the effect of cellulolytic rumen bacteria or their cell
TRANSCRIPT

Western Kentucky UniversityTopSCHOLAR®
Masters Theses & Specialist Projects Graduate School
7-1987
The Effect of Cellulolytic Rumen Bacteria or TheirCell-Free Extracts on the Germination ofEuonymus americanus L.Gary Thomas Howard
Follow this and additional works at: http://digitalcommons.wku.edu/theses
Part of the Biology Commons
This Thesis is brought to you for free and open access by TopSCHOLAR®. It has been accepted for inclusion in Masters Theses & Specialist Projects byan authorized administrator of TopSCHOLAR®. For more information, please contact [email protected].
Recommended CitationHoward, Gary Thomas, "The Effect of Cellulolytic Rumen Bacteria or Their Cell-Free Extracts on the Germination of Euonymusamericanus L." (1987). Masters Theses & Specialist Projects. Paper 1431.http://digitalcommons.wku.edu/theses/1431

THE EFFECT OF CELLULOLYTIC RUMEN BACTERIA OR THEIR CELL-FREE
EXTRACTS ON THE GERMINATION OF EUQnymus americanus L.
A Thesis
Presented to
the Faculty of the Department of Biol ogy
western Kentucky University
Bowling Green, Kentucky
In Partial Fulfillment
of the Requirements for the Degree
Master of Science
by
Gary Thomas Howard
July, 1987

•
AUTHORIZATION FOR USE OF THESIS
PerlTlission is hereby
~ granted to the Western Kentucky Unive rsity Library to lTlake. or allow to be lTla e photocopies, lTlicrofilm Or other copies o f this the s is for appropriate research or scholarly purposes.
O reserved to the author fo'r the making of any copies of this thesis e xcept for brief sections for research or scholarly purposes.
Signed )1alll~ 1. iJtWlQAJ
Date !D JI.S L 87
P l e ase place an "X" in the appropriate; box.
Thi s form w ill be filed with the original of the thesis and will control fu t ure u s e of t h e thesi s.

THE EFFECT OF CELLULOLYTIC RUMEN BACTERIA OR THEIR CELL-FREE
EXTRACTS ON SEED GERM INATION OF ~nymua americanua L.
Dean of the Graduate lege
Recommended ((/~ a\) 1~~7 Date)

;
DEDICATION
Dedicated to my mother, Mildred Colling s Howard, and to
my father, Hurrol Thomas Howard, who took an active interest
in my work.
11
j

ACKNOWLEDGE ME NTS
I would like to thank Dr. Larry P. Elliott, chairman of
my adviso r y committee, for his guidance and encouragement in
directing me through this project. Thanks are also extended
to the other members of my committee , Drs . Martin R. Houston
an d Joe E. Winst ead.
Other individuals who also deserv e special thanks for
their assistance are Dr. Delmer Davis for his help in
collecting t he rumen fluid, Dr. Donald Bailey for his help
i n obtaining deer rumen fluid, and Mr. Rodney McCurry for
his help in the photographic aspects and assistance in the
electron microscopy.
i11

TABLE OF CONTENTS
Pa ge
ACKNOWLEDGMENTS • • • • • • • • • • • • • • • • • • • iii
LIST OF TABLES • • • • • • • • • • • • • • • • • • • • v
LIST OF FIGURES • • • • • • • • • • • • • • • • • • • • vii
ABSTRACT • • • • • • • • • • • • • • • • • • • • • • • viii
INTRODUCTION • • • • • • • • • • • • • • • • • • • • • 1
MATERIAL A~D METHODS • • • • • • • • • • • • • • • • • 6
RESULTS • • • • • • • • • • • • • • • • • • • • • • • • 15
DISCUSSION • • • • • • • • • • • • • • • • • • • • • • 37
SUMMARY • • • • • • • • • • • • • • • • • • • • • • • • 45
LITERATURE CITED • • • • • • • • • • • • • • • • • • • 46
iv

LIST OF TA BL ES
Table Page
1. The Effect of Age on the Viability of Euonymus americanus Se eds Maintained at Room Temperature. 16
2. Germination of Unscarifi e d Euonymus americanus Seeds After Treatment with Clostridium cellobioparum Grown in Thioglycollate Broth . •• 11
3 . Germination of Unscarified Euonymus After Treatment with Clostridium cellobioparum for 24 h . ...•.
americanus
• • • • • • •
4. Germination of Unscarified Euonymus americanus Seeds After Treatment with Clostridium
18
cellobioparum for 48 h. . • • . . • . . . . • •. 19
5.
6.
Germination Seeds After Clostridium Broth . . •
Germination Seeds After Clostridium Broth . • .
of Unscarified Euonymus americanus Treatment with Serial Dilutions of cellobioparum Grown in Thioglycollate • • • • • • • • • • • • • • • • • • •
of Unscarified Euonymus americanus Treatment with Cell Concentration of cellobioparum Grown in Thioglycollate • • • • • • • • • • • • • • • • • • •
1. Germination of Un sca ri f ied ~onymus americanus Seeds After Treatment wil h Ruminococcus
21
22
flavefaciens for 24 h . . . . . . . • • • . . . . 23
8. Germination of Unscarified Euonymus americanus Seeds After Trea tment with Buminococ cus flayefaciens for 48 h . . . • • . • • . • • . .. 24
9. Red ucing Sugars Produced from Euonymu s americanus Seeds in the Presence of Rumen Bacteria. • . . . 25
10. Detec tion of Reducing Sugars Produced by Various Cell uloses Hydrolyzing Avicel and Carboxymethyl Cellulose . . . . . . . . . . . . . . . . . . . . 28
11. Germination of Unscarified Euonymus americanus Seeds in the Presence of Enzyme Extracts Prepared from Rumen Bacteria . . . . . . . . . • . . . . . 30
v

Table
12. Reduc ing Sugars Produced from EUQnymus americanus Seeds in the Presence of Enzyme Extracts Prepared from Rumen Bacteria • • .
13. Perc ent Seed Germinat i on of Euonymus americanus Seeds After Treatment with an
• • •
Endocellulase . . . . . . . . . . , • • • • • • •
vi
Page
31
33

LIST OF FIGURES
Figure Page
1. Congo red screening for endo ellulase activity of Clostridium cellob i ogarum • • . • • . . • . • 27
2.
3.
Reduc i ng sugars liberated from Euonymus americanus seeds by 10 U of a commercial endocellulase . . . . . . . . • . . . .
Reducing sugars l ibe r ated from Euon~mus americanus seeds by 8 U of a commercial endocellulase . . . . . . . . . . . . .
v 1 1
• • • • 35
• • • • 36

THE EFFECT OF CELLULOLYTIC RUMEN BACTERIA OR THEIR CELL-FREE
EXTRACTS ON THE GERMINATION OF Euonymus americanus L.
Gary Thomas Howard July, 1987 50 pages
Directed by: L. P. Elliott, M. R. Houston and J. E. Winstead
Department of Biology W stern Kentucky University
In past at t empts, the e xperimental germination of the
seeds of Euonymus americanus L. in vitro has had little
success. Howe ver, treatmen t of seeds with rumen fluid con
taining viabl e microflora has been succ essful in stimulating
germination. In the presence of the cellulolytic rumen
bacterium, Clostridium cellobiogarum ATCC 15832, seeds of A.
amecicanus were stimulated to germinate. Subsequent studies
were designed to determine whether the bacterium synthesized
a cellulolytic enzyme responsible for initiating germi
nat i on. The cell-free endocellulase from ~. cellobiogarum
was demonstrated to irlduce germination of the seeds. To
support the hypothesis that endocellulase from ~.
cellQbiogarum was responsible for triggering germination, a
1,4- e -endo-glucanase (EC 3.2.1.4) from Penicillium
fu nic ulosum was used to treat the s eeds. Also, Ruminococcus
fla vefaciens strain FD-1 was found to initiate germination
of A. americanus seeds. Thus, cellulase activity is
indicated in the degradation of the testa of the seed
allowing imbibition and germination.
viii

1
INTRODUCTION
The seeds of most endemic plants of temperate climates
will not germinate immediately after ripening as these seeds
at maturity will enter a state of dormancy; the two most
common mechanisms of dormancy are impermeability of the seed
coat to water and dormancy of the embryo (Wilson et al.,
1971). The impermea bilit y of the seed coat or testa, a
common cause of delayed germination , can be gradually
decreased by freezing, thawing, and the action of micro
organisms in the soil (Wilson et al., 1971). These methods,
coupled with wind action and blowing the seeds against
finely abrasive surfaces, tend to scratch the testa and
permit the absorption of water. Scratching of the testa is
commonly referred to as scarific 3tion. An artificial method
of scarifying seed s is to abrade them wi th fine sandpaper or
with a file.
To mediate the physiological condition of the embryo
causing dormancy, seeds must undergo a series of complex
enzymatic and chemical changes collecti ve ly referred to as
after-ripening. After-ripening can occ u r with moisture and
low temperatures by a process known as stratification.
Dormancy can also be caused by various chemicals that may be
present in the seeds . The chemicals that are inhibitors are
commonly l ~ached out of the seeds in wet soil under natural
conditions. In addition, seeds vary grea tly in the length
of time during which they will retai n their viability and

2
ability t o germinate. The l ongevity of most seeds may be
increased by dry storage a t low temperatures (Wilson et al.,
1911). Plants such as Juniperus communis and Juniperus
viriiniana rely on birds to disperse their seeds
(L i vingston, 1912). Holt huijzen and Sharik (1984) showed
that passage of ~. viriiniana seeds through the gastro
intestinal tract of cedar waxw i ngs and yellow-rumped
warblers i ncreased germination 150 to 350 percent. Seeds of
several different plants that have passed through the
digestive t ract of black bears often show greater germi
nation r ates (Rogers and Applegate, 1983). Janzen (1970)
reported that Acacia tortilis seeds must pass through the
digestive system of animals, i.e. elephants and i mpalas , in
order for the seeds to germinate. The lack of Calvaria
species younger than 300 years is reportedly due t o the
extinction of the dodo bird sinc e germination of the seeds
depended upon passage through the bird's intestinal tract
(Pig l , 1912).
The results of this thesis report the germination
strategy of Euonymus americanus L. This plant is an abun
dant understory shrub in remnant ha rdwood forests of south
central Kentucky with a reported distributional frequency
exceeding 98~ (Bougher and Winstead, 1914). This shrub is a
member of the Celastraceae family and contains over 120
other species of this genus worldwide (Strausbaugh and Core,
1913). However, t, americanus is found exclusively in the
United States.

