the effect of river red gum decline on woodland birds in ... · the effect of river red gum decline...
TRANSCRIPT

The effect of
river red gum decline on woodland birds in the Macquarie MarshesAlice Blackwood, Richard Kingsford, Lucy Nairn, Tom Rayner
Australian Wetlands & Rivers CentreSchool of Biological, Environmental and Earth Sciences
University of New South Wales
May 2010

Acknowledgements
We wish to acknowledge the Wailwan people, traditional owners of the Macquarie
Marshes, and the Eora people, traditional owners of UNSW and Sydney.
This project was made possible by the generous funding of the NSW Department of
Environment, Climate Change and Water, the Australian Geographic Society, the Wildlife
Preservation Society of Australia and the Birds Australia (Stewart Lesley Bird Research
Award).
Thank you to everyone at the Wetlands and Rivers Centre, in particular to Kim Jenkins, Ben
Wolfenden and John Porter for statistics advice and Shiquan Ren for providing flood history
data.
We would like to thank all our fieldwork volunteers: Julia Blackwood, Jenny Spencer, Kathy
Wilk, Heather Hartford, Tessa Dowdell, Fiona Johnson, Sujata Allan, Jo Ocock, Chris
Marshall, Jim Blackwood, Misti de Montfort.
Thanks also to Ray and Sue Jones for sharing their wisdom, Steve Tucker for his general
assistance, Frank Hemmings, Mailie Gall, and Ross McMurtrie for their generous help with
equipment and software.

2
Table of Contents 1 Executive Summary ......................................................................................................................... 3
2 Introduction .................................................................................................................................... 5
3 Study area ....................................................................................................................................... 8
4 Methodology ................................................................................................................................... 9
4.1 Health categorisation .............................................................................................................. 9
4.2 Site selection ........................................................................................................................... 9
4.3 Woodland Bird surveys ......................................................................................................... 13
4.4 Vegetation ............................................................................................................................. 14
4.5 Guilds .................................................................................................................................... 18
4.6 Statistical analyses ................................................................................................................ 18
5 Results ........................................................................................................................................... 23
5.1 Birds ...................................................................................................................................... 23
5.2 Vegetation ............................................................................................................................. 34
6 Discussion ...................................................................................................................................... 47
7 References .................................................................................................................................... 52
Appendix 1: Bird species list- Macquarie Marshes, May-August 2009 ................................................. 59
Appendix 2: Understorey vegetation species list ................................................................................. 64
Appendix 3: Site locations ..................................................................................................................... 65

3
1 Executive Summary The state of woodland birds in Australia is well known with long-term declines attributed to
loss of habitat, mainly through land clearing. Woodland birds are generally passerines (e.g.
honeyeaters and robins) but also include parrots, pigeons, cuckoos and other bird. The
relationship of woodland birds to rivers is not so well known nor is the effect of river
regulation on this group. Yet many woodland birds depend on the rivers and wetlands
because these are often the more productive areas in an otherwise dry environment. Even
when wetlands are dry, they often have larger and more dense trees which provide good
habitat for woodland birds. We found that woodland bird communities changed
substantially depending on the health of floodplain trees. Healthier floodplain communities
had higher abundances of foliage specialist birds, reflecting greater canopy cover, while
floodplain communities in poor health had high abundances of birds that are typical of open
agricultural areas (eg. Crested pigeon, jacky winter), and a community similar to surrounding
terrestrial areas.The number of species and their abundance remained similar across health
categories but the community composition changed.
Regulation of rivers worldwide has reduced the ecological resilience and biodiversity of
riparian and floodplain ecosystems and their dependent biota. Most knowledge of this is of
the effects on aquatic organisms, but some terrestrial animals, such as woodland birds, may
also be dependent on flows. We investigated differences in the woodland bird community
among stands of river red gums in the Macquarie Marshes in three health categories: poor,
intermediate and good. We created a map of river red gum health, categorising stand health
as good, intermediate or poor from aerial photos, for selection of 10 sites for woodland bird
surveys in each health category. We surveyed woodland birds at each site in autumn and
spring over a two-hectare area for 20 minutes, recording species, abundance and
microhabitat preferences. At each site, we surveyed vegetation: species, structure,
abundance and health of understorey and trees. We also surveyed birds at three terrestrial
sites, further out on the floodplain.
There was no difference in the abundance of large (50 – 100 cm and >100 cm diameter at
breast height) trees among the tree health categories (F2,27=2.056, p=0.13 and F=1.979,
p=0.14 respectively), indicating that sites were once in similar condition. Abundance of small
trees (2-10 cm, usually young) was significantly different among health categories
(F2,27=5.88, p=0.004), with high abundance at good sites reflecting more favourable flooding
conditions in recent years. There was no difference in abundance of hollows among the
three health categories (F2,27=1.94, p=0.16), but the number of fruiting river red gums did
differ, favouring healthier trees (F2,27=6.105, p<0.001).
The composition of the woodland bird community was significantly different among health
categories (PERMANOVA PseudoF=2.1538, P(perm)=0.001) even though there was no
difference in species richness, overall abundance or diversity. Furthermore, the bird
communities of the three terrestrial sites were similar to those of the poor and intermediate

4
sites, indicating a ‘terrestrialisation’ of the system. Many of the species that were more
abundant at poor sites, such as rufous songlark and crested pigeon, are typical of open,
agricultural areas. The patterns of the bird community were significantly correlated with
changes in tree health and habitat structure (BEST analysis, Rho=0.405, p=0.01), with
patterns best explained by five variables: canopy density, bare ground, shrub cover,
percentage of dead branches and large coarse-woody debris. Good sites were typified by
trees with dense canopy, more leaf litter and low green herbaceous understorey, while poor
sites had trees with thin or absent canopy, more bare ground and tall, dry, shrubby
understorey.
Foliage specialists, such as striated pardalote, weebill, grey fantail and crested shrike-tit,
were most affected by river red gum decline, with declining canopy cover causing decreased
abundances. Live branch specialists, such as galah and white-breasted cuckoo-shrike were
also most abundant at healthy sites. They were replaced by dead tree specialists, such as
hooded robin, rufous songlark, southern whiteface and crested pigeon, which were more
abundant at sites in poor health. Generalist species, such as magpie lark, willie wagtail and
jacky winter, were most resilient to decline, while understorey specialists (fairy-wrens) were
most abundant at sites of intermediate health, probably due to the combination of a
shrubby understorey with some live trees and herb cover. These changes in the bird
community and vegetation were linked with flood history, reflecting long term degradation.
Habitat quality of poor sites will probably degrade further, as dead trees fall over, reducing
the availability of perches and nesting hollows.
The river red gum forests and woodlands of the Macquarie Marshes Northern Nature
Reserve were severely degraded. At poor sites, an average of 58% of tree basal area was
dead, in comparison to 20% at intermediate and poor sites. Cunningham scores of sites,
which are a composite of crown vigour, canopy density and percentage live basal area,
indicated that no sites were in truly good condition; instead, 17 were classified as ‘poor’,
eight as ‘degraded’ and five as ‘severely degraded’. The dominance of the aggressive white-
plumed honeyeater is a further evidence for this degradation. As river red gum health has
declined, the understorey has changed significantly, with the dominance of dense terrestrial
chenopod shrub species such as Sclerolaena sp., replacing aquatic plant species and herbs.
The Macquarie Marshes are losing their unique ecological value and this is not only
reflected in declining aquatic biota but also a changing woodland bird community. The
woodland bird community is increasingly reflecting one of terrestrial habitats, rather than
an aquatic ecosystem. Reversing this decline requires increases in environmental flows,
currently supported by major government programs.

5
2 Introduction Rivers not only drive aquatic ecosystems, supporting their dependent organisms and
processes but also supply resources for some terrestrial organisms (Baxter et al. 2005;
Kingsford et al. 2006; Likens and Bormann 1974). Resources that move across permeable
boundaries from one system to another in this way are known as ‘subsidies’, ‘donations’ or
‘allochthonous inputs’ (Ballinger and Lake 2006) and may benefit organisms directly or
indirectly (Polis et al. 1997). These resources support higher densities of consumers than
would be possible in purely terrestrial systems, augment productivity (Bastow et al. 2002;
Ballinger and Lake 2006) and increase food web stability (Huxel and McCann 1998). For
example, flooding enhances the survival and dispersal of the yellow-footed antechinus
(Antechinus flavipes), a terrestrial marsupial (Lada et al. 2007). Even passerines at high
densities in Alaskan forests are supported by salmon-spawning streams (Gende and Willson
2001). Nutrients and detritus carried by floodwaters, as well as the water, usually boost
primary productivity of terrestrial systems (Polis et al. 1997). Aquatic insects can be an
important food source for terrestrial consumers (Gray 1993; Ballinger and Lake 2006; Chan
et al. 2008). For example aquatic invertebrates can make up over 25 % of the annual total
energy budget of forest birds in Japan (Nakano and Murakami 2001). Higher densities of
woodland birds, especially insectivores are common in riparian areas, compared to nearby
areas (Stauffer and Best 1980; Ford et al. 2001; Gende and Willson 2001; Palmer and
Bennett 2006).
As our understanding of the importance of river systems for terrestrial species increases,
there is accumulating evidence that freshwater ecosystems are declining in function and
resilience around the world. Over half of the world’s large river systems are now affected by
dams, which are capable of storing about 15% of the global total annual river runoff (Nilsson
et al. 2005). Worldwide impoundment of water has been so extensive that it has even
affected sea levels (Sahagian 2000) and minutely changed the Earth’s axis and increased the
speed of its rotation (Chao 1995). Riverine floodplains, among the most biologically
productive and diverse global ecosystems, are also among the most threatened (Lemly et al.
2000; Klement and Jack 2002). Increasing human populations are driving demand for
irrigated agriculture, power generation and water for urban and industrial use, substantially
increasing numbers of dams, water diversions, weirs, levees and off-river storages (Lemly et
al. 2000; Nilsson et al. 2005). Such river regulation has dramatically changed the flow of
rivers across the land, altering the frequency, duration and extent of floodplain inundation
Other threats are often additive: chemical contamination (Lemly et al. 2000), habitat
degradation, species invasions and climate change (Allan and Flecker 1993). A global
temperature rise of 3-4°C is predicted to eliminate 85% of the world’s wetlands (UNEP-
WCMC 2001).
In Australia, the most heavily regulated system is the Murray-Darling Basin (MDB) (Kingsford
2000), the nation’s most economically important river basin. It yields $15 billion a year in
agricultural products, accounts for more than half of Australia’s water consumption (ABS

6
2008), and contains some of the longest river systems in the world (Walker 2006). Before
regulation, the rivers of the MDB were characterised by high variability, slow flows and
gently sloping land (Reid and Brooks 2000). There are now more than twenty large
headwater dams and 6000 regulating weirs, and the discharge of rivers within the basin has
been reduced by between 20 and 81 % (Thoms 2006). More than 94 % of the flow capacity
of the system can be stored in the 30 largest dams (Walker 2006). Seasonality of flows in the
south has changed from predominantly winter and spring to summer flows, and the
frequency of small and medium sized floods has decreased dramatically (Maheshwari et al.
1995).
Such dramatic changes to flows have affected the ecological integrity of aquatic systems.
Loss of flooding reduces macroinvertebrate biomass (Boulton and Lloyd 1992) and species
richness (Jenkins and Boulton 2007, Sabella 2009) on floodplains. Fish have been affected,
with dams and other man-made structures creating barriers for fish passage, changing flows
and altering aquatic habitat (Gehrke et al. 1995; Kingsford et al. 2006; Rayner et al. 2009).
These changes to food sources along with altered habitat and breeding conditions have had
significant impacts upon waterbird distribution, diversity and breeding (Briggs et al. 1997;
Kingsford and Johnson 1998; Kingsford and Auld 2003; Kingsford et al. 2004; Kingsford and
Thomas 2004). In addition, river regulation has changed the distribution, diversity and
composition of vegetation communities (Shafroth et al. 2002; Brock et al. 2006; Richardson
et al. 2007, Sabella 2009). Perennial flood dependent eucalypts, such as river red gums
Eucalyptus camaldulensis have been particularly affected by major changes to river flows
(Bren 1988; Jansen and Robertson 2001b; MDBC 2003; Cunningham et al. 2007a;
Cunningham et al. 2007b)
River red gums are the most widely distributed eucalypt in Australia (Keith 2004;
Cunningham et al. 2007a) and grow in extensive monospecific stands or as lines of trees
along river banks in riparian areas (Di Stefano 2002). Across large areas of the MDB river red
gums are exhibiting deteriorating health, canopy dieback, insect damage, minimal
regeneration, increased parasitism, and eventually tree death (MDBC 2003; Cunningham et
al. 2007a). Some trees have also been killed by prolonged inundation (Briggs et al. 1997; Di
Stefano 2002). Lack of flooding can almost halve the relative growth rate of river red gums
and decrease average leaf area (Cunningham et al. 2007a). The vegetation understorey may
also change with reduced flooding frequency (Bren and Gibbs 1986; Bren 1988; Nairn and
Kingsford, unpubl. data).
Such major changes to floodplains and flooding regimes alter productivity at the landscape
level, with effects cascading through the food web (Bunn et al. 2006). River red gums play
an important role in nutrient cycling between floodplains and rivers (Cunningham et al.
2007b). They also provide habitat and food for a range of terrestrial species, including a
diverse suite of woodland birds (Di Stefano 2002). This includes the vulnerable superb
parrot, Polytelis swainsonii, which requires large healthy river red gums for breeding

7
(Webster in Di Stefano 2002). Woodland birds, or bush-birds, use river floodplains and
riparian areas for habitat, foraging, breeding, and watering (Johnson et al. 2007). When
compared with other habitat types in the Murray catchment, river red gum forest had the
highest total abundance and species richness of woodland birds (Oliver and Parker 2006).
The Macquarie Marshes are in decline, as a result of a long history of river regulation
(Kingsford and Thomas 2004). River red gum decline is one of the many ways this decline
has manifested. Water diversions support a $400 million agricultural industry, of which
cotton is the dominant irrigation enterprise (DECC 2009). Burrendong Dam, the major
storage structure in the system, was constructed in 1967. Since that time, the frequency,
duration and extent of floods have decreased significantly, while some channels are now
subject to constant low flows. This has significantly impacted upon the ecology of the
Macquarie Marshes. Much of the semi-permanent wetland mapped in the early 1990s no
longer supports wetland vegetation (DECC 2009), and waterbirds have been adversely
affected (Kingsford and Thomas 1995). In addition to river regulation, rainfall and flows have
been extremely low over the past decade. Little information exists about the relationship
between vegetation and woodland birds in the Macquarie Marshes (DECC 2009).
The aim of this study was to assess the impact of river red gum decline in the Macquarie
Marshes on the woodland bird community. Given the importance of river red gum to
woodland birds, we predicted that river red gum forests in poor ecological health would
support a different bird community to river red gums in good ecological health. We
investigated the extent and nature of these changes, through different measures of the
woodland bird community and river red gum community and related these to changed
flooding regimes.

