the epidemiology of pneumococcal infections—the swedish experience
TRANSCRIPT

T
Ja
b
a
ARRA
KSMPS
1w
siiaaoywmasrEibdempam
KT
0d
Vaccine 27S (2009) G27–G32
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
he epidemiology of pneumococcal infections—The Swedish experience
essica Darenberga, Birgitta Henriques Normarka,b,∗
Swedish Institute for Infectious Disease Control, 171 82 Solna, SwedenDepartment of Microbiology, Tumor- and Cellbiology, Karolinska Institutet, SE171 77 Stockholm, Sweden
r t i c l e i n f o
rticle history:eceived 3 September 2009eceived in revised form 10 October 2009
a b s t r a c t
Pneumococcal infections are major contributors to morbidity and mortality world-wide and pose a majorpublic health problem. Despite being a devastating pathogen pneumococci are common colonizers of theupper respiratory tract of healthy children. There is a need for more knowledge on the molecular epidemi-
ccepted 16 October 2009
eywords:treptococcus pneumoniaeolecular epidemiology
ology, and pathogenesis of pneumococcal infections to be able to find better strategies for preventionand treatment of these common infections. Here we discuss trends in the vaccine era of the epidemiol-ogy of pneumococcal carriage, invasive disease and antibiotic resistance development as well as presentnational epidemiology data from Sweden of invasive pneumococcal infections during 1987–2006.
© 2009 Elsevier Ltd. All rights reserved.
athogenesisweden
. Pneumococcal infections—a major health problemorld-wide
Infectious diseases are after cardiovascular diseases theecond most common cause of death world-wide, account-ng for 26% of the overall mortality. Lower respiratory tractnfections are the major infectious cause of mortality withnnually 4.2 million deaths globally according to WHO (Fig. 1,dapted from www.int/healthinfo/global burden disease). More-ver, pneumonia is a major contributor to mortality among theoungest children below 5 years of age (Fig. 2, adapted fromww.int/healthinfo/global burden disease). Streptococcus pneu-oniae (also called pneumococci) is the major cause of community
cquired pneumonia, and a major contributor to invasive infectionsuch as septicaemia and meningitis. It also causes milder mucosalespiratory tract infections such as acute otitis media and sinusitis.ven though we have access to antibiotics and intensive care at leastn the developed world pneumococcal infections remain responsi-le for a huge burden of global morbidity and mortality in infectiousiseases and it has been estimated that over 1 million children dievery year world-wide from a pneumococcal infection [1]. Esti-
ates from the USA show that the fatality rates in pneumococcalneumonia is about 5%, in combination with septicaemia about 20%nd in meningitis as high as 30% [2]. Survivors of pneumococcaleningitis are also at risk to get sequelae such as hearing loss. The
∗ Corresponding author at: Department of Microbiology, Tumor- and Cellbiology,arolinska Institutet, Nobels väg 16, SE171 77 Stockholm, Sweden.el.: +46 8 457 24 13; fax: +46 8 30 25 66.
E-mail address: [email protected] (B. Henriques Normark).
264-410X/$ – see front matter © 2009 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2009.10.088
risk groups for acquiring pneumococcal infections are the youngestchildren, the elderly and those that are immuno-compromised forexample HIV patients, but also individuals that have been splenec-tomised. Furthermore, a prior influenza infection predisposes forpneumococcal infections and it has been shown that several of thedeaths caused during the 1918 pandemic influenza were caused bysuperinfections with bacteria, mainly pneumococci [3–5].
Despite being a devastating pathogen pneumococci are alsocommon colonizers of the upper respiratory tract in children.Healthy children attending day-care centres have been shown tocarry these bacteria in the nasopharynx in up to 60–70% [6,7]. Car-riage rates decrease with age and adult individuals without childrenat home have a low carriage rate. Still it is not clear why thesebacteria sometimes cause disease but usually colonizes harmlesslyyoung children.
