the alleles of the bft gene are distributed differently among
TRANSCRIPT

JOURNAL OF CLINICAL MICROBIOLOGY,0095-1137/00/$04.0010
Feb. 2000, p. 607–612 Vol. 38, No. 2
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
The Alleles of the bft Gene Are Distributed Differently amongEnterotoxigenic Bacteroides fragilis Strains from Human
Sources and Can Be Present in Double CopiesANNA SCOTTO D’ABUSCO,1 MARIA DEL GROSSO,1 STEFANO CENSINI,2
ANTONELLO COVACCI,2 AND ANNALISA PANTOSTI1*
Laboratory of Bacteriology and Medical Mycology, Istituto Superiore di Sanita, Rome,1 andImmunobiological Research Institute of Siena, Chiron Vaccines, Siena,2 Italy
Received 28 June 1999/Returned for modification 11 October 1999/Accepted 10 November 1999
Enterotoxigenic Bacteroides fragilis (ETBF) strains are associated with diarrheal disease in children. Thesestrains produce a zinc metalloprotease enterotoxin, or fragilysin, that can be detected by a cytotoxicity assaywith HT-29 cells. Recently, three different isoforms or variants of the enterotoxin gene, designated bft-1, bft-2,and bft-3, have been identified and sequenced. We used restriction fragment length polymorphism analysis ofthe PCR-amplified enterotoxin gene to detect the isoforms bft-1 and bft-2 or bft-3 borne by ETBF. By sequencingthe portion of the bft gene corresponding to the mature toxin in some strains and applying allele-specific PCRfor strains categorized as bft-2 or bft-3, we found in our collection two strains harboring bft-3, a variant thathad been described for isolates from East Asia. Analysis of 66 ETBF strains from different sources showed thatbft-1 is the most frequent allele, being present in 65% of isolates; it is largely predominant in isolates from fecesof adults, while bft-2 is present in isolates from feces of children. This association is statistically significant (P,0.0064). Sixteen strains were examined by Southern hybridization using, as probes, the bft and secondmetalloprotease genes, both included in a pathogenicity islet. Five strains were found to harbor double copiesof both genes, suggesting that the whole islet was duplicated. Four of these strains, harboring bft-1 (threestrains) or bft-2 (one strain), were found to produce a large amount of biologically active toxin, as determinedby a cytotoxicity assay with HT-29 cells. The strains harboring bft-3, either in a single copy or in double copies,produced the smallest amount of toxin in our collection.
Bacteroides fragilis is the anaerobic microorganism most fre-quently isolated from infectious processes in humans (6) aswell as a common component of the normal flora of the colon(7). Some of the strains belonging to this species are able toproduce an enterotoxin and are therefore termed enterotoxi-genic B. fragilis (ETBF) strains (17). ETBF strains are respon-sible for diarrheal diseases in young farm animals, includinglambs, calves, and foals (17–19). Several studies have alsoshown an association between the isolation of ETBF fromfeces and acute diarrhea in children 1 to 5 years old (27, 28,30). In addition, ETBF can be recovered from the normal fecalflora of healthy subjects, especially adults (25). In some of thesubjects harboring ETBF, the enterotoxin is present in thefeces in a biologically active form and can be detected by acytotoxicity assay (26).
B. fragilis enterotoxin, recently termed fragilysin (20), hasbeen characterized as a 20-kDa zinc-dependent metallopro-tease belonging to the metzicins family, precisely to the sub-family that comprises the eukaryotic collagenases or matrixins(15). Recently, the target of fragilysin has been identified asthe cell surface protein E-cadherin, which is the principalstructural component of the zonula adherens and is responsi-ble for cell-cell adhesion of eukaryotic cells (32).
Two groups of investigators have independently cloned andsequenced the enterotoxin gene from two ETBF strains andhave identified two different allelic forms of this gene. The firstpublished sequence of the enterotoxin gene, bftP (or bft-1), was
obtained from strain VPI 13784, a lamb isolate (12). The otherisoform, bft-2, was sequenced from a porcine isolate (8). Bothisoforms code for a protein much larger than the mature en-terotoxin, a “preprotoxin” of 45 kDa comprising a signal pep-tide and a protoxin from which the mature toxin is releasedupon cleavage. Although the two forms of the enterotoxin geneare 95% identical, they show lower homology in the regioncorresponding to the mature toxin moiety (8). New allelicforms of the enterotoxin gene were found in ETBF isolatedfrom blood in Korea (5) and in fecal isolates from Japan (N.Kato, Final Program of the 2nd World Congress on AnaerobicBacteria and Infections, abstr. 5.002, p. 74, 1998) and weretermed Korea-bft and bft-3, respectively. As the correspondingnucleotide sequences are identical, we refer to the third allelicform of the enterotoxin gene as bft-3 throughout this paper.The bft-3 isoform shares high similarity with the other twoforms but is more related to bft-2 (96% identity) (5). Its fre-quency among ETBF strains outside East Asia remains un-known.
Moncrief et al. (16) found that the B. fragilis enterotoxingene is located in a pathogenicity islet together with a putativesecond metalloprotease (MP II) gene which has a low identity(28%) with the enterotoxin gene. The pathogenicity islet of B.fragilis is a genetic unit that shares some characteristic featureswith the larger pathogenicity islands present in pathogenicstrains of Escherichia coli, Salmonella, or Helicobacter pylori. Itcontains virulence genes (for the enterotoxin and the putativevirulence factor MP II), has a lower G1C content than the B.fragilis chromosome, contains almost identical direct repeats inproximity to its ends, and is inserted in the same chromosomalregion in different strains (16).
