tesis aislado de quinua a9 -...
TRANSCRIPT

UNIVERSIDAD DE CHILE FACULTAD DE CIENCIAS QUÍMICAS Y FARMACÉUTICAS
DEPARTAMENTO DE CIENCIA DE LOS ALIMENTOS Y
TECNOLOGÍA QUÍMICA
Profesor Patrocinante:
Lilian Elizabeth Abugoch James Departamento de Ciencia de los Alimentos y
Tecnología Química, Universidad de Chile
Directores de Memoria:
Lilian Elizabeth Abugoch James Departamento de Ciencia de los Alimentos y
Tecnología Química, Universidad de Chile
Nalda Marcela Romero Palacios Departamento de Ciencia de los Alimentos y
Tecnología Química, Universidad de Chile
OBTENCION, CARACTERIZACIÓN ESTRUCTURAL Y DETERMINACIÓN DE LAS PROPIEDADES FUNCIONALES DE UN AISLADO PROTEICO DE
QUINUA ORGÁNICA (CHENOPODIUM QUINOA)
Memoria para optar al Título Profesional de
Ingeniero en Alimentos
MONICA MIREYA RIVERA FIGUEROA
Santiago, Chile
2006

II
Todo nuestro descontento por
aquello de lo que carecemos procede de nuestra falta de gratitud por lo que tenemos.
Daniel Defoe
A mi amada familia

III
La fase experimental de esta memoria fue realizada en el Laboratorio de
Procesos de Alimentos del Departamento de Ciencia de los Alimentos y Tecnología
Química, dependiente de la Facultad de Ciencias Químicas y Farmacéuticas de la
Universidad de Chile.
El financiamiento para esta investigación fue otorgado por la Universidad de
Chile, a través del concurso interno de tesis del año 2006.

IV
AGRADECIMIENTOS
Agradezco de todo corazón a todos quienes me han apoyado durante mi
carrera y a quienes han hecho posible la realización de esta memoria:
A mi profesora patrocinante y directora de memoria, Lilian Abugoch, por su
dedicación, enseñanzas, consejos y su inagotable buena disposición.
A la profesora Nalda Romero, por sus enseñanzas, sus correcciones y su
valioso aporte a esta tesis.
A la profesora María Eugenia Letelier por facilitar la centrífuga sin la cual
hubiese sido imposible la obtención del aislado. Por su generosa ayuda y tiempo,
gracias.
Al profesor Jorge Chavez, por facilitar el secador spray, y a Alejandra Olave,
por enseñarme a usarlo.
Al profesor Abel Guarda por facilitar el equipo DSC, para la realización de los
análisis.
A todos los miembros de los laboratorios de: Procesos, Química de
Alimentos y Operaciones Unitarias, por su gran disposición y valiosa ayuda.
A las profesoras miembros de la comisión evaluadora de mi tesis, Lucía
Collados y Paz Robert, por su aporte y sugerencias a mi trabajo.
A todos los profesores sin excepción, quienes me entregaron más que
conocimientos durante la carrera.
A mis compañeros, muchas gracias por su ayuda y compañerismo.
A mi muy querido amigo Jorge Silva, por su apoyo, compañía y ayuda
durante el desarrollo de esta tesis, mil gracias.
A mi familia, en especial a mis padres, Waldo y Mireya, por inculcarme tantos
valores (en especial el del estudio), y por su constante apoyo durante toda mi
existencia.
A mi queridísima tía Iris, quien incluso en la lejanía me dio la fuerza para
seguir siempre adelante.
A Sosky, Kitty y Nelson, sin cuyo apoyo hubiera sido imposible culminar esta
etapa de mi vida. Siempre les estaré en deuda.

V
INDICE
RESUMEN ............................................................................................................... VII
SUMMARY .............................................................................................................. VIII
I. INTRODUCCIÓN.....................................................................................................1
1.1 Antecedentes generales ..............................................................................1
1.2 Características de la quinua ........................................................................2
1.2.2 Composición de la semilla de quinua y su valor nutricional.................3
1.2.3 Las Saponinas .....................................................................................5
1.3 Empleos de la quinua ..................................................................................6
1.4 Mercado nacional e internacional ................................................................6
1.5 Los aislados proteicos y sus propiedades ...................................................7
II. HIPÓTESIS .............................................................................................................9
III. OBJETIVOS ........................................................................................................10
3.1 Objetivo general.........................................................................................10
3.2 Objetivos específicos.................................................................................10
IV. MATERIALES Y MÉTODOS................................................................................11
4.1 Materiales ..................................................................................................11
4.1.1 Materia prima.....................................................................................11
4.1.2 Reactivos químicos............................................................................11
4.1.3 Equipos e instrumentos .....................................................................12
4.1.4 Insumos y utensilios...........................................................................14
4.2 Métodos .....................................................................................................15
4.2.1 Obtención de la harina de quinua desgrasada ..................................15
4.2.2 Obtención del aislado proteico de quinua..........................................16
4.2.3 Caracterización química ....................................................................18
4.2.3.1 Composición Proximal……………………….……………………….…. 18
4.2.3.2 Perfil de aminoácidos…………………………………………………… 18
4.2.3.3 Determinación de fitoestrógenos………………………………….…… 19

VI
4.2.4 Caracterización estructural de las fracciones proteicas.....................19
4.2.4.1 Perfiles polipeptídicos……………………………………………...…… 19
4.2.4.2 Fluorescencia……………………………………………………………. 20
4.2.4.3 Espectroscopía UV……………………………………………………… 21
4.2.4.4 Calorimetría Diferencial de Barrido………………………………….… 21
4.2.5 Determinación de las propiedades funcionales .................................21
4.2.5.1 Solubilidad…………………………………………………………………21
4.2.5.2 Capacidad de retención de agua………………………………........... 22
4.2.5.3 Capacidad de absorción de agua……………………………………….22
4.2.6 Análisis estadístico ............................................................................23
V. RESULTADOS Y DISCUSIÓN ............................................................................24
5.1 Caracterización química del aislado proteico de quinua ...........................24
5.1.1 Composición Proximal .......................................................................24
5.1.2 Perfil de aminoácidos.........................................................................25
5.1.3 Contenido de fitoestrógenos ..............................................................27
5.2 Caracterización estructural de las fracciones proteicas ............................28
5.2.1 Perfiles Polipeptídicos del Aislado Proteico de Quinua A9................28
5.2.2 Fluorescencia.....................................................................................31
5.2.3 Espectroscopia de absorción UV.......................................................33
5.2.4 Calorimetría diferencial de Barrido ....................................................35
5.3 Determinación de las propiedades funcionales .........................................37
5.3.1 Solubilidad .........................................................................................37
5.3.2 Capacidad de retención de agua .......................................................39
5.3.3 Capacidad de absorción de agua ......................................................41
VI. CONCLUSIONES ...............................................................................................43
VII. REFERENCIAS BIBLIOGRÁFICAS...................................................................45
ANEXOS ...................................................................................................................53
Anexo 1. Métodos electroforéticos (PAGE)...........................................................54
Anexo 2. Método de Bradford................................................................................58
Anexo 3. Preparación de los buffers .....................................................................60
Anexo 4. Gradientes de concentración para fase móvil de HPLC………..……… 61

VII
RESUMEN
El presente estudio tuvo por objetivo la obtención de un aislado proteico de
quinua a partir de quinua orgánica proveniente de la VI Región. El aislado se
caracterizó desde el punto de vista químico, estructural y funcional. Los resultados
se analizaron estadísticamente mediante análisis de varianza y test de Tukey, al 95
% de confianza.
El contenido de proteínas totales del aislado fue de 77,2%. El perfil de
aminoácidos coincidió con lo descrito en la literatura para la harina de quinua, por lo
que la extracción de las proteínas a pH 9 no afectó el balance de aminoácidos,
Entre los aminoácidos esenciales, destacan el alto contenido en lisina, metionina y
arginina. Los perfiles polipeptídicos en condiciones nativa y desnaturante, sin y con
2-Me, indican que las proteínas del aislado están compuestas principalmente por
albúminas del tipo 2S y globulinas 11S. A través de los resultados de
espectroscopia UV se verificó la presencia de globulinas. En fluorescencia se
observó curvas muy similares para los pHs alcalinos, lo que indicaría que la
estructura de la proteína no sufrió modificaciones en tales condiciones. Los
resultados de la calorimetría diferencial de barrido, señalan un cierto grado de
estructuración de la proteína, presentándose una endoterma de 13,5 mJ y una
temperatura de desnaturalización (td) de 98,1ºC, para el aislado suspendido en
agua. La endoterma fue mayor para la misma muestra suspendida en buffer de pH 9
(29,1 mJ y td de 100,5ºC), lo que señala que a pH 9 se produce una reestructuración
de las proteínas. El estudio de las propiedades funcionales muestran que el aislado
de quinua posee un alto nivel de solubilidad, especialmente sobre el pH 5, con una
solubilidad de 76,6%, la cual aumentó con el pH hasta alcanzar un máximo de
94,6%, a pH 11. La capacidad de retención de agua (WHC) fue buena, con valores
entre 2,5 y 3,0 mL/g aislado proteico, no encontrándose una dependencia del pH.
En absorción de agua se obtuvo resultados relativamente elevados, con una
capacidad máxima de absorción (WIC) de 1,8 g agua/g aislado, y una velocidad
inicial de 1,34 g agua/ g aislado × min. Todas estas características apuntan a que el
aislado es apropiado para la elaboración de productos tales como: bebidas, sopas,
embutidos, geles, productos de panificación, alimentos deshidratados y, en general,
en el desarrollo de productos funcionales altamente proteicos.