2. Molecular epidemiology of pneumococcal infections
Pneumococci can be divided into at least 91 different serotypesdepending on differences in their capsular polysaccharides.Virulence has been shown to differ with capsular type and non-encapsulated pneumococci have been regarded as non-virulent.However, non-encapsulated pneumococci may be found in thenasopharynx of children, the major ecological niche for pneu-mococci. The current pneumococcal vaccines are based on a
limited number of pneumococcal capsular polysaccharide types.The polysaccharide based vaccine, including 23 serotypes, has beenused for decades but due to the low protective value of the vaccineespecially in some of the risk groups, the youngest children underthe age of two years, and persons suffering from immunodeficiency,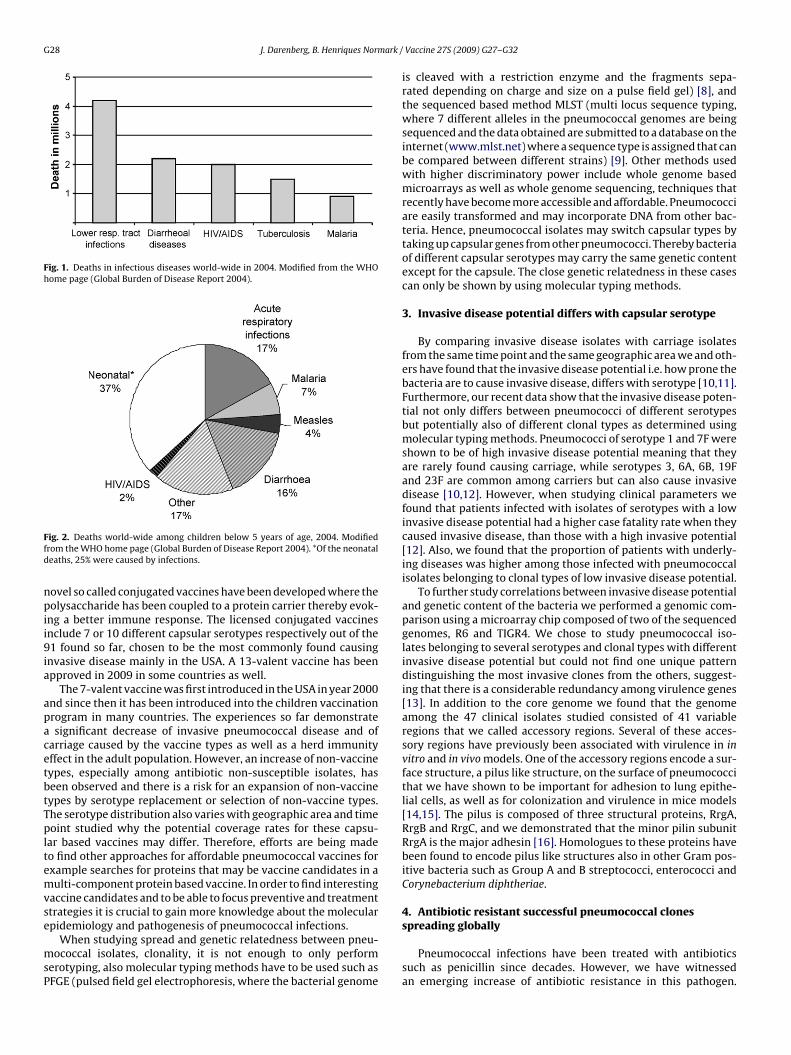
G28 J. Darenberg, B. Henriques Normark /
Fig. 1. Deaths in infectious diseases world-wide in 2004. Modified from the WHOhome page (Global Burden of Disease Report 2004).
Ffd
npii9ia
apacetbtTpltemvse
msP
ig. 2. Deaths world-wide among children below 5 years of age, 2004. Modifiedrom the WHO home page (Global Burden of Disease Report 2004). *Of the neonataleaths, 25% were caused by infections.
ovel so called conjugated vaccines have been developed where theolysaccharide has been coupled to a protein carrier thereby evok-
ng a better immune response. The licensed conjugated vaccinesnclude 7 or 10 different capsular serotypes respectively out of the1 found so far, chosen to be the most commonly found causing
nvasive disease mainly in the USA. A 13-valent vaccine has beenpproved in 2009 in some countries as well.