In a previous study, we used PCR to amplify a portion of the
* Corresponding author. Mailing address: Laboratory of Bacteriol-ogy and Medical Mycology, Istituto Superiore di Sanita, Viale ReginaElena 299, 00161 Rome, Italy. Phone: (39) 06 4990-2331. Fax: (39) 064938-7112. E-mail: [email protected].
607
on March 29, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

bft gene and showed that this gene is present in all ETBFstrains but not in nonenterotoxigenic strains (24). In the pres-ent study, using PCR-restriction fragment length polymor-phism (RFLP) analysis, an allele-specific PCR, and sequencingdata, we have shown the frequency of the two principal iso-forms, bft-1 and bft-2, in a large collection of ETBF strains ofhuman origin. We have also found that the variant bft-3, al-though rare, is present in ETBF isolated in Europe. Moreover,we have found a duplication of both the bft and MP II genes infive strains that probably represents a duplication of the entireislet.
MATERIALS AND METHODS
Bacterial strains. Sixty-eight ETBF strains from different sources were stud-ied; these comprised 28 strains isolated from clinical (extraintestinal) samples, 23strains isolated from the feces of children (18 with diarrhea), and 17 strainsisolated from the feces of adults (10 with diarrhea). The extraintestinal isolatescame from the collection of the Istituto Superiore di Sanita and included isolatesobtained from Italy (18 strains), the United Kingdom (6 strains), and the UnitedStates (4 strains) and characterized in previous studies (22, 23). The fecal isolateswere obtained in Italy during previous (22, 25) and ongoing studies on theprevalence of ETBF in the country and were isolated in different geographicalareas over a span of 5 years. The reference strain ATCC 43858, originallyisolated in the United States from the feces of an infant with diarrhea, was alsoincluded. The bacterial strains were identified as ETBF by previously describedmethods, including a cytotoxicity assay with HT-29 cells (22) and PCR amplifi-cation of an internal fragment of the enterotoxin gene (24). Strain VPI 13784 (agift from T. D. Wilkins, Virginia Polytechnic Institute and State University,Blacksburg) was included in the study as the prototype of the bft-1 genotype. Thenontoxigenic strain B. fragilis NCTC 9343 was used as a negative control.
Chemicals and enzymes. Nylon transfer membranes (Hybond N) were ob-tained from Amersham International (Little Chalfont, Buckinghamshire, UnitedKingdom). Restriction endonucleases were purchased from New England Bio-labs (Beverly, Mass.) and Roche Molecular Biochemicals (Milan, Italy). ForPCR amplification, DynaZyme II (obtained from Finnzyme, Oy, Finland) orAmpliTaq (obtained from PE Applied Biosystems, Roche Molecular Systems,Branchburg, N.J.) DNA polymerase was used. The oligonucleotide primers weresynthesized at Laboratori Genenco, M-Medical, Florence, Italy. Electrophoresis-grade agarose and low-melting-point agarose were purchased from Gibco BRL(Life Technologies Italia, San Giuliano Milanese, Milan, Italy); NuSieve wasobtained from FMC BioProducts (Rockland, Maine); and agarose D-5, used forpulsed-field gel electrophoresis (PFGE), was obtained from Hispanagar (Burgos,Spain). Proteinase K and lysozyme were obtained from Roche. All other re-agents and chemicals were purchased from Sigma Chemical Co. (St. Louis, Mo.).The media for bacterial growth were obtained from Oxoid (Basingstoke, UnitedKingdom).
PCR-RFLP analysis. ETBF strains were processed for PCR amplification aspreviously described (24). Briefly, the bacterial cells were boiled for 10 min,centrifuged at 13,000 3 g for 5 min, and stored at 220°C until used as templates(2 ml in each PCR). All PCRs were performed with a GeneAmp PCR System9600 (Perkin-Elmer) and a reaction volume of 100 ml, containing a 0.5 mM finalconcentration of each oligonucleotide, 200 mM each deoxynucleoside triphos-phate, and 2.5 U of DynaZyme II Taq polymerase. Each amplification waspreceded by 5 min at 94°C and consisted of 35 cycles of 60 s at 94°C, 60 s at 56°C,and 120 s at 72°C, followed by a final 5 min at 72°C.
For PCR-RFLP, the primer pair used, BF5-BF6 (Table 1), was designed toamplify a 976-bp internal fragment of the three isoforms of the enterotoxin gene.The PCR amplification products were purified using a QIAquick PCR Purifica-tion Kit (QIAGEN GmbH, Hilden, Germany) and then digested with the re-
striction enzymes AccI and DraI according to the manufacturers’ recommenda-tions. These enzymes were chosen because they have different recognition sitesin the isoforms bft-1 and bft-2 or bft-3 and therefore generate segments ofdifferent lengths. The sequence identity between bft-2 and bft-3 in these recog-nition sites does not allow them to be distinguished. According to the publishedsequences, digestion of bft-1 with DraI yields two fragments (640 and 336 bp),and digestion of bft-2 or bft-3 yields three fragments (561, 336, and 79 bp);restriction of bft-1 with AccI yields three fragments (701, 213, and 62 bp), andrestriction of bft-2 or bft-3 yields two fragments (914 and 62 bp). The digestionproducts were separated in a 3% agarose gel (2% NuSieve, 1% agarose) con-taining 0.5 mg of ethidium bromide per ml.
Allele-specific PCR. In order to distinguish between bft-2 and bft-3, we devisedallele-specific PCR assays. For bft-2, the primers used were BF5 and BFT2R(Table 1). BFT2R was designed based on the oligonucleotide sequence specificfor bft-2, mapping in a gene region divergent from both bft-1 and bft-3, accordingto Franco et al. (8). For bft-3, the pair used was BF5-BFT3R (Table 1); BFT3Rwas designed in the same position as BFT2R but with a 2-base substitution at the39 end, according to the sequence of bft-3 (5). The PCR conditions used were thesame as those described for PCR-RFLP analysis.