VIII
SUMMARY
OBTAINING, STRUCTURAL CHARACTERIZATION AND DETERMINATION OF THE FUNCTIONAL PROPERTIES OF A PROTEIN ISOLATE OF ORGANIC
QUINUA (Chenopodium quinoa)
The present study had by objective the obtention of a quinoa protein isolate
of organic quinoa, originated at the VI Region. The isolate was characterized from
the chemical, structural and functional point of view. The results were statistically
analyzed by analysis of variance and test of Tukey, at 95 % of confidence.
The total protein content of the isolate was 77.2%. The amino acids
composition corresponded to that described in references for the flour of quinoa,
reason why the extraction of proteins to pH 9 did not affect the balance of amino
acids, between the essential amino acids, emphasize the high content of lisine,
metionine and arginine. The polypeptide profiles in conditions native and denaturing,
without and with 2-Me, indicate that the proteins of the isolate are compound mainly
of albumins of the type 2S and globulins 11S. Through the results of spectroscopy
UV the presence of globulins was verified. In fluorescence it was observed very
similar curves at alkaline pHs, which would indicate that the structure of the protein
did not undergo modifications in such conditions. The results of the differential
scanning calorimetry, indicate a certain degree of structuring of protein, appearing
one endotherm of 13.5 mJ and one temperature of denaturation (td) of 98,1ºC, for
the isolate suspended in water. Endotherm was greater for the same sample
suspended in pH 9 buffer (29.1 mJ and td of 100.5ºC), which indicates that at pH 9 a
reconstruction of proteins takes place. The study of the functional properties
indicates that the isolate one of quinoa has a high level of solubility, especially at pH
5, with a value of 76.6%, which increased with pH until reaching a maximum of
94.6%, at pH 11. The water retention capacity (WHC) was good, with values
between 2.5 and 3.0 mL/g isolate protein, not being a dependency of pH. In water
absorption one obtained relatively elevated results, with a maximum capacity of
absorption (WIC) 1.8 g water/g of isolate, and one initial speed of 1.34 g water/g
isolate × min. All these characteristics aims at that the isolate is appropriate for the
product elaboration such as: drinks, soups, inlays, gels, baking industry, dehydrated
foods, and, in general, in the development of protein high functional products.

1
I. INTRODUCCIÓN
1.1 Antecedentes Generales
Las proteínas son macromoléculas que contienen carbono, nitrógeno,
oxígeno y, casi todas, azufre. En algunas se ha encontrado fósforo, hierro, zinc y
cobre (Robinson, 1991). Son las moléculas más abundantes en el interior de las
células y son fundamentales en cuanto a su estructura y función, proporcionando los
materiales que constituyen los músculos, huesos, glándulas, órganos internos,
sistema nervioso, sangre y otros líquidos del cuerpo, como así también la piel, el
cabello y las uñas. Son los instrumentos moleculares mediante los cuales se
expresa la información genética; además, desempeñan importantes funciones como
la catálisis, la regulación metabólica y los procesos contráctiles (Braverman, 1980;
Pennacchiotti, 1998).
Además de su función nutricional para cubrir necesidades energéticas y de
constitución, las proteínas desempeñan una función esencial en la apetencia del
alimento, es decir, en sus características organolépticas (Braverman, 1980;
Pennacchiotti, 1998). A partir de esta necesidad, se desarrollaron los procesos
necesarios para aislar o extraer las proteínas de sus fuentes orgánicas originales.
Se obtienen así los denominados aislados proteicos, que constituyen un purificado
proteico a partir del alimento o fuente orgánica inicial. De esta manera, se ha llegado
a obtener un macronutriente purificado con papel nutricional en la alimentación
propiamente dicha, y con un papel funcional en la elaboración tecnológica e
industrial de alimentos (Curare, 2006).
Entre los alimentos que entregan proteínas de alta calidad biológica (es
decir, aportan los aminoácidos esenciales) se encuentran la leche, la carne, el
pescado y el huevo (3,2; 20–23; 15-20 y 13,5%, respectivamente). Sin embargo, los
granos de los cereales son una buena fuente de proteína para la mayoría de la
población mundial, siendo consumidos en cantidades relativamente grandes en las
zonas sub-urbanas (FAO, 1970).
En la búsqueda de nuevas fuentes de alimentos que pudieran servir para
solucionar en parte el déficit proteico-calórico, en los años ’60 se inició el estudio de
un pseudo cereal denominado quinoa, quinua o quingua. La quinua (Chenopodium

2
quinoa willd) es un nutritivo pseudocereal que se ha cultivado en forma tradicional
en el área andina desde la época incásica. Fue ampliamente usado en la
alimentación de los pueblos antiguos de Sudamérica como uno de los alimentos
básicos. Sin embargo, el cultivo de la quinua en el altiplano disminuyó después de
la conquista española, cediendo el paso a cereales introducidos como el trigo y la
cebada (Wahli, 1990).
1.2 Características de la Quinua
La quinua es una planta anual herbácea que alcanza alturas entre uno y dos
metros (ver Figura 1); y que presenta acumulaciones de pequeñas semillas en
panojas, ubicadas en los extremos superiores de las ramificaciones. Las semillas
son bastante pequeñas, de alrededor de 1,5 mm de diámetro, aunque es posible
encontrar variedades que llegan hasta 4 mm (Junge y Cerda, 1978).
Figura 1: Planta de Quinua (Chenopodium quinoa willd)
Desde tiempos ancestrales la quinua se cultiva en la región del altiplano
andino de América del Sur. En la actualidad las mayores áreas productivas
corresponden a Perú y Bolivia, aunque también se produce en Colombia, Argentina,
Chile y Ecuador.
Cabe señalar que es un cultivo que se adapta a condiciones muy variables,
pudiéndose cultivar hasta los 3.900 metros sobre el nivel del mar (CIED, 2006). Por
otra parte, y debido a que posee raíces pivotantes y fasciculadas, se adapta bien al
clima frío y a la escasez de humedad, puesto que las raíces pivotantes aprovechan
el agua a mayor profundidad y las raíces fasciculadas el agua superficial (Fontúrbel,
2003).

3
Es interesante notar que la quinua es una planta de la que se aprovecha
todo. Los tallos rojos o amarillos tienen mucha fibra y, como el grano, son buenos
para los animales: ganan peso y producen más leche. Las hojas tiernas de la planta
permiten preparar sopas y ensaladas y, tras sacar el grano de las panojas, la
cascarilla que envuelve a cada quinua se la quema y, con ella, se elabora la “pasa”
o lejía, utilizada en la masticación de coca (Azcui, 2005).
Por otra parte, la quinua es evolutivamente distinta a aquellos cereales que
contienen gluten, fracción tóxica para los celiacos. En la harina de quinua, muy
bajas cantidades de proteínas se encuentran como prolaminas (0,8%), mientras que
albúminas (31%) y globulinas (37%) son predominantes (Berti y col., 2004).
1.2.2 Composición de la Semilla de Quinua y su Valor Nutricional
Tal como se señalara, toda la planta de quinua tiene diferentes usos, sin
embargo el producto primario es la semilla (ver Figura 2). Luego que se realizaran
análisis bromatológicos de la composición del grano y se divulgara esta información,
la quinua ha adquirido importancia internacional por ser uno de los pocos alimentos
de origen vegetal que es rico en proteínas y posee todos los aminoácidos
esenciales para el ser humano (ver la composición del grano en la tabla 1). También
contiene ácidos grasos esenciales como los ácidos grasos insaturados, destacando
su alto contenido de ácido linoleico (50,2-56,1%) y oleico (22,0-24,5%), y moderado
de linolénico (5,4-7%) (Walhi, 1990; Ruales y Nair, 1992). Asimismo, la quinua
posee un alto contenido de vitaminas del complejo B, C y E, además de minerales
tales como: hierro, fósforo, potasio y calcio. Este último se encuentra en la misma
concentración que en la leche descremada, mientras que el fósforo es cuatro veces
más concentrado que el de ésta (Albarrán, 1993).
Figura 2: Semilla de la quinua

4
Tabla 1: Composición Proximal de la semilla de quinua
Contenido g/100 g de semillaCalorías 331 Humedad 9,8 Proteína 13,0* Lípidos 7,4 ENN 64,1** Fibra cruda 2,7 Cenizas 3,0 * N x 5,7 ** Por diferencia
Fuente: Schmidt-Hebbel y col., 1992
El alto valor nutritivo de la semilla de quinua se debe tanto a su composición
química, como a la cantidad y calidad de sus proteínas, que fluctúa entre un 12 y
22%.Como se puede apreciar en la tabla 2, los aminoácidos que contiene la quinua
son en mayor cantidad, ácido glutámico, ácido aspártico, isoleucina, lisina,
fenilalanina, tirosina y valina por unidad de nitrógeno con respecto a otros cereales.
Tabla 2: Contenido de aminoácidos en el grano de quinua, el trigo y la leche (% de aminoácidos/100 g de proteínas)
Aminoácido Quinua Trigo LecheHistidina * 4,6 1,7 1,7 Isoleucina * 7,0 3,3 4,8 Leucina * 7,3 5,8 7,3 Lisina * 8,4 2,2 5,6 Metionina * 5,5 2,1 2,1 Fenilalanina * 5,3 4,2 3,7 Treonina * 5,7 2,7 3,1 Triptofano * 1,2 1,0 1,0 Valina * 7,6 3,6 4,7 Acido Aspártico 8,6 - - Acido Glutámico 16,2 - - Cisteina 7,0 - - Serina 4,8 - - Tirosina 6,7 - - Argina * 7,4 3,6 2,8 Prolina 3,5 - - Alanina 4,7 3,7 3,3 Glicina 5,2 3,9 2,0
Fuente: IESN-Chile, 2001.

5
La lisina que es uno de los aminoácidos más escasos en los alimentos de
origen vegetal, se muestra en la quinua en una proporción que al menos duplica la
contenida en los otros cereales. Esta ha sido la base para considerar el reemplazo
de las harinas de trigo con quinua a fin de ofrecer un alimento popular con un mejor
contenido de este importante aminoácido (Tapia y col., 1979).
La distribución de las proteínas entre los diversos tejidos que constituyen el
grano no es uniforme, las concentraciones mayores se encuentran en las capas
más externas del endospermo, en la aleurona y en el germen. Tampoco se
distribuyen uniformemente los diferentes tipos de proteínas, las prolaminas y las
glutelinas se encuentran localizadas principalmente en el endospermo; las
albúminas y globulinas están en las cubiertas exteriores y en el germen (Primo,
1979).
Las múltiples características de la quinua hacen de éste un alimento natural
que ayuda al desarrollo y crecimiento del organismo, es fácil de digerir, no contiene
colesterol y por ende no forma grasas en el organismo, forma una dieta completa y
balanceada y se presume que ayuda a prevenir enfermedades como la
osteoporosis, el cáncer de mama, las enfermedades al corazón y otras alteraciones
femeninas ocasionadas por la falta de estrógenos durante la menopausia (Zamudio,
2003).
1.2.3 Las Saponinas
Las pequeñas semillas de quinua están recubiertas de una delgada
membrana o pericarpio que contiene hasta un 4% de saponina, sustancia
sumamente amarga y que produce abundante espuma al agitar la semilla en agua
(Junge y Cerda, 1978).
Las saponinas son sustancias orgánicas de origen mixto, ya que provienen
tanto de glucósidos triterpenoides (de reacción ligeramente ácida), como de
esteroides derivados de perhidro 1,2 ciclopentano fenantreno. En las formas
silvestres y las variedades amargas de quinua, el contenido máximo (aproximado)
de saponina es de un 2.8% (aunque el rango es variable de acuerdo a la especie y
al ecotipo), valor extremadamente alto comparado con las exigencias actuales del
mercado, que fijan como valor límite 0,05% (Fontúrbel, 2003).

