the 3d arrangement of the 23 s and 5 s rrna in the escherichia coli 50 s ribosomal subunit based on...
TRANSCRIPT

doi:10.1006/jmbi.2000.3635 available online at http://www.idealibrary.com on J. Mol. Biol. (2000) 298, 35±59
The 3D Arrangement of the 23 S and 5 S rRNA in theEscherichia coli 50 S Ribosomal Subunit Based on aCryo-electron Microscopic Reconstruction at7.5 AÊ Resolution
Florian Mueller1, Ingolf Sommer1, Pavel Baranov2, Rishi Matadeen3
Matthias Stoldt4, Jens WoÈ hnert4, Matthias GoÈ rlach4, Marin van Heel3
and Richard Brimacombe1*
1Max-Planck-Institut fuÈ rMolekulare Genetik, Ihnestrasse73, 14195 Berlin, Germany2A.N. Belozerksy Institute ofPhysico-Chemical Biology &Department of ChemistryMoscow State UniversityMoscow, 119899, Russia3Imperial College of ScienceMedicine and TechnologyDepartment of BiochemistryLondon, SW7 2AY, UK4Abteilung MolekulareBiophysik/NMR-SpektroskopieInstitut fuÈ r Molekulare Bio-technologie, Postfach 10081307708, Jena, Germany
Present address: P. Baranov, EcclHuman Genetics, University of UtaUT 84112, USA.
Abbreviations used: EM, electronimmuno-EM; PIP, principal interfac
0022-2836/00/010035±25 $35.00/0
The Escherichia coli 23 S and 5 S rRNA molecules have been ®tted helixby helix to a cryo-electron microscopic (EM) reconstruction of the 50 Sribosomal subunit, using an un®ltered version of the recently published50 S reconstruction at 7.5 AÊ resolution. At this resolution, the EM densityshows a well-de®ned network of ®ne structural elements, in which themajor and minor grooves of the rRNA helices can be discerned at manylocations. The 3D folding of the rRNA molecules within this EM densityis constrained by their well-established secondary structures, and furtherconstraints are provided by intra and inter-rRNA crosslinking data, aswell as by tertiary interactions and pseudoknots. RNA-protein cross-linkand foot-print sites on the 23 S and 5 S rRNA were used to position therRNA elements concerned in relation to the known arrangement of theribosomal proteins as determined by immuno-electron microscopy. Thepublished X-ray or NMR structures of seven 50 S ribosomal proteins orRNA-protein complexes were incorporated into the EM density. The 3Dlocations of cross-link and foot-print sites to the 23 S rRNA from tRNAbound to the ribosomal A, P or E sites were correlated with the positionsof the tRNA molecules directly observed in earlier reconstructions of the70 S ribosome at 13 AÊ or 20 AÊ . Similarly, the positions of cross-link siteswithin the peptidyl transferase ring of the 23 S rRNA from the aminoacylresidue of tRNA were correlated with the locations of the CCA ends ofthe A and P site tRNA. Sites on the 23 S rRNA that are cross-linked tothe N termini of peptides of different lengths were all found to lie withinor close to the internal tunnel connecting the peptidyl transferase regionwith the presumed peptide exit site on the solvent side of the 50 S sub-unit. The post-transcriptionally modi®ed bases in the 23 S rRNA form acluster close to the peptidyl transferase area. The minimum conservedcore elements of the secondary structure of the 23 S rRNA form a com-pact block within the 3D structure and, conversely, the points corre-sponding to the locations of expansion segments in 28 S rRNA all lie onthe outside of the structure.
# 2000 Academic Press
Keywords: cryo-electron microscopy; computer modelling; cross-links andfoot-prints; functional sites on rRNA; atomic structures of ribosomalcomponents
*Corresponding authores Institute ofh, Salt Lake City,
microscopy; IEM,e protrusion.
Introduction
The ribosome is a highly complex macromolecu-lar machine, and the elucidation of its structure hasbeen the object of intensive research for several
# 2000 Academic Press

36 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
decades. In addition to the determination of theprimary amino acid and nucleotide sequences ofthe many ribosomal protein and rRNA com-ponents, and the derivation of secondary structuresfor the rRNA molecules, a great deal of effort hasbeen devoted to the investigation of the three-dimensional (3D) arrangement of the ribosomalconstituents. Many different physical and bio-chemical approaches have been applied to this pro-blem, including the use of cross-linking, neutronscattering or immuno-electron microscopic (IEM)techniques to determine the spatial distribution ofthe ribosomal proteins, the analysis of interactionsbetween ribosomal proteins and rRNA by a varietyof foot-printing and cross-linking methods, and thestudy of neighbourhoods within or between therRNA molecules by intra and inter-RNA cross-link-ing (e.g. see Brimacombe, 1995; Green & Noller,1997). Similar methodologies have been used toanalyse the interactions between the ribosomalcomponents (proteins and/or rRNA) and func-tional ligands such as tRNA, mRNA or the variousfactors involved in the protein biosynthetic process(e.g. see Brimacombe, 1995; Wilson & Noller,1998a). Most of the available data from theseapproaches have been obtained with ribosomesfrom the eubacterium Escherichia coli, and a num-ber of research groups (e.g. Brimacombe et al.,1988; Nagano et al., 1988; Stern et al., 1988; Mitchellet al., 1990; Malhotra & Harvey, 1994) have madeuse of different combinations of the data sets toderive 3D models for the E. coli 16 S or 23 S rRNAmolecules. However, these models are essentiallyonly ``cartoons'' of the ribosome, and it is abun-dantly clear that on its own this type of molecularmodelling approach could never lead to a structureof the ribosome at atomic resolution.
On a different plane, dramatic advances haverecently been achieved in the study of ribosomestructure by both cryo-electron microscopy (cryo-EM; Agrawal et al., 1996; Stark et al., 1995, 1997a,b;Malhotra et al., 1998; Matadeen et al., 1999) and X-ray crystallography (Ban et al., 1998, 1999; Cateet al., 1999; Clemens et al., 1999; Harms et al., 1999;Tocilj et al., 1999). As a result, electron densitymaps of ribosomes or their subunits are becomingavailable at increasingly high resolution, and it isgenerally assumed that, once the resolution hasreached a suf®cient level, an atomic solution to theribosome structure problem will automatically fol-low. In practice, however, an interpretation of elec-tron density data at the atomic level will beimmensely dif®cult, due to the sheer complexityand total lack of symmetry of the ribosome. Thebacterial 70 S ribosome carries about 55 differentprotein molecules, as well as over 150 elements ofdouble-helical rRNA. Many of the latter will be vir-tually indistinguishable from one another in theelectron density maps, even at quite high resol-ution. Furthermore, it is to be expected that local¯exibilities and/or structural heterogeneities willoccur in many areas of the rRNA molecules, lead-ing to local disorder in the electron density and
hence to apparent discontinuities in the rRNAbackbones. The dif®culty of interpreting the elec-tron density maps is particularly acute in the caseof the large 50 S ribosomal subunit, where there isno obvious correlation between the morphologicalarrangement of the particle and the secondarystructural domains of the 23 S rRNA.
The magnitude of this problem is borne out bythe recent crystallographic studies reported by Banet al. (1999) and Culver et al. (1999). Ban et al.(1999),although they could see double-helical rRNA den-sity in their 5 AÊ map of the 50 S subunit corre-sponding to about 300 base-pairs, have so far beenable to assign only those elements of the rRNA forwhich crystallographically derived atomic struc-tures already exist, totalling only 87 nucleotides or3 % of the 23 S rRNA. Similarly, in the study byCulver et al. (1999) at 7.8 AÊ resolution, only 18nucleotides or 0.6 % of the rRNA were assigned. Incontrast, in the 30 S subunit, where the three-domain morphology of the particle correspondsapproximately to the secondary structural domainsof the 16 S rRNA, Clemens et al. (1999) wereable to assign a substantially higher proportion(ca 25 %) of the rRNA to their electron densityat 5.5 AÊ .
We have been convinced for some time that theonly effective way to solve the ribosome structureproblem is to combine the molecular modellingapproach with the electron density maps obtainedby cryo-EM (or X-ray crystallography) by a processof successive approximation. The ®rst step is tomodify the biochemically-derived ``cartoon''models of the rRNA molecules so that they ®t asaccurately as possible to an appropriate corre-sponding electron density map. As the resolutionof the latter improves, and as more biochemicalcorrelations become known, the rRNA structurescan be re®ned progressively, and errors and ambi-guities eliminated step by step. Crystallographic orNMR structures of individual ribosomal proteinsor of small sections of the rRNA can be incorpor-ated into the overall structure as and when theybecome available. In this way, we published twoyears ago a 3D structure for 16 S rRNA (Mueller &Brimacombe, 1997a,b; Mueller et al., 1997) that was®tted to the 30 S moiety of a cryo-EM reconstruc-tion at 20 AÊ resolution of the 70 S ribosome carry-ing tRNAs at the ribosomal A and P sites (Starket al., 1997a); subsequently, the crystal structure ofribosomal protein S7 was added (Tanaka et al.,1998). Despite the relatively low resolution of thecryo-EM reconstruction, the ®t of the protein struc-ture was suf®ciently accurate to enable us to pre-dict and experimentally con®rm the site on S7 thatis able to be cross-linked to mRNA (Greuer et al.,1999).
A similar cryo-EM reconstruction of E. coli70 S ribosomes carrying N-formylmethionyl-tRNA at the P site together with an EF-Tu/GTP/tRNAPhe ternary complex stalled at the``pre-A'' site with the antibiotic kirromycin (Starket al., 1997b) has recently been re®ned to a

The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 37
resolution of 13 AÊ (H. Stark, M. Rodnina, F.Zemlin, W. Wintermeyer & M. van Heel, unpub-lished results), and now a reconstruction of theE. coli 50 S subunit at signi®cantly higher resol-ution has become available (Matadeen et al.,1999). Here, we describe a 3D structure for thecomplete 23 S and 5 S rRNA molecules, ®tted tothis latter reconstruction. The resolution of the50 S structure published by Matadeen et al.(1999) was 7.5 AÊ . However, we found it moreappropriate (see Materials and Methods) to usean un®ltered version of their reconstruction atslightly lower resolution.
The rRNA fragments and ribosomal proteinsthat were ®tted to the 50 S electron density mapat 5 AÊ by Ban et al. (1999) have been incorpor-ated into our structure, and their list has beenextended to include ribosomal proteins L9(Hoffman et al., 1996) and L25 (Stoldt et al.,1998), the latter in the form of an NMR struc-ture of the protein bound to a fragment of 5 SrRNA (Stoldt et al., 1999). In addition, thepositions of tRNAs bound to the ribosomal A, Pand E sites, as well as of EF-Tu and tRNA atthe pre-A site were interpolated by comparisonwith the 20 AÊ and 13 AÊ cryo-EM reconstructionsmentioned above. This enabled us to make useof the available tRNA foot-printing and cross-linking data on the 23 S rRNA (e.g. see Moazed& Noller, 1989; Steiner et al., 1988; Rinke-Appelet al., 1995) to help re®ne the structure of thelatter. The 3D locations of these tRNA-bindingregions in our structure are described, as well asother regions of functional interest, such as thesites on 23 S rRNA that have been cross-linkedto the N termini of nascent peptides of variouslengths (e.g. see Choi & Brimacombe, 1998) orthe sites of interaction with elongation factors(Moazed et al., 1988; Wilson & Noller, 1998b).We also consider the locations of sites of post-transcriptional modi®cation in the 23 S rRNA,and discuss the conserved central core of the23 S rRNA both in terms of helices that aremissing in ribosomes having a smaller rRNAmolecule (as exempli®ed by the mammalianmitochondrial ribosome), and in terms of thelocations of the expansion segments in mamma-lian 28 S rRNA.
As before (Mueller & Brimacombe, 1997a), therRNA modelling was carried out using the pro-gramme ERNA-3D. In addition, we have nowapplied energy minimization procedures (I.S. &R.B., unpublished results), so as to eliminate vander Waals con¯icts within the structure and tooptimize the conformations of those single-stranded regions that contain inter or intra-RNAcross-links. The complete network of structures,including 23 S rRNA, 5 S rRNA and the ®ttedribosomal proteins, has been deposited with theRCSB Protein Data Bank (see Materials andMethods).
Results and Discussion
Biochemical data relating to the 5 S and23 S rRNA
The secondary structures of the E. coli 5 S and23 S rRNA molecules, together with the biochemi-cal data relevant to the modelling process, are illus-trated in Figure 1. These data consist of bothpurely structural information, such as intra andinter-rRNA cross-links or RNA-protein cross-linksand foot-prints, and of more functional infor-mation, such as cross-link and foot-print sites totRNA or cross-links from the nascent peptidechain. The various data sets, together with theappropriate literature references, are available as aseries of Tables in the Supplementary Material.(The Supplementary Material also contains adetailed description of the rRNA structure, as wellas two further ®gures). The cross-linking data canbe found in the Database of Ribosomal Cross-Links(Baranov et al., 1999).
The secondary structure of the 23 S rRNA (modi-®ed from Brimacombe, 1995) is divided into foursections (A to D, Figure 1), with the secondarystructure of the 5 S rRNA (cf. Fox & Woese, 1975)being included in the upper right-hand corner ofthe diagram. The helices of the 23 S rRNA arenumbered from 1 to 101, as was done by Lefferset al. (1987), and those of the 5 S rRNA from I to Vin the usual manner. In addition, the 23 S rRNAhelices are divided into numbered sub-sections,wherever the regular base-pairing (G-C, A-U orG-U pairs) is interrupted; for example, helix 38 (insection B, Figure 1) is divided into ten sections,which we refer to as h38.1 to h38.10. Such a num-bering scheme is essential in order to identify thecorresponding helical regions in the 3D model; thesingle-stranded regions between the helix sub-div-isions were used where necessary to introducebends into the structure during the modelling pro-cedure. The 23 S rRNA is divided into domains,with domain I (section A) comprising h1-h25,domain IIa (section B) h26-h36 and h46, domainIIb (also section B) h37-h45, domain III (section C)h47-h60, domain IV (also section C) h61-h71,domain V (section D) h72-h93, and domain VI(also section D) h94-h101.
Intra-RNA cross-links determined in our labora-tory within the 23 S molecule are denoted inFigure 1 with Roman numerals (I-XVII), whereascross-links between 5 S and 23 S rRNA aredenoted by 5x together with an Arabic numeral(5x1-5x5). A further set of cross-links, where4-thiouridine residues were introduced at speci®cnick sites in isolated 23 S rRNA followed by recon-stitution of the 50 S subunit and UV-irradiation toinduce intra-RNA cross-linking (Baranov et al.,1998), is denoted by NSx (NSx1-NSx2). RNA-pro-tein interactions are identi®ed by the protein name(L1-L33), followed by x for a cross-link site or p fora foot-print site. Corresponding classical bindingsites on the 23 S rRNA for single proteins or