Sequencing of the bft gene. The portion of the enterotoxin gene correspondingto the whole mature moiety was amplified using the primer pair BFTF-BFTR.These primers were designed based on consensus regions for the three isoforms(Table 1).
Sequencing reactions were performed with the PCR products as templates andwith a Perkin-Elmer ABI 370A DNA Sequencer and an ABI Prism Dye Termi-nator Cycle Sequencing Ready Reaction Kit (PE Applied Biosystems).
DNA extraction and Southern hybridization. B. fragilis cells were pelletedfrom 5 ml of an overnight culture in Wilkins-Chalgren broth. ChromosomalDNA was extracted as follows. Lysis was performed with 50 mM Tris–50 mMEDTA buffer (pH 8.0) containing 50 mM glucose, 5 mg of lysozyme per ml, 40mg of proteinase K per ml, and 10 mg of RNase A per ml at 37°C for 30 min. TheDNA was extracted three times with phenol-chloroform, the aqueous phase wascollected, and the DNA was recovered by ethanol precipitation and resuspendedin water. The DNA was digested with restriction enzymes according to themanufacturers’ recommendations, run on an 0.8% agarose gel at 30 V for 20 h,and transferred to a nylon membrane by capillary blotting.
Two probes were used; both were generated by amplification of the chromo-somal DNA of VPI 13784. The bft probe is the product of the primer pairBF5-BF6; the MP II probe is the product of the primer pair MP1-MP2 andcorresponds to a 1,050-bp internal fragment of the MP II gene. Probe labelingand filter hybridization were performed using a nonradioactive method based onenhanced chemiluminescence (Amersham).
PFGE. For the preparation of genomic DNA suitable for PFGE, ETBF strainswere grown in 10 ml of Wilkins-Chalgren broth to late log phase (optical densityat 600 nm, 0.8 to 0.9). The bacterial cells were placed in agarose plugs and lysedby standard procedures (13).
Digestion of the DNA-containing agarose plugs was performed with the re-striction enzyme NotI (1) by use of a 150-ml reaction mixture containing bovineserum albumin (100 mg/ml), NotI (20 U), and the buffer provided by the man-ufacturer; the mixture was incubated at 37°C overnight.
The restricted chromosomal DNA was separated on 1% agarose gels using theCHEF Mapper Pulsed-Field Electrophoresis System (Bio-Rad Laboratories,Milan, Italy). Electrophoresis was performed in the two-state mode with a 120°pulse angle at 5.4 V/cm for 34 h. The switch times were increased from 1 to 40 sby the ramping factor 0.357. Following electrophoresis, the gels were stained withethidium bromide, and the DNA bands were photographed under UV light. ForSouthern hybridization, DNA was transferred to nylon membranes by vacuumblotting, and hybridization was performed as described above.
RESULTS
Detection of the enterotoxin gene alleles. With the primerpair BF5-BF6, the expected product of 976 bp was amplifiedfrom all of the ETBF strains examined. The nonenterotoxi-genic strain NCTC 9343 yielded no amplification product (datanot shown).
By PCR-RFLP, we were able to classify the enterotoxingenes of all the ETBF strains examined as belonging to isoformbft-1 or isoform bft-2 or bft-3, as the observed sizes of thefragments generated agreed with the predicted sizes of thefragments and no anomalous profile was observed (Fig. 1).Strain VPI 13784 was found to harbor the isoform bft-1, asexpected, while strain ATCC 43858 was found to harbor theisoform bft-2; these findings were subsequently confirmed bysequencing. These two strains were therefore used as proto-types of the two isoforms throughout the study.
To confirm the results of PCR-RFLP, the portion of the bft
TABLE 1. Oligonucleotide primers used to amplifythe bft and MP II genes
Primer DNA sequence Accession no.a
BF5 59-GATGCTCCAGTTACAGCTTCCATTG-39 U67735BF6 59-CGCCCAGTATATGACCTAGTTCGTG-39 U67735MP1 59-CAGCATGTGCTGACGATCTT-39 AF038459MP2 59-ATCCACATGTTCCGCTCCTA-39 AF038459BFTF 59-CCTCAAGTACCTCATGGAAT-39 U67735BFTR 59-ATTCCATTAATCGAACTTCG-39 U67735BFT2R 59-TTGGATCATCCGCATGCCT-39 U90931BFT3R 59-TTGGATCATCCGCATGGTT-39 AF081785
a Primers were derived from the published sequences in the GeneBank data-base under the indicated accession numbers.
608 SCOTTO D’ABUSCO ET AL. J. CLIN. MICROBIOL.
on March 29, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

gene corresponding to the mature toxin of 14 ETBF isolates, 9with bft-1 and 5 with bft-2 or bft-3, was analyzed by sequencing.The nucleotide sequence of the bft gene of 13 strains showedcomplete identity with the published sequence of either bft-1 orbft-2, and the isoforms deduced by sequencing were in accor-dance with the PCR-RFLP results. However, strain MT 2showed a number of divergences from the sequences of bothisoform bft-1 and isoform bft-2, while it showed 100% identitywith the sequence of bft-3.
In order to verify whether in our series other strains har-bored bft-3, using an assay simpler than sequencing, we devisedPCR assays specific for bft-2 and bft-3. In the bft-2-specificPCR, strain ATCC 43858 yielded the expected amplificationproduct, while neither VPI 13784 (bft-1) nor MT 2 (bft-3)yielded any amplification product. With the bft-3-specific PCR,MT 2 yielded the expected amplification product (Fig. 2). Allthe strains classified as bft-2 or bft-3 by PCR-RFLP were sub-mitted to the bft-2-specific PCR. Besides MT 2, another strain,UK 5312, was negative in the amplification reaction for bft-2
but yielded an amplification product in the reaction for bft-3.Sequencing analysis confirmed these findings.