6
Además del fuerte sabor amargo, se ha descubierto que las saponinas son
ligeramente tóxicas para los animales y el ser humano, y por ello deben ser
eliminadas antes del consumo del grano. En el organismo, las saponinas ocasionan
dolor estomacal, nauseas, ligera diarrea y problemas en la digestión, puesto que la
fase jabonosa producida al mezclarse con el agua y al ser agitada por los
movimientos peristálticos de las vísceras, hace que se rompan las fuerzas de
tensión superficial de las fases líquidas que intervienen en el proceso de digestión
(Fontúrbel, 2003).
La eliminación de esta sustancia jabonosa se puede realizar en forma
manual (lavando el grano con agua y frotándolo hasta que no salga espuma) o
mecanizado (usando una peladora que pasa el grano por paletas friccionándolo
contra una malla trenzada) (Sepúlveda y col., 2004).
1.3 Empleos de la Quinua
La quinua se consume como el arroz, en grano; sus hojas tiernas se comen
guisadas como las acelgas y espinacas; su tallo y hojas verdes se aprovechan como
ensalada o en sopas. La agroindustria de los países con las mayores producciones
de quinua (entre ellos Bolivia y Perú), transforma este grano preferentemente en
hojuelas y harina, debido a que la fécula es un excelente alimento panificable. Esta
se usa para elaborar panecillos y galletas y para enriquecer harinas de panificación
en la elaboración de: galletas, barritas, tartas, batidos, pasteles y espaguetis, entre
otros. Por otra parte, los granos de segunda clase como los subproductos de la
cosecha pueden ser empleados en la alimentación de monogástricos, aves, cerdos
y rumiantes en condiciones especiales (Ccbolgroup, 2006).
1.4 Mercado Nacional e Internacional
Bolivia y Perú son los mayores exportadores con cerca de un 88% de la
producción a nivel mundial, le sigue Estados Unidos con un 6%. La producción en
Argentina es usualmente para uso doméstico como semilla o harina. Sin embargo y
pese a que su origen es sudamericano, la quinua posee un importante potencial

7
agronómico, debido a que puede adaptarse para producir altos rendimientos bajo
condiciones adversas. Es así como algunos países europeos como España están
estudiando su adaptación al clima Mediterraneo y otros como Dinamarca y Finlandia
están interesados en su cultivo (Vilches y col., 2003).
El consumo en Chile es bajo, comparado con otros alimentos ricos en
carbohidratos como: trigo, arroz y maíz. Esto se debe a que no se conocen las
ventajas nutritivas de la quinua y a que existe una oferta irregular del producto
(Sepúlveda y col., 2004). Sin embargo, existe un mercado creciente del cereal en
vías de un amplio mercado exportable. Es así como en julio del 2002, se realizó la
primera exportación de quinua a Estados Unidos. Ese mismo año, los productores
agrupados en cooperativas ofrecían sus volúmenes productivos del 2003 a
supermercadistas de Canadá, Australia, Inglaterra, España, México y Japón; a la
vez una cooperativa de la VI Región suscribía acuerdos comerciales y de marketing
con la Sociedad Francesa ABCD, con el objetivo de efectuar una serie de acciones
y estrategias destinadas a introducir el uso de quinua chilena en el mercado del
Viejo Continente (Diario Pyme, 2003).
1.5 Los Aislados Proteicos y sus Propiedades
Se considera aislado proteico aquel cuyo contenido de proteínas es mayor al
70%. En él las proteínas constituyentes deben ser exactamente las que se
encontraban en la fuente orgánica inicial, sin haber sufrido procesos de degradación
o hidrólisis no deseables (Curare, 2006).
Los aislados proteicos pueden ser utilizados en la elaboración de diferentes
productos en la industria alimentaria, tales como productos horneados, en la
elaboración de bebidas para deportistas, de embutidos, para la preparación de
alimentos para bebés, por mencionar algunos. Las proteínas se usan como aditivos
en suplementos nutricionales para mejorar el perfil de aminoácidos e incrementar el
contenido de proteínas, pero también para aportar beneficios funcionales como
emulsificación, estabilización e incremento de viscosidad, mejoramiento de la
apariencia, del gusto, la textura y la absorción de agua o aceite (Giese, 1994).
En este sentido, el estudio de las propiedades funcionales es muy importante
para determinar los usos que pueden darse a las proteínas y aislados proteicos en

8
la industria alimentaria. Básicamente, las propiedades funcionales son aquellas
propiedades no nutricionales que son capaces de impartir una característica
tecnológica específica deseable a un producto dado (Cheftel y col., 1989).
Las propiedades funcionales de las proteínas, pueden clasificarse en tres
grupos principales (Cheftel y col. 1989):
Propiedades de hidratación: dependientes de las interacciones proteína-
agua. Incluye propiedades tales como la absorción y retención de agua,
hinchado, adhesión, dispersibilidad, solubilidad y viscosidad.
Propiedades texturales: dependientes de las interacciones proteína-proteína,
y que intervienen en fenómenos tales como la precipitación, la gelificación y
la formación de estructuras diferentes.
Propiedades superficiales: las cuales están relacionadas con la formación de
emulsiones y espumas.
Dentro de las propiedades funcionales que se busca modificar a través de
las proteínas se encuentran: solubilidad, absorción de agua y aceite,
comportamiento reológico, capacidad emulsificante y espumante (Giese, 1994).
Existen muchos estudios respecto a aislados proteicos, los que van desde el
popular aislado de soja, hasta alimentos menos difundidos como la mucuma
(Adebowale y Lawal, 2003). En el caso del aislado de soja, se ha estudiado muy
ampliamente cómo modificar sus propiedades funcionales (Añón y col., 2001).
Situación muy distinta es la del aislado proteico de quinua, en cuyo caso
prácticamente no se han reportado publicaciones. De hecho, el estudio de la quinua
se ha limitado a la composición y caracterización funcional de la semilla y su harina.
En este estudio se obtendrá un aislado proteico a partir de quinua orgánica y
se investigará las propiedades de hidratación de las proteínas del aislado, las que
dependen de las interacciones proteína-agua. Específicamente se estudiará la
solubilidad y la capacidad de retención y absorción de agua. Todo ello con el
objetivo de producir conocimiento e información que permitan incorporar estas
proteínas, de alto valor nutricional, en alimentos destinados al consumo humano.

9
II. HIPÓTESIS
La preparación de aislados proteicos con características funcionales
conocidas constituye un paso preliminar para la formulación de alimentos
tecnológicamente funcionales que los incluya como ingredientes. En el caso de los
aislados proteicos en general, y en específico de la quínoa, el conocimiento de
dichas características es posible a través del discernimiento de la relación
estructura-funcionalidad de las fracciones proteicas, la que se podría determinar por
medio del estudio de los cambios estructurales de la proteína, frente a distintas
condiciones de procesamiento físicas y/o químicas.

10
III. OBJETIVOS
3.1 Objetivo General
Obtener un aislado proteico a partir de quinua orgánica y estudiar sus
proteínas desde el punto de vista estructural y funcional, para producir conocimiento
que permita incorporar estas proteínas, de alto valor nutricional, en alimentos
destinados al consumo humano.
3.2 Objetivos Específicos
Obtener un aislado proteico de quinua orgánica.
Determinar el contenido de proteínas de la harina de quinua, a partir de la
cual se obtendrá el aislado.
Realizar un análisis proximal y determinar el perfil de aminoácidos al aislado
proteico.
Determinar el contenido de Fitoestrógenos en el aislado proteico, mediante
el uso de cromatrografía líquida de alta resolución (HPLC).
Caracterizar los perfiles polipeptídicos de las proteínas del aislado de quinua,
mediante electroforesis (PAGE).
Estudiar los cambios conformacionales de los aislados proteicos a distintos
pHs, mediante el uso de técnicas espectrofotométricas (UV y fluorescencia).
Determinar la estabilidad térmica de las proteínas del aislado proteico,
mediante la técnica de calorimetría diferencial de barrido (DSC).
Determinar las propiedades funcionales que exhiben las proteínas
integrantes del aislado proteico de quinua, mediante el estudio de tres
propiedades de hidratación: solubilidad, capacidad de retención de agua y
capacidad de absorción de agua, a distintos pHs.
Analizar las relaciones existentes entre las características estructurales que
exhiben las proteínas y sus propiedades funcionales.