The 7-valent vaccine was first introduced in the USA in year 2000nd since then it has been introduced into the children vaccinationrogram in many countries. The experiences so far demonstratesignificant decrease of invasive pneumococcal disease and of
arriage caused by the vaccine types as well as a herd immunityffect in the adult population. However, an increase of non-vaccineypes, especially among antibiotic non-susceptible isolates, haseen observed and there is a risk for an expansion of non-vaccineypes by serotype replacement or selection of non-vaccine types.he serotype distribution also varies with geographic area and timeoint studied why the potential coverage rates for these capsu-
ar based vaccines may differ. Therefore, efforts are being madeo find other approaches for affordable pneumococcal vaccines forxample searches for proteins that may be vaccine candidates in aulti-component protein based vaccine. In order to find interesting
accine candidates and to be able to focus preventive and treatmenttrategies it is crucial to gain more knowledge about the molecular
pidemiology and pathogenesis of pneumococcal infections.When studying spread and genetic relatedness between pneu-ococcal isolates, clonality, it is not enough to only perform
erotyping, also molecular typing methods have to be used such asFGE (pulsed field gel electrophoresis, where the bacterial genome
Vaccine 27S (2009) G27–G32
is cleaved with a restriction enzyme and the fragments sepa-rated depending on charge and size on a pulse field gel) [8], andthe sequenced based method MLST (multi locus sequence typing,where 7 different alleles in the pneumococcal genomes are beingsequenced and the data obtained are submitted to a database on theinternet (www.mlst.net) where a sequence type is assigned that canbe compared between different strains) [9]. Other methods usedwith higher discriminatory power include whole genome basedmicroarrays as well as whole genome sequencing, techniques thatrecently have become more accessible and affordable. Pneumococciare easily transformed and may incorporate DNA from other bac-teria. Hence, pneumococcal isolates may switch capsular types bytaking up capsular genes from other pneumococci. Thereby bacteriaof different capsular serotypes may carry the same genetic contentexcept for the capsule. The close genetic relatedness in these casescan only be shown by using molecular typing methods.
3. Invasive disease potential differs with capsular serotype
By comparing invasive disease isolates with carriage isolatesfrom the same time point and the same geographic area we and oth-ers have found that the invasive disease potential i.e. how prone thebacteria are to cause invasive disease, differs with serotype [10,11].Furthermore, our recent data show that the invasive disease poten-tial not only differs between pneumococci of different serotypesbut potentially also of different clonal types as determined usingmolecular typing methods. Pneumococci of serotype 1 and 7F wereshown to be of high invasive disease potential meaning that theyare rarely found causing carriage, while serotypes 3, 6A, 6B, 19Fand 23F are common among carriers but can also cause invasivedisease [10,12]. However, when studying clinical parameters wefound that patients infected with isolates of serotypes with a lowinvasive disease potential had a higher case fatality rate when theycaused invasive disease, than those with a high invasive potential[12]. Also, we found that the proportion of patients with underly-ing diseases was higher among those infected with pneumococcalisolates belonging to clonal types of low invasive disease potential.
To further study correlations between invasive disease potentialand genetic content of the bacteria we performed a genomic com-parison using a microarray chip composed of two of the sequencedgenomes, R6 and TIGR4. We chose to study pneumococcal iso-lates belonging to several serotypes and clonal types with differentinvasive disease potential but could not find one unique patterndistinguishing the most invasive clones from the others, suggest-ing that there is a considerable redundancy among virulence genes[13]. In addition to the core genome we found that the genomeamong the 47 clinical isolates studied consisted of 41 variableregions that we called accessory regions. Several of these acces-sory regions have previously been associated with virulence in invitro and in vivo models. One of the accessory regions encode a sur-face structure, a pilus like structure, on the surface of pneumococcithat we have shown to be important for adhesion to lung epithe-lial cells, as well as for colonization and virulence in mice models[14,15]. The pilus is composed of three structural proteins, RrgA,RrgB and RrgC, and we demonstrated that the minor pilin subunitRrgA is the major adhesin [16]. Homologues to these proteins havebeen found to encode pilus like structures also in other Gram pos-itive bacteria such as Group A and B streptococci, enterococci andCorynebacterium diphtheriae.
4. Antibiotic resistant successful pneumococcal clones
spreading globallyPneumococcal infections have been treated with antibioticssuch as penicillin since decades. However, we have witnessedan emerging increase of antibiotic resistance in this pathogen.

ark / Vaccine 27S (2009) G27–G32 G29
RlbtnsrlI(mcp1ig
btcttnbotCPraacIisteop2vo
Sbbidtr1icMbscsiSntwwcb
the collected isolates per year. The most commonly found serotypesamong all patients were type 3, 4, 6B, 7F, 9V, 14, and 23F. However,serotype 1 was very common in 1997 but decreased tremendouslyduring 2006. During these years in Sweden we found, as could be
J. Darenberg, B. Henriques Norm
esistance has developed against most antibiotic classes and theast resort consists of vancomycin to which no resistance haseen found yet. Multi-resistance is emerging potentially creatingreatment problems of these common infections. The Europeanetwork EARSS studies resistance rates in Europe among inva-ive pneumococcal isolates and they have observed that resistanceates vary between European countries where the rates areower in Northern Europe as compared to the Southern part.n some countries the resistance rates to penicillin exceeds 50%http://www.rivm.nl/earss/database). The spread of resistance is
ainly due to the successful spread of specific pneumococcallones. One such example is the spread of the serotype 6B clone thatrobably emanated from Spain and emerged on Iceland during the990s creating treatment problems of common respiratory tract
nfections such as otitis media, since the bacteria carried resistanceenes to several antibiotics [17].
