the dinerentiai migration lymphoeytos€¦ · dr. hay has fiiied the lab with many incredible...
TRANSCRIPT

The Dinerentiai Migration oCBlood andLymph Lymphoeytos
Timothy James Seabrook
A thesis submitted in conformity with the requirements
for the degree of Doctor of Philosophy
Graduate Department of Laboratory Medicine and Pathobiology
University of Toronto, 2000
O Copyright by Timothy James Seabrook 2000

Acquisîions and Acquisitions et 8ibliographTc Services services bibliographques 395 WellYigton SMntt 395. lue wdlington OüawaON KlAONI O(tawaON KlAONC Canada Canada
The author has granted a non- exclusive licence allowing the National Lib- of Canada to reproduce, loan, distniute or seil copies of this thesis in microfonn, paper or electronic formats.
L'auteur a accordé une licence non exchsive permettant à la Bibliothècpe nationale du Canada de reproduire, prêter, distniuer ou vendre des copies de cette thèse sous la fome de mictofiche/film, de reproduction sur papier ou sur fonnat électronique.
The author retains ownership of the L'auteur conserve la propriéte du copyright in this thesis. Neither the droit d'auteur qyi protège cette thèse. thesis nor substantid extracts fiom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced wïthout the author's ou autrement reproduits sans son permission. autorisafion.

Abstmct
Ih sheep, a pool of lymphocytes resides in the bIood that does not recirculate as
efficiencly as lymph lymphocytes. However, there is limited information on the
differential migration of blood and lymph lymphocytes into wnlymphoid tissues or
during infiammation. Therefore, experïments in this thesis were designed to investigate
the migration of blood and Lymph Lymphocytes after splenectorny, antigen challenge to a
single lymph node, into n o d cerebral spinal fluid (CSR, and aftet TNFa induced
inflammation-
Following neoaatal splenectomy, no merence in the number or phenotype of
lymphocytes was observed, However, splenectorny did sesuit in an increased migration
of lymph lymphocytes into lymph nodes and a trend towards a longer residence time in
blood for the blood pool of lymphocytes. Whüe splenectomy has little effect on the
development or distribution of lymphocyte subsets in blood and lyrnph, evidence was
obtahed that it affects the rate of Lymphocyte rec~cdation.
Lymph node shutdown was induced by PPD in a BCG immunised sheep. Both
lymph and blood pool CD4 lymphocytes increased in efferent lymph dwing Lymph node
shutdo wn. IFN-y and IL-6 IeveIs were uicreased in efferent lymph plasma during l p p h
node shutdown and may have a role in both the recruitment and retention of lymphocytes-
Lymph lymphocytes preferenwy migrate h to CSF and &erent lymph under
normal conditions, implying that iymph lymphocytes have a greater role in immune
surveillance as compared to blood lymphocytes. After the intracerebrovenaicular
infusion of TNF-a, blood lymphocytes are found in a greater percentage as compared to

l p p h lymphocytes. In both CSF and the perivascuiar spaces of the brain, an
innammatory innltrate composed of CD4, CD8 and y6 lymphocytes was observed.
Together these data demonstrate that under iaflammatory conditions and
fobwing splenectomy there are dinerences in the migration of blood and lymph
lymphocytes. The redts in this thesis provide some basic data into the mi-gration of the
blood and lymph pools of lymphocytes and provides the background for further
investigations into the migration and functional differences between blood and lymph
lymphocytes.

Dedication
This thesis is dedicated to my grandparents
Floyd and Gladys Seabrook
Ferman and Alice Hanes

Firstly, 1 must thank Dr* Jack Hay for hisencouragement and guidance over the
Iast 5 years- He is a wonderfiil mentor, in both science and mee 1 feeI privileged to have
had the opportunîty to work with him,
Dr. Miles Johnston has helped with many discussions on various papers,
experiments and letters of reference. 1 must also thank the rest of my commîttee for their
pidance over the years including Dr. M- Cybulsky, Dr. L. Becker and Dr. R. Midha.
Dr. Hay has fiiIed the lab with many incredible people over the iast 5 years and 1
have benefited fkom working with them- Without the assistance of WiU Andrade, Binh
Au, Jodi Dickstein, Bill Ristevski and Me1 Boulton, this thesis wodd not have k e n
completed. Catherine Munroe deserves special thanks for her help with figures,
references and editing. Diana Armstrong gave mmy hours of her time helping me with
swgeries.
In addition, 1 must thank Dr. A Young and Mk. Lisbeth Dudler at the Basel
Institute for Immunology. Because of hem, 1 had the chance to experïence not o d y a
wonderfiil research institute but also Switzerland.
Cheryl Smith was responsible for teaching me flow cytometry. But even more
important she shared coffee with me every morning at the MSB.
Frank, Ranier, Angela, Wendy and aii of the other people in the animal facitity
took excellent care of my animals.
Lastly, 1 must thank my family for their support over the years. Most of ail
without Darren this thesis would have k e n impossile. His understanding and patience
with my long nights in the lab and talkùig about science over dinner is incredible. Thank
you.

Table of contents
INTRODUCTION ......................................................................................................... 1 1-1 HISTOWCALBACKGROUND .............................................................................. 1 1.2 SIZE OFTHE LYMPHOCYTE POOL ............................................................................. 2
1.3 LYMPHOCYTE SUBSET DISTRIBU~ON OF LYMPHOCYTES IN SWEEP. ...................... ......3
1.4 MOLECULES WOLVED IN LYMPHOCYTE MIGRATION: AN OVERViEW ....................... 3
1.5 LYMPHOCYTE MIGRATION INTO LYMPH NODES ........................................................ 5
1.6 LYMPH~~YTEMIGRATIoN INTO THE SP LEEN .......................................................... 10 1 -7 LYMPHOCVTE MIGRAIION INTO THE CNS AND CSF ...............................-............ - 1 1
1.8 LYMPHOCYTE SUBPOPZ~LATIONS: PHENOTYPE vs . MIGRATION PATERBIS ............... 12
.................... 1.9 DEFIMTION OF BLOOD LYMPHOCYTES AND LYMPH LYMPHOCYTES .,... 14 1-10 RATIONALEFORTHEEXPERIMENTS~THISTHESIS .............................................. 17 1- 1 1 EXPERIMENTAL SYSTEM ................................................................................ 1 8
1.12 MAIN HYPOTEESIS ............................................................................................. 20
1.13 S T R U ~ OFTHETHESIS AND STATEMENT ..................................................... 2 1
2.1 ANIMALS ............................................................................................................. 22
2.2 S ~ G E R Y ............................................................................................................. 23
2.3 CELL LASELLJNG ................................................................................................. 24
2.3. 1 11 1-h .................................................. ,.. ................................................ -24
.......................................................................................................... 2.3.2 Sl.Cr... 25
2.3.3 FlTC labelling of blood ................................................................................. 25 2.3.4 FITC labelling of lymph lymphocytes ......... ... .......................................... 26 2-35 CFSE labeiiing of lymphocytes ...................................................................... 26 2.3.6 DiI-DS Iabelling of efferent lymphocytes ...................................................... 26
2.4 ANTIBODIES USED IN THIS TfIESIS .......................................................................... 28 ............................................................. 2.5 IMMUNOPHENOTYPING OF LYMPHOCYTES 28

2.6 CrTOMETRY ............................................................................................... 29
.................................................................................... 2-7 ~ O E ~ T O C H E M I S T R Y 29
2.8 REAGENTS ........................................................................................................... -30
2.8.1 Hank's balanced salt solution (EIBSS) ........................................................... 30
.............................. 2.8 -2 Phosphate buffered saiïne (PBS) without divalent cations 31
2.8.3 Dulbecco's phosphate buffered saline ............................................................ 31
2.8.4 PBSflween buffer .................................................................................. 3 1
2.8.5 ParaformaIdehyde solution ............................................................................ 31
2.8.6 Erythrocyte lysis solution ............................................................................. .32
2.8.7 Acid Citrate Dextrose ..................................................................................... 32
INVESTIGATIONS INTO THE MIGRATIONPA'ITERN OF BL AND LL INTO
AFFERENT LYMPH AND AETER SPLENECTOMY weoeeew.moe.~eew~woeeeweeo.eweowewoeeeee35
3.1 A~STRACT ........................................................................................................... 35
3.2 INTRODUCTION ..................................................................................................... 36
.................................................................................. 3 -3 MATE RIAL^ AND M~THODS 37
3.3.1 Animals and surgery ..................................................................................... 38
3.3 -2 Lymphocyte labeiiing ......................... ..... ............................................... 38
3.3.3 Sarnphg of blood and lymph for trackùig fluorescent labeUed celis .............. 39
3.3.4 Statistical analysis .............................................................*........................... 39
3.4 RESULTS .............................................................................................................. 40 3.4.1 Repetitive sampling of blood and lymph ........................................................ 40
3.4.2 Aflerent lymph .............................................................................................. 40
3.4.3 Splenectomized sheep migration .................................................................... 41
.................. 3.4.4 Immunophenotyping of lymphocytes in splenectomized sheep .... 4 L
3.5 DISCUSSION ......................................................................................................... 54

4.3.3 Ceilcollection andphenotyping ............................................................... ......60
.............................................................................. 4.3 -4 hunohistochemistry 6 1
4.4.4 Cytokine EUS& .......................................................................................... 61 . . 4.4.5 Stawtics ......................................................................................................... 62
4-5 RESULT~ .............................................................................................................. 62
4.5.1 Lymphocyte Migration ............................................................~..................... 62
4.5.2 Cytokine Levels ............................................................................................. 63
4.5.3 'Immunohistochemistry .................................................................................. 63
........................................................................................................ . 4.6 D~scvss~o~ 70
LYMPHOCYTE IN CEREBROSPINAL FLUID ARE PART OF THE
............................................................ RECIRCULATING LYMPHOCYTE POOL 74
5.3.1 Animais and surgery ...................................................................................... 76
5-3-2 Determining normal CSF ceii counts ............................................................. 76
................................................................................................. 5.3.3 Ceil l abehg 77
5.3.4 Sample collection of blood, lymph, CSF and lymph nodes .............................. 77
5.3.4 Intracerebroventricular infusions of 1 11-In labeiled lymphocytes ................... 78
..................... 5.3.5 Intracerebroventricuiar infusions of J3TC labeiled lymphocyte -78
5.3.6 Flow cytometry .............................................................. ... 5.4 m m .. .............................................-...................................... ................. 79
5.4.1 Cells in normal CSF of sheep ......................................................................... 79
5.4.2 Repeated infusion of labelled lymphocytes .................................................... 79

5.4.3 Singe bolus of labelled efferent Lymphocytes ................................ ............... 80
5.4.4 Kuietics of FITC labelleci ce& in CSF and efferent lymp h. ............................ 84
5.4.5 Lymphocyte egress fkomCSF ........................................................................ 84
......................................... 5.4.6 LL and BL migration into CSF and afferent Lymph 85
......................................................................................................... 5.5 Drscussro~ 93
TNF-a INJECTIONS INM CSF. BUT NOT THE BRAIN PARENCHYMA.
RESULTS IN LEUKOCYTE RECRUiTMENT ....................................................... 98
........................................................................................................... 6.1 A~STRACT 98
6.2 INTRODUCTION ..................................................................................................... 98
6-3 MATERIALS AND MEIHODS ................................................................................. 100
6.3.1 Animais and surgery ................................................................................... 100
............................................................................................... 6.3.2 Cell labeiling 100
6.3.3 TNF-a injections ..................................................................................... 1 0 0
6.3.4 CSF collection, differential and phenotyping ............................................... 101 . . 6.3.5 Bram tissue collection .................................................................................. 101
6.3.6 Immunohistochemistry ............................................................................. 101
6-4 RESULTS ............................................................................................................ 102
6.4.1 Leukocyte number, differential and phewtype after the intracerebroventrïcular
infusion of TNF-a ............................................................................................... 102
6.4.2 CSF leukocyte numbers after parenchyma injections of TNF-a ................... 102
6.4.3 BL and LL migration înto CSF after TNT-a injection .................................. 103
................................................................................ 6.4.4 Immunohistochemistry 103
. 6.4.5 The effect of intracerebrovenvicular injection of TNF-a on cervical Lymph .. 1 19
6.5 DISCUSSION ....................................................................................................... 119
GENERAL DISCUSSION ........................................................................................ 125

7-2 ~ T I G A ~ ~ N S INTO T H E M I W O N PA- OFBL AND LL NCO AEFERENT
LYMPH AND AFER SPLENECTOMY ............................................................................. 126
7-2-1 Repetitive samphg ..................................................................................... 126
............................................................................................ 7.2.2 Merent lymph 127
7.2.3 Splenectomized sheep ............................................................................... 128
7-3 THE MIGRATION OF BL AND LL TEiRoUGH ANTIGEN STIMULATED LYMPH NODES . 130
..................................................... 7 -4 LYMPHOCYTES IN CSF ARE PART OF THE RLP 132
7.4 TNF-a IJSDUCED CSF LEUKOCYTOSIS ................................................................. 133
7-5 FUllJRE EXPE~UMENTS TO D- THE EMS'ENCE OF A RAPIDLY RECIRCULATING
............................................................................................. EQOL OF LYMPHOCYTES 135
7.6 SUMMARY- ......................................................................................................... 139
REFERENCE LIST ............................................................... w........................~......... 140

LET OF FIGURES
F~GURE 1 NUMBER OF LYMPH~~YTES IN VARIOUS TISSUES AND POOLS IN S~EEP ........... --.-.6
~ G U R E 2 P H E N o ~ E OF S- L Y M P H ~ INLYMPH, BLOOD AND LYMPH NODES ..... -8
FIGURE 3 ~RESENTATIVE EXPERTMENT OFREPETLTIVE SAMPLING OVER A ~ ~ H R P E R I O D ~ ~
4 RATIOS OF BL AND LL LABELLED CELLS IN = E L Y M P H AND BUX)D OVER
A 27 HR P m O D ..................................................-..............o...... 9,.-.-..------.----..---.--.----45
FIGURE 5 SHORT-TERM MIGRATiON OFLABELLED LYMPHOCYTES IN SPLENECTOMIZED
SHEEP ....................................o................................................................-.......... --..--48
FIGURE 6 TISSUE LOCAUSATION OF RADIOLABELED LL AND BL LYMPHOCYTES. ,. .- .,. .---- 50
FIGURE 7 PPD INDUCED LYMPH NODE SHUTDOWN -,*.,,-----.,--..-,. ~~~..~~~----.---.--..-------.--------65
FIGURE 8 PHENOTYPE OF LYMPHOCYTES IN LYMPH DURING LYMPH NODE
SHUTDOWN ..................................................................................... ---.-------..-.--.----.--66
FIGURE 9 C Y T O ~ LEVELS DURTNG LWH NODE SHUTDOWN ...................................... 69 RGURE 10 PERCENTAGE OF LABELT.ED c m IN nssms AFIER CONSTANT REINFUSION -8 1
FIGURE 1 1 PERCENTAGE OF LABELLED LYMPHoCXES 2 4 ~ ~ AFïER A SINGLE INFUSION OF
c m .....-........................................................................................................ -.--.--Ai6
FIGURE 12 APPEARANCE OF FIII'C LABET.T.ED LYMPHOCYTES IN CSF AND LYMPH ,,.,.,-.,--88
FIGURE 13 ~~RACEREBROVENTRICULAR NEcïEJ3 1 1 1-1. LAB- CELLS MIGRATE TO
LYMPH NODES KNOWN TO DRAIN CSF ....................................o.............................--... 89
FIGURE 14 CSF C E U ~ ~CREGSES m THE INJECTION OF TNF-a .................. 105
FIGURE 15 BOTH BL AND LL INCREASE AFïER THE INTRACEREBROVENTRTCULAR
INJECTION OFTNF-a ......................... -, ................................................................ 108
FIGURE 16 CNS PARENCHYMA AFER AN INTRACEREBROENTRICULAR INJECTION OF
................................................................................................................. TNF-a.. 1 1 1
FIGURE 17 CHOROID PLEXUS A F E R AN INTRACEREBROVENTRICULAR INJECIION OF
--a .............................................................................................................. 113
FIGURE 18 CNS PARENCHYMA AETERTHE INTRACEREBRAL INJECITON OF m - a 1 15
~ G U R E 19 ~ C E R E B R O V E N T R I C U L A R INJECIION OF --a HAS NO EFFECT ON
CERVICAL LYMPH FLOW OR CELLULARITY ......................................... . . . . . . . 1 1 7
- fi-
- - -

List of Tables

List of ab breviations
Ag
ANOVA
APC
BCG
BBB
Bn:
BL
BLC
CD
CFSE
CM-DiI
C N S
CSF
ELISA
DiI-DS
DMSO
EAE
FITC
GlyCAM
HBSS
HEV
antigen
analysis of variance
diop hycocyanin
Bacillus Calmette- gué^
blood brain barrier
Basel Institute for lmmunology
blood lymphocyte
B-lymphocyte chemoattractant
cluster of differentiation antigen
carboxy-îiuorescein diacetate succinimidyl ester
CeliTracker carboc yanine fluorescent dye
central nervous system
cerebral spinal fluid
enzyme iinked immunoassay
lipophilic, carbocyanine fluorescent dye
dimet hylsulfoxide
experimental autoimune encep halo myelitis
fluorescein isothiocyanate
glycosylation-dependent ceU adhesion molecule
Hank's balanced salt solution
high endothelia1 venule

ICAM-i
IFN-Y
IL
LFA- 1
LL
mAb
MAdCAM
MHC
MS
OCT
PBS
PKH
PPD
rhTNF-a
RLP
SLC
TNF-a
VCAM- 1
interceiIular adhesion molecule
interferon gamma
interleukin
leukocyte hinction antigen- 1
lymph Lymphocytes
monoclonai antiïdy
mucosal addressin ceU adhesio n mo lecule
major histocompatibility complex
multiple sclerosis
optimal cooling temperature
phosphate bUnered saline
lipophiiïc, carbocyanine fluorescent dye
purified protein derivative
recombinant human tumour necrosis factor-a
recirculating Lymphocyte pool
secondary lymphoid tissue chemokine
tumour necrosis factor alpha
vascular cell adhesion moiecule-1

Chapter 1 Introduction
Lymphocytes are unique in their ability to continuously recircuiate fkom blood,
into tissues and retum to blood via the Ipphatic system (Young et aI., 1993a). This
process is important for the dissemination of immunological memory and immune
suweiiiance. Lymphocytes recirculate through Lymph wdes thereby allowing the
presentation of antigen (Ag) to a large number of Lymphocytes. Once a lymphocyte
recognises its Ag and receives the relevant costimulatory signals, an Ag specsc h.mune
response begins. Effector celis exit the lymph node via the efferent lymphatic and
migrate to sites of inflammation (Picker and Siegelman, 1993). Once the immune
response has been resolved, a subset of Lymphocytes become memory cells which
concinuously recirculate through the body ailowing for a rapid secondary immune
response (Sprent, 1994; Butcher and Picker, 1996).
1.1 Historical Background
Gowans, in a series of seminal experiments elucidated the fùndamentals of
lymphocyte recirculation. He and colieagues demonstrated that cannulating the thoracic
duct in rats and diverting the Lymph resulted in a decrease in the number of lymphocytes
in both blood and lyrnph (Gowaas and Knight, 1964). When lymphocytes were Iabelled
with radioisotopes, and intravenously injected, labelled celis could be found in l p p h
(Gowans, 1959). Taken together these experiments demonstrated that the lymphatic
system was responsible for the retum of lymphocytes to blood. Morris and severai
collaborators developed surgical protocols that alio wed the cannulation of Lymphatics and

the continuous collection of lymph in sheep (LasceIles & Mom-s 1961; Smith et al.
1970). This development permïtted the quantincation of lymphocyte t r a c through an
isolated lymph node under normal and inflammatocy conditions. This led to severaï
important discoveries including the preferentiai recirculation of memory lymphocytes
through tissues (Mackay et al., 1990) and the quantincation of lymphocytes produced and
exported de novo in a lymph node during an immune response (Hall and Morris7 196%).
1.2 Site of the lymphocyte pool
There is approximately 1 kg of lymphoid tissue in an adult sheep, containing
approximately 10" lymphocytes (Chin et ai., 1985). %y canndating the thoracic duct
lymphatic, it was demonstrated that approximately 10% of the total lymphocyte
population, or approximately 10'' cek , compose the recirculating lymphocyte pool
(RLP) (Schnappauf and Schnappauf. 1968) (Figure 1). The RLP is comprised of
Lymphocytes that conùnuously recircuiate through the body, while the remainder may be
fixed in tissues or do not recircuiate under normal conditions.
One defi t ion of "pool" is a common supply of a commodity for sharing amongst
a group e.g. a pool of money for a department. Pool in this thesis refers to a population
of lymphocytes that preferentially localise, migrate or are retained in a specific tissue
cornpartment. For example, ali Lymphocytes that recirculate are part of the RLP but this
pool is further subdivided based on preferential tissue migration includuig a lymph pool
of lymphocytes (discussed in detail in section 1.8).
Blood contains approximately 1% of the total lymphocyte pool at any one time
(Figure 1) (Schnappauf and Schnappauf U, 1968). When arterial blood passes through a

lymph node approxmiately one quarter of aü Lymphocytes are extracted by the poa
capiUary vendes B a y and Hobbs, 1977). These Lymphocytes migrate into the lymph
node and exit via the eEerent Lymphatic. The Lymph node also receives Lymphocytes
fiom the tissues via afferent lymphatics, which contain approximatefy 1 x 106 ceWmL
Efferent lymph bas ten times the amount of Lymphocyteslml. Previous studies by Hail
and Morris have demonstrated that fewer then 4% of Lymphocytes present in efferent
lymph are produced de novo in the lymph node (HaU and Morris, 1965a). Therefore,
approximately 90% of lymphocytes present in efferent Iymph migrate directly from the
blood-
1.3 Lymphocyte subset distribution of lymphocytes in sheep
In sheep, there are differences not only in the number of Lymphocytes but also the
phenotype of lymphocytes in the blood, efferent and afferent lymph (Figure 2) (Mackay
et al., 1988). Approximately 10% of afferent lymph in sheep is composed of dendritic
ceiis and other ceiIs of monocytic origin (Haig et aL, 1999). Efferent lymph is virtually
1 0 % lymphocytes, the majority king small resting ceils. In sheep, lymphocytes rnake
up approximately 5060% of the Ieukocytes present in blood (BluntJ975 and my own
observations). The topic of lymphocyte phenotypes is fbrther discussed in section 1.8.
1.4 Molecules involved in lymphocyte migration: an ovewiew
Lymphocytes migrate into tissues fkom the vascular system ushg various
adhesion molecules includïng selectins, integruis and their ligands in a CO-ordinated

series of events (Butcher et ai., 1999; SpMger, 1994). Broadly speaking this can be
divided into discrete steps ùicluding, primary adhesion ( r o b g and tethering). integrin
activation, firm adhesion and tran.smïgration.
Primary adhesion is a reversible process in which lymphocytes transientiy adhere
to endothelial ceiis, allowing lymphocytes to be acted upon by chemokines and other
activating substances. Selectins (Gaiiaùn et ai., 1983) and a4 containing integrins
(Berlin et al., 1995) are responsible for primary adhesion. Selectins are membrane bound
Iectins present on the surface of both endothem ceUs and lymphocytes and bind to
mucin like proteins (Vestweber and Blanks, 1999). L selectin is expressed on the
micovilli of lymphocytes (Stein et ai., 1999) and binds to endothelial cells via several
Ligands, including CD34 and glycosylation-dependent celi adhesion molecule (GlyCAM)
(Vestweber and Blanks, 1999). E and P-selectin are expressed on endothelid celis and
have an important role in the recruitment of Lymphocytes into inflmed skin (Austmp et
ai., 1997).
Integrin activation is a critical step in lymphocyte migration that involves
chemokines bound to glycosaminoglycans on endothelial cells (Koopmann et al., 1999)
and other activating substances including platetet activating factor (Kim and Broxmeyer,
1999). Chemokines are small chernotactic proteins produced by a number of c e k
(Campbell et ai., 1996). They interact with G protein coupled receptors, causing the
activation of integrins on the Lymphocyte surface (Taub and Mqhey, 1997). Ligation of
L-selectin after binding its ligand also activates integrins (Hwang et al., 1996). htegrins
are composed of noncovaiently associated a and chahs and are critical in the firm
adhesio n and transmigration of lymphocytes. Upon activation integrins undergo a

conformational change, thereby increasing their amty for Iigands which are members
of the immunoglobuiin superfamily (Springer, 1995). Lenkocyte fimctionai antigen- 1
(a&) which binds to intracellular adhesion mlecuie-1. 2 and 3, is important in
lymphocyte m*gration into ineammatory sites and Iymph nodes (Butcher et ai.. 1999)
Transmigration involves the lymphocytes migrating between endothelial ceiis and
adhering to structural proteins. This step involves CD31 and some cytoskelatai
rearrangements on the part of the endothelial cells (Zocchi et al, 1996; Allport et al-,
1997). The lymphocyte now migrates into the tissue underlying the endothelid ceiL
1.5 Lymphocyte migration into lymph nodes
Migration of lymphocytes into lymph nodes is dso highly regulated-
Lymphocytes continuously recirculate through Lymp h nodes using various adhesion
molecules but there are several ciifferences fiom the general pattern as discussed above.
Firstly, in most species Lymphocytes migrate into lymph nodes using high endothelial
venules (HEV) which are h e d by cuboidal endothelial ceiis that are n o d l y present
only in iymph nodes (Girard and Springer, 1995). Recent studies using molecular
biology techniques have demonstrated that these endothelial cells preferentiaIly express
unique genes (Izawa et al., 1999; Girard et al., L999). L-selectin is a critical adhesion
molecule for the normal migration of lymphocytes into lymph nodes as demonstrated by
the impaked traffic in L-selectin kwckout mice (Asbones et aL, 1994). GlyCAM and
CD34 are expressed on the HEV of peripheral lymph nodes and are ligands for L-selech
(Vestweber and BI&, 1999).

Figure 1 Number of lyrnphocfles in various tissues and poois in sheep
The data to construct this figure carne fiom several different sources as outluied in the
discussion. Data representing bone rnarrow is not available. Although some data has
k e n descriid in young Iambs for thymic output (Cahili, personal communication) it has
k e n excluded here because older animals were used.

