the function of msx1 gene in promoting meiosis of dairy goat male germline stem cells (mgscs)
TRANSCRIPT

RAPID COMMUNICATION
The function of Msx1 gene in promoting meiosis of dairy goat malegermline stem cells (mGSCs)
Hailong Mu1, Jiang Wu1, Haijing Zhu1, Na Li1, Furong Tang1, Xi Yao1, Churong Yang1, Sha Peng1,Guangpeng Li2* and Jinlian Hua1*1College of Veterinary Medicine, Shaanxi Centre of Stem Cells Engineering and Technology, Key Lab for Animal Biotechnology ofAgriculture Ministry of China, Northwest A&F University, Yangling, China2Key Laboratory for Mammalian Reproductive Biology and Biotechnology, Ministry of Education, Inner Mongolia University, Hohhot, China
During sequential stages of meiosis, numerous cytoplasmic and nuclear events take place in which many germline and non-germline genesinvolved. It is demonstrated that the germline gene Stra8 and synaptonemal complex protein 3 (Scp3) play an important role in the meiosis.Recently, studies showed Msx1, a DNA-binding protein taking part in the skeletal development, also having a functional attractive factor toStra8 and Scp3 in the meiosis. In this study, we cloned the gene Msx1 then transfected the Msx1 constructed recombination plasmid,pMsx1-Ires2-AcGFP, into the dairy goat germline stem cells (male germline stem cells) and analysed the effects of Msx1 on the expressionof Stra8 and Scp3. The results showed that Msx1 could enhance the expression of Stra8 and Scp3 and promote the meiosis in goat testicularcells. Bmp4 activated the expression ofMsx1 and Stra8. This study suggests that Msx1 plays an important role in spermatogenesis and meiosis.Copyright © 2013 John Wiley & Sons, Ltd.
key words—Msx1 Stra8 Scp3 Meiosis mGSCs Bmp4
INTRODUCTION
Msx1 belongs to homeobox family, which is very importantfor the embryonic development. In 1989, Hill first clonedthe Msx1 gene from the human genomic library, which islocated on the short (p) arm of chromosome 4 at position16.2.1 The MSX1 protein was required for the normal devel-opment of many organs or tissues such as limbs, teeth,neural crest and gonads.2,3 Msx1 can bind to multi-site inthe upstream promoter of dentin sialophosphoprotein (DSPP),dentin matrix acidic phosphoprotein I (DMPI), osteocalcin,cyclinDI and MyoDI and can up-regulate cyclinDI and candown-regulate osteocalcin.4 In vitro tissue culture, bonemorphogenetic protein 4 (Bmp4) and retinoic acid (RA) canpromote the expression of Msx1 in mice testis.3,5 During themeiosis initiation, the Msx1 gene is strongly expressed in fetalovaries, and the number of germ cells inmeiosis is significantly
reduced in the fetal mouse ovary with Msx1 and Msx2 nullmutants.5 On the basis of the existing studies, it is imperativethat further understanding on the function of Msx1 in malegerm cells’ meiosis.
The gene Stra8 (stimulated by RA 8) was first clonedfrom mice in 1996, and both the transcriptional and transla-tional levels of Stra8 were regulated by the RA.6 The fetalfemale mice knocked-out Stra8 lead to the DNA replicationarrested before the meiosis and prevent synapsis and recom-bination of the chromosome during the meiosis so that theovum can not be generated.7 It also has the same functionin the male juvenile mice.8 Meiosis is a cell division uniqueto germ cells. During meiosis, homologous chromosomesare paired and recombined, a process essential for generat-ing genetically distinct haploid cells. Male mice carrying anull-mutation in the synaptonemal complex protein 3(Scp3) gene are sterile with complete loss of spermato-cytes,9 and female mutant mice produce aneuploid oocytesthrough defective meiotic chromosome segregation, al-though they are fertile.10 These phenotypes indicate thatSYCP3 is required for the normal progression of meiosis.
The Msx genes are the downstream effectors of BMPsignalling in several developing structures.11–13 The BMPsignalling is both upstream and downstream of Msx genes,depending on the context and the developing structure.