8
3 Study area The Macquarie Marshes is a large floodplain wetland at the lower end of the Macquarie
River (Figure 1), covering an area of >200 000 ha, about 10% of which is within a Nature
Reserve. The Marshes are listed under the Ramsar Convention as a wetland of international
importance and are also listed by the National Trust of Australia and Australian Heritage
Commission as having conservation importance(DECC 2009). There is a complex system of
branched channels conveying water to a range of wetland types, ranging from lagoons and
semi-permanent marshes to permanent wetlands, providing essential habitat for a large
range of biota (DECC 2009). A total of 76 waterbird species have been recorded in the
Macquarie Marshes, and some of the larger waterbird breeding events in Australia have
been recorded there (DECC 2009). River red gums are an important habitat for a diverse
suite of woodland birds (over 120 species) in the Macquarie Marshes (Appendix 1). Eighteen
of these species are declining (Reid in DECC 2009), and four are listed as vulnerable under
the NSW Threatened Species Conservation Act 1995: the brown treecreeper, diamond
firetail, hooded robin and grey-crowned babbler.
The Macquarie Marshes have a diverse mosaic of vegetation types created by variable flows
and a complex network of channels. These include river red gum woodlands, which are
typically on floodplain clays close to watercourses, with blackbox (Eucalyptus largiflorens)
and coolibah (E. coolabah) further out (Shelly 2005). Other major vegetation communities
include cumbungi (Typha domingensis), water couch (Paspalum distichum), lignum
(Muehlenbeckia florulenta) and common reed (Phragmites australis). The understorey is
variable, from aquatic species, reeds, rushes and sedges in the wetter parts, to grasses and
forbs in dry areas (DECC 2009). River red gums in the Macquarie Marshes cover an area of
40 000 ha (Wilson in DECC 2009), constituting the largest northern area of river red gums
(EPA 1995). Already affected by clearing and ring-barking early in the 20th century (Brander
1987), large areas of river red gums have declined in health recently, due to changes in the
flood regime (Bacon 1994; Nairn 2008). One third of river red gum sites in the northern part
of the Macquarie Marshes have died because of river regulation, abstraction and drought
(Steinfeld and Kingsford 2008). Some of these areas only began to show signs of stress in
2001 (R. Jones, NPW, pers. comm.).

9
4 Methodology
4.1 Health categorisation
This project focused specifically on woodland bird communities in river red gum woodlands
within the Northern Macquarie Marshes Nature Reserve, an area where effects of livestock
grazing would not confound research results. Areas of river red gums were identified from
existing vegetation survey data (Wilson 2008) and then divided into three categories of
condition: poor, intermediate and good. This was achieved by visual assessment of digital
aerial photos of the Northern Nature Reserve (resolution 40 cm; 2006) which had been
rectified and georectified (projection GDA94, MGA zone 55). Visual interpretation was
preferred over automated classification as it allowed for use of texture, shape and pattern
of vegetation (Danby and Hik 2007). We created polygons around areas of river red gums in
similar condition, based on similarities in canopy cover, texture and colour. To classify
health we then generated random points within these polygons, and estimated the
percentage of dead or extremely stressed trees, identified by their lack of canopy, within a
50m circle of each point (Figure 2). Random points (n=100 total) were generated using
Hawth’s Tools (Beyer 2004) in ArcGIS (ESRI 2006). Polygons were classified into one of three
condition categories based on the number of dead or extremely stressed trees at the points
within each polygon: good 0-10%, intermediate 20-70%, poor 80-100 %. The result of this
process was a map of the three categories of river red gum health within the Macquarie
Marshes Northern Nature Reserve (Figure 3). Existing vegetation maps (Wilson 2008) were
checked to ensure tree density was standardised across the three categories.
4.2 Site selection
We randomly selected 33 sites (10 good, 10 intermediate, 13 poor) within the three
categories. We created a 500m buffer around all access roads, within which we used
Hawth’s tools (Beyer 2004) to generate random points in the three health categories. This
buffer ensured accessibility and enabled me to survey woodland birds at three sites each
morning. The unbalanced design (Figure 4) reflected the size and accessibility of the
respective areas. We ensured that the minimum distance between sites was 500m to avoid
pseudoreplication. For comparison we also conducted bird surveys at three terrestrial sites,
located in ‘Ninnia’, part of the Macquarie Marshes nature reserve further out on the
floodplain. Power analysis using means from pilot surveys indicated that 10 sites of each
category would give a power > 0.8 to detect differences in both species richness and overall
abundance.

10
Figure 1 Site map showing the location of the Murray Darling Basin, Macquarie River catchment and Macquarie Marshes (Rayner 2009). The Macquarie Marshes Nature Reserve is shown in grey; this study was based in the northern nature reserve. ‘Ninnia’ is the smaller nature reserve area located to the east.

11
50 m a b
Figure 2 Examples of selected areas (50 m radius) assessed for percentage of dead trees. Figure a has 0 % dead, and so is classified as good, and figure b has 100 % dead or extremely stressed trees, identified by the spider-like forms and shadows of trees with bare branches, and so is classified as poor.

12
Figure 3 Map of the Macquarie Marshes Northern Nature Reserve showing areas of river red gums categorised by health and survey sites

13
4.3 Woodland Bird surveys
Using a repeated-measures design sites were surveyed in late autumn (May-June 2009,) and
early spring (August 2009). These periods were expected to coincide with non-breeding and
breeding periods respectively. Rainfall interrupted the non-breeding survey period, so
surveys were completed in two phases (May 5-10, June 15-19). We resurveyed three sites
surveyed in May to ensure that pooling these two phases was appropriate, and found no
significant difference in the bird community detected between these two phases (ANOSIM
global R=0.407, p=0.2). For logistical reasons, sites near each other were surveyed together,
but to minimise temporal bias, the order of visits was varied between sites and groups of
sites.
Woodland birds were surveyed at each site using the two-hectare search method (Barrett et
al. 2003). This is a standardised method for surveying woodland birds in Australia (eg. Oliver
and Parker 2006; Johnson et al. 2007; Martin and McIntye 2007). To standardise search
effort, we followed a repeatable modified line transect (marked with flags the day before)
through each site, along an inner rectangle, 25 m from the boundary, ensuring that the
entire sample area was covered. Line transects are often preferred over point counts as
they are efficient for data collection, more accurate, less susceptible to bias due to bird
movement, and easier to analyse (Bibby et al. 2000). Although this method may
underestimate the presence of cryptic species (Pyke and Recher 1984), it is effective for
detecting differences in species richness among habitats and broad differences in
community composition (Bibby et al. 2000). All birds heard and seen in a two-hectare site
(100x200m rectangle) during 20 minutes were recorded. Care was taken not to record birds
twice.
To test for differences in detectability, pilot surveys were conducted for one hour at one site
of each health category. Although more species were detected with more time spent at
each site, the rate of detection was similar at all health categories, indicating that
detectability did not differ (Figure 5).
Season (2)
Replication (10-13)
Health (3)
River red gums
Good
n=10
Autumn Spring
Intermediate
n=10
Autumn Spring
Poor
n=10
Autumn Spring
n=3
Autumn
Figure 4 Hierarchical surveying design for woodland bird and understorey surveying showing the two factors Health and Season. There were three health treatments (Good, Intermediate and Poor), and two surveying periods (autumn and spring). Ten sites each were surveyed within each health category. Each site was surveyed once in each survey period, with the exception of three poor sites, which were unable to be surveyed in spring due to time constraints (total n=63)

14
For each woodland bird observed, four variables of habitat use were recorded: 1) position in
the vegetation (on ground, in understorey, or in tree); 2) whether trees occupied were dead
or alive, and 3) position in tree (dead or live branch, trunk or foliage), 4) height of the bird in
occupied tree (0-5 m, 5-10 m, >10 m) and 5) the species of the tree. Bird activity at first
observation (feeding/foraging, perching, calling, preening, darting, flying or other) was also
recorded. Surveys were conducted in a 3 hour window beginning at dawn. Surveys were
only conducted in fine weather and low winds (maximum wind 8.4 km hr-1, measured at
each site) (Bibby et al. 2000). Two surveyors (myself and another) were used in autumn to
maximise sample sites coverage. To minimise inter-observer differences, the two observers
did training surveys in the area (Maron et al. 2005). Additionally, we tested for differences
between the two observers with three independent simultaneous surveys over five minutes,
and found no significant difference.
Figure 5 Species accumulation over time for the three health categories
4.4 Vegetation To investigate relationships between woodland birds and river red gum communities, we
surveyed understory structure and floristics as well as tree health at each site using a system
of small and large quadrats positioned along transects (Figure 5, Table 1). Different
organisms vary over different scales, necessitating the use of quadrats of varying sizes. The
variables measured using the different survey methods are summarised in Table 1.
Understorey was measured using 10 (2x10 m) quadrats spaced evenly along two transects.
The percentage cover of bare ground, leaf litter, and dominant vegetation were recorded.
All vegetation was identified and recorded, except species covering < 5% of each quadrat.
Some plants, such as Chenopodiaceae and Asteraceae were only identified to genus as this
gave sufficient information on plant form and habitat structure. These quadrats were also
0
2
4
6
8
10
12
14
16
18
20
0 20 40 60 80
Cu
mu
lati
ve s
pe
cie
s co
un
t
Time (min)
Good
Int
Poor

15
used to estimate understorey height, the abundance of coarse woody debris (CWD) (5-50
cm, > 50 cm) and river red gum seedlings, an understorey greenness index (1 = mostly dry,
i.e. 0 – 30 % green, 2 = 30 – 70 % green, 3 = 70 – 100 % green) and any flowering or fruiting.
Understorey surveys were conducted in both survey periods in order to investigate seasonal
changes. Another observer was used for 11 sites in autumn and so observer differences
were estimated with independent surveys of two sites in different condition. There was no
difference in understorey composition as assessed by the two observers (ANOSIM global R =
-0.064, p=0.85).
To obtain estimates of percentage live basal area, tree density and demographics, trees
were counted and identified in 10 (20 x 20 m) quadrats evenly spaced along transects
(Figure 5) in each survey site, recording size class by diameter at breast height (DBH): > 100
cm, 50–100 cm, 10-50 cm and 2–10 cm (Jansen and Robertson 2001b). Trees were also
classified as live (with green leaf growth and intact bark) or dead (no green leaves and bark
peeling away) (Cunningham et al. 2007a). Given that trees are long lived and there were no
unusual weather patterns between surveys, these data were only collected in autumn.
To measure the health of trees, ten trees were assessed at each site for nine characteristics:
crown vigour; canopy density (% projective cover; percentage of dead branches; size class
and height class (0-5 m, 5-10 m, 10-20 m, >20m); and the presence or absence of hollows,
mistletoe, flowering and insect damage. Crown vigour was the proportion of potential
crown size, based on existing branches by observing the tree from a distance and using a
visual aid (Figure 7). Along with percentage live basal density and plant area index, crown
vigour is one of the more reliable and objective measures of river red gum health
(Cunningham et al. 2007a). Trees were selected using an adaptation of the ‘zig-zag selection
method’ (Figure 8) (Hnatiuk et al. 2006). As the focus of this study was long-term health
decline, only trees with a DBH greater than ten centimetres were assessed, disregarding
saplings, which may not yet have been fully established (George et al. 2005). These surveys
were also only conducted in autumn.

16
Table 1 Summary of the variables measured using the two different quadrat sizes and individual tree assessments. Ten of each survey method were carried out at each site.
Survey method Variables recorded
2 x 10 m quadrat Percentage cover of understorey components Height of understorey covers Understorey green-ness Number of coarse woody debris (CWD)- small (5-50 cm) and large (> 50 cm) Number of river red gum saplings
20 x 20 m quadrat Number and species of live and dead trees of four different size classes determined by diameter at breast height (DBH): 2 - 10 cm, 10 - 50 cm, 50 – 100 cm, > 100 cm (Jansen and Robertson 2001b)
Individual tree assessment Canopy density Crown vigour Percentage of dead branches Presence of insect damage (> 25 % of leaves) Size (DBH) Height Presence of hollows Presence of mistletoe
20 m
2 m
20 m
10 m
Figure 6. Large quadrats (20x10m) to survey vegetation were spread evenly along the transect and understorey recorded in smaller 2x10m quadrats. The blackened circle represents the tree chosen for health assessment.

17
Figure 7 Visual aid for estimating crown vigour (DSE inSRA 2007).
Figure 8 The zig-zag method for selecting trees for assessment. After assessing one tree, the next must be ahead of and towards or on the other side of the transect (Hnatiuk et al. 2006). In addition for this study, successive trees were spaced at least 40 m apart.

18
4.5 Guilds
To analyse broad differences in community structure, woodland birds were grouped into
guilds (Wilson 1999).Guilds are often used to detect community shifts (Croonquist and
Brooks 1991; Fischer et al. 2007). These may be based on distribution according to
environmental conditions (Beta guilds) or resource type used, within communities (alpha
guilds) (Wilson 1999). For instance, birds may be classified into alpha guilds based upon
their food type and the substrate on which they forage (Fischer et al. 2007). On the other
hand, beta guilds may be formed by measuring certain characters of each species and using
a cluster analysis to form groups of species that occur together or have similar niches
(Wilson 1999). By grouping species into guilds and examining changes in guild structure we
can gain perspective on the functional resilience of communities (Fischer et al. 2007) and
the ecological processes behind community change (Roberts 1987). For the present study,
birds were classified into broad guilds on the basis of habitat use data collected during field
surveys (using cluster analysis, discussed further below).
4.6 Statistical analyses we tested for the effect of health and season on bird species richness, abundance and
diversity (Shannon diversity index, which takes into account relative abundances), and on
abundances of the 10 most common species and all listed threatened species, using a
repeated-measures one-way analysis of variance (ANOVA) (Table 2).Health and season were
both fixed factors with three and two levels respectively, and sites were the replicates. For
understorey variables, which were measured in both seasons, repeated measures ANOVAs
were run (Table 3), with site as a random factor nested within health and quadrats as
replicates. We ran one-way ANOVAs to test for differences in whole-site vegetation
variables between health categories (Table 4) and nested ANOVAs to test variables
measured only in one season but with quadrats as replicates (Table 5).Where there were
significant differences, we ran pairwise comparisons with Tukey’s post hoc tests. All
univariate analyses were run in SYSTAT v11.0. The variables tested under each ANOVA
design are summarised in Table 6.
In order to assess whether the red gum community was once in similar condition at all
categories, abundances of river red gums in the four size classes was tested, pooling live and
dead trees to indicate what the community may have been like in the absence of water
stress. Counts of live and dead trees were still used to calculate percentage live basal area.
The major measure of whole-site health used was the “Cunningham” index, which is a
composite of crown vigour, canopy density and percentage live basal area, each
contributing 0-5 points to produce a maximum health index of fifteen (Cunningham et al.
2007a). These scores are divided by Cunningham et al. into five health classes: good (12.1-
15), declining (9.1-12), poor (6.1-9), degraded (3.1-6) and severely degraded (0.2-3) (not to
be confused with the health categories derived from aerial photo analysis used in this
study).