? nurnber of lymphocytes Non HEV containing tissue
Blood 1 x 10 El 0 total lymphocytes in blood
Recirculating lymphocyte pool approx 1 x lOEll
Total lymphocytes in 30kg sheep approx. 1 x 10E12
1 ? Number of lymphocytes
Spleen
5 x 10E10 in total
afferent ly mphatics
2 x 1 0E9 per hour migrate into the lyrnph node via HEV
1
70 g in total
efferent lymphatics - Thoracic duct
2 X 10E9 per hour

Figure 2 Phenotype of sheep lymphocytes in lymph, blwd and lymph c d e s
The phenotypic data in this diagram was obtained nom this thesis and firom the literature
referenced in the text above-
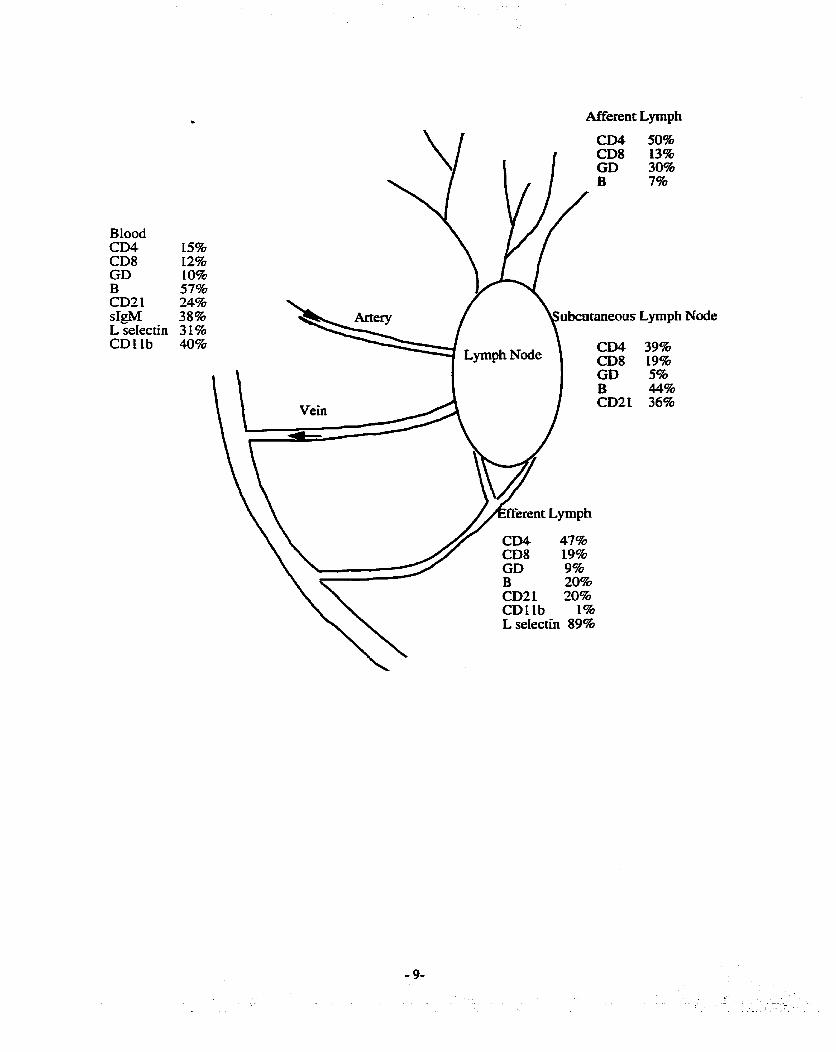
BIood CD4 CDS GD B CD2 1 sIgM L seIectin CDLlb
Afferent Lymph
CD4 50% CD8 13% GD 30% B 7%
15% 12% 10% 57% 24% 38% ubcataneous L p p h Node 31% 40% CD4 39%
CD8 19% GD 5% B 44% CD21 36%
CD4 47% CD8 19%
CDL1b 1% L selectin 89%

Pemissis toxin inhibits the migration of lymphocytes into Lymph nodes, thereby
implicating a G protein coupled receptor in the migration of Lymphocytes into lymph
nodes (B argatze and Butcher, 1993). Recently a chemokine, secondary lymp hoid tissue
chemokine (SLC), was shown to be expressed in E V and T ceil areas within the lymph
node (Warnock et aL, 2000): This chemokuie, with others, may CO-ordhate the migrafion
of Lymphocytes into and their position within the lymph w d e
To date little is known about the mechanismi and adhesion molecules involved in
the migration of lymphocytes within Lymph nodes. AdditionaUy, the signals that are
responsible for the exit of Lymphocytes are relatively unknown.
1.6 Lymphocyte migration into the spleen
The spleen receives fiom and rehuns to the blood more lymphocytes than any
other organ in the body (Pabst and Westermann, 1991). Lymphocytes migrate directly
out of the blood into the white puip without crossing HEV. Unlike lymph nodes
lymphocytes migrate directly back into the blood with only a srnail proportion entering
splenic lymphatics (Pellas and Weiss. 1990). Therefore, lymphocyte migration into the
spleen and their retum to blood is not considered recirculation as defmed in this thesis-
SLC, B-Lymphocyte cheumattractant (BLC) and other chemokùies act in concert
to regulate the migration of lymphocytes nom blood into the white pulp of the spleen
(Liadhut et al, 1999). Chemokines also CO-ordinate the migration of lymphocytes into
their respective niches, for example BLC attracts B celis into follicles (GUM et al., 1998).

1.7 Lymphocyte migration into the CNS and CSF
There is Iittle normai migration of nonactivated Iymphocytes h o the centrai
nervous system (CM) across the intact blood brain barrier (BBB) (Hickey, 1991;
Wekerle et al., 1986). T'bis has been examined using a variety o f experlmentai protocols
including immunohistochernistry (Hickey et ai., 199 1) and radiolabeled (Raine et aL,
1990a) ceus, both of which demonstrate few lymphocytes within the CNS parenchyma.
However, there are a smaU number of Iymphocytes present in normal cerebrai spinal fluid
(CSFI-
During most infiammatory conditions there is a ciramatic increase in lymphocytes
and monocytes within the C N S (Raine, 1994; Anthony et al., 1998). In both multiple
sclerosis and its animal model experimental autoimmune encephdomyeiîtis (EAE),
lymphocytes and monocytes accumulate in perïvascuiar spaces of the brain. The initial
migration of Lymphocytes across the idamed BBB uulises the same adhesion molecules
as seen in the periphery with the exception of selectins (Cannella and Raine, 1995;
Engelhardt et ai., 1994). To date only one putative BBB specifc endothelial adhesion
molecule, recognised by the 4A2 antibody, has been demonstrated (Male et al., 1995).
Leukocyte entry into the CSF has not been as extensively studied as entry into the
CNS. Indeed, the exact pathway by which lymphocytes enter the CSF has not been W y
eluc idated. Under inflammatory conditio os, it appears that Lymphocytes c m directly
enter the subarachnoid space by traverskg venules in this area Another potential route is
across the BBB in the CNS parenchyma into the perivascular sheath of fluid that
surrounds the vessels and eventually joins the CSF (Weller, 1998). Another pathway

may be via the c~cumventricular orgaos and the choroid plexus, which lack a typical
BBB (Engelhardt and Risau, 1995)-
The adhesion moIecdes required for Lymphocyte migration into CSF may wt be
the same as those required for CNS migratioa It has been demnstrated that in EAE,
selectuis are not important (Engelhardt et al-, 1997), however in meningitis the blocking
of selectins attenuates the number of cells in CSF (Tang et ai., 1996). Injections of
cytokines into the CSF causes an influx of ceus (Sankkonen et al., 1990; Quagliareiio et
al., 1991; RamiIo et aL, 199û), but when the same cytokines are injected into the CNS
parenchyma few leukocytes are recruited (Scbneil et aL, 1999; Andersson et al., 1992)-
Taken together these data demonstrate that the CSF and the rest of the CNS differ in their
response to infiammation,
1.8 Lymphocyte subpopulations: phenotype vs. migration patterns
Lymphocytes can be divided into subpopulations based on different attributes
inc luding the tissue or organ of extraction, phenotype, funct ion, migration patterns,
expression of adhesion molecules, etc. Immunophenotyping is a common method that
uses the expression of specific surface markers to classify lymphocytes by using
monoclonal anubodies against these marken. This method is used in the cluster of
differentiation (CD) classification of surface markers. Ho wever, heterogeneity ofien
exists amongst Lymphocytes defhed on the basis of a single phenotypic marker.
Combinations of attniutes are often used to subdivide lymphocytes based on multiple
characteristics.

The function of lymphocytes is utiiised to classify cells and is often used in
conjunction with their phenotypic profile, an example is the Thl, Th2 systern This
system is based on the heterogeneity of cytokuie production by T helper c e h and their
effect on the immune response. The CD4 antigen is the e s t defrnlng charactenstic and
then fuaher subdivision is based on functioa For exaniple, Th1 CD4 c e k are
Lymphocytes that express CD4 and influence other immune ceiIs by producing various
cytokines including IFN-y. This cytokine production has a role in skewhg the immune
response to wards a ceU-mediated immune response (Sailusto et al., 1999). Th2 CD4 cells
have a role in inducing a humoral respoose.
Tissue specific is another marner to subdivide lymphocytes and is
based on the observation that Lymphocyte recirculation is not random Lymphocytes
isolated fiom the efferent lymph of lymph nodes draining specific tissues preferentiaiiy
r e m to those lymph nodes. There are populations (or pools) of gut and skin migrating
lymphocytes (Cam et al., L977; C6in and Hay, 1980). W e this type of lymphocyte
migration has been studied more extensively in sheep than in other species (Abemethy
and Hay, 1992), results fkom mice confirmed these hdings. It has been detennuied that
most of the organ specifc migration is due to memory T ceus (Williams and Butcher,
1997)-
Several papers have used the expression of adhesion molecules to d e h e
migration patterns (Mackay et al., 1992a; Abitorabi et al. 1996)- Investigators have
identifïed the a4B7 integrin as being cntical for the ability of Lymphocytes to migrate
through gut associated lymph nodes, whilst L selectin is important for their abiüty to
migrate into nibcutaoeous lymph nodes (Mackay et al., 1996 Abitorabi et al-, 1996)-

Thus, these studies have further defïned tissue fioming using adhesion molecules.
However, few studies bave examined if lymphocytes are 'fiozen' with these adhesion
molecules or if they can change.
There are reports to suggest that in rats the expression of specific adhesion
molecules may not be an absolute predictor for tissue or lymph node homing
(Westermann et ai-, 1994a; Wdter et al., 1995;). The dlscrepancy in findings maybe due
to the different species (rat vs. mice and sheep), the source of ceils (Lymph nodes vs.
lymph), the age of the animel (young vs. aged) and living conditions (pathogen free vs.
normal housing).
Taken together these data demonstrate that there is tissue specific migration of
lymghocytes in both mice and sheep. Much of this migration is the property of memory
T ceils, which display specific adhesion molecuIes, One hypthesis States that rnemory T
celis home to the tissue in which they 6rst encountered their antigen, dowing for a rapid
secondary immune response (Williams and Butcher, 1997). Ho wever others have
questioned this hypothesis (Westemiann and Pabst, 1996). Nevertheless, the
preponderauce of data in mice and sheep demonstrates that there is a population of
lymphocytes that experiences tissue specific homing and that adhesion molecuIes have a
role in this phenornenon.
1.9 Definition of blood lymphocytes and lymph lymphocytes
Previous studies using sheep have demonstrated a population of lymphocytes
present in blood that does not recirculate as efficientiy through lymph nodes as compared
to efferent lymph lymphocytes. This pooriy recirculating blood pl of lymphocytes
(BL) was detected by labelhg blood and lymph lymphocytes with different fluorescent

dyes and then reinfushg them simultaneously (Young et aL, 1997a; Andrade et al,
1998). These experknents demonstrated that iabeiIed Iymph lymphocytes (LL) were
e ~ c h e d in efferent lymph as compared to bIood, whilst Iabelled BL were concentrated in
blood. Based on these data it was concluded that blood contains a pool of lymphocytes
that does not recirculate as competently through lymph nodes as do LL.
Further studies demonstrated that BL are found in the spleen in greater numbers
than LL (Young et al., 1997a). Phenotypicaiiy, this pool is composed mainly, but not
exclusively, of B c e k that are CD21- and L selectin low, with smaller populations of
CD4, CDS and perhaps y6 T ceiis (Young et ai., 1997a). Recently m e r studies have
demonstrated that BL B ceiis are aiso CDS+, CDLlb+ and surface IgM high (Chevallier
et al., 1998; Gupta et al., 1998). To date, no unique cell surface antigen common to BL
has been found, though this is an area of active research in other laboratories.
Nor has a unique function been found for BL Lymphocytes. It has been speculated
that the B ceiis in BL are BL iïke cells and therefore produce low aff.iinity anti'bodies to
some bacteria (Chevallier, et aI., 1998). Others have theorised that they may have a role
in the imrnunity against blood borne infections (Andrade, 1996). Neither of these
hypotheses has been proven and pnor to the studies in this thesis, any functional
difference between BL and LL remained to be determined-
BLood contains not only BL but also LL in transit. When the experiments in this
thesis were designed various methods were considered to isolate or enrich BL to enable
more definitive experiments. One isolation method examined was panning or magnetic
bead separation protocols. It was decided that this could introduce several problems that
are inherent with ex vivo manipulation of ceiis, such as inadvertent activation of

lymphocytes- As weii, it is difîïcuit to isolate suscient lymphocytes necessary for
effective tracking studies in sheep. Additiondy, isolathg the B ceils alone wodd have
ignored the other subsets present in the BL. Since dead or riamaged lymphocytes do not
recirculate (Andrade 1996), it is important that the isolation procedures are not
excessively long or damagkg to the lymphocytes.
Therefore, similar protocols were foUowed as were used in earlier experïments
(Young et ai., 1997a; Andrade et aL, 1998). S e e s of blood and lymph were coiIected,
Iabeiled with different fluorescent dyes, reinfused intravenously and then subsequently
identified using flow cytometry. This allowed for the examination of many of the subsets
of lymphocytes present within the BL. This approach has been successhilly used in rats
to e x m e the effect of IFN-y on the migration of various lymphocyte subsets
(Westermann et ai., 1994b). However, the extensive number of shared properties
between BL and LL often complicates the simple interpretation of the data
Therefore, in this thesis, the dennition of the blood pool of lymphocytes (BL) is:
Those lymphocyresfkom blood rhat when labelled and reinfused intravenously do not
recirculate a s efficiently as a sample of Iymph lymphocytes simultaneously CO-infused-
The definition of the l p p h pool of lymphocytes (LL) is: Those lymphocytes, that
when labelled and reinfused intravenously c m recirculute and be found in Herent
lymph.
These dennitions are based on migration patterns seen after Iabelling ce& fiom
blood and lymph, reinfushg them and t r achg their migration. This is similar to tissue
specific homing patterns previously described for gut and subcutaneous Lymph node
lymphocytes (a; and Hay, 1980; Cahili et ai., 1977).

1.1 0 Rationak for the expewiments in this thesis
In a clinical setting* blood is ofien sampied to determhe the immunologicd health
of an individual. However, it has been shown that this is not always an accurate
reflection of the immune system as a whole (Westermann and Pabst, 1990). In human
immunodeficiency virus-affected patients, the blood shows a decrease in the number of
CD4+ lymphocytes but in lymph nodes, no decrease in this subset is found (Pabst and
Rosenberg, 1998). Blood contaios only 1% of the total lymphocytes in the body*
therefore the loss of blood CD4 cek may be £imctionally insignificant. Nonetheless,
blood WU continue to be sampled in the foreseeable future due to its ease of collection.
The data fiom this thesis may help in the interpretation of blood samples by detahg
some of the differences between lymphocytes found in blood and the rest of the body.
Using sheep it is possible to retrieve sarnples fiom several different tissues
including spleen, blood, lymph nodes, CSF and both afferent and efferent lymph-
DifTerences may exist in the migration of BL and LL into these tissues, however io date
this migration has not been examined In sheep there is a ciifference in the migration of
lymphocytes obtained fkom iyrnph nodes as compared to efferent lymph Lymphocytes
(Reynolds et al., 1982). Indeed Williams and Butcher (1997) argue that the different
migration pattern of Lpph node and efferent lymph lymphocytes may be partially
responsble for the controversy surroundhg tissue specific homing.
As weIi, several investigators, including our laboratory, are interested in labelling
lymphocytes with contrast media and using them in a clinical setting (Sipe et aL, 1999;
Bulte et al, 1992). This would aliow for the non-invasive trackuig of lymphocytes and

may have a roIe in revealing infIammatory lesions such as early plaques in multiple
sclerosis. However, basic migration patterns of blood derived lymphocytes need to be
elucidated to allow this tracking of blood lymphocytes to pmceed.
Specific studies in this thesis examine the effects of splenectomy on BL-
Splenectomy is known to have an effect on Lymphocyte number and function in patients
(Ferrante et al.. 1987; Sieber et al., 1985). nie experïments in this thesis tested if these
abnormalities maybe a result of changes in BL lymphocyte numbea or migration.
The migration of lymphocytes into the CSF may have a role in the immune
surveillance of the C N S under normal and idammatory conditions, such as menhgitis.
Therefore, experiments were performed to determine the migration of both BL and LL
into this important tissue-
The experiments in this thesis were designed to investigate ciifferences in the
m i m o n of blood and lymph lymphocytes under a variety of conditions. These
experiments were designed to explore both the basic biology of BL and how this may
impact on patients in a clinical setting.
1.1 1 Experimental System
Sheep were utilised in all of the experiments reported in this thesis for a number
of reasons. Foremost amongst them is the ability to chronicdy cannulate lymphatics and
couect lymph tkom an anùnal with a relatively intact lymphatic system (Smith et d,
L970; Cahili et aL, 1974; Young et al., 1997b). This ability allows for experiments that
examine the dynamic nature of lymphocyte recirculation. Lymphocytes c m be re-infused

into a sheep and their migration monitored in severaI co~artrnents both sequentially and
simultaneously, which can not be pedonned in smailer animais-
Mice, rats and rabbits have ail been used to investigate lymphocyte migration into
CSF, but because of their size, limited numbers of cells are avaiiable for examination.
Sheep are large enough that snfficient CSF can be collected and examuied for labeiïed
cells. Lastly, there is a large body of literature examuiing lymphocyte migration in sheep
upon which the present experiments are based (Mackay, 1988; Mackay, et al., 1992a;
Young, et al- 1993b: Seabrook et al-, 1999)-
Fluorescent labehg of cells and their detection using flow cytornetry was chosen
for several reasons. Fluorescent labelled Lipophilic dyes including CM-Da, PKH and
DiI-DS, are retained within the cytoplasm of Lymphocytes for weeks (Salvato et aL 1996;
Andrade et al., L996a; Young and Hay, 1995) and do not affect the migration of labelled
cells from blood into lymph (Teare et al., 199 1). Therefore, these labels allow the Iong-
term tracking of lymphocytes in vivo. Fluorescein isothïocyanate (FITC) has been used
for several years to label lymphocytes and does not impair the migration of Lymphocytes
(Butcher et ai., 1980). Unfortunately, this dye is not amenable for experiments longer
than approximately 2 weeks as its intensity decreases and it can no longer be detected. A
method in which whole blood is labelied with FITC (Andrade et al., 1996b) was selected
and used in this thesis as it allows the rapid labelling of large numbers of BL with little
manipulation.
Radioisotopes have been used in the past as whole tissues can be easily assessed
for the number of labelled tek (Issekutz et aL, 1981). However, the isotopes leach fiom
cells Iuniting the duration of experiments (Issekutz et al., 1980). Radioisotope labels are

superior to fluorescent labels for investigatùig lymphocyte migration into several tissues
simultaneousLy at necropsy and those tissue that are difficult to isoiate lymphocytes ftom
such as skin and Peyerrs patches.
Fluorescent dyes have the advantage that immunophenotyping can be carrïed out
on labeiied ceiis in blood and lymph. However, care must be taken in the labelling of
lymphocytes with fluorescent compounds as prolonged incubation will overlabel the cells
and affect their ability to migrate. Additionaii~~ the PKH class of compounds must be
used with diluents that can aggregate Iymphocytes if the incubation is prolonged (Salvato
et al., 1996). To ensure that the i a b e h g protocol does not adversely affect the migration
of lymphocytes, recovery data is obtained. Recovery data is based on the number of cells
infused and the number which subsequently is recovered fkom a specifïc tissue. This is
often expressed as the percentage injected and ailows the comparisoa between different
labels and experiments. Ali labehg procedures in this thesis were previously published
and/or validated to ensure that they had no effect on the migration of labelled
lymphocytes fiom blood to Iymph.
1.1 2 Main Hypothesis
Published reports have demonstrated that, in sheep, a pool of lymphocytes exists
in blood that does not migrate into the lymphatic system as efficiently as the lymph pool
of lymphocytes. We hypothesised that dinerences in this migration may be exaggerated
by experimental manipulation and therefore could Iead to a better description of the
functional significance of these ciifferences, if any, between these two pools. Therefore,

experiments using splenectomy, antigen challenge of a single Lymph node, examhation
of CSF and r e c r u i e n t with TNF-a were perfomied to test this hypothesis.
1.1 3 Structure of the thesis and statement
Chapter 2 descn i s the methods and materials common to several of the
experiments in this thesis. This limits some of the redundancy fi-om subsequent chapters
but some methods are repeated as 1) there were some minor modifications of standard
protocols and 2) some chapters are presented in manuscript f o m Chapter 3 addresses
some of the basic questions of BL migration including migration into Serent lymph and
the effect of splenectomy. Lymph node shutdown and its affect on BL migration is
discussed in Chaper4. Chapters 5 and 6 investigate the migration of BL and LL into
normal and innamed CSF. Finaily a general discussion, including experiments to
examine the existence of a rapidly recirculating lymphocyte pool is included in Chapter
7.
The experiments in this thesis were performed at Sumybrook Health Science
Centre, Medical Sciences Building, University of Toronto and the Basel Institute for
Immunology. Ail experiments in this thesis were perfonned by myself. However,
surgeries conducted at the Basel Institute for Immunology were perfomied by Dr. W.
Hein, Dr. A- Young and Ms. L. Dudler.

Chapter 2 Methods and Materials
Several methods and techniques are common to many of the experiments and are
discussed in this chapter. Methods that are specific for panicular experiments are
included in subsequent chapters.
Outbred femaie sheep of between 30 and 35 kg were used for ai i experiments
performed in this thesis. Exceptions to this were the older sheep used in the
splenectomized experiments and a single maIe sheep in the antigen stimulation
experiments. Sheep were obtained iÏom 3 sources depending on the site at which the
experiments were performed. Suppiiers included Ledo Farms (Oshawa ON), Bowood
Farms (London ON) and Versuchsbetrieb Semweid (Olsberg, S witzerland). AU s heep
had fiee access to hay, pellets and water at aL times except for the 24 hows imrnediately
preceeding surgery.
Experiments conducted at the Division of Comparative Medicine, University of
Toronto, were approved by the Animal Care Committee of the Faculty of Medicine- The
experiments in Chapter 5 were approved by the Animal Care Committee of Sunnybrook
Heaith Science Centre. Al1 were in accordance with the Canadian Council on Animal
Care and the Anùnals for Research Act of Ontario. For those conducted at the Basel
Institute, Switzerland, handihg and treatment of the animais was accordùig to protocols
approved by the regional government authority, the Kantonales Veteriniiramt.

2.2 Surgery
AU surgicd procedures were pedormed uader steriie conditions. Animais were
anaesthetised with sodium pentothd to effect and were intubated with an endotracheal
tube. A surgical plane of anaesthesia was maintained with either halothane or isoflwane
in oxygen with the aid of a respirator. AU surgical techniques have k e n previously
described (Young et al., 1997b)- Briefly, a catheter attached to a 3-way stopcock, was
surgically placed into one of the jugulas vehs for blood sampluig. Lymphatic vessels
canuiated included the prescapular, prefemoral and cervical efferent lymphatics and
hindlimb merent lymphatics. Anatomicaily these are distinct lymph nodes but the
surgical manipulation required is the same for alI lymph nodes. The efferent lymphatic
was exposed with a minimum of trauma, and a section approximately 3 cm in length
stripped of adherent fascia A silk suture was used to Ligate the efferent lymphatic
downstream fiom the proposed incision site, aiiowing the lymphatic to &te. A second
suture was then loosely placed around the vessel upstream and a srnail incision made in
the vessel. A length of polyvinyl tubing of the appropriate size, previously flushed with a
heparin saline solution, was then gently inserted into the lymphatic and secured in place.
The catheter was carefully extenorised and the wound closed. A bottle holder was
sutured to the animals' skin, a bottie containing approximately 300 IU of heparin saline
was attached and the end of the catheter placed in the bottle allowing the collection of
lymp h.
In those experiments that examined CSF, the following surgeries were performed
at least 5 days prior to lymphatic cannulations. In some experiments, access to the CSF
was required, therefore a laminectomy was performed on vertebrae S2 or S3. A midline

incision was made in the overlying skin and the muscle gently dissected away hrom the
spinous process uncü the vertebral arch was exposed Using a high-speed d d I equipped
with a burr bit the laminae was removed and the dura exposed. An incision was made in
the dura and a catheter then inserted. To aUow the infiision of ceiis and cytokines into the
laterai venuic-les two bilateral 0.3 cm b m holes were made approximately 1.5 cm
anterior and posterior to the posterior fontanelle. A guide screw was then inserted into
the burr holes and a 16 or 2 1 gauge iv, catheter was fed through the guide screw-
Animals were given buprenorphrine (0.005 mglkg i.rn) during surgery and as
required thereafter. Experiments were not carried until the following day to d o w the
animai to recover-
2.3 Cell Labelling
I L 1 -Indium oxine (Amersbarn Coq, Baie d'Urfe. Que.) labelling was performed
as previously descriid (Issekutz et ai., 1980). Briefly lymph was coiiected. Lymphocytes
were then harvested by cenuifugation at 400 g for 10 min and washed twice with HBBS
or PBS. After the finai wash, the c e k were resuspended in either buffer at a
concentration of 1 x 10* ceiis/ml. Ten pCi of 11 1-In was then added, the c e k gentiy
mïxed and incubated at room temperature for 10 min. Ten ml of autologous lymph
plasma was added, the celis suspension washed hvice with either PBS or HBSS and
resuspended in an appropriate volume of saline for infusion.

2.3.2 51-Cr s
Lymphocytes were prepared as above except 50 pCi of ~ a ? ' Cr04 (rm, Costa
Mesa, CA) was added aad the ceils incubated for 30 min at 37°C These ce& were
suspended in saline for reinfûsion-
2.3.3 FITC labelling of blood
A saturated solution of FLTC (Sigma, Oakville, ON) was prepared by adding
0.05g to 500 ml of PBS or HBSS and stimng overnight at 4°C. The solution was then
filtered through a 0.2 micron fiIter immediately prior to Iabelling celis-
A whole blood method developed by Andrade et al. (1996) was used. Briefly,
approxllnately 300 ml of blood was withdrawn fkom the jugular catheter. This represents
approximately 15% of the total blood volume and is well tolerated by sheep. The blood
was added to a sterile beaker containing approximately 70 ml of acid citrate
anticoagulant. The blood was then divided into 2 large centrifuge tubes and cenVifuged
at 400g for 15 min. The plasma was carefüily removed, taking care wt to disturb the
bu- coat and the ce& resuspended in either PBS or HBSS. The c e k were split
between 4 tubes and washed twice to ensure the remval of all plasma. The blood was
then resuspended in saturated FITC and incubated at 4OC for 30 min, after which the ceus
were washed twice with either buffer, suspended in saline and reinfused intravenously.

2.3.4 FlTC labelling of lymph lymphocytes
Efferent lymph was collecte& harvested by cenaifugation at 400 g for 10 min,
washed twice with HBSS or PBS, resuspended in either b a e r and the concentration
adjusted to 1 x IO* ceWml. Seventy pl of saturated FITC was added for every log cells
and încubated at 37OC for 15 min. After this the cell suspension were washed twice with
either buffer and resuspended in saiîne for reinjection.
2.3.5 CFSE labelling of lymphocytes
Approxïmately 300 mi of blood was withdrawn via the indwelhg catheter into 60
cc syringes containing a small amount of EDTA in saline. Mononuclear celis were
harvested by centrihgation over Percoii gradients as previously descrïbed (Young et al.,
1997)-
CFSE (Molecular Probes, Eugene, OR) was diluted to 500 pghl in DMSO and
kept at -20 O C untii required. Mononuclear cells were resuspended at a concentration of 5
x 10'/ml in 37OC PBS containing 1 pglml CFSE for 15 min. The ceils were washed
twice with ice cold PBS contairung 1 8 foetal caif senim. The cells were resuspended in
s terile saline and reinfiised intravenously.
2.3.6 DiI-DS labelling of efferent lymphocytes
Efferent lymph was collected and ceiis harvested by cenaifugation at 400g for 10
min. The ceiis were washed twice with rmm temperature PBS and resuspended at a
concentration of 2 x log ceWd in 37°C Isocoves Modified Dulbecos media (Giko Life

Technologies, Burhgton, ON). The tells were then placed in a 37°C water bath for 10
mis DS-DS (Molecular Probes) was used at a concentration of 12 ~lgl10' c e k and was
weighed and diluted in 300 pl of DMSO immediately pnor to use. The DZ-DSDMSO
solution was added to the s a m ~ amount of tissue culture media as the celIs and gently
mixed with the cells to give a final concentration of log ceWmL The ceiis were aliowed
to incubate at 37OC for 30 min after which the cells were washed twice with room
temperature PBS and resuspended in stede saline for rehfbsion,
As this was a new application for this label it was determined by the author that
lymphocyte subsets were not adversely affected by comparing the phenotype of both the
beginning population and labelled ceiis (data not shown)- Using the above procedure
labeiied lymphocytes were obsecved to migrate fiorn blood to lymph, which dead or
overlabefled cells can not do. Previous work in our Iaboratory bas demonstrated that this
family of dyes efficiently labels c e k with w ceiL to cell transfer. Therefore, this method
of labeliing large numbers of lymphocytes is relatively easy and cost effective. A recent
report has used DiI-DS to label T c e k in mice and found that after intravenous infusion
DiI-DS labelled lymphocytes are found in lymph nodes (Dittel et al., 1999).
For al i of the above labelhg procedures aiiquots of ceiis were retained for
viabiüty staining using 0.4% trypan blue, ceU enurneration and labeIiing efficiency.
most of the experiments the c e k were >95% viable. Additionally the c e k were brightly
labelled with the CFSE and FïïC labelhg procedure. The DZ-DS did not label celis as
brightly as FITC and CFSE but usuaiiy greater than 85% of cells were labelled.

