and oxygen fluxes in an intertidal Zostera noltii meadow. Marine Ecology Progress Series 208: 65-77.

Benthic N and P cycling 26 1
Wolfstein K., J.F.C. de Brouwer and L.J. Stal. (2002). Biochemical partitioning of photosynthetically fixed carbon by benthhic diatoms during short-term incubations at different irradiances. Marine Ecology Progress Series 245:2 1-3 1.
Wulff A,, K. Sundback, C. Nilsson, L. Carlson and B. Jonsson. (1997). Effect of sediment load on the microbenthic community of a shallow-water sandy sediment. Esluaries 20: 547-558
Ziegler S. and R. Benner. (1999). Dissolved organic carbon cycling in a subtropical seagrass-dominated lagoon. Marine Ecology Progress Series 1 80: 149- 1 60
9. AFFILIATIONS
K.J. McGlathery: Department of Environmental Sciences, P.O. Box 400123, University of Virginia, Charlottesville, VI 22904, USA. K. Sundback: Gothenburg University, Department of Marine Ecology, P.O. Box 46 1, S-40530 Gothenburg, Sweden. I.C. Anderson: School of Marine Science, Virginia Institute of Marine Science, College of William and Mary, Gloucester Point, VI 23602, USA.