Distribution of the enterotoxin gene isoforms. Among theETBF isolates examined, bft-1 was found to be more commonthan bft-2, as it was detected in two-thirds of the strains. Thebft-1 isoform was found in almost all isolates from the feces ofadults, including individuals with and without diarrhea; only 1strain out of a total of 17 was found to harbor the bft-2 isoform.However, the bft-2 isoform was as common as the bft-1 isoformin strains isolated from the feces of children. The associationbetween isoform bft-2 and strains from children is statisticallysignificant compared to that between bft-2 and adult strains (P,0.0064) (Table 2). When strains isolated from children withdiarrhea (nine strains bearing bft-1 and nine bearing bft-2) arecompared to strains isolated from adults with diarrhea (ninestrains bearing bft-1 and one bearing bft-2), the association isstatistically significant (P determined by the Fisher exact test,0.04). We found that the bft-3 isoform, originally described forisolates from Korea and Japan, also was present in our collec-tion of strains originating from Europe and the United States,although much less frequently than the other two isoforms.Strain MT 2 was isolated from the stools of a healthy 4-month-old Italian baby in 1994; strain UK 5312 was originally isolatedfrom a blood culture in the United Kingdom.
Southern analysis of the bft and MP II genes. We analyzed16 ETBF strains by Southern hybridization: 10 harboring bft-1,4 harboring bft-2, and 2 harboring bft-3 (Table 3). The intentwas to hybridize the chromosomal DNA with probes for the bftand MP II genes to define whether these two genes were con-sistently associated in all strains, irrespective of the isoformborne. Prior to the experiments, controls were run to verifythat the two probes were specific for the respective genes andthat there was no cross-hybridization.
The chromosomal DNA was digested with the restrictionenzyme EcoRI. Since this enzyme has no recognition site in thebft gene, in the majority of the strains the bft probe hybridizedwith a single band, as expected. This band had a different mo-lecular size in each strain, ranging from approximately 9 to 25kb. However, in five strains, the bft probe hybridized with twobands (Fig. 3A). As the presence of an additional EcoRI siteinternal to the bft gene was ruled out by analysis of the bftsequences, this finding was suggestive of the presence of dou-ble copies of the gene. In the same strains, two hybridizingbands also were obtained when the chromosomal DNA wasdigested with BamHI and SalI, which have no restriction sitesthe bft gene (data not shown).
According to the published sequence of the fragilysin islet,EcoRI has a single recognition site in the MP II gene and asecond site between the MP II gene and the left end of the islet(16). As expected, in the majority of the strains, the MP IIprobe hybridized with two bands. One band, with a high mo-
FIG. 1. Agarose gel electrophoresis of the bft gene PCR products of ETBFstrains and of the RFLPs obtained with DraI or AccI digestion. Lanes: 1, strainVPI 13784; 2, ATCC 43858; 3, PR 838; 4, MT 2. U, unrestricted PCR products;D, PCR products digested with DraI; A, PCR products digested with AccI. M,molecular size markers. In accordance with the fragment lengths indicated in thetext, the isoforms bft-1 and bft-2 or bft-3 can be distinguished in lanes 1 and 3 andin lanes 2 and 4, respectively. The lower-molecular-size fragments are not visible.By sequence analysis and isoform-specific PCR, ATCC 43858 was shown toharbor bft-2 and MT 2 was shown to harbor bft-3.
FIG. 2. Agarose gel electrophoresis of the amplification products obtainedwith allele-specific PCR for the isoforms bft-2 and bft-3. Lanes: 1, VPI 13784; 2,ATCC 43858; 3, B 115; 4, MT 2; 5, UK 5312. M, molecular size markers.
TABLE 2. Distribution of enterotoxin gene isoforms (bft-1 or bft-2)according to the origin of the ETBF isolates
Source No. of isolatestested
No. of isolates withthe following entero-
toxin gene allele:
bft-1 bft-2a
Extraintestinal site 27 17 10Feces
Children 22 12 10*Adults 17 16 1**
Total 66 45 21
a * versus **, chi-square test value, 7.42; P, 0.0064.
