the importance of fibroblasts in remodelling of the human uterine
TRANSCRIPT

The importance of fibroblasts in remodelling of the humanuterine cervix during pregnancy and parturition
Erik Malmstrom1, Maria Sennstrom2, Anna Holmberg1, Helena Frielingsdorf1, Erik Eklund1,Lars Malmstrom3, Ellen Tufvesson1, Maria F.Gomez1, Gunilla Westergren-Thorsson1,Gunvor Ekman-Ordeberg2 and Anders Malmstrom1,4
1Department of Experimental Medical Science, BMC, Lund University, Lund, Sweden, 2Department of Obstetrics and Gynecology,
Karolinska Institute, Stockholm, Sweden and 3Department of Biochemistry, University of Washington, Seattle, USA
4To whom correspondence should be addressed at: Department of Experimental Medical Science, C13, BMC, Lund University, S-221 84
Lund, Sweden. Tel: þ 46 46 2228574; Fax: þ46 46 2223128; E-mail: [email protected]
It is well established that fibroblasts play a crucial role in pathophysiological extracellular matrix remodelling. The aim of this
project is to elucidate their role in normal physiological remodelling. Specifically, the remodelling of the human cervix during
pregnancy, resulting in an enabled passage of the child, is used as the model system. Fibroblast cultures were established from
cervices of non-pregnant women, women after 36 weeks of pregnancy and women directly after partus. The cells were immunos-
tained and quantified by western blots for differentiation markers. The cultures were screened for cytokine and metalloproteinase
production and characterized by global proteome analysis. The cell cultures established from partal donors differ significantly
from those from non-pregnant donors, which is in accordance with in vivo findings. A decrease in a-smooth actin and prolyl-4-
hydroxylase and an increase in interleukin (IL)-6, IL-8 and matrix metalloproteinases (MMP)-1 and MMP-3 were observed in
cultures from partal donors. 2D-gel electrophoresis followed by mass spectrometry showed that the expression of 59 proteins
was changed significantly in cultures of partal donors. The regulated proteins are involved in protein kinase C signalling, Ca21
binding, cytoskeletal organization, angiogenesis and degradation. Our data suggest that remodelling of the human cervix is orche-
strated by fibroblasts, which are activated or recruited by the inflammatory processes occurring during the ripening cascade.
Key words: Fibroblast/human uterine cervix/interleukins/proteome/remodelling
Introduction
Remodelling of the extracellular matrix (ECM) is a prominent feature
during development and normal physiological processes such as
wound healing, pregnancy and growth (Westergren-Thorsson et al.,
1998; Sennstrom et al., 2000; Quan et al., 2004). An intensive remo-
delling, resembling that of post-inflammatory processes, also occurs
in asthma (Westergren-Thorsson et al., 2002), rheumatic diseases
(Westergren-Thorsson et al., 1996; Eklund et al., 2002) and tumour
metastasis (Mousa, 2002). In all these cases, highly active fibroblasts
produce structural ECM-components, cytokines and ECM-degrading
enzymes. In several of these normal remodelling processes, as well
as in diseased states, recruitment of blood-borne fibroblast precursors
(fibrocytes) occurs (Quan et al., 2004).
Fibroblasts in the remodelling process are regulated by cytokines,
both in autocrine and paracrine fashions. The cytokines are produced
by activated fibroblast, macrophages, neutrophils and lymphocytes.
Examples of regulatory cytokines in these processes are transforming
growth factor-b (TGF-b), epidermal growth factor, platelet-derived
growth factor and various interleukins (ILs), where TGF-b is known
as the main enhancer of ECM production with effects on both syn-
thesis and the final structure of the products (Westergren-Thorsson
et al., 1992). The effects of cytokines on fibroblasts, however, differ
considerably depending on tissue and localization within the tissue
(Westergren-Thorsson et al., 1990). Furthermore, several fibroblast
clones, with distinctly different properties, are present in normal,
persisting tissues and tissues undergoing remodelling (Westergren-
Thorsson et al., 2004).
Extensive tissue remodelling occurs during pregnancy. The uterus
undergoes an anabolic process where both smooth muscle and connec-
tive tissue increases (Hjelm et al., 2001a, b).The cervix, on the other
hand, which is mainly composed of fibrous connective tissue, is remo-
delled in a two-step process. During the first 36 weeks, a hormone-
driven decrease in collagen and proteoglycans (Uldbjerg et al.,
1983a) dominates. At the onset of labour, the cervix needs to be a
soft and elastic organ and crucial for the passage of the foetus. This
is obtained by a breakdown and reconstitution of the ECM, achieved
via recruitment of neutrophils (Stygar et al., 2002), an increase in
matrix metalloproteinases (MMPs) (Sennstrom et al., 2003) and
changed ECM production (Westergren-Thorsson et al., 1998).
# The Author 2007. Published by Oxford University Press on behalf of the European Society of Human Reproduction and Embryology. All rights reserved.
For Permissions, please email: [email protected]
The online version of this article has been published under an open access model. Users are entitled to use, reproduce, disseminate, or display the open access version of
this article for non-commercial purposes provided that: the original authorship is properly and fully attributed; the Journal and Oxford University Press are attributed as
the original place of publication with the correct citation details given; if an article is subsequently reproduced or disseminated not in its entirety but only in part or as a
derivative work this must be clearly indicated. For commercial re-use, please contact [email protected] 333
Molecular Human Reproduction Vol.13, No.5 pp. 333–341, 2007
Advance Access publication March 2, 2007 doi:10.1093/molehr/gal117
Downloaded from https://academic.oup.com/molehr/article-abstract/13/5/333/1119687by gueston 23 March 2018

During this final ripening step, cytokines such as IL-6, IL-8 and
G-CSF increase at least a 100-fold (Sennstrom et al., 2000), which
has major effects on ECM production. IL-8 promotes recruitment
and activation of neutrophils, which in turn stimulate IL-6 and IL-8
production from fibroblasts (King et al., 2001). The activated neutro-
phils secrete proteolytic enzymes, such as MMP-8 and leukocyte elas-
tase (Sennstrom et al., 2003), necessary for the final breakdown of the
collagen network. After partus, a rapid restoration of cervix to an
‘unripe state’ occurs (Westergren-Thorsson et al., 1998).
Different cells are involved in this remodelling process, but several
studies indicate that fibroblasts are key players in the final stage by
production of ILs, MMPs and ECM components such as the versican
proteoglycans (Sennstrom et al., 1997; Westergren-Thorsson et al.,
1998; Sennstrom et al., 2000; Sennstrom et al., 2003). In this study,
we established fibroblast cultures from cervices of women that were
non-pregnant, 36-week pregnant or partal to test whether the hypoth-
esis that fibroblasts having different properties are involved in the
cervical remodelling process is valid. Fibroblasts from 36-week preg-
nant or partal women differed markedly form their non-pregnant
counterparts in terms of immunohistological markers, cytokine and
MMP production and also had a dramatically changed proteome.
We conclude that the specific fibroblasts are activated or recruited
during the ripening process and are necessary to fulfil the final cervical
remodelling occurring only hours prior to birth.