11
IV. MATERIALES Y MÉTODOS
4.1 Materiales
4.1.1 Materia Prima:
Quinua orgánica pelada en seco, proveniente de la localidad de La Plaza,
comuna de Pichilemu, ubicada en la VI Región de Chile. La semilla fue
proporcionada por el agricultor Críspulo Leiva.
4.1.2 Reactivos Químicos
Acetato de sodio (CH3COONa) Panreac, Barcelona, España
Acetonitrilo (CH3CN) Panreac. Barcelona, España
Ácido acético glacial (CH3COOH) p.a. Merck. Darmstadt, Alemania
Ácido bórico (H3BO3) Panreac, Barcelona, España
Ácido clorhídrico (HCl) p.a. Merck. Darmstadt, Alemania
Ácido orto-fosfórico (H3PO4) p.a. Merck. Darmstadt, Alemania
Ácido perclórico (HClO4) p.a. Merck. Darmstadt, Alemania
Ácido sulfúrico (H2SO4) p.a. Merck. Darmstadt, Alemania
Acrilamida (C3H5NO) p.a. Merck. Darmstadt, Alemania
Alcohol etílico (Etanol) (C2H60) p.a. Winkler. México
Alcohol etílico absoluto (Etanol) (C2H60) técnico, Comercial Tecnológica
Ltda. Santiago, Chile
Antrona (C14H10O) p.a. Merck. Darmstadt, Alemania
Azul brillante de coomasie G-250 (C47H50N3NaO7S2) Baker. USA
Azul de bromofenol (C19H10Br4O5S) Sigma. USA
Bis-acrilamida (C7H10N2O2) Sigma. USA
β -mercapto etanol (C2H6OS) p.a. Merck. Darmstadt, Alemania
Cloruro de sodio (NaCl) p.a. Merck. Darmstadt, Alemania
Coomassie blue R (C45H44N3O7S2Na) Sigma. USA
Dietil etoximetilenmalonato. Fluka. Buchs, Suiza
Dodecil sulfato de sodio (SDS) (C12H25NaO4S) p.a. Merck. Darmstadt,
Alemania
Estándar de aminoácidos DL-2-Aminobutyric Acid for synthesis, Merck
Schuchardt, Alemania

12
Estándar de proteínas para electroforesis Bio-Rad Precision Plus ProteinTM
Catalog 161-0373 N. (con pesos moleculares de: 250, 150, 100, 75, 50, 37,
25, 20, 15 y 10).
Estándar de Seroalbúmina de bovino (BSA). Sigma. USA
Estándar de fitoestróngenos Daizenine Genist. Sigma. USA
Fosfato de potasio dihidrogeno (KH2PO4) p.a. Merck. Darmstadt, Alemania
Fosfato de sodio monobásico (NaH2PO4*H2O) p.a. Merck. Darmstadt,
Alemania
Glicerol (C3H8O3) Sigma. USA
Glicina (C2H5NO2) Sigma. USA
Glucosa monohidratada (C6H12O6*H2O) p.a. Merck. Darmstadt, Alemania
Hidróxido de sodio (NaOH) técnico. W&Z
Hidróxido de sodio (NaOH) p.a. Merck. Darmstadt, Alemania
Indicador fenolftaleina (C20H14O4) p.a. Merck. Darmstadt, Alemania
Indicador rojo de metilo (C15H15N3O2) p.a. Merck. Darmstadt, Alemania
Persulfato de amonio (PSA) ((NH4)2S2O8) Sigma. USA
Sodio fosfato bibásico anhidro (Na2HPO4) p.a. Merck. Darmstadt, Alemania
Sulfato de cobre pentahidratado (CuSO4*5H2O) p.a. Merck. Darmstadt,
Alemania
Sulfato de potasio (K2SO4) p.a. Purom
TEMED (N,N,N`,N`-Tetramethiletilendiamina) Sigma. USA
Tris (C4H11NO3) p.a. J.T.Baker. USA
4.1.3 Equipos e Instrumentos
Agitador eléctrico vortex Thermolyne, tipo 37600 Mixer, modelo Nº M37610-
26, serie Nº 871570819103
Agitador magnético Magnetic Stirrer HI190M Hanna Instruments Nº S229370
Agitador Heavy Duty KitchenAid Inc. St Joseph, model K5SS, Michigan USA
Balanza analítica Precisa 125 A Swiss Quality
Balanza granataria AND, modelo EK 120-A
Balanza granataria Precisa 1620 D
Baño de agua Haake Tipo FK Nº 751117, fabricado en Alemania
Batería de tamices

13
Calorímetro diferencial de barrido (DSC) Mettler Toledo 822e
Cámara de refrigeración FríoLux
Campana de extracción
Centrifuga Heraeus Sepatech Suprafuge 22, fabricada en Alemania
Conjunto para electroforesis Bio-Rad con kit para preparación de geles, cuba
de corrida y fuente de poder
Desecador
Destilador Büchi 323 tipo B-323 Nº1258804, fabricado en Suiza
Digestor Büchi 426 tipo B-426RC Nº 1270878, fabricado en Suiza
Espectrofotómetro de fluorescencia Perkin Elmer, modelo LS50B, serie Nº
32938 L225-0105
Espectrofotómetro Perkin Elmer Lambda 11 Nº BC11 230V,serie NR30031
Espectrofotómetro UNICAM UV/Vis, tipo UV3-200, Nº UV 3022809, fabricado
en Inglaterra
Estufa Heraeus Instruments D-63450 Hanau, tipo UT 6200, fabricada en
Alemania
Estufa Heraeus, tipo KB 600
Estufa Heraeus W.C. Heraeus GMBH Hanau, tipo TU 60/60, Nº 2760-02,
fabricada en Alemania
Estufa WTB Binder Tipo 1924090000200 Nº 940107 73532 Tuttlingen,
Alemania
Freezer medical Sanyo, modelo MDF-U332, Nº 606033941, fabricado en
Japón
Mechero
Microcentrifuga HermLe Z160M Hersteller Spintron, fabricada en Alemania
Mini secador Spray Dryer B-191, Büchi, fabricado en Suiza
Molino de impacto Retsh Muhle GmbH West-Germany, type SR-2 Nº73454
Mufla Wild Barfiel tipo M254 Nº c2441557, fabricada en Inglaterra
pHmetro microprocesador pH 537 WTW, fabricado en Alemania
Placa calefactora Gerhardt Bonn tipo H52 Nº 443792
Refrigerador Mademsa Evolution 3500 G no frost system
Sistema HPLC Merck Hitachi bomba ternaria L6200 con inyector Rheodyne
7725; loop de 20 µl, Detector espectrofotométrico UV-Visible modelo 484 de

14
longitud de onda variable, Integrador D-2500; Columna Nova Pack RP18 (3,9
x 300 mm, con un tamaño de partícula de 4 µm), acoplado a un computador
con el software Clarity versión preliminar 2.4.1.43, con licencia Waters.
Termómetro
4.1.4 Insumos y Utensilios
Agua destilada y bidestilada
Bandejas de acero galvanizado de 30 mallas de 44 cm x 24 cm
Bolsas de papel Kraft
Cápsula de aluminio
Cápsula de porcelana
Cubetas para electroforesis
Cubetas de cuarzo
Cubetas de plástico
Espátulas y microespátulas
Filtro Millipore membrana 6V (Durapore)
Frascos de vidrio
Fuentes plásticas
Gradillas
Guantes
Hielo
Jeringas desechables Dynatech de 5mL
Material de vidrio (buretas, pipetas de aforo, pipetas gravimétricas, matraces
erlenmeyer, matraces aforados, matraz kitasato, balones, vaso con doble
camisa, dedales, varillas de agitación, tubos de ensayo con tapa con teflón,
varillas de reflujo, etc.)
Micropipetas
Mortero
Nitrógeno
Papel absorbente
Papel Whatman Nº1
Parafilm
Pinzas

15
Plumavit
Potes de plástico
Rejilla de asbesto
Soporte para electroforesis
Trípode
Tubos de digestión
Tubos Ependorf
Utensilios de cocina
Vidrios para polimerización y peineta (plástico dentado)
4.2 Métodos
4.2.1 Obtención de la Harina de Quinua Desgrasada
Se seleccionaron los granos mediante tamizado, retirándose toda materia
extraña. Luego se lavaron con abundante agua corriente y mediante frotación, con
el objetivo de eliminar las saponinas. Dicho proceso se llevó a cabo hasta que no se
produjo más espuma, lo cual indica que las saponinas fueron efectivamente
retiradas.
Posteriormente, se llevó los granos a una estufa, a 50º C, hasta alcanzar la
humedad de acondicionamiento del grano para la elaboración de la harina (15±1%).
Para comprobar que se obtuvo la humedad deseada, se extrajo una muestra de 5 g,
a la que se le determinó humedad por el método rápido, el que consiste en someter
la muestra a 156º C, durante 15 min (Pearson, 1970). Luego, se molió los granos
usándose un molino de martillo-cuchillo (Retsch tipo SR-2), proporcionado por el
laboratorio de Operaciones Unitarias de la facultad.
Para desgrasar la harina, ésta fue suspendida en hexano en una relación
1:10. La suspensión se mantuvo durante 24 horas, a aproximadamente 4º C, bajo
agitación continua. Después, se filtró la harina al vacío con papel Whatman Nº 1 y
se secó a temperatura ambiente hasta la completa eliminación del hexano.
Finalmente se almacenó en bolsas de papel Kraft, dentro de un desecador.

16
4.2.2 Obtención del Aislado Proteico de Quinua
Se suspendió la harina desgrasada en agua al 10 % p/v. Luego, para
solubilizar las proteínas, se ajustó el pH de la solución a 9,0, con NaOH 2N,
manteniéndose a temperatura ambiente y agitación, por 30 minutos. Posteriormente,
se centrifugó durante 60 min. a 3.400 rpm a 15 ºC (ver Figura 3).
Figura 3: Solubilización de las proteínas a pH 9 (izquierda) y posterior
centrifugación (derecha) Para precipitar las proteínas se ajustó el pH del sobrenadante, llevándolo a
pH 5 (cercano al punto isoeléctrico), con HCl 2N. Luego se centrifugó a 3.400 rpm
por 60 min, a 4º C. Inmediatamente después, se resuspendió el precipitado para
neutralizarlo, usándose para ello NaOH 0,1 N (Martínez, 1997).
El secado del aislado se llevó a cabo mediante secado spray (mini secador
Büchi, B-191), bajo las siguientes condiciones:
Temperatura del aire: 120º C
Concentración de la suspensión del aislado: 20%
Flujo del aire: 600
Aspiración: 90%
Alimentación: 15%
Figura 4: Aislado proteico de quinua obtenido a pH 9

17
Figura 5: Diagrama de la Obtención del Aislado Proteico de Quinua a pH 9
Suspensión de la harina de quinua desgrasada en agua al 10 % p/v
Ajuste a pH 9 con NaOH 2N
Agitación por 30 min a temperatura ambiente
Centrifugación a 3.400 rpm por 1 h a 15º C
Ajuste del pH del sobrenadante a 5,0 con HCl 2N
Centrifugación a 3.400 rpm por 1h a 4º C
Resuspensión del precipitado en agua al 20%
Neutralización con NaOH 0,1N
Secado spray a 120º C