Figure 1 (legend opposite)
38 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ

The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 39
groups of proteins are enclosed in Figure 1 bydotted lines. In the case of the functional data, foot-print sites on the 23 S rRNA from tRNA located atthe ribosomal A, P or E sites (Moazed & Noller,1989) are marked by ®lled triangles, circles orsquares, respectively, whereas site-directed cross-links to 23 S rRNA from photoreactive ligandsincorporated into speci®c positions of the tRNA(Rinke-Appel et al., 1995) are all denoted by Txwith an arrow pointing to the site or sites con-cerned. Similarly, cross-links from various labora-tories to the aminoacyl residue of tRNA aredenoted in Figure 1 by Ax, and cross-links that wehave determined from the N termini of in situ syn-thesized peptides of various lengths are markedPx. In addition, Figure 1 includes sites of foot-printing (Moazed et al., 1988) to elongation factorsEF-Tu and EF-G (denoted by the factor name), asite at position 2252 in h80 which has been shown(Samaha et al., 1995) to base-pair with residue C74of P site tRNA (denoted by TB), and a site at pos-ition 2553 in h92 which has been cross-linked(Green et al., 1998) to a photoreactive derivative ofpuromycin (denoted by pux). Sites of post-tran-scriptional modi®cation in the 23 S rRNA aremarked M for methylated residues (Smith et al.,1992) or X for pseudouridine residues (Ofengand& Fournier, 1998). Figure 1 also shows thelocations of the non-canonical three base-pairmotifs described by Leontis & Westhof (1998),denoted by the symbol W. While we are, of course,not able to observe these motifs in the electrondensity at the current level of resolution, theynevertheless impose severe additional constraintson the 3D folding of the 23 S rRNA; the motif atthe end of h36, for example, serves to bring h37and h45 (Figure 1, section B) into close proximitywith one another, by introducing non-canonicalpairs between nucleotides 818-820 and 1186-1189.
Ever since the ®rst derivation of the secondarystructure of the 23 S rRNA (Glotz et al., 1981), ithas been clear that the 23 S molecule is folded in avery compact manner. Subsequent versions of thesecondary structure (e.g. Noller et al., 1981; Gutellet al., 1994), combined with the identi®cation of ter-
Figure 1. Secondary structures of 23 S and 5 S rRNA, witinto four regions; A, comprising domain I; B, domains IIa anThe helices are numbered and divided into numbered subcross-links are indicated by the double-arrowed lines, intracross-links to 5 S rRNA with 5x- and nick site cross-linkarrowed lines showing the protein number together with p (latter case indicate that the cross-link site concerned was onlof proteins are marked by dotted lines enclosing the approprangles, circles and squares are foot-prints for A, P and E siteing from tRNA, the aminoacyl residue of tRNA or the N tand EF-G are sites of foot-printing from the elongation factsite tRNA, pux a cross-link site to a puromycin derivative.sites and X pseudouridine residues. W marks sequences whthe text for further explanation and references.
tiary interactions (Leffers et al., 1987; Gutell et al.,1994) have served to constrain the structure evenfurther. The tertiary interactions are included inFigure 1 and are denoted by the suf®x t (e.g. h6t-2in section A). Particularly noteworthy are the base-pairs between nucleotides 1269-1270/2011-2010and 1262/2017 (Gutell et al., 1994). We have modi-®ed the layout of our secondary structure diagramin Figure 1 so as to emphasize that these base-pairs(denoted h61a in section C of Figure 1) add a verysigni®cant constraint to the structure, by drasticallyshortening the potential separation betweendomain II, domain III, domain IV and domain V.Helix h61a (which also carries a W motif) thuseffectively forms the central junction of the 23 SrRNA, occupying a position of importance similarto that of the central pseudoknot in 16 S rRNA(helix 2; Mueller & Brimacombe, 1997a), withdomains II, III, IV and V radiating outwards fromit. It is furthermore noteworthy in this context thatdomain VI is closely connected to domain V (theconnection from h73 to h94), and that the intra-RNA cross-links XVI and XVII in domain VI bringh94 close to the 50/30-terminal h1. Only domain Ihas relatively long single strands (between h1 andh2, h2 and h25, h25 and h26) in this ``central junc-tion'' area of the molecule.
As was the case with the 16 S rRNA (Mueller &Brimacombe, 1997a), the intra-RNA cross-links inthe 23 S rRNA provide important constraints on the3D folding of the molecule. The level of con®dencein these cross-links is high, because, as with the 16 SrRNA, a substantial proportion of them has beencorroborated by independent data from othersources. For example, cross-link V (connecting h65to the base of h93) involves the same two nucleo-tides as those that were identi®ed as being cross-linked to the N terminus of a nascent peptide chainfour amino acid residues long (Stade et al., 1994).Similarly, the two nucleotides involved in cross-linkVIII (connecting the potential helix 70 to h92) areboth foot-print positions for A-site bound tRNA(Moazed & Noller, 1989). Other cross-links, such asXII, XIII and XV, correspond closely to pairs of phy-logenetically co-variant nucleotides in the 23 S
h biochemical data. The 23 S rRNA structure is dividedd IIb; C, domains III and IV; and D, domains V and VI.
-sections as described in the text. Intra and inter-rRNA-rRNA cross-links being marked with Roman numerals,s with NSx-. RNA-protein interactions are denoted byfor a foot-print) or x (for a cross-link); dotted lines in they partially localized. Binding sites for proteins or groupsiate regions of the structure. Other symbols are: ®lled tri-tRNA, respectively. Tx, Ax and Px are sites of cross-link-erminus of a nascent peptide chain, respectively. EF-Tuor concerned. TB denotes a residue that base-pairs to PPositions marked M are post-transcriptional methylationere non-canonical secondary structural motifs occur. See

40 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
structure (Gutell et al., 1994). The recently deter-mined cross-links from 5 S rRNA to 23 S rRNA(Sergiev et al., 1998; Osswald & Brimacombe, 1999)have also played a pivotal role in the modellingprocess. Three of these cross-links (5x3, 5x4 and5x5, Figure 1) are from the same residue (U89) ofthe 5 S molecule (Sergiev et al., 1998), whereas theremaining two (5x1, 5x2) are from a group of substi-tuted uridine residues in hII-hIII of the 5 S rRNA, toseveral sites within the upper part of h38 (section B,Figure 1) and to sites in the region of h83-h85 (sec-tion D), respectively (Osswald & Brimacombe,1999). Together with the cross-links from nick sitesin the 23 S rRNA (NSx1 connecting h21 and h22 insection A, and NSx2 connecting h41 and h42 in sec-tion B; Baranov et al., 1998), this gives a total of 24cross-link sites de®ning tertiary contacts within orbetween the 23 S and 5 S rRNAs. It is noteworthythat the corresponding number of intra-rRNAcross-links used to model the 16 S rRNA was eight(Mueller & Brimacombe, 1997a), and furthermoreonly one of the latter (crosslink IV) is an ``inter-domain'' cross-link. In contrast, in the 23 S rRNAthe cross-links I-IV, V, VIII and X are all inter-domain cross-links, so that together with the ®vecross-links to 5 S rRNA, there is an effective total of12 such cross-links. These inter-domain cross-linksare obviously of particular importance for the 3Dfolding of the structure and emphasize the compactnature of the 23 S rRNA. Thus, at least in regard tothe rRNA cross-linking data, the widely held viewthat less topographical information is available forthe 23 S rRNA than for the 16 S rRNA is demon-strably incorrect.
In contrast to the 30 S subunit (Capel et al.,1988), there is no complete neutron-scattering mapdescribing the relative positions of the mass centresof the 50 S ribosomal proteins, although a partialmap has been published (May et al., 1992). How-ever, the majority of the 50 S proteins have beenmapped by IEM (StoÈ f¯er-Meilicke & StoÈ f¯er, 1990),and protein-protein cross-linking data have beenused to extrapolate the IEM data so as to provideapproximate locations for most of the remainingproteins (Walleczek et al., 1988). The RNA-proteininteraction data (Figure 1) are thus useful for corre-lating the locations in the 3D model of regions ofthe 23 S or 5 S rRNA identi®ed in interactions witha particular protein with the corresponding generalarea of the 50 S subunit where that protein hasbeen mapped. In some cases, such as protein L11(h42-h43, section B of Figure 1) or L2 (h66-h67, sec-tion C), the cross-link, foot-print and binding sitedata are concentrated in the same small area of the23 S rRNA. In other instances, widely separatedareas of the primary or secondary structure of the23 S molecule are involved. Examples are proteinL4, which has cross-link sites in h19a (section A)and h28 (section B), or L23 where the cross-linksites (in h6 and h9, section A) are at a locationdifferent from that of the foot-print and bindingsite data (h51-h54, section C). In such cases, the
different RNA-protein data sets serve as additional3D folding constraints on the 23 S rRNA.
Fitting NMR or X-ray crystallographicstructures of ribosomal components to thecryo-EM reconstruction
X-ray crystallographic structures are availablefor 50 S ribosomal proteins L1 (Nikonov et al.,1996), the RNA-binding domain of L2 (Nakagawaet al., 1999), L6 (Golden et al., 1993), the C-terminaldomain of L7/L12 (Leijonmarck & Liljas, 1987), L9(Hoffman et al., 1996), L14 (Davies et al., 1996), L22(Unge et al., 1998) and L30 (Wilson et al., 1986).Corresponding NMR structures have been derivedfor protein L25 (Stoldt et al., 1998) and for theRNA-binding domain of L11 (Markus et al., 1997).In both the latter cases, the NMR structure of theprotein complexed with rRNA has also been stu-died, L25 with a fragment of 5 S rRNA (Stoldt et al.,1999) and L11 with a fragment of 23 S rRNA(Hinck et al., 1997). Furthermore, X-ray structuresare now available for the L11-rRNA fragment com-plex (Wimberly et al., 1999; Conn et al., 1999), andX-ray as well as NMR structures have been pub-lished both for the sarcin-ricin loop (h95 of the 23 SrRNA, Figure 1; Szewczak & Moore, 1995; Correllet al., 1998) and for fragments of the 5 S rRNA(Dallas & Moore, 1997; Correll et al., 1997).
In their recent X-ray crystallographic study ofthe 50 S subunit at 5 AÊ resolution, Ban et al. (1999)reported the ®tting to the electron density map ofthe sarcin-ricin loop, the L11-rRNA fragment com-plex, and proteins L2, L6, L14 and (indirectly) L1.We used their PDB coordinates to ®t these sameelements (except for L1) to the 7.5 AÊ cryo-EMstructure (Matadeen et al., 1999); in addition, wehave ®tted the structures of the L25-5 S rRNAcomplex and protein L9, as well as making a direct®t of L1 (cf. Malhotra et al., 1998, and seeFigure 6(b), below). The positions of all these struc-tural elements, viewed from the interface side ofthe 50 S subunit, are illustrated in mono and stereoin Figure 2(a) and (b), respectively. Figure 2(a) alsoidenti®es the structural ``landmarks'' of the 50 Ssubunit, including the central protuberance, the L1and L7/12 protuberances, the A site ®nger, theprincipal interface protrusion (PIP), the interfacecanyon (known also as the peptidyl transferasecleft) and the interface bridge. Figure 2(c) shows aclose-up stereo view of the ®tted L25-5 S rRNAfragment complex. The positioning of L9 has beendescribed in detail already (Matadeen et al., 1999),and it is noteworthy that the location of this pro-tein appears to be signi®cantly different in isolated50 S subunits as opposed to 70 S ribosomes.
The PDB coordinates of the ribosomal com-ponents ®tted by Ban et al. (1999) correspondedremarkably well with the cryo-EM density, andonly minimal adjustments were necessary to opti-mize the ®t of each individual structure to thecryo-EM density (Figure 2). It should be noted,however, that the placement of the L11-rRNA com-
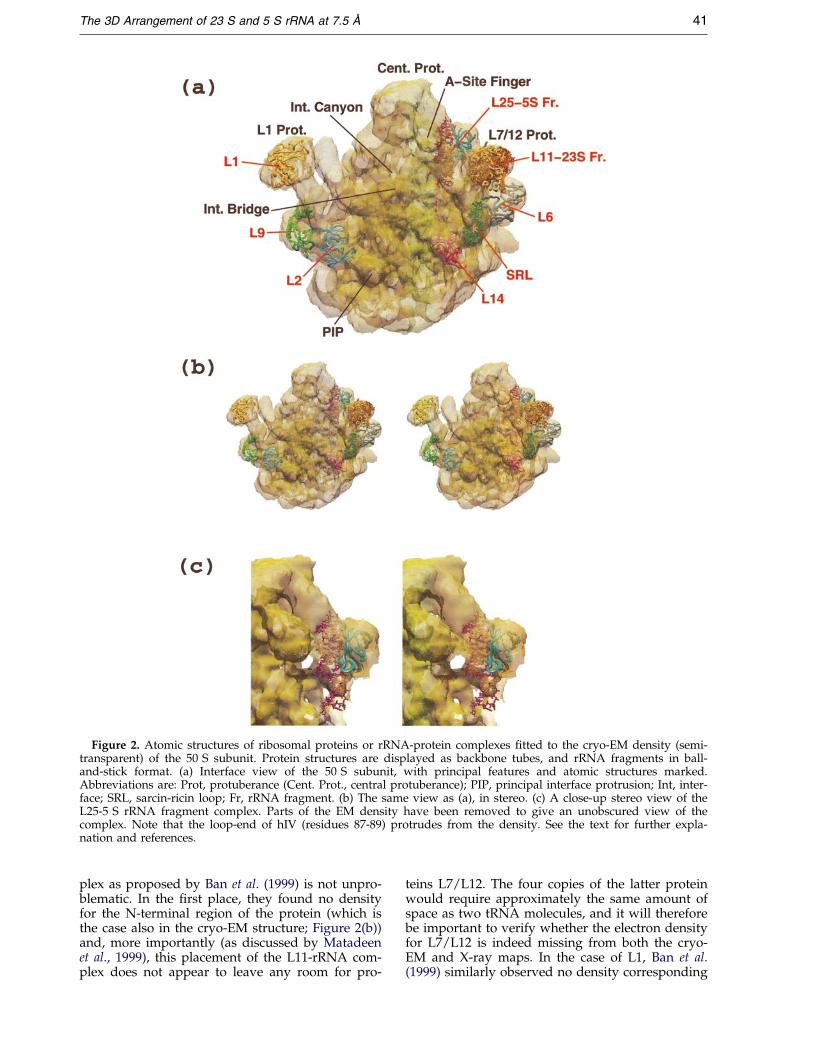
Figure 2. Atomic structures of ribosomal proteins or rRNA-protein complexes ®tted to the cryo-EM density (semi-transparent) of the 50 S subunit. Protein structures are displayed as backbone tubes, and rRNA fragments in ball-and-stick format. (a) Interface view of the 50 S subunit, with principal features and atomic structures marked.Abbreviations are: Prot, protuberance (Cent. Prot., central protuberance); PIP, principal interface protrusion; Int, inter-face; SRL, sarcin-ricin loop; Fr, rRNA fragment. (b) The same view as (a), in stereo. (c) A close-up stereo view of theL25-5 S rRNA fragment complex. Parts of the EM density have been removed to give an unobscured view of thecomplex. Note that the loop-end of hIV (residues 87-89) protrudes from the density. See the text for further expla-nation and references.
The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 41
plex as proposed by Ban et al. (1999) is not unpro-blematic. In the ®rst place, they found no densityfor the N-terminal region of the protein (which isthe case also in the cryo-EM structure; Figure 2(b))and, more importantly (as discussed by Matadeenet al., 1999), this placement of the L11-rRNA com-plex does not appear to leave any room for pro-
teins L7/L12. The four copies of the latter proteinwould require approximately the same amount ofspace as two tRNA molecules, and it will thereforebe important to verify whether the electron densityfor L7/L12 is indeed missing from both the cryo-EM and X-ray maps. In the case of L1, Ban et al.(1999) similarly observed no density corresponding