3
Considerable resea r ch on the genus EUQnymus has been
focused on seed germi nation and embryo development in the
European species E. europaeus (Alekseeva, 1975a, 1975b;
Beranger-Novat and Monin, 1971a, 1971b; Beranger-Novat et
al ., 1977 ; Monin, 1965, 1967a, 1967b, 1967c, 1967d, 1968;
Nikolaeva, 1959; Nikolaeva et al., 1973; and Nikolaeva et
al., 1974). More specifically, Nikolaev~ et al. (1973)
found that B-indolyl-acetic acid and abscisic acid behaved
as growth i nh i bitors while g i boerellin and kinetin played an
important ro le in removal of dormancy. The results 0f this
study also revealed that B-indolyl-acetic acid decreased in
concentration under the influence of co l d strat i fication.
Bayer (1984) showed that E. americanus primarily
reproduces asexually. Runners, produced from the base of
the stem, grew horizontal l y along the substrate. These
runners often developed roots and frequently produced a new
stem. Eventually, such runners are severed and separate
plants formed.
Rudolf (1974) using long, cold stratification periods
reported a 15J increase in seed germination. Bayer (1984)
demonstrated that exhaustive tests us i ng HCl/HOH solutions,
heat treatments , long stratification periods, and different
combi nations of photo-periods were unsuccessful. Howard
(1986) showed a 90J germination rate using rumen fluid
containing viable microflora. Howard's study also revealed
a decrea~e in germination time from 20 days, with no
stratlfication, to three days, wi th a cold stratification.
Scanning electron microscopy by Hill et al. (1985) verified

Howard's wo rk concerning the germ in t ion of £. americanus
seeds by the rumen fluid treatment.
4
Herrera ( 1981) sugg ested that the qual i ty of seed
d ispersal bears an important relationship to the nutritional
rewa rd offered by the plant in the form of fruit pulp.
Those pulps associated with h igh s eed d ispersal had a high
lipid andlor protein content. Har greaves (1971) reported
t he presences 0 lipids and sugars in high concentrations in
the arils of Euonymus. The oil s cont ained the usual range
of C16 and C18 fatty acids but were particularly rich in
unsaturated C18 fatty acids. The sugars were a mixture of
glucose, fructose, and rhamnose. Since seeds of Euonymus
are highly nutritional, they should be di s persed easily.
According t o Halls and Alcaniz (1972), the seeds of £.
am e ricanus are an important food source for deer, and
Blakelock (1951) noted bluebirds and robin s foraged on the
fruit. Sinc e deer forage on this pl ant and Howard's (1986)
study showed inc reased germination of the seeds when treated
with rumen fluid, an hypothes i s that r umen microflora play
an i mportant role in inducing ge rmination of t he seeds of £.
americanus is pl aus ibl e. Windham and Ak i n (1984) reported
that rumen bacte r ia were the most act i ve cellulose and fiber
d i gesting populations in the rumen. Clostridium
cellobioparum and Ruminococcus flaveTaciens are two common
cellulolytic bacteria found in the microbial niche of the
rumen. Thus, the objectiv e of thi s thesis was to elucidate
whether the rumen bacterium k. cellobioparum ATCC 15832
acting alone with the seeds will induce germination and, if

5
so, by what mechanism. Also, B. flayefaciens strain FD-l is
being assayed for initiating germi nation of ~. americanus.

6
MATERIAL AND METHODS
Seed Collection
Fruiting capsules of Euonymu5 americanu5 were collected
i n early October of 1983, 1984, 1985, and 1986 from two
sites in south central Kentucky and maintained for testing.
The first collection site was l ocated east of Interstate 65
on Route 68/80 i n Barren County near the village of Bonayer.
The second site was east of Interstate 65 on Route 31E in
Barren County nea r Barren River Lake at the Brigadoon Nature
Preserve of Western Kentucky University.
Seed Pregaration
Seeds were allowed to dry at room temperature before
removal of the arils. Before treatment, seeds were surfaced
sterilized as described by Kremer et al. (1984), whi oh
involved immersing seeds in 3.0S (w/v) sodium hypochlorite
for five minutes followed by immersing in 70S (w/v) ethanol
for two minutes and then rinsing with distilled deionized
water (DI H20). Subsequently, seeds were then subjected to
various treatments to induce germination. After treatment,
the seed~ wer e rinsed with DI H20 and placed on a single,
sterile nine centimeter circular piece of Whatman #1 filter
paper in petri dishes.
~rtitions for Germination
An Environator Corporation Model 3448 growth chamber
was used to provide the conditions for seed germination.

The chamber was programmed for a cont1nuously cycling 12-h
day temp e rature of 29.4oC and a 12-h night temperatu re of
18 . 30 e wit h cyc1 1ng fluorescent and incandescent lighting.
All seeds were kept moist with DI H20 throughout the
experiments.
Seed Viability Test1n&
7
The two chemicals used in the seed viability testing
were a tetrazo11um sal t and indigo carmine ( Bonner, 1974).
The tetrazolium salt used in this assay was 2,3,5-triphenyl
tetrazolium chloride (Sigma, St . Louis, MO) which may be
used as a vital stain. Indigo carmine (Sigma, St. Louis,
MO) is a non-vital stain. One-hundred and forty seeds from
1983, 1984, 1985, and 1986 were scarified with a dissecting
probe to facilitate entry of the solutions.
One-ha lf of the seeds (70 of 140) were immersed in a 1S
(w/v) aqueous solution of tetrazolium chloride and were eft
in the dark at 300 c for 48 h. The other 70 sceds were
imme rsed in a 1S (w/v) aqueou solution of indigo carmine
and left in the dark at 30 0 C for 48 h. The seeds were then
removed after 48 h and cut in half to observe staining with
a stero binocular dissecting microscope.
Mi croor&anisms and Growth
The cellulolytic rumen bacterium Clostridium
cellobioparum ATCC 15832 was obtained from the American Type
Culture Collection, Rockville, MD. Ruminococcus
flayefaciens FD-1 was a gift from Dr. B. A. White,

University of Illinois, Urbana, IL. The k. cellobiopa~
was grown r outinely at 37°c in Brew er 's Thioglycollate Broth
(Oifco) Ot- Medium 10 (Caldwel l an d Bryant, 1966)
anae r obically in Gas Pak jars (BBL Microbiology Systems,
Cockeysvill e , MO) for 48 h. For seed germination studies,
an inoculum was prepared by pipetting 1.0 mL of culture into
either 250 mL of Brewer's Thioglycollate Broth or Medium 10.
The inoculated broths were incubated anaerobically in Gas
Pak jars at either 39 0 C or 40 0 C for 24 h or 48 h. R.
flayefaciens was grown routinely at 39°C in Gas Pak jars for
24 h or 48 h. In seed germination studies, an inoculum was
prepared by pi petting 1.0 mL of culture into 250 mL of
Medi um 10 (M 10) and incubating anaerobically in Gas Pak
jars for 24 h or 48 h.
Seed Iooculation with Bacterial Cells
One-half of the seeds (1985) in each assay (80 of 160)
were scarif ied by the use of a dissec t ing probe to scratch
the seed coat, and ha lf were unscarified. Each subset of 20
seeds was covered with 20 mL of sterile thioglycollate broth
(control) or one of the following 48-h thioglycollate broth
cultures of k. cellobioparum: (1) incubated anaerobically,
(2) cell-free extra ct cent rifuged at 10 ,000 x g for 5 min
and filtered through a 0.45 ~m membrane, or (3) autoclaved
at 121°C for 15 min. The same assays were repeated as
described above except that the incubation period for growth
of k. cellobioparum was 48 h at 40°C. The same assays were
repeated except that the i ncubation temperature was 39°C for

24 hand 48 h. All experiments were re peated using
~. cellob ~oparum cultures grown in M 10 i nstead of
thioglycol l ate broth.
Seeds of ~. americanus were treated with A.
flavefacien s by those assays described for ~. cellobioparum;
however, only M 10 broth was used for growth , and an
incubation temperature of 39 0 C was used for 24 hand
repeated at 48 h. At the end of the incubation periods, the
seeds were r i nsed with 01 H20 and placed in the
environmental chamber programmed for the proper germination
conditions.
In additi on, ~. americanus seeds were treated with
decimal dilutions of ~. cellobioparum. The dilutions were
made in 90-mL blanks containing 0.1J (w/v) peptone wate in
an anaerobic glove chamber Model AL 12441. One-tenth of a
milliliter of each dilution was then transferred onto
triplicate petri dishes which contained 4J thioglycollate
agar. Glas 3 L-rods were used t o spread the inoculum on the
plates. Plates were incubated anaerobically at 39 0 C for
48 h. After the incubation period, the seeds were
elutriated with 01 H20 and placed in an environmental
chamber programmed as previously descri bed, and the seeds
were check ed for germination daily.
A con~entrated suspension of ~. celloPioparum was also
used to assay for induction of germination in ~. americanus
seeds. An inoculum of 1.0 mL of a 24-h culture of~.
cellobioparum was used to inoculate 250 mL of Brewer's