Materials and methods
Chemicals
Monoclonal antibodies against prolyl-4-hydroxylase (#M0877) and a-smooth
actin (a-SMA) (#M0851) and the alkaline-phosphatase-conjugated secondary
rabbit anti-mouse antibody (#D0314) were purchased from DakoCytomation,
Denmark. The monoclonal antibodies against IL-8 (NAP-1) came from
Miroslav Ceska, Vienna, Austria, and biotinylated goat anti-mouse immuno-
globulin G (IgG) was purchased from Caltag Laboratory, San Francisco,
USA. The secondary horse anti-mouse (IgG), the avidin–biotin detection
system Vectastein ABC-elite and 3,30-diaminobenzidine (DAB) were pur-
chased from Vector Laboratories, Burlingame, USA. The goat anti-mouse sec-
ondary antibody, Alexa FluorTM 594, was purchased from Molecular Probes,
The Netherlands. 3-Amino-9-ethyl-l-carbazole (AEC) came from Sigma
Chemicals Co, St Louis, USA. The Immulitew Immunoassay system was
from Diagnostic Products Corporation, Los Angeles, USA. The sodium
dodecyl sulphate–polyacrylawide gel electrophoresis (SDS–PAGE) was per-
formed on NuPAGE 4–12% Bis Tris gradient gels (#NP0326BOX) from Invi-
trogen Life Technologies, USA. Cell culture materials, including antibiotics,
amino acids and cell culture medium, were purchased from Gibco, Sweden.
All other chemicals were of analytical grade.
Tissue collection
Cervical biopsies (50–300 mg) were obtained transvaginally from the distal
part of the human cervix from pregnant and non-pregnant women. Biopsies
were taken at elective caesarean sections from term (.36þ 6 weeks)-pregnant
women not in labour and with unripe cervices (n ¼ 5) and from women at spon-
taneous vaginal partus (n ¼ 5). Non-pregnant cervical tissue samples obtained
from fertile women, undergoing hysterectomy due to benign disorders such as
myomas or menorrhagia, served as controls (n ¼ 5). The Ethics Committee at
the Karolinska Hospital, Solna, Sweden, approved the study, and all biopsies
were obtained with informed consent.
Cell culture
The biopsies were washed in Earle’s minimal essential medium (EMEM) and
cut into�1-mm3 pieces. These were spread on the bottom of growth flasks and
allowed to adhere, and EMEM with penicillin (1%), streptomycin (1%), gluta-
mine (1%) and calf serum (10%) was added. Cells were grown until the isolated
fibroblast populations were confluent and transferred to new culture flasks three
to four times prior to the experiments. Mycoplasma infections were excluded
using 4,6-diamidine-2-phenyl-dihydrochloride. The cell cultures were studied
between passages 4 and 8.
Immunohistochemistry
For an a-SMA detection, biopsies were fixed for 12–24 h in 4% paraformalde-
hyde, kept in ethanol (70%) until embedded in paraffin and then sectioned at
5 mm. After de-paraffination with xylene and rehydration with ethanol, a mono-
clonal mouse anti-human a-SMA antibody (dilution 1 : 200) was added to the
sections. For detection of prolyl-4-hydroxylase, samples were cryosectioned in
8-mm sections and fixed in acetone for 3 min and acetone:methanol (1 : 1)
before adding a monoclonal mouse anti-human prolyl-4-hydroxylase antibody
(dilution 1 : 50). Horse anti-mouse IgG (dilution 1 : 200) was used as secondary
antibody, both for a-SMA and prolyl-4-hydroxylase. For detection of IL-8
expression, biopsies were treated as described previously (Sennstrom et al.,
2000). In short, the biopsies were sectioned and fixed in 2% paraformaldehyde.
Saponin was used to permeabilize the cell membranes. The sections were
incubated over night at room temperature in a humidified chamber with the
monoclonal antibodies for IL-8 (NAP-1). A biotinylated goat anti-mouse IgG
antibody was used as secondary antibody. Endogenous peroxidase activity
was blocked with 0.5% hydrogen peroxidase in methanol. Non-specific
binding was blocked with 10% horse serum. After staining with the primary
and secondary antibodies the slides were incubated with an avidin–biotin–
peroxidase complex and developed with either AEC for red staining (prolyl-4-
hydroxylase) or DAB for brown staining (a-SMA and IL-8). The slides were
washed in water, counter stained with 10% Mayer’s haematoxylin for 3 min
and then mounted. Two investigators evaluated the staining independently
using a Zeiss Axioskop. Photos were taken with a digital colour video
camera (Sony, Exwave HAD). Negative controls were treated in an identical
manner, except that the primary antibody was omitted. Staining specificity
was tested by incubation with recombinant produced proteins. Placenta
(IL-8) and uterine tissue were used as positive controls.
Non-pregnant, 36-weeks pregnant and partal cervical fibroblasts were
immunostained for prolyl-4-hydroxylase and a-SMA. The cells were fixed
with acetone for 5 min and permeabilized with 0.5% Triton X-100 in phosphate
buffered saline (PBS) for 5 min and unspecific binding was blocked with a 1%
bovine serum albumin solution in PBS for 2 h. The cells were then incubated
with primary monoclonal mouse anti-human a-SMA antibodies (1 : 100 in
blocking buffer), or monoclonal mouse anti-human prolyl-4-hydroxylase anti-
bodies (1 : 100 in blocking buffer), for 1 h at room temperature. The Alexa
Fluor 594 goat anti-mouse antibody was used for detection of the bound
primary antibodies. Between each step, the cells were carefully rinsed with
PBS. A light-scanning microscope (Multiprobe 2001, TM CLSM, Molecular
Dynamics) was used to examine the cells.
Western-blot analysis
Confluent or sub-confluent cultures were washed with ice-cold PBS, scraped in
a homogenization buffer (10% glycerol, 50 mM Tris, 100 mM NaCl, 1%
NP-40, 2 mM MgCl2, 2 mM sodium orthovanadate, 1 mg ml21 PMSF, aproti-
nin and leupeptin) and centrifuged at 48C for 5 min at 1500g. The protein con-
tents were determined in aliquots from the supernatants using Bradford protein
assay provided by Biorad. Equal amounts of protein were separated using
SDS–PAGE. The samples were blotted to PVDF-P membranes and processed
as described (Larsen et al., 2006), using primary antibodies directed towards
human a-SMA and prolyl-4-hydroxylase. The intensity of the bands on the
membrane were analysed using the Gel-ProTM Analyser software (Media
Cybernetics, Silver Spring, MD, USA).
Cytokines in fibroblast cultures
Levels of ILs-6 and -8 were analysed in medium using a solid-phase, two-site
chemiluminescent enzyme immunometric assay with the IMMULITE Auto-
mated Analyzer, as described earlier (Berthier et al., 1999). For IL-8, the
intra-assay variation was calculated to 3.6–3.8%, inter-assay variation 5.2–
7.4% and sensitivity 2 pg ml21. There was no cross-reactivity with IL-1b,
IL-2, IL2R, IL-4, IL-6, interferons or TNF-a and no interference with EDTA
or heparin. For IL-6, the intra-assay variation was 3.0–8.4%, inter-assay vari-
ation 6.2–9.7% and sensitivity 1 pg ml21. There was no cross-reactivity with
IL-2, IL-2R, IL-8, IL-1b and TNF-a and no interference with EDTA.
E.Malmstrom et al.