18
4.2.3 Caracterización Química
4.2.3.1 Composición Química:
Se realizó análisis proximal, lo que comprendió la determinación del
contenido de:
a. Proteínas. Se determinó proteínas totales mediante el método de Kjeldahl
(Official Method AOAC 920.53, 1995). El método consiste básicamente en la
digestión ácida total de las proteínas y conversión cuantitativa del nitrógeno
a NH3, el que se valora por retrotitulación.
El factor de conversión utilizado tanto para la harina de quinua como para el
aislado proteico es 5,77 (Schmidt-Hebbel, 1981).
b. Carbohidratos. A través del método colorimétrico de reacción de Antrona, el
cual implica digerir 1,0 g de muestra con ácido perclórico. Posteriormente se
procede a una serie de diluciones, para finalmente tomar 1 mL de la solución
la que se hace reaccionar con 5 mL de Antrona. Los almidones hidrolizados,
junto con los azúcares solubles, se determinaron colorimétricamente
mediante espectrofotómetría a una longitud de onda de 630 nm, utilizándose
patrones de glucosa de 0,05 y 0,1 mg/mL (Osborne y Voogt, 1986).
c. Humedad. Se empleó el método gravimétrico propuesto por la AOAC
(Official Method 935.29, 1995), lo que implicó colocar la muestra en estufa a
105º C, hasta peso constante.
d. Cenizas. Se realizó de acuerdo al método oficial de la AOAC (Official
Method 923.03, 1995), el cual consistió en calcinar la muestra a 550º C,
determinándose gravimétricamente el contenido de cenizas.
4.2.3.2 Perfil de Aminoácidos:
Se hidrolizó las muestras (aproximadamente 2 mg) con ácido clorhídrico 6N,
durante 24 horas, a 110ºC. Posteriormente, el ácido se evaporó a sequedad y los
aminoácidos se solubilizaron con tampón de borato de sodio 1M (pH 9,0), aforando
a 25 mL. Seguidamente, se derivatizó los aminoácidos, para lo cual se colocó 5 mL
de la solución anterior en un tubo de ensayo con tapa de teflón más 4 µl de
dietiletoximetilenmalonato. La derivatización se llevó a cabo en un baño de agua a
50ºC, durante 50 min, bajo agitación constante.

19
Finalmente, 20 µl de la muestra derivatizada se inyectaron en un
cromatógrafo HPLC con detector UV-Visible Merck Hitachi, modelo 484, longitud de
onda 280 nm, acoplado a un computador con el software Clarity versión preliminar
2.4.1.43, con licencia Waters. La resolución de los derivados de aminoácidos se
logró usando una columna Nova Pack RP18 (3,9 x 300 mm, con un tamaño de
partícula de 4 µm) y un sistema de gradiente binario a temperatura ambiente,
empleando como fase móvil tampón de acetato de sodio 25mM (pH 6,0) y
acetonitrilo, de acuerdo al detalle descrito en el Anexo 4 (Alaiz y col., 1992).
4.2.3.3 Determinación de Fitoestrógenos:
El método utilizado consistió en hidrolizar las muestras (1 g) con 24 mL de
ácido clorhídrico 1N en baño de agua, a 98ºC, durante 2 horas. Luego se extrajo los
fitoestrógenos con 100 mL de acetonitrilo. Enseguida, se diluyó 1 mL del
sobrenadante obtenido con 1 mL de agua destilada, dilución que luego de agitada
se filtró, utilizándose para ello una jeringa con porta filtro Millipore.
La determinación de los Fitoestrógenos se realizó mediante cromatrografía
líquida de alta resolución (HPLC Merk Hitachi), utilizando el mismo equipamiento
señalado en la sección 4.2.3.2 y un sistema de gradiente binario de metanol y agua
como fase móvil, de acuerdo al detalle descrito en el Anexo 4 (Wang y col., 1990).
4.2.4 Caracterización Estructural de las Fracciones Proteicas
4.2.4.1 Perfiles Polipeptídicos:
Mediante Electroforesis se analizó y caracterizó los polipéptidos que integran
el aislado proteico de quinua. Ello fue realizado de acuerdo al método descrito por
LaemmLi U. K. (1970), el cual detalla la técnica PAGE, la que se realiza en función
del estado de las proteínas (nativo o desnaturalizado).
En la electroforesis nativa (PAGE-nativa), se somete a las proteínas a
migración en un campo eléctrico. En esta situación las proteínas migran en función
de su carga, su tamaño y forma, mientras que en la electroforesis desnaturalizante
(PAGE-SDS), la más común, las proteínas son sometidas a migración en un campo
eléctrico en presencia de un detergente aniónico, lo que asegura la completa

20
desnaturalización (pérdida de la estructura tridimensional). En esta situación la
migración es proporcional al tamaño molecular de la molécula, pero no a su carga.
El aislado proteico de quinua fue sometido a ambos tipos de electroforesis,
además de la electroforesis desnaturante con β-mercaptoetanol (2-Me).
Para la realización de estas pruebas se elaboró geles de 1 mL de espesor.
En el caso de la electroforesis desnaturante, sin y con β-mercapto etanol, se usó gel
concentrador al 5% y gel separador al 12%. Se cargó 10 µl de muestra y 7 µl de
estándar.
Las masas moleculares de los polipéptidos fueron calculadas usando un
estándar de proteínas con los siguientes pesos moleculares: 250, 150, 100, 75, 50,
37, 25, 20, 15 y 10 kDa.
En el caso de la electroforesis nativa se usó gel separador al 5%, y no se
carga estándar (ver el detalle de estos métodos en el anexo 1).
4.2.4.2 Fluorescencia:
Se preparó dispersiones al 1% del aislado proteico en buffer a los siguientes
pHs: 3, 5, 7, 9 y 11, en tubos ependorf (ver detalle de los buffer en anexo 3).
Posteriormente se sometió los tubos a agitación intensa en vortex, a intervalos de
15 minutos, durante 1 hora, a temperatura ambiente. A continuación, se centrifugó
los tubos a 10.000 rpm, durante 30 minutos, a 15° C. Luego, se midió la absorbancia
del sobrenadante a 260 y 280 nm, datos que se utilizaron en la siguiente fórmula,
para conocer los mg de proteína por mL de solución:
Proteína = 1,55 * A280 – 0,76 * A260 [mg/mL]
El propósito del paso anterior fue estandarizar la concentración de las
dispersiones de los distintos pHs, diluyéndolas, para que todas tuvieran una
concentración de 0,02 mg/mL.
Una vez estandarizadas las muestras, se midieron en un espectrofotómetro
de fluorescencia (Perkin Elmer, modelo LS50B) con una excitación de 270-290 nm,
para obtener el espectro de emisión en el rango de 300 a 500 nm, con una
velocidad de barrido de 300 nm/min (Permyakov, 1993; Mathews y col., 2002;
Abugoch, 2006).

21
4.2.4.2 Espectroscopia UV:
Se preparó diluciones del aislado al 1% en buffers de pHs: 3, 5, 7, 9 y 11 (ver
detalle de los buffers en anexo 3). Posteriormente se agitó las muestras
vigorosamente en vortex cada 15 min, hasta completar 1 hora. Concluida la etapa
anterior, se centrifugó las muestras por 30min a 10.000 rpm. Para realizar la
medición se diluyó 5 veces el sobrenadante obtenido de la centrifugación (Mathews
y col., 2002; Abugoch, 2006).
Finalmente, se midió la absorción de las muestras en el espectrofotómetro
UNICAM UV/Vis, efectuándose un barrido de longitudes de onda, que cubrió desde
los 250 hasta los 450 nm.
4.2.4.4 Calorimetría Diferencial de Barrido:
Se efectuó una caracterización térmica de las proteínas, para lo cual se
preparó dispersiones al 20% con buffer de pH 9 y agua. Se colocó entre15 y 20 mg
de dicha mezcla en una cápsula, previamente pesada y tarada la cual se selló y
llevó al equipo de calorimetría diferencial de barrido (Mettler Toledo 822e). Se
trabajó desde los 10 a los 120º C, con un régimen de calentamiento de 10º C por
min. Como referencia se utilizó aislado proteico de quinua con la proteína
previamente desnaturalizada.
4.2.5 Determinación de las Propiedades Funcionales
4.2.5.1 Solubilidad:
Se preparó dispersiones proteicas al 1% p/v en buffers que cubrieron desde
el pH 3 al pH 11 (detalle de las fórmulas de los buffers en el anexo 3). Las muestras
se sometieron cada 15 minutos a agitación intensa y breve en vortex a temperatura
ambiente, durante 1 hora. En seguida, se centrifugaron los tubos a 10.000 rpm,
durante 30 minutos, a 15° C (Scilingo, 2000). La proteína que quedó en el
sobrenadante se cuantificó a través del método descrito por Bradford, M. M. (1976)
(ver detalles de este método en el anexo 2).

22
4.2.5.1 Capacidad de Retención de Agua (WHC):
En tubos ependorf, previamente pesados, se elaboró dispersiones proteicas
al 1% p/v en agua y en buffers de pH: 3, 5, 6, 7 y 9. A continuación, los tubos se
sometieron a agitación intensa en vortex, a intervalos de 15 minutos, durante 1 hora,
a temperatura ambiente. Posteriormente, se centrifugaron los tubos a 10.000 rpm,
durante 30 minutos, a 15°C. Luego se determinó la masa de precipitado obtenido,
para lo cual antes se dejó invertidos los tubos durante 10 min sobre papel
absorbente. Paralelamente, se cuantificó la cantidad de proteína solubilizada en el
sobrenadante después de la centrifugación, lo que se efectuó utilizándose el método
de Bradford M. M. A (1976) (ver detalles de este método en el anexo 2).
La capacidad de retención de agua, se obtuvo a través de la siguiente
fórmula:
WHC = m2 – (m1 – m3) [mL agua / g muestra] m1 * d Donde:
m1: masa de muestra pesada en gramos
m2: masa del precipitado obtenido en gramos
m3: masa de la proteína soluble en gramos
d: densidad del agua a 25° C
4.2.5.3 Capacidad de Absorción de Agua (WIC):
La WIC es la cantidad máxima de agua que un gramo de aislado es capaz
de absorber espontáneamente, a una temperatura definida. Para determinarla se
utilizó un equipo similar al diseñado por Baumann (1967), el que consiste en un
embudo buchner conectado, a través de una manguera flexible, a una pipeta de 1
mL graduada (ver Figura 6).
Una vez instalado el equipo, se llenó con agua, evitando la incorporación de
burbujas. Luego se colocó el papel filtro dentro del embudo, dejándolo que
embebiera agua durante aproximadamente 5 min. Posteriormente, se eliminó el
exceso de agua sobre el papel filtro, con el objetivo de enrasar a cero la columna de
agua en la pipeta. Se verificó que la nivelación del equipo no variara durante un
lapso de 10 min, tras lo cual se esparció rápidamente 60 mg del aislado sobre el
papel filtro húmedo, para formar una monocapa uniforme. Al mismo tiempo, se

23
disparó un cronómetro para realizar mediciones a través del tiempo, registrándose el
retroceso de la columna de agua en la pipeta, hasta que no hubo más variación
(Añón y col., 2000).
Figura 6: Esquema del equipo de absorción de agua
Los datos obtenidos se graficaron colocando el tiempo en minutos en el eje
de las abscisas y los mL de agua absorbidos por gramo de aislado en el eje de las
ordenadas. A partir de él se obtuvo la WIC, el tiempo de equilibrio (te) y la velocidad
inicial de absorción de agua (vi) (Abugoch, 2006).
4.2.6 Análisis Estadístico
Se evaluó estadísticamente los resultados obtenidos usando la media
aritmética y la desviación estándar. Asimismo, se efectuó análisis de varianza, con
un nivel de confianza del 95%, para determinar si existen diferencias significativas
entre los distintos pHs estudiados. Este último análisis se llevó a cabo empleándose
el programa StatGraphics Plus 4.0.