10
Thioglycollate Broth, and the inocul a ted broth wa s
anaerobic~lly incubated for 48 h at 39 0 C. Afte r the
incubation period, the broth was centrifuged at 10,000 x g
for 5 min. The supernatant was removed, a nd 40 mL of saline
solution (0.9%) were added to the pellet. The cell
suspensi on was pipetted into two petri dishes, 20 mL/plate.
To these petri dishes, 20 seeds of t. americanus were added
per dish. One petri dish was incubated anaerobically at
39 0 C for 24 h, and the other was i ncubated anaerobically at
39 0 C for 48 h. Then the seeds were elutriated with Dr H20
and placed in an environmental chamber programmed as noted
bef ore and checked daily fo r germination.
ScreeninB Cultures for Cellulase Actiyity
~. ~ ellobioparum was assayed for the production of
cellulase by two met hods . The fjrst method consisted of
inoculating 0.1 mL of a 48-h culture from thioglycollate
broth into 10 mL of Omelianski Both which contained a strip
of Whatman #1 f il ter paper as t he sole source of carbon
(Omelianski , 1902). Three cultures at each temperatu re were
incubated anaerobically at 39 0 C and 40 0 C for 21 days.
The second method used was Teather a ll d Wood's (1982)
technique of congo red screening. A 48-h culture of ~.
cellobioparum grown in Brewer's Thioglyc0!la t e Broth was
used to spot inoculate 0.05 mL of inoculum on 45 plates of
Medium 10 Agar. Three sets of tripl i cate ~lates (15
plates/set) were placed at 37 0 C, 380 e, 3S J C, 400 C and 41 0 C
and were incubated anaerobically for 48 P. After 48 h, the

11
plates were overlayed with M 10 which lacked the normal
carbohydrates but contained 0.8~ agar a nd O . l~ carboxymethyl
cellulose (CMC). One set of plates was reincubated for 24 h
at the same tempera tu re while another set was reincubated
for 48 h and the third set reincubated for 72 h. After
these time periods, the plates were flooded with an aqueous
solution of congo red (1 mg/mL) for 15 min. The congo red
was decanted, and the plates we re flooded with 1M NaCl for
15 min . This was fo llowed by decanting the NaCl solution
and flooding the plates with 1M HC1. The clear zones around
the colon i e s were then measured in millimeters.
Isolation of Cell-Free Cellulase
A 48-h culture of ~. cellobiogarum grown in 250 mL of
Brewer' s Th i oglycollate Broth and Medium 10 Broth and a 48-h
culture of R. flavefaciens grown in Medium 10 Broth were
used to prepare cell-fre~ extracts. Cells were sedimented
by cen~rifugation a t 10,000 x g for 5 min . The supernatant
was decanted and proteins concentrated by precipitation with
ammonium sulfate at an 80~ (wt/vol) concentration. The
preCipitate was removed by centrifugation at 22,000 x g for
20 min at 40 C. The protein was resuspended by addition of
5 mL of 0.05 M sodium acetate buf fe r, pH 5.2. The ammonium
sulfate was remov ed from the resulting enzyme extract by
dialysis in 1.0 1 of 0.05 M sodium acetate buffer, pH 5.2,
which was changed twice in a 24-h period. The crude enzyme
extract was stored at _200 C until needed.

2
Enzyme Assays
(a) Endog l ucanase activity
The reaction mixture containing 1% CHC (w/v ) (Sigma, St.
Lou i s, HO) in 4.0 mL of 0 . 05 H sodium acetate buffer, pH
5.2, and 1. 0 mL of enzyme extract was s haken in a New
Brunswick Scientific Incubator shaker at 90 strokes per min
at 37°C for 2 h (Hayashida and Ho, 1986). Reducing sugars
formed were determined by the method of Nelson (1944).
(b) Exocellulase activity
Assays were conducted using a 1% (w/v) Avicel (Baker,
Phillipsburg, NJ) in 4.0 mL of 0.05 H sodium acetate buffer,
pH 5.2, and 1.0 mL of enzyme extract. The reaction mixture
was shaken in a New Br unswick Scientific Incubator shaker at
90 strokes per min at 37 0 C for 2 h. Reducing sugars f ormed
were determined by the method of Nelson (1944).
(c) Cel l obiase act i vity
Assays were conducted using a 1% (w/v) D (+) cellobiose
(Sigma, St. Louis, HO) in 4.0 mL of 0.05 H sodium acetate
buffer pH 5 .2 and 1.0 mL enzym~ extract. The reaction
mix ture was shaken in a New Brunswick Scientific Incubator
shaker at 90 strokes per min at 37°C for 2 h. Reducing
sugars formed were determined by the method of Nelson
(1944) .
Protein Determination
Protein was est i mated by the Bio-Rad protein assay with
bovine serum albumin as a standard (Bradford, 1976).

13
Enzyme Inhibition
Ass a ys were conducted using a 11 CMC (w/v) in 4.0 mL of
0.05 M ~od i um acetate buffer, pH 5.2, and 1.0 mL enzyme
e xtract of ~. cellobiopa rum were incubated a t 37oC, 45 0 C,
and 600 C for 2 h. In addition, an enzyme reaction mixture
was inactivated with 1.0 mL of 0.001 M HgCl 2 and incubated
at 370 C for 2 h. This assay was repeated except that 1.0 mL
of 0.01 M HgC1 2 was used. Reduci ng s ugars were measured by
the method of Nelson (1944).
Seed Assay with the Enzyme Extracts
Enzyme extract s from ~. cellobioparum and B.
flavefaciecs obtained as described earlier were used in thi s
assay. One milliliter of cell-free extract prepared from
each bacte rial species was used to treat ten seeds of ~.
americanus for 24-h and 48-h incubation periods with shaking
at 37oC. After each time period, the amount of reducing
sugar libera ted was measured by t he method of Ne l son (1944).
The seeds were removed, e l utriated with OI H20, placed in
the environmental chamber programmed as previously described
and observed daily for germination.
Seed Assa y with an Endoglucanase
A 1,4- 8-0 glucan glucanohydrol ase (EC 3.2.1.4) from
Penicillium funiculosum was obtained from Si gma Chemical
Co., St. LOUis, MO. This endocellulase was a purified
enzyme extract with a specif i c activity of 11 U/mg of
protein. In this assay, quantities of 10 U and 8 U of the

14
endocellulase was buffered with 2 mL, 1 mL, and 0.5 mL of a
0.05H s odium acet ate buffer, pH 5.2.
The buffe. ed endocellulase was ster ilized by filtrat i on
using a Hillex HA SLHA0250S ul t ra c lean 1ng filter wh i ch was
attached to a s t erile syringe. The filtrate was collected
in sterile test tubes. To seven test tubes (10 x 100 mm),
ten seeds were added and i ncubated with shaking at 370 C for
2 h, 4 h, 6 h, 8 h, 12 h, and 48 h. Af t er e ach time
i nterval, the amount of glucose liberat ed was measured by
the method of Nelson (1944), the seeds removed, elutriated
with 01 H20, placed in the programmed environmental chamber,
and checked dai l y for germination.
See d Assay with an ExoKlucanase
A 1,4-8-0 glucoside glucohydrolase (EC 3.2.1.91) was
obtai ned from Sigma Chemical Co., St. Louis, HO. This
exocellulnse was purified from almonds and had a unit
act i vity of 5 U/mg. In th i s assay, quantities of 5 U and
8 U of the exocellulase was buffered with 1 mL and 0.5 mL of
a 0.05 M sodium acetate buffe r , pH 5.2. The buffered
exoce l lulase was sterilized by filtering through a Hillex HA
SLHA0250S ultracleaning filter into sterile test tubes. To
each test tube, ten seeds were placed and incubated with
shaking at 37°C for 6 h, 8 h, and 12 h. After these time
periods, the amount of glucose liberated was measured by the
method of Nelson ( 1944), the seeds removed, elutriated with
01 H20, placed in a programmed e nvironmental chamber, and
observed daily for germination.

15
RESULTS
Seed Viability Assay
Seeds of Euonymus amerioanus tested with indigo
carmine and tetrazolium showed an average of 20.7S decrease
i n viability in a three-year period when stored at room
temperatu re (Table 1). The method us i ng tetrazolium
reflected a 2.2S higher viability of the seeds per year than
the ind igo carmine method. Seeds three years old had an
average of 79.3S viab i lity as determined by both of the
assays.
Seed Germination Assay with Baoterial Cells
Seeds treated with ~. oellobioparum germinated only i n
the presence of viable cells. When the bacterium was grown
i n Brewer's Thioglycollate Broth, germination was induced at
a 65S rate within 9 days after 48 h of i ncubation with ~.
oellobioparum (Tab le 2). Seeds did not germinate when
treated with Brewer's Thioglycollate Broth. Also, seeds did
not germinate whe n subjected to ~. oellobioparum f or only
24 h nor d id germination occur when they were scarified.
When ~. oellobioparum was grown in M 10 broth, seeds were
induced to germinate at a 20~ rate within 9 days after a
24-h incubat i on period with the bacteria (Table 3). An
i ncrease of germination to 70S was achieved when the seeds
were treated with ~. oellobioparum for 48 h (Table 4).
1

16
Table 1 . The Effect of Age on the Viability of Euonymus
americanus Seeds Maintained at Room Temperature
ASSAY
Indigo Carmine Tetrazoli um
Year of Collection Vi a ble ( J) Nonviable ( J) Vi3ble (S) Nonviabl e( S)
1986 98.0 2.0 100 0
1985 88.89 11. 11 91. 43 8.57
1984 84.29 15.71 88 .57 11.43
1983 78.34 21.66 80 .19 19.81
170 seeds per year were assayed in each method .

• Ii
Table 2. Germination of Unscarified Euonymys ~ricanys
Seeds 1 after Treatment 2 with Clostridiym
cellobioparym Grown in Thioglycollate Broth
17
Trea tment Germination Time (Days)
~ of Germination
Anaerobically Grown Culture 6-9 65
Ce ll-Free Extract
Autoclaved Culture
'20 seeds per assay.
o
o
2Seeds were held anaerobica l ly for 48 h at 40 0 C i n a Brewer's Thioglycollate Broth culture preparation of Clos t ridiym cellobiQparym. The culture preparations were treated as indicated.
o
o

Table 3. Germination 0 ' Unscarified Euonymus americanus
Seeds 1 after Treatment2 with Clostridium
cellobioparum for 24 h
18
Treatment Germination Time (Day s)
J of Germination
Anaerobically Grown Culture
Cell-Free Extract
Autoclaved Culture
6-9
o
o
20
o
o
120 seeds per assay.
2Seeds were held Medium 10 broth cellobillparum.
anaerobically for 24 h at 39 0 C in a culture preparation of Clostridium The culture preparations were treated
as indicated.