19
we tested assumptions of independence, homogeneity of variances and normality. We
tested for spatial and temporal autocorrelation with matrix correlation tests, similar to
Mantel tests (Legendre and Legendre 1998), using the BEST function within BIOENV in
PRIMER (Clarke and Gorley 2006). This tested similarity matrices of geographic coordinates
(eastings and northings) against both bird community composition and Cunningham score
(Clarke and Gorley 2006). There was no significant spatial autocorrelation of the bird
community (Rho =0.083, p = 0.318). However, there was spatial autocorrelation of tree
health (Rho = 0.291, p = 0.01). This was probably due to variation from east to west, which is
to be expected, as most poor sites are in the east and most good sites are in the west of the
study area (Figure 3). Accordingly, a conservative significance value of p ≤ 0.025 was used
for vegetation variables.
We ran Levene’s tests (Levene 1960) to test for homogeneity of variances. Most variables
had homogeneous variances. Transformation (log or fourth root) successfully stabilised
some variances, but where it did not, we again used a conservative p value of 0.025 (Sokal
and Rohlf 1995), to decrease the probability of a Type 1 error. We used the Shapiro-Wilk
normality test to test for normality (Shapiro and Wilk 1965), and found that about half the
variables were normally distributed. Only some variances were successfully normalised by
transformation (Table 6). For other non-normal data, we continued analysis under the
assumption that ANOVA was robust to non-normality (Quinn and Keough 2002; Underwood
1997). The transformations used are summarised in Table 6.
We grouped bird species into habitat-use guilds based on occupancy in the vegetation, using
group-average hierarchical cluster analysis. First, we summed the number of individuals in
each microhabitat category (ground, understorey, live branch, dead branch of live tree,
branch of dead tree, trunk of live or dead tree, foliage) for each species, irrespective of site
or health category. This was converted to a proportion of the total observations, so that all
species could be analysed on the same scale. We then constructed a Bray-Curtis similarity
matrix, followed by cluster analysis. Group names reflected major microhabitat preferences
of their members. We used linear regression to test the relationship between the
abundance of the habitat use guilds and Cunningham score.
To test for changes in bird species’ abundances, habitat-use guilds and understorey
community, we used permutational multivariate analysis of variance (PERMANOVA).
PERMANOVA runs an ANOVA on the basis of any resemblance measure, using permutation
methods (Anderson 2001a; b). We used a repeated measures design (Table 2) in
PERMANOVA+ for PRIMER (Anderson and Gorley 2007), using Bray-Curtis similarity
matrices, which are suitable for species abundance data, which typically include many zeros
(Clarke and Gorley 2006). Data were fourth root transformed to reduce the influence of the
most abundant species (Clarke and Warwick 1994). PERMANOVA has no explicit
assumptions about the distribution of the original data, but assumes homogeneity of
dispersion (Anderson and Gorley 2007). So, if one health category has a more variable

20
community composition than the other categories, this violates this assumption. We tested
for homogeneity of dispersions using the PERMDISP routine in PERMANOVA+ (Anderson
and Gorley 2007).
For significant PERMANOVA results, we explored community change using similarity
percentages (SIMPER) in PRIMER to identify the species or guilds contributing most to
similarities within and dissimilarities between health categories . We used non-metric multi-
dimensional scaling to construct ordinations of the bird and understorey communities at
each site and Principal Component Analyses (Clarke and Gorley 2006) to represent the
differences in habitat structure among health categories and sites.
We also tested for correlations between the bird community, habitat structure, understorey
and flood history. Flood history was the frequency of flooding 2000-2006, and time since
last flood at 2006 . We used BEST analysis (part of BIO-ENV in PRIMER) on pairs of similarity
matrices (Table 7), which produced variables that best explained the patterns in the
dependent matrix. BEST analysis is based on permutation methods, and produces a Rho
statistic, which is a measure of the strength of the relationship between the two matrices,
similar to the R2 value of linear regression (Clarke and Gorley 2006). We also used linear
regression to test for relationships of tree health and guild abundances with flood frequency
and time since last flood.

21
Table 2 Repeated measures statistical design testing the effects of tree health (H), which has i=a levels where a=3 and season (T) which has j=b levels where b=2. All combinations were replicated n times (r=n). Factor H and T are fixed. Formulae to determine estimated means squares (E(MS)), denominators (Den.) and variance components are shown.
Source Multipliers E (MS) Den. df V.C. (%)
BETWEEN SUBJECTS i J r
1. Hi 0 - n δe2 + nδH
2 2 2 [MSH -MSe]/n
2. er(i) 1 - 1 δe2 27 MSe
WITHIN SUBJECTS i J r
3. Tj a 0 n δE2 + anδT
2 5 1 [MST –MSE]/an 4. H x Tij 0 0 n δE
2 + nδHT2 5 2 [MSTH –MSE]/n
5. Er[ij] 1 1 1 δE2 27 MSE
Table 3 Repeated measures statistical design testing the effects of tree health (H), which has i=a levels where a=3; site (S) which has j=b levels where b=10 and season (T) which has k=c levels where c=2. All combinations were replicated n times (r=n), where n=10. Factor H and T are fixed. Formulae to determine estimated means squares (E(MS)), denominators (Den.) and variance components are shown.
Source Multipliers E (MS) Den. df V.C. (%)
BETWEEN SUBJECTS i J k r
1. Hi 0 b - n δe2 + nδSH
2 +
bnδH2
2 2 [MSH –MSSH]/bn
2. S(H)j(i) 1 1 - n δe2 + nδSH
2 3 27 [MSSH-MSe]/bn
3. er[ij] 1 1 - 1 δe2 540 MSe
WITHIN SUBJECTS i J k r
4. Tk a B 0 n δE2 + abnδT
2 7 1 [MST –MSE]/abn 5. H x Tik 0 B 0 n δE
2 + bnδHT2 7 2 [MSTH –MSE]/bn
6. S(H) x Tj(i)k 1 0 0 n δE2 + nδSHT
2 7 27 [MSST –MSE]/bn
7. Er[ij] 1 1 1 δE2 540 MSE
Table 4 Statistical design testing the effects of tree health (H), which has i=a levels where a=3 and is a fixed factor. There are r=n replicates, where n=10. Formulae to determine estimated means squares (E(MS)), denominators (Den.) and variance components (VC) are shown.
Source Multipliers E (MS) Den. df V.C. (%) i R
1. Hi 0 N δe2
+ nδH
2 2 2 [MSH -MSe]/n
2. er[i] 1 1 δe2 27 MSe
Table 5 Nested statistical design testing the effects of tree health (H), which has i=a levels where a=3 and site (S), which has j=b levels where b=10. Health is a fixed factor and site is random There are r=n replicates, where n=10. Formulae to determine estimated means squares (E(MS)), denominators (Den.) and variance components (VC) are shown.
Source Multipliers E (MS) Den. df V.C. (%)
i j r 1. Hi 0 b n δe
2 + nδSH2 + bnδH
2 2 2 [MSH –MSSH]/bn
2. S(H) (i)j 1 1 n δe2 + nδSH
2 3 27 [MSsH –MSe]/n
3. er[ij] 1 1 1 δe2 540 MSe

22
Table 6 Summary of the variables tested under each ANOVA design and whether they had homogeneous variances (H.V.) and normal distributions (N.D.)
Analysis Variable/s H. V. (Y/N) N. D. (Y/N) Transformation
One-way analysis of
variance (Table 4)
Cunningham index Y Y None
Percentage live basal area Y N None
Tree abundance in each size class Y N Log*** Nested analysis of variance
Crown vigour N N** Log
Canopy density Y Y None Percentage dead branches N N None* Total number of trees N N None* Presence of hollows Y Y None Presence of red gum buds or fruit Y N None* Number of river red gum saplings Y N None* One-way repeated
measures(Table 2)
Bird abundance Y N** Log Bird species richness Y N None*
Bird diversity (Shannon Index) Y N None* White-plumed honeyeater abundance Y N None* Brown tree creeper abundance Y N None* Jacky winter abundance Y N None* Willie wagtail abundance Y N None* Fairy-wren abundance Y N None* Australian ringneck abundance Y N None* Grey shrike-thrush abundance Y N None* Magpie-lark abundance N N None* Red-rumped parrot abundance Y N None* Peaceful dove abundance N N Log Nested repeated measures (Table 3)
Understorey species richness N N None* Shrub cover N N None*
Bare ground Y N None* Leaf litter N N None* Herb cover N N None* Understorey height N N Log
*indicates that no transformation was successful in stabilising variances, **indicates that data became normal after transformation, *** indicates that data were transformed to reduce influence of outliers
Table 7 Summary of pairs of data matrices tested in BEST analysis
Explanatory matrix Dependent matrix
Tree health and habitat structure* Bird community
Tree health Understorey composition
Flood history Bird community Tree health and habitat structure*
*variables used in the tree health and habitat structure data set were: crown vigour, canopy density, percentage of dead branches, hollows, mistletoe, tree size, tree height, average understorey height, bare ground cover, leaf litter cover, shrub cover, herb cover and CWD (large and small)

23
5 Results
5.1 Birds
A total of 87 woodland bird species were observed (Appendix 1): 58 during systematic
surveys and the rest incidentally. White-plumed honeyeater was the most abundant,
recorded at 90 % of all 33 sites, making up 20 % of all birds in surveys. Jacky winter, brown
treecreeper and willie wagtail were recorded at > 70% of all sites and made up respectively
12%, 7% and 5% of total birds observed. A total of 44 species were observed at good sites,
37 at intermediate sites and 47 at poor sites. Maximum species richness (18) and abundance
(92) occurred at good sites. Species accumulation curves showed that the number of sites in
the survey was sufficient to detect most species of the bird community (Figure 9). There
were no differences in species richness, abundance or diversity indices among health
categories or between seasons (Table 8). For two hectare sites, mean species richness was 8
(± 2.8 SE, range 3-18); mean abundance 27 (± 1.9 SE, range 4- 92) and; mean diversity index
was 1.75 (±0.05SE, range 0.79-2.52).
Of the ten most abundant bird species, white-plumed honeyeater was the only species
where abundance significantly varied among different health categories of river red gum
and seasons (Table 9), declining from good to poor sites during spring but not autumn
(Figure 10). Jacky winter and red-rumped parrot abundances varied significantly with
season (Figure 10,Table 9). The vulnerable hooded robin (Appendix 1) was only recorded at
poor sites. The bird community was highly variable, with variation among sites within river
red gum health categories, accounting for >85% of the variation for all species except white-
plumed honeyeaters (Table 9).
Despite the lack of difference in univariate variables there were significant differences in
community composition among health categories and between seasons (PERMANOVA,
Table 10). This significance was due to the difference between good and poor sites (P (perm)
= 0.005), good and intermediate sites (P (perm) = 0.002), but not between poor and
intermediate sites (P (perm) = 0.157). Good sites formed the most distinct group in three
dimensional space, while intermediate and poor overlapped significantly (Figure 11). The
three terrestrial sites were more similar to intermediate and poor sites than to good sites
(Figure 11). Health category accounted for 10% of the variation between subjects, with
differences between sites within health categories making up the remaining 90%. Season
contributed only 3% of the within variation (Table 10).
The differences in the woodland bird community were significantly related to river red gum
tree health and habitat structure (BEST analysis; Rho = 0.405, p = 0.01). This relationship was
best explained by five vegetation variables: canopy density, bare ground, shrub cover,
percentage of dead branches and large coarse-woody debris. Tree height and crown vigour
also featured in combinations of variables that explained more than 35 % of the variation in
the woodland bird community. Bird community composition was also significantly related to
flood history (BEST analysis, Rho = 0.269, p = 0.037)

24
Figure 9 Accumulation of woodland bird species for the three health categories across the 10 sites surveyed in autumn and spring 2009, using randomised data.
Figure 10 Mean abundances (±SE ) of white-plumed honeyeaters among different health groups of river red gum in spring (top left) and autumn (top right), and jacky winters and red-rumped parrots between seasons
0
5
10
15
20
25
30
35
40
45
50
0 5 10 15 20
Spe
cie
s
Sites
Good
Int
Poor
Good Int Poor
HEALTH
0
10
20
30
AW
HIT
EP
LU
ME
Good Int Poor
HEALTH
0
1
2
3
4
5
6
7
8
9
MW
HIT
EP
LU
ME
Autumn Spring
SEASON
0
5
10
15
JA
CK
YW
INT
ER
Autumn Spring
SEASON
0
1
2
3
4
5
6
7
8
9
RE
DR
UM
PE
DP

25
Table 8 Repeated measures ANOVA results for bird abundance, species richness, and Shannon diversity index, showing variance components (V.C.) Factors were river red gum health with three levels (poor, intermediate and food) and season (autumn and spring). Signficant (p<0.05) results in bold.
Variable Source df MS F P V.C. (%)
Bird Abundance (log transformed)
BETWEEN SUBJECTS
Health 2 0.057 0.330 0.722 0
Error 27 0.173 100
WITHIN SUBJECTS
Season 1 1.566 3.778 0.062 7.95
Health x Season 2 0.107 0.259 0.773 6.35
Residual 27 0.414 85.70
Total 54
Species BETWEEN SUBJECTS richness Health 2 5.417 0.563 0.576 0 Site(Health) 27 9.626 100 WITHIN SUBJECTS Season 1 0.067 0.009 0.924 3.02 Health x Season 2 2.817 0.391 5.56 Residual 27 7.196 91.42 Total 54
Shannon BETWEEN SUBJECTS diversity Health 2 0.166 0.939 0.403 0 Site(Health) 27 0.177 100 WITHIN SUBJECTS 1 0.080 0.687 0.415 1.04 Season 2 0.123 1.048 0.364 0.50 Health x Season 27 0.117 98.46 Residual 54 Total
Table 9 Variance components (%) of factors health (H) Season (T) and residuals (e) from repeated measures ANOVAs for the ten most abundant bird species. Significant results are indicated: * (p < 0.05), ** (p < 0.01) River red gum health had three levels (poor, intermediate and food) and season had two (autumn and spring).
Source Au
stra
lian
Rin
gne
ck
Fair
y W
ren
Mag
pie
lark
Wh
ite
-
plu
me
d
ho
ne
yeat
er
Pe
ace
ful
Do
ve
Will
ie
wag
tail
Jack
y
win
ter
Bro
wn
tre
ecr
ee
per
Gre
y sh
rike
-
thru
sh
Re
d-
rum
pe
d
par
rot
BETWEEN SUBJECTS
H 0 12.2 12.9 35.2* 0 0.8 13.1 0 0 7.2
e 100 87.8 87.1 64.8 100 99.2 86.9 100 100 92.8
WITHIN SUBJECTS T 1.0 2.9 0.7 33.3** 3.2 2.9 17.5* 0.8 2.4 13.4*
T x H 0.5 7.8 4.4 22.6* 0.9 4.9 3.1 12.6 7.2 5.5
e 98.5 89.3 94.9 44.1 95.9 92.1 79.4 86.6 90.4 81.1

26
Figure 11 The distribution of study sites of woodland bird communities composition in three categories of river red gum health (poor, intermediate and good) as well as terrestrial sites, defined by multidimensional scaling (MDS) in three dimensional ordination space . The bottom graph is rotated 90 degrees to show a view based on a third axis of ordination space.