24
V. RESULTADOS Y DISCUSIÓN
5.1 Caracterización Química del Aislado Proteico de Quinua
5.1.1 Composición Proximal
A partir de harina de quinua desgrasada con 12,7 ± 0,4 % de proteína, se
obtuvo un aislado proteico con un contenido de proteína en base húmeda del 72,5 ±
0,14 %. En la tabla 3 se observa la composición proximal del aislado.
Tabla 3: Composición proximal del aislado proteico de quinua A9
Componente % Proteínas* 77,2 ± 0,15Humedad 6,1 Hidratos de carbono* 18,8 ± 0,52Cenizas* 3,0
*En base seca
El aislado fue tamizado con el objetivo de determinar el tamaño de partícula,
estableciéndose que un 2,73% del aislado se encontró entre 0,25 y 0,5 mm, un
76,6% entre 0,125 y 0,250 mm y un 20,67% presentó un tamaño de partícula inferior
a 0,125mm.
La actividad de agua (aw), determinada en un equipo Novasina, fue de 0,326,
medida a 25º C.
El aislado de quinua fue obtenido solubilizando las proteínas a pH 9, por lo
que en adelante se denominará A9. De esta manera se diferenciará de otro aislado,
obtenido en condiciones similares, pero a pH 11 (Silva, 2006) el que se designará
como A11. El rendimiento de A9 fue similar al de A11, esto es próximo al 5 %,
aunque el contenido de proteínas para A11 fue mayor, siendo de un 83,5 %, en
base seca.
El contenido de proteínas también fue inferior que el obtenido en aislados de
amaranto (otro pseudocereal), en cuyo caso se han reportado contenidos proteicos
del 90% (Martínez y Añón, 1996) y del 84,4%, en condiciones de extracción muy
similares a las de la quinua (Abugoch, 2006).
25
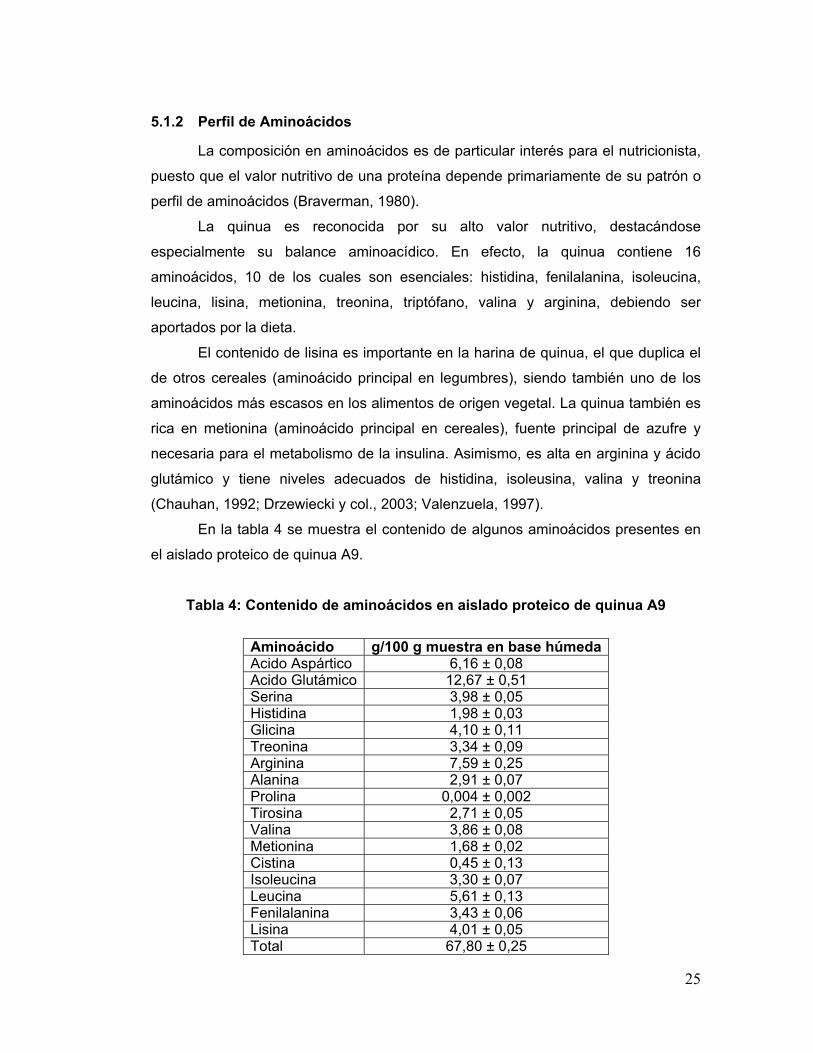
5.1.2 Perfil de Aminoácidos
La composición en aminoácidos es de particular interés para el nutricionista,
puesto que el valor nutritivo de una proteína depende primariamente de su patrón o
perfil de aminoácidos (Braverman, 1980).
La quinua es reconocida por su alto valor nutritivo, destacándose
especialmente su balance aminoacídico. En efecto, la quinua contiene 16
aminoácidos, 10 de los cuales son esenciales: histidina, fenilalanina, isoleucina,
leucina, lisina, metionina, treonina, triptófano, valina y arginina, debiendo ser
aportados por la dieta.
El contenido de lisina es importante en la harina de quinua, el que duplica el
de otros cereales (aminoácido principal en legumbres), siendo también uno de los
aminoácidos más escasos en los alimentos de origen vegetal. La quinua también es
rica en metionina (aminoácido principal en cereales), fuente principal de azufre y
necesaria para el metabolismo de la insulina. Asimismo, es alta en arginina y ácido
glutámico y tiene niveles adecuados de histidina, isoleusina, valina y treonina
(Chauhan, 1992; Drzewiecki y col., 2003; Valenzuela, 1997).
En la tabla 4 se muestra el contenido de algunos aminoácidos presentes en
el aislado proteico de quinua A9.
Tabla 4: Contenido de aminoácidos en aislado proteico de quinua A9
Aminoácido g/100 g muestra en base húmeda Acido Aspártico 6,16 ± 0,08 Acido Glutámico 12,67 ± 0,51 Serina 3,98 ± 0,05 Histidina 1,98 ± 0,03 Glicina 4,10 ± 0,11 Treonina 3,34 ± 0,09 Arginina 7,59 ± 0,25 Alanina 2,91 ± 0,07 Prolina 0,004 ± 0,002 Tirosina 2,71 ± 0,05 Valina 3,86 ± 0,08 Metionina 1,68 ± 0,02 Cistina 0,45 ± 0,13 Isoleucina 3,30 ± 0,07 Leucina 5,61 ± 0,13 Fenilalanina 3,43 ± 0,06 Lisina 4,01 ± 0,05 Total 67,80 ± 0,25
26
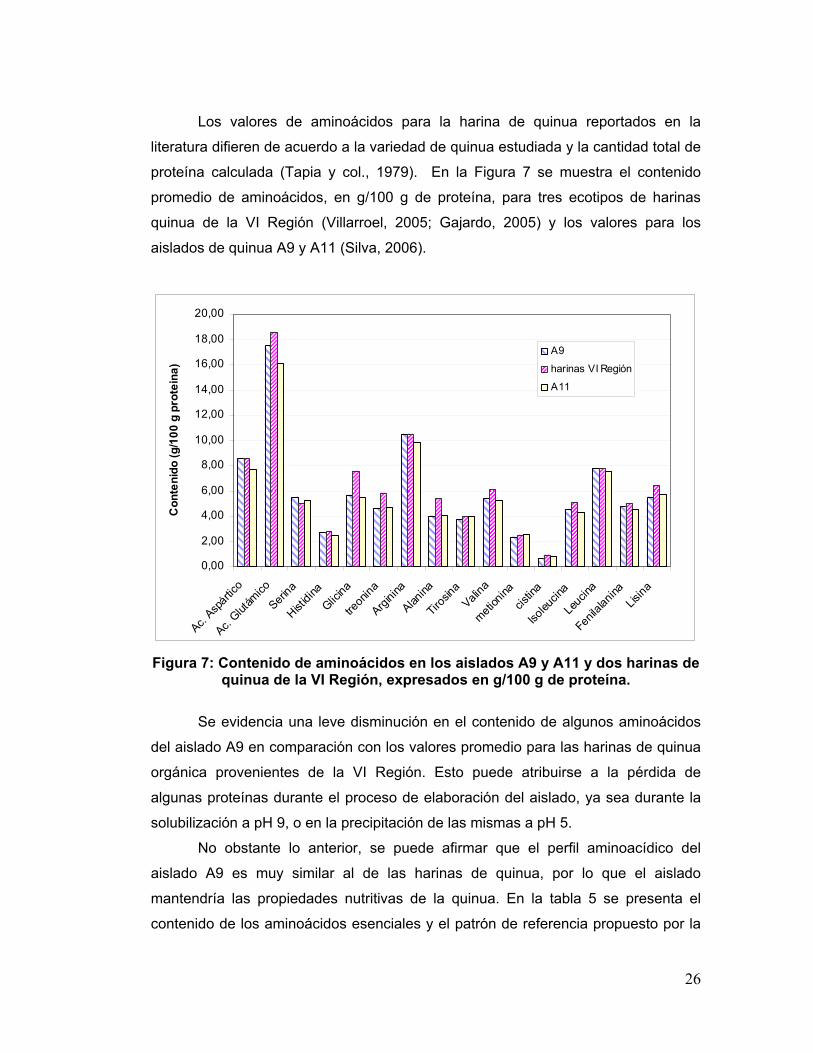
Los valores de aminoácidos para la harina de quinua reportados en la
literatura difieren de acuerdo a la variedad de quinua estudiada y la cantidad total de
proteína calculada (Tapia y col., 1979). En la Figura 7 se muestra el contenido
promedio de aminoácidos, en g/100 g de proteína, para tres ecotipos de harinas
quinua de la VI Región (Villarroel, 2005; Gajardo, 2005) y los valores para los
aislados de quinua A9 y A11 (Silva, 2006).
0,00
2,00
4,00
6,00
8,00
10,00
12,00
14,00
16,00
18,00
20,00
Ac. Asp
ártico
Ac. Glut
ámico
Serina
Histidi
naGlic
ina
treon
ina
Arginin
a
Alanina
Tiros
inaVali
na
metion
inacis
tina
Isoleu
cina
Leuc
ina
Fenil
alanin
aLis
ina
Con
teni
do (g
/100
g p
rote
ina)
A9
harinas VI Región
A11
Figura 7: Contenido de aminoácidos en los aislados A9 y A11 y dos harinas de
quinua de la VI Región, expresados en g/100 g de proteína.
Se evidencia una leve disminución en el contenido de algunos aminoácidos
del aislado A9 en comparación con los valores promedio para las harinas de quinua
orgánica provenientes de la VI Región. Esto puede atribuirse a la pérdida de
algunas proteínas durante el proceso de elaboración del aislado, ya sea durante la
solubilización a pH 9, o en la precipitación de las mismas a pH 5.
No obstante lo anterior, se puede afirmar que el perfil aminoacídico del
aislado A9 es muy similar al de las harinas de quinua, por lo que el aislado
mantendría las propiedades nutritivas de la quinua. En la tabla 5 se presenta el
contenido de los aminoácidos esenciales y el patrón de referencia propuesto por la