2.4 Antibodies used in this thesis
Most a n t i i e s and ceii h e s were generously shared by Dr. A Young of the
Basel Institute for hunology. These antiidies have k e n extensively characteriseci
and the results published (Table 1). For the fist senes of experiments, antiidy
supernatants were provided by Dr. Young. In 1998 we received the cell h e s and began
to culture the hybrïdomas ourselves using standard tissue culture techniques. AU
antibodies are antiovine muruie aatlbodies and used as ceU cuiture supemtants-
Exceptions to this was the antiovine CD25 (VMRD) and antiVCAM (gift fiom T-
Tedder), both of which were ascites.
2.5 lmmunophenotyping of lymphocytes
Blood samples were collected fkom the jugular catheter in a syrïnge containhg a
s m d amount of heparin (approximately 50U). Erythrocytes were lysed with either
distilled water or Tris:N&CI Iysis solution. The Ieukocytes were then pelleted by
centrifugation at 400g and washed twice with either PBS or HBSS. Samples of lymph
were washed twice with either buffer. The ceil count was determined using a ZN mode1
(Coulter Electronics, Hialeah, Fl) and 2 x 1o6 ceiis added per weii of a 96 weli U bottom
tissue culture plate (Becton Dickinson, San Jose. CA). The volume was adjusted to 100
pl with ice cold buffer and 50 fl of primary antiidy added. A 10 min incubation on ice
was foiiowed by centrifugation at 450 g. The supernatant was decanted and the cells
washed twice. Fifty pl of appropriately diluted secondary (see Table 1) antifbody was
added, the volume adjusted to 150 pi and incubated for 10 min on ice. After this fiaai
incubation the cells were washed twice with buffer and resuspended in 1%

paraformaldehyde. If flow cytometcy was not perf'ioffned immediately the plates were
wrapped in tin foil and stored at 4°C.
Celis were examined by fiow cytomeq wïthia 1 day of staining- In some
experiments, the buffer had 1% bovine aibumin added and the cells were preincubated
with goat IgG as a b l o c h g agent. These steps did not make a significant ciifference in
the background staining. In ai i experiments negative celis and ceus incubated with
secondary mAb and/or m u s e IgG were used as controis,
2.6 Flow cytometry
Either a FACScan or FACScahbur (Becton Dickinson) with Celiquest software
was used to perform the flow cytometry analysis. The instruments were checked with
quality convol beads by an operator every day to ensure its proper function.
Additionaily, if the second laser was required, calibrating beads were immediately nui
before the instrument was used. An eiectronic gate was drawn around the lymphocyte
population based on their typical side and forward light scatter properties. Lymphocytes
were then examuied using the relevant detector depending on the expeeen t and the
secondary anti'body used.
2.7 lmmunohistochemistry
Tissue was harvested and as soon as possible placed in a medium size cryomold,
embedded in OCT and fiozen in liquid nitrogen. Blocks were wrapped in aluminium foil
and stored at -70°C. Eight pm sections were cut using a cryostat and placed on

~ i ~ a n i z e d siides. The slides were dried ovedght, fixed in cold acetone for 5 mùi. air
dried and stored at -20°C
Siides were allowed to corne to m m temperature in the slide box and then
endogenous peroxidases were blocked with a solution of PBS, 0.1% hydrogen peroxide
and 1% sodium azide. AU incubations were at room temperature in a humid container.
The slides were rinsed in PBS, biotin and avidin blocked using a commercial blocking kit
(Vector Laboratones, Burluigton, ON) and incubated with 10% foetal calf senun for 30
min. The undiluted primary antibody was added (Table 1) and the slides incubated for 2
hr. PBSî'ïween was used to wash the siides twice, biotin labelIed goat antirnouse
antibody added and incubated for 30 min, The siides were washed twice, the commercial
avidin/biotin complex (Vector Laboratories) added and incubated for 30 min- The slides
were rinsed twice and DAB solution (Vector Laboratories) added for 5 min. Several
rinses of distüied water were used and a Light nuclear counterstain of haemotxylin
applied. The slides were dried overnight and pemianently coverslipped with mounting
media (Perrnount),
2.8 Reagents
2.8.1 HanKs balanced salt solution (HBSS)
For imrnunophenoîyping, sterile 1X Ca and Mg containhg buffer without phenol
red was used (Gibco Canada, Burhgton, ON). For a general wash buffer 1OX
concentrated buffer without divatent cations was used and reconstituted with stede
distilied water.

2.8.2 Phosphate buffered saline (PBS) without divalent cations
This was made using the foUowuig protocol; 2L d&O. 400g NaCl, log KCl,
57-58 N a m 4 and log L(H2 PO4. The LOX-concentrated buffer was prepared in 4 L
batches, autoclaved and stored a m m temperature until use. It was then reconstituted
using distilled water.
2.8.3 Dulbecco's phosphate buffered saline
This was prepared ushg the foliowing protocol;
solution 1 - 8.0g NaCl, O.2g KCI, 1.15g Na2HP04, 0.2g KH2P04 and 800 mi Hfi
solution II- O. lg CaC12 and LOO mi Hfl
solution III- O. lg MgC12 *6 H20 and 100 ml H20
The three solutions were made separately, autoclaved and mixed immediately
pnor to use. This buffer was used for immunophenotyping and to dilute secondary
antibodies-
2.8.4 PBSrrween buffer
10 X PBS was reconstituted with distilied Hfl and 0.05% Tween added-
2.8.5 Parafonnaldehyde solution
100 ml of PBS without divalent cations was preheated to 53-57OC in a fumehood
using a hotplate equipped with a stirrer. One gram of paraformaldehyde was added and
ailowed to stir until dissolved. The solution was then cooled and filtered through a 0.2
micron filter to remove any undissolved particles. The solution was then aliquoted and
stored fiozen until required.

2-8.6 Erythrocyte lysis solution
solution 1 - Tris O- 17M was dissolved in dH20 with continuous s t idg
solution II - O.83g NE&CL was dissolved in 1OOmls dEF20
The solutions were stored at 4°C mtiI needed- When the solution was required
they were mixed at a ratio of 19, (HI) and prewarmed to 37°C. The solution was then
added at a ratio of 4:L blood
2.8.7 Acid Citrate Dextrose This was prepared as followed C&&as07 2Sg, C&Nag 1.5g and -1206 2-08 were
added to LOO ml H20- It was used at a ratio of 1:6 (acid citrate dextrose to blood).

Table 1 Antibodies used in this thesis
Antihdies obtained fiom the Basel Institute for Immunology @II) were either cell
culture supernatants or hybridomas. Severai of the hybridoma hes were grown in
Toronto by TS- Cederlane= Cederlane, Hornby, ON

Antigen Distribution Clone# Supplier Reference
Recognised
PrÏrnarv Anhibodies
CD4
CD8
YS B cell
CD2 1
CD25
CD45 RA
L selectin
SIEN VCAM
Secondarv
Antibodies
FITC-GAM IgG
PE-CAM-Ifi
APC-GAM-I,@
T ce11 subset
T ce11 subset
T cell subset
Ail B cells
Absent from T celk
some B c e k &
monocytes
Subset B cells
Activated T celis
B celldnaive T cells
recirculatïng
lymehocyt=
B cells
endothelial cells
DU2-128 BII
VMRD
p z 0 BII
DU2945 BII
13-30 BII
HAE-2 T- Tedder
Cederlane
Cederlane
Cederlane
(Mackay et a1, 1986)
(Mackay et al., 1986)
(Mackay et al, L99 1)
(Young et al., 1997a)
unpublished
(Young et al., 1997a)
(Mackay et al., 1990)
(Spertini et al, 199 1)
unpublished
(Mackay et al, 1992b)

Chapter 3 Investigations into the migration pattern of BL and LL
into afferent lymph and after splenectomy
3.1 Abstract
Previous data demonstrate a relative preponderance of BL to localise in the spleen
and blood as compared to LL (Young et al. 1997). Sequentki repeitive sampling of
blood and lymph was performed to test the stability of both pools in blood and lymph-
Normal afferent lymph was also examined for labe1Ied BL and LL to determine if either
preferentially migrated through tissues. Experiments were performed to investigate the
effect of splenectomy on the maintenance and migration of BL. The relative proportions
of labeUed BL and LL remained constant in both blood and efferent lymph over the 27 hr
samphg period- LL are found in afferent lyrnph in greater numbers than BL, and one
may postulate that LL have a greater role in immune surveillance of peripheral tissues-
Neonatal splenectomy did not result in a change in the number or phenotype of
lymphocytes in either lyrnph or blood indicating that the spleen is not necessary for the
long-term maintenance of BL. However, its removai resulted in an increase in the
migration of LL into peripheral lymph nodes and a retention of labelled BL in blood as
compared to intact sheep. Together these data provide information requirrd for the
interpretation and design of subsequent experiments in this thesis.

3.2 Introduction
Previous experiments using fiuorescent labels and flow cytometry demonstrated
the existence of the BL pool (Young, et al., 1997a; Andrade, et aL, 1998). This pool does
not migrate into lymph nodes as efficiently as LL, but instead localises in the spleen
(Chevallier et al., 1998). The majority of BL are B cells that do not express either CD21
or L selectin, however CD4 CD8 and perhaps y6 T cek are also present in the BL pool
(Young et al., 1997a). Two recent publications have demonsaated tbat BL B cells are B 1
like ceils and express CDS, CDllb and high levels of surface IgM (Chevallier et ai.,
1998; Gupta, et al.. 1998). Separating these ceils and perfomiing tracking studies
confirmed that BL B cells are excluded fiom the lymphatic system (Gupta, et al., 1998).
Investigations into the existence of BL were begun in 1994 by Young (Young,
1994). However Iittle data exist about these cells beyond their anatomic location,
phenotype and some preliminary data on their behaviour (Chevallier, et aL, 1998). TO
Leam more about BL, the present experiments were designed to investigate the migration
of BL into afferent lyrnph, and after splenectomy. Additionally, repetitive sampling of
blood and lymph was perforrned to examine the stability of the two pools.
To date BL have not k e n examined in normal afferent lymph. There are
significant ciifferences in the cellular composition of blood, afferent and efferent lpph.
This led to the speculation that BL and LL may daer in their migration into afferent
lymph. The appearance of labelled cells in aEerent lymph implies migration through
tissue and hence immune surveillance (Schieiffenbaum and Fehr, 1996).
The spleen is an important site of lymphocyte migration, with BL localising in
this tissue to a greater extent than LL (Young et al., 1997a). Previous snidies have

demonstrated that in splenectomized rats a lymphocytosis, composed oCB celis and CD8
T celis, developed (Westernmm et al, 1990). Using FïïC hbe1Ied thoracic duct ce&
Westennana et ai. (1989) demnstrated that splenectomy caused an hcrease in labelied B
cells as compared to T c e k in blood and Lymph nodes. Based on these data it was
concluded that splenectomy changed the migration pattern of both B and T ceiis.
However, no experiments have been perfomied that examines the migration of the BL
and LL in splenectomized animals.
The increased B ceils in the blood of splenectomùed subjects may possibly be
due to an accumulation of BL (Sieber et aL, 1985). If splenectomy increases BL in
blood, this could be a method to selectively expand thk pool for hiture experiments.
Therefore, the eexperiments in this chapter were performed to gain a better
understanding of the basic biobgy of BL. These data are important for the design and
interpretation of subsequent experiments.
3.3 Materials and Methods
The experiments involving the splenectomized sheep were performed at the Basel
Institute for Immunology. The repetitive sampling and afferent lymph experiments were
performed at the University of Toronto.
Lymphocyte migration in splenectomized sheep was £irst investigated ushg
fluorescent labeiled celis and immunophenotyping. After 4 days this was foliowed by
tissue distribution e x p e e n t s using radiolabeled cells. Sheep used in the repetitive
samphg experiments had prescapular and prefemord efferent lymphatics cannulated.

Both BL and LL were labelled with fluorescent dyes, intravenously infused and then
blood and lymph were repetitively sampled
3.3.1 Anirnals and surgery
Sheep used in the splenectomy experiments had their spleens removed between
the ages of 19 and 2L days by Dr. W. Heia The animals were then retumed to the fann
for the foIlowing 2 years. At the tïme of the experiments the animals were of a n o d
size and weight and did not appear to s&er fkom disease. The control sheep were fiom
the same supplier and were approximately the same age but were not splenectomïsed.
For aii experiments, prescapular and prefemoral lymphatics were canulated as
described in Chapter 2. Afferent lymphatics were canulated in some experiments- A
jugular catheter was also surgically placed at the same time to d o w access for blood
samphg.
3.3.2 Lymphocyte labelling
For the splenectomized sheep experiments peripheral blood mownuclear cells
were labelled with CFSE. For aii other experiments, the whole blood labelling procedure
using FITC was used. In al l experiments, lymph Lymphocytes were labelled with DiI-DS.
Radiolabeling of lymphocytes with 111-lk and 51-Cr was as outiined in Chapter 2.
Blood lymphocytes were isolated using Percoli and labeled with 11 1-In, while lymph
lymphocytes were labeled with 5 1-Cr. The radiolabeled cells were infused intravenously
and allowed to migrate for 8 hours before tissues were harvested. The tissues were

3.3.3 Sampling of blood and lymph for tracking fluorescent labelled cells
Samples of blood were cokcted and the erythrocytes lysed. The samples of
lymph and blood were washed twice with buffer and resuspended in paraformaldehyde.
A FACScalibur was used to determine the percentage of IabeIIed cek in the samples.
Lymphocytes were examined by using thek typical side and forward Light-scattering
properties.
3.3.4 Statistical analysis
To ailow pooling of the data two methods were employed. In the experïtnents
involving the repetitive sampluig of blood and lymph the average percentage of FITC+
and DiI-DS+ was determined for aII samples. AU values were divided by this number to
give a ratio and these ratios fiom al l experiments poded. The second method was used
for splenectomized sheep, in which the percentage injected was calcuIated and the results
pooled. GraphPad Instat software was used to perform one way ANOVA and appropriate
post-hoc tests. Student t-tests were used were appropriate-

3.4 Results
3.4.1 Repetitive sampling of blood and lymph
Labeiled ceiis were infbsed and allowed to equilibrate for 20 hours and blood and
Lymph were then sampled houriy. As previously docamented lymphocytes isolated nom
lymp h, labelled and reinfused intravenously are found in higher concentrations in lymp h
as compared to blood Figure 3 is a representative experiment fiom 5 experiments
performed and shows that FïïC labelled blood Lymphocytes were approximately 3 times
enriched in blood as compared to lymph. Conversely, DiI-DS labeiied Lymph
lymphocytes were approximately 1.5 times enriched in lymph as compared to blood
Pooling of the data fkom the 5 separate experiments demonstrated that there was no
significant change in the concentration of BL and LL over t h e in either blood or lymph
(Fi-gure 4).
3.4.2 Afferent lymph
Samples of afferent and efferent lymph and blood were collected in 3 normal
anhals. The percentage of labelied ceUs in afferent and efferent lymph was divided by
their percentage in blood, Blood was chosen as the denominator as cells were migrating
kom this tissue. As previously reported there is an encichment of FLTC+ lymphocytes in
blood and DiI-DS+ cells in efferent lymph (Table 2). In afferent lymph FlTC labelled
BL were not e ~ c h e d as the ratio was approxirnately 1. Ln contrast, the ratio of Da-DS
labelled lymphocytes was approximately 3, indicatuig a selective migration of LL into
afferent lymph.

3.4.3 Splenectomized sheep migration
In 4 splenectomized sheep the migration of BL and LL was examined using both
fluorescent and radioiabeled lymphocytes. Using fluorescent c e k the short-term
migration (4 hr) was examined and compared to control animals (n=3). In the blood of
intact sheep DiI-DS labeiied LL ceiis decreased to approximately 0.00246 injected by 1
hour p o s ~ i o n , whilst the CFSE iabeIied BL were retained in the blood (Figure 5)- In
splenectomized sheep, there was no significant difference in the migration pattern as
cornpared to control sheep. However, there was a trend towards BL king retained withui
the blood in greater numbers as compared to intact sheep 0.004 % injected vs. 0.002 %
injected-
Radiolabeled cells were used to determine which tissues BL and LL migrated into
after splenectomy. No tissue had a higher concentration of labelied BL as compared to
LL; therefore, no tissue appeared to be a preferential site of BL homing (Figure 6). When
the data is compared to that previously reported (Young et ai., 1997a) for intact sheep it is
seen that LL enter lymph nodes and tiver in signincantly higher amounts and BL are
partiaiiy excluded fiom the lungs (Table 3).
3.4.4 lmmunophenotyping of lymphocytes in splenectomized sheep
There was no lymphocytosis or difference in blood and lymph phenotypes in
splenectomized sheep (Table 4) as compared to contml sheep.

As seen in intact animais there is an enrichment of CFSE labeiied BL In blood and
Da-DS IabeUed LL in lymph. Table 4 summarises the phewtypic data of IabeIIed cek
and udabeled c e h at 24 hours pst-infusion. The resuits are sunüar to those previously
reported by Andrade (1996) in intact animals, There is an enrichment of sIgM and
CD1 Lb+ aad B ceils in the CFSE labelled lymphocytes in blood. CD4 and CD8 CFSE
labeiled celis are present in a lower number in blood. In Iymph, y6 T cek are enriched in
both populations as compared to uniabeIed tells-

Figure 3 Representative expehent of repetitive snmpling over a 27 br pend
One representative experiment is shown. FiTC labelled BL are e ~ c h e d in blood as
cornpared to lymph, whilst DE-DS Iabeiled LL are concentrated in lymph.
A - Flow cytoxnetric detection of labelled ceiis in blood
B - Flow cytometric detection of IabeIled ceh in efferent lymph

-% FITC
Time (hours)
B Efferent lymph
-% FITC -cl- % DiI-DS

Wgure 4 Ratios of BL and LL lnbeiied c e k in efferent lymph and blood over a 27 hr period
There is little variation of FITC labelled blood lymphocytes in blood or efferent lymph
over 27 hr. SimilarIy, DE-DS Iabeiied lymph lymphocytes exhibit little variation. Ratios
were determined by dividing the percentage of labelied ce& at each tirnepoint by the
average percentage obtained during the entire experiment (n=5)-
A - ratio of Iabeiied cells in efferent lymph
B - ratio of labeiled c e k in blood

A Efferent lymph
Time (hours)
0 . 5 0 ! , 1 I 1 l I l I L 1 l I I 1 1 1 1 I L L L t i i '
O 2 4 6 8 10 12 14 16 18 20 22 24 26
T ime (hours)

Table 2 Perrentage and ratio oPIabeIled ceL in bbod, efferent and aerent lymph
The percentage of labeiled cells is shown with the ratios in parenthesis. A ratio was
determined by dividing the percentage in lymph by the value in blood DS-DS labelleci
LL is enriched in both afferent and efferent lymph as compared to FITC labeiïed BL.
Experiment % btood % efferent (ratio) % afferent (ratio)
HTC DIT-DS FITC DiI-DS FITC DiI-DS
Expt. #l 1.22 0.30 0.28 (0.2) 0.54 (1.8) 0.54 (0.4) 0.80 (2.7)
Expt- #2 0.48 0.09 0.20 (0.4) 0.30 (3.3) 0.62 (1.3) 0.44 (4.9)
Expt. #3 1-15 0.88 0.49 (0.4) 0.90 (1.0) 0.87 (0.7) 1.41 (1.6)
average ratio f SEM 0.33 k 0.07 2.0 f: 0.67 0.8 t 0.26 3.1 & 0.96

Figure 5 Short-tenn migration oflpbelled lymphocytes in the blood of splenectomized sheep
Blood was sampled every 15 min during the E s t 4 hr d e r the infusion of fluorescentfy
labelied cells. In splenectomized sheep (A) (n4) there was a trend for Da-DS IabelIed
LL to disappear fiom the blood at a faster rate than in control sheep (B) (n=3). CFSE
labeiied BL were enriched in blood in both splenectomized (A) and control (B) sheep- A
trend for splenectomized sheep to retain BL in blood is seea

A Splemectodzeà sheep
1 -Avg CFSE
+Avg DiI-DS
O 15 30 45 60 7s 90 105 120 135 150 165 T80 195210 225280
T ime (min)
B Control sheep
-Avg CFSE
+ Avg D il-OS
1 ime (min)

Tissue
Figure 6 T'me localisation of radiokbeied LL and BL lymphocytes
AU lymphoid and control tissues in splenectorrüzed sheep contain greater amounts of LL
than BL (n=4),
LL - 5 1 -Cr labeiied Lymphocytes (solid bacs)
BL - 1 1 1-In iabeiied Lymphocytes (open bars)

Table 3 Tissue localisation h splenectornized sheep
Peripherai blood lymphocytes and efferent lymph lymphocytes were labeiled with
radioisotopes (see figure 6) and their migration into ciifferent tissues determined. There is
a signincant increase in the amount of LL in prescapular Lymph nocies of splenectomized
sheep (n=4) as compared to intact sheep. The lung has fewer BL in the splenectomized
sheep as compared to the controI subject-
* = percentage of injected c e k I gram of tissue t SEM
# = significantly different (pc 0.05) fiom the correspondhg ceii migration in the control
animal
S denectodzed shee~ Contrai s h e e ~
Tissue BL LL BL LL
PrescapuIar LN .04 f .()os* -204 t -03" .O24 tOO8 -043 & .O1 1
Mesenteric LN .O2 + -005 . 107 + -0 19 .O21 f -008 -065 I -020
Lung -03 ,t ,005' , 106 + -007 ,065 I .O 12 -075 t .O 16
Peyer's patch ,008 f ,004 .O12 k -002 .O05 & .O02 .O20 I .O05
Liver .O12 t .O04 -020 I .w# .O14 + .O03 ,011 t .O01
Blood cells .006 f .O01 ,002 f .O003 .004+.001 .O02 I .O01

Table 4 Immunophenotypic data h m splenectomized sheep
S plenectomized s heep ( n 4 ) demonstrate no merence in the phenotype of Lymphocytes
in blood and lymph as compared to intact sheep. BIood Lymphocytes were labelled with
CFSE and LL with DiI-DS,
A = p henotypes in splenectomized and intact animals
B = phenotypes of CFSE and DI[-DS labelled ceils in blood
C = phenotypes of CFSE and DiI-DS Iabelled cells in lymph
* = average + SEM
7 = significantly different (p < 0.05) than total (unlabeiied) Lymphocytes
§ = ~i~onificantly different (p < 0.05) than CFSE labeiled B L

A Cornparison o€lymphocytes in bbod and lympb
S~lenectom*zed sheee htact s h e e ~ Cell Type Blood Efferent Lymph Blood Efferent Lymph £3 cells 56.7 4.L* 23.4 * 3.2 43.8 I 7.5 24.4 I 3.5 CD4 14.6 * 1-7 42.8 * 1.5 18.0 & 4-1 35.0 * 3-4 CD8 12-2 & 1.3 14-0 I 1.3 14.2 I 3.3 15.1 i3.8 y5 9.9 k2.1 L 1.3 * 1.0 12.3 I 7.5 13.5 * 2.1
B Phenotype O€ IabeUed cells in blood
Subset Unlabeled C'SE ~abemed CD4 15 I 1.7 4 t 0.8' CD8 12 t 1-3 2 k 0.2~
10 t2.1 7 + 1-3 B 57 & 4-1 75 I 6.5
CDllb 40 t 4.7 75 I 3.2' CD21 24 t 6.3 25 -t 8.9
L selectin 31 I3-3 16 t 2.7' sIgM 38 t 2.6 63 + 5.4'
C Phenotype of kbeUed ceL in lymph
Subset CD4 CD8 YS B
CD1 lb CD2 1
L selectin
SI@
DZ-DS labeiled
Lvm~hocvte ~ o ~ u h t i o n s in lvmah Unlabeled CFSE IabeLled DiI-DS labeiied 43 k 1.5 16 I 1.5' 38 t 3.4' 14 I 1.3 4 & 0.3' 8 + 0-9'' 11 I 1-0 40 I 3.4' 3 1 + 2.7': 23 I 3.2 28 f 1.4 16 k 0.9~ 1 k 1.0 5 + 1-9 1.2 I0.3 23 33.2 31 13.2 19 & 2.7 89 I 1.4 89 ,t 1.8 90 k 1.3 22 f 2.7 31 12.9 17 t 2.2"

3.5 Oiscussion
The data presented in this chapter confïrm and extend previous hdings of BL as
compared to LL. These basic data are also important in the interpretation of the
expeaents in subsequent chapters.
Previous investigators have used fluorescentiy labelled Lymphocytes to examine
the migration of lymphocytes obtained nom blood and lymph (Young et al., 1997a;
Andrade et A, 1998). There are several advantages to labehg the whole population of
blood and efferent lymph and reinfusing all cells. Fkstly, both blood and lymph
lymphocytes undergo Little in vitro manipulation, thereby reducing any chance of
inadvertent activation or change in adhesion molecules. However, the main advantage is
that the whole population is retucned to the blood and the subpopulations of lymphocytes
can migrate into their respective compartments (Westemiann et al., 1993).
Repetitive samplîng of both blood and lymph confirmed that a pool of
lymphocytes exists that is preferentially retained in blood as compared to LL, as seen in
Figure 3. The BL are found in blood at a percentage 3 times that in simultaneously
coiiected efferent lymph. Figure 4 demonstrates that there is Little variation of the two
populations in either blood or lymph during the entire 27 hr sampiing period.
Immune surveillance is a putative function of recirculating lymphocytes
(Schleiffenbaum and Fehr, 1996; Butcher et al., 1999). In afferent lymph, the ratio of
labelled LL is 3 times higher than in blood but BL are found in a similar percentage
(Table 2). The appearance of IabelIed cells in aiferent lymph impiies that they have
migrated through tissues. However, Lymphocytes may migrate into tissues and be
retained or deleted, therefore not entering afferent lymph. Prior snidies using

radiolabeled BL and LL have demonstrated greater numbers of LL in slcin (Young et al.,
1997)- Taken together these data demonstrate that LL m i t e into nonlymphoid tissues
in larger numbers than BL. Fiutber experiments are required to investigate transit times
and quanti@ the trafGc of these pools into tissues.
In splenectomized sheep, no Iymphocytosis or differences in the phenotype of
blood and Lymph lymphocytes was observed as compared to control sheep (Table 4).
These results disagree with previous reports in rats (Westemiann et ai., 1990) and humaos
(Ferrante et aL, 1987). One explanation for this discrepancy maybe that the effects of
splenectomy are species specific. However, 1 believe a more plausible explanation is that
the animals in this study were s p l e n e c t o ~ d at approximately 3 weeks of age as
compared to the other studies, which used adult subjects splenectomized as adults.
Tracking studies reveal no statisticd diffferences between the disappearance of the
two pools in intact and splenectomized sheep, either in the short-tenn @ours) or up to 4
days af3er the infusion of ceils (Figure 5). As weil, there were no differences in the
phenotype of labelied ceils in blood and lymph (Table 4) as compared to previously
reported data (Gupta et al., 1998). Those CFSE labelled Lymphocytes retaioed in the
blood were mainly B celis, CD L Lb+ and L selectin-, whilst those in lymph were CD1 Lb-
and L selectin+.
In agreement with a previous report (Young et al., 1997a), both CFSE and DiI-DS
labelled cells were enriched for y6 ceils in Lymph 40% f 3.4 and 3 LI + 2.7 respectively
as compared to 11% t 1.0 for unlabelled Lymph. In blood there is no enrichment of
labeiled y6 BL as compared to unlabelled lymphocytes, 7% f 1.3 vs. 10% f 2.1, but LL
y6 are signifcantly increased (29% k 3.2). Thecefore, y6 celis may rapidly recirculate

nom blood into lymph. This conclusion disagrees with previous results by Andrade
(1998) and others (Witherden et aL, 1990), who suggest that a pool of wnrecircuIating ya
T ceus is present in blood. An explanation for this discrepancy maybe the different ages
of the sheep used in these experiments as Young et aL (personal communication) have
demonstrated a difference in the migration and number of y6 T cells in young and adult
sheep.
Experiments using radioactive IabeIled BL and LL were executed to determine the
migration pattern of the two pools in splenectomized sheep. No examuled tissue in
splenectomized sheep had a hîgher concentration of BL as compared to LL 6).
This is in contrat to intact sheep in which BL preferentiaiIy home to the spleen (Young
et al., 1997a). Peripherd lymph nodes and Iiver had a greater percentage of injected LL
as compared to control sheep. In splenectoniized sheep significantly fewer BL were
detected in the lung compared to control sheep, 0.03 k 0.005 and 0.065 + 0.012 %
injected (Table 3).
These radioactive and fluorescent tracking data demo as trate a ciifference in the
circulation of BL and LL in splenectomized sheep. An explanation of these data is that in
splenectomized sheep Li, spend a shorter time in blood aad quickly enter the lymphatic
system and iïver. This is shown by the decrease in labelied cells in the short term
t r a c h g studies and the higher numbers of radiolabeled ceils in peripheral lymph nodes.
The spleen therefore appears to act as an organ of localisation of the BL but retards the
migration of the LL in its migration into lymph nodes.
In conclusion, these experiments contriiute to the increasing amount of data that
demonstrates the existence of a pool of lymphocytes in blood that does not recirculate as

efficiently as LL. The spleen is not necessary for the maintenance of these BL but its
removai does have an effect on the migration of both pools, resulting in enhanced
migration of LL Ïnto perip heral lymph nodes. In afferent lymph, there is a preponderance
of LL as compared to BL. This in combination with previous resdts using radioisotopes
implies that LL have a greater role in the immune surveillance of tissues as compared to
BL. The BL and LL exhiiit littie variation in their concentration in blood and Iyrnph
over time. Taken together these data provided the basis for the design and anaiysis of
subsequent experiments presented in this thesis.