VOL. 38, 2000 ALLELES OF THE bft GENE 609
on March 29, 2018 by guest
http://jcm.asm
.org/D
ownloaded from
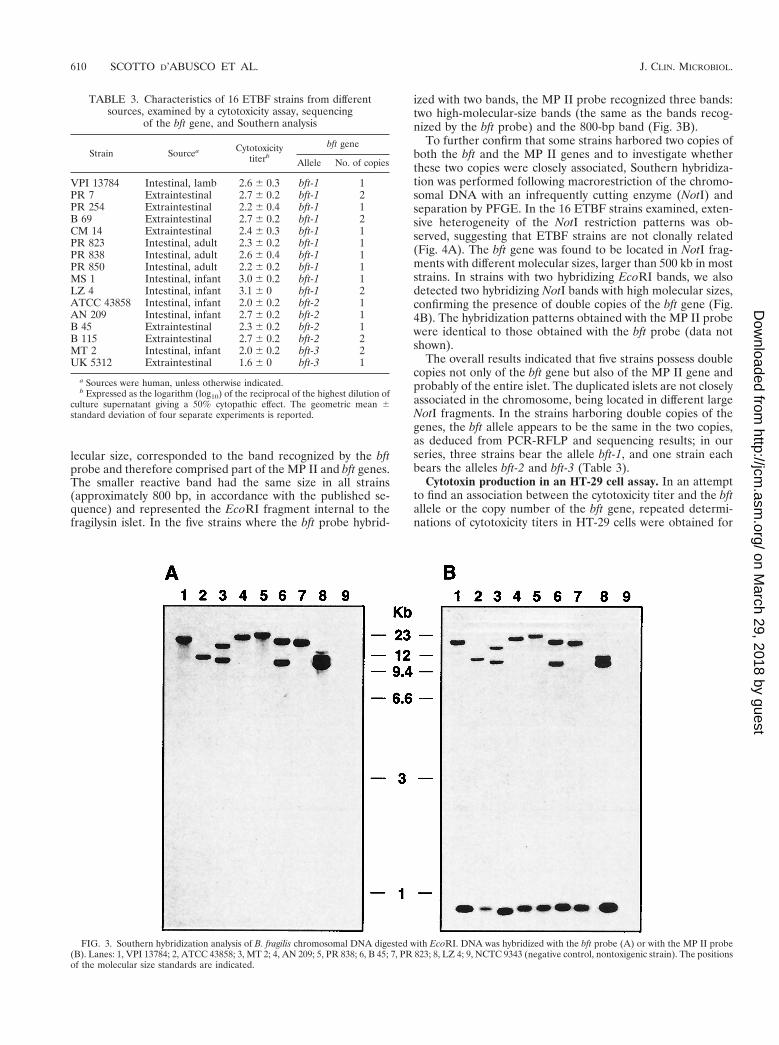
lecular size, corresponded to the band recognized by the bftprobe and therefore comprised part of the MP II and bft genes.The smaller reactive band had the same size in all strains(approximately 800 bp, in accordance with the published se-quence) and represented the EcoRI fragment internal to thefragilysin islet. In the five strains where the bft probe hybrid-
ized with two bands, the MP II probe recognized three bands:two high-molecular-size bands (the same as the bands recog-nized by the bft probe) and the 800-bp band (Fig. 3B).
To further confirm that some strains harbored two copies ofboth the bft and the MP II genes and to investigate whetherthese two copies were closely associated, Southern hybridiza-tion was performed following macrorestriction of the chromo-somal DNA with an infrequently cutting enzyme (NotI) andseparation by PFGE. In the 16 ETBF strains examined, exten-sive heterogeneity of the NotI restriction patterns was ob-served, suggesting that ETBF strains are not clonally related(Fig. 4A). The bft gene was found to be located in NotI frag-ments with different molecular sizes, larger than 500 kb in moststrains. In strains with two hybridizing EcoRI bands, we alsodetected two hybridizing NotI bands with high molecular sizes,confirming the presence of double copies of the bft gene (Fig.4B). The hybridization patterns obtained with the MP II probewere identical to those obtained with the bft probe (data notshown).
The overall results indicated that five strains possess doublecopies not only of the bft gene but also of the MP II gene andprobably of the entire islet. The duplicated islets are not closelyassociated in the chromosome, being located in different largeNotI fragments. In the strains harboring double copies of thegenes, the bft allele appears to be the same in the two copies,as deduced from PCR-RFLP and sequencing results; in ourseries, three strains bear the allele bft-1, and one strain eachbears the alleles bft-2 and bft-3 (Table 3).
Cytotoxin production in an HT-29 cell assay. In an attemptto find an association between the cytotoxicity titer and the bftallele or the copy number of the bft gene, repeated determi-nations of cytotoxicity titers in HT-29 cells were obtained for
TABLE 3. Characteristics of 16 ETBF strains from differentsources, examined by a cytotoxicity assay, sequencing
of the bft gene, and Southern analysis
Strain Sourcea Cytotoxicitytiterb
bft gene
Allele No. of copies
VPI 13784 Intestinal, lamb 2.6 6 0.3 bft-1 1PR 7 Extraintestinal 2.7 6 0.2 bft-1 2PR 254 Extraintestinal 2.2 6 0.4 bft-1 1B 69 Extraintestinal 2.7 6 0.2 bft-1 2CM 14 Extraintestinal 2.4 6 0.3 bft-1 1PR 823 Intestinal, adult 2.3 6 0.2 bft-1 1PR 838 Intestinal, adult 2.6 6 0.4 bft-1 1PR 850 Intestinal, adult 2.2 6 0.2 bft-1 1MS 1 Intestinal, infant 3.0 6 0.2 bft-1 1LZ 4 Intestinal, infant 3.1 6 0 bft-1 2ATCC 43858 Intestinal, infant 2.0 6 0.2 bft-2 1AN 209 Intestinal, infant 2.7 6 0.2 bft-2 1B 45 Extraintestinal 2.3 6 0.2 bft-2 1B 115 Extraintestinal 2.7 6 0.2 bft-2 2MT 2 Intestinal, infant 2.0 6 0.2 bft-3 2UK 5312 Extraintestinal 1.6 6 0 bft-3 1
a Sources were human, unless otherwise indicated.b Expressed as the logarithm (log10) of the reciprocal of the highest dilution of
culture supernatant giving a 50% cytopathic effect. The geometric mean 6standard deviation of four separate experiments is reported.
FIG. 3. Southern hybridization analysis of B. fragilis chromosomal DNA digested with EcoRI. DNA was hybridized with the bft probe (A) or with the MP II probe(B). Lanes: 1, VPI 13784; 2, ATCC 43858; 3, MT 2; 4, AN 209; 5, PR 838; 6, B 45; 7, PR 823; 8, LZ 4; 9, NCTC 9343 (negative control, nontoxigenic strain). The positionsof the molecular size standards are indicated.