42 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
to the protein, although in the cryo-EM map(Figure 2(a)) density for L1 is clearly present.
In order to ®t the L25-5 S rRNA fragment com-plex, the structure presented by Stoldt et al. (1999)(which includes nucleotides 70-82 and 94-106 ofthe 5 S molecule) was superimposed on the NMRstructure for nucleotides 70-106 described byDallas & Moore (1997), and this complex was then®tted to the cryo-EM density. The NMR structurefor the 5 S rRNA fragment was more useful forour purposes than the similar X-ray structurereported by Correll et al. (1997), which comprisedhI, hIV and hV of the 5 S rRNA; in the latter struc-ture, the three-way junction between hI, hII andhV is destroyed, and the loop-end of hIV (whichcontains several important cross-link sites; Figure 1)is missing. The position of the L25-5 S rRNA com-plex in the cryo-EM density (Figure 2(c)) providesan important focus for locating a number ofelements of the 23 S rRNA, and a more detaileddescription of the ®tting of this complex will bepresented elsewhere (T. Pape, R.M., M. Cuff &M.v.H., unpublished results).
It should be noted that we did not apply anyenergy minimization procedures (see Materials andMethods) to the interfaces between the protein andrRNA structures. This is because induced ®ttingbetween the bound protein and its site on therRNA is likely to modify the conformations of bothcomponents to an extent that is not possible toassess at our current level of resolution. Thus, it ispreferable for the moment not to allow any altera-tions to the X-ray or NMR structures of the(bound) proteins and, in consequence, to toleratesome van der Waals con¯icts between the proteinand the rRNA. These can be removed at a laterdate when an appropriate resolution for the rRNAstructure (or for the rRNA-protein complex)becomes available, as is already the case for bothL11 and L25.
Fitting the rRNA to the cryo-EM reconstruction
The cavities and channels in the cryo-EM struc-ture (Figure 2) impose enormous constraints on thepossible 3D folding of the 23 S rRNA, and the ®t-ting of the latter to the EM density in such a wayas to satisfy the biochemical constraints (Figure 1)is not a trivial task. We adopted the followingstrategy. A ``wire-and-tube'' model, previouslyused to construct a preliminary 3D model of the23 S rRNA (Mitchell et al., 1990), was taken as thestarting point, and this was adapted so as to incor-porate the more recent biochemical information(Figure 1). (With large molecules such as the 23 SrRNA, ``physical'' wire-and-tube models still havecertain advantages over computer-generatedmodels, in particular for gaining an overview ofthe complete structure.) The wire-and-tube modelwas then re-generated as faithfully as possible inthe computer, using the programme ERNA-3D(Mueller & Brimacombe, 1997a), and the computerversion was inserted (together with the corre-
sponding model for the 5 S rRNA) into the EMdensity. Next, the detailed ®tting process wasbegun, moving each helix of the 23 S and 5 S mol-ecules so as to ®t an appropriate density element,at the same time keeping track of (a) the interheli-cal connections within the secondary structure, (b)the intra-rRNA cross-links and tertiary interactionsin the rRNA, (c) the general locations of the var-ious RNA-protein interaction sites, and (d) thelocations of the atomic structures of ribosomalcomponents described in the foregoing section. Asalready noted above, the intra-helical loops andbulges were used where necessary to introducebends into the helices so as to ®t the curves in theEM density elements. A preliminary ®tting of the23 S and 5 S rRNA had been made (Brimacombe,1999; Brimacombe et al., 2000) to the cryo-EMreconstruction at 13 AÊ mentioned in the Introduc-tion of 70 S ribosomes carrying a P-site tRNA anda kirromycin-stalled EF-Tu/tRNA complex (H.Stark, M. Rodnina, F. Zemlin, W. Wintermeyer &M.v.H., unpublished results; and see Stark et al.,1997b); the structure presented here represents are®nement of this preliminary ®tting.
A special ``helix-bump'' programme was devel-oped as part of the energy minimizing procedure(see Materials and Methods) to check for van derWaals con¯icts between the rRNA helices. Thisprogramme was primarily intended to be used for®ne tuning of the structure, but in fact proved tobe extremely useful in the early stages of the mod-elling. Since only a small part of the overall 23 Smolecule can be selected at any one time for the ®t-ting process, it was not uncommon at ®rst for thesame density element to be inadvertently used toaccommodate two different helices from differentareas of the rRNA; such errors were immediatelyregistered by the helix-bump programme. Severalrounds of re®nement of the ®tted structure werecarried out, until we were satis®ed that the entire23 S molecule, as well as the 5 S rRNA, were opti-mally incorporated into the EM density. In the®nal structure, there was only one case where ahelix or a single strand of the rRNA had to cross acanyon or gap in the EM density (cf. Figures 5 and6, below, and see the discussion relating toFigure 6(b)), and furthermore the intra-RNA cross-links were all at positions in the structure thatappeared plausibly accessible for a cross-linkingreagent to reach. The last stage in the modellingprocedure was energy minimization, using pro-grammes especially developed for the purpose (seeMaterials and Methods). First, the single strands ofthe rRNA were automatically manipulated, keep-ing the helical elements (and the NMR-derivedRNA structures) ®xed, so as to remove van derWaals con¯icts, optimize the positions of single-stranded nucleotides involved in cross-links, andto keep the single strands within the EM density asfar as possible. Any remaining violations wereeliminated by allowing the helical elements tomove slightly. It should be emphasized, however,that in the case of the single-stranded regions this

The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 43
energy minimization procedure is still only anapproximation, primarily intended to demonstratethe plausibility of the connections between helices.Furthermore, for a structure of this complexity theprocedure is not entirely error-free, with the resultthat the minimized structures contain occasionalincorrect bond lengths or bond angles, or potentialknots where two long single strands have fortui-tously crossed one another during the modellingprocedure. We anticipate that these and othererrors will be ironed out in subsequent versions ofthe structure at higher resolution.
The 3D arrangement of the double-helicalelements of the 23 S and 5 S rRNA structures, rep-resented as cylinders, and with appropriate helicesnumbered as in Figure 1, is illustrated in Figures 3and 4. Figure 3 shows views from the L7/L12 sideof the 50 S subunit ((a) and (b)) and from the L1
side ((c) and (d)), respectively. In Figure 3(a) and(c), the complete rRNA structure is shown, and allof the visible helices are numbered, whereas inFigure 3(b) and (d), the layer of helices nearest tothe viewer has been removed so as to reveal thearrangement of helices in the interior of the sub-unit; in the latter cases, only these interior helicesare numbered (together with a few ``landmark''helices from the numbering of Figure 3(a) and (c)).Figure 4 is constructed in a similar manner, show-ing views from the interface side of the subunit ((a)and (b)) and from the solvent side ((c) and (d)),respectively; again, the outer layers of helices havebeen removed in Figure 4(b) and (d). The cylindersin Figures 3 and 4 are colour-coded according tothe secondary structural domains.
Some details of the rRNA structure ®tted to thecryo-EM density are given in Figures 5 and 6.
Figure 3. The 3D arrangement ofthe double-helical elements of the23 S and 5 S rRNA molecules inthe 50 S subunit. The helices arerepresented as cylinders, numberedas in Figure 1. The colour-codingis: domain I, red; domain IIa, yel-low; domain IIb, orange; domainIII, green; domain IV, light blue;domain V, magenta; domain VI,dark blue; and 5 S rRNA grey-brown. (a) View of the structurefrom the L7/12 side of the subunit.(b) The same view as in (a), butwith the layer of helices nearest tothe reader removed. (c) View fromthe L1 side of the subunit. (d) Thesame view as in (c), but with theouter layer of helices removed as in(b).