Table 4. Germination of Unscarified Euooymus americanus
Seeds 1 after Treatment 2 with Clostridium
cellobioparum for 48 h
19
Treatment Germination Time (Days)
J of Germination
Anaerobically Grown Culture
Cell-Free Extrac t
Autoclaved Culture
120 seeds per assay.
6-9 70
o o
o o
2Seeds were held anaerobically for 48 h at 390 C in a Medium 10 broth culture preparation of Clostridium ~_llobioparum. The culture preparations were treated as indicated.

20
Seed3 did not erminate when they were only treated with
M 10 broth.
To determine if concentrated cells were needed for
germ i nation, seeds were treate with serial dilutions of ,.
cellobioparum. Germination was induced with 10- 1 to 10-3
dilutions of the culture. In addition, lOS germination of
the seeds occurred within 10 days with the 10-8 dilution
(Table 5) . When cells of ,. cellobioparum were
concentrated, increased ge r mination of E. americanus seeds
occurred. A 24-h treatment of seeds with concentrated cells
resulted in 75S germination in 9 days; however, a 48-h
treatment resulted in an 85S germination rate in 7 days
(Table 6).
The question arises whether other ruminant bacteria
would stimulate germination. Seeds treated with E .
flavefaciens germinated only in the presence of viable
cells. When li. flavefaci€ns was grown in M 10 broth and
used to treat seeds for 24 h, 70S of the seeds germinated
within 9 days (Table 7). An increase of 80% germination was
achieved when the seeds were treated with E. flayefaciens
for 48 h (Table 8). Again, seeds treated with M 10 did not
germinate.
RedUCing sugars liberated f r offi E . americanus seeds by
the rumen bacteri a were detected by the method of Nelson
(Table 9). There was an increase in the glucose
concentration released from the seeds as time of treatment
was increased f rom 24 h to 48 h indicating the hydrolysis of

Table 5. Germination of Unscarified Euonymus americanus
Seeds' after Treatment2 with Serial Dilutions
Dilut ion
of Cl ostridium cellobiQparum grown in
Thioglycollate Broth3
Germination S of
21
of Culture Time (Days) Germination
100 7-9 70
10-1 6-9 30
'0-2 6-9 40
10-3 6-9 '0
'0-4 0 0
'0-5 0 0
10-6 0 0
10-7 0 0
'0-8 10 10
'20 seeds pel" assay.
2Seeds were held anaerobically for 48 h at 390 C in Brewer' s Thioglycollate Broth prepa r at i on of Clostridium cellobioparum. The culture was d i l u~ ed as indicated.
3An average count of 49 x 108CFU/mL was obtained on three plates of thioglycollate agar.
i

22
Table 6. Germination of Unscarified Euonymus americanus
Seeds' after Treatment 2 with Cell Concentration
of Clostridium cellobioparum grown in
Thioglycollate Broth
Incubation Time (h)
24
48
'20 seeds per assay.
Germina t ion Time (Days)
6-9
5-7
~ of Germination
75
85
2Seeds were held anaerobically at 39 0 C in a concentrated cell suspension of Clostridium cellobioparum. The culture preparations were incubated as indicated.

Table 7. Germ i nation 0 Unscar i fied EUQnymus americanus
Seeds' after Treatment 2 with Ruminococcus
f layefaciens for 24 h
23
Treatment Germination Time (Days)
S of Germination
Anaerobically Grown Culture
Cell-F r ee Extra ct
Autoclaved Culture
'20 seeds per assay.
6-9
o
o
2Seeds were held anaerobically for 24 h at 390 C in a Medium '0 broth culture preparation of Ruminococcus flayefaciens. The culture preparations were treated as indi cated.
70
o
o

Table 8. Germination of Unscarified Euonymus americanus
Seeds' after Treatment 2 with Ruminococcus
flavefaciens for 48 h
24
Treatment Germination Time (Days)
S of Germination
Anaerobically Grown Culture
Cell-Free Extract
Autoclaved Culture
'20 seed s per assay.
6-9
o
o
2Seeds were held anaerobically for 48 h at 39 0 C in a Medium '0 broth culture preparation of Ruminococcus flavefaciens. The culture preparations were treated as indicated.
80
o
o
25
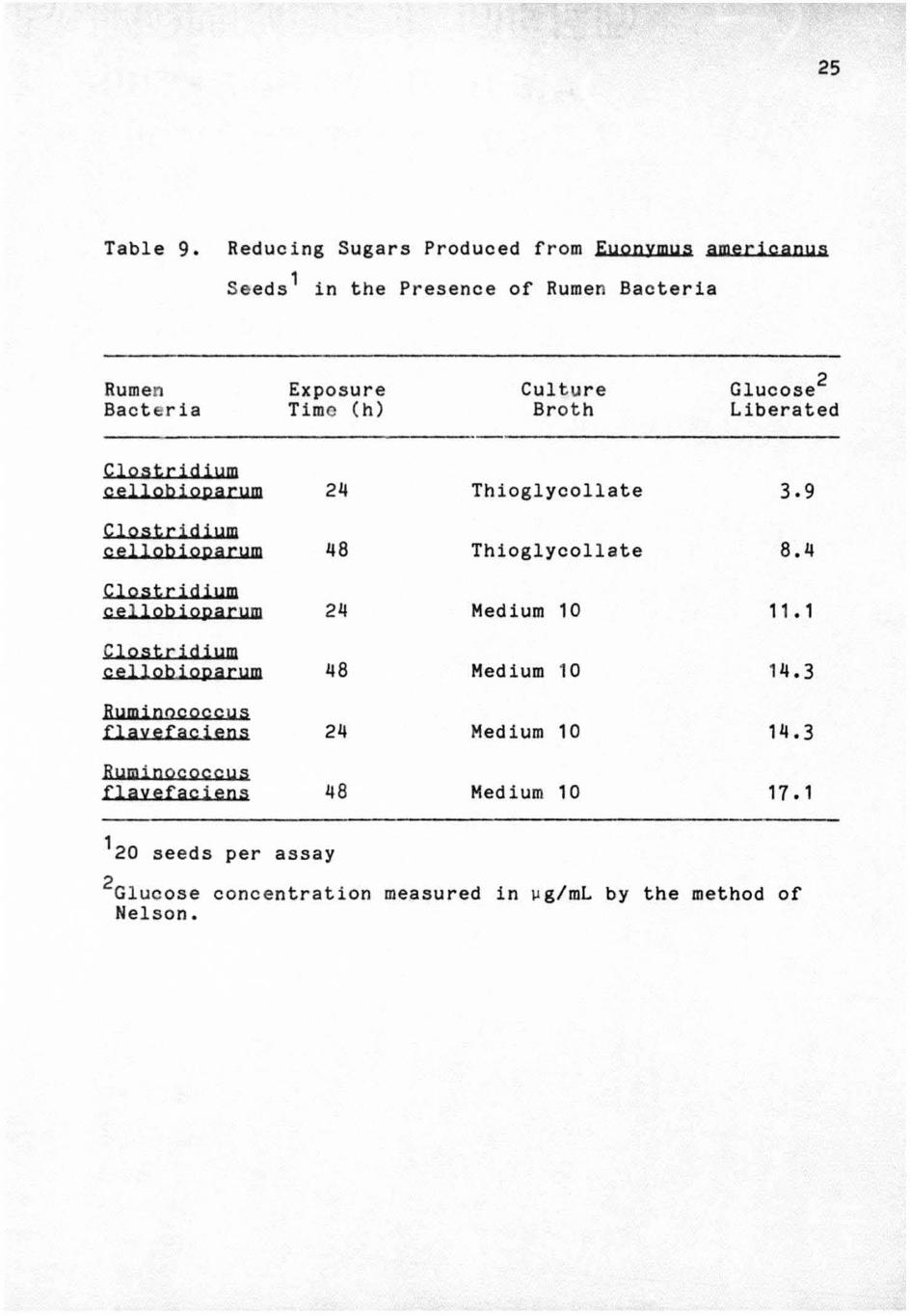
Table 9. Reducing Sugars Produced from Euonymus americanus
Seeds 1 in the Presence of Rumen Bacteria
Rumen Bact eri a
Clostridium cellobioparum
Clostridium cellobioparum
Clostridium c ellob ioparum
Clostridium cellobioparum
Ruminncoccus flayefaciens
Ruminococcus flayefaciens
Exposure Time (h)
24
48
24
48
24
48
120 seeds per assay
Culture Broth
Thioglycollate
Thioglycollate
Medium 10
Medium 10
Medium 10
Medium 10
Glucose2 Liberated
8.4
11. 1
14 .3
14.3
17 • 1
2Glucose concentration measured in vg/mL by the method of Nelson.

cel l ulose by cel lulase. In addition, more glucose was
liberated from the seed s that were subjected to
26
~. cellobioparum grown in M 10 ~roth as opposed to growth in
Brewer's Thiogl ycollate Broth.
Screenini Cultures for Cellulase Activity
Initially the ~. cellobioparum bacterium was screened
for cellulase activ i ty in Omelianski Broth. The organism
was found t o produce cellulase both at 39 0 C and 40 0 C as
indicated by the paper strips being decomposed.
Subsequently, the congo red method was used to detect
endocellulase activity. The halo around colonies of ~.
cellobioparum by the congo red staining procedure indicated
that the bacterium was positive for endocellulase synthesis.
The zone diameter of the cellulase enzyme increased with
time; however, ther e was a slight decrease i n zone diameter
when the temperature was increased or decreased f rom 39 0 C
(Figure 1).
Comparison of Enzyme Activi t ies
To prove that cellulase synthesis by the rumen bacteria
i s involved in triggering seed germinat i on, enzyme extracts
from the bacteria must be assayed. The extent of hydrolysis
of cellulose from concentrated enzyme extracts of ~.
cellobioparym and R. flavefaciens was measured by the amount
of reducing sugar released from CMC and Avicel (Table 10).
Commercial purified cellulase enzymes were also tested. The
commercial exocellulase showed activity on Avicel; but no
activity was obtained on the CHC. The commercial