610 SCOTTO D’ABUSCO ET AL. J. CLIN. MICROBIOL.
on March 29, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

16 ETBF strains. In strains bearing either bft-1 or bft-2, thetiters varied from 2 to 3 log10 units. Both strains bearing bft-3showed low toxin titers (1.6 and 2 log10 units). Four out of fivestrains harboring a duplication of the bft gene (three strainsbearing bft-1 and one strain bearing bft-2) were found to havecytotoxicity titers in the high range of the series. The onlyexception was the strain bearing double copies of bft-3, whichhad a low toxin titer (Table 3). Due to the limited number ofstrains examined, it was not possible to demonstrate statisti-cally significant differences.
DISCUSSION
Although ETBF strains have been associated with diarrheain children under 5 years of age (27, 28, 30), the pathophysi-ological mechanisms linking enterotoxin production to diar-rhea are not completely understood. B. fragilis toxin, or fragi-lysin, cannot be considered a classical enterotoxin, being ametalloprotease similar to eukaryotic collagenases (15). Invitro and in vivo studies have shown that this toxin is able todamage the intestinal mucosa (both ileal and colonic) of var-ious animal species, including humans (21, 26, 29), and to elicitfluid accumulation through increased permeability (26) andactive chloride secretion (4). Recently, the zonula adherensprotein E-cadherin has been recognized as the substrate forB. fragilis enterotoxin. It has been hypothesized that cleavageof the extracellular domain of E-cadherin can lead to alter-ation of the cytoskeletal structures and to increased intestinalpermeability. This hypothesis represents a novel mechanism ofaction for a bacterial enterotoxin (32).
The presence of ETBF in the gut does not necessarily indi-cate disease, as the organism has been frequently isolated fromthe feces of healthy individuals. Asymptomatic carriage is par-ticularly high in adults, but children also can harbor both themicroorganism and the toxin without intestinal disturbances(25). This finding suggests that other factors that can be relatedto either the host or the microorganism are necessary for thedevelopment of diarrhea. A recent study has shown unrespon-siveness of the colon mucosa of some subjects to B. fragilistoxin (29).
ETBF can produce different isoforms of the enterotoxin, themost common being those encoded by the bft-1 and bft-2 genes(8, 12). Although these isoforms display the same biological ac-tivities, Wu and coworkers have suggested that their potenciesmight be different (32). We examined several ETBF strainsisolated from different sources (extraintestinal infections andfeces of adults and children) by PCR-RFLP analysis to distin-guish between the isoforms bft-1 and bft-2 or bft-3 and subse-quently by an isoform-specific PCR to distinguish betweenbft-2 and bft-3. We found that the majority (65%) of the strainsinvestigated harbor the bft-1 isoform. This allele is largely pre-dominant in strains from the feces of adults (with or withoutintestinal symptoms), while strains isolated from children har-bor either bft-1 or bft-2. The rarity of the isoform bft-2 in adultsindicates that strains bearing this isoform are more apt to col-onize (and consequently induce diarrhea in) children than adults.The factors responsible for this association are unknown, butthey might consist either of different properties of the bft-2toxin itself or of characteristics of the strains which allow betterproliferation in the colon of children than in that of adults.
By sequence analysis and an allele-specific PCR, we foundthat two strains in our collection harbor the isoform bft-3,originally described for blood isolates from Korea (5) and forfecal isolates from Japan (Kato, Final Program of the 2ndWorld Congress on Anaerobic Bacteria and Infections). Thetwo strains in our collection were isolated in the United King-dom and in Italy; this finding indicates that the bft-3 allele, al-though rare, is present in geographical areas outside East Asia.
Interestingly, the sequences of the bft genes examined were100% identical to the published nucleotide sequences of thethree alleles, without a single base substitution. Although thisobservation is limited to the portion of the genes coding for themature toxin, this lack of variation suggests that bft is a recentacquisition of B. fragilis, as already proposed by Smith andCallihan (31), and that each isoform is conserved because of anevolutionary advantage. The amino acid substitutions amongthe three deduced proteins are not abundant: bft-2 toxin di-verges from bft-1 toxin in 25 amino acids out of 186, and bft-3(which is more similar to bft-2 than to bft-1) diverges from bft-1in 20 amino acids and from bft-2 in 8 amino acids. However,these substitutions cluster in two regions adjacent to the activesite of the metalloprotease and the zinc-binding motif (8).Therefore, the three variants could exhibit subtly different re-ceptor-binding preferences that could result in differences inhost range and/or pathogenic potential. Similar differenceshave been described for other allelic proteins. For instance, thealleles speA1 and speA3 of Streptococcus pyogenes code fortoxins which differ only in one amino acid; however, the prod-uct of speA3 has significantly greater mitogenic activity andaffinity for the class II major histocompatibility complex and isassociated with clinical cases of streptococcal toxic shock syn-drome (11). The papG alleles of Escherichia coli, which codefor variant forms of the P adhesin, are associated with differentclinical syndromes, such as pyelonephritis and cystitis (10).
In our collection, the two strains carrying bft-3 were found toproduce low levels of biologically active toxin in the HT-29 cellcytotoxicity assay. One explanation is that a smaller amount ofthe protein is produced. An alternative possibility is that thebft-3 toxin is less active on the cell system used, although Chunget al. have demonstrated that the purified bft-3 toxin fromKorean isolates cleaves E-cadherin at a concentration similarto that observed with the other purified enterotoxins (5).
We cannot rule out the possibility that other enterotoxinvariants with substitutions in areas not explored by the RFLPand PCR assays used in this study exist. Sequencing of morestrains from different sources and geographical regions is nec-
FIG. 4. PFGE restriction patterns generated by NotI digestion (A) andSouthern hybridization with the bft probe (B) of ETBF chromosomal DNA.Lanes: 1, VPI 13784; 2, ATCC 43858; 3, MT 2; 4, AN 209; 5, PR 838; 6, B115;7, PR 823; 8, B 69; 9, LZ 4. Lanes M1 and M2, molecular size markers. Thehybridization pattern obtained with the MP II probe was identical to that ob-tained with the bft probe.