27
Table 10 Results of PERMANOVA based on bird community composition, relative to river red gum health , with three levels (poor, intermediate and good) and season, with two levels (autumn and spring).
Source
df MS Pseudo-F P (perm) Unique perms
Variance component (%)
BETWEEN SUBJECTS Health 2 5802.8 2.1538 0.001 998 10.3 Error 27 2694.2 89.7
WITHIN SUBJECTS Season 1 3598.3 2.074 0.033 999 3.4
Health x Season 2 1705.4 0.98295 0.48 998 0.2
Residual 27 1735 96.4
Total 59
There were a similar number of species contributing to most (90%) of the similarity within
good (10 species), intermediate (9 species) and poor (9 species) health categories of river
red gum (Table 11). Of these, eight species were common to all categories: white-plumed
honeyeater, jacky winter, willie wagtail, brown treecreeper, Australian ringneck, grey shrike-
thrush, fairy-wrens (unidentified and superb) and magpie lark. Eastern yellow robin and red-
rumped parrot also contributed to similarities among assemblages at good sites, while
weebill and hooded robin contributed to similarities among assemblages at intermediate
and poor sites respectively. Thirty-three species accounted for 90% of the dissimilarity
between bird assemblages of good and poor sites of river red gum health (Table 11).
Crested shrike-tit, galah, red-winged parrot and little friarbird were nearly always found at
good sites. Contrastingly, hooded robin and weebill were mostly at poor sites, while
diamond dove and rufous songlark were found exclusively at poor sites (Table 11). The
species contributing most to dissimilarities between intermediate and good sites included
jacky winter and fairy wren, more abundant at intermediate sites, and white-plumed
honeyeater and brown treecreeper, more abundant at good sites (Table 11). Fairy wrens
were more abundant at intermediate than poor sites and the most significant contributor to
dissimilarity between these groups (Table 11).
To investigate habitat selection based on microhabitat preferences, we identified seven
habitat-use guilds with a cluster analysis of the bird species (Figure 12). Quail species
(ground specialists), fairy-wrens (understorey specialists) and brown treecreeper (trunk
specialists) formed single-taxon guilds related to the proportion of individuals observed in
different microhabitats (Table 12). There were 10 species that predominantly used the
foliage (foliage specialists): striated pardalote, white plumed honeyeater, weebill, yellow-
rumped thornbill, mistletoebird, yellow-throated miner, grey fantail, crested shrike tit, little
friarbird and rufous whistler (Figure 13, Table 12). The rufous whistler and little friarbird
were also grouped in this guild, with strong preference for both foliage and live branches
(Table 12). Most observations of eight species were in dead trees: diamond dove, yellow
thornbill, hooded robin, rufous songlark, southern whiteface, cockatiel, white-winged
chough and crested pigeon (Table 12). Contrastingly, galah, pied butcherbird, woodswallows

28
(masked and white-browed), grey butcherbirds and white-bellied cuckoo shrike showed a
strong preference for live branches (Table 12). The remaining species were generalists, with
no one microhabitat accounting for more than 51% of observations. Within this group,
magpie lark, restless flycatcher, willie wagtail and jacky winter were often observed on dead
branches irrespective of tree health; Australian ringneck, eastern yellow robin and grey
shrike-thrush were often on live branches; and red-winged parrot, black-faced cuckoo-
shrike, peaceful dove and red-rumped parrot were often in dead branches of live trees
(Table 12).
Table 11 Mean abundances (No. ±SE) and SIMPER contributions to similarities (%C) for all species contributing > 1% similarity within (first three columns) or dissimilarity (last three columns) between different health categories of river red gums (Good, Intermediate and Poor).*Fairy-wren includes superb, white-winged and variegated fairy-wrens.
Species Good Intermediate Poor GvsI GvsP IvsP No. (%C) No. (%C) No. (%C) %C %C %C
White-plumed honeyeater
8.05±1.7 (27.16) 4.30±1.0 (25.44) 2.61±0.6 (17.22) 5.80 6.16 5.66
Brown treecreeper 2.20±0.6 (18.29) 1.70±0.5 (6.71) 1.74±0.4 (13.38) 5.78 4.75 5.83 Willie wagtail 1.65±0.4 (12.79) 1.00±0.3 (10.57) 0.91±0.3 (6.08) 4.88 4.97 5.02 Eastern-yellow robin
0.85±0.3 (6.43) 0.25±0.2 0.17±0.1 4.43 4.29 2.19
Magpie lark 1.40±0.4 (6.38) 0.20±0.1 0.70±0.3 (3.15) 4.56 4.63 3.58 Australian ringneck 0.90±0.3 (6.07) 1.75±0.7 (4.48) 1.30±0.5 (7.21) 5.20 4.85 5.67 Fairy-wren* 1.10±0.4 (4.71) 3.85±1.1 (15.48) 2.17±0.6 (7.43) 6.46 5.29 6.74 Red-rumped parrot 1.90±0.6 (4.18) 0.65±0.3 0.74±0.5 4.51 4.28 3.12 Grey shrike-thrush 0.45±0.1 (3.87) 0.70±0.2 (3.86) 0.57±0.2 (5.21) 4.10 3.90 4.45 Jacky winter 1.30±0.5 (2.19) 3.85±0.9 (17.73) 4.26±0.1 (29.01) 7.05 7.81 5.77 Black-faced cuckoo-shrike
0.25±0.1 0.65±0.2 (4.83) 0.22±0.2 3.89 2.09 4.00
Weebill 0.05±0.05 1.55±0.9 (2.66) 0.43±0.3 3.46 1.66 4.13 Hooded robin 0.05±0.05 0.00 0.52±0.2 (2.44) 2.62 2.73 Peaceful dove 0.75±0.3 0.40±0.2 0.52±0.3 3.49 3.26 3.08 Crested pigeon 0.30±0.1 0.50±0.3 2.57±2.1 2.36 3.46 3.70 Rufous whistler 0.20±0.1 0.30±0.1 0.09±0.06 2.35 1.45 2.27 Quail sp. 0.15±0.1 0.35±0.2 0.22±0.2 2.26 1.45 2.32 Crested shrike-tit 0.40±0.2 0.00±0 0.09±0.1 2.10 2.20 Yellow-throated minor
0.25±0.2 0.15±0.1 0.13±0.13 2.04 1.81 1.45
Grey fantail 0.15±0.1 0.30±0.2 0.00 2.03 1.53 Pied butcherbird 0.05±0.05 0.35±0.2 0.13±0.1 1.89 1.06 2.37 Galah 0.25±0.2 0.30±0.2 0.04±0.04 1.86 1.17 1.55 Little friarbird 0.25±0.1 0.00 0.05±0.05 1.57 1.68 Grey butcherbird 0.10±0.06 0.15±0.1 0.05±0.05 1.46 0.94 1.22 Striated pardalote 0.15±0.1 0.10±0.1 0.26±0.2 1.22 1.82 1.96 Woodswallows (masked and white-browed)
2.00±2.0 0.00 0.65±0.6 1.20 1.64
Red-winged parrot 0.15±0.1 0.05±0.5 0.00 1.12 Australian raven 0.20±0.2 0.05±0.05 0.05±0.05 1.01 White-breasted woodswallow
0.30±0.3 0.00±0 0.00 1.48
Rufous songlark 0.00 0.10±0.1 0.48±0.2 2.43 2.92 Restless flycatcher 0.25±0.2 0.00 0.26±0.2 1.79 1.38 Diamond dove 0.00 0.00 0.57±0.4 1.02 1.11 Yellow thornbill 0.20±0.1 0.13±0.1 1.00

29
US
GS
DT
LB
TS
GL
FS
Microhabitat preference cluster analysis
Figure 12 Cluster analysis of 37 bird species by microhabitat preferences (frequency of observation in each microhabitat type as a proportion of all observations of that species), producing seven habitat-use guilds: generalists (GL), trunk specialist (TS), live branch specialists (LB), dead tree specialists (DT), foliage specialists (FS), understorey specialist (US) and ground specialist (GS).

Table 12 Percentage of observations of each species in the various microhabitats: ground (Gr), understorey (U), live branch (LB), dead branch (DB), trunk of live (LT) and dead (DT) trees, and foliage (F).
Species Live Tree Dead tree
Gr U LB DB LT F DB DT
Black-faced cuckoo shrike 0 0 31 50 0 0 19 0
Brown treecreeper 6 2 6 14 29 0 30 13
Cockatiel 0 0 0 0 0 0 100 0
Crested pigeon 9 0 3 17 3 0 68 0
Crested shrike -tit 0 0 50 0 0 50 0 0
Diamond dove 0 46 0 0 0 0 54 0
Eastern-yellow robin 0 8 35 23 4 12 15 4
Fairy wren- unid. and superb 5 74 5 0 5 1 7 3
Galah 0 0 88 13 0 0 0 0
Grey butcherbird 0 0 67 33 0 0 0 0
Grey fantail 0 0 44 0 0 56 0 0
Grey shrike-thrush 2 3 29 29 12 6 12 6
Hooded robin 0 0 0 0 0 0 67 33
Jacky winter 2 14 2 24 1 5 51 1
Little friarbird 0 0 50 17 0 33 0 0
Magpie lark 16 11 3 35 0 3 32 0
Mistletoebird 0 0 0 0 0 100 0 0
Peacful dove 23 0 12 47 0 0 18 0
Pied butcherbird 0 0 60 30 0 0 10 0
Quail sp. 100 0 0 0 0 0 0 0
Red-rumped parrot 18 0 21 40 8 4 8 0
Red-winged parrot 0 0 25 50 0 0 25 0
Restless flycatcher 0 27 18 27 0 0 27 0
Ringneck 5 11 26 33 0 5 18 3
Rufous songlark 0 0 8 8 0 0 85 0
Rufous whistler 0 8 42 8 0 42 0 0
Southern whiteface 0 0 0 0 0 0 100 0
Striated pardalote 0 0 0 0 0 75 25 0
Weebill 0 0 0 5 0 90 5 0
White plumed honeyeater 0 0 5 9 1 80 4 1
White-bellied cuckoo-shrike 0 0 67 33 0 0 0 0
White-winged chough 0 0 0 40 0 0 60 0
Willie wagtail 3 19 13 19 0 8 37 1
Woodswallow 0 0 73 25 0 0 0 2
Yellow rumped thornbill 0 0 0 0 0 100 0 0
Yellow thornbill 0 0 0 0 0 43 57 0
Yellow-throated minor 0 0 29 0 0 57 14 0

31
The relative abundances of habitat guilds differed significantly for the different river red gum
health categories but did not vary between seasons (Table 13, Figure 13). There were significant
differences between poor and good (P (perm) = 0.011), poor and intermediate (P (perm) =
0.016) but not between intermediate and good (P (perm) = 0.201). River red gum health and
season contributed 17.7% and 2.7% of the variation between and within subjects respectively.
Generalists, foliage specialists and live trunk/dead branch specialists contributed to 94% of the
similarity among good sites(Table 14). These same guilds also contributed to similarity within
the intermediate and poor categories, while dead tree specialists contributed to similarity
within poor sites, and understorey specialists to similarity within intermediate sites (Table 14).
Dead tree specialists and understorey specialists contributed most to the dissimilarities
between good and poor sites (Table 14). Foliage specialists were most abundant at good sites,
and least abundant at poor sites; this pattern was reversed for dead tree specialists (Figure 13,
Table 14). Abundances of generalists and live trunk specialists were relatively constant among
the three river red gum health categories, while live branch specialists were more abundant at
good sites (Figure 13, Table 14).
Abundance of foliage specialists was positively related to increasing health score (R2 = 0.524, p
< 0.001, Figure 14). Contrastingly, there was a significant negative relationship between dead
tree specialists and health score (R2 = 0.651, p < 0.001, Figure 14). Furthermore, there was a
significant relationship between flood frequency and both foliage specialists and dead tree
specialists (Figure 15). There were no significant relationships between Cunningham score and
abundances of generalists, live branch specialists, live trunk specialists or understorey
specialists.
Table 13 Results of PERMANOVA based on the bird community grouped into habitat-use guilds, showing variance components (V.C.) Factors were river red gum health with three levels (poor, intermediate and food) and season (autumn and spring). Signficant (p<0.05) results in bold.
Source df MS Pseudo-F P (perm) Unique perms
Variance component (%)
BETWEEN SUBJECTS Health 2 2400.7 3.15 0.006 999 17.7 Error 27 760.99 82.3
WITHIN SUBJECTS Season 1 892.99 1.93 0.157 998 2.7 Health x Season 2 271.3 0.59 0.718 8.7 Residual 27 12513 463.46 88.6 Total 54

32
Table 14 Average abundances (No.) and SIMPER contributions to similarities (%C) for all habitat-use guilds except understorey specialists, which did not contribute to similarities within or dissimilarities between groups.
Guild Good Int Poor G vs I G vs P I vs P
No. (%C) No. (% C) No. (% C) % C % C % C
Dead tree specialists
0.65 0.60 4.74 (12.82) 11.06 23.23 21.63
Foliage specialists
9.55 (36.94) 6.80 (38.13) 3.78 (25.39) 10.8 13.56 9.90
Generalists 9.85 (39.91) 9.50 (37.61) 9.65 (41.59) 12.04 8.26 8.82
Live branch specialists
2.50 0.85 0.91 15.71 13.42 13.61
Live trunk specialist
2.20 (17.16) 1.70 (6.41) 1.74 (11.51) 20.73 16.18 17.80
Understorey specialist
1.10 3.85 (12.93) 2.39 22.63 18.69 19.89
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Good Int Poor
Ave
rage
ab
un
dan
ce
HEALTH
Understorey specialists
Live trunk specialists
Live tree branch specialists
Ground specialists
Generalists
Foliage specialists
Dead tree specialists
Figure 13 Relative abundances of habitat-use guilds in the three health categories.