Pneumococci with a reduced susceptibility to penicillin (PNSP)ecame mandatory to report to the Swedish Institute for Infec-ious Disease Control (MIC ≥ 0.5 mg/L) in 1996. Since 1998 we haveollected all PNSP isolates from the Swedish microbiological labora-ories for further characterization. The dominant serotype has beenype 9V since many years back [18,19] and with molecular tech-iques, PFGE and MLST, we have demonstrated that the isolates allelong to the same clone, CC156. Recently we observed an increasef serotype 14 isolates and with molecular typing we concludedhat the increase was due to an increase in the same clonal complexC156 but of another capsular serotype [19]. Also among the 19FNSP isolates we found CC156 isolates. CC156 is an internationallyecognized clonal complex that has been found on most continentsnd represents one of the most successfully spread pneumococcalntibiotic resistant clones in the world. It carries reduced sus-eptibility to both penicillin and trimetoprim–sulfametoxazole.nterestingly, we have found that all CC156 isolates except for onesolate that we have tested so far harbour pili genes and express thetructure. We propose that one reason for the successful spread ofhis clone is that it carries pili that facilitate colonization. When wexamined the other Swedish PNSP isolates we found that severalf the other dominating serotypes in Sweden, 6B and 35B, also hadili and totally more than 70% of the PNSP strains in Sweden in003 were piliated [19]. This is the first example where a bacterialirulence factor has been suggested to be important for the spreadf antibiotic resistant clones [15].
After the introduction of the conjugated vaccine in the Unitedtates, non-vaccine isolates, including drug-resistant strains, haveeen identified in patients in increasing numbers. An increase haseen observed in serotypes such as types 3, 15, 19A, 22F and 33F
n children and the non-vaccine serotype 19A has become the pre-ominant cause of invasive pneumococcal disease in children inhe United States [20]. Also, a high proportion of the multidrug-esistant strains have been shown to be of the non-vaccine serotype9A [21,22]. Recently a S. pneumoniae 19A strain was discovered
n the United States that is resistant to all FDA—approved antimi-robial drugs for treatment of acute otitis media in children [23].olecular typing with MLST showed that this strain was of ST2722,
elonging to the same clonal lineage as CC156. In addition toerotype 19A this multidrug-resistant lineage was found with otherapsular types, 9V, 14, 11A, 15C, and 19F [24]. Other penicillin non-usceptible clones of non-vaccine serotypes have also expandedn the post-vaccine era in the United States, one example beingT558 of serotype 35B [25]. Several of these emerging penicillinon-susceptible, non-vaccine isolates, belong to clonal complexes
hat have been shown to possess pneumococcal pili [26]. Hence,e suggest that we might witness a clonal selection of clones thatere minor lineages in the carriage population, but which have theapacity to expand potentially promoted by properties mediatedy pili [15].
Fig. 3. Number of reports sent to the Swedish Institute for Infectious disease Controlper year 1989–2006 (www.smittskyddsinstitutet.se). It became mandatory to reportinvasive pneumococcal infections in Sweden the 1st of July 2004, hence, year 2004is lacking since the system changed this year.