*Correspondence to: Dr. Jinlian Hua, College of Veterinary Medicine,Shaanxi Centre of Stem Cells Engineering and Technology, Key Lab forAnimal Biotechnology of Agriculture Ministry of China, Northwest A&FUniversity, Yangling, Shaanxi, 712100 China. E-mail: [email protected]; Dr. Guangpeng Li, Key Laboratory for Mammalian ReproductiveBiology and Biotechnology, Ministry of Education, Inner Mongolia Univer-sity, Hohhot 010021, China. E-mail: [email protected]
Received 26 June 2013Revised 21 September 2013
Accepted 23 September 2013Copyright © 2013 John Wiley & Sons, Ltd.
cell biochemistry and functionCell Biochem Funct 2013; 31: 629–635.Published online 14 October 2013 in Wiley Online Library(wileyonlinelibrary.com) DOI: 10.1002/cbf.3010

The possibility that Bmp2 and Bmp4 participate in a Dlx–Msxsignalling loop between the limb bud apical ectodermal ridge(AER) and mesoderm is clearly in line with previously identi-fied roles of these molecules.14
In our study, we cloned the Msx1 gene, and analysed theover-expression of Msx1 in mGSCs and induced theexpression of Msx1 with Bmp4; the results suggested thatMsx1 gene involved in the spermatogenesis and meiosis.This study provided a foundation of study in goat spermato-genesis and meiosis.
MATERIALS AND METHODS
Construction of recombination plasmid
TheMsx1 primers were designed according to the reported BosTaurus b-Msx1 cDNA sequence (NM_174798). Msx1 is1091bp. Primers were synthesized by Peking SanBoYuanZhicompany (Beijing, China):
Msx1: Forward: 5’-CTAGCTAGCGGCTGACATGA-3’Reverse: 5’-ACAGGATCCCTGTTGCTTTCTGG-3’.
The gene was amplified from cattle testis using reverse tran-scription polymerase chain reaction (RT-PCR). Then thespecific fragments were cloned into pMD18-T cloning vector(TaKaRa, Dalian, China). After white-blue screening, thepositive clones were identified by PCR, restriction enzymedigestion and sequencing. Then the fragments were clonedinto pIRES2-AcGFP eukaryotic expression vector to obtainpMsx1-IRES2-AcGFP recombination plasmid. The plasmidwas verified by PCR and restriction enzyme digestion.15
Evolutionary relationship among the Msx1 coding sequence
The cattle Msx1 coding sequence (CDS) was obtained,sequenced by Peking SanBoYuanZhi company (Beijing,China). Then the sequence was aligned with the knownsequences in GenBank by DNAMAN software, and the phy-logenetic tree was depicted with MEGA4.1 (Center for Evo-lutionary Functional Genomics, Tempe, USA).15
Cell transfection male germline stem cells
The dairy goat mGSCs were cultured in our lab and previ-ous studies.16 The pMsx1-IRES2-AcGFP recombinationplasmid and pIRES2-AcGFP were transfected into mGSCsin a 48-well plate, respectively. The plasmid DNAwas dilutedto 200 ng in 100μl Opti-MEM (Invitrogen, Carlsbad, CA,USA) reduced serum medium. Mixed gently, then added0.5μl PLUS™ reagent (Invitrogen) directly to the dilutedDNA and incubated the mixedmedium for 5min at room tem-perature. Mixed Lipofectamine™ LTX reagent (Invitrogen)gently before using, then added 1 μl directly to the dilutedDNA. Mixed gently and incubated for 30min at room tem-perature. The 50 μl DNA-Lipofectamine™ LTX complexeswere added and incubated the cells for 4–6 h at 37 °C in aCO2 incubator. The green fluorescent protein (GFP) posi-tive cells were examined after 48 h, and the total RNAs