Figure 4. Further views of the3D arrangement of the 23 S and 5 SrRNA, displayed as in Figure 3. (a)View from the interface side of thesubunit. (b) The same view as in(a), but with the outer layer ofhelices removed (cf. Figure 3(b)).(c) View from the solvent side ofthe subunit. (d) The same view asin (c) but with the outer layer ofhelices removed as in (b).
44 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
Figure 5(a) shows the complete atomic structuresof both the 5 S and 23 S molecules in a semi-trans-parent rendering of the 50 S subunit, viewed fromthe L1 side. The rRNA elements are colour-codedaccording to their locations in double-helicalregions, hairpin loop-ends, intra-helical loops orinter-helical strands. It can be seen that the rRNAis distributed uniformly throughout the electrondensity, with many of the hairpin loop-endsdirected towards the periphery of the structure.Areas of empty density are present also, whichcould plausibly accomodate ribosomal proteins (inaddition to those described in Figure 2(a)). A num-ber of helices from widely separated regions of thesecondary structure appear to come together at thebottom of the subunit (cf. Figures 3 and 4). Theseinclude h5-h7, h46, h50-h54, h62-h63, h96 andh101, and their positions are illustrated inFigure 5(b), which indicates the location of the exitend of the putative peptide channel (cf. Figure 8(b),below). It should be noted that our positioning ofh96 differs from that tentatively proposed by Banet al. (1999). However, the loop-ends of the helices
depicted in Figure 5(b) are good candidates forapplication of the ``nick site'' cross-linking methodof Baranov et al. (1998), and experiments to verifythe juxtaposition of these helices using the lattermethod are in progress.
Figure 6(a) shows details of rRNA helices locatedon the interface side of the 50 S subunit, with pro-teins L2 and L14 included for reference. Helixh34.2 is placed in the PIP (Culver et al., 1999;Matadeen et al., 1999), whereas h69 forms the inter-face bridge, by virtue of the cross-link from theloop-end of this helix to h44 of the 16 S rRNA(Mitchell et al., 1992; Figure 1). Helix h69 connectsto h68.1-h68.3, which lie along the outer rim of thepeptidyl transferase cleft to the left of h69, andh68.1 connects in turn to h66 and to h67. Helix h66represents the binding site for protein L2 (Figure 1),and it is noteworthy that the amino acid residuesin L2 that have been predicted to be in contactwith the 23 S rRNA (Nakagawa et al., 1999) are onthe ``inside'' face of the protein (i.e. the side facingaway from the viewer in Figure 6(a)), reasonablyclose to the loop-end of h66. Figure 6(a) shows h92
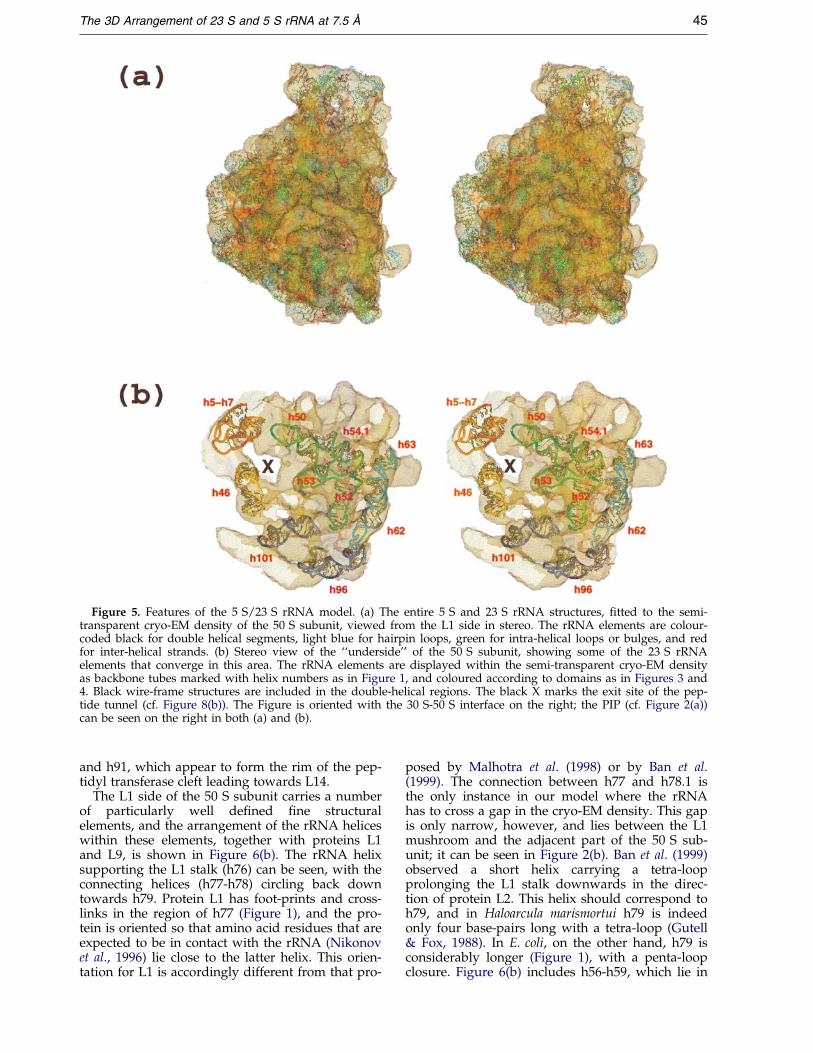
Figure 5. Features of the 5 S/23 S rRNA model. (a) The entire 5 S and 23 S rRNA structures, ®tted to the semi-transparent cryo-EM density of the 50 S subunit, viewed from the L1 side in stereo. The rRNA elements are colour-coded black for double helical segments, light blue for hairpin loops, green for intra-helical loops or bulges, and redfor inter-helical strands. (b) Stereo view of the ``underside'' of the 50 S subunit, showing some of the 23 S rRNAelements that converge in this area. The rRNA elements are displayed within the semi-transparent cryo-EM densityas backbone tubes marked with helix numbers as in Figure 1, and coloured according to domains as in Figures 3 and4. Black wire-frame structures are included in the double-helical regions. The black X marks the exit site of the pep-tide tunnel (cf. Figure 8(b)). The Figure is oriented with the 30 S-50 S interface on the right; the PIP (cf. Figure 2(a))can be seen on the right in both (a) and (b).
The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 45
and h91, which appear to form the rim of the pep-tidyl transferase cleft leading towards L14.
The L1 side of the 50 S subunit carries a numberof particularly well de®ned ®ne structuralelements, and the arrangement of the rRNA heliceswithin these elements, together with proteins L1and L9, is shown in Figure 6(b). The rRNA helixsupporting the L1 stalk (h76) can be seen, with theconnecting helices (h77-h78) circling back downtowards h79. Protein L1 has foot-prints and cross-links in the region of h77 (Figure 1), and the pro-tein is oriented so that amino acid residues that areexpected to be in contact with the rRNA (Nikonovet al., 1996) lie close to the latter helix. This orien-tation for L1 is accordingly different from that pro-
posed by Malhotra et al. (1998) or by Ban et al.(1999). The connection between h77 and h78.1 isthe only instance in our model where the rRNAhas to cross a gap in the cryo-EM density. This gapis only narrow, however, and lies between the L1mushroom and the adjacent part of the 50 S sub-unit; it can be seen in Figure 2(b). Ban et al. (1999)observed a short helix carrying a tetra-loopprolonging the L1 stalk downwards in the direc-tion of protein L2. This helix should correspond toh79, and in Haloarcula marismortui h79 is indeedonly four base-pairs long with a tetra-loop (Gutell& Fox, 1988). In E. coli, on the other hand, h79 isconsiderably longer (Figure 1), with a penta-loopclosure. Figure 6(b) includes h56-h59, which lie in
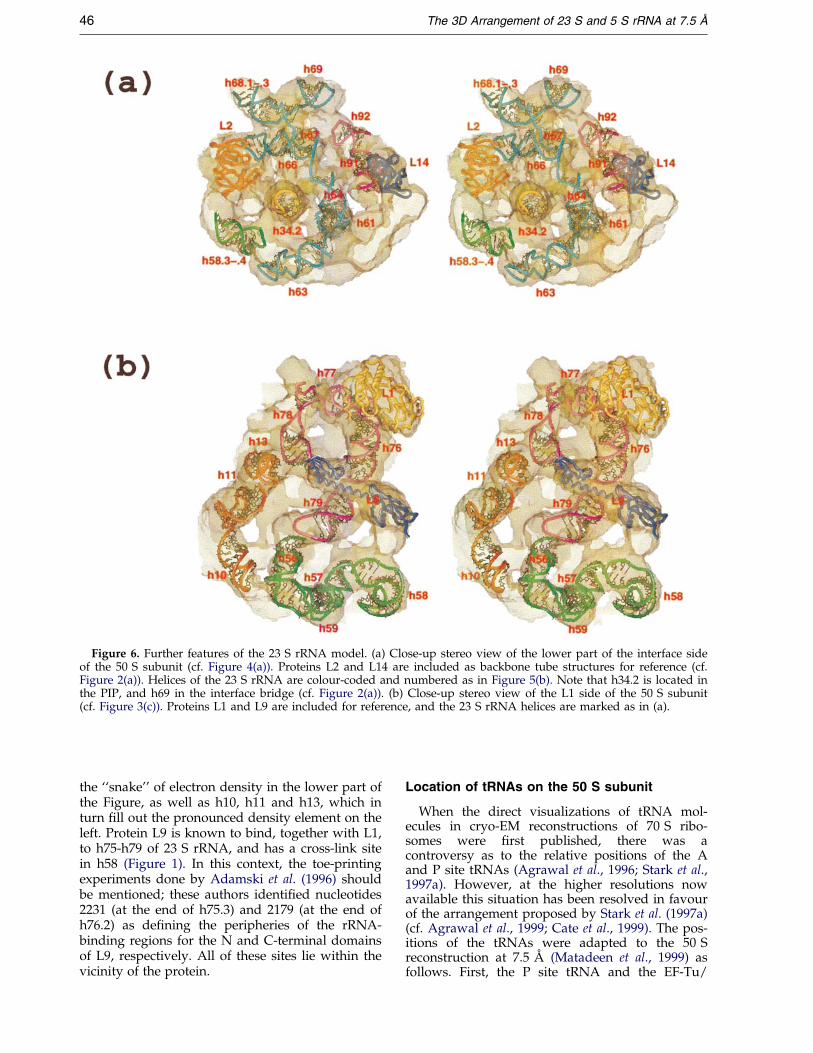
Figure 6. Further features of the 23 S rRNA model. (a) Close-up stereo view of the lower part of the interface sideof the 50 S subunit (cf. Figure 4(a)). Proteins L2 and L14 are included as backbone tube structures for reference (cf.Figure 2(a)). Helices of the 23 S rRNA are colour-coded and numbered as in Figure 5(b). Note that h34.2 is located inthe PIP, and h69 in the interface bridge (cf. Figure 2(a)). (b) Close-up stereo view of the L1 side of the 50 S subunit(cf. Figure 3(c)). Proteins L1 and L9 are included for reference, and the 23 S rRNA helices are marked as in (a).
46 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
the ``snake'' of electron density in the lower part ofthe Figure, as well as h10, h11 and h13, which inturn ®ll out the pronounced density element on theleft. Protein L9 is known to bind, together with L1,to h75-h79 of 23 S rRNA, and has a cross-link sitein h58 (Figure 1). In this context, the toe-printingexperiments done by Adamski et al. (1996) shouldbe mentioned; these authors identi®ed nucleotides2231 (at the end of h75.3) and 2179 (at the end ofh76.2) as de®ning the peripheries of the rRNA-binding regions for the N and C-terminal domainsof L9, respectively. All of these sites lie within thevicinity of the protein.
Location of tRNAs on the 50 S subunit
When the direct visualizations of tRNA mol-ecules in cryo-EM reconstructions of 70 S ribo-somes were ®rst published, there was acontroversy as to the relative positions of the Aand P site tRNAs (Agrawal et al., 1996; Stark et al.,1997a). However, at the higher resolutions nowavailable this situation has been resolved in favourof the arrangement proposed by Stark et al. (1997a)(cf. Agrawal et al., 1999; Cate et al., 1999). The pos-itions of the tRNAs were adapted to the 50 Sreconstruction at 7.5 AÊ (Matadeen et al., 1999) asfollows. First, the P site tRNA and the EF-Tu/

The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 47
tRNA complex were ®tted to the 13 AÊ reconstruc-tion of kirromycin-stalled 70 S ribosomes men-tioned in Introduction (unpublished results; Starket al., 1997b; Brimacombe et al., 2000). In order tooptimize this ®t, the CCA end of the P site tRNAwas turned upwards slightly in relation to the X-ray structure of tRNA (Kim et al., 1974), and theangle between the EF-Tu molecule and its attachedtRNA was also adjusted in relation to the X-raystructure of the complex (Nissen et al., 1995; cf.Stark et al., 1997b). The 13 AÊ cryo-EM reconstruc-tion (including these structures) was then superim-posed on the 7.5 AÊ 50 S reconstruction, so that thepositions of the P site tRNA and the EF-Tu/tRNAcomplex could be transferred to the 50 S recon-struction. The tRNA in the EF-Tu/tRNA complexis not at the A site, but rather at a ``pre-A'' site(Stark et al., 1997b), referred to also as the T site.The position of the A site tRNA as such wasaccordingly interpolated by taking the coordinatesof the A and P site tRNA molecules from the 20 AÊ
reconstruction presented by Stark et al. (1997a),and superimposing them on the 50 S reconstruc-tion (with P site tRNA added as just described) sothat the P site molecules from the two reconstruc-tions coincided. The exit site tRNA was similarlypositioned, making use of the coordinates of the Pand E site tRNAs from the same 20 AÊ reconstruc-tion (Stark et al., 1997a).
The locations of the various tRNA positions, andthat of EF-Tu, are illustrated in Figure 7(a), whichshows the 50 S subunit from the interface side. Itcan be seen that the CCA ends of the A and P sitemolecules lie close together to enable peptidyltransfer to occur and, similarly, the two anticodonloops are close enough for simultaneous codon-anticodon interaction to take place. In contrast, thepre-A site position is quite different from that ofthe A site, with the CCA end of the tRNA rotatedtowards the L7/12 side of the 50 S subunit. Never-theless, the anticodon loops of the A and pre-A sitetRNAs overlap one another, which is as would beexpected; the incoming EF-Tu/tRNA complexmust be able to read the codon at the A site. Theexit site position in Figure 7(a) correspondsapproximately to the E2 site observed by Agrawalet al. (1999), the E site itself as de®ned by theseauthors lying in the gap between the E and P sitesas depicted in Figure 7(a). In the E site describedby Agrawal et al. (1999), the anticodon loops of theE and P site tRNAs are potentially able to under-go simultaneous codon-anticodon interaction, asproposed by Gnirke et al. (1989), whereas in theE2 site (or E site of Figure 7(a)) this is clearly nolonger possible.
It is not appropriate here to enter into a discus-sion of the de®nition of the exit site or sites, ratherthe purpose of Figure 7(a) is to demonstrate that,as the tRNAs move through the ribosome duringthe protein biosynthetic cycle, they describe a widearc traversing the whole face of the 50 S subunit,from the L7/12 protuberance to the L1 protuber-ance. Furthermore it is interesting that at each pos-
ition the elbow region of the tRNA appears to lieclose to a speci®c feature of the 50 S subunit; in thepre-A site it is the L7/12 protuberance, in the Asite the A site ®nger, in the P site, the underside ofthe central protuberance, and in the E (or E2) sitethe L1 mushroom. In contrast, at the A and P sitesthe CCA ends of the tRNAs lie deep in the pepti-dyl transferase cleft or interface canyon (describedby Wagenknecht et al., 1989) at the entrance to thepeptide tunnel.
Locations of tRNA foot-print andcross-link sites
The foot-print sites for tRNA bound at the A, Por E site described by Moazed & Noller (1989) arelocated within domains IV and V of the 23 S rRNA(Figure 1), with the exception of the group of Asite foot-prints in h42-h43, which were in¯uencedby the presence of EF-Tu (Moazed & Noller, 1989).The locations of the A and P site foot-prints in the23 S rRNA model (except for this latter group of Asite foot-prints) are given in Figure 7(b). The corre-sponding E site foot-prints lie in the region of h77and h88 (Figure 1) at positions that correspond tothe E site tRNA location described by Agrawal et al.(1999) (compare Figures 4(a) and 7(a), and see theforegoing discussion of the E site); these will notbe considered further here.
In Figure 7(b) the P and A site tRNAs are shownand, as noted above, the CCA of the P site tRNA isturned up slightly in relation to its position in theX-ray structure (Kim et al., 1974). This movementbrings C74 of the tRNA into potential base-pairingdistance from G2252 of the 23 S rRNA in h80 asshown by Samaha et al. (1995), G2252 being knownfrom NMR studies to be exposed to the solvent(Puglisi et al., 1997). C74 and G2252 are illustratedby the CPK nucleotides in Figure 7(b), whichclearly lie close together (just below the acceptorstem of the tRNA), although for technical reasonsthey are not displayed as an actual Watson-Crickpair. G2252 is also a foot-print site position for theP site tRNA, and is located on the inner wall of theinterface canyon. In contrast, the other A and P sitefoot-print positions are distributed on both sides ofthe canyon. Moazed & Noller (1991) demonstratedthat all of the P site foot-prints, with the exceptionof those at positions 1916, 1918, 1926 and 2505, aredependent on the presence of C75 and A76 of thetRNA. The ®rst three of these positions are those inor close to h69, which as already noted is directedtowards the decoding site of the 30 S subunit.Among the P site foot-prints, these sites in h69 arethus the farthest away from the CCA of the P sitetRNA (Figure 7(b)) and presumably the least likelyto be in¯uenced by the absence of the 30-terminalnucleotides. Figure 7(b) indicates the location ofthe site that has been cross-linked to a photoreac-tive derivative of puromycin (Green et al., 1998);this site (G2553) can be regarded as a ``marker'' forthe CCA end of the A site tRNA.