27
FAO/OMS, los que corresponden a los requerimientos para niños preescolares entre
2 y 5 años (FAO, 1970).
Tabla 5: Contenido de aminoácidos esenciales en el aislado de quinua A9 y
patrón de referencia FAO (mg/g proteína)
Aminoácidos Aislado de quinua A9 Patrón FAO/OMS Histidina 19,8 19 Isoleucina 33 28 Leucina 56,1 66 Lisina 40,1 58 Metionina + Cisteina 21,3 25 Fenilalanina + Tirosina 61,4 63 Treonina 33,4 34 Triptófano * 11 Valina 38,6 35
* No determinado
Se observa en la tabla 5 que algunos valores de contenidos de aminoácidos
para el aislado de quinua se encuentran por debajo del patrón de referencia,
especialmente la leucina y la lisina, mientras que para los otros aminoácidos los
valores son muy cercanos o los superan. Por lo tanto, si bien la proteína no es
completa para el grupo de niños de 2 y 5 años, grupo que posee la mayor demanda
de requerimientos de aminoácidos, sí lo sería para los adultos los cuales poseen
requerimientos bastante inferiores a los de los preescolares.
5.1.3 Contenido de Fitoestrógenos
La determinación de fitoestrógenos en el aislado proteico de quinua mostró
su ausencia. Sin embargo tampoco se hallaron estas sustancias en la harina a partir
de la cual se elaboró el aislado. Cabe señalar que en los tres ecotipos de quinua
utilizados anteriormente, también procedentes de la VI Región, se encontró
fitoestrogenos del grupo isoflavonas, específicamente Genisteina y Daidzeina
(Araneda, 2004), por lo tanto, en estudios posteriores deberá determinarse si estos
compuestos quedan en el aislado o se pierden en el proceso de extracción de la
proteínas.
Son conocidas las propiedades benéficas para la salud de los fitoestrógenos,
los que se han asociado principalmente a una menor incidencia de ciertas

28
patologías, como la enfermedad cardiovascular y algunos cánceres
hormonodependientes, sin embargo también se ha planteado que su consumo
puede producir efectos adversos como interferencia en los procesos reproductivos
(Wang y col., 1990), por lo que la posibilidad de obtener aislados proteicos de
quinua sin fitoestrógenos no es del todo desfavorable.
5.2 Caracterización Estructural de las Fracciones Proteicas
5.2.1 Perfiles Polipeptídicos del Aislado Proteico de Quinua A9
5.2.1.1 Electroforesis Nativa (PAGE-Nativa)
En la electroforesis nativa las proteínas migran en función de su carga, de su
tamaño y de su forma. Además se mantienen en ciertos casos las interacciones
entre subunidades y entre proteínas, separándose los complejos (UB, 2006). En la
Figura 8 se presenta el gel del aislado proteico de quinua (A9) obtenido en
condiciones nativas. En él se visualizan principalmente 3 bandas, dos de ellas (a y
b), con menor nivel de migración, debieran tratarse de proteínas de mayor tamaño o
menor carga negativa, mientras que la tercera banda (c) correspondería a moléculas
más pequeñas o con mayor carga negativa, lo cual les permitió mayor movilidad en
el gel.
Figura 8: PAGE-Nativa de los aislados proteicos de quinua A9 y A11

29
Por otra parte, debido a que la primera banda (a) es de mayor intensidad que
las restantes, se prevé una mayor concentración de este tipo de proteínas. De
acuerdo a lo señalado por Martínez y Añón (1996), para aislados de amaranto
extraídos en condiciones similares, dichas proteínas con baja movilidad (tanto a,
como b), podrían corresponder al tipo albúminas y globulinas, las cuales están
presentes en la quinua. En condiciones nativas las proteínas de quinua están
compuestas principalmente por dos tipos de polipéptidos, albúminas del tipo 2S y
globulinas 11S, ambas estabilizadas por puentes disulfuro, con masas moleculares
de 3 - 4 kDa y 7 - 9 kDa (Brinegar y Goundan, 1993; Brinegar y col., 1996).
Estos resultados son prácticamente los mismos que los obtenidos por el
aislado proteico A11 (Silva, 2006), esto es: el mismo número de bandas, con un
nivel de movilidad muy similar (ver Figura 8).
Por otra parte, en pruebas de electroforesis nativa realizada a muestras de
harina de quinua, también de la VI Región (Gajardo, 2005), se obtuvo sólo 1 banda
y con un alto grado de movilidad (mayor a “c”, en la Figura 8). En este sentido,
podría conjeturarse que en el proceso de obtención del aislado proteico de quinua
hubo agregación proteica, dando como resultado proteínas de mayor tamaño y por
ende con menor movilidad.
5.2.1.2 Electroforesis Desnaturante (PAGE-SDS)
En la electroforesis en gel con detergente SDS, la cadena polipeptídica se
despliega y se rodea de moléculas de SDS, cargándose negativamente. Dicha
carga es proporcional a su longitud, y por lo tanto a su peso molecular, de modo que
las moléculas migran con movilidades relativas de acuerdo a su peso molecular
(Mathews y col., 2002).
En la Figura 9, se muestra el gel realizado para el aislado proteico de quinua
(A9) en condiciones desnaturantes. En el primer carril, de izquierda a derecha, se
observa el estándar de peso molecular; en el siguiente (a) el perfil proteico sin la
presencia de β-mercaptoetanol (2-Me); mientras que en la última línea (b) se
visualiza el perfil polipeptídico en presencia del agente reductor de enlaces disulfuro.
Este procedimiento se efectúa para distinguir entre las subunidades que se
mantienen juntas mediante puentes –S-S- y las que se mantienen unidas sólo
mediante fuerzas no covalentes.

30
Figura 9: PAGE-SDS del aislado A9 (a) sin 2-Me y (b) con 2- Me
En lo que respecta a los perfiles sin 2-Me, son 11 las bandas más intensas, y
por lo tanto las proteínas presentes en el aislado corresponden en su mayoría a
dichos pesos moleculares. Las masas moleculares de dichas bandas son: 111,9;
74,9; 52,5; 32,4; 26,7; 19,8; 19,0; 14,8; 14,3; una banda con peso molecular mayor a
250 kDa y una aglomeración entre 13,1 y 10,9 kDa.
Las globulinas tipo 11S se caracterizan por tener dos grupos heterogéneos
de polipéptidos a 30 - 40 kDa (subunidades ácidas) y a 20 - 25 kDa (subunidades
básicas), las cuales son ligadas por puentes de disulfuro (Brinegar y Goundan,
1993). Por lo tanto, las bandas de A9 sin 2-Me (ver carril a en la Figura 9), con
pesos moleculares de 32,4; 30,3; 22,8 y 20,7 kDa, y que en presencia de 2-Me
desaparecen (ver carril b), corresponderían a dichas globulinas.
En lo referente a la electroforesis con 2-Me, se aprecian menos bandas y de
menores pesos moleculares que sin 2-Me. Con ello se evidencia que las bandas con
masas superiores a 15 kDa están unidas por puentes disulfuro. Los enlaces de
dichas proteínas (los que ayudan a estabilizar la estructura terciaria y cuaternaria de
la proteína intra e intermolecular), en presencia del agente reductor se rompieron
para convertirse en proteínas de menor peso molecular (ver Figura 9, carril b),
específicamente de: 14,8; 12,5; 11,5 y 10,2 kDa.