33
Figure 14 Linear regression of foliage specialists (Slope=0.201, R2=0.524) and dead tree specialists (Slope= - 0.291, R
2=0.424)
Figure 15 Linear regressions of foliage specialists and dead tree specialists score against flood frequency (years flooded from 2001-2006) (Slope = 0.145, R
2 = 0.226; Slope = - 0.217, R
2 = 0.222 respectively)
0 1 2 3 4 5 6 7 8 9
Cunningham Score
1
2
3
4F
olia
ge s
pecia
lists
(lo
g)
0 1 2 3 4 5 6 7 8 9
Cunningham Score
-1
0
1
2
3
Dead t
ree s
pecia
lists
(log)
0 1 2 3 4 5 6 7 8
Flood frequency
1
2
3
4
Folia
ge
spe
cia
liasts
(lo
g)
0 1 2 3 4 5 6 7 8
Flood frequency
-1
0
1
2
3
Dead t
ree s
pecia
lists
(lo
g)

34
5.2 Vegetation
There were no significant differences in abundances of large trees (50–100cm and >100cm
DBH) among health categories (Table 17).There were significant differences in the two smallest
size classes of river red gum, with trees of 10-50cm DBH most abundant at poor sites, and trees
of 2-10cm DBH most abundant at good sites (Table 17, Figure 18). Also, there was no significant
difference in the presence of tree hollows among river red gum health categories (F2,27 = 1.949,
p = 0.16),(Figure 19). There was negligible flowering of river red gums or their understorey in
any categories but the number of river red gums with fruits or buds differed significantly among
river red gum health categories (F2,27 = 6.105, p = 0.0006), with significantly more in good
compared to poor sites and a general downward trend (Figure 19). Mistletoe was only found at
four good sites and one intermediate site.
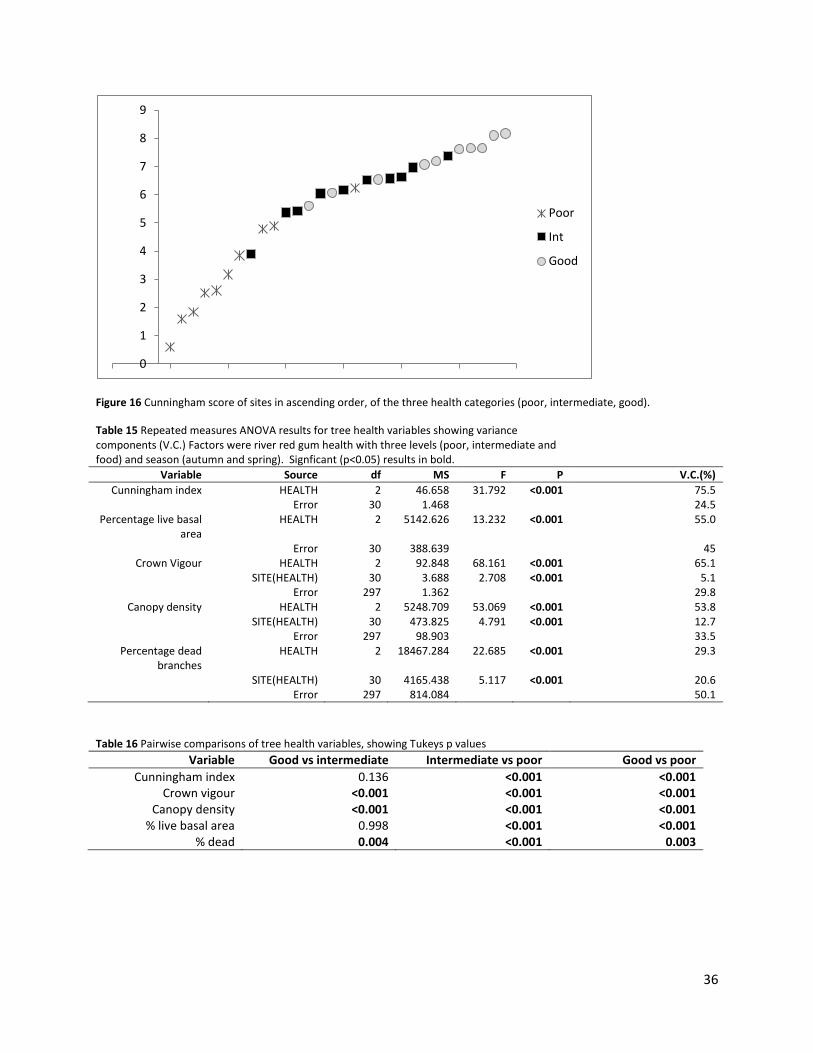
Cunningham score for river red gum health averaged 5.3 (± 0.4 SE, range 0.6-8.2). There was a
continuum of scores, with some overlap: nine out of the ten lowest scored sites were in the
poor category, and seven out of the ten best sites were in the good category (Figure 16).There
was a significant difference in the health score among river red gum health categories (Table
15), with higher scores in good compared to poor sites (Tukeys p < 0.001) and intermediate
compared to poor sites (Tukeys p < 0.001), but not between good and intermediate sites (p =
0.136). Health accounted for 75.5% of the variability in Cunningham scores among sites.
Percentage live basal area averaged 66 (± 4.6SE, range 4.2 - 97.2 %), with a significant
difference among river red gum health categories (Table 15). Again, this was higher in good
compared to poor, good compared to intermediate, but not between good and intermediate
(Table 16, Figure 17). Crown vigour, canopy density also differed among all river red gum
categories showing denser more full canopies at sites of better health (Table 15, Table 16). The
percentage of dead branches was also higher at poor sites (Table 15, Table 16).
Further, there were clear differences in habitat structure among river red gum health
categories with a two-dimensional PCA explaining 46.3 % of the variation among sites (Figure
19). The different river red gum health categories were differentiated along the PC1 axis, with
leaf litter, canopy density, understorey green-ness, canopy height and herb cover
corresponding to negative loadings on PC1, while bare ground, shrub cover and understorey
height had positive loadings (Table 18). Good sites were typified by tall trees with dense
canopy, more leaf litter and low green herbaceous understorey, while poor sites had small
trees with thin canopy, more bare ground and tall, dry, shrubby understorey (Figure 19, Figure
21, Figure 22, Figure 23). Flood history was significantly correlated with tree health variables
(BEST analysis, Rho = 0.337, p = 0.02). There was also a significant negative relationship
between the Cunningham index and time since last flood and flood frequency (Figure 24, Figure
15).

35
Twenty seven plant taxa were recorded with a minimum of five percent cover in at least one
quadrat (Appendix 2). Aquatic species were only found at good sites, and herbs were mainly
found at good sites (Figure 28). Larger water dependent plants (lignum and common reed)
were most abundant at good sites (Figure 28). These differences in overall understorey
structure are summarised in Figure 28. The genus Einadia, a generally low shrub, was the most
common species, found at 93 percent of sites. The next most common taxa were Sclerolaena,
Atriplex and Maireana (Figure 25). Bare ground and leaf litter were also significant components
of the ground cover at most sites. Five good sites had standing water during the autumn survey,
which had mostly subsided by spring. Understorey community composition varied significantly
between all pairs of river red gum health categories (Table 19, Figure 26). Understorey of good
sites was much more variable than intermediate and poor sites (PERMDISP average deviations
from centroid 37.48, 27.27 and 23.83 respectively). There was a significant correlation between
tree health variables and understorey composition (BEST Rho = 0.519, p = 0.001), best
explained by crown vigour and the presence of mistletoe. Four cover types, Einadia, bare
ground, leaf litter and Sclerolaena, contributed > 90% of the similarity among poor sites.
Contrastingly, six and seven cover types drove patterns at intermediate and good river red gum
health respectively (Table 20). Einadia, bare ground and leaf litter were common to all river red
gum health categories. Bare ground was more abundant at poor sites, while leaf litter was more
abundant at good sites. Lippia, clover and water contributed to similarities among good sites
and were not found at any poor sites while Maireana contributed only to intermediate, and
Atriplex to good and intermediate sites (Table 20). Sclerolaena was found at all poor and
intermediate sites but only at 20% of good sites. The same cover types were among a more
extensive list of species contributing to the dissimilarities among river red gum health
categories (Table 20). Atriplex, Maireana, Enchylaena and Chenopodium were most abundant
at intermediate sites (Table 20). Average and maximum understorey height differed
significantly among all river red gum health categories (F2,27 = 25.374, p = 0.001; F2,27 = 3.936, p
= 0.021) (Figure 27).
36
Figure 16 Cunningham score of sites in ascending order, of the three health categories (poor, intermediate, good).
Table 15 Repeated measures ANOVA results for tree health variables showing variance components (V.C.) Factors were river red gum health with three levels (poor, intermediate and food) and season (autumn and spring). Signficant (p<0.05) results in bold.
Variable Source df MS F P V.C.(%)
Cunningham index HEALTH 2 46.658 31.792 <0.001 75.5 Error 30 1.468 24.5
Percentage live basal area
HEALTH 2 5142.626 13.232 <0.001 55.0
Error 30 388.639 45 Crown Vigour HEALTH 2 92.848 68.161 <0.001 65.1
SITE(HEALTH) 30 3.688 2.708 <0.001 5.1 Error 297 1.362 29.8
Canopy density HEALTH 2 5248.709 53.069 <0.001 53.8 SITE(HEALTH) 30 473.825 4.791 <0.001 12.7 Error 297 98.903 33.5
Percentage dead branches
HEALTH 2 18467.284 22.685 <0.001 29.3
SITE(HEALTH) 30 4165.438 5.117 <0.001 20.6 Error 297 814.084 50.1
Table 16 Pairwise comparisons of tree health variables, showing Tukeys p values
Variable Good vs intermediate Intermediate vs poor Good vs poor
Cunningham index 0.136 <0.001 <0.001 Crown vigour <0.001 <0.001 <0.001
Canopy density <0.001 <0.001 <0.001 % live basal area 0.998 <0.001 <0.001
% dead 0.004 <0.001 0.003
0
1
2
3
4
5
6
7
8
9
Poor
Int
Good

Figure 17 Mean values (±SE) for tree health variables across the health categories.
Good Int Poor
HEALTH
0
10
20
30
40
50
60
70
80
90
100
Pe
rce
nta
ge
liv
e b
asa
l a
rea
Good Int Poor
HEALTH
0
10
20
30
40
Ca
nop
y d
en
sity
Good Int Poor
HEALTH
0
10
20
30
40
50
60
Cro
wn
vig
ou
r
Good Int Poor
HEALTH
0
1
2
3
4
5
6
7
8
9
Cu
nn
ing
ha
m s
co
re
Good Int Poor
HEALTH
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
Ho
llow
s

38
Table 17 ANOVA results for abundances of the four size classes of river red gum(R.R.G.) (dead and live trees have been pooled), showing variance components (V.C.) Health has three levels (good, intermediate and poor).
Source R.R.G. 2-10cm (log) R.R.G. 10-50cm (log) MS F p V.C. MS F P V.C.
Health 9.571 5.88 0.004 31.3 2.998 3.888 0.022 22.4 error 1.722 68.7 0.771 77.6
Source R.R.G. 50-100cm (log) R.R.G. >100cm (log) MS F p V.C. MS F P V.C.
Health 0.855 2.056 0.130 9.5 0.533 1.979 0.140 8.9 error 0.416 90.5 0.269 91.1
Figure 18 Mean abundances of trees in each of eight size classes by diameter at breast height (cm), both alive (L) and dead (D).
Figure 19 Average proportion of river red gums with hollows, mistletoe and fruit or buds by health category (good, intermediate and poor) (±SE)
0
5
10
15
20
25
Good Int Poor
Ave
rage
ab
un
dan
ce p
er
40
0sq
.m
HEALTH
D100+
L100+
D50-100
L50-100
D10-50
L10-50
D2-10
L2-10
Good Int Poor
HEALTH
0.0
0.1
0.2
0.3
0.4
0.5
0.6
Mis
tle
toe
Good Int Poor
HEALTH
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
Riv
er
red
gu
m f
ruits

39
Figure 20 PCA of sites based on habitat structure, showing structural changes with health decline along the PC1axis. PC1 and PC2 account for 46.3% of the variation between sites. The variables are bare ground (bare), canopy density (CD), understorey green-ness (Gr), insect damage (ID), herb cover (herb), leaf litter (litter), shrub cover (shrub), tree height (TH) and understorey height (UH)
Table 18 Eigenvector values of habitat structure measured by 12 variables against principal component axes one and two.
Variable PC1 PC2
Bare ground 0.322 -0.005 Leaf litter -0.324 -0.078 Shrub cover 0.440 -0.131 Understorey height 0.214 0.364 Canopy density -0.399 -0.242 Hollows -0.188 0.547 Small coarse woody debris 0.002 -0.227
Large coarse woody debris 0.069 0.056 Understorey green-ness -0.252 -0.396 Insect damage -0.022 -0.443 Canopy height -0.340 0.191 Herb cover -0.416 0.207

40
Figure 21 Examples of sites in the 'good' health category showing low green herb cover, leaf litter, and relatively dense canopy, providing habitat for foliage-specialist woodland birds.

41
Figure 22 Examples of sites in the 'intermediate' health category showing increasing shrub cover, thinning canopies.

42
Figure 23 Examples of sites in the 'poor' health category, showing dense shrubby understorey, and dead trees providing abundant perches for birds such as jacky winter, hooded robin.

43
Figure 24 Relationships between flood frequency (2001-2006) and Cunningham score (Slope = 0.628, R2 = 0.354), and time
since last flood (at 2006) (Slope = - 0.461, R2 = 0.322), showing regression lines.
Figure 25 Number of sites (maximum 20) at which understorey taxa were found, terrestrial (T) and flood dependent (F) in the three river red gum health categories (poor, intermediate, good).
0 1 2 3 4 5 6 7 8
Flood frequency
0
1
2
3
4
5
6
7
8
9C
un
nin
gh
am
Score
0 1 2 3 4 5 6 7
Time since last flood
0
1
2
3
4
5
6
7
8
9
Cun
nin
gh
am
Score
0
2
4
6
8
10
12
14
16
18
20
Ein
adia
(T)
Scle
rola
ena
(T)
Mai
rean
a (T
)
Atr
iple
x (T
)
Ench
ylae
na
(T)
Lign
um
(F)
Sals
ola
(T)
Spro
uts
Clo
ver
Sto
rksb
eak
(T)
Ree
ds
(F)
Ch
eno
po
diu
m(T
)
No
ogo
ora
Bu
rr (
T)
Dai
sy 2
(T)
Gra
ss (
T)
This
tle2
(T)
Lip
pia
(F)
Ran
un
culu
s (F
)
Flea
ban
e
This
tle
Per
sica
ria
dec
ipie
ns
(F)
Dai
sy1
(T)
Ch
aro
ph
yte
(F)
Dai
sy3
(T)
Nar
do
o (
F)
Nu
mb
er
of
site
s
Species
Good
Int
Poor

44
Table 19 PERMANOVA results based on understorey composition showing variance components (V.C.) Factors were river red gum health with three levels (poor, intermediate and food) and season (autumn and spring). Signficant (p<0.05) results in bold.
Source df MS Pseudo-F P (perm) Unique perms V.C. (%)
BETWEEN SUBJECTS
Health 2 42435 8.6819 0.001 999 35.7
Site(Health) 27 4887.7 7.72 0.001 998 4.0
Residual 270 633.13 60.2
WITHIN SUBJECTS
Season 1 10821 5.0162 0.004 999 1.5
Health x Season 2 4219.4 1.9559 0.066 999 1.2
Site (Health) x Season 27 2157.3 1.1301 0.168 997 0.1
Residual 569 1908.9 97.2
Total 599
Figure 26 MDS of sites based on understorey composition for both spring (s) and autumn (a) surveys. The distribution of study sites of woodland bird communities composition in three categories of river red gum health (poor, intermediate and good) in autumn and spring defined by multidimensional scaling (MDS) in three dimensional ordination space .