Chapter 4 Lymph and blood CD4 cells increase in efferent
lymph during antigen induced lymph node shutdown
4.1 Abstract
Lymph node shutdown is a period of decreased cell output in efferent Iymph after
the introduction of certain antigens. Previous studies have demoostrated that CD4 ceUs
are not retained within the Iymph node to the same extent as other Lymphocyte subsets
during shutdown. However, the effect of shutdown on the migration of LL and BL has
not been investigated. Cytokines may mediate Lymphocyte retention within the lymph
node by upregulating adhesion molecules; therefore, IL-1P, IL-6, IL-8, IFN-y and TNF-a
were measured in efferent Lymph during shutdown. The percentage of CD4 Lymphocytes
f?om both pools increased in efferent Lymph during lymph node shutdown. IFN-y and IL-
6 increased in efferent lymph when tell output reached its nadir, implying they have a
role in the recruitment &or retention of Lymphocytes within the Lymph node d u ~ g
shutdown.
4.2 Introduction
Certain antigens, when introduced into lymph nodes, uivoke a penod of decreased
cellularity in efferent lymph, which has been called "lyrnph node shutdown" (Bujdoso et
al, 1989). Hall and Morris (1065b) reported that antigens ranging from viruses to
globulins induced Lymph node shutdown for various Iengths of tirne. This is fiequently
foliowed by a period of increased lymphocyte output, including antigen specinc blast

ceus (Cahill et aL, 1974). TNFa (Young et ai, 2000) and IFN-a (Hein and Supersaxo,
1988; E;alaaji e t aL, 1989) also invoke a period of shutdown demnstrating that this
phenornenon does not require antigen. The physiological relevance o f lymph node
shutdown is unknown but one speculation is that it aiIows the presentation of Ag to large
numbers of recruited lymphocytes in a shoa time (Haii and Morris, 1965b).
Previous studies have investigated lymphocyte output and subsets during
shutdown. (Bujdoso et ai., 1989; Mackay et al., 1992b) but the migration of BL and LL
through an antigen s h l a t e d Lymph node has not been examioed The present
experiments may begin to detennine if different fiuictions exist for the two pools.
Various cytokines can increase adhesion molecules on the blood vascuiar
endotheiium, thereby contributing to lymphocyte migration (Juda et ai, 1989; Raine et al,
1990b). Cytokines produced during lymph w d e shutdown may increase the t r a c of
lymphocytes into Lymp h nodes and t heû nibsequent retention. Lymphocyte retention
may be due to the upregulation of adhesion molecules on lymph node stroma1 ceus,
lymp hatic endothelium and other lymphocytes. Interleukin- 1 f3, IL-6, IL-8, IFN-y and
TNF-a levels were measured as they are produced during BCG induced grandomas and
intravesical BCG infusions (Sugisalci et al, 1998; O'DonnelI et al, 1999). These same
cytokines are known to increase adhesion molecules on endotheliai ceiis and may have a
role in lymph node shutdown,

4.3 Material and Methods
4.3.1 Animals and Surgeries
These experiments were performed at the Basel Lnstinite for Immunology and aJI
surgeries were performed by Dr. A Young and Ms. Lisbeth Dudler. Prescapular efferent
Lymphatics were canulated as descriid in Chapter 2.
The sheep had been immunised with 5 times the nomial human dose of BCG
(Institut Serotherapique et Vaccinal Suisse, Berne) at least 21 days prior to nirgev For
antigen stimulation 50 pg PPD (Staîens Seniminstitut, BaseI) was suspended in 1 mi of
salùie with 2% Evan's blue and htradermally injected in several sites located in the
drainage area of the canulated lymph node. The appearance of Evan's blue in the efferent
lyrnph allowed the confirmation that PPD had reached the lymph node. Evan7s blue has
been used in our laboratory and others for several years to locate lymph nodes and does
not have a detrimental effect on lymphocyte output-
4.3.2 Cell labelling
Twenty-four hours after surgery, blood cells were coiiected and labelled with
CFSE and lymph Lymphocytes labelled with DiI-DS as in Chapter 2 Both celi
suspensions were intravenously reinfused.
4.3.3 Cell collection and phenotyping
M e r 48 hours sampies of efferent Lymph were collected every 4 hours for 16
houn to obtain baseline Ievels of A) lymphocyte subsets, B) percentage iabeiled ceff s and
C) cytokine levels. Lymph plasma for cytokine levels was harvested after cenuifugation
and fiozen at -80°C until required. Lymphocytes were ïmmunophenonlped and analysed

ushg APC as a secondary antibody. After the injection of PPD, samples of lymph were
collecced at vai5ous t h e s over the foIlowing 48 hours.
After the last collection of Lymph, PPD and saluie were separately injected i'to
the drainage area of contralateral prefemoral lymph nodes in two sheep. Sixteen hours
later, during the period of maximum shutdown, the sheep were sacrificed and the lymph
nodes excised. They were bisected and half was miaced in coM media, fikered, washed
twice with PBS and immunophenotyping performed as previously outlined- The other
haif was fiozen in OCT and stored at -80°C until immunofiistochemistry was performed-
4.3.4 Imrnunohistochemistry
Murine antiovine MF-a, IL-1$, IL-6 and IL-8 were used to stah the excised
lymph nodes for cytokine production. Optunal concentrations were determined and the
procedure in Chapter 2 was followed. A lymph node fiom a normal sheep was used as a
control.
4.4.4 Cytokine ELlSAs
ELISAs for ovine cytokines were performed as previously descnbed with minor
modifications. Briefly, matched pairs of antiovine cytokine antibdies for L I $ , IL-6,
IL-8 (Serotec, Oxford, U.K) and TNF-a (Centre for Aaimal Biotechnology, Melbourne
Aus.) were obtained. Each pair consisted of a monoclonal murine antihdy and
polyclonal rabbit senun. 0.1% alpha casein was used as the blocking agent and
PBSflween as the washing buffer. The muruie anti idy was used as the capture
antibody, followed by efferent lymph plasma, then rabbit senun, which was detected by
horse radish peroxidase-conjugated goat anti rabbh IgG (Southern Biotechnology,

Birmingham, AL) and visualised using tetramthylbenzidlne iiquid substrate (Sigma,
Basel, S witzerland),
IFN-y was detected using the BOVIGAM commercial kit obtained fÎom CSL
Vete~ary (Australia) and the instructions were foilo wed exactly, This kit cross-reacts
with ovine IFN-y (BOVIGAM insert and ourown unpublished observations)
Duplicate samples £iom each experiment were nui on the same day and each nin
Ïncluded a standard curve or controis. The Iowest amount of cytokine that could be
reliably detected was 0.1 nglml, therefore this was used as the cut-off point. No W - a
of a known concentration was available, therefore a standard cuve could not be
constnicted. This cytokine was therefore measured as a change in absorbance over the.
A sample of tissue culture supernatant fiom a TNF-ût producing ceiI line (gift of J
Hopkins) was used a positive control,
4.4.5 Statistics A Students two taiied paired t-test with a signiticance of pe0.05 and a repeated
measures ANOVA using a Dumets p s t test were used to determine statistical
4.5 Results
4.5.1 Lymphocyte Migration
Lymph node shutdown was detected as previously reported (n=3), with efferent lymph
ceii output decreasing to approximately 20% of baseline vahes at 16 hour postinjection

of PPD (Figure 7). These experiments were terminated at 48 hours posîk.jection,
therefore, the subsequent typical increase in Iymphocyte output was not observed.
The number of CD4 CDS, y5 T celis and CD2h B ce& ail decreased during
lymph node shutdown to at least 50% of basehe, However, when the percentage of c e k
exiting the lymph node was determiaed CD4 cek did not decrease as compared to the
other subsets. Using three-colour flow cytornetry, CD4 ceiis fkom both pools appear to
be retained less efficientiy. This clifference was not st atistically significant . Nonetheless.
it appears that BL and LI, CD4 celIs exit the lymph node easier then the other subset
du~511g lymph node shutdown (Figure 8).
PPD ïnjected lymph nodes excised during shutdown (n=2) did not statistically
m e r from saline injected lymph nodes with respect to the percentage of CD4, CD8 y6 T
cells and B celis (Table 5)-
4.5.2 Cytokine Levels
IL-lp, IL-6, IL-8 and IFN=y could not be detected in efferent lymph during the
pre-injection baseline period Both IFN-y and IL-6 levels had significantly increased in
concentration at 9 and 12 hr postinjection (Figure 9), occurring just prior to the period of
rnïnïxnal ceil output in the efferent lymph. TNF-a, IL-lp and IL-8 levels were not
increased over baseline measurements.
IL-6 was found in both PPD and saline injected lymph nodes but not in normal
lymph node. The IL-6 appeared to be Iocated near endothelid and smooth muscle ceh,

both of which have been reporteci previously. IL-@, IL-8 and TNF-a were not found in
the PPD or saline injected nodes,

Time (hours)
Figure 7 PPD induced lymph node shutdown
There was a substantial decrease in the ceiiularity of efferent lymph after the injection of
PPD hto the drainage area of a prescapular Lymph node (n=3). Ratios for individual
experiments were determined by dividing aU values by the average obtained during the
baseLine period. These ratios were then pooled for all three experiments.

Figure 8 Phenotype of lymphocytes in efferent lyrnph during lymph n d e shutdown
There is a decrease in the number of all measured subsets during lymph node shutdown
(A). The percentage of CD4 c e k demonstrate a trend to an increase (B), whilst other
subsets decrease. Both the CFSE iabeiied blood lymphocytes CD4 ceils (C) and DiI-DS
IabeiIed CD4 lymph lymphocytes (D) increase-
Ratios were produced by d e t e e g the average value obtained during the preinjection
baseline and dividing a i i values by this average. Therefore, a value of greater then 1
indicates an increase. (n=3)
A = number of ceiis as compared to baseline (ratio)
B, C, D = percentage of ceiis as compared to baseline(ratio)
+- CD4
A - y 5
m- CD8
O- CD21


Table 5 Immunophenotype of PPD and saline hjected Iymph nodes
There is no merence between lymph nodes excised 16 hours after the injection of saline
or PPD. (n=2)
* average percent t SEM
Subset PPD injected Saiine injected
CD4 41.5 f 7.0* 37.1 & 2.9
CD8 L8.9 t 0.6 18.8 t 0-6
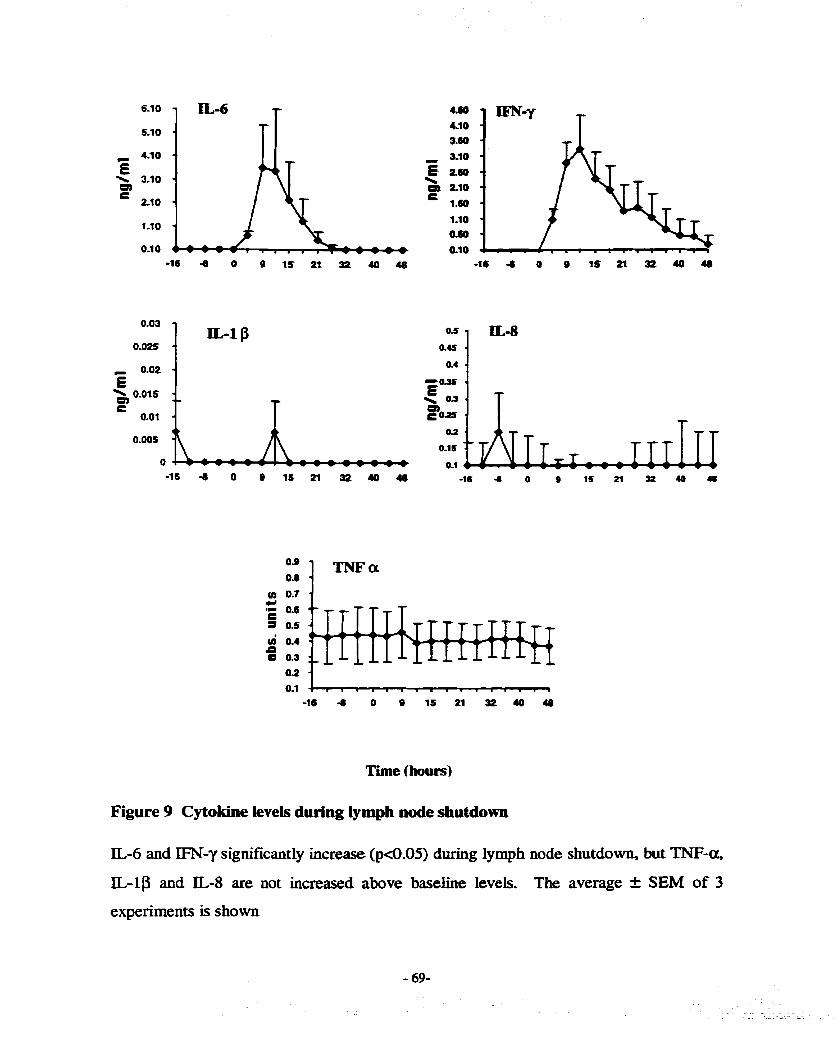
T i e (hourd
Figure 9 Cytokine levels during lymph node shutdown
IL6 and IFN-y significantly increase (p4.05) during lymph node shutdown, but TNF-ot,
IL-1P and IL-8 are not increased above baseline levels. The average i SEM of 3
experiments is shown

4.6. Discussion
The exact mechanïsm(s) and reason(s) for lymph node shutdown remains obscure.
Ic may have a role in the propagation of an efficient immune response by presenting Ag
to large numbers of lymphocytes (Mackay et al., 1992b; HaU and Morris, 196%)-
However, it does appear to be a valid p hysiological response to certain antigeas based on
the reproducibb shutdown observed by independent researchers.
Durhg shutdown, the lymph node output of aii measured lymphocyte subsets
decreased to at Ieast 50% of the^ basehe measurements (Figure 8). However, when the
data were examùied using the percentage of subsets as compared to baseline. it was found
that CD4 lymphocytes increased whilst CD8, y5 and B cells decreased. That is, the
overaii number of lymphocytes decreased, but of those present, the percentage of CD4
ceus increased by approximately 50%. This is not the fkst report of an increase in C m
cells during shutdown. Bujdoso et al (1989) reported that 24 hours after the injection of
PPD, CD4 cells increased and then seturned to normal after 3 days. Mackay et al (1992)
also noted an increase in the percentage of CD4 ceiis in efferent lymph during shutdown
and a decrease in the number of CD4 celis exiting the lymph node, similar to the present
report. Therefore, in a l l three studies a change in the output of CD4 lymphocytes in
efferent Lymph during lymph node shutdown was observed.
Two explanations are possible for the finding that CD4 ceils are less efficiently
retained during shutdown: A) CD4 ceils are migrating into the lymph node faster and
therefore present in a greater percentage than other subsets or B) non CD4 lymphocytes
are k i n g retained in the lymph node in a greater percentage. To determine if the tranic

of CD4 cells was increased, lymph nodes were excised dwing shutdown. There was no
difference in the percentage of CD4 cells between lymph nodes injected with saline or
PPD (Table S), arguhg that CD4 ceils were not migrating into Lymph nodes in a greater
percentage. Therefore, it was concluded that the change in CD4 ceiis was due to
regulation within the Lymph node.
Three-colour flow cytometry was used to examine the subsets of BL and LL in
the efferent lymph during shutdown. An increase the percentage in CFSE and Da-DS
CD4 cells was seen (Figure 8). This trend towards an increase in CD4 cells was
nonsignificant but was observed for both labeiled and nonlabelled cek. Though the
difference was not statisticaiiy significant there maybe a biological basis for the increase.
Therefore, it appears CD4 lymphocytes may be retaioed within the lymph node less
efficiently then other measured subsets,
Only efferent lymph was examined in these experiments. By dennition, ail
lymphocytes present in efferent lymph are part of the RLP, including blood derived
lymphocytes labelied with CFSE. The increase in both CFSE and Dit-DS labelled CD4
cells may indicate a population of celis that can quickly recirculate. pool would be
found in lymph and in transit through the blood, therefore it would be labelied with both
dyes. Under inflammatory conditions, this population of rapidly recirculating celis
enter the lymph node. If this specuiation is correct, these cells may have different
adhesio n rnolecule profiles thus facilitating more rapid recirculation. This population
may be quite small and therefore wouldn't be detectable in the recirculation of
lymphocytes under normal conditions. This population may be composed of memory T
cells as reported by others (Mackay et al., 1992b). However, this explanation does not

account for the increased migration of BL CD4 cells as the number of memory T ce& in
blood is similar to that in efferent Iymph @a@ et aL, 1999). A possible expianation is
that blood memory ce& have a greater prepouderance to migrate into infiamed lymph
nodes. This is fùrther discussed in section 7.5.
A Iess Likely explanation is that labelled c e k are preferentially recruited. This is
the least Likely of the possibilities as several studies have demonstrated no difference
between labelled and uniabeled ceh (Davenpeck et ai, 1995; Samlowski et al, 1991).
This was determined by performing flow cytomeuy to measure the amount of various
adhesion and activation molecules and kding no ciifference between unlabeled and
labelled ceils.
There is evidence in the Literature to support the specdation of multiple
populations of lymphocytes with varying degrees of migratory abilities. Mita and
colieagues have demonstrated a subset of y6 T cells in calves that can preferentially
localise in lymph nodes after the injection of TNF-a (Wilson et al, 1998). In other
experiments, this same group demonstrated an inabiliry of a subset of y6 T cells to
migrate into a skh site of idammation (Wilson et al, 1999). The y6 T cells were found
to express varying amounts of L-selectin and E-selectin ligands, resulting in differing
abilities to migrate into tissues. These data indicate that calves have multiple subsets of
y6 T ceils that Vary in their migratory capacity. Therefore, it is conceivable that sheep
also contain similar subsets, varying in their degree to migrate into sites of inflammation.
Some cytokines have an effect on the migration of lymphocytes when injected
into the drainage areas of lymph nodes (Young et aL, 2000; Hein and Supersaxo. 1988)-
IL-@, IL-6, IL-8, IFN-y and TNF-a levels were measured in efferent lymph aiter the

injection of PPD. IL-6 and IFN-y increased reaching maximum Levels at 12 hr, just prior
to the ceii output attaining its nadir at 16 hr. (Figure 9) EN-y has been demonstrated to
have a proadhesive effect for Lymphocytes on cultured skep lymphatic and bIood vesse1
endothelia1 ceils (Borron, 199 1). Therefore, it may increase adhesion molecules on p s t
capillary venules and lymphatic endothelid ceiis withui the lymph node, causing an
increase in both lymphocyte migration and retention. IL-6 has been demonstrated to be
important in the migration of lymphocytes into areas of inflammation by increasing the
expression of chemokines (Romano et al, 1997)- It is reasonabIe to assume that it has a
similar role in the infiamed lymph node and increases the amount of chemokines thereby
enhancing the entry of Lymphocytes. Conceivably, wndetected cytokines, iacluding
W-CX, could be produced in the lymph node but not secreted into efferent Lymph.
Immunohistochemistry on excised lymph nodes provided no evidence for their
production.
Taken together these data demonstrate that there is a decrease in Lymphocqtte
output of CD4 CD8, y6 T cells and CD21+ B cells. However, CD4 c e h may be the Ieast
affected, indicating that a subset of c e k may rapidly respond to the antigenic stimulation
of lymph nodes. IFN-y and IL6 are increased in lymph plasma and may have a role in
the increased migration and retention of lymphocytes during lymph node shutdown.

Chapter 5 Lymphocytes in cerebrospinal f luid are part of the
recirculating lymphocyte pool
5.1 Abstract
CSF contains a small number of lymphocytes under normal circumstances. To
determine if these lymphocytes are part of the normal recirculating pool of iymphocytes
efferent lymph was labelled, reinfused and samples of blood, lymph and CSF obtained.
Similar concentrations of IabelIed ce& were found in all three tissues. Based on these
data, it was concluded that wnactivated lymphocytes migrate into the CSE TO
determine if lymphocytes egress from the CSF, labeiled ceUs were infused h o the lateral
ventricle. Retropharyngeal lymph nodes contaiaed higher numbers of 1 1 1 -In labelled
lymphocytes as compared to other lymph nodes. Further experiments using labelied BL
and LL conclusively demonstrated that CSF lymphocytes belong to the RLP. This may
have implications for the immune surveillance of the CNS.
5.2 Introduction
The CNS was thought to be an immunologicaiy pnvileged site based on the
paucity of lymphocytes present in the brain parenchyma, lack of a lymphatic system and
the delayed rejection of transplanted tissue (Selmaj, 1996). However, studies have
demonstrated that there is immunological surveillance of the CNS, albeit at a lower level
than other tissues (W-iams and Hickey, 1995; Hickey, 199 1).

Activated T c e k enter the CNS parenchyma regardless of their antigen specincity
( Bauer et aI, 1998; aickey et aL, 1991). Activated Lymphocytes that encounter their
antigen in the CNS are retained and initiate an immune respome causing a non-specific
migration of Lymphocytes (Knopf et al 1998; Ludowyk et ai, 1992; Mor and Cohen,
L992;Cross et al, L99 1). Most investigations have not examined the entry of lymphocytes
into the CSF but rather have concentrated on the brain parenchyma, especially during
inflanimatory conditions such as multiple sclerosk (MS) (Hickey, 1991). Studies have
revealed that CSF and brain parenchyma dBer in their immunological potentid
therefore, resuks fiom one tissue can not be extrapolated to the other (Matyszak and
Perry, 1996).
The CSF under normal physiologicai situations contains a small number of
lymphocytes, with the majoricy behg CD4 T c e k (Vrethem et al, 1998; Svenningsson et
ai, 1995). Previous reports suggest CSF Lymphocytes are not a static population but may
enter and exit the CSF, irnplying that they are part of the RLP. Hafler and Weiner (1987)
in a study of MS patients, found that in vivo 1abeiIed CD4 Lymphocytes enter the CSF.
Lymphocytes and other cells injected into CSF and brain parenchyma migrate into
cervical iymph nodes (Carson et al, 1999; Oehmichen et al, 1979)- These same lymph
nodes drain fluids and proteins fiom the CNS (Boulton et ai, 1997; Cserr and Knopf,
1992), therefore, it is possible that Lymphocytes also use these pathways to exit CSE
Therefore, experiments were performed to detemine if lymphocytes in m~mal
CSF belong to LL. As well, cervical lymphatics were canulated and monitored for the
appearance of IabeiIed ceiis infused into CSF.

5.3 Materials and methods
The £irst experirnents examine the normal ceU concentraiion in CSF. Experiments
were then performed to determine if FïTC labelled lymphocytes migrate into the CSF and
detected ushg fiow cytomtry. The next experiments investigated the migration kioetics
of lymphocytes into the CSF and their concentrations as compared to efferent lympb. As
well, the migration of lymphocytes out of the CSF iato Lymph nodes and cervical lymph
was detennined,
5.3.1 Anirnals and surgery
Jugular veùis, prescapular, prefewral and ceMcal lymphatics were canulated as
described in Chapter 2. As weii, catheters were placed into the lateral ventricles to allow
the infusion of cells. Two sheep had successful laminectomies perfonned and catheters
introduced into the subdural space to allow seriai collections of CSF-
5.3.2 Detemiining normal CSF cell counts
The normal oumber and differential of ceIIs in the CSF of sheep was the first
experiment performed. This involved coilecting CSF ftom sheep involved in other
nonrelated studies as weii as this snidy. A lumbar puncture was performed by inserting a
needle between lumbar vertebrae 3 and 4 and aiiowing approximately 2 mi of CSF to drip
into a sterile tube. The CSF was immediately placed on ice and a ceU count performed
using a Neubaur chamber. Slides were prepared using a cytocentrifuge (S handon),
stained with a modified Wright stain and a differential perforrned. Samples contaminated
with red blood cells were discarded.

Efferent lymph ceiis were collected and labeiied with FITC as d e s c n i d in
chapter 2 and reinfused intravenously. In al1 experiments, greater than 80% of c e k were
viable using trypan blue exclusion and greater than 95% of the celis were labeiled with
FITC.
In some experiments, lymph lymphocytes were labeiIed with I l 1-Ih and infused
into the lateral ventricles. h experiments in which LL and BL lymphocytes were king
simultaneously investigated, blood was labelled with FITC and lymp h Lymphocytes wit h
DiI-DS.
5.3.4 Sample collection of blood, lymph, CSF and lymph nodes
Samples of blood, lymph and if required CSF were coiiected as needed. If
anùnals did not have catheters in the subdural space, CSF was coilected by performiag a
lumbar puncture. For kinetic studies, in which serial samples of CSF were required,
samples of between L and 2 ml were removed. For terminal sampling at the end of
experiments 2 to 10 ml of CSF was collected.
Samples of blood and lymph were prepared as descnid in Chapter 2. CSF
samples required more care, as few lymphocytes were present. To avoid loss and
maintain the integrity of cells, CSF was immediately placed on ice and an equal volume
of paraformaldehyde added after an aliquot was removed for a cell count. Cenaifugation
was avoided if less then 2 ml of CSF was coliected, instead the whole sample was
analysed by flow cytometry.