5. Invasive pneumococcal infections in Sweden
Reports on invasive pneumococcal cases have been sent fromthe clinical microbiological laboratories in Sweden to the SwedishInstitute for Infectious Disease Control every year since the 1980son a voluntary basis. However, since the 1st of July 2004 it becamemandatory to report the number of invasive infections (i.e. pneu-mococci isolated from sterile sites such as blood, and meninges)caused by pneumococci from the clinicians as well as from theclinical microbiology laboratories to the Swedish Institute for Infec-tious Disease Control. Fig. 3 shows the number of reports per yearsince 1989 (www.smittskyddsinstitutet.se). During the 1990s weobserved a tremendous increase in the number of reports whichcould not be solely explained by an increased blood sampling orchanged routines at that time [27,28]. To clarify the reason for thisincrease we collected most of the corresponding invasive pneu-mococcal isolates from the clinical microbiological laboratories inSweden and performed molecular typing of invasive pneumococ-cal isolates collected during 1987, 1992 and 1997. We found thatthe increase observed between 1987 and 1992 was partly due tothe expansion of one specific clonal type of serotype 14 (ST124)and between 1992 and 1997 we found a corresponding increaseof a clone of serotype 1 (ST306), as determined using Box finger-printing, PFGE and MLST [29]. These data showed that the serotypedistribution differs with time period studied where changes maybe observed in as short time periods as 5 years. To follow theserotype distribution over time we collected all Swedish invasivepneumococcal isolates also from 2002 and 2006. Fig. 4 shows theserotype distribution for the 15 most prominent serotypes among
Fig. 4. Serotype distribution among adults and children with invasive pneumococ-cal infections in Sweden during 1987, 1992, 2002 and 2006.
G30 J. Darenberg, B. Henriques Normark / Vaccine 27S (2009) G27–G32
Fig. 5. Age distribution among patients with invasive pneumococcal disease in Swe-den (based on the isolates that have been sent to the Swedish Institute for InfectiousDisease Control).
Fsi
emsiiam2tIis2cFftti
6p
iiItiabi
Table 1Potential coverage rates (%) for the 7-, 10- and 13-valent vaccines among invasivepneumococcal isolates from adults and children collected from the microbiologicallaboratories in Sweden.
Year 1987 1992 1997 2002 2006
No of isolates 148 464 525 964 1137PCV7 (%) 47 51 38 49 54PCV10 (%) 60 63 60 63 67PCV13 (%) 71 77 74 76 80
Table 2Potential coverage rates (%) for the 7-, 10- and 13-valent vaccines among isolatesfrom pediatric invasive cases ≤2years of age in Sweden.
Year 1987 1992 1997 2002 2006
No of isolates 11 10 17 49 73PCV7 (%) 64 50 35 65 78PCV10 (%) 82 60 41 82 85PCV13 (%) 82 60 65 89 95
Table 3Potential coverage rates (%) for the 7-, 10- and 13-valent vaccines among isolatesfrom children with invasive disease >2 years of age in Sweden. (Only 2 invasiveisolates were collected from year 1987 in this age group).
Year 1987 1992 1997 2002 2006
No of isolates 2 11 25 15 31
ig. 6. Serotype distribution among isolates from children ≤2 years of age with inva-ive pneumococcal disease collected from the clinical microbiological laboratoriesn Sweden.
xpected, that pneumococcal invasive infections were most com-on in the youngest children as well as in the elderly, as has been
hown in studies from other countries. However, also middle agedndividuals could get a pneumococcal infection (Fig. 5). Interest-ngly we found that the serotype distribution differs between thedult population and the children. Serotypes 6B and 14 was theost commonly found serotypes among the youngest children in
006 while in addition serotypes 3, 4, 7F, 9V and 23F also belongedo the most commonly found types in the adult population (Fig. 6).n a previous study where we collected invasive pneumococcalsolates from children in Sweden during 1998–2001 we demon-trated that serotype 1 pneumococci infected older children overyears of age, while serotype 1 was lacking among the youngest
hildren below 2 years of age as a cause of invasive disease [30].urthermore, in a study of adult patients with invasive disease weound that serotype 1 isolates mainly caused invasive disease inhe younger adult population, while serotype 23F was common inhe elderly [12]. Hence, the serotype distribution among invasivenfections differs with age.
. Potential vaccine coverage rates for conjugatedneumococcal vaccines
The 7-valent pneumococcal conjugated vaccine (PCV7), includ-ng serotypes 4, 6B, 9V, 14, 18C, 19F, 23F, has now been includedn several countries’ childhood vaccination programmes in Europe.n Sweden it was introduced in the national vaccination programhe 1st of January 2009. A 10-valent conjugated vaccine (PCV10),
ncluding in addition to the 7 serotypes above, serotypes 1, 5 and 7Fs well as a protein carrier derived from H. influenzae, has now alsoeen licensed and a 13-valent conjugated vaccine (PCV13) includ-ng also serotypes 3, 6A, 19A is expected to come soon.