45
Table 20 Average abundances (N.) and SIMPER contributions to similarities (%C) for all understorey cover types contributing at least 1% similarity within groups or dissimilarity between river red gum health groups: good (G), intermediate (I) and poor (P)
Cover Good Intermediate Poor G vs I G vs P I vs P N (% C) N (% C) N (% C)
Leaf litter 37.05 (42.78) 33.38 (28.39) 16.72 (18.85) 7.39 11.48 15.65
Bare ground 13.83 (16.92) 18.35 (23.29) 18.58 (25.36) 8.88 8.70 6.42
Einadia sp. 12.83 (14.48) 26.68 (24.59) 41.83 (40.89) 12.06 15.16 11.86
Lippia 9.98 (7.69) 3.35 0.00 10.21 9.47 3.20
Atriplex sp. 3.58 (4.04) 1.57 (5.22) 1.35 6.61 6.14 9.30
Clover 4.80 (2.69) 3.62 0.85 4.41 3.48 3.15
Water 6.80 (2.65) 0.15 0.00 6.32 6.14
Sclerolaena sp. 0.25 2.24 (58.22) 12.72 (11.39) 9.71 11.75 13.17
Maireana sp. 0.65 1.95 (4.06) 0.89 4.33 3.02 6.42
Sprouts 1.90 1.23 0.48 4.38 3.67 5.17
Common reed 1.35 1.13 0.15 3.71 2.65 4.03
Chenopodium sp. 1.10 1.9 0.11 3.38 1.77 3.81
Salsola sp. 0.00 0.78 0.35 3.62
Enchylaena sp. 0.18 0.48 0.1 0.96 3.28
Lignum 0.45 0.00 0.5 1.77 2.55
Ranunculus sp. 4.10 0.53 0.00 5.75 4.89
Fleabane 0.375 0.15 0.00 2.57 2.03

46
Figure 27 Mean understorey height (±SE) among three river red gum health categories
Figure 28 Average cover of the main understorey components across the three health categories.
Good Int Poor
HEALTH
0
20
40
60
80
100
120U
nders
tore
y h
eig
ht
(m)
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Good Int Poor
Water dependant plants
Terrestrial shrubs
Herbs
Aquatic plants
Leaf litter
Bare ground
Un
der
sto
rey
hei
ght
(cm
)

47
6 Discussion River red gum decline in the Macquarie Marshes is causing a shift in the woodland bird
community composition, consistent with a ‘terrestrialisation’ of the system. Similar abundances
of large river red gum trees across all health categories suggest these woodlands were all once
in good condition, probably supporting similar woodland bird communities to those in good
sites. The changes in woodland bird community were probably linked to changes in the flooding
regime via effects on vegetation (BEST analysis, Figure 15, Figure 24), providing more evidence
for the effects of river regulation and highlighting the importance of floodplain health and
complexity. The dominance of terrestrial shrubs such as Sclerolaena sp. (Table 17) in the river
red gum forests provide further evidence that the ecosystem is becoming more terrestrial.
Simultaneously, the range of flood dependent understorey shrank, occupying only 10% of sites,
although this probably reflected the dry period.
The decline and death of river red gums significantly changed habitat structure for woodland
birds (Figure 19). If loss of structural complexity occurred, a critical factor for avian biodiversity
(Ford 1985b; Gilmore 1985; Recher 1985; Johnson et al. 2007), then species richness, diversity
and abundance should have decreased. It did not except through changes in community
composition (Table 8, Table 10, Figure 11). Rather, the habitat structure changed, favouring a
different community of woodland birds. As canopy cover, herb cover and leaf litter (an
important foraging substrate) decreased, dense terrestrial shrub cover and abundant dead
limbs increased (Figure 19) favouring species such as hooded robin, jacky winter, southern
whiteface, willie wagtail and white-winged chough (Table 11). These species use perches to
detect terrestrial predators and survey the ground layer for prey (Antos et al. 2008). Fairy-
wrens benefited from the invasion of terrestrial shrubs, although living trees and grassy areas
are also important habitat features for them (Parsons et al. 2008). This may explain why this
group was more abundant at intermediate sites, with live trees and terrestrial shrubs, rather
than at poor sites (Table 14, Figure 13). It is also likely that lignum, a flood dependent shrub
that is good habitat for fairy wrens, has decreased in abundance in the Macquarie Marshes.
These findings confirm the importance of managing for structural complexity and recognising
the value of all parts of the habitat (Shelly 2005).
Foliage specialist birds declined as river red gums lost their canopy (Figure 14). Similar patterns
are seen in terrestrial woodlands, with decreasing abundances of foliage-gleaning insectivores
correlated with declining tree health (Ford and Bell 1981; Ford 1985a; Gilmore 1985). River
regulation is probably contributing to declines already occurring in this group. Rufous whistler,
crested shrike-tit, grey fantail, weebill and striated pardalote are already declining in general
across the western-division of NSW (Smith et al. 1994). Live branch specialists such as white-
bellied cuckoo-shrikes and grey butcherbirds are also sensitive to river red gum decline but
there was another group of generalist woodland birds not sensitive to changing river red gum

48
health. This group included red-rumped parrot, peaceful dove, willie wagtail and magpie lark ,
which use a variety of different habitats and have already benefited from open habitat created
by agriculture and land clearing (Smith et al. 1994).
Some woodland bird species have specific habitat or foraging requirements which help to
explain their response to river red gum decline. Crested shrike-tits forage by probing and
gleaning insects from underneath bark (Smith 1985). When river red gums die they shed their
bark, explaining why crested shrike-tits were found almost exclusively at good sites, and only in
live trees. Mistletoe-birds were also only observed in live trees, and are constrained by the
distribution of mistletoe-bearing trees, found almost exclusively at sites in the good health
category. In general parrots were more abundant at good and intermediate sites, possibly
reflecting the higher abundance of river red gum fruits and other seeds. The lower fertility of
declining trees is also likely to be affecting nectarivores, with fewer flowering trees to supply
their high energy needs. Only five honeyeater species were detected in this study, out of a
recorded ten species for the area (Appendix 2); three of these were isolated sightings, and the
others were only observed feeding on insects, not nectar.
River red gum decline may threaten species already listed as threatened or vulnerable. The
vulnerable brown treecreeper was relatively unaffected by river red gum health, although
highest abundances occurred at good sites. Its distribution may be driven by other habitat
variables, such as the availability of large trees (Antos et al. 2008), which may be a long-term
problem where recruitment is low and dead trees fall over. River red gum sites in poor
condition were favoured by the vulnerable hooded robin, contrasting other work indicating an
association with large patches of healthy trees (Oliver and Parker 2006). The superb parrot,
observed incidentally and at one intermediate site, needs mature red gums next to box
woodlands (Ford et al. 2001) and large hollow-bearing trees for nesting (Webster 1998 in Di
Stefano 2002).
Further data could help better define these changes. In this study river red gum health was
treated as a categorical variable with inevitable overlap across the continuum, even though
there were significant differences in canopy health. More measures of this continuum would
help. Longer temporal analysis of woodland bird communities would also determine whether
patterns changed over seasons and years, improving the power to detect smaller differences in
species richness and abundance and enabling analysis of bird breeding, with tree fruiting and
flowering. There could be long term effects of decline in river red gums on woodland birds.
Many woodland birds are long-lived, so the full effects may not be realised, as decreased
breeding success may not be immediately apparent (Ford 1985b). A few nests were observed
incidentally during spring, all near good sites, indicating that poor sites may not provide suitable

49
nesting habitat. Although there was no difference in hollow bearing trees, important nesting
sites for many birds, this will change as dead trees burn, eventually fall over or break down.
Other impacts could also affect the woodland bird community; changes to groundwater access,
exotic species, grazing and prey abundance. River red gums access water from groundwater
resources and so identifying the role of groundwater in river red gum health is particularly
important (Bren 1988; MDBC 2003; Cunningham et al. 2007a). The high variability in tree health
at infrequently flooded sites may reflect different access to groundwater. Groundwater levels
of some of the areas of poor health have dropped over eight metres in the past decade (Jones,
R. NPW, pers. comm). Groundwater and surface-water are probably interconnected. The
potentially interactive effect of grazing may also be important; this study was only on non-
grazed areas. Grazing significantly alters understorey composition, possibly reducing the
structural complexity of vegetation (Jansen and Robertson 2001b), affecting many grazing-
sensitive bird species (Jansen and Robertson 2001a; Martin and McIntye 2007). Grazing could
amplify the effects of river red gum decline on woodland birds. Invasive species are also
changing woodland bird habitat. For example, lippia, an exotic spreading groundcover which
needs moist conditions, was found at 90% of all sites, yet it is unknown to what extent it
excludes native herbs and grasses or how its presence affects woodland birds. Finally, our
understanding of the mechanisms behind the shift in the bird community would be greatly
improved by information on invertebrate prey abundance at sites in different conditions.
Unfortunately, most river red gum forest and woodland areas in the Macquarie Marshes are
already degraded (Figure 16). Due to a lack of baseline data it was not possible to know the full
extent of changes, but woodland bird communities have probably changed throughout the
Macquarie Marshes. Out of 123 woodland bird species previously recorded in the Macquarie
Marshes (excluding nocturnal birds, see Appendix 1), this study detected 85, and one species,
the red-backed kingfisher, not previously recorded. Some of the species not recorded, such as
finches and chats, are more commonly found in more open areas, but it is possible that some
have declined. No sites were in truly good condition, as under the Cunningham system,
seventeen sites would actually be classified poor (6.1-9), eight degraded (3.1-6), and five
severely degraded (0.2-3). The low variability of understorey composition at poor sites
indicated that the Macquarie Marshes have lost the variability of vegetation that once
supported a diverse range of organisms. This study contributes to a growing body of knowledge
around the ecological decline of the Macquarie Marshes, a decline recently prompting the
Australian Government’s Environment Minister Peter Garrett to make a public statement about
‘a likely adverse change in the ecological character of the Macquarie Marshes Ramsar
Site’(Garrett and Tebbutt 2009).

50
Further evidence of a degraded system was the dominance of white-plumed honeyeater across
all sites (Ford et al. 2001). They can exclude small insectivorous birds especially if trees are
stressed and not flowering, aggressively defending lerp patches (Woinarski 1985). Most good
sites of river red gums were in small patches (8 – 65 ha) of relatively healthy trees. Only four
good sites were in a large healthy patch of forest (1,105 ha). Even relatively healthy areas were
probably suffering from increased edge effects, and associated habitat fragmentation. This
represents a further threat to some woodland bird species, due to increased risk of predation,
nest parasitism and decreased prey abundances with fragmentation (Loyn 1985; Zanette et al.
2000;Ford et al. 2001; Oliver and Parker 2006). For example, nesting success of eastern-yellow
robins is lower in small sites ( 55 ha), compared to large sites ( > 3400 ha) (Marchant 1985). My
results may underestimate changes to the woodland bird community, as more sites in truly
good condition would be needed for a full assessment.
There is widespread understanding of the need for environmental flows for aquatic ecosystems
and their dependent biota, including the Macquarie Marshes. We have shown that there
appears to be significant changes to the woodland birds as a result of decreased flows and
declining health of river red gums. The top priority should be to maintain core areas of river red
gums in good condition. They may need to be flooded every one to three years with inundation
lasting four to seven months (Roberts and Marston 2000). The next priority should be to flood
river red gums that are stressed but still alive to improve canopy health and maintain habitat
for foliage dependent bird species.
Significant progress has been made in recent years towards more sustainable river
management, but major impediments still exist. The introduction of environmental flows was a
major achievement, stepping away from the paradigm that water is wasted if it reaches lakes,
swamps or wetlands (Johnson 2005). The introduction of government buyback of water
licences is also positive. For example, 70 000ML have been bought from willing sellers for the
Macquarie Marshes (DECC 2009). Current levels of environmental flow allocations are only
sufficient to sustain parts of systems, forcing managers to make triage decisions, choosing
which elements to save at the cost of others (DECC 2009). These are complex social-ecological
systems (Johnson 2005), that can be hindered by conflicts of interest and institutional lethargy.
The findings of this study have broader conservation implications outside of the Macquarie
Marshes. Firstly, this study emphasises the importance of river red gum forests and riparian
forests in general for maintaining woodland bird diversity. Similar changes in the woodland
bird community are probably occurring in other areas where river red gum health is declining.
On the South Australian River Murray floodplain, about 80% of the trees are stressed, with 20-
30% being severely stressed (MDBC 2003). On the Victorian River Murray floodplain, only about
30% of river red gum stands are in good condition (Cunningham et al. 2006). Only on the

51
unregulated Lower Ovens River are most river red gum stands in good condition. As the extent
of river red gum health decline becomes evident, there is increasing public concern about the
health of these iconic trees (Ker 2009; Morton 2008; Plane 2003). Woodland birds are already
declining across agricultural south-east Australia, largely because of habitat loss, degradation
and fragmentation (Ford et al. 2001). Remnant riparian forests form critical habitat (Jansen and
Robertson 2001b), and so widespread river red gum decline poses a serious threat, especially
for foliage dependent species. Positively, recent moves have been made to protect remaining
areas of river red gum in national parks (Clennell 2009), but this does not address the issue of
water stress. Only when water diversion is reduced to sustainable levels will the maintenance
of river red gum woodlands, and their dependent woodland bird communities, and other
ecological values of river and floodplain systems become possible.

52
7 References ABS. (2008) Water and the Murray-Darling Basin- A Statistical Profile, 2000-01 to 2005-06. Australian Bureau of Statistics, Canberra.
Allan J. D. & Flecker A. S. (1993) Biodiversity Conservation in Running Waters. Bioscience 43, 32-43.
Anderson M. J. (2001a) A new method for non-parametric multivariate analysis of variance. Austral Ecology 26, 32-46.
Anderson M. J. (2001b) Permutation tests for uivariate or multivariate analysis of variance and regression. Canadian Journal of Fisheries and Aquatic Sciences 58, 626-39.
Anderson M. J. & Gorley R. N. (2007) PERMANOVA+ for primer: guide to statistical methods. PRIMER-E, Plymouth.
Antos M. J., Bennett A. F. & White J. G. (2008) Where exactly do ground-foraging birds forage? Foraging sites and microhabitat selection in temperate woodlands of southern Australia. Emu 108, 201-11.
Bacon P. E. (1994) The importance of water availability to river red gum (Eucalyptus camaldulensis) in the Macquarie Marshes. In: Proceedings of the Macquarie Marshes Workshop 1994 pp. 13-9. Macquarie Marshes Total Catchment Management Subcommittee, Dubbo.
Ballinger A. & Lake P. S. (2006) Energy and nutrient fluxes from rivers and streams into terrestrial food webs. Marine and Freshwater Research 57, 15-28.
Barrett G., Silcocks A., Barry S., Cunningham R. B. & Poulter R. (2003) The new atlas of Australian birds. Royal Australasian Ornithologists Union, Hawthorn East, Vic.
Bastow J. L., Sabo J. L., Finlay J. C. & Power M. E. (2002) A basal aquatic-terrestrial trophic link in rivers: algal subsidies via shore-dwelling grasshoppers. Oecologia 131, 261-8.
Beyer H. (2004) Hawth's Analysis Tools for ArcGIS. Available at http://www.spatialecology.com/htools.
Bibby C., Burgess N., Hill D. & Mustoe S. (2000) Bird Census Techniques. Academic Press, London.
Boulton A. J. & Lloyd L. N. (1992) Flooding frequency and invertebrate emergence from dry floodplain sediments of the River Murray, Australia. Regulated Rivers-Research & Management 7, 137-51.
Brander D. (1987) Environmental changes in the southern Macquarie Marshes: 1934-1987. BSc (Applied Geography) thesis, University of NSW, Sydney.
Bren L. J. (1988) Effects of river regulation on flooding of a riparian red gum forest on the River Murray, Australia. Regulated Rivers-Research & Management 2, 65-77.
Bren L. J. & Gibbs N. L. (1986) Relationships between Flood Frequency, Vegetation and Topography in a River Red Gum Forest. Australian Forest Research 16, 357-70.
Briggs S. V., Thornton S. A. & Lawler W. G. (1997) Relationships between hydrological control of River Red Gum wetlands and waterbird breeding. Emu 97, 31-42.