Figure 1. Congo Red Screening for Endocellulase Activity
of Clostridium cellobioparum
27
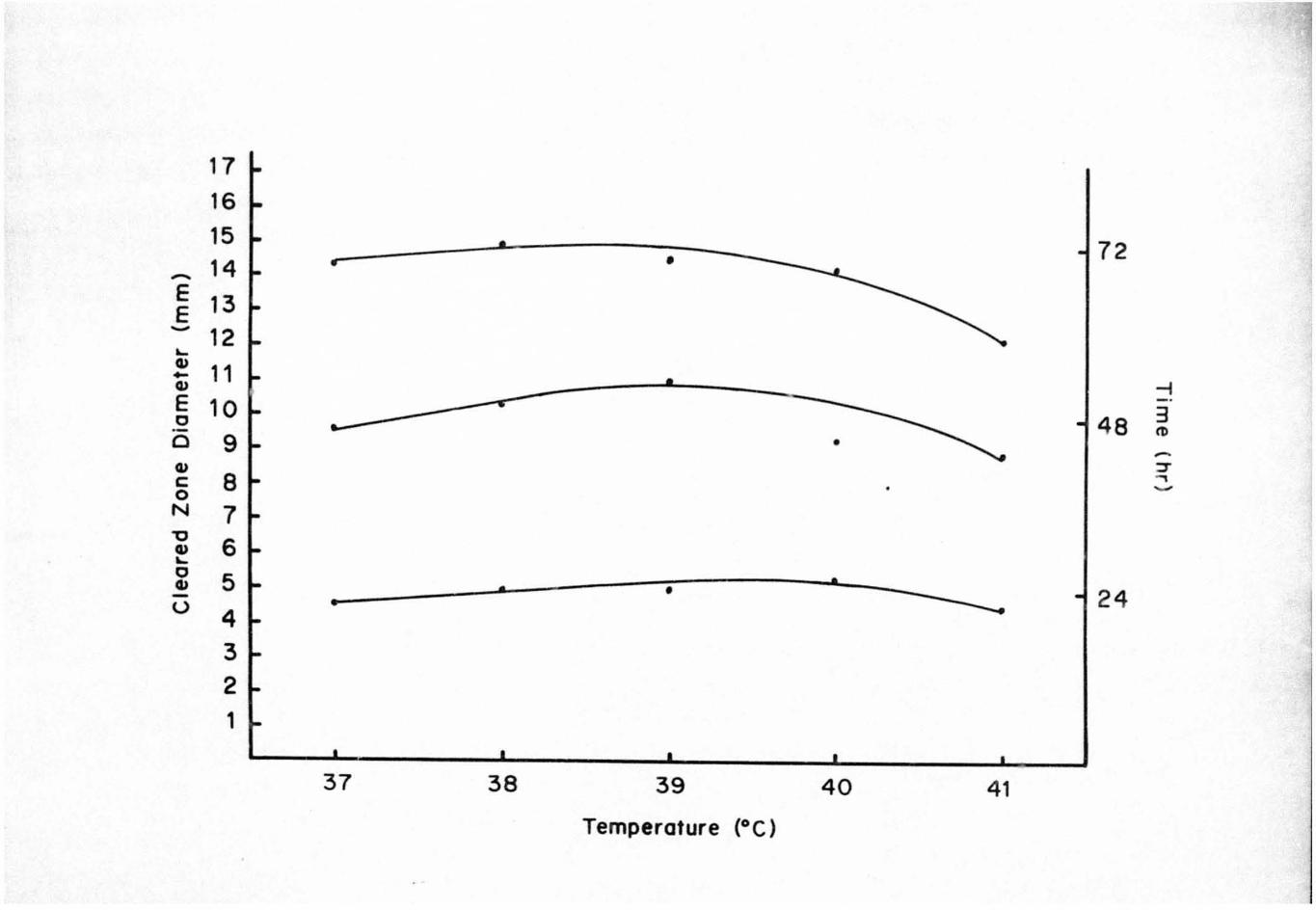
17 16 15
72 • ~ 14 E 13 E ~
12 .... 41
11 - -i 41 - . E 10 48 3 c
tD .-• 0 9
~
::r 41 , c: 8 ~
0 N 7 "0 6 41 .... c 5 • 41 • • ----- 24 -u •
4 3
2 1
37 38 39 40 41 Temperature (OC)

28
Table 10. Detection of Reduc i ng Sugars Produced by Various
Cellulases Hydrolyzing Avicel and Carboxymethyl
Cellulose
Reduced Sugars Produced (\lg/mL)
Enzyme Avicel Carboxymethyl
cellulose
Exocellulase 1
EC 3.2.1.91
Endocellulase2
EC 3.2. 1 .4
Extract from 3 Ruminococcus flavefaciens
Extract from 3 Clostridium cellobiQparum
3.0
0.8
9.5
4.2
1preparation from Almonds.
2preparation from Penicillium funiculosum.
15
20
15.5
3Ex~ract from cells grown anaerobically 48 h in H 10 at 39 C prepared by centrifugation, membrane filtration and ammonium sulfate precipitation.

29
endocellulase showed more activity on CHC than on the
Av icel. The enzyme extract fro m l . flav efaci ens had
activity on both Av i cel and CHC; however, the activity on
the Avicel was one-half that of the activity on the CHC.
The enzyme extract from C. cellobigparum showed activity on
CMC and slight activity on Avicel (Table 10). The enzyme
extracts, prepared from cultures of C. cellobioparum and l.
flavefaciens grown in H 10 broth, revealed no cellobiase
activ i ty.
Seed Germination Assay with Enzyme Extracts
Seeds treated 24 h with an enzyme extracts from l.
flayefaciens enhanced germination compared to seeds treated
for 48 h. Seeds treated with an enzyme extract from C.
cellobigparum germinated optimumly when the cells were grown
in H 10 broth and when treated for 48 h as indicated in
Table 11.
The reducing sugars liberated from the seeds by the
enzyme extracts were quantitated as indicated in Table 12.
An increase in reducing sugars was obtained by an increase
in contact time between seeds and enzyme extract. Also,
more reducing sugars were liberated from the act i vity of the
enzyme extract from C. cellobioparum grown in H 10 broth
than from the enzyme extract prepared from cells grown in
th10gl ycollate broth. The enzyme extract prepared from l.
flayefaciens liberated more reducing sugars from the seeds
than the enzyme extract prepared from C. cellobioparum.

Table ". Germination of Unscarified Euonymus americanus
Seeds' in the Presence of Enzyme Extracts
Prepared from Rumen Bacteria
Species
Clostridium cellobioparum
Clostridium c ellobioparum
Rumi nococcus flavefaciens
Ruminococcus flavefaciens
Time Held in Extract (h)
24
48
48
Culture Broth
Thioglycollate M '0
S of Germination
40 70
60 80
-- 80
20
' Qc curring within 4-8 days in an environmental chamber.
2'0 seeds per assay.
30

31
Table 12. Reducing Sugars Produced from Euonymus americanus
Seeds 1 in the Presence of Enzyme Extracts
Prepared from Rumen Bacter ia
Clostridium cellobioparum
Clostrid i um cellobioparum
Ruminococcus flayefaciens
Ruminococcus flavefaciens
Medium Cells Grown In
Thiogl ycollate Medium 10
Th ioglycollate Medium 10
Medium 10
Medium 10
1'0 seeds per assay.
Exposure Time (It)
24 24
48 48
24
48
Glucose2
Liberated
2.8 9.4
10.2 11.1
12.6
15.3
2Glucose concentration measured in ~g/mL by the method of Nelson.

32
Protein Determination from Enzyme Ex tracts
When ~. cellQbiogarum was grown 24 h an a erob ically in
250 mL of M 10 broth, the cell-free extract was fo und to
contain a concentration of 122 ~g of protein per milliliter.
However, if the incubation period was increased to 48 h,
the pro tein concentration of a cell-free extract from ~.
flavefaciens grown i n M 10 broth for 48 h was 380 ~g/mL.
Enzyme Inhibition
The amount of red ucing sug~r liberated from CMC by the
cellulase activity of the enzyme extract was decreased as
the incubation temperature was increased. The amount of
reducing sugar released at the 45 0 C incubation temperature
was one-half of that released at the 37°C incubation
temperature and the amount of reducing sugar released a t
600 C was one-tenth the amount released at 37°C. In
addition, 0 . 001 M Hg C1 2 and 0.01 M HgC1 2 inhibited all
oe l l ulase activity of the enzyme extract. No reducing
sugars were l i be r nted from the CMC when these concentrations
of HgC1 2 were present.
Seed Germination Assay with Commercial Endocellulase
The total germination percent of seeds exposed to a
purified endocellu l ase from Penicillium f uoiculosum depended
on time of exposure to the enzyme and concentration of the
enzyme (Table 13). Generally, there was an increase in
pe rcent seed germination with more contact time between
seeds and enzyme and when a more concentrated enzyme
suspension was used. However, the percent seed germination

33
Tab l ~ 13 . Percent Seed Germination 1 of Euonymus americanus
Seeds after Treatment with an Endocellulase2
Exposure Time (h)
2
4
6
8
10
12
24
48
10ccurring
2 mL
0
0
0
30
30
30
30
40
within 4-6
2Purified enzyme from
Units of Enzym e
10 U 8U
1 mL 0.5 mL 2 mL 1 mL 0.5 mL
0 0 0 0 0
40 60 0 0 20
60 70 0 40 60
80 90 0 60 80
70 50 0 60 10
60 0 0 50 0
60 0 20 50 0
20 0 20 30 0
days in an environmental chamber.
Penicillium funiculosum.

34
did not increase after 8 h of exposure nd decreas ed
the r eafter with all concentrations of t he enzyme . The more
concentrated enzyme preparation ca sed the greate s t percent
seed germination in the shortest amount of exposure
time, while the les s concentrated enzyme preparation
stimulated less seed germination.
Reducing sugars liberated from the seeds during various
ex posure times with commercial enzym s are indicated in
Figures 2 and 3. There was an increase in the amount of
reducing sugar liberated from the seeds by the different
concentrations up to 8 h of exposure. After 8 h exposure
the amount of reducing sugars leveled off and remained at
the same level throughout the assay.
Seed Germination Assay with Commercial Exocellulase
No germination was obtained from seeds treated with the
commercial exocellulase enzyme. In addition, no redu ing
s ugars were liberated from the seeds treated with the
commerc i al exocellulase enzyme preparation.