334
Downloaded from https://academic.oup.com/molehr/article-abstract/13/5/333/1119687by gueston 23 March 2018

MMP production in fibroblasts cultures
The MMPs were analysed in medium from fibroblasts by immunoassay 24 h
after fresh medium was added. For MMP-1, this assay determines total
MMP-1 (i.e. free MMP-1 and MMP-1 in complex with tissue inhibitor of
metalloproteinase-1 (TIMP-1), but not MMP-1 bound to a-2-macroglobulin).
According to the manufacturer, there is no cross-reactivity with
a-2-macroglobulin. The MMP-3 assay measures total MMP-3, including
proMMP-3, active MMP-3 and MMP-3/TIMP complexes, but not MMP-3
bound to a-2-macroglobulin. There is no cross-reactivity with MMP-1,
MMP-2 or MMP-9 or TIMP-1 or TIMP-2, according to the manufacture.
The detection limit was 6.25 ng ml21 for MMP-1 and 3.75 ng ml21 for
MMP-3. Standard MMP-1 and MMP-3 were purchased from Amersham,
Bucks, UK. The assay was performed according to the instructions of the man-
ufacturers, and all samples were assayed in duplicates. The results are
expressed as percent of non-pregnant control and the median levels in the
groups are expressed as nanogram per millilitre medium.
Expression profiles of abundant proteins by 2-Dgel electrophoresis
The cervical fibroblasts were cultured in six-well plates (6 � 8.3 cm2) and were,
at confluence, rinsed twice with ice-cold PBS and scraped into 200 ml of 7 M
urea, 2 M thiourea and 4% CHAPS as described previously (Malmstrom
et al., 2002a). The protein extracts were precipitated with four volumes of
ice-cold acetone and re-dissolved in scraping buffer. About 150 ml of the cell
extract was mixed with 200 ml of solubilization solution containing 7 M
thiourea, 2 M urea, 4% CHAPS, 10 mM DTT and 0.5% IPG 4–7 buffer and
added to Immobiline Dry strips (180 mm, pH 4–7 NL) (Amersham Pharmacia
Biotech, Uppsala, Sweden) for overnight dehydration. The isoelectric focusing
step was performed at 208C in a MultiphorTM (Amersham Pharmacia Biotech)
and run according to the following schedule: 0–300 V for 1 min, 300–3500 V
for 1.5 h and finally 3500 V for 23 h, until 83 kV h21 was reached. The strips
were reduced for 10 min in 65 mM DTT, 6 M urea, 30% (w/v) glycerol, 2%
(w/v) SDS and 50 mM Tris–HCl, pH 8.8, and alkylated for 10 min in the
same solution except for DTT, which was replaced with 0.259 M iodoaceta-
mide. The strips were soaked in electrophoresis running buffer (24 mM Tris
base, 0.2 M glycine and 0.1% SDS) just before the 2D-gel electrophoresis and
the strips were applied on 14% homogeneous Duracryl slab gels. The strips were
overlaid with a solution of 1% agarose in electrophoresis buffer (kept at 608C)
and electrophoresis was carried out in a HoeferTM DALT gel apparatus (Amer-
sham Pharmacia Biotech) at 208C with constant voltage (100 V) for 18 h. The
gels were stained with a silver solution according to a protocol described by
Shevchenko et al. 1996) and scanned using a Gs-710 Calibrated Imaging Den-
sitometer (Bio-Rad Laboratories, Sundbyberg, Sweden). Spot analysis was per-
formed using the PDQUEST (version 7.0) 2D-gel analysis system (Bio-Rad
discovery series, Bio-Rad Laboratories). Spots with a significant Wilcoxon
P-value were validated by visual inspection using a gel-slicing feature of the
2DDB software. Every spot on all the gels were assigned an integrated
optical density (IOD) value by the software program. This value was divided
by the total IOD for all detected spots. Thus, each spot is expressed as parts
per million of the total IOD of all valid spots. The gels were run in triplicate,
thus each experiment comprised nine gels (n ¼ 3, i.e. 27 gels). Match sets con-
taining 9 gels and 27 gels were made and analysed separately as a control of the
analysis schema. After matching, the data were exported to an SQL relational
database, where the individual experiments were further matched by x,y
co-ordinates and statistical analysis was performed. Spots with a P-value
,0.05 were sliced out and transferred to small Eppendorf tubes, followed by
washing (40% acetonitrile, 60 mM ammonium bicarbonate, pH 7.8). The gel
plugs were dried down in a Speed Vac for 15 min (Savant Speed Vac, DNA
110) and digested with trypsin (Promega Porcine, acetic acid) in 25 mM
ammonium bicarbonate and incubated overnight at 378C. The digestions were
stopped by adding 0.2% trifluoroacetic acid.
Mass-spectrometry identification
Ziptips (Millipore, Bedford, MA, USA) were used to concentrate and desalt the
protein digests according to the manufacturer’s instructions. After washing, the
peptides were eluted on an Anchor-chipTM plate (Bruker Daltonics, Bremen,
Germany) and mixed with 0.5 ml of 2.5–dihydroxybenzoic acid matrix
(2.8 mg ml21). The matrix-assisted laser desorption ionization time of flight
(MALDI-TOF) instrument used was a Bruker REFLEX (Bruker-Daltonics)
operated in a reflector mode at an accelerating voltage of 20 kV. The data
were analysed using both the Swissprot and NCBI database using Profound
(http://prowl.rockefeller.edu/profound_bin/WebProFound.exe).
Statistical methods
Spot data and gel images were exported from PDQUEST and imported into an
in-house-developed analysis software (Malmstrom et al., 2002b). In short, the
software integrates PDQUEST data with bioinformatical and statistical tools.
PDQUEST-matched spots from the same treatment were compared with
PDQUEST-matched spots from the two other treatments using Wilcoxon
signed-rank test. Spots with a quality score below 25 and quantity score
below 100 were omitted. Because of the relative measure (IOD), the spot inten-
sities were normalized to non-pregnant cultures. Identified proteins were, when
possible, assigned GO terms, either from the proteins themselves or from close
orthologues. For IL-6, IL-8, MMP-1 and MMP-3 analyses, the Mann–Whitney
U and Kruskal–Wallis tests were used to estimate P-values.