PCV7 (%) 50 36 8 33 45PCV10 (%) 100 64 64 60 65PCV13 (%) 100 82 68 60 71
The potential coverage rates in Sweden for these vaccines duringyears 1987–2006 are shown in Tables 1–3. The potential cover-age rates among all invasive cases ranges between 38% and 54%for PCV7 (Table 1). However, we found that the potential coveragerates differs with age and was highest among the youngest chil-dren below 2 years of age where up to 78% (in 2006) would havebeen covered by PCV7 (Table 2). Since serotype 1 and 7F are com-mon serotypes in Sweden during the years studied we find thatthe coverage rates for PCV7 are affected by the fluctuations espe-cially among serotype 1 isolates. Hence, we find low coverage ratesduring 1997 depending mainly on the expansion of serotype 1 bythis time point. Calculations on potential coverage rates for PCV10and PCV13 show that these vaccines have higher vaccine coveragerates than PCV7 depending on the inclusion of common serotypesin Sweden (types 1, 7F in the 10-valent and 3, 6A and 19A in the13-valent) in theses vaccines (Tables 1–3).
The data presented above shows that in Sweden we know theserotype distribution and burden of disease before the introduc-tion of the pneumococcal conjugated vaccines in Sweden. We willnow follow the development after vaccine introduction and are cur-rently collecting all invasive pneumococcal isolates continuouslyfrom the whole of Sweden as well as clinical information aboutthe patients. It will be interesting to monitor the effect of the vac-cine introduction on the rate of invasive pneumococcal disease,on the clinical picture among the patients as well as on the pos-sible changes in the serotype distribution. These data will be mostimportant for optimal decisions by policy makers regarding futurevaccine strategies against pneumococcal infections.
7. Epidemiological aspects on a future protein basedpneumococcal vaccine
Since the conjugated vaccines so far include a limited amount ofpneumococcal serotypes, other approaches for making new pneu-mococcal vaccines are under investigation, such as attempts to findprotein antigens that could be included into a multi-component

ark /
pptvmahits(atbpc
8
8
I4C
8
dGzM
8
Hr1itaMsdt[
8
E[a(
A
dIJtm
[
[
[
[
[
[
[
[
[
[
[
[
[
J. Darenberg, B. Henriques Norm
neumococcal protein based vaccine. In the search for suitableneumococcal antigens most efforts have been devoted to protec-ive antigens that are expressed by 100% of analyzed pneumococcalaccines i.e. belong to the core genome and that do not appear inany allelic variants [31]. Protective antigens that are encoded on
ccessory genes only present in a subset of pneumococcal clonesave therefore usually been discarded. However, our epidemiolog-
cal data from Sweden shows that children isolates belonging tohe dominating clonal complexes comprise a majority of all inva-ive isolates from the Stockholm area during the period 1997–2004unpublished data). A search for protective antigens encoded by theccessory genome of each of these dominating clones might formhe core of a multi-protein based vaccine, that however need toe modified as the clonal spectrum changes over time and vaccineressure, much in the same way as is currently the case for theonjugated polysaccharide vaccines.
. Materials and methods
.1. Serotyping
All invasive isolates were serotyped at the Swedish Institute fornfectious Disease Control (Solna, Sweden) using gel diffusion with6 serotype/group antisera obtained from Statens Seruminstitut inopenhagen, Denmark [7].
.2. Antibiotic susceptibility testing
Isolates were tested for antibiotic susceptibility using the disciffusion method and the guidelines from the Swedish Referenceroup for Antibiotics (www.srga.org). Isolates with an inhibition-one of <18 mm for 1 �g of oxacillin were subjected to analysis ofIC for benzylpenicillin using Etest (AB BIODISK).
.3. Pulsed field gel electrophoresis
The PFGE method was adapted from the procedure described byermans et al. [8]. The chromosomal DNA were cleaved with the
estriction enzyme Apa1. The fragments were then separated on a% agarose gel, by use of PFGE performed for 22 h at 14 ◦C at 6 V/cm
n 0.5× Tris-borate EDTA, with pulse time of 2–30 s. The gels werehan stained with 1 mg/mL of ethidium bromide for 30 min andnalyzed using the programme Bionumerics version 4.5 (Appliedath), with Dice and UPGMA dendogram type. The optimization
ettings were 1% and position tolerance of 1.5%. The clones wereefined using the criteria described by Tenover et al. i.e. isolateshat differ with ≤3 bands were considered to be closely related32].