Figure 2. Reducing Sugars Liberated from Euonymus
americanus Seeds by 10 U of a Commercial
Endocellulase
35
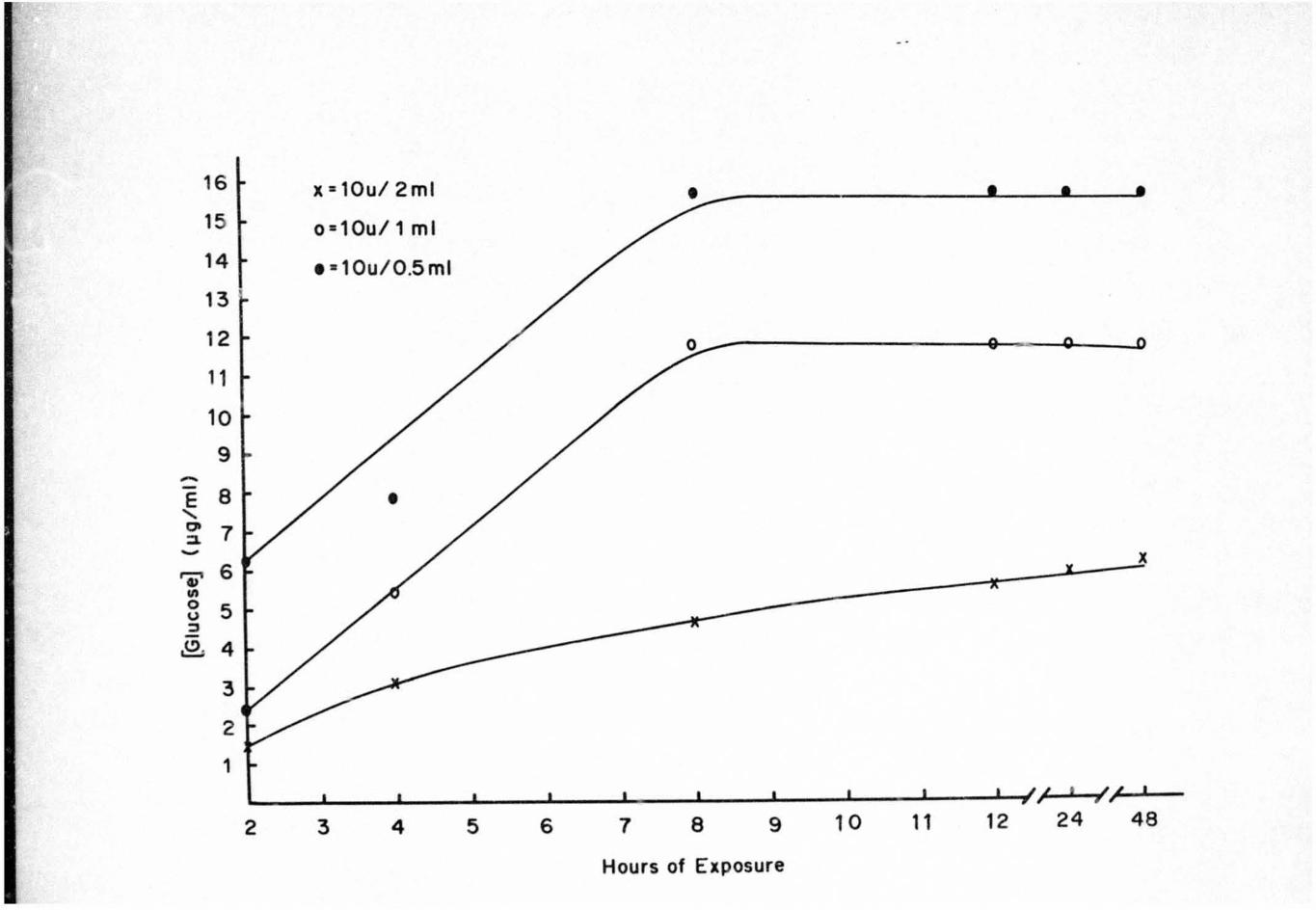
16 It = 10u/ 2ml • 15 0-10u/l ml
14 , · 10u/0.5ml
13
12
1 1
10
9 ~
E 8 • "-co 7 ::a. ~
It
'Ii' 6
"' 0 5 <) ::s
,I:>. 4
3
2
1 ; , I ;; y , I
2 3 4 5 6 7 8 9 10 11 12 24 48
Hours of Exposure

Fi gure 3. Reducing Sugars Liberated from Euonymus
americanus Seeds by 8 U of a Commercial
Endocel lulase
36
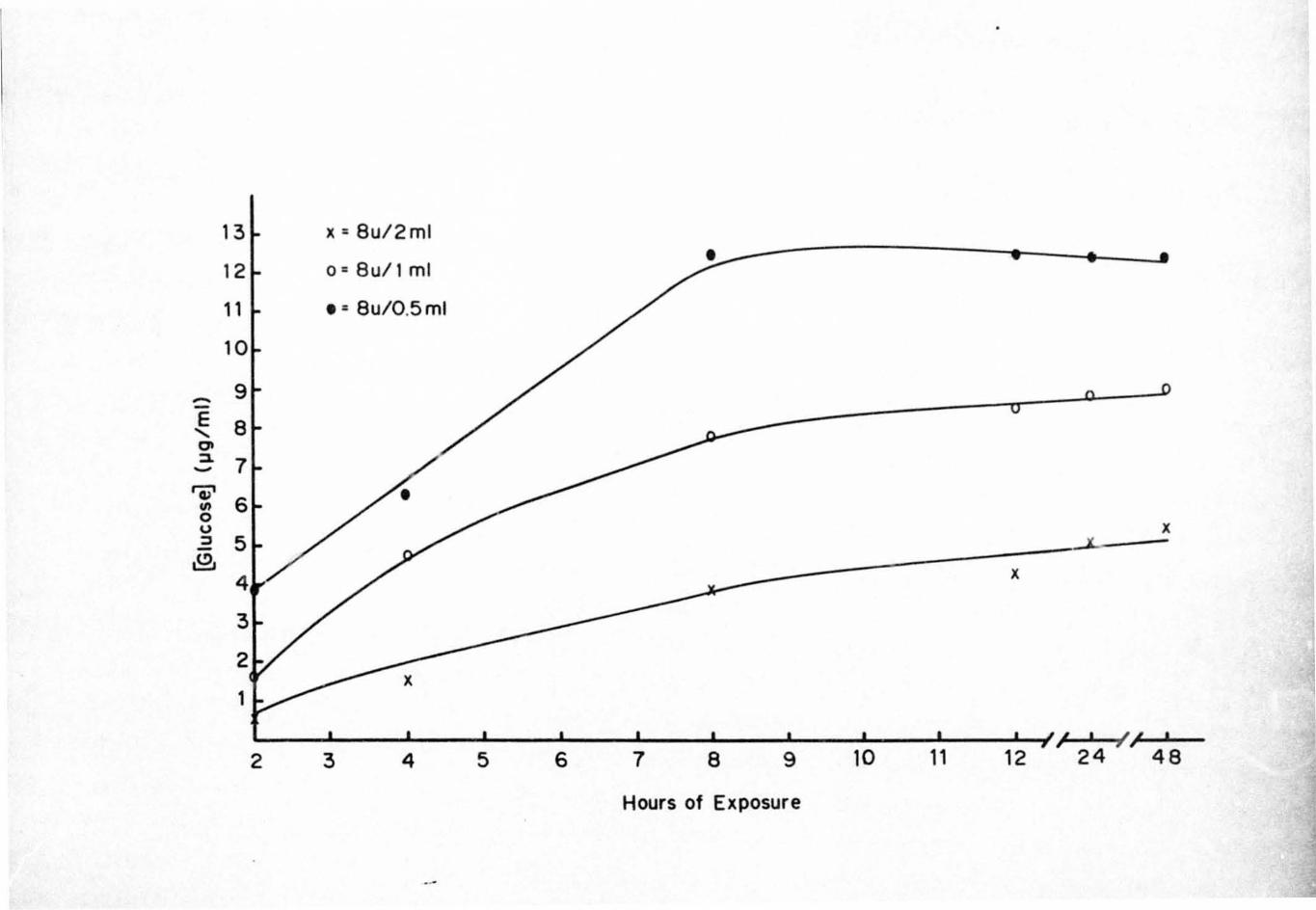
13 x = Bu/2ml
• 12 o=Bu/lml
11 • = Bu/0.5ml
10
~ 9
E B "-CJI ::1 7 -'V
6 III 0 U X ::l 5 X -,l!),
4 x
3
2 x
1
I , I I , , y, 2 3 4 5 6 7 B 9 10 11 12 24 4B
Hours of Exposure

37
DISCUSSION
Seed Viabi l ity Testini
The seed viability of ~. americanus tested by the
indigo c a rmi ne and tetrazolium methods indicated
approximately the same results and provided confidence that
the seeds used were consistent for testing germination.
Seeds from 986 had a 99J viability wh i le three year old
seeds had an 80J viability. The percent of the seeds that
were viable after three years shows an ample longevity of
the seeds which ensures a greater chance of survival of the
plant.
Seed Germination with Bacterial Cells
Howard (1986) indicated that germination of seeds of ~.
americanus is improved by passage through the rumen of
animals. Once seeds have been ingested by a r uminant
animal, the microflora component of the rumen fluid is the
key facto r i n breaking down th c dormancy and stimulating
germination. The microbial population of the rumen is
composed of a complex mixture of bacteria, protozoa, molds
and yea s ts. Each of these groups comprises a collection of
species whose carbohydrate substrate utilization spectra
overlap ( Leedle et al., 1982). The digestive secretions of
animals are unable to hydrolyze major plant cell wall
components i.e., cellulose and hemicellulose (Russell,
1985), but rumen bacteria are the mo s t active cellulose and