VOL. 38, 2000 ALLELES OF THE bft GENE 611
on March 29, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

essary for a comprehensive analysis of the frequency of thedifferent enterotoxin alleles.
Following the recent discovery of the pathogenicity islet ofB. fragilis, which includes the fragilysin gene, B. fragilis hasbeen added to the growing list of microorganisms carryingpathogenicity islands. This islet is much smaller than classicalpathogenicity islands. It comprises only the enterotoxin geneand the MP II gene and lacks the genes coding for secretionsystems necessary for the delivery of the toxin directly to targetcells (14). However, the B. fragilis islet shares with the largerstructures the property of transforming a typical commensalorganism into a virulent organism (9).
An unexpected finding was that some ETBF strains possessdouble copies of the enterotoxin gene, associated with doublecopies of the MP II gene and possibly of the entire islet.Bacteria harboring more than one pathogenicity island havebeen described before: for instance, in the same strain of uro-pathogenic E. coli serotype O4, PAI I and PAI II are presenttogether. However, the virulence genes carried by the two is-lands are different (2). In some Helicobacter pylori type I strains,the cag pathogenicity island appears as two regions separatedby a long stretch of chromosomal DNA, containing differentgenes and probably derived from rearrangement driven by in-sertion sequences (3).
For B. fragilis, the whole islet, including the enterotoxin andMP II genes, appears to have undergone duplication in somestrains, as deduced from Southern analysis; consequently, thetwo islets carry the same enterotoxin gene isoform, as con-firmed by PCR-RFLP and sequencing. The islet duplicationappears to be quite stable, as the Southern hybridization pat-terns were reproducible when the strains were examined afterstorage and repeated subculturing. Interestingly, with the ex-ception of the bft-3 strain, the cytotoxicity titers obtained fromsupernatants of strains with double copies of the gene wereamong the highest in our series; however, the limited numberof strains examined does not allow conclusions to be drawn.
Our observations indicate that there are complex microbio-logical properties of ETBF that need to be elucidated for abetter understanding of the pathogenic potential of this micro-organism.
ACKNOWLEDGMENTS
We are grateful to Alessandra Carattoli and Alfredo Caprioli forhelpful and stimulating discussions; to Maria Grazia Menozzi andMonica Malpeli for providing recent ETBF isolates; and to FabioD’Ambrosio and Patrizia Chinzari for experienced technical assis-tance.
This work was funded in part by Consiglio Nazionale delle Ricerche,Rome, Italy (grants 96.03301.CT04 and 98.00493.CT04).
REFERENCES1. Bedzyk, L. A., N. B. Shoemaker, K. E. Young, and A. A. Salyers. 1992.
Insertion and excision of Bacteroides conjugative chromosomal elements.J. Bacteriol. 174:166–172.
2. Blum, G., V. Falbo, A. Caprioli, and K. Hacker. 1995. Gene clusters encodingthe cytotoxic necrotizing factor type 1, Prs-fimbriae and a-hemolysin formthe pathogenicity island II of the uropathogenic Escherichia coli strain J96.FEMS Microbiol. Lett. 126:189–196.
3. Censini, S., C. Lange, Z. Xiang, J. E. Crabtree, P. Ghiara, M. Borodovsky,R. Rappuoli, and A. Covacci. 1996. cag, a pathogenicity island of Helicobacterpylori, encodes type I-specific and disease-associated virulence factors. Proc.Natl. Acad. Sci. USA 93:14648–14653.
4. Chambers, F. G., S. S. Koshy, R. F. Saidi, D. P. Clark, R. D. Moore, and C. L.Sears. 1997. Bacteroides fragilis toxin exhibits polar activity on monolayers ofhuman intestinal epithelial cells (T84 cells) in vitro. Infect. Immun. 65:3561–3570.
5. Chung, G. T., A. A. Franco, S. Wu, G. E. Rhie, R. Cheng, H. B. Oh, and C. L.Sears. 1999. Identification of a third metalloprotease toxin gene in extraint-estinal isolates of Bacteroides fragilis. Infect. Immun. 67:4945–4949.
6. Duerden, B. I., and B. S. Drasar. 1991. Anaerobes in human disease. EdwardArnold, London, England.
7. Finegold, S. M., V. L. Sutter, and G. E. Mathisen. 1983. Normal indigenousintestinal flora, p. 3–31. In D. J. Hentges (ed.), Human intestinal microflorain health and disease. Academic Press, Inc., New York, N.Y.
8. Franco, A. A., L. M. Mundy, M. Trucksis, S. Wu, J. B. Kaper, and C. L.Sears. 1997. Cloning and characterization of the Bacteroides fragilis metal-loprotease gene. Infect. Immun. 65:1007–1013.
9. Groisman, E. A., and H. Ochman. 1996. Pathogenicity islands: bacterialevolution in quantum leaps. Cell 87:791–794.
10. Johnson, J. R., T. A. Russo, J. J. Brown, and A. Stapleton. 1998. papG allelesof Escherichia coli strains causing first-episode or recurrent acute cystitis inadult women. J. Infect. Dis. 177:97–101.
11. Kline, J. B., and C. M. Collins. 1996. Analysis of the superantigenic activityof mutant and allelic forms of streptococcal pyrogenic exotoxin A. Infect.Immun. 64:861–869.
12. Kling, J. J., R. L. Wright, J. S. Moncrief, and T. D. Wilkins. 1997. Cloningand characterization of the gene for the metalloprotease enterotoxin ofBacteroides fragilis. FEMS Microbiol. Lett. 146:279–284.