Prescapular, prefemoral and popliteal lymph nodes were harvested at the end of
severai experiments. They were immediately bisected, wmpped in saIine soaked gauze
and placed on ice. To harvest Lymphocytes, the tissues were muiced in cold media,
fdtered and washed twice in PBS. These ceUs were mtered, paraformaidehyde added and
processed using identical methods as for lymph samples.
5.3.4 Intracerebroventricular infusions of 1 1 1 -In labelled lymphocytes
To determine if labelied lymphocytes can migrate from CSF hto cenical lymph
nodes, 11 1-In labelled c e k were infused into the lateral ventricIes in 3 sheep. Between
2-5 x 10' ceils were infused. At 24 hrs (nd) and 48 hrs (n=2) the sheep were sacriuced
and various lymph nodes and control tissues excised. These tissues were weighed,
counted on a y-spectrometer and the counts per minutefgram of tissue deterrnined TO
ensure that the radioactivity was cell associated, the retropharyngeal lymph nodes were
minced, washed with PBS and the radioactivity associated with the cells detemiined. The
majority of the radioactivity (>go%) was celi associated.
5.3.5 Intracerebroventricular infusions of FlTC labelled lymphocyte
To determine if cervical lymphatics are a path of exit for CSF Lymphocytes, these
lymp hatics were canulated. Lymphocytes were coilected labelled wit h FJTC, suspended
in 1 ml of saline and infbsed into the lateral verticais. Samples of cervical lymph were
collected for flow cpometry. To ensure that the celi infusions were sterile, agar was
inoculated with samples of celi solution. No samples tested grew bacteria after 4 days.

5.3.6 Flow cytometry
AU samples were analysed using a FACScan (Becton Dickinson). Srnail
lymphocytes were analysed based on their olpicai forward and side Light scatter
properties. Aliquots of iabeiied and unlabeled lymphocytes were used as controis.
The percentage of FKC, and if required, DiI-DS labeiled tek in each sample
were determined For blood and lymph, at least 1 6 lymphocytes were analysed but
because of low numben of ceils present in CSF, oniy 1o3 lymphocytes codd be
examined.
5.4 Results
5.4.1 Cells in normal CSF of sheep
Samples of CSF were coliected from 7 sheep and a cell count deterrnined. The
mean of these counts was 3.0 + 0.3 x 10~/ml, No red blood cells were seen in the those
samples used to determine this count, therefore the samples were not contaminate with
blood. The differential showed that the majority of these cells were small lymphocytes
but, because of the low number of ceils present, an accurate differential could not be
performed.
5 A.2 Repeated infusion of labelled lymphocytes
The next series of experiments was carried out to determine if FITC labelied
lymphocytes could migrate into CSF and be detected using flow cytometry. Ail coliected

efferent lymph was iabelled with HTC and intravenously reinfused at regular intervals
(approximately 12 hr) for a minimum of 3 days in 3 sheep. Between 5-10 x 10' ce&
were infused. Samples of blood, lymph, CSF and lymph nodes were coiiected 24 hotus
after the last infusion and the percentage of FITC labeiled c e k determined. Slmilit~~
concentrations of labelled cells were found in all examined tissues within the same
experiment. Because of the large differences between the nmber of injected ceh. these
data could w t be pooled. Representative histograms h m one experiment c m be seen in
Figure 10. These data demonstrate that JTC labelled lymphocytes migrate into and are
detected in CSF. Next experiments using a single bolus of labeiied lymphocytes were
performed, allowing kinetic studies.
5.4.3 Single bolus of labelled efferent lymphocytes
To determine the percentage of FXTC labelled efferent lymphocytes in tissues of
interest after a single infusion of ceiis, samples of CSF, blood. lymph and lymph nodes
were collected 24 hours postinfusion. The percentage of labelled cells was as follows
CSF 0.6 1 % I 0.26. efferent Lymph 0.50% I O. L6 and lymph nodes 0.35% f 0.13. There
was no signifrcant ciifference between any of the tissues sampled both in the same
experiment and in the Gnal average (Table 6). Fiepre LI is composed of representative
histograms fIom one experiment. Subcutaneous efferent lymph and CSF appear to share
the same population of lymphocytes.

Figure 10 Percentage of IabeMed ceiis in tissues afkr constant reinfusion
Representative histograms nom one of three experiments in which lymphocytes were
coliected and intravenously infüsed over three days. LabeUed cells are clearly seen in the
CSF, blood, efferent lymph and lyrnph node,
A - FITC labded ceils and negative control
B - CSF before the infiision of cells
C - efferent subcutaneous lymph
D - CSF
E - blood
F - popLiteai Lymph node

Log fluorescence intensity

Table 6 Percentage FITC lnbelled efferent lymphocytes in CSF, Iymph and lymph nodes after a single uifiision
There is no signifïcant merence (p < 0.05) in the percentage of FïTC hbeiled cek 24 hr
after a single intravenous infusion of ceils. ND = not determined
* = percentage labeiied cells & SEM
Exp. number CSF Lymph Lymph node 1 O. 16 + 0-04 0.23 I 0.05 0.17 + 0.04 2 0.75 + 0-05 0.75 t 0.05 0.60 + 0.05 3 0.5 L t 0.08 0.49 I 0.02 0.58 t 0.02 4 0.1 1 t 0-02 0.07 f 0.02 0.05 + 0-0 I 5 1.55 t 0.11 0.97 f 0.03 ND
Average 0.6 1 + 0-26 0.50 f O- L6 0-35 k O. 13

5.4.4 Kinetics of FlTC labelled cells in CSF and efferent Lymph
Multiple samples of CSF and eEerent lymph were collected over a 24 hr sampling
period. In both sheep, in which successful Iaminectomies were performed, labened
lymphocytes appeared in approoximately the same concentrations and at similar times in
CSF and efferent lymph (Figure 12). Using regession analysis the correlation between
appearance of lymphocytes in lymph and CSF in the separate experiments was 0.6L and
0.98. These data indicate that subcutaneous efferent lymph and CSF share the same pool
of recirculating Lymphocytes.
5.4.5 Lymphocyte egress from CSF
If Lymphocytes in the CSF are part of the recirculating Lymphocyte pool then they
must be able to exit the CSF- To investigate this, Ill-In labelled cells were Uifused into
the laterai ventricles and radioactivity determined in tissues, In 3 experiments, lymph
nodes known to drain fluids and proteins fiom CSF had greater radioactivity than distal
lymph nodes or control tissues (Figure 13). These data were not pooled as different
amounts of IabeUed cells were infirsed and tissues harvested at 24 and 48 hr-
Lymphocytes appear to migrate fiom the CSF and enter retropharyngeal lymph nodes,
perhaps using the same pathways descn id for proteins.
To determine if labelled c e k migrate from the CSF using cervical lymphatics,
FITC labelled ce& were infused into the lateral ventricles and cervical lymph mnitored
for their appearance. In only 2 of 6 experiments codd ETïC labelled cells be detected in
cervical lymph (data not shown). However, retropharyngeal lymph nodes hamsted at

the termination of experïments, contained FITC IabeIIed ceiis. Therefore, E7ITC Iabeiied
c e k were able to migrate from the CSF. Experïments were perforxned to confiml that
the canulated cervicaI lymphatics were communicating with the CNS. In 2 experiments
in which iabelled c e k couid not be detected in cervical lymph radioactive albumin was
infused into the lateral ventricles. In both experlments, radioactive albumin was detected
in cervical lymph, connrming that the correct lymphatics were canulated.
5.4.6 LL and BL migration hto CSF and afferent lymph
Table 7 shows that LL (0.52% f 0.15) are present in CSF at greater
concentrations than BL (0.036 k 0.02). There was no significant dinerence in the
percentage of labeiied LL in CSF (0.52% + 0.19, blood (0.454 & 0.10) and efferent
lymph (0.79% + 0.19). However, labelled BL were found to be ~ign~caatly lower in
CSF (0.03% + 0.02) as compared to blood (0.72% f 0.13)-
Expressing the data as percentage injected ceils corrects for the daerence of
infused cells. Using this method LL are present at 1.07 x 106% injected I 0.44 and BL
1-15 x 10-~% injected & 0.86 (n=S)(Figure 15). This coafïrms that LL enter CSF in
approxbmtely 10X greater numbers than BL.
Both LL and BL are present in Herent lymph in higher concentrations than CSF.
Nevertheless, the RLP is present in both tissues at a higher percentage than BL.

Figure 11 Percentage of iabeüed lymphocytes 24 hr &et a single inhision of cells
Representative histograms 24 hr after a single intravenous uihision of FITC iabeiied
subcutaneous efferent lymphocytes. Similar concenaations of FïïC labelled cells are
seen in di tissues examined-
A - FITC labeiied and negative lymphocytes
B - efferent lymph
C - CSF D - blood
E - prefemoral lymph node

Log fluorescence intensity

8 14 19 24
Time (hrs)
Figure 12 Appeanuiee of EïïC iabeUed lymphocytes in CSF and lymph
In two experiments, CSF and lymph were monitored for the appearance of labeiled ceik
after a single infusion of FITC labelled lymphocytes. Labeiied cells were detected in
sirnilar concentrations and the in both compartrnents. One experiment is s hown above.

Figure 13 Intracerebmventcic~~r injectecl Ill-In bbelled ceUs migmte to lymph nodes known to drain CSF
After the intracerebroventricular infusion of 11 1-In iabelled lymphocytes, they are found
at higher concentrations in lymph nodes known to drain CSF.
Retro : retropharyngeal lymph node
Mand: submandibdar lymph node
Prefem: prefemord lymph node
Mesent: mesenteric lymph node


Table 7 Percentage labelled LL and BL in CSF, blood, and lymph
A The Di[-DS IabeUed LL were aiways present in higher concentrations than FITC
labeiied BL in CSE The percentage of FiTC labeiied cek in blood was sigoificantly
different than in CSF (pd.0 1). No significant difference (pM.05) was seen when the
percentage DiI-DS labeiled ceils was compared between tissues.
B In normal CSF and afferent lymph the DiI-DS labelled RLP is present in a higher
percentage. There is no signincant ciifference (p>0.05) between the percentage of
labeiied ceiis in afferent lymph and CSF,

A Percentage iabelled in blood, CSF andefferent lymph
Percenta~e FITC labelIed Percentaee DiI-DS labelled Exp. number Lymph Blood CSF Lymph Blood CSF
1 0.33 0.58 0.09 0.79 0.60 0.79
6 0. 15 0.4 1 O 1.56 0.58 1-10 Avg- t SEM 0.25 f 0.02 0.72 I0.13 0.03 k0.02 0.79 i0.19 0.45 &0.10 0.52 I0.15
B Percentage Iabeiied in Pnerent lymph and CSF
Afferent lvm~h - CSF Exp. number 9% FiTC % Dïi-DS % FITC % DiI-DS
4 0.54 0.80 O O. 17 5 0.62 0.44 O 0.59 7 0.87 1-41 0.44 0.44
Avg. t SEM 0.67 t 0.10 0.88 +, 0.28 0-15 2-14 0.38 +0.11

5.5 Discussion
Tissues are continuously patroiled by Lymphocytes, but it was believed that the
CNS was spared this surveiiiance (Selmaj, 1996). However, it has become increasingly
obvious that activated Lymphocytes migrate into the CNS, albeit at lower nurnbers than
most tissues (Hîckey, 1999). Normal CSF contains a srnail number of lymphocytes
(Svenningsson et ai., 1995), but brain parenchyma is vùnially devoid of Lymphocytes.
Matyszak and Perry (1996) injected BCG into the brain parenchyma and the CSF and
demonstrated that these two compartments dBer in their immune response. The
inflammatory response was Iimited in the parenchyma but the CSF immune response was
similar to that seen in the periphery. These data together illustrate that the CSF and brain
parenchyma can not be cousidered as the same tissue with respect to their interactions
with the immune system,
Several connections exist between the CNS and immune system, It has been
clearly demonstrated that antigeos infused into the CNS drain Uito cervical Lymph nodes
and stimulate an immune response (Knopf et al, 1995)- Cytokines in the periphery can
enter the CNS (Pan et al, 1997a), while those in the CSF exit via the cervicai lymphatics
(Dickstein et al, 1999). Recent experiments have demonstrated that tbe removal of
cervical Lymph nodes lessens the impact of EAE in rats (Philiïps et al, 1997). Taken
together these data prove that communication exists between the CNS and the immune
systern.
If CSF lymphocytes belong to the RLP, this would be yet another connection
between the CNS and the immune system. Therefore, experiments were desigoed and

perfomd to test if this connection is present. First, the normal concentration of cells
within the CSF of sheep was determuied to be 3.0 x 1o3 celWrnl f 0.3 which is similar to
humans (Svennhgsson et al., 1995). The major* of celis are srnail, nomial appearing
lymphocytes.
M e r repeated intravenous infusions of FITC Iabelled efferent LL over severai
days, labelled ceiis were identifîed in CSF- In 3 separate experiments, the percentage of
1abeIIed ce& was similar in blood, CSF, efferent lymph and Lymph nodes (Figure 10).
These preliminary experiments were required to eosure that iabelled cells could be
detected using our methodologies and to confirm that lymphocytes migrate into the CSF.
In agreement with previous reports @LUC et aL, 1994)- it was critical to place CSF
samples on ice and avoid prolonged cenvifugation to maintain the integrïty of the ceh.
Next, experiments using a singie bolus of labelied cells were performed allowing
for kinetic studies (Table 6). Efferent lymph and CSF were found to contain similar
percentages of labelled cek, 0.50% f 0.16 and 0.618 k 0.26 respectively (n=S)-
Additionally, in 2 experiments, multiple samples of CSF and efferent lymph were
coiiected over a 24 hr period and found to contain sunilar percentages of labelied cells
(Figure 12). Taken together these data ïmply that lymphocytes in CSF are part of the
EUP. Further experiments labelling both blood and efferent lymph lymphocytes
confirmed this (Table 7). Di[-DS labeiled LL were found in higher percentage than FI'ïC
labelied BL, (0.52% f 0.15 vs. 0.03% +, 0.02). The few FLTC labelled cells detected in
CSF rnay belong to the RLP but were labelled as they were in transit through blood- A
siagpSicznt ciifference (pc0.01) in the percentage of FITC labelied cells in blood and CSF,
0.72% f 0.13 vs. 0.03% I0.02. This implies that BL do not migrate weii fiom blood into

CSF. Expressing the data as percentage of injected ceiis codirmed that U are found in
greater numbers than BL (Figure 17)- arefore, m normal CSF LL m-grate into the CSF
in greater numbers than BL.
These data imply that lymphocytes are wt reqoired to be activated to migrate into
n o r d CSE Previous studies have examined the phenotype of Ipphocytes in CSF and
concluded that the majority of celis in CSF are activated CD4 celis (Vrethem et ai.,L998;
Scoloui et ai., 1992; Mix et aL, 1990). Others have disputed these kdings and argue
that CSF cells do not need to be activated (Svenningsson et ai., 1995; Kleine et al., 1999).
Several reasons may account for these discrepancies including the selection of patients.
Often subjects are recruited fiom patients seen in hospitals and cluiics and have
underlying non-inflammatory neurological problems but are considered as 'cnormai".
However, Svenningsson et ai. (1995) examined healthy volunteers without a history of
neurologicai disease and did not îind that Lymphocytes isolated from CSF expressed
activation markers. Another explanation for these discrepancies is the variations in
critena that different investigators use to determine activation, Vrethem et ai. (1998)
w d the expression of CD45RA+ to assign activation status, whilst Svenningsson et aL
(1995) utilise CD25 (IL-2 receptor). The results repted here support the findiags that
nonactivated cells migrate into CSF. This statement is based on the fact that normal
efferent lymph contains approdtely 5% activated celis as measured by CD25
expression (Haig et al., 1999). As weii, the majority of lymphocytes in efferent lymph
are small noncycling lymphocytes, thereby implying a low level of activation- Therefore,
if oniy activated lymphocytes migrated into CSF, we would expect to fhd Iow ievels of

k h d e d ce&. W e fouad sirnilar levels of labeiled cells in both lymph and CSF implying
that nonactivated lymphocytes do migrate into CSF.
If these Lymphocytes are tculy part of the RLP, they must be able to exit the CSF.
Experiments using 111-In labelied lymphocytes infused into the lateral ventricles found
amounts of radioactivity in retmpharyngeal lymph nodes that was 5 times higher than
other lymph nodes (Figure 13). Previous reports have also demonstrated this (Carson et
ai., 1999; Oehmichen et al., 1979).
Boulton and colleagues (1997) demonstrated that cervical lymphatics in sheep are
responsible for transporting approximately 50% of a protein tracer infused into CSF-
These same pathways may be used by lymphocytes to migrate into the retropharyngeai
lymph node. To inveaigate this, cervical lymphatics were canulated and monitored for
intracerebroventricular infused FITC labeiied cells. In only 2 of 6 experiments labelled
celis were detected in cervical Lymph, nonetheless, in every experiment labelled cefi
were found in retropharyngeal lymph nodes at necropsy. These data demonstrate that
labeiled cells appear to migrate using the cervical lymphatics and enter retmpharyngeal
lymph nodes. There are severai reasons that iabelled cells were not consistently found in
cervical lyrnph. First, the number of labelled c e k would have ken a minor population
of lymphocytes in cervical lymph and possibly below our of detection. Secondly,
the infusion of a large number of lymphocytes (approximately 2 - 5 x 108j, initiated an
inIlammatory response as seen by an increase in neutrophils in the CSE Lastly, the large
number of celis may cause a physical obstruction of the pathways used by the
lymphocytes to exit, as under normal conditions oniy thousands of cells would migrate
every day.

hmrine surveiiIance of CSF occurs under normal conditions as shown by the
ingress and egress of lymphocytes. The data presented in this chapter are the moa
complete nirvey of this dynamic process to date and demonstrates that U are found in
CSF in greater numbers than BL. hdeed LL are also found in a greater percentage in
&erent lymph, indicating that this pool has a larger role in tissue surveillance, includuig
the CSF. Percentages of labelied ceiis in tissues is not a perfect method to determine
migration, as differing amounts of infûsed cells will impact this measmement but
measuring percentage of injected ceIls accounts for this varïability. Expressing the data
with this method contirms that LL are found in a greater percentage than BL in CSF.
Unfortunately, the ce11 concentration of afferent lymph was not measured, so the direct
cornparison of these two tissues can not be made. Nevertheless, the data clearly shows
that U migrate into CSF in a greater numbers then BL,
Taken together, these data demonsuate that those lymphocytes present in CSF
under normal physiological conditions are mainly from the RLP. Lymphocytes do
recirculate through the CSF and perhaps have a role in the immune surveillance of the
CNS -

Chapter 6 TNFa injections into CSF, but not the brain parenchyma, results in leukocyte reccuitment
No tissue outside of the Lymphatic system has been investigated for the migration
of LL and BL under idlammatory conditions. Under normal conditions, lymphocytes
belonging to the RLP are present in CSF. htracerebroventricular injections of TNF-a
were used to recnùt lymphocytes into CSF and labeiled ceils fiom both pools were
quantified. TM-a induced a . inEIammatory response in CSF, which by 48 hours
postinfusion was predominately composed of CDS+ lymphocytes. Lymphocytes fiom
blood were present in the infiamed CSF in greater percentage than LL. Injections of the
same amount of TNF-a into the parenchyma of the brain did not hcrease CSF cell counts
and resdted in minimal leukocyte recruitment. These data provide evidence that BL
respond to inflammation in the CSF in a greater percentage as compared to LL. As well,
these data provide fiirther evidence that the CSF and CNS respond differently to
inflammatory stimuli.
6.2 Introduction
During meningitis and sorne infl-tory reactions within the CNS the number
of leukocytes in CSF hcreases. These ieukocytes may be a mixture of neutrophils,
monocytes or lymphocytes dependhg on the stimuli (Schoning et ai, 1999; Bamborschke

et 1990)- Cytokines, chemokines and adhesion molecules orchestrate tk migration of
leukocytes into CSF (Spelierberg and Tuornuien, 1994).
The CSF and brain parenchyma differ in their immuw Iogical response to antigen.
BCG injected ioto the CSF resuits in a robust immune response, but has a minimai effect
when injected into the parenchyma (Matyszak and Perry, 1996). Injections of TNF-a
into CSF resdts in increases in leukocytes (Schoning et aL, 1999) but has v h & y no
effect when injected into the brain parenchyma (Andersson et aL, 1992). This maybe due
to varying concentrations of neuropeptides, immunosuppressant factors or ciifferences in
vasculature (Phillips and Lampson, 1999). Therefore, the CSF and the brain parenchyma
must be considered as separate compartments with respect to inflammation and leukocyte
migration.
Dickstein et al. (1999) have shown that radioactive labelled TNF-a can be
transported to the retropharyngeal lymph nodes and cem-cd lymph. The function of this,
if any, bas not been elucidated. The sanie lymph nodes are important in the immune
response to CNS infused antigen (Harling-Berg et al 1999; Harhg-Berg et al., 1989). It
is conceivable that TNF-a kv ing in the lymph node with antigen may potentiate the
immune response by increasing lymphocyte tranic into the lymph node.
Studies were performed to determine if inuacerebrovenuicular injections of Tb?F-
a înduced inflammatory conditions in sheep. Additionally, the effect of TNF-a on BL
and LL migration into the CSF, lymphocyte subsets in the idammatory infiltrate and its
effect on cervical lymph was investigated. The migration of lpphocytes into CSF
injections of TNFa into brain parenchyma was also examined.

6.3 Materials and methods
6.3.1 Animais and surgery
AU experiments were carrïed out in the Division of Comparative Medicine at the
University of Toronto. Sheep of approximately 6-8 months and approxbately 30 kg
were obtained from Boxwood famis. Surgeries were performed as outlined in Chapter 2
including cenical, prescapular and prefemoral efferent Iymphatic cannuiation- Guide
screws were irnplanted at least 5 days prior to the lymphatic cannulations to allow
adequate healing.
6.3.2 Cell labelling
Blood c e k were labelled with FITC and lymph lymphocytes with DiI-DS as
explained in chapter 2, Cells were reinfused and allowed to equili'brate for at least 24 hrs.
6.3.3 TNF-a injections
Five hundred ng of recombinant human TNF-a (Cederiane Hornby ON) was
diluted in 1 ml of sterile saline and infiised into the lateral ventricles using the implmted
guide screws. Two Werent lots of TNF-a were used, both giving similar results-
Previous studies have shown human TNF-a WU induce an inflammatory response in
sheep (Kallaaji et al., 1989).
If the cytokine was to be injected into the brain parenchyma, 500 ng was diluted
in 25 pl of saline. It was injected approximately 1 cm into the cortex overlying the lateral
ventricles using a Hamilton syringe. The syringe was slowly removed after king left in

place for a few minutes to prevent backaow dong the needle track The opposite side of
the brain was lnjected with saline to serve as a control-
6.3.4 CSF collection, differential and phenotyping
CSF was coiiected by lumbar puncture as previously descnibed in Chapter 5. A
cell count was determined and a slide made by cytosph This slide was stained with a
modified Wright stain and a differential performed. CSF ceiis were washed twice with
PBS and at l e s t 20 000 cells immunophenotyped as per Chapter 2.
6.3.5 Brain tissue collection
Sheep were sacrficed and the braia immediately removed- If TNF-a had been
hjected ïnto the brain, the areas under the guide screws were exckd dong with control
areas. If the cytokine had been infused into the CSF, representative areas of cortex,
choroid plexus and areas surroundhg the ventricles were removed. Brains fiom control
animais were obtained which had never experïenced any CNS surgery and therefore
served as normal controls.
The tissue was placed in saline soaked gauze and placed on ice as soon as it was
removed It was then embedded in OCT, fiozen in liquid nitrogen and stored at -80°C.
6.3.6 lmmunohistochemistry
Blocks of brain and control tissues were cut at 6-8 pm using a cryostat and ptaced
on poly-L Lysine coated slides. These slides were ailowed to dry overnight and stored at -
20°C until use. hunohistochemistry was perfonned as outlined in Chapter 2.

6.4 Results
6.4.1 Leukocyte number, differential and phenotype after the
intracerebroventricular infusion of TNF-a
M e r the infusion of 500 ng of TNF-a into the iateral ventricle, there was a ciramatic
bcrease in leukocyte numben. At 24 and 48 hm postinfusion the leukocyte count was
5.7 x 10' t 1.4 (n=8) and 1.3 x 106 + 0.5 (n=7) respectively as compared to the n o d
count of 3 -0 x ld/ml (p c 0.05) (Figure 14).
Under normal circumstances, the leukocytes present in CSF are predominately
smdl Lymphocytes. Twenty-four hours after the infusion of TNF-a the majonty of
leukocytes in the CSF were neutrophils and monocytes with Lymphocytes king a s d
population (Table 8). However, by 48 hrs postinjection lymphocytes composed
approximately 70% of the leukocytes and neutrophils had decreased.
Using flow cytometry the lymphocyte phenotype was determîned. At 24 hr (nd)
CD4 cells were the predominate lymphocyte subset present (38.5% f 8.0) and CD8 celis
were lower (14.6% k 4.0). However 48 hr postinhision CD8 cells had increased to 37.2%
t 6.0 and CD4 c e k had decreased to 14.6% + 4.0 (n=3). B c e k and y6 T ceils were
minor populations of approximately 15 4b at both tirnes (Table 9).
6.4.2 CSF leu kocyte numbers after brain parenchymal injections of TNF-a
Forty-eight hours after the parencymal injection of 500 ng of rhTNF-a there was
no increase in the CSF cellulanty (2.0 x lo3/ml f 0.5 (n=3)) as compared to normal CSF
(3 -0 x 10' f 0.3 cells/d. Because of this low ceil number no phenotype or dBerentia.1
couid be performed.

6.4.3 BL and LL migration into CSF after TNF-a injection
After the infusion of T M - a into the CSF, FKC labelled BL increased and
surpassed the percentage injected of Dit-DS IabelIed LL, 2.7 x 10-~% I 1.3 vs. 81) x 10-
4 % f 3 (n=6). The ciifference between the two pools was not statisticaiiy signincant
(Figure 15). Forty-eight houn was chosen as the samphg t h e as previous experiments
had shown that this was the tune of maximum Lymphocyte recruitment. h 5 of 6
experiments, the percentage of m C labeued celk was higher in CSF as compared to
lymph but was aiways lower than blood (Table 10). DI[-DS labeiied LC in 5 of 5
experiments were lower in CSF as compared to lymph but varied as compared to blood.
Due to various technical difficulties including poor labelling of ceils and blood
contaminated CSF collections, the perceotages of the two pools could not be determined
after the parenchpal injections of TNF-a.
6.4.4 Irnmunohistochemistry
The ~unohistochernical analysis of the brain removed after the CSF infusion of
500 ng of TNF-a demonstrated a widespread infiammatory infiltrate surroundhg vessels
(n4). The phenotype of this appeared to mirror that found in the CSF, that is CD8
lymphocytes predominated but populations of CD4 and y6 T ceils were present (Figure
16). B cells and CD25+ cells were rarely detected The anti-CD25 aotibody used in
these studies has not k e n previously characterised for immunohistochemïstr~ and
therefore the results should be interpreted with caution. Lectin stalliing ushg GSA 1-B4
was perfonned for the detection of macrophages, wbich were located in small numbers in

the infiltrate. EndotheIIal ceils were &O stained with this Iectin as previousiy reported
(Kmeger et al, 1995).
The choroid plexus was examuied 48 hours after the intracerebroventricular
infusion of TNF-a. In control choroid plexus, macrophages were seen as were CD4
CD8 and y6 T ceiis (Figure 17). M e r the intracerebroventcicular injection of TNF-a the
numbers of ceiis appeared to increase and were located around the vendes and in the
stroma of the choroid plexus. However, without quantitative measures, the results must
be interpreted with caution, but it appears that TNF-a injections increase the number of
lymphocytes in the choroid plexus,
In contrast, the injection of TNF-a into the cortex of 3 sheep demonstrated very
different results. At the site of injection there was iïttle increase in the number of
leukocytes over that found in the saline control injection site (Figure 18)- In control areas
distant fkom the site of injection, no Lymphocyte infiltrate was seen.
There appeared to be an increase in the amount of VCAM on small vesseis in the
brain and the choroid plexus afier the CSF infusion of TNF-a. However, it was diffcult
to quantify.
Control brain tissue removed from sheep with no CNS surgery had vimidy no
CD4, y6, or B ceus present but some CD8+ stainuig ceiis were scattered throughout the
parenchyma. It seems unlikely that CD8 lymphocytes are present in the normal sheep
brain as lymphocytes are rarely seen in the <3NS of other animals but there are no prier
studies in sheep for cornparison. Ail tissuas with normal muse IgG or without primary
anti'body exhibited minimal background staining.

normal 24 hours 48 hours
Figure 14 CSF celiularity increases after the injection of TNF-a
Mer the intracerebrovenuicu1ar injection of 500 ng of rhTNF-a the number of
leukocytes increases significantly (@.OS) at both 24 ( 0 ~ 8 ) and 48 hr (n=7).