Figure 7 (legend opposite)
48 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
The cross-linking data for tRNA (Rinke-Appelet al., 1995; see also Table 4 of the SupplementaryMaterial) are predominantly from positions in thecentral fold or elbow region of the tRNA molecule(positions 8, 20:1 and 47), and the effective brid-
ging distance of the cross-linking agents used was10-15 AÊ . An exception in both respects is the cross-link reported by Wower et al. (1989), which is fromthe 30 terminus of the P site tRNA (position 76)and involves a zero-length cross-link from an

The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 49
azido-adenine residue at that position. Several ofthe cross-links from the elbow region (Rinke-Appelet al., 1995) could not be localized precisely and,furthermore, the site-speci®city (A, P or E) was notclear in some cases, particularly with regard to theE site. The locations of the A and P site cross-linksthat were both site-speci®c and localized to a pre-cise nucleotide are given in Figure 7(c). Especiallyinteresting are the cross-links from position 47 ofthe P site tRNA to nucleotides in h83-h85 at thebase of the central protuberance of the 50 S sub-unit, and those from position 47 of the A sitetRNA to h89 as well as from position 8 of the latterto the loop-end of h38. Notably, cross-links frompositions 20:1 and 47 of the A site tRNA wereobserved to the loop-end of h38 (Rinke-Appel et al.,1995; Osswald et al., 1995), although these wereamong the cross-links that could not be pinpointedprecisely.
Given the lengths of the cross-linking agentsused, in principle it should not be possible forcross-links to be formed from the elbow region ofthe tRNA to sites on the 23 S rRNA that are closeto the CCA end of the tRNA. The presence of suchcross-links (Figure 7(c)), together with the overalldistribution of the cross-link sites in the latterFigure, suggests that (as was the case in our stu-dies with the 16 S rRNA (Mueller et al., 1997))there is some ¯exibility in the tRNA-binding sitesunder the conditions used for the cross-linking. Itis highly probable that conformational changes inthe 23 S rRNA occur under the differing conditionsthat are required by the various experiments, par-ticularly in the functionally important areas, and itis already evident (Matadeen et al., 1999) that the50 S subunit undergoes signi®cant changes when it
Figure 7. (a) Positions of tRNA sites on the 50 S subunit, vis displayed as an opaque object, with the tRNA moleculesthe tRNA in the EF-Tu/tRNA ternary complex (the pre-A sitand the E site tRNA (or E2 site) green. The A and E site tRNthat these two tRNA positions were interpolated from differof A and P site tRNA foot-print positions in the 23 S rRNAshown, semi-transparent and viewed from the L7/12 side;tRNAs are displayed as light blue and green backbone tubesplayed as dark blue backbone tubes, with black wire-framebered as in Figure 1. The foot-print sites are marked by colothe 23 S rRNA: A site foot-prints (light blue spheres) at nucl(green spheres) at 1916, 1918, 1926, 2253, 2505, 2506, 2584 anat 2439 and 2451. Nucleotide 2553 is displayed as a yellowthe P site tRNA as red CPK nucleotides (see the text). (c)rRNA model. The A and P site tRNA molecules, and relevatransparent contour of the 50 S subunit as in (b), except thatunit. Cross-links from the elbow region of the tRNAs are ifrom position 8, orange from position 20:1 and magenta frsmaller diameter. Sites of cross-linking on the 23 S rRNA aresite, nucleotides 889 (light blue), 2586 (orange), and 2469, 242574 (orange), and 1852, 2296, 2309, 2321, 2326, 2334, 2452tRNA to nucleotide 1945 of the 23 S rRNA is indicated simiand h74, only the relevant parts of the helix concerned are d
becomes part of the 70 S ribosome. In this context,it is noteworthy that at the base of h71 there is across-link from nucleotide 1963 to amino acid resi-due 67 of protein L2 (Thiede et al., 1998; Figure 1)as well as the cross-link from nucleotide 1945 toposition 76 of the P site tRNA (Wower et al., 1989);from a comparison of Figures 2(a) and 7(c), it isclear that these two cross-links cannot be satis®edsimultaneously.
A further data set that needs to be mentionedin the context of the tRNA foot-printing andcross-linking results concerns the identi®cation ofcleavage sites in the 23 S rRNA induced in the pre-sence of Cu2� by ribosome-bound tRNA deriva-tized with phenanthroline. Using this technique,Bullard et al. (1995) have identi®ed an extensive setof cleavage sites covering a large area of the 23 Smolecule. Whereas many of their sites are consist-ent with the foot-printing and cross-linking data indomains IV and V of the 23 S rRNA, a number ofother cleavage sites were observed in domains I, IIIand VI, which in our model are located on the sol-vent side of the 50 S subunit. This discrepancyremains to be clari®ed.
According to current estimates, there are 24post-transcriptionally modi®ed nucleotides inE. coli 23 S rRNA, 16 of them being base methyla-tions (Smith et al., 1992) and eight pseudouridineresidues (Ofengand & Fournier, 1998); theirlocations in the 23 S rRNA secondary structure areincluded in Figure 1. It has been clear for sometime that the modi®ed bases in both 16 S and 23 SrRNA are concentrated into clusters in the 3Dstructure (Brimacombe et al., 1993; Mueller &Brimacombe, 1997a), and in our 23 S rRNA modelthis clustering is around the CCA ends of the
iewed from the interface side in stereo. The 50 S subunit(and EF-Tu) as backbone tubes. The EF-Tu is magenta,e) is blue, the A site tRNA is white, the P site tRNA redAs are displayed with smaller tube diameters to indicateent cryo-EM reconstructions (see the text). (b) Locations
model. A close-up in stereo of part of the 50 S subunit isthe interface canyon is clearly visible. The A and P site, respectively. Relevant regions of the 23 S rRNA are dis-structures in the double-helical regions; helices are num-ured spheres at the positions of the phosphate groups ineotides 1941, 2254, 2555, 2602 and 2609; P site foot-printsd 2585; sites common to both tRNAs (magenta spheres)
CPK nucleotide, and 2252 together with nucleotide 74 ofLocations of A and P site tRNA cross-links in the 23 Snt regions of the 23 S rRNA, are displayed on the semi-in this case the view is from the interface side of the sub-ndicated by the coloured phosphate spheres (light blueom position 47); the spheres for the P site tRNA are of
marked by spheres corresponding in size and colour: A75, 2480 (magenta); P site, nucleotides 2428 (light blue),(magenta). The cross-link from position 76 of the P site
larly by green spheres. (Note that in the case of h38, h68isplayed.)

Figure 8 (legend opposite)
50 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
A and P site tRNAs. The only modi®ed nucleotidethat lies somewhat outside this cluster is the meth-ylated residue at nucleotide 1618 (h59a); this site isclose to the peptide cross-link at nucleotide 1614(Figure 1), which is located in the peptide tunnel(see below).
The peptidyl transferase ring and thepeptide tunnel
A number of cross-linking studies have beenmade using photoreactive reagents attached tothe a-NH2 group of an amino acid, either as the

The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 51
aminoacyl residue of tRNA (Steiner et al., 1988;Mitchell et al., 1993) or at the N terminus of agrowing peptide chain. In the latter type of exper-iment (Stade et al., 1994, 1995; Choi & Brimacombe,1998) peptides of up to 30 amino acid residues inlength were synthesized in situ on the ribosomeusing suitable mRNA analogues, and the sites ofcross-linking to 23 S rRNA from the N termini ofthe various peptides were analysed. The resultsshowed that, with increasing peptide length, the Ntermini of the peptides became progressively cross-linked to sites ®rst in domain V of the 23 S rRNA(in the peptidyl transferase ring), then in domainIV (h65), then domain II (h35, h35a-h37), thendomain III (h49-51, h59a), and ®nally in domain I(h7) (Figure 1). However, at the same time it wasclear that the nascent chain must be very ¯exible,as in every case the longer peptides were able to``reach back'' and become cross-linked to almost allof the sites observed with the shorter peptides. Thedata from these experiments (Stade et al., 1994)also included cross-links from peptides ``oneamino acid long'', i.e. from the aminoacyl residueof P-site bound tRNA.
All of the cross-links from the aminoacyl residueare located within or close to the peptidyl transfer-ase ring (Vester & Garrett, 1988), which is enclosedby helices h73, h74, h89, h90 and h93 (Figure 1).This was an intriguing part of the 23 S rRNA tomodel, because, apart from the large amount offunctional data in the area, other modelling con-straints dictate that the peptidyl transferase ringhas to run around both sides of the interface can-yon. This region of the 23 S rRNA, together withthe cross-links from the aminoacyl residue oftRNA is illustrated as a close-up stereo view fromthe interface side of the 50 S subunit in Figure 8(a).There is in fact a total of 30 single-stranded resi-dues in the peptidyl transferase ring, which, ifextended, would have a length of about 200 AÊ . Itcan be seen from Figure 8(a) that the peptidyltransferase ring can be accommodated without vio-
Figure 8. (a) The peptidyl transferase ring, with cross-linregions of the 23 S rRNA are displayed with helices numberred backbone tube. (Note that in the case of h73, h89 and h9the peptidyl transferase ring are displayed.) The close-up stesemi-transparent 50 S subunit, from the interface side of thgroups in the 23 S rRNA indicate the cross-link sites from2451, 2505, 2584 (Steiner et al., 1988); green sphere at 2439 (M(Stade et al., 1994). The black x marks the peptide tunnel. (side of the 50 S subunit. The subunit is rendered opaque, wto reveal the tunnel. The P site tRNA is shown as a red batides of different lengths are shown by coloured spheres at ppeptide with three amino acid residues); orange, nucleotideturquoise, nucleotide 1614 (12 or 25 residues; see Choi & BThe dark blue arrow highlights the path of the tunnel. (c) Tthe semi-transparent 50 S subunit is shown, viewed from thtube, with the L11 binding region of the 23 S rRNA (nucle2646-2674) as orange and light blue tubes, respectively. Dostructures as in Figure 6. Nucleotides of particular interest aEF-Tu complex (green); nucleotide 1067 (light blue); nucleotid
lating the constraints of the EM density and, at thesame time, it corresponds to the entrance to thepeptide tunnel, which lies just beneath the CCAend of the P site tRNA molecule in the EM recon-struction. It is noteworthy that several of the cross-links from the aminoacyl residue in Figure 8(a)coincide with tRNA foot-print or cross-link sites(Figure 1); nucleotide 2506 is particularly ubiqui-tous. It should be noted that at the current level ofresolution we make no distinction between thecross-links from aminoacyl tRNA at the A and Psites (cf. Steiner et al., 1988), since the 30 termini ofthe two tRNAs are closely juxtaposed.
Details of the peptide tunnel, together with thepeptide cross-links (Figure 1), are shown inFigure 8(b), which is a stereo view from the L7/12side of the 50 S subunit. The subunit is rendered asan opaque object, and the EM density has been cutaway so as to reveal the tunnel, which runs fromthe interface canyon to a point on the solvent sideof the subunit, close to the location for the exit sitededuced from IEM studies by Bernabeu & Lake(1982). The tunnel exit also corresponds to thatobserved in yeast ribosomes by Beckmann et al.(1997), which they demonstrated by cryo-EM to bein line with the doughnut-shaped Sec61 complexthat is involved in the translocation of newly syn-thesized proteins across the endoplasmic reticulummembrane. The tunnel is not straight, and inFigure 8(b) the EM density has been cut away soas to reveal it optimally as a continuous corridor;this continuity is not evident from a planar sectionthrough the subunit as shown by Beckmann et al.(1997), (cf. also Stark et al., 1995; Malhotra et al.,1998).
The sites of cross-linking to the 23 S rRNA fromthe N termini of the nascent peptides are indicatedin Figure 8(b) by the coloured balls. It can be seenthat all of the sites lie within or close to the tunnel,those from the progressively longer peptides lyingfarther from the peptidyl transferase centre andcloser to the exit site. The peptide cross-link sites
k sites from the aminoacyl residue of tRNA. Relevanted as in Figure 7(b), and the P site tRNA is shown as a0, only those parts of the helices immediately adjacent toreo view shows the peptidyl transferase cleft area of thee latter. Coloured spheres at the positions of phosphatethe aminoacyl residue: light blue spheres at nucleotides
itchell et al., 1993); magenta spheres at 2062, 2506, 2585b) The peptide tunnel, viewed in stereo from the L7/12ith the EM density cut away (leaving the black ``scars'')
ckbone tube. Cross-link sites from the N termini of pep-hosphate positions as in (a): red, nucleotide 2609 (from a1781 (four residues); green, nucleotide 750 (six residues);rimacombe, 1998), magenta, nucleotide 91 (30 residues).he environment of EF-Tu in the 50 S subunit. A part of
e L7/12 side. EF-Tu is displayed as a magenta backboneotides 1051-1108) and the sarcin-ricin loop (nucleotidesuble-helical regions are marked with black wire-framere highlighted in CPK format: the GTP analogue in thee 2660 (red).