of the two groups were extracted and the germ cell andmeiosis markers – Stra8 and Scp3, Dazl, Nanos2 and Plzf– were analysed by RT-PCR and quantitative RT-PCR(QRT-PCR).17
RT-PCR and quantitative RT-PCR analysis
Total RNAs for RT-PCR analysis were extracted frommGSCs using TRIzol (Tiangen Biotech Co. Ltd, Beijing,China). The cDNA was synthesized on the basis of 500 ngRNA with a commercially available kit (TaKaRa, Biotech.Co. Ltd, Dalian, China). The PCR steps included denaturationat 94 °C for 5min, followed by 35 cycles at 95 °C for 30 s,55–58 °C for 30 s and 72 °C for 30–60 s and a final extensionat 72 °C for an additional 10min. The primers were designedon the basis of the sequences of the open reading frame fromthe National Center for Biotechnology Information (NCBI)GenBank and synthesized by AuGCT Biotechnology com-pany (Beijing, China). The PCR primers and the lengths ofthe amplicons are shown in Table 1. The PCR products wereanalysed in 2% agarose (Invitrogen, Carlsbad, CA, USA) gelelectrophoresis, stained with ethidium bromide (Invitrogen),and visualized under UV illumination.16,18
Quantitative RT-PCR was set up in 15μl reaction mix-tures containing ddH2O 6.3 μl, 2 × SYBR (Bioer Co. Ltd.,Hangzhou, China) 7.5μl, cDNA 0.5μl, sense primer0.3μl, antisense primer 0.3μl, TaqDNA polymerase 0.1μl.Reaction conditions were as follows: 94 °C for 5min andthen 40 cycles and 94 °C for 20 s, 58 °C for 30 s, and 70 °C for 10 s. All expression levels were normalized to β-actinin each well. The 2-[delta][delta]Ct method was used tomeasure the expression alteration.17 The fluorescence signalwas collected every 0.5 °C for 10 s. The primers are listed inTable 1.
Immunofluorescence staining
The testicular tissues were paraffined, deparaffinization andrehydration following the standard methods. Slides weredipped in three changes of xylene for 6min each, two changesof 100%, 95%, and 75% alcohol for 3min, respectively, after-wards rinsed twice in deionized water for 5min. The slideswere soaked in the boiling citrate buffer for 15–25min,followed by three washes in cold phosphate-buffered saline(PBS), each for 5min. Washed slides were blocked withblocking solution (PBST+1% bovine serum albumin(BSA)) for a minimum of 30min and incubated with primaryantibody to Msx1 (1:100, Bioss Biotechnology Inc., Beijing,
Table 1. The primer sequences
Gene Name Sense primer Antisense primer
β-actin acggcatcaccaact aggaaggaaggctggaagagMsx1 cactcggtgtcaaagtggagga gaaggggacactttgggcttStra8 cggccatctcccatctgtg tgtccttcacgctgccctcScp3 gtatggaggacttggaga gagactttcggacacttgcDazl caagttcaccagttcaggttatcac gacaacggagtttctcagtctattcNanos2 ggaaggactacttcaacctgagcc ataatgccgcagaatgggacacPlzf Caccgcaacagccagcactat cagcgtacagcaggtcatccag
630 H. MU ET AL.
Copyright © 2013 John Wiley & Sons, Ltd. Cell Biochem Funct 2013; 31: 629–635.
China) overnight at 4 °C. Tissues were washed in PBS forthree times and then incubated with secondary antibody(1:500, Chemicon International, Inc., Temecula, CA, USA)following the manufacturer’s manual. The nuclei of cells werestained by Hoechst 33342.