53
Brock M. A., Capon S. J. & Porter J. L. (2006) Disturbance of plant communities dependent on desert rivers. In: Ecology of Desert Rivers (ed R. T. Kingsford). Cambridge University Press, Cambridge.
Bunn S. E., Balcombe S. R., Davies P. M., Fellows C. S. & McKenzie-Smith F. J. (2006) Aquatic productivity and food webs of desert river ecosystems. In: Ecology of Desert Rivers (ed R. T. Kingsford). Cambridge University Press, Cambridge.
Chan E., Yu Y., Zhang Y. & Dudgeon D. (2008) Distribution patterns of birds and insect prey in a tropical riparian forest. Biotropica 40, 623-9.
Chao B. F. (1995) Anthropogenic impact on global geodynamics due to reservoir water impoundment. Geophysical research letters 22, 3529-32.
Clarke K. R. & Gorley R. N. (2006) PRIMER v6: User Manual/ Tutorial. Plymouth Marine Laboratory, PRIMER-E Ltd, Plymouth.
Clarke K. R. & Warwick R. M. (1994) Change in Marine Communities: an Approach to Statistical Analysis and Interpretation'. Natural Environment Research Council, U. K.
Clennell A. (2009) National park plan to save red gums, gain green votes. In: The Sydney Morning Herald. John Fairfax Holdings, Sydney.
Croonquist M. J. & Brooks R. P. (1991) Use of Avian and Mammalian Guilds as Indicators of Cumulative Impacts in Riparian-Wetland Areas. Environmental Management 15, 701-14.
Cunningham R. B., Mac Nally R., White J. G., Read J., Baker P., Thomson J. & Griffioen P. (2006) Mapping the current condition of river red gum (Eucalyptus camaldulensis Dehnh.) stands along the Victorian Murray River floodplain: A report to the northern Victorian Catchment Management Authorities and the Department of Sustainability and Environment. Australian Centre for Biodiversity, Melbourne.
Cunningham S., Read J., Baker P. & Mac Nally R. (2007) Quantitative assessment of stand condition and its relationship to physiological stress in stands of Eucalyptus camaldulensis (Myrtaceae). Australian Journal of Botany 55, 692-9.
Danby R. & Hik D. (2007) Evidence of Recent Treeline Dynamics in Southwest Yukon from Aerial Photographs. Arctic 60, 411-20.
DECC. (2009) Draft Macquarie Marshes Adaptive Environmental Management Plan. Department of Environment and Climate Change, Sydney.
Di Stefano J. (2002) River red gum (Eucalyptus camaldulensis): a review of ecosystem processes, seedling regeneration and silvicultural practice. Australian Forestry 65, 14-22.
EPA. (1995) New South Wales State of the Environment 1995. New South Wales Environmental Protection Authority, Sydney.
ESRI. (2006) ArcGIS version 9.2.

54
Fischer J., Lindenmayer D. B., Blomberg S. P., Montague-Drake R., Felton A. & Stein J. A. (2007) Functional Richness and Relative Resilience of Bird Communities in Regions with Different Land Use Intensities. Ecosystems 10, 964-74.
Ford H. A. (1985a) The bird community in eucalypt woodlands and eucalypt dieback in the northern tablelands of New South Wales. In: Birds of Eucalypt Forests and Woodlands: Ecology, Conservation, Management (eds A. Keast, H. Recher, H. A. Ford and D. A. Saunders). Surrey Beatty and Sons, Chipping Norton, NSW.
Ford H. A. (1985b) A synthesis of the foraging ecology and behavious of birds in eucalypt forests and woodlands. In: Birds of Eucalypt Forests and Woodlands: Ecology, Conservation, Management (eds A. Keast, H. Recher, H. A. Ford and D. A. Saunders). Surrey Beatty and Sons, Chipping Norton, NSW.
Ford H. A., Barrett G. W., Saunders D. A. & Recher H. F. (2001) Why have birds in the woodlands of Southern Australia declined? Biological Conservation 97, 71-88.
Ford H. A. & Bell H. (1981) Density of birds in eucalypt woodland affected to varying degrees by dieback. Emu 81, 202-8.
Garrett P. & Tebbutt C. (2009) Joint press release: planning for the future of the Macquarie Marshes. http://www.environment.gov.au/minister/garrett/2009/mr20090820.html accessed 10/9/2008.
Gehrke P. C., Brown P., Schiller C. B., Moffatt D. B. & Bruce A. M. (1995) River-regulation and fish communities in the Murray-Darling River system, Australia. Regulated Rivers-Research & Management 11, 363-75.
Gende S. M. & Willson M. F. (2001) Passerine densities in riparian forests of southeast Alaska: Potential effects of anadromous spawning salmon. Condor 103, 624-9.
George A., Walker K. F. & Lewis M. (2005) Population status of eucalypt trees on the River Murray floodplain, South Australia. River Research and Applications 21, 271-82.
Gilmore A. M. (1985) The influence of vegetation structure on the density of insectivorous birds. In: Birds of Eucalypt Forests and Woodlands: Ecology, Conservation, Management (eds A. Keast, H. Recher, H. A. Ford and D. A. Saunders). Surrey Beatty and Sons, Chipping Norton, NSW.
Gray L. J. (1993) Response of insectivorous birds to emerging aquatic insects in riparian habitats of a tallgrass prairie stream. Am. Midl. Nat. 129, 288-300.
Hnatiuk R. J., Thackway R. & Walker J. (2006) Vegetation Field Methods, Yellow Book. CSIRO.
Huxel G. R. & McCann K. (1998) Food web stability: The influence of trophic flows across habitats. Am. Nat. 152, 460-9.
Jansen A. & Robertson A. I. (2001a) Relationships between livestock management and the ecological condition of riparian habitats along an Australian floodplain river. J. Appl. Ecol. 38, 63-75.
Jansen A. & Robertson A. I. (2001b) Riparian bird communities in relation to land management practices in floodplain woodlands of south-eastern Australia. Biological Conservation 100, 173-85.

55
Johnson M., Reich P. & Mac Nally R. (2007) Bird assemblages of a fragmented agricultural landscape and the relative importance of vegetation structure and landscape pattern. Wildlife Research 34, 185-93.
Johnson W. J. (2005) Adaptive management of a complex social-ecological system: the regulated Macquarie River in south-eastern Australia. M. Res. Sci. Thesis. University of New England, Armidale.
Keith D. (2004) Ocean Shores to Desert Dunes: the native vegetation of New South Wales and the ACT. Department of Environment and Conservation, Hurstville NSW.
Ker P. (2009) $30m rescue bid for parched forest. In: The Age. John Fairfax Holdings, Melbourne.
Kingsford R. T. & Auld K. M. (2003) Waterbird breeding and environmental flow management in the Macquarie Marshes, Arid Australia. In: 9th International Symposium on Regulated Streams pp. 187-200, Albury, AUSTRALIA.
Kingsford R. T., Georges A. & Unmack P. J. (2006) Vertebrates of desert rivers: meeting the challenges of temporal and spatial unpredictability. In: Ecology of Desert Rivers (ed R. T. Kingsford). Cambridge University Press, Cambridge.
Kingsford R. T., Jenkins K. M. & Porter J. L. (2004) Imposed hydrological stability on lakes in arid Australia and effects on waterbirds. Ecology 85, 2478-92.
Kingsford R. T. & Johnson W. (1998) Impact of water diversions on colonially-nesting waterbirds in the Macquarie Marshes of arid Australia. Colonial Waterbirds 21, 159-70.
Kingsford R. T. & Thomas R. F. (1995) The Macquarie Marshes in arid Australia and their waterbirds- a 50 year history of decline. Environmental Management 19, 867-78.
Kingsford R. T. & Thomas R. F. (2004) Destruction of wetlands and waterbird populations by dams and irrigation on the Murrumbidgee River in arid Australia. Environmental Management 34, 383-96.
Klement T. & Jack A. S. (2002) Riverine flood plains: present state and future trends. Environmental Conservation 29, 308.
Lada H., Mac Nally R. & Taylor A. (2007) Genetic reconstruction of the population dynamics of a carnivorous marsupial (Antechinus flavipes) in response to floods. Molecular ecology 16, 2934-47.
Legendre P. & Legendre L. (1998) Numerical Ecology. Elsevier Science, Amsterdam.
Lemly A. D., Kingsford R. T. & Thompson J. R. (2000) Irrigated agriculture and wildlife conservation: Conflict on a global scale. Environmental Management 25, 485-512.
Levene H. (1960) A robust approximate confidence-interval for components of variance. Annals of Mathematical Statistics 31, 534-5.
Loyn R. H. (1985) Birds in fragmented forests in Gippsland, Victoria. In: Birds of eucalypt forests and woodlands: ecology, conservation, management (eds A. Keast, H. Recher, H. A. Ford and D. A. Saunders). Surrey Beatty and Sons, Chipping Norton, NSW.

56
Maheshwari B. L., Walker J. & McMahon T. A. (1995) Effects of river regulation on the flow regime of the River Murray, Australia. Regulated Rivers-Research & Management 10, 15-38.
Marchant S. (1985) Breeding of the eastern yellow robin. In: Birds of eucalypt forests and woodlands: ecology, conservation, management (eds A. Keast, H. Recher, H. A. Ford and D. A. Saunders). Surrey Beatty and Sons, Chipping Norton, NSW.
Maron M., Lill A., Watson D. M. & Mac Nally R. (2005) Temporal variation in bird assemblages: How representative is a one-year snapshot? Austral Ecology 30, 383-94.
Martin T. G. & McIntye S. (2007) Impacts of livestock grazing and tree clearing on birds of woodland and riparian habitats. Conserv. Biol. 21, 504-14.
MDBC. (2003) Preliminary investigations into observed river red gum decline along the River Murray below Euston. Technical Report 03/03. Murray Darling Basin Commission, Canberra.
Morton A. (2008) Action sought on river gums. In: The Age. John Fairfax Holdings, Melbourne.
Nairn L. (2008) Redgum Health in the Macquarie Marshes Nature Reserve. In: Wetlands and Rivers, University of New South Wales.
Nakano S. & Murakami M. (2001) Reciprocal subsidies: dynamic interdependence between terrestrial and aquatic food webs. Proceedings of the National Academy of Sciences of the United States of America 98, 166-70.
Nilsson C., Reidy C., Dynesius M. & Revenga C. (2005) Fragmentation and Flow Regulation of the World's Large River Systems. Science 308, 405-8.
Oliver D. L. & Parker D. G. (2006) Woodland birds of the NSW central Murray catchment: measuring outcomes of the Greening Australia fencing and tree planting program. NSW Department of Environment and Conservation, Queanbeyan.
Palmer G. C. & Bennett A. F. (2006) Riparian zones provide for distinct bird assemblages in forest mosaics of south-east Australia. Biological Conservation 130, 447-57.
Parsons H., French K. & Major R. E. (2008) The vegetation requirements of Superb Fairy-wrens (Malurus cyaneus) in non-urban edge and urbanised habitats. Emu 108, 283-91.
Plane T. (2003) The bare fact- iconic gums plainly hurting. In: The Australian. Nationwide News.
Polis G. A., Anderson W. B. & Holt R. D. (1997) Toward an integration of landscape and food web ecology: The dynamics of spatially subsidized food webs. Annual Review of Ecology and Systematics 28, 289-316.
Pyke G. & Recher H. (1984) Censusing of Australian birds: A summary of procedures and a scheme for standardisation of data presentation and storage. Department of Vertebrate Ecology, The Australian Museum.

57
Quinn G. P. & Keough M. J. (2002) Experimental design and data analysis for biologists. Cambridge University Press, Cambridge.
Rayner T. S., Jenkins K. M. & Kingsford R. T. (2009) Small environmental flows, drought and the role of refugia for freshwater fish in the Macquarie Marshes, arid Australia. Ecohydrology In press.
Reid M. A. & Brooks J. J. (2000) Detecting effects of environmental water allocations in wetlands of the Murray-Darling Basin, Australia. Regulated Rivers-Research & Management 16, 479-96.
Richardson D. M., Holmes P. M., Esler K. J., Galatowitsch S. M., Stromberg J. C., Kirkman S. P., Pysek P. & Hobbs R. J. (2007) Riparian vegetation: degradation, alien plant invasions, and restoration prospects. Diversity and Distributions 13, 126-39.
Roberts J. & Marston F. (2000) Water regime of wetland and floodplain plants in the Murray-Darling Basin: a sourcebook of ecological knowledge. CSIRO Land and Water, Canberra.
Roberts T. H. (1987) Construction of Guilds for Habitat Assessment. Environmental Management 11, 473-7.
Sabella A. (2009) Aquatic plant and macroinvertebrate community responses to extended dry periods in the Macquarie Marshes (honours thesis). School of Biological, Earth and Environmental Sciences, Faculty of Science UNSW, Sydney.
Sahagian D. (2000) Global physical effects of anthropogenic hydrological alterations: sea level and water redistribution. Global and Planetary Change 25, 39-48.
Shafroth P. B., Stromberg J. C. & Patten D. T. (2002) Riparian vegetation response to altered disturbance and stress regimes. Ecological Applications 12, 107-23.
Shapiro S. S. & Wilk M. B. (1965) An analysis of variance test for normality (complete samples). Biometrika 52.
Shelly D. (2005) Flora and Fauna of the Macquarie Marshes Region (Draft Report). Department of Infrastructure, Planning and Natural Resources, Dubbo.
Smith P. J. (1985) Woodchip logging and birds near Bega, NSW. In: Birds of eucalypt forests and woodlands: ecology, conservation, management (eds A. Keast, H. Recher, H. A. Ford and D. A. Saunders). Surrey Beatty and Sons, Chipping Norton.
Smith P. J., Pressey R. L. & Smith J. E. (1994) Birds of particular conservation concern in the western division of New South Wales. Biological Conservation 69, 315-38.
Sokal R. & Rohlf R. (1995) Biometry: the principles and practice of statistics in biological research. W. H. Freemand and company, New York.
SRA. (2007) The sustainable rivers audit vegetation theme level 2 assessment pilot study: field protocol (draft). Sustainable Rivers Audit.