52 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
were not used as primary constraints in the model-ling of the 23 S rRNA, and their locations thus pro-vide independent support for the peptide tunnelconcept, as opposed to the alternative proposal(Ryabova et al., 1988) of a peptide channel alongthe outer surface of the 50 S subunit. The width ofthe tunnel, combined with the ¯exibility of the nas-cent peptide chain already mentioned above, arefurthermore consistent with the concept of co-translational folding of the newly synthesized pro-tein chains, as demonstrated by Kudlicki et al.(1995) and Makeyev et al. (1996). As was the casewith the tRNA cross-linking data (above), some ofthe peptide cross-link sites (Stade et al., 1995; Choi& Brimacombe, 1998) could not be localized pre-cisely (notably those encompassing h35a-h37 andh49-h51); these are not considered here, althoughat least parts of the regions concerned lie along thepeptide tunnel.
Locations of EF-Tu, the sarcin-ricin loop andthe L11 binding region of 23 S rRNA
The locations of the sarcin-ricin loop and of theL11 binding region (Ban et al., 1999) in our currentmodel of the 23 S rRNA are taken from the data ofBan et al. (1999) (see Figure 2(a)). These locationsdiffer from those that we had previously proposed(Brimacombe, 1999; Brimacombe et al., 2000),where both rRNA regions were placed lowerdown in the 50 S subunit. The reason for this wasthat we had ``reserved'' the density now occupiedby the L11 binding region (Figure 2(a)) for proteinsL7/12 and, as discussed above, the situation withregard to the placement of these proteins stillneeds to be clari®ed (Matadeen et al., 1999). Thepositions of the sarcin-ricin loop (h95) and the L11binding region (h42.4, h43, h44), together with thatof Ef-Tu (cf. Figure 7(a)), are illustrated inFigure 8(c). Moazed et al. (1988) localized sites offoot-printing from EF-Tu to h95 of the 23 S rRNA,and from EF-G to both h95 and h43. The loop-endof h95 carries the site of action of the toxinsa-sarcin and ricin at nucleotide 2660 (Endo &Wool, 1982; Endo et al., 1987), whereas the site ofresistance to the antibiotic thiostrepton is located atnucleotide 1067 in the loop-end of h43 (Thompsonet al., 1982). These positions are highlighted as CPKnucleotides in Figure 8(c), as is the GTP analoguein the EF-Tu molecule (Nissen et al., 1995).
Ef-Tu (Stark et al., 1997b) and EF-G (Agrawalet al., 1998) occupy essentially similar positions inthe ribosome, and the close contact between h95and the EF-Tu (Figure 8(c)) is thus consistent withthe direct binding data for EF-G of Munishkin &Wool (1997). On the other hand, the L11 bindingregion is not in such close contact with the factor,and it is noteworthy that this placement of h42.4within the L11 binding area (Ban et al., 1999) doesnot allow the ``nick site'' cross-link suggested byBaranov et al. (1998) connecting h42 with h41(Figure 1) to be satis®ed. It should be borne inmind, however, that this area of the 50 S subunit is
subject to considerable conformational changes indifferent functional states (e.g. see Stark et al.,1997b). Another data set that is relevant here con-cerns the mapping by Wilson & Noller (1998b) ofFe(II)-induced cleavage sites on 23 S rRNA fromtethered EDTA molecules at various positions inEF-G. They found sites in h43-h44, h69 and h95 (cf.Figure 4(a)). In their Figure 7, Wilson & Noller(1998b) designate the sites as 1070, 1920 and 2660,respectively, and a comparison of our Figure 4(a)with their Figure 7 shows a broad level of agree-ment.
The conserved secondary structural core, andeukaryotic expansion segments
Ribosomal RNA molecules from different organ-isms vary considerably in length, and it is of inter-est to see where the ``missing'' or ``extra'' helicesrelative to the E. coli molecule are located in the23 S rRNA model. We have arbitrarily chosen thelarge subunit rRNA from the human mitochondrialribosome and the rat liver ribosome to make thiscomparison. Figure 9 shows the secondary struc-ture of the E. coli 23 S rRNA, with those helicesthat are still remaining in the human mitochondrialrRNA (Gutell & Fox, 1988) marked by bars. Thepoints in the structure at which the principalexpansion segments in the mammalian 28 S rRNAoccur are indicated (Gutell & Fox, 1988).
In the human mitochondrial rRNA (Figure 9),the whole of domain I is missing, with the excep-tion of the small helix h23. Domain IIa is largelypresent, as well as a signi®cant fraction of domainIIb; h43 and h44 are still present in the latterdomain, although h41 and h42 connecting them tothe rest of the structure are destroyed. Most ofdomain III is absent, except for h48, h49 and h59a,whereas the majority of domain IV is conserved. Indomain V, a lot of the rRNA corresponding to theL1 protuberance and the central protuberance ismissing, but the rest of the domain is largely pre-sent. In domain VI only the sarcin loop (h95) hassurvived. Notably, this conserved core in the mito-chondrial rRNA contains all the sequence regionswhere post-transcriptionally modi®ed nucleotidesare located (Figure 1). The conserved core ofhelices that is preserved in the human mitochon-drial rRNA is illustrated in Figure 10, in which thehelices are represented by cylinders, colour-codedaccording to the secondary structural domains asin Figures 3 and 4. The cylinders are superposedupon the semi-transparent contour of the 50 S sub-unit. It is clear that the core helices are clusteredaround the peptidyl transferase and L11 bindingareas of the 50 S subunit, whereas the peripheralregions are empty.
The locations of the expansion segments in ratliver 28 S rRNA would conversely be expected tolie on the periphery of the 50 S subunit. It isimmediately obvious from Figure 9 that the inser-tion points of these expansion segments are all out-side (or on the fringe of) the conserved secondary

Figure 9. The conserved core of the 23 S rRNA, and locations of eukaryotic expansion segments. The secondarystructure of the 23 S rRNA is shown, with black bars indicating the conserved core of secondary structure, as rep-resented by the rRNA from the human mitochondrial ribosome. Arrows with numbers indicate the approximateinsertion points of the principal expansion segments in rat liver 28 S rRNA, with the numbers giving the approximatenet size (in nucleotides) of the expansion segment concerned.
The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 53

Figure 10. Distribution of the conserved core of the 23 S rRNA in the 50 S subunit. The helices of the conservedcore (Figure 9) are displayed as cylinders, numbered and colour-coded as in Figures 3 and 4. The EM density of the50 S subunit is shown semi-transparent. (a) View from the L7/12 side of the subunit. (b) View from the interfaceside. (c) View from the L1 side. (d) View from the solvent side.
54 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
structural core, and their locations in the 23 SrRNA model can be seen by referring to Figures 3and 4. The largest expansion segment is in h25,and the position of this helix on the solvent side ofthe 50 S subunit is visible (coloured red) inFigure 4(c). Helix h18 (red) carrying a 60 nucleo-tide insertion, and h30 (yellow) with 90 extranucleotides, are visible in the same Figure, whereash38 (orange) with two 30 nucleotide insertions canbe seen in Figure 4 (d). Similarly, h45 (orange, with125 extra nucleotides), h52 (green, with 35) andh101 (dark blue, with 20) can be seen in Figure 4(c),with h98 (dark blue, with 215 nucleotides) inFigure 3(a) or Figure 4(c). The remaining insertionpoints, h54 (green, with 40 extra nucleotides), h63(light blue, with 560), h78 (magenta, with 30) andh79 (magenta, with 65) are visible in Figure 3(c).
In a recent publication (Dube et al., 1998) we cor-related the positions of the eukaryotic expansionsegments in a preliminary version of the 23 S
rRNA model with extra density elements relativeto the E. coli ribosome that were observed in anEM reconstruction at 21 AÊ resolution of the rabbitreticulocyte 80 S ribosome. The insertion sites weregrouped and named according to the positions ofE. coli 50 S ribosomal proteins that were cross-linked to sites on the 23 S rRNA close to the inser-tion points. A re-examination of these assignmentsshows that the extra EM density in the 80 S ribo-some described as the L13/L21 area by Dube et al.(1998) carrying the major insertion in h25 and thesmall insertions in h38 is in good agreement withour current 23 S rRNA model. The same applies tothe L19 area (h63), the L3/L32 area (h45, h98 andh101), and most of the L4 area (h18 and h30). Onthe other hand, h52 and h54 (with small insertionsassigned to the L4 area) and h78 and h79 (withrelatively small insertions assigned to the L1 area)are now at locations lower down in the 50 Ssubunit.

The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 55
Conclusions
The 3D arrangement for the 5 S and 23 S rRNAthat we have described here is not the ®nal struc-ture, but rather a step in the process of successiveapproximation to the structure, as outlined in theIntroduction. It will be evident to the reader thatsome areas of the rRNA molecules are more rigor-ously placed than others, and that complex regionssuch as the peptidyl transferase area or the groupsof helices connected by tertiary interactions (h5-h7,h82-87, etc.) will require a much higher level of res-olution for an accurate description of their struc-ture. Nevertheless, in our opinion it is veryimportant to be able to demonstrate a plausiblearrangement for the entire rRNA, including thoseareas that are less certain. Expressed in anotherway, the most common reason for rejecting apotential arrangement for the rRNA helices duringthe course of this work (and this happened manytimes) was when that arrangement, for one particu-lar group of helices, left no available space in theEM density for a contiguous helix or group ofhelices. In the structure described here, all of the5 S and 23 S rRNA could be accommodated in theEM density and, conversely, there is no case wherea helix or single strand has to traverse a canyon orgap in the density map (with the single exceptiondiscussed in relation to Figure 6(b)).
In terms of the interactions with functionalligands and other related topics, our structuregives a picture of the organization of the rRNA inthe 50 S subunit that is largely coherent and inwhich the great majority of the biochemical infor-mation could be incorporated. It is important, how-ever, to remember in this context that the variousbiochemical data sets have been collected under awide variety of experimental conditions, and as aresult a number of different conformational statesof the ribosome are likely to have been involved.Our primary concern here was to ®t the rRNA tothe EM density, which a priori re¯ects a single con-formational state. In order to ful®l this criterion, itwas occasionally unavoidable that the residuesinvolved, for example, in a particular cross-linkhad to be separated by a few AÊ ngstroÈm units. Inthis context, our structure helps to reveal areaswhere conformational changes in the rRNA arelikely to occur, and/or where there are potentialcon¯icts between different biochemical data points.The situation pertaining to proteins L7/12 is a casein point here because, as already mentioned, thereremains a direct controversy between the interpret-ations of the cryo-EM and X-ray density maps inthe L7/12 region of the subunit (Ban et al., 1999;Matadeen et al., 1999; M. van H., unpublished),which could have major consequences for thearrangement of the rRNA in this area; as notedabove, we have used the interpretation offered byBan et al. (1999) in our current model.
For the future, we can expect that the continueddevelopment of the cryo-EM techniques will enableus to improve our understanding of the ribosomal
function, by following conformational changes inthe ribosomes, or by observing the positions offunctional ligands at different stages in the proteinbiosynthetic cycle. The cryo-EM methodology canbe exploited to test directly the accuracy of therRNA structures, by making reconstructions ofmodi®ed ribosomes carrying helix extensions. Theviability of this method has been demonstrated(Spahn et al., 1999), and similar experiments are inprogress in our laboratories, in collaboration withDrs A. Bogdanov and O. Dontsova (Moscow). It isencouraging that the placements in the 50 S sub-unit of the two helices (h63 and h98) described bySpahn et al. (1999) are in precise agreement withthose that we have (entirely independently)derived here. Areas of the structure where moreinformation is needed are being investigated byapplication of the ``nick site'' cross-linking tech-nique (Baranov et al., 1998).
Materials and Methods
Ribosomal RNA model generation
The 5 S and 23 S rRNA structures were modelledinteractively into the 50 S cryo-EM reconstructionreported by Matadeen et al. (1999) using the programmeERNA-3D, exactly as described for 16 S rRNA byMueller & Brimacombe (1997a). Conversion of EM den-sity data into contour data for use with ERNA-3D wasperformed as described (Mueller & Brimacombe, 1997a).The modelling was conducted using a version of thecryo-EM reconstruction without high-pass ®ltering(Matadeen et al., 1999), as the high-pass ®ltering processappears to cause some attenuation of the electron densityin the internal regions of the 50 S subunit. Althoughomitting the ®ltering results in a small loss of resolution,it enabled us to correlate the connectivities in the EMdensity with the rRNA structure more easily in theseinternal regions.
Energy minimization
When the positions of helical elements of the rRNAare manipulated using ERNA-3D, the single strands con-necting them remain automatically connected. This hasthe consequence that in the interactively generated struc-ture the conformations of the single strands are arbitrary.The only exceptions are where structures of speci®crRNA regions are available at atomic resolution (such asthe sarcin-ricin loop, or the L11-rRNA and L25-rRNAbinding regions), in which case the structures concernedwere substituted into the appropriate section of the PDB®le of the 23 S or 5 S molecules and were manipulatedas rigid objects. For the computational re®nement of theinteractively generated model, we aimed at two goalssimultaneously: (1) computing a stereochemically plaus-ible structure for the single-stranded regions, and (2)incorporating the biochemical information such as cross-linking data or the cryo-EM density.
Technically, this was done by minimizing aCHARMM-like potential function E (Brooks et al., 1983),with additional terms for the biochemical informationadded. For the stereochemical plausibility we introducedbond-length, bond-angle and van der Waals terms (omit-ting for the sake of simplicity the usual dihedral-angle