The mGSCs transfected by pMsx1-IRES2-AcGFP andpIRES2-AcGFP were washed two times with PBS, each for3min, and fixed in 4% paraformaldehyde for 15min at roomtemperature. Afterwards, rinsed twice in PBS for 3min,followed by 0.1% triton-100 for 10min, PBS washed twicefor 3min. Washed plates were blocked with blocking solution(PBST+1% BSA) for a minimum of 30min and incubatedwith primary antibodies to Msx1(1:100, Bioss BiotechnologyInc.), Stra8(1:200, Abcam), Scp3(1:200, Abcam) overnight at4°C; the plates were washed in PBS for three times and thenincubated with secondary antibody (1:500,Chemicon Interna-tional, Inc., Temecula, CA, USA.) following the manufacturer’s manual. The nuclei of cells were stained by Hoechst 33342.19
Western blot
Total cell extracts were prepared from mGSCs in thetrsnsfected Msx1 or not, and proteins were extracted in 1×sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) sample loading buffer. Total cell proteins wereresolved by SDS-PAGE, transferred to Polyvinylidenedifluoride membrane and probed with β-actin (1:1000,Beyotime, Haimen, Jiangsu, China), Stra8 (1:1000, Abcam),horse-radish peroxidase-conjugated anti-rabbit was used as asecondary antibody (1:1000, Beyotime). The detection wasperformed using the Thermo Scientific Pierce enhanced chemi-luminescence western blotting substrate (Thermo Scientific).The results were analysed by Tanon-410 automatically gelimaging system (Shanghai Tianneng Corporation, China).
Luciferase reporter assay
The pStra8-luciferase (LUC), the 1.4-kb fragment of Stra8promoter was inserted into SacI/HindIII sites of the pGL3-basic
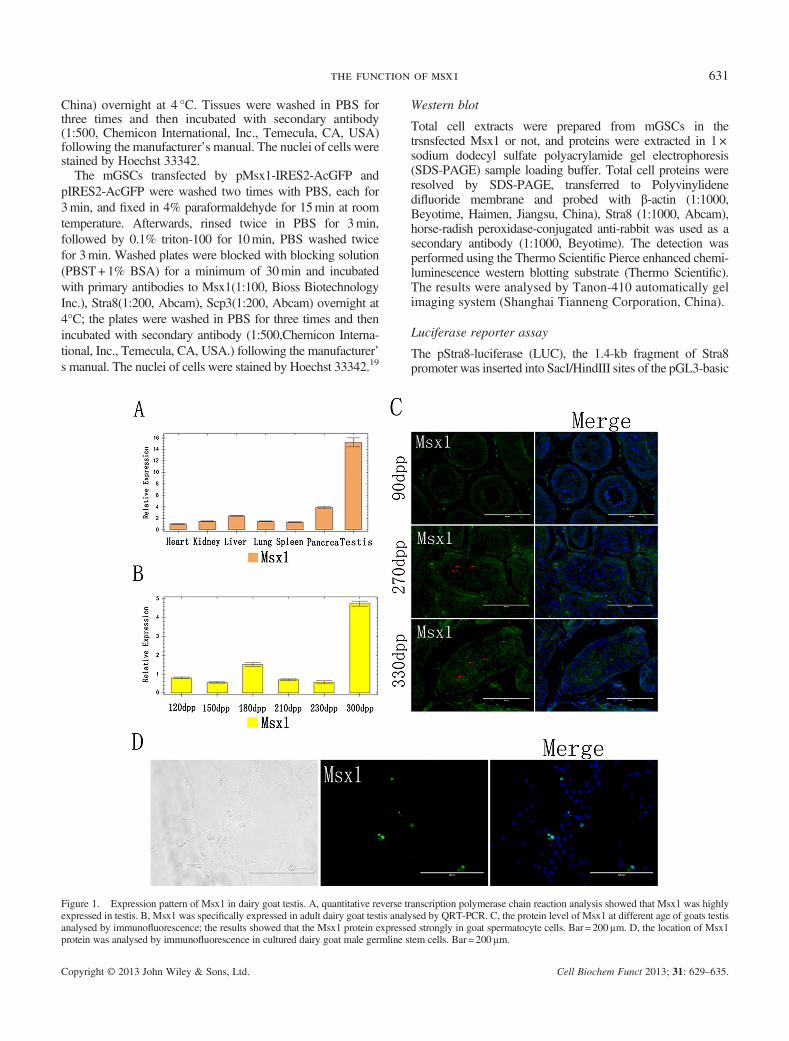
Figure 1. Expression pattern of Msx1 in dairy goat testis. A, quantitative reverse transcription polymerase chain reaction analysis showed that Msx1 was highlyexpressed in testis. B, Msx1 was specifically expressed in adult dairy goat testis analysed by QRT-PCR. C, the protein level of Msx1 at different age of goats testisanalysed by immunofluorescence; the results showed that the Msx1 protein expressed strongly in goat spermatocyte cells. Bar = 200μm. D, the location of Msx1protein was analysed by immunofluorescence in cultured dairy goat male germline stem cells. Bar= 200μm.