13. Maslow, J. N., A. M. Slutsky, and A. R. D. Arbeit. 1993. Application ofpulsed-field gel electrophoresis to molecular epidemiology, p. 563–572. InD. H. Persing, T. F. Smith, F. C. Tenover, and T. J. White (ed.), Diagnosticmolecular microbiology: principles and applications. American Society forMicrobiology, Washington, D.C.
14. Mecsas, J., and E. J. Strauss. 1996. Molecular mechanisms of bacterialvirulence: type III secretion and pathogenicity islands. Emerg. Infect. Dis. 2:271–285.
15. Moncrief, J. S., R. Obiso, L. A. Barroso, J. J. Kling, R. H. Wright, R. L. VanTassell, D. M. Lyerly, and T. D. Wilkins. 1995. The enterotoxin of Bacte-roides fragilis is a metalloprotease. Infect. Immun. 63:175–181.
16. Moncrief, J. S., A. J. Duncan, R. L. Wright, L. A. Barroso, and T. D. Wilkins.1998. Molecular characterization of the fragilysin pathogenicity islet of en-terotoxigenic Bacteroides fragilis. Infect. Immun. 66:1735–1739.
17. Myers, L. L., B. D. Firehammer, D. S. Shoop, and M. M. Border. 1984.Bacteroides fragilis: a possible cause of acute diarrheal disease in newbornlambs. Infect. Immun. 44:241–244.
18. Myers, L. L., D. S. Shoop, and T. D. Byars. 1987. Diarrhea associated withenterotoxigenic Bacteroides fragilis in foals. Am. J. Vet. Res. 48:1565–1567.
19. Myers, L. L., D. S. Shoop, B. D. Firehammer, and M. M. Border. 1985.Association of enterotoxigenic Bacteroides fragilis with diarrheal disease incalves. J. Infect. Dis. 152:1344–1347.
20. Obiso, R. J., A. O. Azghani, and T. D. Wilkins. 1997. The Bacteroides fragilistoxin fragilysin disrupts the paracellular barrier of epithelial cells. Infect.Immun. 65:1431–1439.
21. Obiso, R. J., D. M. Lyerly, R. L. Van Tassell, and T. D. Wilkins. 1995.Proteolytic activity of the Bacteroides fragilis enterotoxin causes fluid secre-tion and intestinal damage in vivo. Infect. Immun. 63:3820–3826.
22. Pantosti, A., M. Cerquetti, R. Colangeli, and F. D’Ambrosio. 1994. Detectionof intestinal and extra-intestinal strains of enterotoxigenic Bacteroides fragilisby the HT-29 cytotoxicity assay. J. Med. Microbiol. 41:191–196.
23. Pantosti, A., R. Colangeli, A. O. Tzianabos, and D. L. Kasper. 1995. Mono-clonal antibodies to detect capsular diversity among Bacteroides fragilis iso-lates. J. Clin. Microbiol. 33:2647–2652.
24. Pantosti, A., M. Malpeli, M. Wilkins, M. G. Menozzi, and F. D’Ambrosio.1997. Detection of enterotoxigenic Bacteroides fragilis by using PCR. J. Clin.Microbiol. 35:2482–2486.
25. Pantosti, A., M. G. Menozzi, A. Frate, L. Sanfilippo, F. D’Ambrosio, and M.Malpeli. 1997. Detection of enterotoxigenic Bacteroides fragilis and its toxinin stool samples from adults and children in Italy. Clin. Infect. Dis. 24:12–16.
26. Riegler, M., M. Lotz, C. Sears, C. Pothoulakis, I. Castagliuolo, C. C. Wang,R. Sedivy, T. Sogukoglu, E. Cosentini, G. Bischof, W. Feil, B. Teleky, G.Hamilton, J. T. LaMont, and E. Wenzl. 1999. Bacteroides fragilis toxin 2damages human colonic mucosa in vitro. Gut 44:504–510.
27. Sack, R. B., M. J. Albert, K. Alam, P. K. B. Neogi, and M. S. Akbar. 1994.Isolation of enterotoxigenic Bacteroides fragilis from Bangladeshi childrenwith diarrhea: a controlled study. J. Clin. Microbiol. 32:960–963.
28. Sack, R. B., L. L. Myers, J. Almeido-Hill, D. S. Shoop, W. C. Bradbury, R.Reid, and M. Santosham. 1992. Enterotoxigenic Bacteroides fragilis: epide-miologic studies of its role as a human diarrhoeal pathogen. J. DiarrhoealDis. Res. 10:4–9.
29. Sanfilippo, L., T. J. Baldwin, M. G. Menozzi, S. P. Borriello, and Y. R.Mahida. 1998. Heterogeneity in responses by primary adult human colonicepithelial cells to purified enterotoxin of Bacteroides fragilis. Gut 43:651–655.
30. San Joaquin, V. H., J. C. Griffis, L. Christopher, and C. L. Sears. 1995.Association of Bacteroides fragilis with childhood diarrhea. Scand. J. Infect.Dis. 27:211–215.
31. Smith, C. J., and C. J. Callihan. 1992. Analysis of rRNA restriction fragmentlength polymorphisms from Bacteroides spp. and Bacteroides fragilis isolatesassociated with diarrhea in humans and animals. J. Clin. Microbiol. 30:806–812.
32. Wu, S., K.-C. Lim, J. Huang, R. F. Saidi, and C. L. Sears. 1998. Bacteroidesfragilis enterotoxin cleaves the zonula adherens protein, E-cadherin. Proc.Natl. Acad. Sci. USA 95:14979–14984.
612 SCOTTO D’ABUSCO ET AL. J. CLIN. MICROBIOL.
on March 29, 2018 by guest
http://jcm.asm
.org/D
ownloaded from