56 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
and electrostatic terms). For the cross-link data, springswere added, to pull identi®ed residues into neighbouringpositions; and for the EM density, an additional potentialterm was interpolated from the voxel data. The interac-tive manipulation leads to a model in which the cross-link and EM density terms are satis®ed to a ®rst approxi-mation, and the helices do not signi®cantly overlap oneanother. The internal structures of the A-form helicalelements are correct, but there are many bond angles inthe backbone of the single strands that are distorted andthe highest energies result from Van der Waals con¯ictsbetween accidentally overlapping strands. In order tofacilitate the computational re®nement, groups of atomsare clustered into rigid objects. The potential E de®nesforces acting on the atoms, thus implying forces andmoments of forces acting on the rigid objects. By numeri-cal integration over time, motion of the objects is thusgenerated. This integration is adaptive, i.e. carried outwith a variable timestep, as the forces involved aredifferent by orders of magnitude. Since minimization isthe goal, the energy of the system needs to be reduced;this is done by frictional damping. Two single overlap-ping nuclei, yielding a potentially in®nite energy, might``explode'' the whole system. Accordingly, control of theoptimization process is important, and several moleculardynamics runs are started with different parameters.Initially, helices, residues and known structures (such asthat of the sarcin-ricin loop) are clustered, the helices arekept ®xed, and a very high frictional constant is used,which is then gradually reduced. At a certain point, thesingle-stranded residues are divided into subgroups(base, sugar, phosphate group and single oxygen atoms).Finally, the helices are released for minor adjustment.The whole process of clustered adaptive moleculardynamics simulation will be described in detail else-where.
Protein Data Bank accession numbers
The complete network of structures has been depos-ited with the RCSB PDB: accession numbers, 1C2W forthe 23 S rRNA, 1C2X for the 5 S rRNA and 487D for the®tted proteins.
Acknowledgements
We thank all members, past and present, of ourresearch groups, whose work in collecting the biochemi-cal data and developing the EM methodology have con-tributed to this study. Our thanks are due to thelaboratories of Drs A. Bogdanov and O. Dontsova (Mos-cow) and Dr D. Bochkariov (California) for the collabor-ations that have played an important role in the cross-linking studies. We are grateful to the Max-Planck-Gesellschaft for providing the computer graphics hard-ware. We gratefully acknowledge the ®nancial supportof the Deutsche Forschungsgemeinschaft (grant BR 632/3-4), the European Commission (Biotechnology Pro-gramme, no. BI04-CT97-2188) and the Volkswagen Foun-dation (grant nos I/72 338 and I/74 598). Last but notleast, we thank Frau Gabriele Eder for her patient helpin preparing the manuscript.
References
Adamski, F. M., Atkins, J. F. & Gesteland, R. F. (1996).Ribosomal protein L9 interacts with 23 S rRNA: theuse of a translational bypass assay to study theeffect of amino acid substitutions. J. Mol. Biol. 261,357-371.
Agrawal, R. K., Penczek, P., Grassucci, R. A., Li, Y.,Leith, A., Nierhaus, K. H. & Frank, J. (1996). Directvisualization of A-, P- and E-site transfer RNAs inthe E. coli ribosome. Science, 271, 1000-1002.
Agrawal, R. K., Penczek, P., Grassucci, R. A. & Frank, J.(1998). Visualization of elongation factor G on theE. coli 70 S ribosome: the mechanism of transloca-tion. Proc. Natl Acad. Sci. USA, 95, 6134-6138.
Agrawal, R. K., Penczek, P., Grassucci, R. A., Burkhardt,N., Nierhaus, K. H. & Frank, J. (1999). Effect of buf-fer conditions on the position of tRNA on the 70 Sribosome as visualized by cryoelectron microscopy.J. Biol. Chem. 274, 8723-8729.
Ban, N., Freeborn, B., Nissen, P., Penczek, P., Grassucci,R. A., Sweet, R., Frank, J., Moore, P. B. & Steitz,T. A. (1998). A 9 AÊ resolution X-ray crystallographicmap of the large ribosomal subunit. Cell, 93, 1105-1115.
Ban, N., Nissen, P., Hansen, J., Capel, M., Moore, P. B.& Steitz, T. A. (1999). Placement of protein andRNA structures into a 5 AÊ -resolution map of the50 S ribosomal subunit. Nature, 400, 841-847.
Baranov, P. V., Gurvich, O. L., Bogdanov, A. A.,Brimacombe, R. & Dontsova, O. A. (1998). New fea-tures of 23 S ribosomal RNA folding: the long helix41-42 makes a ``U-turn'' inside the ribosome. RNA,4, 658-668.
Baranov, P. V., Kubarenko, A. V., Gurvich, O. L.,Shamolina, T. A. & Brimacombe, R. (1999). Thedatabase of ribosomal cross-links: an update. Nucl.Acids Res. 27, 184-185.
Beckmann, R., Bubeck, D., Grassucci, R., Penczek, P.,Verschoor, A., Blobel, G. & Frank, J. (1997). Align-ment of conduits for the nascent polypeptide in theribosome-Sec61 complex. Science, 278, 2123-2126.
Bernabeu, C. & Lake, J. A. (1982). Nascent polypeptidechains emerge from the exit domain of the largeribosomal subunit: immune mapping of the nascentchain. Proc. Natl Acad. Sci. USA, 79, 3111-3115.
Brimacombe, R. (1995). The structure of ribosomal RNA:a three-dimensional jigsaw puzzle. Eur. J. Biochem.230, 365-383.
Brimacombe, R. (1999). The structure of ribosomal RNA.ASM News, 65, 144-151.
Brimacombe, R., Atmadja, J., Stiege, W. & SchuÈ ler, D.(1988). A detailed model for the three-dimensionalstructure of E. coli 16 S ribosomal RNA in situ inthe 30 S subunit. J. Mol. Biol. 199, 115-136.
Brimacombe, R., Mitchell, P., Osswald, M., Stade, K. &Bochkariov, D. (1993). Clustering of modi®ednucleotides at the fuctional center of bacterial ribo-somal RNA. FASEB J. 7, 161-167.
Brimacombe, R., Greuer, B., Mueller, F., Osswald, M.,Rinke-Appel, J. & Sommer, I. (2000). Three-dimen-sional organization of the bacterial ribosome and itssubunits: the transition from low-resolution modelsto high-resolution structures. In The Ribosome; Struc-ture, Function, Antibiotics and Cellular Interactions(Garrett, R. A., Douthwaite, S. R., Liljas, A.,Matheson, A. T., Moore, P. B. & Noller, H. F., eds),ASM Press DC, Washington. In the press.

The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 57
Brooks, B. R., Bruccoleri, R. E., Olafson, B. D., States,D. J., Swaminathan, S. & Karplus, M. (1983).CHARMM: a program for macromolecular energy,minimization and dynamics calculations. J. Comput.Chem. 4, 187-217.
Bullard, J. M., van Waes, M. A., Bucklin, D. J. & Hill,W. E. (1995). Regions of 23 S ribosomal RNA proxi-mal to transfer RNA bound at the P and E sites.J. Mol. Biol. 252, 572-582.
Capel, M. S., Kjeldgaard, M., Engelman, D. M. & Moore,P. B. (1988). Positions of S2, S13, S16, S17, S19 andS21 in the 30 S ribosomal subunit of E. coli. J. Mol.Biol. 200, 65-87.
Cate, J. H., Yusupov, M. M., Yusupova, G. Z., Earnest,T. N. & Noller, H. F. (1999). X-ray crystal structuresof 70 S ribosome functional complexes. Science, 285,2095-2104.
Choi, K. M. & Brimacombe, R. (1998). The path of thegrowing peptide chain through the 23 S rRNA inthe 50 S ribosomal subunit; a comparative cross-linking study with three different peptide families.Nucl. Acids Res. 26, 887-895.
Clemens, W. M., May, J. L. C., Wimberly, B. T.,McCutcheon, J. P., Capel, M. S. & Ramakrishnan, V.(1999). Structure of a bacterial 30 S ribosomal sub-unit at 5.5 AÊ resolution. Nature, 400, 833-840.
Conn, G. L., Draper, D. E., Lattman, E. E. & Gittis, A. G.(1999). Crystal structure of a conserved ribosomalprotein - RNA complex. Science, 284, 1171-1174.
Correll, C. C., Freeborn, B., Moore, P. B. & Steitz, T. A.(1997). Metals, motifs and recognition in the crystalstructure of a 5 S rRNA domain. Cell, 91, 705-712.
Correll, C. C., Munishkin, A., Chan, Y. L., Ren, Z.,Wool, I. G. & Steitz, T. A. (1998). Crystal structureof the ribosomal RNA domain essential for bindingelongation factors. Proc. Natl Acad. Sci. USA, 95,13436-13441.
Culver, G. M., Cate, J. H., Yusupova, G. Z., Yusupov,M. M. & Noller, H. F. (1999). Identi®cation of anRNA-protein bridge spanning the ribosomal sub-unit interface. Science, 285, 2133-2135.
Dallas, A. & Moore, P. B. (1997). The loop E-loop Dregion of E. coli 5 S rRNA: the solution structurereveals an unusual loop that may be important forbinding ribosomal proteins. Structure, 5, 1639-1653.
Davies, C., White, S. W. & Ramakrishnan, V. (1996). Thecrystal structure of ribosomal protein L14 reveals animportant organizational component of the transla-tional apparatus. Structure, 4, 55-66.
Dube, P., Bacher, G., Stark, H., Mueller, F., Zemlin, F.,van Heel, M. & Brimacombe, R. (1998). Correlationof the expansion segments in mammalian rRNAwith the ®ne structure of the rabbit reticulocyteribosome at 21 AÊ resolution. J. Mol. Biol. 279, 403-421.
Endo, Y. & Wool, I. G. (1982). The site of action of alphasarcin on eukaryotic ribosomes. J. Biol. Chem. 257,9054-9060.
Endo, Y., Mitsui, M., Motizuki, M. & Tsurugi, K. (1987).The mechanism of action of ricin and related toxiclectins on eukaryotic ribosomes. The site and thecharacteristics of the modi®cation in 28 S ribosomalRNA caused by the toxins. J. Biol. Chem. 262, 5908-5912.
Fox, G. E. & Woese, C. R. (1975). 5 S RNA secondarystructure. Nature, 256, 505-507.
Glotz, C., Zwieb, C., Brimacombe, R., Edwards, K. &KoÈssel, H. (1981). Secondary structure of thelarge subunit ribosomal RNA from E. coli, Z. mays
chloroplast, and human and mouse mitochondrialribosomes. Nucl. Acids Res. 9, 3287-3306.
Gnirke, A., GeigenmuÈ ller, U., Rheinberger, H. J. &Nierhaus, K. H. (1989). The allosteric three-sitemodel for the ribosomal elongation cycle: an anal-ysis with a heteropolymeric mRNA. J. Biol. Chem.264, 7291-7301.
Golden, B. L., Ramakrishnan, V. & White, S. W. (1993).Ribosomal protein L6: structural evidence of geneduplication from a primitive RNA binding protein.EMBO J. 12, 4901-4908.
Green, R. & Noller, H. F. (1997). Ribosomes and trans-lation. Annu. Rev. Biochem. 66, 679-716.
Green, R., Switzer, C. & Noller, H. F. (1998). Ribosome-catalyzed peptide-bond formation with an A-sitesubstrate covalently linked to 23 S ribosomal RNA.Science, 280, 286-288.
Greuer, B., Thiede, B. & Brimacombe, R. (1999). Thecross-link from the upstream region of mRNA toribosomal protein S7 is located in the C-terminalpeptide; experimental veri®cation of a predictionfrom modelling studies. RNA, 5, 1521-1525.
Gutell, R. R. & Fox, G. E. (1988). A compilation of largesubunit RNA sequences presented in a structuralformat. Nucl. Acids Res. 16, r175-r269.
Gutell, R. R., Larsen, N. & Woese, C. R. (1994). Lessonsfrom an evolving rRNA: 16 S and 23 S rRNA struc-tures from a comparative perspective. Microbiol.Rev. 58, 10-26.
Harms, J., Tocilj, A., Levin, I., Agron, I., KoÈ lln, I., Stark,H., van Heel, M., Cufe, M., SchluÈ nzen, F., Bashan,A., Franceschi, F. & Yonath, A. (1999). Elucidatingthe structure of ribosomal particles: an interplaybetween electron cryo-microscopy and X-ray crys-tallography. Structure, 7, 931-941.
Hinck, A. P., Markus, M. A., Huang, S., Grzesiek, S.,Kustonovich, I., Draper, D. E. & Torchia, D. A.(1997). The RNA binding domain of ribosomal pro-tein L11: three-dimensional structure of the RNA-bound form of the protein and its interaction with23 S rRNA. J. Mol., Biol. 274, 101-113.
Hoffman, D. W., Cameron, C. S., Davies, C., White,S. W. & Ramakrishnan, V. (1996). Ribosomal pro-tein L9: a structure determination by the combineduse of X-ray crystallography and NMR spec-troscopy. J. Mol. Biol. 264, 1058-1071.
Kim, S. H., Suddath, F. L., Quigley, G. J., McPherson,A., Sussman, J. L., Wang, A. H. J., Seeman, N. C. &Rich, A. (1974). Three-dimensional tertiary structureof yeast phenylalanine transfer RNA. Science, 185,435-440.
Kudlicki, W., Chirgwin, J., Kramer, G. & Hardesty, B.(1995). Folding of an enzyme into an active confor-mation while bound as peptidyl tRNA to the ribo-some. Biochemistry, 34, 14284-14287.
Leffers, H., Kjems, J., éstergaard, L., Larsen, N. &Garrett, R. A. (1987). Evolutionary relationshipsamongst archaebacteria: a comparative study of23 S ribosomal RNAs of a sulphur-dependentextreme thermophile, an extreme halophile and athermophilic methanogen. J. Mol. Biol. 195, 43-61.
Leijonmarck, M. & Liljas, A. (1987). Crystal structure ofthe C-terminal domain of the ribosomal protein L7/L12 from E. coli at 1.7 AÊ . J. Mol. Biol. 195, 555-579.
Leontis, N. B. & Westhof, E. (1998). A common motiforganizes the structure of multihelix loops in 16 Sand 23 S ribosomal RNAs. J. Mol. Biol. 283, 571-583.