Table 8 Leukoryte difEereotia1 M e r the intracerebroventricular injection of TNF-a
Twenty-four hours after the injection of TNF-a the leukocyte idtrate is dominated by
neutrophils and monocytes (n=9). However, at 48 hours lymphocytes predominate (n=!5)-
Leukocyte type 24 hours 4û ~OULS
neu trophils 68 t4 10t4
monocytes 2 L I l 17f 6
lymphocytes 10tL 72 & 10

Table 9 Lymphocyte subsets present after the intracerebmventriculr hjection of TNF-a
There is a change in the subsets of lymphocytes present in the CSF after the injection of
TNF-a. At 24 hours postinjection (UA) CD4 c e k are the predominate lymphocyte
present, but by 48 hours (n=3) CD8 ce& predominate.
Note that due to the s d amount of CSF coilected only 1 sample was
immunophenotyped for y8 T ceiis at 48 hours. There is no signifcant ciifference (p
0.05) between any of the subset percentages at 24 and 48 hr-
Lymphocyte Subset 24 hours 48 houls

Figure 15 Both BL and LL increase after the intracerebroventricular injection of TNF-a
DZ-DS Iabeiled LL are present in a greater amount in normal CSF as compaced to FIT%
labeiied BL (n=5). Both DiI-DS IabeIled and FlTC labeiled lymphocytes increase
significantly after the injection of 500 ng rhTNF-a (pc0.05). FITC labeiied BL increase
to a greater extent than Da-DS IabeUed LL after the injection of TNF-a (nt@. The
percentage of iojected celis was used to quantify the ce& as it accounts for the clifferhg
amounts of infUsed IabelIed lymphocytes.
Samples of CSF were coliected 48 hours &er the infusion of 500 ng of rhTNF-a into the
lateral ventricIe.
A - normai CSF
B - CSF after the rhTNF-a injection

% injected DiI-OS O h injected FlTC
B CSF 48 hr &ter a TNF-a injection
% injected Dii-DS % iniected FKC
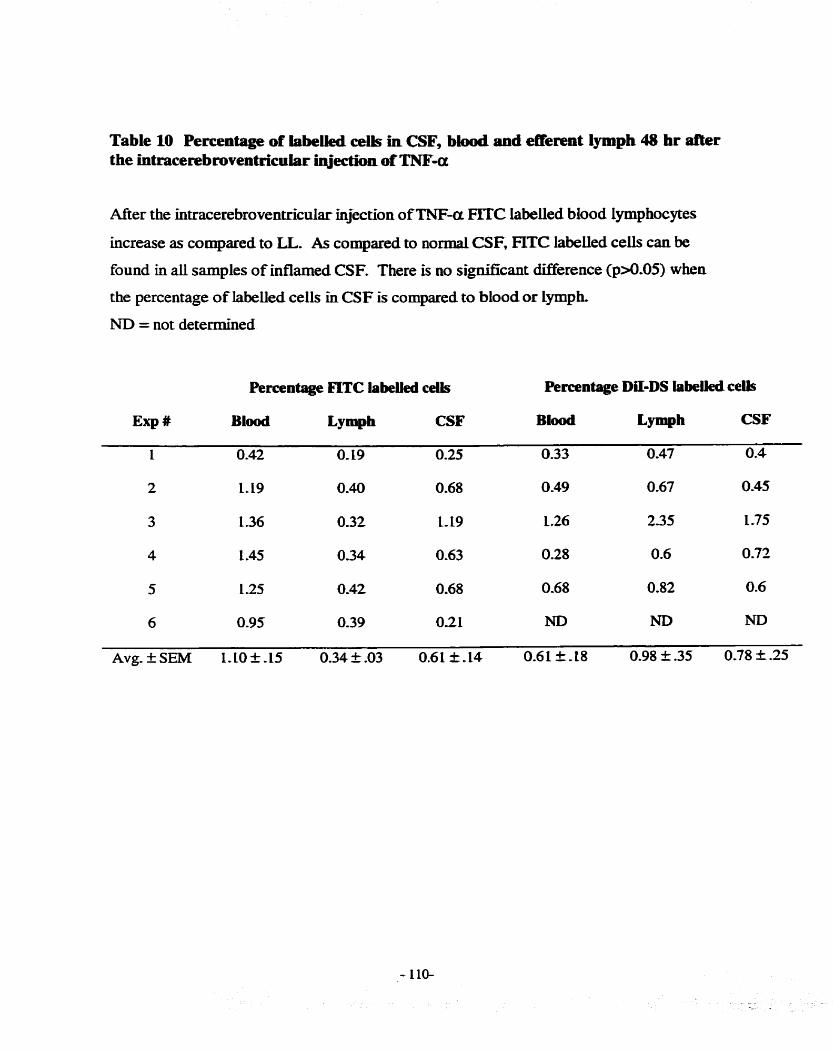
Table 10 Percentage of iabeiïed c e b in CSF, biood and efferent lymph 48 hr after the intracerebroventcicular îqjection of TNF-a
After the intracerebr~vent~cular uijection of TNF-a FITC labeiied blood Lymphocytes
increase as compared to LL. As compared to nomial CSF, FïTC labeiied c e k c m be
found in aU sarnples of inflamed CSF. There is no signincant difference (p>O-05) when
the percentage of labeiied ceils in CSF is compared to blood or Lymph-
ND = not determined
Percentage FLTC labeiled ceüs Percentage DiI-DS iabelled ceUs
E ~ P # B l d L Y ~ P ~ CSF Blood L Y ~ P ~ CSF
1 0.42 0, 19 0.25 0.33 0.47 0.4

Figure 16 CNS parenchyma after an intr~rerebroventn*calar injection of TNF-OC
An idammatory infiltrate is seen in the perivascular spaces throughout the CNS 48 h .
after the intracerebroventricular infusion of rhTNF-cm CD4 ceiIs are present in the
infiltrate (A) but CD8t cells predominate (B). y6 T c e k (C) are also present. MHC II is
upre,oulated on what appear to be endotheliai ceils (D), even in areas with few leukocytes
in the perîvascular space. There is a scattering of unidentined ceiis in the b r d
parenchyma that express CD8 (E) but there are no y6 T cek (F) in normal sheep. AU
photos were printed at 200x mgnification-
A - CD4 ceiis
B - CD8 ceils
C - y6 T ceiis
D-MHCII
E - CD8 staining in control (normal) brain
F - y6 staîning in control ( n o d ) brain
- ILL-


Figure 17 Choroid plexus Pfter an intracerebroventricular iqjection of rhTNF-a
In n o d choroid plexus, CD8 stalliing c e k are present in the stroma (A), but after 48 hr
after the injection of TNF-a there appears to be a slight increase in numbers (B). MHC II
staining is also present in stromal ceiis under normal conditions (C). ikely due to resident
macrophages. Forty-eight hours after the injection of T3JF-a the number of ceh appears
to increase, but this may be due to an upregulation of the amount of MHC II on celis @)-
There is a s d amount of VCAM stainuig on seromal cells (E), which appears to
increase after the injection of TNF-a.
A - CD8 normal choroid plexus (2ûûx magnification)
B - CD8 alter the injection of TNFa (2ûûx magnification)
C - MHC II normal choroid plexus (400~ magnification)
D - MHC II post TNF-a injection (400x magnification)
E - VCAM normal choroid plexus (400x magnification)
F - VCAM after TNF-a injection (200x magnification)


Figure 18 CNS pareachyma d e r the intmcerebral iqiection of rbTNF-a
- - There is minrmal hfiammatory infiltrate 48 hr after the injection of rhTNF-a into the
parenchyma as compared to the injection of saline. Few CD4+ ceils are seen in the area
after TNF-a (A) or saline (B). TNF-a (C) and saluie @) sites of injection attract
minimal numbers of CD8+ cek or y6 T cek. AU photos were p ~ t e d at 400x
magnific atio n,
A - CD4 cells afier a parenchymai injection ofTNF-a
B - CD4 ceiis after a control injection of saLine
C - CDS+ ceUs post TNF-a injection
D - CD8+ cek d e r a saline injection
E - y6 T ceils after TNFa
F - y6 T ceiis post saline injection


Figure 19 Iatracerebroventncuiar injection ofrhTNF-a has no effect on cervical lymph flow or ce11uiarity
Five hundred ng of rhTNF-a injected at O hr did not significantly affect the Lymph flow
or ceilularity of cervical lymph (n=3). Ratios were determined by dividing al i values by
the average value obtained during the 6 hr baseline.
A - cervical lymph flow
B - cervical lymph ceii concentration

A Cervical Iymph flow
Time (hr)
B Cervical lymph cell concentration
4 - 2 1 3 5 7 9 1 1 1 3 2 0 2 4
Time (hr)

6.4.5 The effect of intracerebroventncular injection of TNF-a on ceMcal lymph
In 3 sheep, the cervical Lymph was sampled after the injection of TNF-a into CSF.
The percentage of CD4, CD8, y6 T cek, B cells, CD45 and L selectin positive
lymphocytes were monitored. There was no significant change in any of the subsets (not
shown), nor in Lymph flow or ceii output (Figure 19). Lymph flow did decrease at
approximately 2 hours afier the injection but this may be due to the animai k i n g in a
recumbent position due to increased sleep as been previously demonstrated by Dickstein
et al, (1999).
6.5 Discussion
TNF-cf is present in a number of inflammatory diseases in the CNS includuig
menïngitis and MS. Previous reports have demonstrated that an infusion of =-a into
CSF causes a leukocytosû (Paris et al., 1995; RamiIo et al., 1990). Many of these studies
were performed under anaesthesia (Saukkonen et al., 1990) and/or the experhents were
limited to 24 hours or less (Quagliareilo et al., 1991). As mice and rats were used in
previous studies, immunopheno typing of recruited lymphocytes was not possible (Tang et
al., 1996). Therefore, experiments were perfonned to determine which pool of
lymphocytes responds to TNF-a and the subsets recruited. Additionally, the ciifference
between parenchymal and ventricular injections of TNF-a was investigated with respect
to Lymphocytes recruited and CSF ceilularity.

The infusion of 500 ng of rhTNF-a caused a 1000 fold iocrease in the number of
CSF leukocytes at both 24 and 48 hours. M o u s snidies dexnonstrated a peak in
leukocyte numbers within 12 hr after the infusion of TNF-a Cparis et al-, 1995; RamiIo et
al., 1990) with a decrease by 24 hr. In contrast, we found a sustained CSF ceudarity,
which lasted unti148 hr postinntsion (Figure 14). There are several explmations for this
discrepancy. Firstly, other midies used rabbits, rats and mice, whilst we used sheep. AS
w e k human recombinant TNF-a was used in our experiments instead of species specific
cytokine. Prior studies have demonstrated that human TNF-a recruits Lymphocytes into
skin sites in sheep (Kalaaji et al., 1989) and induces lymph node shutdown (Young et ai.
2000). Therefore, this cytokine appears to cross react in sheep and is unlikely to be the
cause of the dflerences observed. The dosage of injected TNF-a varied widely between
different studies. We used a bolus injection of 500 ng of TNF-a, whilst QuagliareUo et
al. (1992) used up to 20 pg in a rats and Angstwurm (1998) used injections ranging fiom
5 pg to 280 pg in rats. Similar results to our study were obtained by others using a
continuous infusion of TNF-a over 48 hrs (Schoning et aL, 1999). There is evidence to
suggest that TNF-a acts as an anti-inflammatory cytokine during EAE (Liu et al., 1998).
Therefore, it is conceivable that the larger doses of TNF-a resulted in a decreased
recruitment of leukocytes into CSF.
A study to examine the effiects of various TNF-a doses on leukocyte migration in
sheep was not perforrned. Five hundred ng of TNF-a was injected in di experiments
based on previous work by Dickstein (1999), which demonstrated that this dose of
cytokine induced sleep in sheep. Additionally, this dose was chosen to mimic levels
found in CNS infiammatory conditions. Udortunately, there was no method to measure

the levels of CSF TM-a afier injectioa Theceforefore, we can not ascertain if the levek
were comparable to those found in CNS inflammatory conditions.
Few snidies have examined the type of Ieukocytes present in CSF afier TNF-a
infusion. Tang et al. (1996) demonstrated an increase in neutrophils in response to TNF-
a - L p injections at eariy tirnepoints (2-6 hr) and mownuclear celis at 8 hr. In sheep
neutrophils and monocytes predominate in the h s t 24 hrs after TNF-a. By 48 hrs, these
Ieukocytes were a minor population with lymphocytes predominating amongst recmited
ceiis (Table 8). CD4 cells are the most nunmerous subset at 24 hrs but CD8 lymphocytes
predominate by 48 hrs (Table 9). A previous in vitro snidy suggested that CD8 ce& do
not migrate across brain e n d o t h e h as quickiy as the CD4 subset of Lymphocytes (Pryce
et al., 1994). These in vivo data reported here confirm this, as CW ceils predominate
early but are surpassed by CD8 Lymphocytes at 48 hr.
Lymph Lymphocytes are present in CSF in a greater percentage under normal
ckumstances therefore; 1 hypothesised that this pool would aiso migrate in response to
TNF-a in a greater percentage as compared to BL. However, cornparing the percentage
injected of labelied BL to labelled LL, 2.7 x 10-~ & 1.3 vs. 8.0 x 1 0 ~ t 3 (Figure 15), it îs
clear that BL migrate in greater percentage. The merence between BL and LL was not
statistically significant, however the increased percentage of BL in the infiamed CSF may
have biological signifïcance. As well, FïïC Iabelled BL are present in a higher
percentage in CSF as compared to lymph (0.61 f 0.14 vs. 0.34 10.3) (Table 10). This
indicates that BL were not non-specificaiiy migrating into aii tissues but were selectively
recruited into the ïnflamed CSF. This could be accomplished in one of two ways.
Firstly, there may be a pool of rapidly recirculating lymphocytes recruited into the CSF as

in the antigen stimulated Lymph node (Chapter 4). An alternative explanation is that the
idamed CSF preferentially attracts BL. Perhaps blood lymphocytes have higher levels
of adhesion molecules or chemokiw receptors ailowing them to enter the CSF- This
should be tested in funire experiments.
Due to smaU numbers of labelled ceh, immunophenotyphg of the recruited FJTC
and DiI-DS Lymphocytes was not performed. This would have allowed the direct
cornparison between the antigen stimulated lymph node experiments and these
experiments.
Intracerbrovenuicular TNF-a injections resulted in a widespread infiammatory
infiltrate within the perivascular spaces of the brain. The recmited lymphocytes did not
appear to migrate into the parenchyma of the CNS. This may be due to lymphocytes
undergo ing apo ptosis outside of the perivascular space as previously reported (Bauer et
al., 1998). Imrnunohistochemistry demonstrated that the perivascular infiitrate was
similar to that seen in the CSF with respect to lymphocyte subsets, that is
CD8>CD4>ybB cells. There were rare CD2k lymphocytes in the infiltrate indicating
that the majority of celis were not fully activated. This is similar to other studies that
investïgated MS (Hofinan et aL, 1986), EAE (Korner et aL, 1997) and toxopiasmosis
infection of the brain (Deckert-Schluter et al., 1994). If few activated lymphocytes are
present in these autoimmune and parasitic diseases it wodd be expected that few
lymphocytes would be activated after a cytokine injection. MKC II is present in small
amounts in nomial control brauis but is upregulated after the intracerebroventnfular
injection of TNF-a, simiiar to results obtained in rats after IFN-y injections (Vass and
Lassrnaan, 1990). The infT1trate included MHC II positive cells, which may be

macrophages, either recruited fkom blood or activated CNS macrophages. The
endotheIium of blood vesseis also appeared to be positive for MAC II as previously
descnibed in visna Wus infected sheep (Torsteiosdottir et al., L992).
Choroid plexus samples were taken to determine if Lymphocytes migrate dicectiy
into the CSF fiom blood as suggested by others (Engeihardt, 1997). The data obtained
was not dennitive but there appeared to be an increase in lymphocytes within the choroid
plexus and VCAM was expressed on choroid plexus bbod vessels. These data m e r
kom previous reports tbat demonstrated an increase of VCAM on the epitheiial c e h of
the choroid plexus but not on the endothelia1 cells (Wolburg et al, 1999; Steffen et al,
1996). This maybe due to diffierences in experimental protocois (EAE vs. TNF-a
injections) and animai species (mice vs. sheep). Further experiments are required to
determine if choroid plexus can support leukocyte extravasation into the CSF.
Afier an injection of 500 ng ofTNF-a into the cortex of the braïn, no h e a s e in
CSF cellularity was seen. There was Little inaammatory ianltrate at the site of injection
as compared to saline injected controls. This is in contrast to other reports (WdIenborg et
al., 1993; Wright and Merchant, 1992), which demonstrated that TNF-a injections
resulted in leukocyte innltratioe The discrepancy maybe due to a species ciifference or
arnounts of TNF-a injected. The dose of TNF-a used in the study by Wright and
Merchant was 5 tirnes the dose used in the present study. Schnell and colleagues (1999)
demonstrated that TNF-a injected into the spioal cord of rats resulted in an inflammatory
infiltrate, but the brain parenchyma was resistant. WiUenborg et al. (1993) used the
spinal cord and Wright and Merchant (1992) the parietal lobe, while the present snidy
injected TNF-a into the cortex overlying the lateral ventricles.

TNF-a has been shown to into the rettopharyngeal Lymph nodes but the
function of this is stili unknown (Dickstein et aL, 1999). Its injection into CSF did not
have a discernible effect on cemecai Lpph with respect to Lymphocyte subset, ffow or
cellularity. Perhaps the parameters exûmuied in this series of experiments were not able
to detect subtle changes that may be present. TNF-a causes lymph node shu tdom
which may have a role in the immune response to some Ag (Young et al., 2000). It is
conceivable that TNF-a draining lÏom the CSF may increase the immune response to Ag
arrïving fiom the CSF (Harhg-Berg et al, 1989). The drainage of proinfiamatory
cytokines into retropharyngeal lymph nodes may assist in the activation of T c e b
specifc for antigens sequestered in the CNS. This is speculation at this point and fuaher
studies are required.
In conclusion, the data in this chapter fuaher demonstrates that a pool of
lymphocytes present in blood may be able to rapidly enter sites of uinarnmation.
Intracerebroventricular TNF-a injections resulted in a rapid increase in ieukocytes b ~ t h
in CSF and brain parench- Simi1a.r injections into the cortex did not increase
ieukocyte numbers in either cornpartment.

Chapter 7 General Discussion
7.1 Introduction
The recirculation of lymphocytes aliows for the dissemination of immuno1ogica.l
memory and immune surveillance (Butcher et ai., 1999). Recently, in sheep, a pool of
blood lymp hocpes has been descnibed whic h recirculates poorly through lymp hatic tissue
(Andrade et al., 1998; Chevallier et al., 1998; Young et ai., 1997a). The main objective
of this thesis was to investigate the migration of the blood pool of Lymphocytes in
cornparison to the lymph pool of lymphocytes in a variety of compartments under n o n d
and hfhnmatory conditions. Experiments were conducted to compare the migration of
LL and BL into CSF under both normal and TNF-a induced inflammation- As well,
experiments were performed to determine the migration of the two pools after
splenectomy and through an antigen stimulated lymph node. Together these data extend
earlier studies and show that a functional dinrerence may exist between BL and LL.
Sheep were used in aii experiments in this thesis, as they permit chronic
collections of lyrnph. Additionaiiy, a large body of Iiterature exists on lymphocyte
migration in this animal, allowing a critical cornparison of my data to others. Indeed it
has been said that more is kno wn about the in vivo physio logicai dynarnics of lymphocyte
recirculation in sheep than in any other animal (Abemethy and Hay, 1992)
In this discussionT the main conclusion fkom each major study will be re-
examined, the limitations of each wiU be considered and funher experiments suggested-
At the end of ehis chapter, a series of experiments to investigate the existence of a rapidly
recirculating pool of lymphocytes is proposed.

7.2 Investigations into the migration pattern of BL and LL into afferent
lymph and after splenectomy
The main objectives of this senes of experiments was to extend our basic
knowledge of BL migration. The h t report of BL appeared in 1994 (Young, 1994) but
Little was kaow about these Lymphocytes beyond thek retention in blood (Andrade et al.,
1998) and that B ce& compose the largest population of BL (Gupta et aL, 1998; Young
et al., 1997a).
Repetitive sequential sarnpling of blood and lymph was performed to gain insight
into the stability of the relative proportion of BL and LL. The migration and cornparison
of LL and BL into normal afferent lymph was determined adding an additional
cornpartmentai analysis. Since there are significant ciifferences in the cellular
composition of blood and aEerent lymph (Haig et al. 1999)- it was thought that the two
pools may differ in their migration. Additionally, prior studies had demonstrated changes
in lymphocyte subpopulations in blood d e r splenectomy (Sieber et ai. 1985) and it was
hypothesised that this maybe due to changes in the migration or number of BL-
7.2.1 Repetitive sampling
The relative proportions of the labeiied BL and LL remained constant in both
blood and efferent Lymph over the 27 hour sampling period (Figure 4).
A Iunitation of the repetitive sampiing experiments was the method used to
coilect sarnples as the animal was disturbed hourly during the expriment. This =y
result in stress due to alterations in the normal sleep pattern of the sheep, which

affect lymphocyte migratioa A remote sampling technique to which the sheep was
oblivious, as used by Dickstein (1999), wodd have avoided this problem Nonetheless,
these experiments confkmed previoas fïndings that BL and U are enriched in blood and
Lymph, respectively (Andrade et aL, 1998; Young et al., 1997a). Based on these hdings
the tirne of day of sampling was not considered a confounding factor ùi subsequent
experiments.
LL IabelIed with the fluorescent compouod, DiI-DS, recirculated fiom blood into
efferent lymph in percentages comparable to c e k labelied with other Lipophilic dyes
(Andrade et al., 1996a). This was considered a validation for the subsequent use of this
tracking label in sheep.
7.2.2 Afferent lymph
In afferent lymph, a ratio was determined by dividing the percentage of labelied
lymphocytes in afferent lymph by the percentage in blood Lymph lymphocytes are
enriched in aeren t lymph as compared to BL (3.1 + .96 vs. 0.8 & 0.26) (Table 2) The
appearance of labeiled cells in aerent lymph implies that they have migrated fiom blood
through tissues. Therefore, the population of lymphocytes in afferent lymph may reflect
that present in tissues.
A limitation of this study is that these samples were acquired at varyiag times
after the iofusion of labelled ceils. However, ail samples were obtained at least 3 days
postiofusion at which t h e previous studies (Andrade et al. 1998) have demonstrated that
the BL and LL have equihibrated between blood and efferent Lymph. Experiments to
examine the kiwtics of migration and standardised recoveries are required to compare the

two pools in afferent Lymph. As weli, the appearance of labeUed ceils in afferent Lymph
implies that lymphocytes have migrated into tissues but tbir is wt a direct measure-
Further experiments are required to detennine if BL migrating into tissues become
resident ceils or are deleted in greater numbers as compared to U.
7.2.3 Splenectomized sheep
In splenectornized sheep, no differences were detected in the relative proportions
of the analysed lymphocyte mbsets (Table 4)- Furthemore, the concentration of
lymphocytes was not different ikom intact sheep. The migration experiments
demonstrated that LL exit blood quicker as compared to intact sheep and enter lymph
nodes and liver in significantly greater numbers. This is demonstrated by the trend
towards fluorescently labelied LL to exit the blood quicker in splenectomized sheep
(Figure 5) and by a greater number of radioisotope labeiled celis localising in lymph
nodes and liver (Table 3). Blood lymphocytes demonstrated a trend to an increase in
blood and are found in Lesser numbers in the lung of splenectomized sheep. This may
indicate that the marginating pool in the lung is saturated and excluding BL fiom entering
in the normal m e r , thereby enriching this population in blood.
However, these experiments should be repeated in sheep that undergo
splenectomy at a later age. Previous reports have demonstrated that there is extensive
replacement and/or expansion of foetal T celis during the fkst week of life (Cahill et aL,
1997). The sheep in these experiments were splenectomized between 19 and 21 days
after birth, which may allow the newly forrned lymphocytes to adapt to the absence of the
spleen. Therefore, the data fiom these studies may not refiect the changes in migration of

BL and LL that result after adult splenectomy. As well, fkther snidies are required to
investigate B ceii function as there are abnofmalities in humorai immunïty in humans
folio wing spIenectorny.
Conclusion
The cepetitive sampling experiments demonstrated that BL and LL are maintained
at steady levels after equiii'brium and that the time of day did not &ect their
concentrations,
There are no other reports comparing BL and LL in aEerent lymph. Lymph
lymphocytes. which belong to the weii-characterised RLP (Young, 1999), are found in a
greater ratio in afferent lymph as compared to BL. These results indicate that LL migrate
through tissues in greater numbers than BL and therefore may have a greater role in
immune surveilIance.
The BL pool is present in normal numbers 2 years after neonatal splenectomy
ïmplying that the spleen is not necessary for its maintenance. DBerences are seen in the
migration of bo th BL and LL in splenectomized sheep, with LL migrating into peripheral
lymph nodes and the liver in greater numbers than in intact sheep. These data indicate
that after splenectomy LL may exit the blood in greater numbers to maintain lymphocyte
homeostasis,
These data extend previous reports regarding the migration of BL and provide a
foundation for subsequent experiments-

7.3 The migration of BL and LL through antigen stimulated lymph nodes
These were the first experiments to examine the m-gration of BL and LL
lymphocytes during ï p p h wde shutdown. They were part of a larger series examining
lymphocyte retention during lymph node shut down but much of that data was wt
included in this thesis as it was performed by collaborators (manuscript in preparation).
In agreement with previous studies (Mackay et al, 1992b; Bujdoso et al., 1989) CD4 T
cells increased over baseline in their migration through inflamed lymph nodes (Figure 8).
An increase in IFN-y and IL-6 in efferent Lymph during Lymph node shutdown was
observed (Figure 9) and may have a role in the recniitment and retention of Lymphocytes.
IFN-y can recruit lymphocytes when injected into the skin (Colditz and Watson, 1992)
and has been shown to retain naive lymphocytes withui the lymphatic tissue of rats
(Westermann et al., 1994b). IL-6 induces the production of chemokines in skin
infiammatory conditions (Romano et al., 1997) and may have a similar role in the lymph
node.
An increase in both CFSE (blood) and Di[-DS (lymph) iabelIed CD4 cells was
observed (Figure 8), indicating that both BL and LL contributed to the increase in CD4
cells. However, BL CD4 ceils increased to a greater extent than LL. One explanation is
that memory T cells are increasing as previously shown durhg antigen induced lymph
node shutdown (Mackay et ai., 1992b). However, this does not account for the greater
hcrease amongst CD4 BL unless blood memory CD4 cells preferentidy migrate into
inflamed lymph nodes. An alternative explmation is the existence of a rapidly
rec irculating poo 1 of lymphocytes. This hypothesis is M e r explored and experiments
proposed in section 7.5.

Previous experiinents have demonstrated that labelbg and infirsing a whole
population of lymphocytes is an effective method to study subset migration in rats
(Westernmm et al., 1993) and sheep (Andrade et ai., 1998). This avoids the separation of
subsets using various in vitro methods such as anh'body/compiement, magnetic bead
separation etc- Our methodology minimises Ln vitro rnaIumaIupulation of cells thereby
decreasing my chance of inadvertent activation and allows ail subsets to be examined
after in vivo migration. Using these techniques we observed several subsets present Ï n
BL and demonstrate that CD4 ceiis are not retained within the l p p h node to the same
extent as other measured subsets.
A Limitation of the present studies is that only one antigen, PPD, has been
examined for its effect on LL and BL migration. Other antigens including vinises and
bacteria should be investigated to ensure that this phenomenon is not unique to PPD-
Additionaiiy, the direct effect of cytokines including IFN-y and IL-6 on the retention of
lymphocytes within a single lymph node should be examined Previous reports have
demonstrated that both EN-a (Hein and Supersaxo, 1988; Kalaaji et aL, 1988)and N-
a (Young et al., 2000) induce lymph node shutdown.
Conclusion
These experiments were undertaken to examine BL and LL under infiammatory
conditions. Both pools appeared to contriiute to the increase in CD4 celIs during lymph
node shutdown but BL CD4 ceus increased to a greater extent. As weil, these data
suggest the existence of a rapidy recirculating pool of lymphocytes.