38
fiber digesting populations in the r umen (Windham and Akin,
1984). Thus , the rumen bacteria could degrade the testa of
s eed s al lowing imbibition to occur leading to germination.
The r esults of this study indicate that rumen bacteria
acting alone could induce germination of seeds of S.
americanua by degradation of the seed coat.
Unacarified seeds treated anaerobically for 48 h with
~ . cellobioparum grown in thioglycollate broth resulted in
ge r mi nation while treatment for 24 h i nduced no germination.
Also, seeds did not germinate if scarified and treated with
the bacterium. In addition , unscarified seeds treated
anaerobically for 24 h with ~. cellobioparum grown in M 10
broth resulted in a low percent germination while a 48 h
contact time enhanced the ge r mination of the seeds.
Apparently, a 24-h contact time with the seeds i s not
sufficient time for the bacterium to effectively degrade the
seed coat and a l low germination to occur. A 48-h contact
time allowed the bacterium to sufficiently degrade the testa
resulting in seed germination; however, scarification of the
seeds inhibited germination by allowing the bacteria a
direct pathway to the inside of the seed which could result
in degradation of the embryo.
Seeds treated with ~. cellobiQP·a rUJII grown in M 10 broth
had a higher germination rate than those treated with ~ .
cellobioparum grown in thioglycollate broth. The M 10
broth, which contains cellobiose, enhanced cellulase
production whereas thioglycollate broth did not stimulate

cellulase production to the same extent. Thus, bacterial
cells grown in H 10 would have a higher concentration of
cellulase resulting in Quicker degradation of the seed's
testa leading to more germination.
The congo red screening of ~. cellobioparum for
39
endocellulase i ndicated maximum synthesi~ of cellulase at
39 0 C with the production decreasing as the temperature
inc r eased or decreased. This somewha t correlates with the
fact that ~. cellobioparum does not grow at either 25 0 C or
45 0 C and its optimum temperature is 39 0 C (Gibson and Gordon,
1911). It is i nteresting that maximum endocellulase
synthesis occurred at the temperature of the rumen (39 0 C)
(Hungate, 1966).
A decrease in the number of bacterial cells should
result in a decrease in the amount of cellulase which woul d
lead to a decrease in the amount of cellulase which would
lead to a decrease in seed germination. This hypothesis was
tested when seeds were treated with serial logarithmic
d i lutions of ~. cellobioparu~ . The results obtained from
-8 the study supported the hypothesis; however, the 10
dilution resulted in one seed germinating. The reason why
the seed germinated is unknown. The results demonst rated
that an inc r ease in cell concentration of the bacter i um
results in an increase i percent seed germina tion.
A parallel study was done using ~. flavefaciens grown
in H 10 broth to treat seeds for 24 hand 48 h. This •
bacterium also triggered ger minat on of £. amerlcanus seeds

40
as did the treatment of seeds with k. c ellobioparum.
Although a 24 h contact time was adequa te for seed
germination , a 48 h contact time im roved the germination
rate. The bacterium H. flavefaciens is one of the most
important cellulose decomposers found in the rumen (Fox et
RI., 1980). Since H. flayefaciens is a more efficient
cellulase producer than k. cellobioparum, using H.
flayefaciens to treat ~. americanus seeds should decrease
the a mount of contact time needed for triggering germination
of the seeds. The results of this study support this
conclusion.
One of th e most common ways to measure cellulase
activity is to measure the amount of reducing sugar or
glucose liberated from the cellulose polymer. The method of
Nelson (1944) is a colorimetric method using a copper
reag e nt to measure the concentration of glucose liberated
spectophotometrically. With this method, the amount of
gl ucose released f r om the seed could be c ompared to the
percent germ i nation. The amount of reducing sugar produced
from the seeds in the presence of rumen bacteria increased
as exposure time increased. Seeds treated with k.
cellobioparum, grown in thioglycollate broth, liberated less
reduc i ng sugars t h an se ed s treated wi th k . cellobioparum
grown in H 10 broth. The results indicated that H 10 broth
stimulates the bacterium to synthesize more cellulase than
when grown in thioglycollate broth. This increase in
cellulase synthesis results in more degradation of the testa

41
leading to higher germination. Also, seeds treated with B.
flayefaciens libe~~ted more reducing sugar s than those seeds
treated with ~. cellobiogarum which is co related with the
fact that B. flayefaciens produces higher concentrations of
cellul se than does ~. cellobiogarum. One of the most
obvious observations from this experiment is the fact that
an increase of exposure time between the bacteria and the
seeds inc reased the amo unt of reducing sugar liberated.
This increase in exposure time allows the bacteria tv
produce more cellulase which can degrade the cellulose
polymers and libera t e more glucose .
Seed Germination with Enzyme Extracts
The cellulase system of B. flavefaciens has been
studied extensively (Halliwell and Bryant, 1963; Bryant,
1973; and Gardner, 1987 ) . These investigators have shown
that B. flayefaciens produces both an endocellulase and an
exocellulase. Other studies have shown that the
cellulolytic activity of B. flave f aciens is constitutively
expressed, and the degradation of both cel l ulose and
cellobiose occurs simultaneously (Hiltner and Dehority, 1983
and Pettiphen and Latham, 1979). As for ~. cellobiogarum,
characterization of cellulase it synthesi zes has not been
accomplished. ~. cellobiogarum is an isolate from the rumen
(Hungate, 1966) and has cellulolytic activity (Hungate,
1950). The result of the present study showed that ~.
cellobiogarum can degrade CHC, filter paper and to some
extent Avicel. These results imply that ~. cellobiogarum

definitely has an endocellulase activ i ty and to a lesser
degree an exocellulase activity.
42
Endocel lulase is produced by all cellulolytic micro
organisms and is also referred to a s endo- B-1,4-g1ucanase,
B -1,4-g1ucan glucanohydrolase, CMCase or Cx cellulase.
Acid-swollen cellulo s e and carboxymethyl cellulose (CMC) are
the preferred substrate of endocellulase and does not
degrade crystalline cellulose with any efficiency.
Endoc ellulases degrade cellulose polymers by random internal
cuts and produce cellod extrins, cellobiose and glucose.
Exocellulase has been described in mycelial fungi;
however, the presence of the enzyme in bacterial species is
somewhat controversial. The enzyme is also referred to as
exo- B-1,4-g1ucanase, B-1,4-g1ucan cellobiohydrolase,
Avicelase, C1 cellulase or cellobiohydrolase. The prefe r red
substrate for exocellulase is crystalline cellulase and
degrades CMC ineffectively. Exoc ellulase degrades cellul ose
through end-wise cleavage generating cellobiose (written
communication, from Dr. B. A. White, University of Illinois,
Urbana, Illinois).
The cell-free enzyme extract prepared from ~.
cellobioparum grown in thioglycollate broth had a positive
effect upon seed germination. The germ i nation rate of the
seeds increased as exposure time with the enzyme extract
increa s ed. In addition, more seeds germin~ted when treated
with the cell-free extract prepared from ~. cellobioparum
grown in M 10 broth than cells grown in thioglycollate broth

43
indicating that a greater concentration of c llulase was
present in the extract prepared from cells grow in H 10
than in the thioglycollate broth. In suppo r t of this
conclusion, the amount of reducing sugar l I berated from the
seeds by the cellulase extract was sim i lar to that from the
rumen bacteria. With the cell-free enzyme extract of R.
flavefaciens, an increase in contact time between seeds and
extract liberated more reducing sugar; however, there was a
decrease i n germination s exposure time was i ncreased.
Thi ~ suggested excessive seed degradation occurred due to a
greater concentration of the cellulase enzyme and greater
length of exposure t i me. The inhibition assays, using HgC1 2
and increased incubation temperature, reduced the amount of
reducing sugars liberated or completely stopped the
cel l ul ase act i vity. These results support the conclusion
that a cel l ulase enzyme was present in the extract.
Seed Germination with Use of Commercial Cellulases
The results obtained from the concurrent use of the
purified commercial endocellulase pr oved that the cellulase
from the rumen microorganisms is involved in stimulating
germination of E. americanus seeds. Also, it was shown that
the endocellulase, not the exocellulase, is the key enzyme
that induced germina tion. As the data indicated from the
endocellu ase assay, a more dil ute enzyme preparation yields
less reducing sugar and takes longer contact time with the
seeds for the induction of germination. A more concentrated
enzyme preparation yields more reducing s ugar and results in

44
less contact time needed with the seeds fo germination to
be induced. The optimum exposure time of th e endocellulase
with the seeds was between 4 hand 10 h . After 10 h there
was a plateauing of the r educing sugar liberated and a
reduction of seed germination . Apparently the endocellulase
had exhaus ted its supply of subst rate resul ti ng in extreme
degradation of the seed leading to the death of the embryo.
The cellulolytic microflora in the r umen possibly are
synthesizing cellulases that are degrading the cellulose of
the seed coat causing sufficient scarifica ti on of the seed
resulting in imbibition. This process leads to normal
germination of t he seed. By germinating the seeds,
different genotypes of t he plant can be grown and possibly
lead to a better var iety for landscaping. Also, the
cellulolytic bacter i a tested in this research could possibly
be used to stimulate germination in other plants. Other
cellulolytic micro flora of the rumen could possibly induce
germinat ion of E. americanus seeds.

45
SUMMARY
Cellulolyt i c bacter i a of rumen fluid were shown to be
the key fa c tor i n breaking down dormancy and stimulating
germina tion of £. americanus seeds. Onl y seeds treated with
v i able cultures of ~. cellobioparum and l. flayefaciens
germinated. Unconcentrated c ell-free e xt r act and autoclaved
cel l s d i d not stimul ate germination. Concentrated enzyme
extracts prepared from bacteria grown i n M 10 broth and
thiog l ycoll a te broth were tes t ed for stimulation of seed
germination. These enzyme extracts were successful in
induc i ng germinat i on of the seeds and liberating reducing
sugars from the seed coat. Apparently the endocellulase
synthesized by both cellulolyt i c bacteria is induc i ng
germ i na ti on. The purified endocellulase from £. funiculo s um
was a l so successful i n s timulating germination of the seeds.
The concentration of reducing sugars l iberated from the seed
coat and the percent of seed germinating were directly
re la ted to exposure t i me of seeds to the endocellulase.