.4. Multi locus sequence typing (MLST)
This method was adapted from the procedure described bynright and Spratt [9] and performed as previously described7]. Pair-wise comparisons of how many alleles differed betweenny pair of isolates was performed using the MLST websitewww.mlst.net).
cknowledgements
We thank all the clinical microbiological laboratories in Swe-
en for sending in invasive pneumococcal isolates to the Swedishnstitute for Infectious Disease Control. We also thank Christinaohansson, Ingrid Andersson and Gunnel Möllerberg for excellentechnical support and Staffan Normark for critically reading the
anuscript.
[
[
Vaccine 27S (2009) G27–G32 G31
Conflict of Interest
The authors state that they have no conflict of interest.
References
[1] Cartwright K. Pneumococcal disease in western Europe: burden of dis-ease, antibiotic resistance and management. Eur J Pediatr 2002;161(April(4)):188–95.
[2] Tomasz A. Antibiotic resistance in Streptococcus pneumoniae. Clin Infect Dis1997;24(January (suppl. 1)):S85–8.
[3] Klugman KP, Madhi SA. Pneumococcal vaccines and flu preparedness. Science2007;316(April (5821)):49–50.
[4] McCullers JA. Insights into the interaction between influenza virus and pneu-mococcus. Clin Microbiol Rev 2006;19(July (3)):571–82.
[5] McAuley JL, Hornung F, Boyd KL, Smith AM, McKeon R, Bennink J, et al. Expres-sion of the 1918 influenza A virus PB1-F2 enhances the pathogenesis of viraland secondary bacterial pneumonia. Cell Host Microbe 2007;2(October (4)):240–9.
[6] De Lencastre H, Tomasz A. From ecological reservoir to disease: the nasophar-ynx, day-care centres and drug-resistant clones of Streptococcus pneumoniae. JAntimicrob Chemother 2002;50(December (suppl. S2)):75–81.
[7] Henriqus Normark B, Christensson B, Sandgren A, Noreen B, Sylvan S, BurmanLG, et al. Clonal analysis of Streptococcus pneumoniae nonsusceptible to peni-cillin at day-care centers with index cases, in a region with low incidence ofresistance: emergence of an invasive type 35B clone among carriers. MicrobDrug Resist 2003;9(Winter (4)):337–44.
[8] Hermans PW, Sluijter M, Hoogenboezem T, Heersma H, van Belkum A, de GrootR. Comparative study of five different DNA fingerprint techniques for molec-ular typing of Streptococcus pneumoniae strains. J Clin Microbiol 1995;33(June(6)):1606–12.
[9] Enright MC, Spratt BG. A multilocus sequence typing scheme for Streptococcuspneumoniae: identification of clones associated with serious invasive disease.Microbiology 1998;144(November (Pt 11)):3049–60.
10] Sandgren A, Sjostrom K, Olsson-Liljequist B, Christensson B, Samuelsson A, Kro-nvall G, et al. Effect of clonal and serotype-specific properties on the invasivecapacity of Streptococcus pneumoniae. J Infect Dis 2004;189(March (5)):785–96.
11] Brueggemann AB, Peto TE, Crook DW, Butler JC, Kristinsson KG, Spratt BG.Temporal and geographic stability of the serogroup-specific invasive diseasepotential of Streptococcus pneumoniae in children. J Infect Dis 2004;190(October(7)):1203–11.
12] Sjostrom K, Spindler C, Ortqvist A, Kalin M, Sandgren A, Kuhlmann-Berenzon S,et al. Clonal and capsular types decide whether pneumococci will act as a pri-mary or opportunistic pathogen. Clin Infect Dis 2006;42(February (4)):451–9.
13] Blomberg C, Dagerhamn J, Dahlberg S, Browall S, Fernebro J, Albiger B, et al.Pattern of accessory regions and invasive disease potential in Streptococcuspneumoniae. J Infect Dis 2009;199(April (7)):1032–42.
14] Barocchi MA, Ries J, Zogaj X, Hemsley C, Albiger B, Kanth A, et al. A pneumo-coccal pilus influences virulence and host inflammatory responses. Proc NatlAcad Sci USA 2006;103(February (8)):2857–62.
15] Henriques-Normark B, Blomberg C, Dagerhamn J, Battig P, Normark S. Therise and fall of bacterial clones: Streptococcus pneumoniae. Nat Rev Microbiol2008;6(November (11)):827–37.
16] Nelson AL, Ries J, Bagnoli F, Dahlberg S, Falker S, Rounioja S, et al. RrgAis a pilus-associated adhesin in Streptococcus pneumoniae. Mol Microbiol2007;66(October (2)):329–40.