7.4 Lymphocytes in CSF are part of the RLP
CSF is the fîrst noniymphoid tissue to k examined for the differentiai migration
of BL and LL. Using FïïC Iabeiied efferent LL it was found that labelled cens appeared
in similar concentrations and times in both CSF and efferent Lymph (Table 6 & Figure
12). This impLies that CSF lymphocytes belong to LL or the RLP-
The data fiom this series of experiments argue that activation is not required for
lymphocytes to enter n o d CSF as previously reported (Vrethem et aL, 1998; ScoIozzi
et al., 1992; Mïx et al, L990). Less than 5% of efferent LL express the activation marker
CD25 (IL-2 receptor) (Haig et al., 1999). Therefore, ifonly activated T ceils migrate into
CSF it would be expected that the number of labeiied ce& wouId be Iow. Svenningsson
et al. (1995) argues that activation is not required for lymphocyte migration into CSF
based on a series of phenotypic markers on Lymphocytes obtained £tom normal human
CSF. They did not find any upregulation of CD25 or HLA-DR, both markers indicating
lymphocyte activation. Reasons for the discrepancy between snidies maybe due to
staining protocols or subject selection as often patients with tension headaches or other
neurologicai abnormalities are used as normal controls. A more likely explanation is that
the definition for activated T c e k is different in a i l studies- For example, Vrethem et al-
(1998) use the expression of CD45RO as the sole indicator of activation, whilst
Svenningson et al. use CD25+ and HLA-DR to determine activation of T cells. Based on
these ciifferhg dennitions of activation it is not surpriskg that fidings confiict. The data
in this thesis irnply that Lymphocytes are not required to be activated to migrate into the
CSF.

Studies were undertaken to examine the migration of LL and BL into CSF. AS
measured by percentage of injected cek , LL are present in greater numbers in normal
CSF as compared to BL, confinning that CSF lymphocytes are part of the RLP (Figure
15)- These results are similar to that found in affierent lymph (Chapter 3), implying that
LL have a greater role in immune surveillance of tissues as compared to BL.
A limitation to these studies is that due to problems m a i n t d g a patent catheter
in the subdural space, multiple samples of CSF over a 24 br period were not obtaiwd to
determine kinetic data for BL. As weil, fluorescently conjugated primary antibdies were
not avaïlable to perform immunophenotyping of Lymphocytes present in normal CSF- If
activation markers were examined it may ~ ~ p p o a the fbdings that nonactivated T c e k
migrate into CSF-
Conclusion
These data support previous reports that lymphocytes do not need to be activated
to migrate into the CSF- Lymph lymphocytes migrate into the CSF in greater numbers as
compared to BL. These data in conjunction with data from the afferent lymph
experiments, imply that LL have a greater role in immune surveiüance of tissues.
7.4 TNFa induced CSF leukocytosis
Several other studies have examined the proidlamatory effects of
intracerebroventriculat injections of TNF-a but in most of these studies, cytokine was
injected in amounts several times higher than the 500 ng injected in the present snidy.
For example, Angstwurm et al. (1998) injected up to 280 pg of TNF-a in adult rats, while

QuagliareIlo et aL (1991) used 20 ug. Both of these injected amounts are thousands of
fold higher than levels of TNF-a measured during CNS inflainmation. Glimaker e t ai.
(1993) reported levels of 1.16 n g h i of CSF during purulent meningitis, whilst in MS
values of 10-1 pg/ml of CSF are found (Druuiovic et ai. 1997). Previous work In our
laboratory demonstrated that an intracerebroventricular injection of 500 ng of rhTNFa
induced sleep in sheep (Dickstein et al., 1999) and produced infiammation in CSF.
Therefore, th% amount was used as it was determlned to cause inflammation in
preliminary experiments and had a physiological effect in vivo. As welI, it may be
similar to TNF-a levels measured during meningitis.
The concentration of leukocytes in CSF after the infusion of TNF-a was similar
to those reported in bacterial rneningitis 5.75 x ld celldml vs. 3.40 x 1o3 cellslml
(Ghaker et al., 1993). The phenotypic subset of recnUted lymphocytes changed over
the 48 hr observation period, with CD4 ceiis king the largest subpopulation at 24 hr but
by 48 hr, CD8 cells predominated (Table 9). These rii vivo data confirm a previous in
vitro report that CD4 ceiis migrate across cultured cerebral endothelhm more rapidly
than CD8 lymphocytes (Pryce et al., 1994)-
An intracerebroventricular infision of TNF-or was used to induce inflammation in
the CSF. This resulted in a 1000 fold increase in both BL and LL into the inflamed CSF
as compared to normal (Figure 15). However, BL were present in 4 times the
concentration as LL. At les t two explanatioas for the increase in FITC labeiled blood
c e k in the infiamed CSF are possible. 1) a rapidly recirculating pool of T cells is present
that enters sites of inflammation 2) blood CD8 cells are better able to migrate into CSF as

compared to other lymphocytes. Neither of these expianations can be discounted nom
these data. E x p e k n t s are proposed in section 7.5 to test between these alternatives.
CSF inflammation was induced with TNF-a in these studies but to ensure that
these fhdings are not unique to this cytokine fuaher experiments are required-
Intracerebroventricular injections of other cytokines should be examllied for their ability
to attract BL and LL hto CSF, As welI, viruses and bacteria can be used to mimic
meningitis to determine if these conditions cause the preferential migration of BL-
Conclusion
TNF-a induced inflammation in the CSF results in FITC labelied BL migrahg in
a greater percentage as coqared to Da-DS labelled LL. The FLTC Iabelied cells may be
part of a rapidly recirculating pool or a BL subset may be preferentially recnùted to the
inflamed CSF. Further experiments are required to determine which is correct.
7.5 Future experiments to determine the existence of a rapidly recirculating
pool of lymphocytes
Findings fio m the experiments examining Lymphocyte migration into the inflamed
CSF and during lymph node shutdown may be due to either of two explanations; A)
increased migration of memory T ceils. B) a pool of Lymphocytes may exist which
rapidly recirculates as compared to the majority of the RLP.
The increased CD4 and CD8 cells, in lyrnph and CSF respectively, d-g
inflammation may be memory T cells. They have been demonstrated to have different
migration kinetics through antigen stimulated lymph nodes (Mackay et al., 1992b) and

after the infusion of IFN-y pestermiann et &, 1994~). The increase in BL may indicate *
that blood memory T c e k respond to inflammation and migrate into antigen stimuiated
l p p h nodes or uiflamed CSF in greater numbers than LL memory T cek.
An alternative hypothesis is that a population of Lymphocytes exists which rapidly
recirculates and is found in transit through blood and lymph. As this population may be
present in both blood and lymph, the labelling protocols Ï n this thesis would label it with
either fluorescent dye. This population of c e k may be functionaily different fiom both
BL and LL and/or may express different adhesion moIecdes or chemokuie receptors.
There are data in the Literature to both support and refute this hypothesis of a population
of lymphocytes that rapidly recirculates.
In sheep, 11 1-In labelied lymphocytes are detected in efferent and afferent Lymph
within 5 hours of intravenous injection (Issekutz et al. 1981). As weil, in rats
Westennann et al. (1994b) demonstrated that FITC Iabeiied Lymphocytes are found in
thoracic duct lymph within 12 hours after intravenous injection. Both the sheep and rat
studies detect labelied lymphocytes in lymph preceding the t h e of maximum levels of
detected ceiis, 2 1 - 23 and 24 - 36 hours respectively. Fuahermore, there is a population
of lymphocytes in rats, that is rapidiy mobilised after thoracic duct lymphatic cannulation
(Westermann et al. 1994b). These studies hply that a population of rapidly recirculating
lymphocytes is present within the RLP,
However, Ford and Simmoads (1972) reported that in rats RLP lymphocytes
could not be aivded into "fast" and "slow" populations. This was based on a series of
experiments in which labelied lymphocytes were obtained either 12 or 36 hours &es
thoracic duct cannulatioa The two populations were then differentially labelied and

injected into syngeneic recipients. No merence in the kinetic appeacance of the two
popdations in thoracic duct lymph of the recipients was found. However, the rats
underwent a splenectomy less than 24 hours before the experiment was begun.
Splenectomy may affect the normal migration patterns of lymphocytes in adult subjects
by removing this important lymphoid orgaa Additionaüy, the shidy was performed
before the use of monocIonal antiidies to detect lymphocyte phenotypes- Westermann et
aL (1993) using monoclonai antiidies reponed rbat CDS lymphocytes recirculated with
slower kinetics than CD4 celfs. Therefore, there is data to suggest that the RLP is
composed of several populations of lymphocytes with varying migration khetics and
may include a rapidiy recirculating population of ce&.
Experiments to determine the existence of thû putative recirculating population
with rapid migratory properties are as foiiows;
1) Label blood and lymph ceils with fluorescent dyes and intravenously reinfuse.
Coilect efferent lymph hourly and phenotype the c e k for adhesion molecules includuig L
selectin, CD44 and a4 integrin. Phenotyping for activation and memory markers such as
CD45R.A would demonstrate if the labelled ceffs that first appear in lymph were memory
ceiis or activated. I hypothesise that the c e h recovered in lymp h on the ascending part of
a recovery curve may have higher levels of adhesion molecules than those recovered
during the descendhg phase.
2) It remains to be determined if the labeiied BL seen in efferent lymph d m g
Lymph node shutdown and those in CSF after =-a induced inflammation both belong
to a rapidly recirculating pool. 1 propose the following two experimental protocols to

examine this question. The h t series of experiments wouid require inaadermal
injections TNF-a and PPD in BCG primed sheep. Both have been used in separate
experiments in this thesis but have not been dïrectly compared to determine if they resdt
in the preferentiai recruïtment of BL. Prior midies by Waaji and coiieagues (1989)
demonstrated that both PPD and TNF-a recruit LL into the skin of BCG primed sheep.
Therefore, this protocol wiii aliow the direct comparison of the ability of TNF-a and PPD
to recruït BL into slon. Blood and lymph Lymphocytes can be labelled with different
radioisotopes, such as 11 1-In and 5 1-Cr, and infüsed intravenously. The sites of injection
cari be removed and the anmount of radioactivity determined. These results WU indicate if
the BL is preferentiaily recded into the skui by both PPD and TNF-a.
A second method w u examine the detailed kinetics of the two Iabeiied pools in
the aerent lymph drainlog a delayed type hypersensitivity site. ABerent lymphatics will
be canulated, blood and efferent lymph cells fluorescentiy IabeUed and reinhised. PPD
injections into the drainage area of the &erent lymphatic c m then be performed and the
Lymph monitored for labelled ceils- The colIected afferent lymph lymphocytes can then
be immunophenotyped for several adhesion molecules and activation markers. Thû
would al10 w the direct comparison of BL and LL labelled cells, which was not possible in
the CSF inaarnmation experiments.
The proposed experiments might document a rapidly recirculating population of
lymphocytes, if the hypothesis is correct. Alternatively, if the increase is due to the
increased tr&c of memory T celis these proposed experiments would also determine if
this is correct,

7.6 Summary
Tbe experiments in this thesis were designed to investigate the migration of the
LL and BL under normal and infiammatory conditions. It appears that LL may have a
greater role in immune su~veiliance as compared to BL. TNF-a induces inflammation in
sheep CSF with an infiammatory infiltrate that changes in composition over time. The
experiments in this thesis provide the first in vivo experimental evidence that BL and LL
may have Heren t functions under both normal and infiammatory conditions. Further
experiments are required to determine if these differences c m be -pulated to gain
m e r insight into their roles in immune system.

Reference List
Abernethy, N.I. and Hay, J.B- (1992) The recircuiation of lymphocytes fiom blood to lymphr physiological considerations and molecular mechanrsms. Lymphology, 25: 1-30.
Abitorabi, M.A., Mackay, CR, Jerome, E H , Osorio,O., Butcher,E.C., and Erle,D.J. (1996) Differentiai expression of ho- molecules on recirculating lymphocytes fÏom sheep gut, peripheral, and lung lymph- J.ïmmunoL 156:3 1 1 1-3 1 17.
ALlport, J.R., Ding,H., Collins,T., Gerritsen,ME., and Luscinskas,F.W. (1997) EndotheLial-dependent mechanism regulate leukocyte transmigration: a process invo Lving the proteasorne and disrup tioa of the vascular endotheliai-cadhe~ compiex at endothelial celi-to-ceii junctions. JExpMed., 186:s 17-527.
Andersson, PB., Perry,V.H., and GordosS. (1992) Intracerebral injection of proidammatory cytokines or leukoc yte chemtaxins induces mlliimal myelomonocytic ceil recruitment to the parenchyma of the central nervous system. I.Exp.Med., 176:255- 259.
Andrade, W. The relationship of blood lymphocytes to the recirculating lymphocyte pool- 1996. University of Toronto. 1996. PhD Thesis
Andrade,W., Seabrook,T.J., Johnston,M.G., and HayJB. (1996a) The use of the Lipophilic fluorochrome CM-DiI for tracking the migration of Lymphocytes. J.Tmmunol.Methods, 194: 18 1-189.
Andrade, W.N., Johnston,M.G., and Hay, J.B. (1996b) A method for t racbg the migration of blood lymphocytes. Iinmunolhvest, 25:455-467.
Andrade, W.N., JO hnston,M.G., and Hay, J.B. (1998) The relationship of blood lymphocytes to the recirculating Lymphocyte pool. Blood, 9 1: 1653- 166 1.
Angstwurm,K., Freyer,D., Dirnagl,U., Hanisch,U.K, Schumann,R.R.. EinhaupLKM., and Weber, J-R. (1998) Tumour necrosis factor alpha induces only minor idammatory changes in the centrai nervous system, but augments experimental meningitis. Neuroscience, 86:627-634.
Ant hony,D .C., Miller,K.M., Feam,S . , To wnsend,M. J., Opdenakker,G., Wells,G.M., CIements,J.M., Chandler,S., Gearing,A.L, and Perry,V.H. (1998) Ma& rneta.Uoproteinase expression in an experimentally-induced DTH mode1 of multiple sclerosis in the rat CNS . JNeuroimmuno l., 87 :62-72.
Arbones,M.L., Ord,D.C., Ley,K, RatechB., Maynard-Curry,C., Otten,G., CapoaD. J., and Tedder,T.F. (1994) Lymphocyte homing and leukocyte rolling and migration are impaired in L- selectin-deficient mice- Immunity., 1:247-260.

AustnipS., Vestweber,D., Borges,E., Lohning,M-, Brauer&., Herz,U., ReozB, Hallmann,R., Scheffold,A, Radbruch,&, and Kamann,k (1997) P- and E-selectin mediate recruitment of T-helper- 1 but not T-helper-2 cells into idamed tissues. Nature, 385:8 1-83,
Bamborschke,S., WulieaT., and Beil,C. (1990) Quantitation of lymphocyte subsets in cerebrospinal fluid and blood during the clinical course of aseptic and bacterial meningitis- Eur.Neurol., 30% 1-295-
Bargatze,RE and Butcher,E.C. (1993) Rapid G protein-regdated activation event invo lved in lymphocyte binding to high endothelid vendes. JExpMed-, 178:367-372-
B auer, J., Brandi,M., Hickey,W-F., Forss-Petter,S ., BreitschopfJI., LiningtonC, Werkele,H., and Lassman,H. (1998) Tcell apoptosis in inflarnmatory brain lesions. AnzJ.Pathol., 153:715-724.
Berh,C., Bargatze,R.F., Campbeil, J.L, von Andrian,UH., Szabo,M-CC, Hasslen,S P., Nelson,R.D., Berg,E-L., Erlandsen,S L., and Butcher,E.C. (1995) alpha 4 integrins mediate lymphocyte attachment and r o h g under physiologie flow. Ce& 80r413-422.
Blunt,M.H. (1975) Cellular elements of ovine blood. In: The bloodof sheep. M.H. Blunt, ed. Springer-Vrlag, New York
Borron,P. (1991) The role of lymphatic endothelium in the lymph nodes' response to a delayed hypersensitivity stimulus. MSc Thesis University of Toronto
B oulton,M-, Ftessner,M., Armstrong,D., Hay,l., and JohnstonM- (1 997) Lymphatic drainage of the CNS: effects of Lyrnphatic diversionlligation on CSF protein transport to plasma. Am.J Physiol, 272:R16 13-R1619.
Bujdoso,R., Young,P., HopkuisJ., AUen,D., and McConneilJ- (1989) Non-raadom migration of CD4 and CD8 T ceus: changes in the CD4: CD8 ratio and interleukin 2 responsiveness of efferent lymph cells foilowing in vivo antigen challenge. Eur.J.ImmunoL, 19: 1779- 1784.
Bulte,J.W., Hoekstra,Y., Kammaa,R.L., Magin,R.L., Webb,A.G-, Bnggs,R.W-, Go,KG-, Hulstaert,C.E., Miltenyi,S., and The J.H. (1992) Specific MR imaging of human lymphocytes by monoclonal antibody-guided dextran-mgnetite particles. Magn Resoa-Med., 25: 148-157.
Butcher,E.C. and Picker,L. J. (1996) Lymphocyte homuig and homeostasis. Science, 272:60-66,
Butcher,E.C., Scoilay,RG., and Weissman,I.L. ( 1980) Direct fluorescent iabeling of celIs with fluorescein or rhodamine isothiocyanate. II. Potential application to studies of Lymphocyte migration and maturation. J.ImmunoI.Methods, 37: 109-121.

CahiU,R-, Hay,H3., Frost,H., and Tmka,Z- (1974) Changes in Lymphocyte circulation afier administration of antigen. Haematologia (Budap-). 8% 1-334.
CahiLl,RN., Kirnpton.W.G., Washington,E.A., DudlerJ.,., and Tdca,Z- (1997) An immune system switch in T cell Lifespan at birth results in extensive Ioss of naive fetal T cells during the fist week of postnatal Mee Tiit.ImmunoI., 9: 1253- 1258-
CahiU,RN., Poskitt,D., Frost,H., and Trnka,Z, (1977) Two distinct pools of recirculating T lymphocytes: migratory characteristics of nodal and intestinal T lymphocytes. J-ExpMed., 145:428,
Campbell, J.J., Qin,S ., Bacon,= ., Mackay,CE., and Butcher,E.C. ( 1996) Biology of chemokine and classicai chemoattractant receptors: differential requirements for adhesion-trïggering versus chemotactic responses in Iymphoid ceils. J.Cell Biol., l34:255-266.
Cannella,B. and Raine,C.S. (1995) The adhesion molecule and cytokine profile of multiple sclerosis lesions [see comments] . AnnNeuroL, 37:424-435.
Carson,M.J., ReiiIy,C.B -, Sutcliffe,J.G., and Lo,D. ( 1999) Disproportionate recruitment of CD8+ T cells into the central nervous system by professional antigen-presenting celIs. Anl J-Pathol., 15448 1-494.
Chevallier,N., Berthelemy,M., Laine,V., Le Rhun,D., Fernenia$., PolackB., Naessens,J., Levy,D., and Schwartz-Conzil,I. (1998) B- 1-like c e h exist in sheep. Characterization of t heir p heno type and be haviour. Immuaology, 95: 178-1 84.
Chin,G.W., Pearson,L.D., and HayJB. (1985) Cens in sheep lymph and their migratory characteristics. In: Experimental Biology of the Lymphatic Circulation. M.G. Johnston, ed. Elsevier Science, Tarrytown, pp. 141-164.
Chin,W. and Hay,J.B. (1980) A comparison of lymphocyte migration through intestinal Lymph nodes, subcutaneous Lymph nodes, and chronic inflammatory sites of sheep. Gastroenterology, 79: 123 1-1242.
Co1ditzJ.G. and Watson,D.L. (1992) The effect of cytokiues and chemotactic agonists on the migration of T lymphocytes hto skin. Immunology, 76:272-278.
Cro ss,A.H., CanneUa,B ., Brosnan,C.F., and Raine,C-S - (199 1) Hypo thesis: Antigen- speci6c T cells prime centrai nervous system endothelium for recruitment of nonspecinc inflammatory cells to effect autoimmune demyebation. J.Neuroimmunol., 3 3:237-244.
Cserr,H.F. and Knop f,P.M. (1992) Cervical lymphatics. the blood-brain barrier and the immunoreactivity of the brain: a new view. Immunol.Today, 13507-5 12.

DavenpeckKL., Chrest,F.J., Sterbinsky,S.k, BickelC.k, aad Bochner,B.S. (1995) Carboxyfluorescein diacetate labelhg does not afEect adhesion mlecule expression or function in human neutrophils or eosinophils. J ~ u n o L M e t h o d s , 188:79-89.
Deckert-SchluterM., Schluter,D., HOCH., Wiestler,OD., and LassmanSI, (1994) Differential expression of ICAM- L, VCAM-1 and their Iigands LFA-L, MAC-1, CD43, VLA-4 and MHC class II antigens in murine toxoplasma encepbalitis: a Light microscopie and ultrastructural immunohistochernicai study. J~europathoLExp.NeuroL. 53:457-468.
Dickstein, J. B. The effects ofsleep and tumour necrosis factor-alpha on the lymphatic system 1999- University of Toronto. P h . Thesis
Dickstein,JB ., Moldo fsky,H-, Lue,F.A-, and Hay, JI3 . ( l999) htracerebroventrïcular injection of TNF-alpha promotes sleep and is recovered in cervical lymph. Am.J.PhysioI, 276:RlO 18-R1022,
Dittel,BN., VisintinJ., Merchant,RM-, and Janeway,C.A. (1999) Presentation of the self antigen myelin basic protein by dendntic ceiis leads to experimental autoimmune encephalomyeiitis. J. ImmunoL, 163:32-39
Druulovic,J., Mostarica-S tojkovi~~M.~ Levic9Z., S tojsavljevic,N-, Pravica,V., and Mesaros,S. (1997) Interleukia-12 and tumour necrosis factor-alpha levels in cerebrospinai fluid of multiple sclerosis patients. J.Neurol.Sci, 147: 145- 150.
DUX~R-, Kindler-Rohrbom, A., Annas,M., Faustmann,P., Lennartz,K, and Zimmermann,C.W. (1994) A standardized protocol for flow cytometrïc analysis of ceils isolated irom cerebrospinal fluid, J.Neurol.Sci., 12 1:74-78.
EngeIhardtB., Conley,F.K, and Butcher,E.C. (1994) Ceii adhesion molecules on vessels during infiammation in the mouse central nervous system. JNeuroimmunoL, 51:199-208.
Engelhardt-B. and Risau,W. (1995) Development of the blood-brain barrier. In: New concepts of a blood-brain barrier. KGreenwood, DBegley, and M.Segal, eds. Plenum Press, New York, pp. 1 1-3 1.
Engelhardt,B ., Vestweber,D., Hallmann,R., and Schulz,M. (1997a) E- and P-selectin are not involved in the recmitment of infiammatory cells across the blood-brain barrier in experimental autoimmune encephalomyelitis. Blood, 90:4459-4472.
Engelhardt,B. (1997b) Lymphocyte tranicking through the central nervous system. In: Adhesio n mo lecules and c hemokines in lymphocyte trafnc king. A.Hamann, e d Harwood Academic miblishers, Amsterdam, pp, 173-200.
Fawwaz,R.A., Oluwole,S ., Wang7T.S, Kuromoto9N., Iga,C., Hardy,M-A., and Alders0n~P.0. ( 1985) Biodistribution of radiolabeled lymphocytes. Radiology, 155:483- 486.

Ferrante,k, Drew,P.A, Kiroff?G.K., and Zola,H. (1987) Peripherial blood leukocyte subpopulations in patients splenectomized for traumauma Clin.Exp_Tmmunol., 70: 158-163-
Ford,W.J. and iuun0~ds~S.J- (1972) The tempo of lymphocye recirculation fiom blood to lymph in the rat, Ceil Tissue iCi.net, 5: 175-189
Gallatin, WM., Weissman,I.L,, and Butcher,E.C- (1983) A cell-surface moIecde involved in organ-specific homing of lymphocytes. Nature? 30430-34.
Girard, J.P., Baekkev01d~E.S ., Yamanaka,T,, Haraidsen,G., Brandtzaeg,P., and AmalricS: (1999) Heterogeneity of endothelid cek: the specialized phenotype of human high endothelid venules characterized by suppression subtractive hybridizatioa ArilLPathoL, t SS:2043-2055-
Girard, J.P. and Springer,'ï.A. (1995) High endo theliai venules (HEVs) : specialized endotheiium for lymphocyte migration, ïmmunoLToday, 16:449-457.
Glimaker,M., Kmgsbjerg,P., Forsgren,M., and Olcen,P. (1993) Tumor necrosis factor- alpha (TNF alpha) in cerebrospinal fîuid fiom patients with meningitis of different etiologies: high levels of TNF alpha uidicate bacterial menin@. JJnfect.Dis., 167:882- 8 89.
GowansJ-L. (1959) The recirculation of lymphocytes ftom blood to lymph in the rat. J-Physiol (Land) , 14654-69.
Gowans J.L. and Knight,E.J. (1964) The route of re-circulation of Lymphocytes in the rat- Proc.R.Soc.Lond B BioLSci., 159:257-282.
Gum,MD., Ngo,V.N., AnseI,ICM., Elciand,E.H., CysterJ-G., and Wiiliams,L.T. (1998) A B -ceIl-ho ming c hemokine made in lymp ho id fo iiicles activates Burkitt's l p p ho IM receptor- 1. Nature, 39 1:799-803.
Gupta,V.K, McConnellJ., DalWeI,R.G., and Hopkins,l. (1998) Two B ceU subpopulations have distinct recûculation characteristcs. Eur.Jhmunol., 28: 1597- 1603-
Ha£ier,D.k, Fox,D.A., Manning,M,E., SchIossman,S.F., Reinherz,EL., and Weiner,H.L. (1985) In vivo activated T lymphocytes in the peripheral blood and cerebrospinal fluid of patients with multiple sclerosis, N.Eng1. J-Me&, 3 12: L4OS- 141 1.
Haig,D., HopkinsJ., and Miller,H. (1999) Local immune responses in aEerent and efferent lymp h. Immunology, 96: 155- 163.
Hail,l.G. and Morris,B. (1965b) The immediate effect of antigens on the ce11 output of a lymph node. BrJExpPathol., 46:450-454.
Hall,J.G. and Morris,B. (1965a) The orïgins of the cells ih the efferent Lymph fkom a single lymph node. J.Exp.Med., 12 1390 1-9 10.