58 The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ
Makeyev, E. V., Kolb, V. A. & Spirin, A. S. (1996).Enzymatic activity of the ribosome bound nascentpeptide. FEBS Letters, 378, 166-170.
Malhotra, A. & Harvey, S. C. (1994). A quantitativemodel of the E. coli 16 S RNA in the 30 S riboso-mal subunit. J. Mol. Biol. 240, 308-340.
Malhotra, A., Penczek, P., Agrawal, R. K., Gabashvili,I. S., Grassucci, R. A., JuÈ nemann, R., Burkhardt, N.,Nierhaus, K. H. & Frank, J. (1998). E. coli 70 Sribosome at 15 AÊ resolution by cryo-electronmicroscopy: localization of fMet-tRNAf
Met and ®t-ting of L1 protein. J. Mol. Biol. 280, 103-116.
Markus, M. A., Hinck, A. P., Huang, S., Draper, D. E. &Torchia, D. A. (1997). High resolution solutionstructure of ribosomal protein L11-C76, a helicalprotein with a ¯exible loop that becomes structuredupon binding to RNA. Nature Struct. Biol. 4, 70-77.
Matadeen, R., Patwardhan, A., Gowen, B., Orlova, E.,Stark, H., Pape, T., Cuff, M., Mueller, F.,Brimacombe, R. & van Heel, M. (1999). The E. colilarge ribosomal subunit at 7.5 AÊ resolution. Struc-ture, 7, 1575-1583.
May, R. P., Nowotny, V., Nowotny, P., Voss, H. &Nierhaus, K. H. (1992). Interprotein distances withinthe large subunit from E. coli ribosomes. EMBO J.11, 373-378.
Mitchell, P., Osswald, M., Schueler, D. & Brimacombe,R. (1990). Selective isolation and detailed analysis ofintra-RNA cross-links induced in the large riboso-mal subunit of E. coli; a model for the tertiary struc-ture of the tRNA binding domain in 23 S RNA.Nucl. Acids Res. 18, 4325-4333.
Mitchell, P., Osswald, M. & Brimacombe, R. (1992).Identi®cation of intermolecular cross-links at thesubunit interface of the E. coli ribosome. Biochemis-try, 31, 3004-3011.
Mitchell, P., Stade, K., Osswald, M. & Brimacombe, R.(1993). Site-directed cross-linking studies on the E.coli tRNA-ribosome complex; determination of siteslabelled with an aromatic azide attached to the vari-able loop or aminoacyl group of tRNA. Nucl. Acids.Res. 21, 887-896.
Moazed, D. & Noller, H. F. (1989). Interaction of tRNAwith 23 S rRNA in the ribosomal A, P and E sites.Cell, 57, 585-597.
Moazed, D. & Noller, H. F. (1991). Sites of interaction ofthe CCA end of peptidyl-tRNA with 23 S rRNA.Proc. Natl Acad. Sci. USA, 88, 3725-3728.
Moazed, D., Robertson, J. M. & Noller, H. F. (1988).Interaction of elongation factors EF-G and EF-Tuwith a conserved loop in 23 S RNA. Nature, 334,362-364.
Mueller, F. & Brimacombe, R. (1997a). A new model forthe three-dimensional folding of E. coli 16 S riboso-mal RNA. I. Fitting the RNA to a 3D electronmicroscopic map at 20 AÊ . J. Mol. Biol. 271, 524-544.
Mueller, F. & Brimacombe, R. (1997b). A new model forthe three-dimensional folding of E. coli 16 S riboso-mal RNA. II. The RNA-protein interaction data.J. Mol. Biol. 271, 545-565.
Mueller, F., Stark, H., van Heel, M., Rinke-Appel, J. &Brimacombe, R. (1997). A new model for the three-dimensional folding of E. coli 16 S ribosomal RNA.III. The topography of the functional centre. J. Mol.Biol. 271, 566-587.
Munishkin, A. & Wool, I. G. (1997). The ribosome-in-pieces: binding of elongation factor EF-G to oligo-ribonucleotides that mimic the sarcin/ricin and
thiostrepton domains of 23 S ribosomal RNA. Proc.Natl Acad. Sci. USA, 94, 12280-12284.
Nagano, K., Harel, M. & Takezawa, M. (1988). Pre-diction of three-dimensional structure of E. coliribosomal RNA. J. Theoret. Biol. 134, 199-256.
Nakagawa, A., Nakashima, T., Taniguchi, M., Hosaka,H., Kimura, M. & Tanaka, I. (1999). The three-dimensional structure of the RNA-binding domainof ribosomal protein L2; a protein at the peptidyltransferase center of the ribosome. EMBO J. 18,1459-1467.
Nikonov, S., Nevskaya, N., Eliseikina, I., Fomenkova,N., Nikulin, A., Ossina, N., Garber, M., Jonsson,B. H., Briand, C., Al-Karadaghi, S., Svensson, A.,ávarsson, A. & Liljas, A. (1996). Crystal structureof the RNA binding ribosomal protein L1 fromThermus thermophilus. EMBO J. 15, 1350-1359.
Nissen, P., Kjeldgaard, M., Thirup, S., Polekhina, G.,Reshetnikova, L., Clark, B. F. C. & Nyborg, J.(1995). Crystal structure of the ternary complex ofPhe-tRNAPhe, EF-Tu and a GTP analog. Science, 270,1464-1472.
Noller, H. F., Kop, J. A., Wheaton, V., Brosius, J., Gutell,R. R., Kopylov, A. M., Dohme, F., Herr, W., Stahl,D. A., Gupta, R. & Woese, C. R. (1981). Secondarystructure model for 23 S ribosomal RNA. Nucl.Acids Res. 9, 6167-6189.
Ofengand, J. & Fournier, M. J. (1998). The pseudouridineresidues of rRNA: number, location, biosynthesis,and function. In Modi®cation and Editing of RNA(Grosjean, H. & Benne, R., eds), pp. 229-253, ASMPress, Washington, DC.
Osswald, M. & Brimacombe, R. (1999). The environmentof 5 S rRNA in the ribosome: cross-links to 23 SrRNA from sites within helices II and III of the 5 Smolecule. Nucl. Acids Res. 27, 2283-2290.
Osswald, M., DoÈring, T. & Brimacombe, R. (1995). Theribosomal neighbourhood of the central fold oftRNA; cross-links from position 47 of tRNA locatedat the A, P or E site. Nucl. Acids Res. 23, 4635-4641.
Puglisi, E. V., Green, R., Noller, H. F. & Puglisi, J. D.(1997). Structure of a conserved RNA component ofthe peptidyl transferase centre. Nature Struct. Biol. 4,775-778.
Rinke-Appel, J., JuÈ nke, N., Osswald, M. & Brimacombe,R. (1995). The ribosomal environment of tRNA:cross-links to rRNA from positions 8 and 20:1 inthe central fold of tRNA located at the A, P or Esite. RNA, 1, 1018-1028.
Ryabova, L. A., Selivanova, O. M., Baranov, V. I.,Vasiliev, V. D. & Spirin, A. S. (1988). Does the chan-nel for nascent peptide exist inside the ribosome?Immune electron microscopy study. FEBS Letters,226, 255-260.
Samaha, R. R., Green, R. & Noller, H. F. (1995). A base-pair between tRNA and 23 S rRNA in the peptidyltransferase centre of the ribosome. Nature, 377, 309-314.
Sergiev, P., Dokudovskaya, S., Romanova, E., Topin, A.,Bogdanov, A., Brimacombe, R. & Dontsova, O.(1998). The environment of 5 S rRNA in the ribo-some: cross-links to the GTPase-associated area of23 S rRNA. Nucl. Acids Res. 26, 2519-2525.
Smith, J. E., Cooperman, B. S. & Mitchell, P. (1992).Methylation sites in E. coli ribosomal RNA: localiz-ation and identi®cation of four new sites of methyl-ation in 23 S rRNA. Biochemistry, 31, 10825-10834.
Spahn, C. M. T., Grassucci, R. A., Penczek, P. & Frank,J. (1999). Direct three-dimensional localization and

The 3D Arrangement of 23 S and 5 S rRNA at 7.5 AÊ 59
positive identi®cation of RNA helices within theribosome by means of genetic tagging and cryo-electron microscopy. Structure, 7, 1567-1573.
Stade, K., Riens, S., Bochkariov, D. & Brimacombe, R.(1994). Contacts between the growing peptide chainand the 23 S RNA in the 50 S ribosomal subunit.Nucl. Acids Res. 22, 1394-1399.
Stade, K., JuÈ nke, N. & Brimacombe, R. (1995). Mappingthe path of the nascent peptide chain through the23 S RNA in the 50 S ribosomal subunit. Nucl.Acids Res. 23, 2371-2380.
Stark, H., Mueller, F., Orlova, E. V., Schatz, M., Dube,P., Erdemir, T., Zemlin, F., Brimacombe, R. & vanHeel, M. (1995). The 70 S E. coli ribosome at 23 AÊ
resolution; ®tting the ribosomal RNA. Structure, 3,815-821.
Stark, H., Orlova, E. V., Rinke-Appel, J., JuÈ nke, N.,Mueller, F., Rodnina, M., Wintermeyer, W.,Brimacombe, R. & van Heel, M. (1997a). Arrange-ment of tRNAs in pre- and post-translocationalribosomes revealed by electron cryomicroscopy.Cell, 88, 19-28.
Stark, H., Rodnina, M., Rinke-Appel, J., Brimacombe, R.,Wintermeyer, W. & van Heel, M. (1997b). Visualiza-tion of elongation factor Tu on the E. coli ribosome.Nature, 389, 403-406.
Steiner, G., Kuechler, E. & Barta, A. (1988). Photo-af®-nity labelling at the peptidyl transferase centrereveals two different positions for the A- and P-sites in domain V of 23 S rRNA. EMBO J. 7, 3949-3955.
Stern, S., Weiser, B. & Noller, H. F. (1988). Model forthe three-dimensional folding of 16 S ribosomalRNA. J. Mol. Biol. 204, 447-481.
StoÈ f¯er-Meilicke, M. & StoÈ f¯er, G. (1990). Topographyof the ribosomal proteins from E. coli within theintact subunits as determined by immunoelectronmicroscopy and protein-protein cross-linking. In TheRibosome; Structure, Function and Evolution (Hill,W. E., Dahlberg, A., Garrett, R. A., Moore, P. B.,Schlessinger, D. & Warner, J. R., eds), pp. 123-133,ASM Press, Wahington, DC.
Stoldt, M., WoÈhnert, J., GoÈrlach, M. & Brown, L. (1998).The NMR structure of E. coli ribosomal protein L25shows homology to general stress proteins and glu-taminyl-tRNA synthetases. EMBO J. 17, 6377-6394.
Stoldt, M., WoÈhnert, J., OhlenschlaÈger, O., GoÈrlach, M.& Brown, L. R. (1999). The NMR structure of the5 S rRNA E-domain-protein L25 complex showspre-formed and induced recognition. EMBO J. 18,6508-6521.
Szewczak, A. A. & Moore, P. B. (1995). The sarcin/ricinloop, a modular RNA. J. Mol. Biol. 247, 81-98.
Tanaka, I., Nakagawa, A., Hosaka, H., Wakatsuki, S.,Mueller, F. & Brimacombe, R. (1998). Matching thecrystallographic structure of ribosomal protein S7 toa three-dimensional model of the 16 S ribosomalRNA. RNA, 4, 542-550.
Thiede, B., Urlaub, H., Neubauer, H., Grelle, G. &Wittmann-Liebold, B. (1998). Precise determinationof RNA-protein contact sites in the 50 S ribosomalsubunit of E. coli. Biochem. J. 334, 39-42.
Thompson, J., Schmidt, F. & Cundliffe, E. (1982). Site ofaction of a ribosomal RNA methylase conferringresistance to thiostrepton. J. Biol. Chem. 257, 7915-7917.
Tocilj, A., SchluÈ nzen, F., Janell, D., GluÈ hmann, M.,Hansen, H., Harms, J., Bashan, A., Bartels, H.,
Agmon, I., Franceschi, F. & Yonath, A. (1999). Thesmall ribosomal subunit from Thermus thermophilusat 4.5 AÊ resolution. Pattern ®ttings and the identi®-cation of a functional site. Proc. Natl Acad. Sci. USA,96, 14252-14257.
Unge, J., AÊ berg, A., Al-Kharadaghi, S., Nikulin, A.,Nikonov, S., Davydova, N. L., Nevskaya, N.,Garber, M. & Liljas, A. (1998). The crystal structureof ribosomal protein L22 from Thermus thermophilus:insights into the mechanism of erythromycin resist-ance. Structure, 6, 1577-1586.
Vester, B. & Garrett, R. A. (1988). The importance ofhighly conserved nucleotides in the binding regionof chloramphenicol at the peptidyl transferasecentre of E. coli 23 S ribosomal RNA. EMBO J. 7,3577-3587.
Wagenknecht, T., Carazo, J. M., Radermacher, M. &Frank, J. (1989). Three-dimensional reconstruction ofthe ribosome from E. coli. Biophys. J. 55, 455-464.
Walleczek, J., SchuÈ ler, D., StoÈ f¯er-Meilicke, M.,Brimacombe, R. & StoÈ f¯er, G. (1988). A model forthe spatial arrangement of the proteins in the largesubunit of the E. coli ribosome. EMBO J. 7, 3571-3576.
Wilson, K. S. & Noller, H. F. (1998a). Molecular move-ment inside the translational engine. Cell, 92, 337-349.
Wilson, K. S. & Noller, H. F. (1998b). Mapping the pos-ition of translational elongation factor EF-G in theribosome by directed hydroxyl radical probing. Cell,92, 131-139.
Wilson, K. S., Appelt, K., Badger, J., Tanaka, I. & White,S. W. (1986). Crystal structure of a prokaryoticribosomal protein. Proc. Natl Acad. Sci. USA, 83,7251-7255.
Wimberly, B. T., Guymon, R., McCutcheon, J. P., White,S. & Ramakrishnan, V. (1999). A detailed view of aribosomal active site: the structure of the GTPasecenter at 2.6 AÊ resolution. Cell, 97, 491-502.
Wower, J., Hixson, S. S. & Zimmermann, R. A. (1989).Labeling the peptidyl transferase center of theE. coli ribosome with photoreactive tRNAPhe deriva-tives containing azidoadenosine at the 30 end of theacceptor arm; a new model of the tRNA-ribosomecomplex. Proc. Natl Acad. Sci. USA, 86, 5232-5236.
Edited by D. E. Draper
(Received 7 January 2000; received in revised form15 February 2000; accepted 15 February 2000)
http://www.academicpress.com/jmb
Supplementary material comprising Tables of datasets, two Figures and a detailed description of thetRNA structure is available from JMB Online