17] Soares S, Kristinsson KG, Musser JM, Tomasz A. Evidence for the introductionof a multiresistant clone of serotype 6B Streptococcus pneumoniae from Spainto Iceland in the late 1980s. J Infect Dis 1993;168(July (1)):158–63.
18] Hogberg L, Henriques Normark B, Ringberg H, Stenqvist K, Fredlund H, Geli P,et al. The impact of active intervention on the spread of penicillin-resistantStreptococcus pneumoniae in Swedish day-care centres. Scand J Infect Dis2004;36(9):629–35.
19] Sjostrom K, Blomberg C, Fernebro J, Dagerhamn J, Morfeldt E, Barocchi MA, etal. Clonal success of piliated penicillin nonsusceptible pneumococci. Proc NatlAcad Sci USA 2007;104(July (31)):12907–12.
20] Hicks LA, Harrison LH, Flannery B, Hadler JL, Schaffner W, Craig AS, et al. Inci-dence of pneumococcal disease due to non-pneumococcal conjugate vaccine(PCV7) serotypes in the United States during the era of widespread PCV7 vac-cination, 1998–2004. J Infect Dis 2007;196(November (9)):1346–54.
21] Mera RM, Miller LA, Daniels JJ, Weil JG, White AR. Increasing prevalence ofmultidrug-resistant Streptococcus pneumoniae in the United States over a 10-year period: Alexander project. Diagn Microbiol Infect Dis 2005;51(March(3)):195–200.
22] Pelton SI, Huot H, Finkelstein JA, Bishop CJ, Hsu KK, Kellenberg J, et al.Emergence of 19A as virulent and multidrug resistant pneumococcus in Mas-sachusetts following universal immunization of infants with pneumococcalconjugate vaccine. Pediatr Infect Dis J 2007;26(June (6)):468–72.
23] Pichichero ME, Casey JR. Emergence of a multiresistant serotype 19A pneumo-coccal strain not included in the 7-valent conjugate vaccine as an otopathogenin children. JAMA 2007;298(October (5)):1772–8.
24] Xu Q, Pichichero ME, Casey JR, Zeng M. Novel type of Streptococcus pneumoniaecausing multidrug-resistant acute otitis media in children. Emerg Infect Dis2009;15(April (4)):547–51.

G ark /
[
[
[
[
[
[
[
32 J. Darenberg, B. Henriques Norm
25] Hanage WP, Huang SS, Lipsitch M, Bishop CJ, Godoy D, Pelton SI, et al. Diversityand antibiotic resistance among nonvaccine serotypes of Streptococcus pneu-moniae carriage isolates in the post-heptavalent conjugate vaccine era. J InfectDis 2007;195(February (3)):347–52.
26] Moschioni M, Donati C, Muzzi A, Masignani V, Censini S, Hanage WP, et al.Streptococcus pneumoniae contains 3 rlrA pilus variants that are clonally related.J Infect Dis 2008;197(March (6)):888–96.
27] Hedlund J, Svenson SB, Kalin M, Henrichsen J, Olsson-Liljequist B, Mollerberg G,
et al. Incidence, capsular types, and antibiotic susceptibility of invasive Strep-tococcus pneumoniae in Sweden. Clin Infect Dis 1995;21(October (4)):948–53.28] Normark BH, Ortqvist A, Kalin M, Olsson-Liljequist B, Hedlund J, SvensonSB, et al. Changes in serotype distribution may hamper efficacy of pneu-mococcal conjugate vaccines in children. Scand J Infect Dis 2001;33(11):848–50.
[
Vaccine 27S (2009) G27–G32
29] Henriques Normark B, Kalin M, Ortqvist A, Akerlund T, Liljequist BO, HedlundJ, et al. Dynamics of penicillin-susceptible clones in invasive pneumococcaldisease. J Infect Dis 2001;184(October (7)):861–9.
30] Hedlund J, Sorberg M, Henriques Normark B, Kronvall G. Capsular types andantibiotic susceptibility of invasive Streptococcus pneumoniae among childrenin Sweden. Scand J Infect Dis 2003;35(8):452–8.
31] Giefing C, Meinke AL, Hanner M, Henics T, Bui MD, Gelbmann D, et al. Discov-ery of a novel class of highly conserved vaccine antigens using genomic scale
antigenic fingerprinting of pneumococcus with human antibodies. J Exp Med2008;205(January (1)):117–31.32] Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, etal. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol1995;33(September (9)):2233–9.