Harhg-Berg,C.J.. Park,J-T. and I(nopE PM. (1999) Role of cervical lymphatics in the TM-type hierarchy of CNS h u n e regulation. L Neurommunol. 10 1 : 1 1 1- 127
Harling-Berg,C., Knopf$.M., Merriam,J., and Cserr,ELF. (1989) Role of cervicd lymph nodes in the systemic humoral immune response to human serumalbumin microinfused into rat cerebrospïnal £luid, L.Neuroimmunol., 25:L85-193.
Hay,J-B. and Hobbs,BB. (1977) The flow of blood to Lymph nodes and its relation to lymphocyte traffic and the immune response, J-ExpMed., 145:3 1-44
Hein, W.R. and Supersaxo,k (1988) Effect of interferon-alpha-2a on the output of recirculating Lymphocytes fkom single lyrnph nodes. Immunology, 64469474.
Hickey,WS. (199 1) Migration of hematogenous ceils through the blood-brain barrEe~ and the initiation of CNS infiammation, Brain Pathol., 1:97-105.
Hickey,W.F. (1999) Leukocyte trafnc in the central nervous systenï the participants and their roles. Semin.ImrnunoL, 1 1 : 125-138.
Hickey,W.F., Hsu,B.L., and Kimura,H. (199 1) T-lymphocyte entry into the central nervous s YS tem J.Neurosci.Res., 28:254-260.
Hofinan,F.M., von Hanwehr,R.I., Dinareilo,C.A., Mize1,S.B-, and Merrïll,J.E- (1986) Immunoregulatory molecules and IL 2 receptors identined in multiple sclerosis braia J.ImmunoI.,3239-3245.
Hwang,S.T.. Shger,M.S., Giblin,P.A., Yednock,T.A., Bacoo.,ICB., Simon,S.I., and Rosen,s.D. (1996) GlyCAM-1, a physiologie ligand for L-selectin, activates beta 2 integrins on naive peripheral lymphocytes. J-Exp-Med., 184: 1343 - 1348-
Issekutz,T., Chh, W., and Hay, J.B. (1980) Measurement of lymphocyte tranic with indium-1 11. Ch.Exp.ImmunoL, 39:215-221.
Issekutz,T.B., Chin,G.W., and Hay,J.B. (198 1) Lymphocyte t r a c tbrough chronic inflarnrnatory lesions: differential migration versus differential retention. Ch-Exp.Immunol., 45:604-6 14.
Izawa,D., Tanaka,T., Saito,K, Ogihara,H., Usuï,T., Kawamo to,S ., MatsubaraK, Okubo ,KT and Miyasaka,M. (1999) Expression profile of active genes in mouse lymph node high endothelial cells. Int.Tmmunol., 11: 1989-1998.
Jahn,S ., Bauer,B ., Schwab, J., KirchmairJ,, Neuhaus,K-, Kiessig,S .T., Volk,H-D-, Mau,H., von Baehr,R., and Specht,U. ( 1993) h u n e restoration in chilken after partid splenectomy. Immunobiol. 188:370-378
Jongen,P. J.H., Lamers,K J.B., Doesburg, W.H., Lemmens,W.A. J.O., and Hommes,O.R. ( 1997) Cerbrospùial fluid analysis differentiates between relapsing-remitting and secondary progressive multiple sclerosis. J.Neurol.Neurosurg.Psychiatry, 63r446-45 1.

Jutila,M.k, Berg,E.L., Kishumto,T.K, PickerA J., Bargatze,RF., Bishop,D.K, Orosz.C.G., Wu,N.W., and Butcher,E.C. (1989) Mammation-induced endothelid ceii adhesion to lymphocytes, neutrophils, and monocytes Role of homing receptors and other adhesion molecules. Transplantation, 48~727-73 1.
Kdaaji,lw., Abemethy,N.J., McCuUough,KT and Hay,J.B. (1988) Recombinant bovine interferone-a inhibits the migration of Iyphocytes from lymph nodes but not into lymph nodes, Reg- Immunoi. 156
Kallaaji,A.N.. McCulIough,IC, and Hay,l.B. (1989) The enhancement of Lymphocyte localkation in skïn sites of sheep by tumour wcrosis factor alpha. Immun01 Letters 23: 143-148
KimC.H. and Broxrneyer,H.E. (1999) Chemoklnes: signai Iarnps for traficking of T and B cells for development and effector fiinction- ILeukoc_BioI., 65:6-15.
Kleine,T.O., Aibrecht, JO, and Zofe/P. ( 1999) Flow cytometry of cerebrospinal fluid (CSF) lymphocytes: alterations of bloodlCSF ratios of lymphocyte subsets in inflammation disorders of human cenual nervous system (CNS). Clin-ChemJab Med., 37:23 1-241.
KnopEP-M., Harling-Berg,C..J., Cserr,H.F., Basu,D., Sirulnick,E.J., Nolan,S-C., Park,J.T., Keir,G., Thompson, E.I. and Hickey,WF. (1998) Antigen-dependent intrathecal anti idy synthesis in the normal rat brak tissue entry and local retention of antigen-specinc B cells. J. ~ u n o l . 16 13692-70 1
Knop&P.M., Cserr,H.F., N01an~s.C.. Wu J.Y., and Harling-Berg,C.J. (1995) Physiology and immunology of iymphatic drainage of interstitial and cerebrospinai fluid fiom the brain. NeuropathoI.Appl.Neurobiol., 2 1 : 175- 180.
Kooprnann,W., Ediriwickrema,C., and Krangel,M.S. (1999) Structure and b c t i o n of the glycosaminoglycan binding site of chemokiae macrophage-infhmmatory protein-1 beta. J.Immunol., 163:2120-2127,
Komer,H., Lemckert,F.A., Chaudhri,G., EtteIdorf,S., and Sedgwick,J.D. (1997) Tumor necrosis factor blockade in ac tively induced experimental autoimmune encephalo myelitis prevents clinical disease despite activated T celi infiltration to the central nervous system Eur.J.lmmunol., 27: 1973-198 1,
Kniegers., Low,C., and Donachie,W. (1995) Pheaotypic charactekation of the ceUs of the inflamed response in ovie encepbalitic listeriosis. J.Comp.Pathol., 1 13:263-75.
Lascelles,A.K. and Morris,B. (1961) Surgical techniques for the collection of Lymph nom unanaesthetized s heep. Qum. I. Exp. Physiol.46: 199-205
Lindhutz., Visers,J.L.M., Figdor,C.G., and Adema,G.J. (1999) Chemokines and lymphocyte migration. The hunologist , 7: 147-152.

LiuJ.. Marin0,M.W-. Wang-G., Graü,D., Dunn,k, Bettadapura,J., Slavin,AJ., Old&., and Bemard,CC. (1998) TNF ïs a potent anti-infiammatory cytokine in autoimmune- mediated demyelination. Nat-Med., 478-83,
Ludowyk,P.A-, Wïenborg,D.O., and Parish,CK (1992) Selective localisation of neuro- specific T lymphocytes in the central nervous system. I.Neuroimmuno1.. 37:237-250-
Mackay,C. (1988) Sheep Ieukocyte moIecuIes: a review of their distribution, stmcture and possible fiiaction. Vet.Immunol.Imtnunopath01., 19: 1-20.
Mackay,C.R-, Andrew,D.P., Briskin,M., Ringler,DJ., and ButcherE-C- (1996) Phenotype, and migration properties of three major subsets of tissue homing T c e h in sheep. Eur. J .buno1-, 26:2433-2439,
Mackay,C.R.. Kimpton,W.G., Brandon,MR., and Cahill,R.N. (1988) Lymphocyte subsets show marked ciifferences in their distri'bution between blood and the afferent and efferent lymp h of peripheral lymph nodes. IExpMed., 167: 1755- 1765.
Mackay,C.R., Maddox,J.F., and Brandon,MX. (1986) Three distinct subpopulations of sheep T lymphocytes. Eur.Jhmunol., 16: 19-25.
Mackay,C.R., Marston,W., and Dudler,L. (1992b) Altered patterns of T celi migration through lymph nodes and skin following antigen challenge. Eur.J.Immuno1-, 22:2205- 22 10.
Mackay,C-R., MarstogW.L., and Dudler,L. (1990) Naive and memory T cells show distinct pathways of lymphocyte recirculation. JExpMed., 17 M O 1-8 17.
Mackay,C.R., Marston,W.L., Dudler,L., and Heh, W.R. (199 1) Expression of the "T 19" and "null ceW markers on gamma delta T cells of the sheep. Vet.ImmunoLImmunopathol., 27: 183- 188,
Mackay,C-R., Marston,W.L., DudlerJ,., Spertini,O., Tedder,T.F., and Hein, W.R. (1992a) Tissue-specinc migration pathways by phenotypically distinct subpopulations of memory T cells. Eur.J.Immunol., 22:887-895.
Male,D., RahmanJ., Linke,k, Zhao,W., and Hickey,W. (1995) An interferon-inducible molecule on brain endotheliurn which controls lymphocyte adhesion mediated by Uitegrins. Tmmunology, 84:453-460.
Matyszak,M.K and Perry,V.H. (1996) A comparison of leucocyte responses to heat- killed baciüus Calmette- Guerin in dBerent CNS compattments. Neuropathoi.Appi.Neurobioi., 22:44-53.
Mix,E., Correale,O. J., Kostulas,V., and Li&,& (1990) CM+, CD8+, and CD4-CD8- T cells in CSF and blood of patients with muitipie sclerosis and tension headache. Scand. J.Immunol., 3 1 :493-SO 1.

Mor,F- and CohenJX (1992) T c e k in the lesion of e x p e b n t a i autoimmune encephalomyelitis. Enrichment for reactivities to myelin basic protein and to heat shock proteins- J,ChJnvest, 90:2447-2455.
Navikas,V. and Link,H- (1996) Review: cytokines and the pathogenesk of multiple scierosis. JNeurosci.Res., 42322-333,
ODonneIi,M-A, Luo,Y., Chen,X., Szilvasi,A, Hunter,S.E., and Clinton,S-K- (1999) Role of IL-12 in the induction and potentiation of IFN-gamma in response to bacillus Calmette-Guerin. J.Iuununol., 163 r4246-4252.
Oehmichen&Z., Grunhger,H., WiethoIter,H.. and Gencicw. (1979) Lymphatic efflux of intracerebrally injected cells. Acta NeuropathoI.(Berl), 45:6 1-65.
Pabst,R. and Rosenberg.Y.J. (1998) Interpethg data on Lymphocyte subsets in the blood of HIV patients - organ distriiution, proliferation and migration kioetics are criticai factors. Pathobiology, 66: 1 17- 122.
Pabst,R md Westermann,J. (1991) The role of the spleen in Lymphocyte Scanning Microsc., 5: 1075- 1079,
Pan, W.. Banks, W.A., and Kastin,kJ. (1997a) PermeabiIity of the blood-brain and blood- spinal cord barriers to interferons. JNeuroirnmunol., 76: 105- 1 1 1.
Pan,W-, ZadinaJE., Harlan,R.E., Weber,J.T., Banks,W.A., and Kastin,A.J- (1 997b) Tumor necrosis factor-alpha: a neuromoduiator in the CNS. NeurosciBiobeha~Rev., 21:603-613.
Paris,M.M., FriedlandJ-R, Ehrett,S., Hicke y,S .M.. Olsen, KD.. Hansen,E., ThonarE-J., and McCracken,G.H., Ir. (1995) Effect of interleukin- l receptor antagonist and soluble tumor necrosis factor receptor in animal models of infection. J.Infect.Dis., 17 1: 161- 169.
Parish,C.R. (1999) Fluorescent dyes for lymphocyte mi-gration and proliferation snidies. Irnmunol.Celi B iol., 77:499-508.
Peilas,T.C. and WeissL. (1990) Deep splenic lymphatic vessels in the mouse: A route of splenic exit for recirculating lymphocytes. Am. J. Anat .. 187:347-354.
PhillipsL.M. and Lampson,L.A* (1999) Site-specific control of T cell M c in the brain: T cell entry to brainstem vs. hippocampus aftet local injection of IFN-gamma J.NeuroimmunoI., 96:2 28-227.
Phillips,M.J-, Needham,M., and WeIier,R.O. (1997) Role of cervical lymph nodes in autoimmune e ncephalomyelitis in the Lewis rat. J-Pathol., l82:457-461.
Picker,L. and Siegelman,M. (1993) Lymphoid tissues and organs. In: Fundamental Immunology. W.Paul, ed. Raven press, New York, pp. 145-197.

Pryce,G, Santos,W., and Male,D. (1994) An assay for the aoalysis of lymphcyte migration across cerebrd e n d o t h e h in vitro, J~unoLMethods , 16755-63-
Quagliarelio,V.J., WispeIweyJI., Long,\N.J., and Scheld,WM. (199 1) Recombinant human interleukin-1 induces meningitis and blood-brain bamer injury in the rat- 1-Clinhvest, 87: 1360-1366,
Raine,C.S. (1994) The Dale E. McFarlia Mernorial Lecture: the immunology of the multiple sclerosis lesion. AnaNeuroI., 36 SuppkS6 1472.
Raine,C.S., CanneUa,B., Duijvestijn,AM., and Cross,A.K (1990a) Homing to central nervous s ystem vasculature by aocigen-specifîc lymphocytes. II. Lymp hocytelendothelial celi adhesion during the initiai stages of autoimmune demyelination. Lab Invest, 63:476- 489,
Raine,C.S., Lee,S.C., Scheinberg,L.C., Duijvestin,A.M.. and Cr0ss.A.H- (1990b) Adhesion molecules on endotheliai c e k in the central nervous system: an emerging area in the neuroimmunoiog y of multiple sclerosis. Ciin.Immuw LImmunopathoL, 57: 173- 187.
Ram;io,O., Saez-Llorens,X., MertsolaJ., lafari&., Okn,ICD., HansenE. J., YoshinaghM., Ohkawara,S., Nariuchi,K, and McCracken,G.H., Jr. (1990) Tumor necrosis factor aIp hdcachectin and kterleukin 1 bet a initiate rnenhgeal inflammation. J.Exp.Med-, 172:497-507.
Reynolds,J., HeronJ., DudIerJ., and Trnka,Z. (1982) T-ceU recirculation in the sheep: migratory properties of cells fiom lymph nodes. Immunology, 47:415-421-
Romano,M-, Sironi,M., Toniatti,C., Polentanitti,N., Fruscella,P., Ghezzi,P., FaggionLR., Luini,W., van,H., V, Sozzanï,S., Bussolino,F., Poi.i,V., Cih'beao,G., and MantovanLA- (1997) Role of IL-6 and its soluble receptor in induction of chemokines and leukocyte recruitment. Immunity., 6:3 15-325.
Sallusto,F., Palermo,B., Ho y , k , and Lanzavecchia,A. (L999) The role of chemokine receptors in directing trafic of naive, type 1 and type 2 T cells. Curr.Top.Microbiol.Immunol., 246: 123-128.
Salvato,M.S., Rater,M., and Pauza,CD. (1996) Tracking of dye-labelied lymphocytes in rhesus macaques. I.Med.PrimatoI., 25: 1 12-121.
Samlowski,W.E., Robertson,B.A., Draper,B.K, Prystas,E., and McGregor,J.R. (1991) Effects of supravitai fluorochromes used to analyze the in vivo homing of murine lymphocytes on cellular function. J.ImrnunoLMethods, le 10 1- 115.
Saukkonen,IC, Sande,S., Cioffe,C., Wolpe,S., Sheny,B., Ceramï,k, and Tuamuien,E. (1990) The role of cytokines in the generation of inflammation and tissue damage in expimental grain-positive menùigitis. J.Exp.Med., 17 1:439-448.

SchteiBenbaum,B. and Fehr,L (1996) Regdation and selectivity of leukocyte emigration. JLab.Clin.Med., 127: 168.
Schoappauf,B and SchnappaufU (1968) Drainage of the thoracic duct and amount of the "easily mobilued" Lymphocytes in calves, sheep and dogs. Blut, 4209-220.
Schne&L., Fearn,S., Schwab,M.E., PenyfV.H., and Anth0nyS.C. (1999) Cytokine- induced acute iaflammation in the brain and spinal cord. INeuropathoIE~Ne~1:ol., 5 8 :245-254,
Schoning&., ElepfmdtJ?., Lanksch,WX., Vok,H.D., and Woiciechowsky,C- (1999) Conîinuous inhision of proinfiammatory cytokines into the brain to study brain cytokine induced local and systemic immune effects. Brah Res.Protocol, 4217-222-
ScoIozzi,R,, Boccafogii,A., Tola,M.R., Viceati'ni_L., Camerani,A., Degani,D., Granieri,E., Caniatti,L.. and Paolino,E. (1992) T-celi phenotypic promes in the cerebrospinai faid and peripheral blood of muitiple sclerosis patients. JNeuroLSci*, 108:93-98.
SeabrootT., AuB., DicksteinJ., Zhang,X., Ristevskï,B.. and Hay,J.B. (1999) The traffic of resting Lymphocytes through delayed hypersensitivity and chronic inflammatory lesions: a dynamic equili'brium, Semin-Tmmunol., 11: 115- 123 - Selmaj, K. ( 1996) Pathophsiology of the blood-brain barrier. In: Immunoneurology. M-Chofflon and L-Steinman, eds. S p ~ g e r , Berlin, pp. 175-19 1.
Sieber,G., Breyer,H.G., HerrmaM,F., and RuM,H. (1985) Abnormaüties of B-ceil activation and immunoregulation in splenectomized patients. Immunobiology, 169:263- 27 1.
Sirnpson-Morgan,M. W., Trevella,W., Hugh,AR., McClure,S.J., and Morris,%. (1985) The Long-term CO llection of lymp h from single lymph nodes of foetal Iambs in utero. Aust.J.Exp.Biol.Med.Sci., 63 ( Pt 4):397-409.
S ipe,J.C., FilippLM., Martino,G., Furlan,R., Rocca,M.A., Rovaris,M-, Bergami,A., Zyro ff, J., Scotti,G., and Comi,G. (1999) Method for ktraceiiular magnetic labeling of human mononuclear celis using approved iron contrast agents. Magn Resonhging, 17:1521-1523.
SmithJB., McIntosh,GX., and Morris,B. (1970) The traffk of ceils through tissues: a study of peripherai lymph in sheep. J.Anat., lO7:87- 100.
Spellerberg,B. and TuomanenE.1. (1994) The pathophysiology ofpneumococcd meningitis. Ann.Med-, 26:411-418.
Spertini,O., Luscinskas,F. W., Kansas,G.S ., Munro,I.M., Griffui,J.D., Gimbrone,M.A, Jr., and Tedder,T.F. (1991) Leukocyte adhesion molecule-1 (LAM-1, L-selectin) interacts

with an inducible endothelial cell ligand to support Ieukocyte adhesioe JImmunoL. l47:2565-2573,
SprentJ. (1994) T and B mmory cek. CeIl, 76:3 15-322.
Springer,T.A. (1994) Traffic signais for lymphocyte recirculation and Ieukocyte emigration: the muitistep paradigm Ceil, 76101-3 14.
S pringer,T.A. ( 1995) Traffic signals on endothelium for lymphocyte cecirculation and Ieukocyte em-gration. A~u.Rev.Physiol,57:827-872.
Steffen,B.J., Breier,G., Butcher,E.C., Schuiz,M., and EngelhardtB. (1996) ICAM-1, VCAM- 1, and MAdCAM-1 are expressed on choroid plexus epithelium but not endotheliam and mediate bmding of lymphocytes m vitm AmLPathoL. 148 : 18 19-1838-
S tein,J.V., Cheng,G., Stockton,BM, Fors,B.P., Butcher,E.C., and von Andrh,U.H. (1999) L-selectin-mediated leukocyte adhesion in vivo: microvilious distribution determines tethering eficiency. but not rolling velocity. J-ExpMed., 189:37-50.
Sugisaki,K, Dannenberg,A.M., Jr., Abe,Y., TsurutaJ., Su,W.J., Said, W., Feng,L., Yoshimwa,T,, Converse,P.J., and Mounts,P. (1998) Nonspecific and immune-specinc up-regulation of cytokines in rabbit d e d tuberculous (BCG) lesio ns. J.LeukocB iol., 63 :440-4SO-
Svenningsson,k, Andersen.O., Edsbagge,M., and Stemme,S. (1995) Lymphocyte phenotype and subset distribution in normal cerebrospinai fluid. J.NeuroirnrnunoL, 63:39- 46.
Svenningsson,A, Hansson,GK, AnderseaO., Andersson,R., Patarroyo&l., and S temme,S . ( 1993) Adhesio n rnolecde expression on cerebrospinal fluid T lymphocytes: evidence for common recnritment mechanisms in multiple scierosis, aseptic rneningitis, and normal controls. Ann.Neurol., 34155-161.
Tang,T., Frenette,P.S., Hynes,R.O., Wagner,DS., and Mayadas J-N. ( 1996) Cytokine- induced rneningitis is dramaticaliy attenuated in mice deficient in endothelial selectins. J.Ch.Invest, 97:2485-2490.
Taub,D.D. and Murphey, W. (1997) Chemokines as regulators of leukocyte adhesion and migration. In: Adhesion Molecules an Chemokines in Lymphocyte Trafockuig. A.Hnm;inn, ed. Harwood Academic Publishers, Amsterdam, pp. 89-128.
Teare,G.F., Hocan,P.K., Slezak,S E ., Srnith,C., and HayJB. (199 1) Long-term t rachg of lymphocytes in vivo: the migration of PKH-labeiied Lymphocytes. Ceii Immuw l, 134: lS7-l7O.
Torsteinsdottir,S .. Georgsson.G., GisladottV,E., Rafoar,B ., Palsson,P.A., and Penirsson,G. (1992) Pathogewsis of central nervous system lesions in visna: ce&

mediated immunity and lymphocyte subsets in blood, brain and cerebrospinal fluid- JNeuroimmunol., 41~149-158,
Vass,K. and Lassmann,H. (1990) htrathecal application of interferon gamma Progressive appearance of MHC antigeos withio the rat nervous system Am.JJ?athol., 137:789-8Oow
Vestweber,D. and Blanks, J.E. (1999) Mechanisms that regulate the finction of the selectins and their Ligands. Physiol Rev., 79:l8 1-2 13.
Vrethem&l., Dahie,C., Ekerfelt,C., Forsberg,P., Danielsson,O., and Emerudh,J. (1998) CD4 and CD8 lymphocyte subsets in cerebrospinal fluid and peripheral blood hom patients with multiple sclerosis, meningitis and normal controls. Acta Neurol.Scand., 97~2 15-220.
Walter,S., Micheel& Pabst,R., and Westermann,J. (1995) Interaction of B and T lymphocyte subsets with high endothelîal vendes in the rat: binding in vitro does not reflect homing in vivo. Eur.Jmunol., 25:1199-1205.
Waniock,R.A., Campbeli,J.J.. Do~,M.E., Matsuzawa,A., McEvoy,L.M., and Butchers-C- (2000) The roIe of chemokines in the microenvironmental control of T versus B cell arrest in Peyer's patch high endothefial vendes Process Citation]. J-ExpMed., 19 1~77-88.
Wekerle,H., Linington,H., Lassmann_H., and MeyenmmJL (1986) Cellular immune reactivity within the CNS. Trends Neurosci., 9:27 1-276.
Weller,R.O. (1 998) Pathology of cerebrospinai fluid and interstitiai fluid of the CM: signincance for Alzheimer disease, prion disorders and multiple sclerosis. J.Neuropathol.Exp.Neurol., 57:885-894.
Westermann,l-, Nagahori,Y., Walter,S., Heerwagen,C., Miyasaka,M., and Pabst,R- (1994a) B and T Lymphocyte subsets enter peripheral lymph nodes and Peyer's patches w ithout preference in vivo: no correlation occurs between their localization in different types of high endotheliai venules and the expression of CD44, VLA-4, LFA-1, ICAM-1, CD2 or L-selectin. Eur.J-Tmmunol., 2423 12-23 16.
Westermann,J. and Pabst,R. (1990) Lymphocyte subsets in the blood: a diagnostic windo w on the lymphoid system? [see comments] . ImmunoLToday, 1 MO64 10.
Westermann,l. and Pabst,R (1996) How organ-specific is the migration of 'naive' and 'memory' T celis? [see comments]. ImmunoLToday, 17:278-282.
WestermannJ., Persin,S ., MatyasJ., van der,M.P., and Pabst,R. (1993) IFN-gamma influences the migration of thoracic duct B and T lymphocyte subsets in vivo. Random hcrease in disappearance fkom the blood and differential decrease in reappearance in the lymph. J.Immunol., 150:3843-3852.

WestermumJ-, Persin,S., Matyas,J., van der,M.P., and Pabst& (1994b) Migration of so- c a e d naive and memory T lymphocytes fiom blood to lyrnph in the rat. The influence of IFN-gamma on the circulation pattern. JJmamnoL, 152: 1744-1750.
Westemam,J.~ PershS., Matyas,J., van der,M.P., andPabst,R- (1994~) Migration of so- c d e d naive and mernory T Lymphocytes fiom blood to Lyrnph in the rat. The influence of FN-gamma on the circulation pattern. LImmunoI., 152:1?44-1750.
Westermam,J., SchwiBzer,R., Jecker,P., and Pabst& (1990) Lymphocyte subsets in the blood. The influence of splenectomy, splenic autotransplantation, ageing, and the site of blood samphg on the number of B, T, CD4+, and CD8+ lymphocytes in the rat. S~~d.J.Tmmun~l., 3 1:327-334.
Westermann,J., Willfuhir,KU., RothkotterST. J., Fritz,F.J., and Pabst,R. ( 1989) Migration pattern of lymphocyte subsets in the normal rat and the influence of splenic tissue. Scand J-Imrriunol-, 29: 193-20 1.
W'ienborg,D-O., Simmons,R.D., Tamatani,T., and Miyasaka,M. (1993) ICAM-1- dependent pathway is not cnticaily involved in the inflammatory process of autoimmune encephalomyelitis or in cytokine-induced inflammation of the central nervous system. J.Neuroirnrnunol., 45: 147- 154.
Williarns,KC- and Hickey,W.F. (1995) T r a c of hematogenous ceils through the central nervous system Curr.TopMcrobiol_ImmunoL, 202:22 1-245.
Wiarns,M.B- and Butcher,E.C. (1997) Homuig of naive and memory T lymphocyte subsets to Peyer's patches, lymph nodes, and spleen, J_Immunol., 159: 1746- 1752.
Wilson,E., Aydintug,M.K, and Juùla,M.A- (1999) A circulating bovine gamma delta T ceU subset, which is found in large numbers in the spleen, accumulates inefficiently in an artificial site of infiammation: correlation with lack of expression of E-selectin Egands and L-selectin- J.Immunol., 162:49 14-49 19.
Wilson,E., Walcheck,B., Davis,W.C., and Jutila,M.A. (1998) Preferential tissue localkation of bovine gamma delta T ceii subsets defined by anti-T celi receptor for antigen antiidies- ImmunoLLett ., 64:39-44.
Witherden,D.A., Kimpton,W.G., Washuigton,E.A., and CahiU,R.N. (1990) Non-random migration of CD%, CD€!+ and gamma delta+T19+ lymphocytes through peripheral lymph nodes. Immunology, 70:235-240.
Wolbur,o,KI, Gerhardt,H., Schulz,M., Wolburg,H., and Engelhardt,B . (1999) Ultrastructural localization of adhesion molecules in the healthy and inflarned choroid plexus of the mouse. CeU Tissue Res., 296:259-269.
Wright,J.L. and Merchant,R.E. (1992) Histopathological eecs of intracerebral injections of human recombinant tumour necrosis factor alpha hthe rat. Acta Neuropathol., 85:93- 100.

Young, k J. Steady-state analysis of recircuiating Lymphocyte p i s in the sheep. 1994. University of Toronto. P h . Thesis
Young,A.J. and Hay,J.B. (1995) Rapa turnover of the recirculating lymphocyte pool in vivo. I n t h u n o L , 7: 1607-16 15.
Young,A. J-, I3ayJ.B ., and Mackay,CR (1993) Lymphocyte recirculation and life span in vivo. CurrTop-Microbio 1.Immuao II, 184~16 1- 173.
Young,A.L, Heïn,W.R., and HayJ-B. (199%) Cannulation of iymphatic vessels and its use in the study of lymphocyte traffic- In: Immunology Methods Manual. 1.Lefkovits. ed- Academic Press, San Diego. pp. 2039-2059.
Young,A. J., Marston,W.L., Dessing,M., Duder&., and Hein,W-R- (1997a) Distinct recirculating and non-recircuiating B-Lymphocyte pools in the peripheral bIood are dehed by coordinated expression of CD2 1 and L- selectin. Blood, 90:4865-4875-
Young,A. J., Seabrook,T. J., Marstoa,W.L,., Dudler,L., and Hay,JB, (2000) A role for lymphatic endothelium in the sequestration of recirculating gamma delta T ceh in TNF- alpha-stimulated lymph nodes. Eur.J.Immunol., 30:327-334.
Zocchl,MIR., Ferrero$., Leone,B.E., Rovere,P., Bianchi,E., Tonineiii,E., and PardkR. ( 1996) CD3 1PECAM-1-driven chemokine-independent transmigration of human T lymphocytes. E u r . J ~ u n o l . , 26:759-767 -