Results
Expression of fibroblast markers in cervical tissue andfibroblast cultures
The human cervix undergoes an extensive remodelling during preg-
nancy and at parturition. During the final stages of pregnancy, a weak
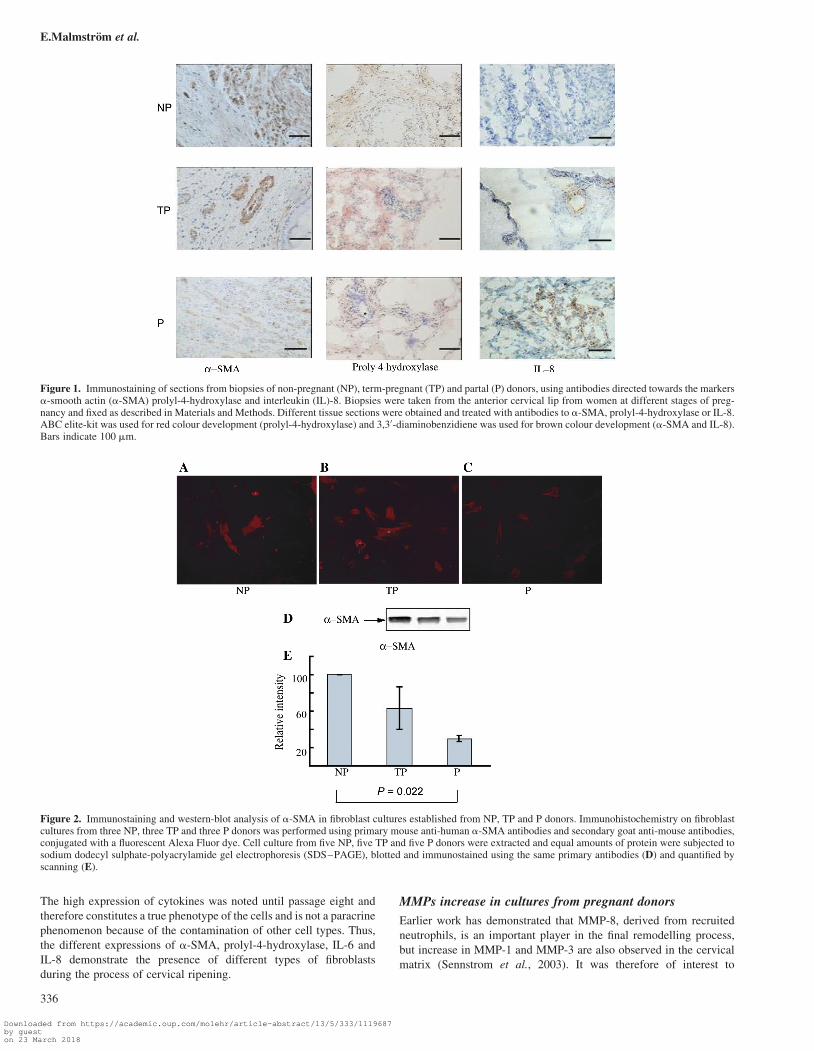
decrease in prolyl-4-hydroxylase staining is observed, which is in agree-
ment with the substantial decrease of collagen concentration observed at
term and in partal cervices (Figure 1). Since it is seen in tissues from
both 36-week-pregnant and partal women, changed collagen turnover
is a feature of the slower, more long-lasting phase of the cervical remo-
delling. Staining for a-SMA, a marker for myofibroblast differentiation,
is, however, not clearly decreased until the very last phase (Figure 1).
This may indicate that contractile elements in the fibroblasts of cervix
are needed to retain the fetus in the uterus until the delivery process
is initiated. To investigate whether these changes in expression patterns
also occur in fibroblast cultures from the different stages of pregnancy,
we established several cultures from each state. The expressions of
a-SMA (Figure 2A–D) and prolyl-4-hydroxylase (Figure 3A–D)
were then studied by western-blot analysis and immunohistochemistry.
The data were in accordance with the tissue sections, with an �70 %
decrease in the prolyl-4-hydroxylase expression in fibroblasts from
both 36-week-pregnant and partal women, whereas a-SMA expression
was only significantly decreased in partal fibroblast (40% of control).
Expression of inflammatory markers in cervical tissueand fibroblasts cultures
In vivo, the amounts of IL-6 and IL-8 (Sennstrom et al., 2000) increase
100–1000-fold during the late phase of pregnancy. It was therefore of
interest to examine the production of these cytokines in corresponding
cervical tissues and fibroblast cultures. Immunohistochemistry
revealed an elevated expression of IL-8 term-pregnant material,
obtained at 36 weeks of pregnancy, and an even stronger expression
in partal tissue (Figure 1). This staining is found close to both fibro-
blasts and neutrophils, especially in partal tissue, but their site of syn-
thesis, is unclear. To further collect information of sites of synthesis,
we studied whether the fibroblasts in cultures established from non-
pregnant, term pregnant and partal tissues produce IL-6 and IL-8,
using the IMMUNOLITE methodology. IL-6 showed a significant
increase in partal samples. In comparison with non-pregnant cultures,
a mean 2.5-fold increase in cultures from term-pregnant patients was
seen, however not statistically significant. In partal cultures, a signifi-
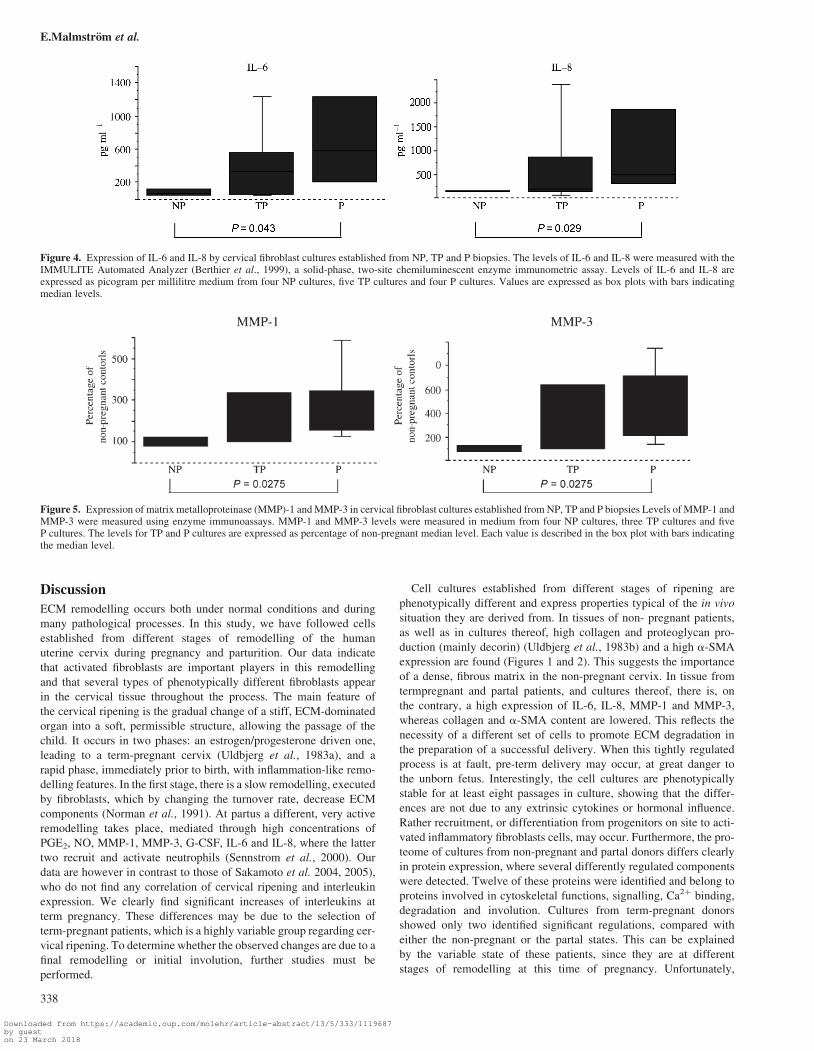
cant 3.5-fold increase was recorded (Figure 4, P ¼ 0.0433). The same
pattern was noted for IL-8, where term-pregnant donors increased on
an average 3.5-fold compared with controls, whereas those from partal
donors increased significantly 5–6-fold (Figure 4, P ¼ 0.0209).
Role of fibroblast in remodelling
335
Downloaded from https://academic.oup.com/molehr/article-abstract/13/5/333/1119687by gueston 23 March 2018
The high expression of cytokines was noted until passage eight and
therefore constitutes a true phenotype of the cells and is not a paracrine
phenomenon because of the contamination of other cell types. Thus,
the different expressions of a-SMA, prolyl-4-hydroxylase, IL-6 and
IL-8 demonstrate the presence of different types of fibroblasts
during the process of cervical ripening.