631the function of msx1
Copyright © 2013 John Wiley & Sons, Ltd. Cell Biochem Funct 2013; 31: 629–635.

vector (Clontech Laboratories, Inc., CA, USA).20 Fiftynanograms of pStra8-LUC, pMsx1-IRES2-AcGFP orpIRES2-AcGFP (negative control) was co-transfected intoHeLa cells in a 48-well plate using Lipofectamine2000(Invitrogen). After 48 h, the target validation assays wereperformed with LUC reporter system (Beyotime, Jiangsu,China) according to the manufacturer’s instructions.
RESULTS
Expression pattern of Msx1 in dairy goat testis
Quantitative RT-PCR analysis showed that Msx1 was highlyexpressed in testis (Figure 1A). The expression level of Msx1was significantly increased in adult dairy goat testis and hadan extremely lower expression level in juvenile testis analysedby QRT-PCR and immunofluorescence (Figure 1B and C).Immunofluorescence analysis showed that the Msx1 proteinwas expressed strongly in spermocyte and in the testis at270 and 330 dpp old goat. And the Msx1 was also expressedin the cultured round large spermatocytes-like cells(Figure 1D). The results manifested that theMsx1 was locatedin the cytoplasm of spermatocyte cells.
The construction of recombination plasmidpMsx1-Ires2-AcGFP
Msx1 was cloned by PCR from the adult cattle testes andwhich is 946 bp (Figure 2A and B). Furthermore, thesequence was submitted to National Center for Biotechnol-ogy Information and obtained the formal sequence number
(JX982767). Then we aligned the CDS of Msx1 betweendifferent species and found that Msx1 is a conserved gene(Figure 2C).
The effects of Msx1 on the expression of Stra8 and Scp3 inmGSCs
The Msx1 CDS was obtained, and the pMsx1-IRES2-AcGFPplasmid was constructed and verified by NheI, BamHI doubledigestion (Figure 3A). Then, pMsx1-IRES2-AcGFP andpIRES2-AcGFP were transfected into mGSCs. The resultsdemonstrated that pMsx1-IRES2-AcGFP was successfullytransfected into mGSCs (Figure 3B). The expression level ofMsx1 in pMsx1-IRES2-AcGFP transfected group was signif-icantly higher than that in the pIRES2-AcGFP group analysedby QRT-PCR (Figure 3C). Simultaneously, the expressionlevels of meiosis genes, Stra8, Scp3 and Dazl, were remark-ably up-regulated; however, the expression of self-renewalmarkers (Plzf and Nanos2) of SSCs was down-regulated inover-expression of Msx1 compared with control (Figure 3C).The protein level of Stra8 was up-regulated by Msx1over-expression analysed by western blot (Figure 3D). Theresults of immunofluorescence also manifested Stra8, andScp3 was up-regulated by Msx1 over-expression (Figure 3F).However, the results of the pStra8 promoter-LUC assayshowed that the Msx1 had no direct effect on Stra8 promoter.
The expression of Msx1 and Stra8 in male germline stemcells induced by Bmp4
There were more round or globate cells in the group inducedby Bmp4 (12.5 ng/ml) than control (Figure 4A). The
Figure 2. Identification of Msx1 gene. A, the Msx1 was cloned. B, the phylogenetic trees of Msx1 showed that Msx1 was conserved. C, the sequence ofMsx1.
632 H. MU ET AL.
Copyright © 2013 John Wiley & Sons, Ltd. Cell Biochem Funct 2013; 31: 629–635.