LITERATURE CITED
Alekseeva, Kh. A. 1975a. Ultrastruc ture of embryos of Euocymys eyropaea L. dormant seeds. Bot. Zh. 60: 40-46.
46
1975b. Effect of temperature on the ultrastructure of em br yo cells of Eyonymys eyropea L. during seed stratification and germinat i on. Bot. Zh. 60:1448-1456.
Bayer, P. E. 1984. Phenotypic differences in populations of Eyonymys americanys L. from the central United States and autecological studies of this species in a deciduous forest of Kentucky . Haster Thesis: Western Kentucky University, Bowl ing Green, Kentucky. 35p.
Beranger-Novat, N. and J. Honin. 1971a. Action de differentes gibberellines sur l'elimination de la dormance des embryons d'Eyonymys europaeys L. C. R. Seances Soc. BioI. Fil. 165:237-240.
__________ 1971b. The breaking of dormancy in Eyonymys eyropeays embryos by gibberellic acid. C. R. Hebd. Seances Acad. Sci. Ser. D. Sci. Natur. 272:1368-1371. (Abstract).
Beranger-Novat, N., J. Honin, and C. P. Singh. 1977. Es ai de definition des etats physiologiques les plus favorables a l'action d'un traitement gibb~rellinique de levee de dorma nce chez l'embryon d'Eyonymys eyropaeys L. C. R. Seances Soc. BioI. Fil. 171:82-36.
Blakelock, R. A. 1951. A synopsis of the genus Eyonymys L. Kew. Bul l . 1951:210-290.
Bonner, F. T. 1974. Seed Testing . p. 143-149. In C. S. Schopmeyer (ed.), Seeds of Woody Plants in the United States. Forest Service, U. S. Department of Agriculture, Washington, D. C.
Bougher, C. K. and J. E. Winstead. 1974. A phytosociological study of a relict ha rdwood forest in Barren County, Kentucky. Trans . Kentucky Acad. Sci. 35 : 44-54.
Bradford, H. 1976. A rapi d and sensitive method for the quantitation of microgram quantities of protein utilizing the pr inc i ple of protein-dye bindi ng. Anal. Biochem. 72:248-254.

47
Bryant, H. P. 1973. Nutr i tional reqviremen t s of the predominant r umen cellulol ytic bacte 1 . Fed. Pro~ . 32:1809-1813.
Ca ldwel l, D. R. and H. P. Bryant, 1966. Hedium without rumen f l ui d f o r t he nonselective enumera t ion and isola t ion of rumen bacteria. App l . Hi crobiol . 14 :794-80 1.
Fox, G. E., E. Stackebrandt, R. B. Kespell, J. Gibson, J. Man i loff, T. A. Dyer, R. S. Wolfe, W. E. Balch, R. S. Tanner, L. J. Magrum, R. Blakemore, R. Gupta, L. Bonen, B. J. Lewis, D. A. Stahl, K. R. Luehrsen, K. N. Chen, and C. R. Woese. 1980. The Phy l ogeny of prokaryotes. Sc i ence . 209:457-463.
Gardner, R. H., K. C. Doerner, and B. A. White. 1987. Pu r ifica ti on and character izat i on of an exoglucanase from Rum i nococcus flayefaciens FD-1. 87th Annual Meeting of the AS H. p.56 (Abstract).
Gibson, T. and R. E. Gordon. 1974. Bergey's Manual of Dete r mi native Ba ter i ology, 8th Ed. Williams and Wilkins Co., Baltimore , Maryla nd. p 565.
Hall i wel l , G. and M. P. Bryant. 1963. The cellulolytic ac tivity of pure s t rains of bacteria from the rumen of cattle. J . Gen. Microbiol. 32:441-448.
Hall s , L. K. and R. Al caniz. 1972. Growth patterns of deerbrowse plants in southern forests. U.S. For. Servo Re s . paper. so- 75:1-14.
Hargreaves, K. R. 1971. Const j tuent~ of Euonymous eurogeas. Phytochem. 10:898-899.
Haya s hi da, S. and K. Ho. 1986. Production and characterist i cs of Av i cel di s i ntegrat i ng endoglucanase from a protease-negative Humicola irisea var. thermoidea mutant. Appl. Envi r on. Mi crob i ol. 51 : 1041-1046.
Herrera, C. M. 1981. Are tropical fruits more rewarding to d i sperse rs than temperate one s ? Am. Nat. 118:896-907.
Hi l l, D. R. , J. R. McCurry, L. P. Elliott, and G. T. Howard . 1985. SEM observ ation s on the germin ~ti on of Euonymus amer i canus. El ec t ron Mi croscopy SOCiety of America. 43:508-509. (Abstract).
Hi ltner, P. and B. A. Dehority. 1983. Effect of soluble carbohydrates on digestion of cellulose by pure cultures of rumen bacteria. Appl. Environ. Hicrobiol. 46:642-648.

48
Holthuij zen, A.M.A. and T. L. Sharik. 1984. The avian seed dispersal system of eastern r ed cedar (Juniperus ylriniana). Can. J. Bot. 63:1508-1515.
Howard, G. T. 1986. Germi nation of the seeds of Euonymus arnericBnus L. with the us e of rumen fluid of cows. BIOS. 57:14-18.
Hungate, R. E. 1950. The anaerobic, mesophilic, cellulolytic bacteria. Bacteriol. Rev. 14:1-49.
__________ . 1966. The Rumen and its Microbes. Academic Press, Inc., New York. p. 8-91.
Janzen, D. H. 1970. Herbivore and the number of tree species in tropical forests. Am. Nat. 100:25-35.
Kremer, R. J., L. B. Hughes, and R. J. Aldrich. 1984. Examination of microorganisms and deterioration resistance mechanisms associated with velvetleaf see~. Agron. J. 76:745-749.
Leedle, J. A. Z., M. P. Bryant, and R. B. Hespell. 1982. Diurnal variations in bacterial numbers and fluid parameters in ruminal contents of an i mals fed low- or high-forage diets. Appl. Environ. Microbiol. 44:402-412.
Livingston, R. B. 1972. Influence of birds, stones and soil on the establishment of pasture juniper, Juniperus communis, and red cedar, ~. yirainiana in New England pastures. Eco. 53:1141-1147.
Monin, J. 1965. Comparison of the effect of freezing on the subsequent viability of embryos of Euonymus yulaaris Miller. Experientia. 21:213-214.
__________ • 1967a. Study on the formation of gibberellins in embryos of Euonymus europaeus in the course of stratification of seeds. Bull. Soc. Franc. Physiol. Veglt. 13:171-177.
__________ • 1967b. Effects of different dormancy breaking agents during the stratification of Euonymus europaeus L. seeds. C. R. Hebd. Seances Acad. Sci . Ser. D. Sci. Natur. 264:300-302. (Abstract).
________ ~. 1967c. Study of an inhibitor existing in dormant embryos of Euonymus europaeus L. C. R. Hebd. Seances Acad. Sci. Ser. D. Sci. Natur. 264:2367-2370.
__________ • 1967d. Action of some agents on the dormancy of Euonymus europaeus. C. R. Seances Soc. BioI. 161:576-579.

~ ________ • 1968. Study of the growt h substance content on Euocymus europaeus L. embryos in relat i onship to t heir dormancy. Bul l. So,c. Franc. Physiol. Veg~t . 14:25-29.
Nelson, N. 1944. A photometric adapt at i on of the Somogyi method f o r the determination of glucose. J. BioI . Chem . 53:375-380.
Nikolaeva, H. G. 1959. Fiziologiya prorastaniya semyan bereskleta. Referat. Zhur. BioI. No. 83858 .
Nikola eva, H. G., T. V. Daletzkaya, H. V. Razumova, and N. N. Kofanova. 1973. Effect of gibberellin and kinetin on growth of embryos and germination of seeds of EuoDymus europaeas and Acer tataricum. Fiz iol. Rast. 20:714-720.
Niko laeva, H. G. , E. N. Polyakova, T. V. Daletzkaya, V. N. Petrova, and N. S. Vo r ob'eva. 1974. Effect of beta-indoleacetic acid on seed germination and embryo growth and its interaction with other hormones. Fiziol . Rast. 21:919-926.
49
Omel ianski, W. 1902. Uber d i e Garung der cellulose. Zenlbl. Bakt. Parasi t kde. Abt. II. 8: 225.
Pettiphen, G. L. and H. J. Latham. 1979. Production of enzymes degrading plant cell walls and fermentation of cellobiose by Ruminococcus flavefaciens in batch and continuous culture. L. Gen. Microbiol. 110:29-38.
Pigl, L. Van Der. 1972. Principles of Dispersal in Higher Plants. 2nd ed. Spr inge r-Verleg, New York. 161 p.
Rogers, L. L. and R. D. Applegate. 1983. Dispersal of fruit seeds by black bears. J. Mamm. 64:310-311.
Rudolf, P. O. 1974 . Euonymus L. p. 393-397. In C. S. Schopmeyer (ed.), Seeds of Woody Plants in the United 3tates . Forest SerV i ce, U. . Department of Agriculture, Washington, D. C.
Russell, J. B. 1985. Fermentation of cellodextrins by cellulolytic and noncel l ulolytic rumen bacter i a. Appl. Environ. Macrobiol. 49:572-576.
St rau sbaugh, P. D. and E. L. Core. 1973. Flora of West Virginia, 2nd ed., Part III. West Virginia University Bulletin Series 74, No. 2-1 300 p.
Teather, R. H. and P. J. Wood. 1982. Use of congo redpolysaccharide interactions in enumeration and characterizati on of cellulolytic bacter i a from the bovine rumen. Appl. Environ. Hicrobiol. 43:777-780.

Wilson, C. L., W. E. Loomis, nd T. A. Steeves. 1911. Botany. 5th ed. Holt, Rinehart, and Winston, New York. p. 323-331.
50
Windharo, W. R. and D. E. Ak i n. 1984. Rumen fungi and f~rage fi~er degradation. Appl. E viron. Mi orob i ol. 48:413-416.