MMPs increase in cultures from pregnant donors
Earlier work has demonstrated that MMP-8, derived from recruited
neutrophils, is an important player in the final remodelling process,
but increase in MMP-1 and MMP-3 are also observed in the cervical
matrix (Sennstrom et al., 2003). It was therefore of interest to
Figure 2. Immunostaining and western-blot analysis of a-SMA in fibroblast cultures established from NP, TP and P donors. Immunohistochemistry on fibroblastcultures from three NP, three TP and three P donors was performed using primary mouse anti-human a-SMA antibodies and secondary goat anti-mouse antibodies,conjugated with a fluorescent Alexa Fluor dye. Cell culture from five NP, five TP and five P donors were extracted and equal amounts of protein were subjected tosodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS–PAGE), blotted and immunostained using the same primary antibodies (D) and quantified byscanning (E).
Figure 1. Immunostaining of sections from biopsies of non-pregnant (NP), term-pregnant (TP) and partal (P) donors, using antibodies directed towards the markersa-smooth actin (a-SMA) prolyl-4-hydroxylase and interleukin (IL)-8. Biopsies were taken from the anterior cervical lip from women at different stages of preg-nancy and fixed as described in Materials and Methods. Different tissue sections were obtained and treated with antibodies to a-SMA, prolyl-4-hydroxylase or IL-8.ABC elite-kit was used for red colour development (prolyl-4-hydroxylase) and 3,30-diaminobenzidiene was used for brown colour development (a-SMA and IL-8).Bars indicate 100 mm.
E.Malmstrom et al.
336
Downloaded from https://academic.oup.com/molehr/article-abstract/13/5/333/1119687by gueston 23 March 2018

investigate whether the cervical cells secrete these MMPs. Indeed,
cultures from non-pregnant donors did secrete an appreciable
amount of both MMP-1 and MMP-3 (Figure 5). However, cultures
from term-pregnant donors showed 40–70% higher mean secretion
and cultures from partal donors showed an even higher secretion,
with a 2–3-fold increase in MMP-1 and MMP-3 secretion, respect-
ively (Figure 5, P ¼ 0.0275). This feature was also stable in eight
passages, further implying that it is a true endogenous phenotype of
the cells, not exogenously regulated.
Proteome analysis
2D-gel electrophoresis, analysed by PDQUEST and MALDI-TOF,
was used to study the protein expression pattern in the cell cultures
established from the non-pregnant, 36-week-pregnant and partal sub-
jects. A typical gel, with the pI range from 4 to 7, is shown in Figure 6.
Using cell cultures from a total of nine patients and performing 2D-gel
sets in triplicates enabled us to establish a hierarchical gel set with, in
total, 519 matched spots. Forty-four of these were assigned an identity.
The IOD of the spots were normalized against the non-pregnant group,
and a total of 22 significantly regulated components were detected in
partal cultures. In cultures from 36-week-pregnant women, 16 signifi-
cantly regulated spots were found, compared with cultures from
non-pregnant donors. Four regulations were noted when cultures
from term-pregnant and partal donors were compared. Of the regu-
lated components, 12 were identified, and of these, 11 were
up-regulated up to 4–5-fold. Only one protein was down-regulated
by�50% in partal cultures (Table I). The identified proteins were sep-
arated into groups reflecting the function of the proteins (Table I).
Vimentin, which is involved in cytoskeletal organization and related
signalling and is regarded as a marker for fibroblasts, is up-regulated
2–3-fold (Figure 6). Of note is the increased expression of proteins of
the 14-3-3 family, where especially 14-3-3 protein theta influencing
Protein kinase C (PKC)-activity is significantly up-regulated around
5-fold in both term and partal cultures (Figure 6). Also Ca2þ-binding
proteins such as annexin V are clearly induced (Table I). On the other
hand, calreticulin, which is a chaperon taking part in glycoprotein
quality control/folding, is strongly repressed (Table I). The protease
cathepsin B is also up-regulated, which probably is of importance
for final ECM degradation (Figure 6).
Associated with the cervix involution is the increased expression of
angiogenin inhibitor, which is only induced in the partal cultures
(Figure 6). This indicates that these proteins play a role in the final
remodelling, where a decrease in the vascular bed and formation of
ECM are hallmarks (Table I).
Figure 3. Immunostaining and western-blot analysis of prolyl-4-hydroxylase in fibroblast cultures established from NP, TP and P donors. Immunohistochemistry onfibroblast cultures from three NP, three TP and three P donors were performed using monoclonal mouse anti-human prolyl-4-hydroxylase antibodies. The secondaryantibodies were conjugated with a fluorescent Alexa Fluor dye. Cell culture from five NP, five TP and five P donors were extracted and equal amounts of protein weresubjected to SDS–PAGE, blotted and immunostained using the same antibodies (D) and quantified (E).
Role of fibroblast in remodelling
337
Downloaded from https://academic.oup.com/molehr/article-abstract/13/5/333/1119687by gueston 23 March 2018
Discussion
ECM remodelling occurs both under normal conditions and during
many pathological processes. In this study, we have followed cells
established from different stages of remodelling of the human
uterine cervix during pregnancy and parturition. Our data indicate
that activated fibroblasts are important players in this remodelling
and that several types of phenotypically different fibroblasts appear
in the cervical tissue throughout the process. The main feature of
the cervical ripening is the gradual change of a stiff, ECM-dominated
organ into a soft, permissible structure, allowing the passage of the
child. It occurs in two phases: an estrogen/progesterone driven one,
leading to a term-pregnant cervix (Uldbjerg et al., 1983a), and a
rapid phase, immediately prior to birth, with inflammation-like remo-
delling features. In the first stage, there is a slow remodelling, executed
by fibroblasts, which by changing the turnover rate, decrease ECM
components (Norman et al., 1991). At partus a different, very active
remodelling takes place, mediated through high concentrations of
PGE2, NO, MMP-1, MMP-3, G-CSF, IL-6 and IL-8, where the latter
two recruit and activate neutrophils (Sennstrom et al., 2000). Our
data are however in contrast to those of Sakamoto et al. 2004, 2005),
who do not find any correlation of cervical ripening and interleukin
expression. We clearly find significant increases of interleukins at
term pregnancy. These differences may be due to the selection of
term-pregnant patients, which is a highly variable group regarding cer-
vical ripening. To determine whether the observed changes are due to a
final remodelling or initial involution, further studies must be
performed.
Cell cultures established from different stages of ripening are
phenotypically different and express properties typical of the in vivo
situation they are derived from. In tissues of non- pregnant patients,
as well as in cultures thereof, high collagen and proteoglycan pro-
duction (mainly decorin) (Uldbjerg et al., 1983b) and a high a-SMA
expression are found (Figures 1 and 2). This suggests the importance
of a dense, fibrous matrix in the non-pregnant cervix. In tissue from
termpregnant and partal patients, and cultures thereof, there is, on
the contrary, a high expression of IL-6, IL-8, MMP-1 and MMP-3,
whereas collagen and a-SMA content are lowered. This reflects the
necessity of a different set of cells to promote ECM degradation in
the preparation of a successful delivery. When this tightly regulated
process is at fault, pre-term delivery may occur, at great danger to
the unborn fetus. Interestingly, the cell cultures are phenotypically
stable for at least eight passages in culture, showing that the differ-
ences are not due to any extrinsic cytokines or hormonal influence.