Figure 3. Over-expression of Msx1 in mGSCs. A, the pMsx1-IRES2-GFP was identified by restricted enzyme digestion. B, GFP positive malegermline stem cells (mGSCs) were observed with fluorescent microscope. Bar = 200 μm a, b, mGSCs were transfected with pIRES2-AcGFP; c, dmGSCs were transfected with pMsx1-IRES2-AcGFP. C, The expression level of Msx1 in pMsx1-IRES2-AcGFP transfected group was significantlyincreased compared with the pIRES2-AcGFP group analysed by QRT-PCR. Additionally, the expression of meiosis genes, Stra8 and Scp3 remarkably,were up-regulated in pMsx1-IRES2-AcGFP transfected group. D, The expression of Stra8 was increased analysed by western blot. E, The level ofStra8 had no significant difference in pMsx1-IRES2-AcGFP group compared with pIRES2-AcGFP analysed by pStra8 promoter-LUC assay.Control was the lysis buffer and D-Luciferin. F, The protein level of Stra8 and Scp3 was also significantly increased by overexpression Msx1 analysedby immunofluorescence. Bar = 200 μm.
633the function of msx1
Copyright © 2013 John Wiley & Sons, Ltd. Cell Biochem Funct 2013; 31: 629–635.

expression levels of Msx1 and Stra8 were significantly up-regulated by Bmp4; however, Scp3 was not changed clearly(Figure 4B, C and D).
DISCUSSION
Meiosis is a complex and critical process in spermatogene-sis. Recently, more and more genes and molecularmechanisms related to meiosis and spermatogenesis werefound. In these genes, Stra8 and Scp3 played critical rolesin meiosis. Stra8 is one key regulator to switch germ cellsfrom mitosis into meiosis in mammals.21 Stra8 is specifi-cally expressed in premeiotic germ cells.21,22 RA, as aninducer to initiate Stra8, then germ cells enter meiosis stagein mice.23,24
Synaptonemal complex protein 3 is an essential compo-nent of the synaptonemal complex, which is a proteinaceousstructure formed between paired homologous chromosomesduring prophase of the first meiotic division.25,26 Mice thatare deficient in SYCP3 fail to establish synapsis, resultingin meiotic arrest during spermatogenesis in males andchromosome segregation errors leading to aneuploidoocytes in females.27,28
Msx1 plays an important role in the embryonic develop-ment and also was necessary for the ovogenesis. Evidencesshowed that Msx1 and Msx2 genes are essential in murinefetal germ cell meiosis initiation. In vivo, the Msx1/Msx2double-null mutation prevented full activation of Stra8. InF9 cells, Msx1 can bind to Stra8 regulatory sequences, andMsx1 overexpression stimulates Stra8 transcription. Resultsdemonstrate that some homeobox genes are required formeiosis initiation in the female germ line.5
In this study, we for the first time identified theexpression of Msx1 in dairy goat testis at the different agesand tissues. The results suggested that Msx1 was closelyassociated with dairy goat spermatogenesis and meiosis.Furthermore, we transfected the recombination plasmidpMsx1-Ires2-AcGFP into mGSCs, analysed by QRT-PCRand immunofluorescence staining. The results demon-strated over-expressed Msx1 up-regulated the criticalfactors in spermatogenesis and meiosis: Dazl, Stra8 andScp3.8,29 Additionally, the markers of self-renewal, Nanos2and Plzf, were down-regulated by Msx1. These datasuggested that male spermatogenesis and meiosis were alsoregulated by Msx1. Furthermore, we used pstra8 promoter-LUC assay to confirm whether Msx1 directly activate Stra8;however, the results were not supported by this hypothesis.Thus, we assumed that Msx1 can activate the expression ofStra8 indirectly, but not directly.Bone morphogenetic protein 4 is expressed by Sertoli
cells in mice until postnatal day 7, whereas at later ages, itis predominantly expressed in spermatogonia and spermato-cytes.30 Our results showed that Bmp4 induced the expres-sion of Msx1 and Stra8, but there was no effect on Scp3.These results suggested that Bmp4 can enhance theexpression of Msx1 and Stra8 to regulate the meiosis. Incombination with the previous studies, we assumed thatBmp4 may regulated Stra8 by influencing the expressionof Msx1(Bmp4→Msx1→ Stra8).5,31 Further, we need toelucidate the real mechanisms on the interactions betweenthese genes and signal molecules.In conclusion, we clarified the expression pattern of Msx1
in dairy goat testis and found Msx1 is required for the meiosisand spermiogenesis in dairy goat testis. Over-expression ofMsx1 in mGSCs resulted in the elevation of meiosis-related
Figure 4. Bone morphogenetic protein 4 induced Msx1, Stra8 and Scp3 in male germline stem cells (mGSCs). A, the morphological characters of mGSCswere observed with microscope. Bar = 100μm a, mGSCs treated by Bmp4; b, control. B, the expression levels of Msx1 and Stra8 were increased in Bmp4treated group, but Scp3 was not up-regulated. C, the expression of Stra8 was increased and analysed by western blot. D, The protein level of Stra8 andMsx1 was also significantly increased and analysed by immunofluorescence. Bar = 200μm.