58
Stauffer D. F. & Best L. B. (1980) Habitat selection by bird of riparian communities- evaluating effects of habitat alterarions Journal of Wildlife Management. 44, 1-15.
Steinfeld C. & Kingsford R. T. (2008) Floodplain development and vegetation health on the Macquarie River floodplain of the Murray-Darling Basin. School of Biological, Earth and Environmental Sciences. University of New South Wales, Sydney.
Thoms M. C. (2006) Variability, complexity and diversity: the geomorphology of river ecosystems in dryland regions. In: Ecology of Desert Rivers (ed R. T. Kingsford). Cambridge University Press, Cambridge.
Underwood. (1997) Experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge University Press, Cambridge.
UNEP-WCMC. (2001) Biodiversity and climate change. United Nations Environment Programme- World Conservation Monitoring Centre.
Walker K. F. (2006) Serial weirs, cumulative effects, the Lower River Murray, Australia. In: Ecology of Desert Rivers (ed R. T. Kingsford). Cambridge University Press, Cambridge.
Wilson J. B. (1999) Guilds, functional types and ecological groups. Oikos 86, 507-22.
Wilson R. (2008) Vegetation Mapping for the Macquarie Marshes, as part of the Wetland Recovery Program. Unpublished report for the the NSW Department of Environment and Climate Change.
Woinarski J. C. Z. (1985) Foliage-gleaners of the treetops, the pardalotes. In: Birds of eucalypt forests and woodlands: ecology, conservation, management (eds A. Keast, H. Recher, H. A. Ford and D. A. Saunders). Surrey Beatty and Sons, Chipping Norton, NSW.
Woinarski J. C. Z., Brock C., Armstrong M., Hempel C., Cheal D. & Brennan K. (2000) Bird distribution in riparian vegetation in the extensive natural landscape of Australia's tropical savanna: a broad-scale survey and analysis of a distributional data base. J. Biogeogr. 27, 843-68.
Zanette L., Doyle P. & Tremont S. M. (2000) Food shortage in small fragments: Evidence from an area-sensitive passerine. Ecology 81, 1654-66.

59
Appendix 1: Bird species list- Macquarie Marshes, May-August 2009 Order Family Common Name Scientific Name Spatial
guild Number observed
Casuariiformes Casuariidae 1. Emu Dromaius novaehollandiae
na i
Columbiformes Columbidae
2. Common Bronzewing
Phaps chalcoptera
na 1
3. Crested Pigeon Ocyphaps lophotes
Dead tree branch
75
4. Diamond Dove Geopelia cuneata Dead tree branch
13
5. Peacful Dove Geopelia placida Generalist 35
Coraciiformes Alcedinidae 6. Azure Kingfisher Alcedo azurea na 1
Halyconidae 7. Laughing Kookaburra
Dacelo novaguineae
na i
8. Red-backed Kingfisher
Todiramphus pyrrhopygia
na i
9. Sacred Kingfisher Todiramphus sanctus
na i
Cuculiformes Cuculidae 10. Fan-tailed Cuckoo
Cacomantis flabelliformis
na i
11. Horsfield's Bronze Cuckoo
Chalcites basalis na i
12. Pallid Cuckoo Cuculus pallidus na 1
Falconiformes Falconidae 13. Australian Hobby Falco longipennis na i
14. Brown Falcon Falco berigora na i
15. Nankeen Kestrel Falco cenchroides
na 2
16. Spotted harrier Circus assimilis na i
Accipitridae 17. Black-shouldered Kite
Elanus axillaris na i
18. Brown Goshawk Accipiter fasciatus
na i
19. Little Eagle Hieraaetus morphnoides
na i
20. Wedge-tailed Eagle
Aquila audax na i
21. 22. Whistling Kite Haliastur
sphenurus na i
23. White-bellied Sea-Eagle
Haliaeetus leucogaster
na i
Galliformes Phasianidae 24. Brown Quail Coturnix ypsilophora
Ground 6
Passeriformes Alaudidae 25. Rufous Songlark Cincloramphus mathewsi
Dead tree branch
13

60
Order Family Common Name Scientific Name Spatial guild
Number observed
Artamidae 26. Australian Magpie
Gymnorhina tibicien
na 4
27. Grey Butcherbird Cracticus torquatus
Live tree branch
6
28. Masked Woodswallow
Artamus personatus
Live tree branch
55*
29. Pied Butcherbird Cracticus nirgogularis
Live tree branch
11
30. White-breasted Woodswallow
Artamus leucorhynchus
na 2
31. White-browed Woodswallow
Artamus superciliosus
Live tree branch
55*
Campephagidae 32. Black-faced Cuckoo-shrike
Coracina novaehollandiae
Generalist 23
33. White-bellied Cuckoo-shrike
Coracina papuensis
Live tree branch
4
34. White-winged Triller
Lalage tricolor na 2
Climacteridae 35. Brown Treecreeper
Climacteris picumnus
Live trunk/ dead tree branch
118
Corcoracidae 36. Apostlebird Sruthidea cinerea na 0
37. White-winged Chough
Corcorax melanorhamphos
Dead tree branch
10
Corvidae 38. Australian Raven Corvus coronoides
na 6
Dicaeidae 39. Mistletoebird Dicaeum hirundinaceum
na 3
Passeriformes Dicruridae 40. Grey Fantail Rhipidura albiscapa
Foliage 9
41. Magpie Lark Grallina cyanoleuca
Generalist 48
42. Restless Flycatcher
Myiagra inquieta Generalist 11
43. Willie Wagtail Rhipidura leucophrys
Generalist 74
Estrildidae 44. Diamond Firetail Stagonopleura oculata
na i
Hirundinidae 45. Fairy Martin Petrochelidon ariel
na 5
46. Welcome Swallow
Hirundo neoxena na i
Maluridae 47. Superb Fairy-wren
Malurus cyaneus Understorey 149**
48. Variegated Fairy-wren
Malurus lamberti Understorey 3

61
Order Family Common Name Scientific Name Spatial guild
Number observed
49. White-winged Fairy-wren
Malurus leucopterus
Understorey 2
Meliphagidae 50. Blue-faced Honeyeater
Entomyzon cyanotis
na 2
51. Little Friarbird Philemon citreogularis
Foliage 6
52. Spiny-cheeked Honeyeater
Acanthagenys rufogularis
na i
53. Striped Honeyeater
Plectorhyncha lanceolata
na i
54. White-plumed Honeyeater
Lichenostomus penicillatus
Foliage 307
55. Yellow-throated Miner
Manorina flavigula
Foliage 11
Motacilladae 56. Australian Pipit Anthus australis na i
Pacycephalidae 57. Crested Shrike-tit Falcunculus frontatus
Foliage 10
58. Golden Whistler Pacychephala pectoralis
na i
59. Grey Shrike-thrush
Colluricincla harmonica
Generalist 36
60. Rufous Whistler Pacychephala rufiventris
Foliage 12
Pardalotidae 61. Buff-rumped Thornbill
Acanthiza reguloides
na 7
62. Chestnut-rumped Thornbill
Acanthiza uropygialis
na 2
63. Southern Whiteface
Aphelocephala leucopsis
Dead tree branch
3
64. Spotted Pardalote
Pardalotus punctatus
Foliage 2
65. Striated Pardalote
Pardalotus striatus
Foliage 11
66. Weebill Smicrornis brevirostris
Foliage 42
67. Yellow rumped thornbill
Acanthiza chrysorrhoa
Foliage 3
68. Yellow Thornbill Acanthiza nana Dead tree branch
7
Petroicidae 69. Eastern-yellow Robin
Eopsaltria australis
Generalist 26
70. Hooded Robin Melanodryas cucullata
Dead tree branch
13
71. Jacky Winter Micoeca fascinans
Generalist 201

62
Order Family Common Name Scientific Name Spatial guild
Number observed
72. Red-capped Robin
Petroica goodenovii
na 1
Pomatostomidae 73. Grey-Crowned Babbler
Pomatostomus temporalis
na i
Ptilonorhynchidae 74. Spotted Bowerbird
Chlamydera maculata
na i
Sylviidae 75. Australian Reed-Warbler
Acrocephalus australis
na i
76. Little Grassbird Megalurus gramineus
na i
Psittaciformes Cacatuidae 77. Cockatiel Nymphicus hollandicus
na 7
78. Galah Eolophus roseicapillus
Live branch 12
Psittacidae 79. Australian Ringneck
Barnardius zonarius
Generalist 83
80. Blue Bonnet Northiella haematogaster
na i
81. Parrot unid. na
82. Red rumped parrot
Psephotus haematonotus
Generalist 68
83. Red winged parrot
Aprosmictus erythropterus
Generalist 4
84. Superb parrot Polytelis swainsonii
na 2
Turnicidae Turniciformes 85. Quail sp. Ground 15
86. Painted button quail
Turnix varia Ground i
Table 21. Bird species information. ‘i’ indicates incidental observations made outside the dedicated surveys. *masked woodswallows and white-browed woodswallows were observed together in large mixed flocks, totalling 55 birds; **Unidentified fairy-wrens are grouped with superb fairy-wren.

The following woodland bird species have also been recorded at the Macquare Marshes, but were not
recorded on this study (list compiled from Australian Bird Atlas and NPWS atlas of NSW wildlife):
1. Stubble Quail Coturnix pectoralis
2. Bar-shouldered Dove Geopelia humeralis
3. Black Kite Milvus migrans
4. Collared Sparrowhawk Accipiter cirrocephalus
5. Swamp Harrier Circus approximans
6. Black Falcon Falco subniger
7. Peregrine Falcon Falco peregrinus
8. Brolga Grus rubicunda
9. Eastern Rosella Platycercus eximius
10. Pale-headed Rosella Platycercus adscitus
11. Budgerigar Melopsittacus undulatus
12. Blue-winged Parrot Neophema chrysostoma
13. Shining Bronze-Cuckoo Chalcites lucidus
14. Rainbow Bee-eater Merops ornatus
15. Splendid Fairy-wren Malurus splendens
16. Inland Thornbill Acanthiza apicalis
17. Black honeyeater Certhionyx niger
18. Black-chinned honeyeater (eastern subspecies) Melithreptus gularis gularis
19. Singing Honeyeater Lichenostomus virescens
20. Crimson Chat Epthianura tricolor
21. White-fronted Chat Epthianura albifrons
22. Noisy Friarbird Philemon corniculatus
23. White-browed Babbler Pomatostomus superciliosus
24. Chestnut-crowned Babbler Pomatostomus ruficeps
25. Ground Cuckoo-shrike Coracina maxima
26. Crested Bellbird Oreoica gutturalis
27. Olive-backed Oriole Oriolus sagittatus
28. Little Woodswallow Artamus minor
29. Horsfield Bushlark Mirafra javanica
30. Little Grassbird Megalurus gramineus
31. Brown Songlark Cincloramphus cruralis
32. Silvereye Zosterops lateralis
33. Common Blackbird Turdus merula
34. Common Starling Sturnus vulgaris
35. Zebra Finch Taeniopygia guttata
36. Double-barred Finch Taeniopygia bichenovii
37. Plum-headed Finch Neochmia modesta
38. House Sparrow Passer domesticus

64
Appendix 2: Understorey vegetation species list Scientific name Common name
Atriplex Saltbush
Chenopodium Goosefoot, crumbweed, Fat Hen
Conyza sp.* Fleabane
Einadia Saltbush, fishweed
Enchylaena tomentosa R.Br. Ruby saltbush
Erodium cicutarium (L.) L'Her. Ex Aiton* Common storksbill
Maireana Bluebush
Marsilea drummondii A. Braun Nardoo
Muehlenbeckia florulenta Meisn. Lignum
Onopordum acanthium L.* Scotch Thistle
Persicaria decipiens (R.Br.) K.L.Wilson Slender knotweed
Phragmites australis (Cav.) Trin. Ex Steud. Common reed
Phyla nodiflora (L.) Greene* Lippia
Ranunculus sp. Buttercup
Salsola
Sclerolaena Rolypoly, copperburr, poverty bush
Trifolium sp.* Clover
Xanthium occidentale Bertol.* Noogoora Burr
Chara sp. or Nitella sp. Charophytes
Daisy 1**
Daisy2**
Daisy3**
Grass**
Sprouts***
Thistle2**
*indicates introduced species
**indicates species that were not identified but given type names; these species were only
found at one or two sites
***’sprouts ‘ was the name given to small mixed germinating plants, generally in damp
depressions.

65
Appendix 3: Site locations
Name FID Longitude Latitude Health
Struggling Alley 0 147.569385 -30.648705 poor
Poo 2 147.590049 -30.698274 intermediate
Phoenix 4 147.563585 -30.675025 poor
Quail Heaven 6 147.516939 -30.633263 good
Afternoon Tea 9 147.516102 -30.640911 good
Scar Tree 11 147.517708 -30.647026 intermediate
Dinner Plate 12 147.511673 -30.649875 intermediate
Mosquitoe's Playground 18 147.518674 -30.669203 good
Waders Reach 20 147.531451 -30.670616 good
Red gum thicket 23 147.568786 -30.659263 poor
Gentle Giant 25 147.527154 -30.661266 good
Billabong Swans 27 147.532899 -30.658055 poor
Whoopsadaisy 28 147.534764 -30.670341 poor
Coolabah heaven 29 147.519894 -30.625708 good
No complaining 34 147.52622 -30.631488 intermediate
Rice Bubbles 39 147.574885 -30.655873 poor
Morning Tea 45 147.524032 -30.638971 intermediate
Lost binoculars 49 147.52141 -30.635422 good
Mob of goats 56 147.529617 -30.640828 intermediate
Not with a bang 58 147.524202 -30.64928 intermediate
Prickly legs 62 147.540576 -30.671985 poor
Contradiction 64 147.52351 -30.655543 poor
Dead Tree Gully 66 147.529668 -30.654738 poor
Dead Wood 67 147.592926 -30.659596 poor
Nutbush Thistle Limit 68 147.577066 -30.672239 poor
Atadwindi 69 147.571865 -30.637317 poor
Boomanditchydaisy 72 147.52012771700 -30.66441986550 good
Miss Piggy 76 147.51073473000 -30.69938432000 good
Enchanted Puddles 77 147.50802870700 -30.70462367880 good
SpeedyReedy 85 147.58658584300 -30.70445310950 intermediate
All knotted up 87 147.59264818200 -30.65415641490 poor
BogSprout 88 147.51462581800 -30.65731272220 intermediate
Hiccoughs 89 147.53213675000 -30.64754373780 intermediate