Rather recruitment, or differentiation from progenitors on site to acti-
vated inflammatory fibroblasts cells, may occur. Furthermore, the pro-
teome of cultures from non-pregnant and partal donors differs clearly
in protein expression, where several differently regulated components
were detected. Twelve of these proteins were identified and belong to
proteins involved in cytoskeletal functions, signalling, Ca2þ binding,
degradation and involution. Cultures from term-pregnant donors
showed only two identified significant regulations, compared with
either the non-pregnant or the partal states. This can be explained
by the variable state of these patients, since they are at different
stages of remodelling at this time of pregnancy. Unfortunately,
Figure 4. Expression of IL-6 and IL-8 by cervical fibroblast cultures established from NP, TP and P biopsies. The levels of IL-6 and IL-8 were measured with theIMMULITE Automated Analyzer (Berthier et al., 1999), a solid-phase, two-site chemiluminescent enzyme immunometric assay. Levels of IL-6 and IL-8 areexpressed as picogram per millilitre medium from four NP cultures, five TP cultures and four P cultures. Values are expressed as box plots with bars indicatingmedian levels.
Figure 5. Expression of matrix metalloproteinase (MMP)-1 and MMP-3 in cervical fibroblast cultures established from NP, TP and P biopsies Levels of MMP-1 andMMP-3 were measured using enzyme immunoassays. MMP-1 and MMP-3 levels were measured in medium from four NP cultures, three TP cultures and fiveP cultures. The levels for TP and P cultures are expressed as percentage of non-pregnant median level. Each value is described in the box plot with bars indicatingthe median level.
E.Malmstrom et al.
338
Downloaded from https://academic.oup.com/molehr/article-abstract/13/5/333/1119687by gueston 23 March 2018

proteomic analyses using 2D gels have shortcomings concerning low-
expressed proteins as well as extensively modified proteins. Thus,
using this system, we cannot detect the low-expressed ILs and
matrix components as collagens and proteoglycans.
Of great interest is the 5-fold up-regulation of a group of PKC acti-
vator theta proteins (Table I). An increase in these activators could
potentially activate several signal transduction pathways, together
with Ca2þ and/or diacylglycerol. This results in modification of
activity of various groups of PKCs, which contain at least 11 sub-
groups. The downstream signalling involves mitogen-activated
protein kinase (MAPK) (Ueda et al., 1996) and further c-fos and
c-Jun, activating AP-1 sites in promoter sequences. Possible down-
stream targets of this pathway are the genes of MMP-1 and MMP-3,
which are increased 2–3-fold in partal cells. The regulation of
MMP-1 and MMP-3 expressions has earlier been shown to indepen-
dently rely on both the PKC and the p44/42 MAPK pathways
(Reuben et al., 2002). Furthermore, the activation of PKC induces col-
lagenase activity in the cervix of pregnant guinea pigs (Rajabi et al.,
1992), implying its importance in the signal transduction occurring
during cervical ripening.
Up-regulation of IL-8 and IL-6, .100-fold in the human cervix
in vivo, has also been recorded during cervical ripening (Sennstrom
et al., 2000). The results from the present study show that the same
phenomenon occurs in fibroblast cultures, however, more pronounced
in partal cultures compared with term-pregnant cultures. One possible
pathway is induction of IL-8 by increased Ca2þ flux (Marino et al.,
2003), followed by activation of PKC and NF-kB (Cummings
et al., 2004; Page et al., 2003). Induced changes of c-fos and c-Jun
influence the AP-1 site, which is found in the IL-8 promoter region
(Cummings et al., 2004). The up-regulation of activators of PKC
suggests that it is involved in signalling leading to IL-8 production,
both in vivo and in the term-pregnant and partal cell cultures.
Also, IL-6 production is stimulated by increased Ca2þ (Jeong et al.,
2002, Giugliano et al., 2003) and ERK and PKC activation (Graness
et al., 2002). Thus, the up-regulation of PKC activators and a
changed Ca2þ-sensitivity could be one mechanism for IL-6 secretion
in partal cells.
During involution after partus, a remodelling back to the non-
pregnant state occurs rapidly (Westergren-Thorsson et al., 1998) and
is featured by a significant increase in collagen and proteoglycan
production. It is therefore likely that constitutive fibroblasts are
re-activated to produce matrix reconstructing the cervical tissue.
The fate of the inflammatory fibroblasts after partus is uncertain, but
one possibility is that they undergo apoptosis. Furthermore, a hallmark
of ripening is a highly increased level of vessels that disappear after
parturition. Of interest is therefore the observation that the partal fibro-
blasts produce a 2-fold higher amount of the angiogenin inhibitor
(Table I), preparing the tissue for dilatation and involution.
Figure 6. 2D-gel electrophoresis of the proteome of cervical fibroblasts established from a partal biopsy and quantification of relative expression in cultures estab-lished from NP, TP and P donors. Fibroblast cultures from NP, TP and P donors were solubilized and subjected to 2D-gel electrophoresis. After silver staining, thegel was evaluated by PDQUEST. Spots of interest were selected and treated with trypsin. Mass spectrometry was performed using a Bruker Daltonics MALDI-TOFinstrument. The spots surrounded by red rings are identified and SPP numbers given. Identities and relative expressions are given in Table I.
Role of fibroblast in remodelling
339
Downloaded from https://academic.oup.com/molehr/article-abstract/13/5/333/1119687by gueston 23 March 2018

Two major conclusions can be drawn from these data. The first is
the fact that fibroblasts are important cells in tissue remodelling and
are involved both in tissue degradation/remodelling and in attraction
of neutrophils. The second, that to accomplish this remodelling, fibro-
blasts with different properties must be engaged. An important ques-
tion is how these fibroblasts, having inflammatory characteristics,
are recruited or activated and how they maintain their activity
in vitro. During remodelling of other tissues, for instance, during
cancer-induced remodelling, progenitor fibroblasts (fibrocytes) are
recruited from the circulation and can differentiate into inflammatory
fibroblasts (Abe et al., 2001; Barth et al., 2002). It is also well docu-
mented that recruitment and differentiation into active fibroblasts
occur in the lungs during fibrosis associated with asthma, scleroderma
and interstitial lung fibrosis (Schmidt et al., 2003; Larsen et al., 2004;
Postlethwaite et al., 2004). The inflammatory fibroblasts found in the
uterine cervix do, however, not generate fibrosis as observed in asthma
and lung fibrosis. Another possibility is recruitment of mesodermal
progenitors (Pittenger et al., 1999). The results from this study do
not provide evidence that recruitment of such cells occurs, but the pre-
sence of TGF-b in partal cervix (Westergren-Thorsson et al., 1998)
suggests that a mesenchymal stem-cell differentiation may be
involved (Roelen and Dijke, 2003). TGF-b however also promotes
myofibroblast differentiation, which is not the case in this study,
where the expression of a-SMA was decreased in partal tissues and
cultures.