634 H. MU ET AL.
Copyright © 2013 John Wiley & Sons, Ltd. Cell Biochem Funct 2013; 31: 629–635.

genes, Stra8 and Scp3. Additionally,Msx1 and Stra8 were up-regulated by Bmp4. Taken together, Msx1 plays an importantrole in spermatogenesis andmeiosis, and this study provided amodel for further investigation of the mechanisms of Msx1 inspermatogenesis and meiosis.
CONFLICT OF INTEREST
The authors have declared that there is no conflict of interest.
ACKNOWLEDGEMENTS
This work was supported by the grants from the Program(31272518) of National Natural Science Foundation of China,the National Basic Research Program(2013CB947900),Doctoral Fund of Ministry of Education of China (RFDP,20120204110030), Key Laboratory of National the InnerMongolia Autonomous Region Mammalian ReproductiveBiology and Biotechnology of Ministry (Inner MongoliaUniversity) Open Major Basic Research Project (20130902),and the Fundamental Research Funds for the Central Univer-sities (QN2011012).
REFERENCES
1. Hill R, Jones P, Rees A, et al. A new family of mouse homeo box-containing genes: molecular structure, chromosomal location, anddevelopmental expression of Hox-7.1. Genes Dev 1989; 3: 26–37.
2. Lallemand Y, Nicola M-A, Ramos C, et al. Analysis of Msx1; Msx2double mutants reveals multiple roles for Msx genes in limbdevelopment. Development 2005; 132: 3003–3014.
3. Ramos C, Robert B. msh/Msx gene family in neural development.Trends Genet 2005; 21: 624–632.
4. Fux C, Langer D, Fussenegger M. Dual-regulated myoD-and msx1-basedinterventions in C2C12-derived cells enable precise myogenic/osteogenic/adipogenic lineage control. J Gene Med 2004; 6: 1159–1169.
5. Le Bouffant R, Souquet B, Duval N, et al. Msx1 and Msx2 promotemeiosis initiation. Development 2011; 138: 5393–5402.
6. Oulad-Abdelghani M, Bouillet P, Décimo D, et al. Characterization ofa premeiotic germ cell-specific cytoplasmic protein encoded by Stra8,a novel retinoic acid-responsive gene. J Cell Biol 1996; 135: 469–477.
7. Baltus AE, Menke DB, Hu Y-C, et al. In germ cells of mouseembryonic ovaries, the decision to enter meiosis precedes premeioticDNA replication. Nat Genet 2006; 38: 1430–1434.
8. Anderson EL, Baltus AE, Roepers-Gajadien HL, et al. Stra8 and its in-ducer, retinoic acid, regulate meiotic initiation in both spermatogenesisand oogenesis in mice. Proc Natl Acad Sci 2008; 105: 14976–14980.
9. Yuan L, Liu J-G, Zhao J, et al. The Murine SCP3 gene is required forsynaptonemal complex assembly, chromosome synapsis, and male fer-tility. Mol Cell 2000; 5: 73–83.
10. Yuan L, Liu J-G, Hoja M-R, et al. Female germ cell aneuploidy andembryo death in mice lacking the meiosis-specific protein SCP3.Science 2002; 296: 1115–1118.
11. Vainio S, Karavanova I, Jowett A. Identification of BMP-4 as a signalmediating secondary induction between epithelial and mesenchymaltissues during early tooth development. Cell 1993; 75: 45–58.