Cervical remodelling shows similarities to an inflammation-driven
remodelling with the presence of ‘inflammatory’ fibroblast and, in
the final stage, activated neutrophils. This recruitment of neutrophils
is most likely due to the production of ILs-6 and -8, both known attrac-
tants of neutrophils (Sennstrom et al., 2000). The key question is what
factors induce the very large production of these ILs. Using immuno-
histochemistry, we have previously shown that IL-8 production
initially occurs in the cervical epithelium and that it temporally corre-
lates to fetal fibronectin production from the fetus at term (Ekman
et al., 1995). This indicates that other factors, e.g. from fetal tissues,
may be involved in recruitment and activation of fibroblasts.
In conclusion, we have shown that fibroblasts are important in cer-
vical tissue remodelling and that different, phenotypically stable, cell
clones are found. The activation pathways involved could be Ca2þ-
and PKC-dependent and result in a considerable increase in IL-6,
IL-8, MMP-1 and MMP-3. This is of absolute necessity for a
normal softening of the cervical tissue, allowing the dilation and
expulsion of the fetus at parturition.
Acknowledgements
Jonas Hamfors is acknowledged for initial experiments. This work was sup-ported by grants from the Swedish Medical Research Council (11550, 7970),Heart-Lung Foundation, Djuskyddsmyndigheten, Greta and John Kock,Alfred Osterlund, Crafoord Foundations, Gustaf V:s 80 Arsfond, Research-school in Pharmaceutical Science (FLAK) and the Medical Faculty, Lund Uni-versity. Funding to pay the Open Access publication changes for this articlewas provided by . . .
References
Abe R, Donnelly SC, Peng T, Bucala R and Metz CN (2001) Peripheral bloodfibrocytes: differentiation pathway and migration to wound sites. J Immunol166,7556–7562.
Barth PJ, Ramaswamy A and Moll R (2002) CD34(þ) fibrocytes in normalcervical stroma, cervical intraepithelial neoplasia III, and invasivesquamous cell carcinoma of the cervix uteri. Virchows Arch 441,564–568.
Berthier F, Lambert C, Genin C and Bienvenu J (1999) Evaluation of anautomated immunoassay method for cytokine measurement using theImmulite Immunoassay system. Clin Chem Lab Med 37,593–599.
Cummings R, Zhao Y, Jacoby D, Spannhake EW, Ohba M, Garcia JG, WatkinsT, He D, Saatian B and Natarajan V (2004) Protein kinase Cdelta mediateslysophosphatidic acid-induced NF-kappaB activation and interleukin-8secretion in human bronchial epithelial cells. J Biol Chem 279,41085–41094.
Eklund E, Broberg K, Westergren-Thorsson G, Bjardahlen A, Hedlund Mand Malmstrom A (2002) Proteoglycan production in disomic and trisomy7-carrying human synovial cells. Matrix Biol 21,325–335.
Ekman G, Granstrom L, Malmstrom A, Sennstrom M and Svensson J (1995)Cervical fetal fibronectin correlates to cervical ripening. Acta ObstetGynecol Scand 74,698–701.
Giugliano G, Pasquali D, Notaro A, Brongo S, Nicoletti G, D’Andrea F,Bellastella A and Sinisi AA (2003) Verapamil inhibits interleukin-6 and
Table I. Regulated proteins, identified in cultures from non-pregnant (NP), term-pregnant (TP) and partal (P) donors
Protein ID Accesionnumbera
SSPnumberb
MeanNP
MeanTP
SD MeanP
SD P-valueNP–TP
P-valueTP–P
P-valueNP–P
CytoskeletonVimentin P08670 1603 1 1.84 0.7 3.04 0.884 0.286 0.087 0.0052Calpain, small subunit 1 P04632 2206 1 1.42 0.45 1.78 0.075 0.489 0.5457 0.0625
Signalling14-3-3 protein theta P27348 1302 1 5.55 4.75 4.74 0.614 0.004 1 0.015214-3-3 protein zeta/delta P63104 1301 1 2.30 0.511 4.10 1.378 0.259 0.616 0.093914-3-3 proitein beta/alpha P31946 1307 1 3.02 2.7 1.46 1.388 0.094 0.029 0.1119
Calcium binding proteinsCalreticulin P277797 702 1 0.72 0.23 0.54 0.107 0.171 0.8571 0.0476Annexin V P08758 2409 1 1.91 0.734 2.94 0.503 0.236 0.236 0.014
DegradationCathepsin B AAA52125.1 4202 1 1.3 0.94 1.26 0.636 0.195 0.0206 0.0464
InvolutionRibonuclease/angiogenin inhibitor P13489 1703 1 1.57 0.544 1.94 0.467 0.229 0.4 0.028
Proteins associated with increased cellactivity
Nicotinamide N-methyltransferase P40261 5201 1 1.74 0.94 2.09 0.421 0.423 0.1388 0.0078Hypothetical protein XP 057199 1602 1 2.17 0.67 3.26 0.501 0.286 0.2398 0.0056Glutatione Transferase P1-1 Chain B 1EOH_B 5204 1 1.541 0.594 1.664 0.863 0.034 0.888 0.0019Laminin-binding protein CAA3469.1 1509 1 2.62 2.51 1.98 0.221 0.546 0.1615 0.0078Nuclear chloride channel protein AAC25675.1 4301 1 0.92 0.35 1.48 0.218 0.546 0.136 0.0315
aThe Swiss-Prot accession numbers are selected whenever possible.bSSP number is a spot identifier generated by PDQuest.
E.Malmstrom et al.
340
Downloaded from https://academic.oup.com/molehr/article-abstract/13/5/333/1119687by gueston 23 March 2018

vascular endothelial growth factor production in primary cultures of keloidfibroblasts. Br J Plast Surg 56,804–809.
Graness A, Chwieralski CE, Reinhold D, Thim L and Hoffmann W (2002)Protein kinase C and ERK activation are required forTFF-peptide-stimulated bronchial epithelial cell migration and tumornecrosis factor-alpha-induced interleukin-6 (IL-6) and IL-8 secretion.J Biol Chem 277,18440–18446.
Hjelm A, Ekman-Ordeberg G, Barchan K and Malmstrom A (2001a) Changesof the uterine proteoglycan distribution at term pregnancy and during labour.Eur J Obstet Gynecol Reprod 100,146–151.
Hjelm A, Ekman-Ordeberg G, Barchan K and Malmstrom A (2001b)Identification of the major proteoglycans from human myometrium. ActaObstet Gynecol Scand 80,1084–1090.
Jeong HJ, Hong SH, Lee DJ, Park JH, Kim KS and Kim HM (2002) Role ofCa(2þ) on TNF-alpha and IL-6 secretion from RBL-2H3 mast cells. CellSignal 14,633–639.
King AE, Kelly RW, Critchley HO, Malmstrom A, Sennstrom M and PhippsRP (2001) Cd40 expression in uterine tissues: a key regulator of cytokineexpression by fibroblasts. J Clin Endocrinol Metab 86,405–412.
Larsen K, Tufvesson E, Malmstrom J, Morgelin M, Wildt M, Andersson A,Lindstrom A, Malmstrom A, Lofdahl CG and Marko-Varga Get al. (2004)Presence of activated mobile fibroblasts in bronchoalveolar lavage frompatients with mild asthma. Am J Respir Crit Care Med 170,1049–1056.
Larsen K, Malmstrom J, Wildt M, Dahlqvist C, Hansson L, Marko-Varga G,Bjermer L, Scheja A and Westergren-Thorsson G (2006) Functional andphenotypical comparison of myofibroblasts derived from biopsies andbronchoalveolar lavage in mild asthma and scleroderma. Respir Res 7,11.
Malmstrom J, Larsen K, Hansson L, Lofdahl CG, Norregard-Jensen O,Marko-Varga G and Westergren-Thorsson G (2002a) Proteoglycanand proteome profiling of central human pulmonary fibrotic tissue utilizingminiaturized sample preparation: a feasibility study. Proteomics 2,394–404.
Malmstrom L, Malmstrom J, Marko-Varga G and Westergren-Thorsson G(2002b) Proteomic 2DE database for spot selection, automated annotation,and data analysis. J Proteome Res 1,135–138.
Marino F, Cosentino M, Fietta AM, Ferrari M, Cattaneo S, Frigo G, Lecchini Sand Frigo GM (2003) Interleukin-8 production induced by the endozepinetriakontatetraneuropeptide in human neutrophils: role of calcium andpharmacological investigation of signal transduction pathways. Cell Signal15,511–517.
Mousa SA (2002) Anticoagulants in thrombosis and cancer: the missing link.Expert Rev Anticancer Ther 2,227–233.
Norman M, Ekman G, Ulmsten U, Barchan K and Malmstrom A (1991)Proteoglycan metabolism in the connective tissue of pregnant and non-pregnant human cervix. An in vitro study Biochem J 275(Pt 2),515–520.
Page K, Li J, Zhou L, Iasvovskaia S, Corbit KC, Soh JW, Weinstein IB, BrasierAR, Lin A and Hershenson MB (2003) Regulation of airway epithelial cellNF-kappa B-dependent gene expression by protein kinase C delta.J Immunol 170,5681–5689.
Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD,Moorman MA, Simonetti DW, Craig S and Marshak DR (1999)Multilineage potential of adult human mesenchymal stem cells. Science284,143–147.
Postlethwaite AE, Shigemitsu H and Kanangat S (2004) Cellular origins offibroblasts: possible implications for organ fibrosis in systemic sclerosis.Curr Opin Rheumatol 16,733–738.
Quan TE, Cowper S, Wu SP, Bockenstedt LK and Bucala R (2004) Circulatingfibrocytes: collagen-secreting cells of the peripheral blood. Int J BiochemCell Biol 36,598–606.
Rajabi MR, Solomon S and Poole AR (1992) Activation of protein kinase Cstimulates collagenase production by cultured cells of the cervix of thepregnant guinea pig. Am J Obstet Gynecol 167,194–200.
Reuben PM, Brogley MA, Sun Y and Cheung HS (2002) Molecular mechanismof the induction of metalloproteinases 1 and 3 in human fibroblasts by basiccalcium phosphate crystals. Role of calcium-dependent protein kinase Calpha. J Biol Chem 277,15190–15198.
Roelen BA and Dijke P (2003) Controlling mesenchymal stem celldifferentiation by TGFBeta family members. J Orthop Sci 8,740–748.
Sakamoto Y, Moran P, Searle RF, Bulmer JN and Robson SC (2004)Interleukin-8 is involved in cervical dilatation but not in prelabour cervicalripening. Clin Exp Immunol 138,151–157.
Sakamoto Y, Moran P, Bulmer JN, Searle RF and Robson SC (2005)Macrophages and not granulocytes are involved in cervical ripening. JReprod Immunol 66,161–173.
Schmidt M, Sun G, Stacey MA, Mori L and Mattoli S (2003) Identification ofcirculating fibrocytes as precursors of bronchial myofibroblasts in asthma. JImmunol 171,380–389.
Sennstrom MK, Brauner A, Lu Y, Granstrom LM, Malmstrom AL and EkmanGE (1997) Interleukin-8 is a mediator of the final cervical ripening inhumans. Eur J Obstet Gynecol Reprod Biol 74,89–92.
Sennstrom MB, Ekman G, Westergren-Thorsson G, Malmstrom A, Bystrom B,Endresen U, Mlambo N, Norman M, Stabi B and Brauner A (2000) Humancervical ripening, an inflammatory process mediated by cytokines. Mol HumReprod 6,375–381.
Sennstrom MB, Brauner A, Bystrom B, Malmstrom A and Ekman G (2003)Matrix metalloproteinase-8 correlates with the cervical ripening process inhumans. Acta Obstet Gynecol Scand 82,904–911.
Shevchenko A, Wilm M, Vorm O and Mann M (1996) Mass spectrometricsequencing of proteins silver-stained polyacrylamide gels. Anal Chem68,850–858.
Stygar D, Wang H, Vladic YS, Ekman G, Eriksson H and Sahlin L (2002)Increased level of matrix metalloproteinases 2 and 9 in the ripeningprocess of the human cervix. Biol Reprod 67,889–894.
Ueda Y, Hirai S, Osada S, Suzuki A, Mizuno K and Ohno S (1996) Proteinkinase C activates the MEK-ERK pathway in a manner independent ofRas and dependent on Raf. J Biol Chem 271,23512–23519.
Uldbjerg N, Ekman G, Malmstrom A, Olsson K and Ulmsten U (1983a)Ripening of the human uterine cervix related to changes in collagen,glycosaminoglycans, and collagenolytic activity. Am J Obstet Gynecol147,662–666.
Uldbjerg N, Malmstrom A, Ekman G, Sheehan J, Ulmsten U and Wingerup L(1983b) Isolation and characterization of dermatan sulphate proteoglycanfrom human uterine cervix. Biochem J 209,497–503.
Westergren-Thorsson G, Sarnstrand B, Fransson LA and Malmstrom A (1990)TGF-beta enhances the production of hyaluronan in human lung but not inskin fibroblasts. Exp Cell Res 186,192–195.
Westergren-Thorsson G, Schmidtchen A, Sarnstrand B, Fransson LAand Malmstrom A (1992) Transforming growth factor-beta inducesselective increase of proteoglycan production and changes in thecopolymeric structure of dermatan sulphate in human skin fibroblasts. EurJ Biochem 205,277–286.
Westergren-Thorsson G, Coster L, Akesson A and Wollheim FA (1996)Altered dermatan sulfate proteoglycan synthesis in fibroblast culturesestablished from skin of patients with systemic sclerosis. J Rheumatol23,1398–1406.
Westergren-Thorsson G, Norman M, Bjornsson S, Endresen U, Stjernholm Y,Ekman G and Malmstrom A (1998) Differential expressions of mRNA forproteoglycans, collagens and transforming growth factor-beta in the humancervix during pregnancy and involution. Biochim Biophys Acta 1406,203–213.
Westergren-Thorsson G, Chakir J, Lafreniere-Allard MJ, Boulet LPand Tremblay GM (2002) Correlation between airway responsiveness andproteoglycan production by bronchial fibroblasts from normal andasthmatic subjects. Int J Biochem Cell Biol 34,1256–1267.
Westergren-Thorsson G, Sime P, Jordana M, Gauldie J, Sarnstrand Band Malmstrom A (2004) Lung fibroblast clones from normal and fibroticsubjects differ in hyaluronan and decorin production and rate ofproliferation. Int J Biochem Cell Biol 36,1573–1584.
Submitted on October 7, 2006; resubmitted on December 19, 2006; accepted onDecember 29, 2006
Role of fibroblast in remodelling
341
Downloaded from https://academic.oup.com/molehr/article-abstract/13/5/333/1119687by gueston 23 March 2018