12. Bei M, Maas R. FGFs and BMP4 induce both Msx1-independent andMsx1-dependent signaling pathways in early tooth development.Development 1998; 125: 4325–4333.
13. Hollnagel A, Oehlmann V, Heymer J, et al. Id genes are direct targetsof bone morphogenetic protein induction in embryonic stem cells. JBiol Chem 1999; 274: 19838–19845.
14. Vieux-Rochas M, Bouhali K, Mantero S, et al. BMP-mediatedfunctional cooperation between Dlx5; Dlx6 and Msx1; Msx2 duringmammalian limb development. PLoS One 2013; 8: e51700.
15. Li M, Liu C, Zhu H, et al. Expression pattern of Boule in dairy goattestis and its function in promoting the meiosis in male germline stemcells (mGSCs). J Cell Biochem 2013; 114: 294–302.
16. Hua J, Zhu H, Pan S, et al. Pluripotent male germline stem cells fromgoat fetal testis and their survival in mouse testis. Cell Reprogram2011; 13: 133–144.
17. Hu Y, Sun J, Wang J, et al. Characterization of female germ-like cellsderived from mouse embryonic stem cells through expression of GFPunder the control of Figla promoter. J Cell Biochem 2012; 113:1111–1121.
18. Cao H, Chu Y, Zhu H, et al. Characterization of immortalizedmesenchymal stem cells derived from foetal porcine pancreas. CellProlif 2011; 44: 19–32.
19. Liu C, Zhu H-J, Liu W-S, et al. Male germ cells specification ofembryonic gonad from guanzhong dairy goat. Chinese J Vet Sci2011; 9: 018.
20. Jia W, Cheng D, Chen S, et al. Retinoic acid induces myoblaststransdifferentiation into premeiotic stra8-positive cells. Cell Biol Int2011; 35: 356–372.
21. Kee K, Angeles VT, Flores M, et al. Human DAZL, DAZ and BOULEgenes modulate primordial germ-cell and haploid gamete formation.Nature 2009; 462: 222–225.
22. Luetjens CM, Xu EY, Pera RAR, et al. Association of meiotic arrestwith lack of BOULE protein expression in infertile men. J ClinEndocrinol Metab 2004; 89: 1926–1933.
23. Koubova J, Menke DB, Zhou Q, et al. Retinoic acid regulates sex-specific timing of meiotic initiation in mice. Proc Natl Acad Sci U SA 2006; 103: 2474–2479.
24. Maines JZ, Wasserman SA. Post-transcriptional regulation of themeiotic Cdc25 protein twine by the Dazl orthologue Boule. Nat CellBiol 1999; 1: 171–174.
25. Rooij DG, Russell LD. All you wanted to know about spermatogoniabut were afraid to ask. J Androl 2000; 21: 776–798.
26. de Rooij DG. Proliferation and differentiation of spermatogonial stemcells. Reproduction 2001; 121: 347–354.
27. Russell LD, Ettlin RA, Hikim APS, et al. Histological and histopatho-logical evaluation of the testis. Int J Androl 1993; 16: 83–83.
28. Bowles J, Knight D, Smith C, et al. Retinoid signaling determinesgerm cell fate in mice. Sci Signal 2006; 312: 596.
29. Jørgensen A, Nielsen JE, Almstrup K, et al. Dysregulation of themitosis–meiosis switch in testicular carcinoma in situ. J Pathol 2013;10: 588–598.
30. Pellegrini M, Grimaldi P, Rossi P, et al. Developmental expression ofBMP4/ALK3/SMAD5 signaling pathway in the mouse testis: apotential role of BMP4 in spermatogonia differentiation. J Cell Sci2003; 116: 3363–3372.
31. Saadi I, Das P, Zhao M, et al. Msx1 and Tbx2 antagonistically regulateBmp4 expression during the bud-to-cap stage transition in toothdevelopment. Development 2013; 140: 2697–2702.
635the function of msx1
Copyright © 2013 John Wiley & Sons, Ltd. Cell Biochem Funct 2013; 31: 629–635.