31
De acuerdo a un estudio de las proteínas 2S en quinua (Brinegar y col.,
1996), estos polipéptidos tienen masas moleculares entre 8 y 9 kDa bajo
condiciones reductoras. La banda con masa molecular de 10,2 kDa, en el carril b
(ver Figura 9), podría corresponder a este tipo de polipéptido, teniendo en realidad
masas moleculares un poco inferiores a las entregada por la ecuación del estándar
de proteínas.
5.2.2 Fluorescencia
El comportamiento de los residuos triptofano, tirosina y fenilalanina puede
dar información útil para caracterizar y detectar modificaciones en proteínas. El
triptofano cuando es excitado a 295 nm genera fluorescencia que es muy sensible a
los cambios en el entorno vecino a él (Royer, 1995). Sin embargo, la intensidad de
dicho espectro es mayor que la de fenilalanina y tirosina, los que resultan apagados,
de modo que el triptofano se constituye como el fluorófobo dominante. De hecho, la
fluorescencia del triptofano persiste aunque el aminoácido forme parte de la
estructura proteica (Cheftel y col., 1989).
En la Figura 10 se presenta el espectro de emisión del aislado proteico de
quinua (A9), a distintos pH, con una misma fuerza iónica, igual a 0,5. Se graficó el
promedio de cada ensayo y su duplicado.
0
40
80
120
160
200
305 330 355 380 405 430 455 480 505
Longitud de onda (nm)
Inte
nsid
ad d
e flu
ores
cenc
ia
pH 3pH 5pH 7pH 9pH 11
Figura 10: Espectros de fluorescencia de A9 en función del pH,
a una fuerza iónica constante igual a 0,5

32
Se aprecia en la figura anterior una clara similitud entre las curvas a pH
alcalinos, lo que indicaría que la estructura de las proteínas del aislado A9 en dichas
condiciones no se vería modificada, especialmente entre los pH 7 y 11, en los
cuales las curvas son casi coincidentes.
Respecto a la intensidad de fluorescencia (ver Figura 11) se observa que
éstas son mayores a pHs alcalinos, con la excepción del pH 11, donde la intensidad
máxima es prácticamente la misma que a pH 7. Se advierte, además, que la
disminución de la intensidad de la fluorescencia a pHs ácidos (pH 3 y 5) es mayor al
incremento de la misma experimentada a pHs alcalinos. Resultados similares se
registraron en pruebas de fluorescencia a un aislado proteico de quinua obtenido a
pH 11 (A11) (Silva, 2006) y a aislados proteicos de amaranto obtenidos a pHs 9, 11
y 9-11. Dichas variaciones se pueden atribuir a cambios en el entorno del triptofano
inducidos en una misma proteína por efecto del pH o a la presencia de distintas
especies con diferente contenido de triptofano los que tendrían entornos
moleculares diferentes (Abugoch, 2006).
c
d
c
b
a
0,0
50,0
100,0
150,0
200,0
250,0
3 5 7 9 11
pH
Inte
nsid
ad d
e Fl
uore
scen
cia
Figura 11: Intensidad de fluorescencia del aislado de quinua (A9)
a distintos pHs y a una fuerza iónica constante igual a 0,5
En cuanto a los valores de longitud de onda máxima (λmáx, ver tabla 6) no
se observa una tendencia de corrimiento entre los mismos. De hecho, en lo que
respecta a los pHs 5 al 11 no existen diferencias significativas entre los valores

33
obtenidos. Sólo el espectro correspondiente al pH 3 se aleja de los restantes, con un
valor mayor.
Tabla 6: Longitud de onda máxima obtenida para el aislado A9 a distintos pHs e igual fuerza iónica (0,5)
pH Longitud de onda máxima (nm) 3 366,8 ± 0,4a 5 335,5 ± 0,7b 7 334,3 ± 1,1b 9 335,8 ± 1,1b
11 335,3 ± 1,8b Letras distintas indican diferencias estadísticamente significativas,
con un nivel de confianza del 95%
El estudio de los espectros de fluorescencia que se obtiene de los aislados
solubles a distintos pHs será la resultante de la composición de las proteínas en
cada condición y de la influencia de cada pH sobre su conformación (Abugoch,
2006). En este sentido, si se compara los resultados anteriores con los del aislado
de quinua A11 se aprecia que para ambos la forma de los espectros es similar, lo
que podría sugerir que la composición de ambos aislados es también parecida. Sin
embargo, las intensidades de los espectros de A9 son mayores que los de A11, lo
que indicaría que la composición proteica del aislado A9 posee mayor concentración
de fluorófobos que A11.
5.2.3 Espectroscopia de Absorción UV
Los usos más comunes en bioquímica de las técnicas espectroscópicas son
los de la espectroscopía ultravioleta. En dicha región, las proteínas absorben
intensamente. Las absorciones proteicas más fuertes se encuentran en dos
márgenes de longitud de onda dentro de las regiones ultravioleta, en la proximidad
de 280 y 200 nm. En el margen de 270-290 nm, vemos la absorción por las cadenas
laterales aromáticas de fenilalanina, tirosina y triptófano (Mathews y col., 2002).
En este estudio se determinó el espectro de absorción entre 250 y 450 nm a
los sobrenadantes obtenidos de la solubilización del aislado A9 a distintos pHs. En
la Figura 12 se haya el promedio de las absorbancias registradas para cada ensayo
y su duplicado a los distintos pHs estudiados.

34
0,0
0,5
1,0
1,5
2,0
240 280 320 360 400 440
Longitud de onda (nm)
Abs
orba
ncia
pH 3
pH 5pH 7
pH 9pH 11
Figura 12: Espectros de absorción del aislado proteico de quinua (A9), a
distintos pHs y una misma fuerza iónica (0,5)
Tal como se aprecia en la figura anterior, las curvas de los distintos pHs
poseen formas levemente distintas, asemejándose entre sí las correspondientes a
los pHs más ácidos (pHs 3 y 5) y las de los pHs alcalinos (pHs 9 y11), mientras
que la del pH 7 difiere de todas ellas.
En cuanto a los pHs, se puede ver en el mismo gráfico que a pHs alcalinos
las curvas se hayan más elevadas que a pHs ácidos, de modo que prácticamente
en todo el rango de longitud de onda analizado, a mayor pH aumenta la
absorbancia, con la sola excepción del pH 7, cuyo espectro supera a los de los pHs
alcalinos. Los picks encontrados podrían ser de globulinas, proteínas presentes en
la quinua (Tapia y col., 1979), y cuya absorción se encuentra en el rango de los 260
a 282 nm, de acuerdo a estudios realizados en guisantes (Chavan y col., 2001).
En pruebas efectuadas a aislados proteicos de amaranto se visualiza el
mismo tipo de curvas, pero con una absorbancia mayor (Avanza y Añón, 2005). El
motivo de esto último radica en que el amaranto posee un mayor contenido de
proteínas que la quinua (14-19%), y mayor aún si se trata de un aislado de
proteínas, lo que implica que la solución proteica va a absorber más energía, por lo
cual la absorbancia es mayor (Mathews y col., 2002).

35
5.2.4 Calorimetría Diferencial de Barrido
La calorimetría diferencial de barrido (DSC) mide la capacidad calorífica
relativa de un sistema determinado cuando sufre una transición inducida por un
cambio térmico, como por ejemplo la desnaturalizacion de una proteína. Para ello,
se someten dos celdas, una con la muestra y otra con la referencia, a un programa
de calentamiento a una velocidad de flujo de calor controlado. Cuando se produce
una transición inducida por la temperatura, la muestra absorbe parte del calor que
se le está suministrando a la celda. Como resultado, la célula de la muestra tendría
una temperatura algo inferior a la de la celda de referencia, pero el sistema de
control del equipo suministra una potencia calorífica adicional para que ambas
celdas mantengan la misma temperatura. Esta potencia en exceso es proporcional a
la diferencia de capacidades caloríficas entre las dos celdas y sus contenidos
(Beldarraín, 2001).
En la Figura 13 se muestra el termograma del aislado de quinua A9 obtenido
por DSC.
Integral -13.50 mJPeak 98.12 °CLeft Limit 84.80 °CRight Limit 103.44 °CHeating Rate 10.00 °Cmin^-1
A9 en agua, 05.10.2006 11:08:49A9 en agua, 16.4600 mg
mW
-11.5
-11.0
-10.5
-10.0
-9.5
-9.0
-8.5
-8.0
min
°C55.0 60.0 65.0 70.0 75.0 80.0 85.0 90.0 95.0 100.0 105.0 110.0
4.5 5.0 5.5 6.0 6.5 7.0 7.5 8.0 8.5 9.0 9.5 10.0
^exo A9 en agua 1 05.10.2006 11:37:35
Universidad de Santiago de Chile: Lab. Prop. Fisicas SystemeRTAMETTLER TOLEDO S Figura 13: Termograma (DSC) de A9 en una suspensión al 20% en agua
En el termograma se aprecia una endoterma que denota un cierto grado de
estructura de las proteínas del aislado suspendidas al 20 % en agua y una
temperatura de desnaturalización (Td) de la muestra de 98,1 ± 0,1 ºC. En un ensayo,
bajo las mismas condiciones, el aislado proteico de quinua obtenido a pH 11 (A11)
tuvo una endoterma muy pequeña, de 1,35 mJ (Silva, 2006), en comparación con la

36
del aislado A9, cuya endoterma fue de 13,5 mJ, lo que evidencia el efecto de la
obtención de aislados a un pH tan alcalino sobre la estructura de las proteínas, al
mismo tiempo que corrobora el hecho de que las globulinas presentes en la quinua
(Abugoch, 2006; Martínez y Añón, 1996), al ser extraídas a pH 9, mantienen un
mayor grado de estructura.
Para el aislado de quinua suspendido al 20% en buffer de pH 9, en tanto, la
transición térmica es mayor en comparación a la estudiada en medio acuoso, con
una temperatura de desnaturalización (Td) igual a 100,5 ± 5,6 ºC y una endoterma
de mayor área, en la zona que va desde los 87,2 a 112,9 ºC, y equivalente a 29,1
mJ (ver Figura 14). Se pudo observar que el aislado de quinua suspendido a pH 9
aumenta el grado de estructura de sus proteínas. Situación similar ocurre con el
aislado A11, el que aunque de todos modos se presenta prácticamente
desnaturalizado, suspendido en pH 9 muestra una endoterma con un área mayor
que en agua (Silva, 2006).
Integral -29.11 mJPeak 104.44 °CLeft Limit 87.16 °CRight Limit 112.89 °CHeating Rate 10.00 °Cmin^-1
A9 pH 9 2, 05.10.2006 12:44:07A9 pH 9 2, 17.0300 mg
mW
-1.8
-1.6
-1.4
-1.2
-1.0
-0.8
-0.6
min
°C65.0 70.0 75.0 80.0 85.0 90.0 95.0 100.0 105.0 110.0
5.5 6.0 6.5 7.0 7.5 8.0 8.5 9.0 9.5 10.0
^exo A9 pH 9 2 05.10.2006 12:45:24
Universidad de Santiago de Chile: Lab. Prop. Fisicas SystemeRTAMETTLER TOLEDO S Figura 14: Termograma (DSC) de A9 suspendido al 20% en buffer a pH 9
En el análisis calorimétrico realizado a un aislado proteico de amaranto
obtenido a pH 9, y suspendido a distintos pHs, se obtuvo resultados similares a los
de A9, con condiciones de mayor termoestabilidad que para el aislado de amaranto
obtenido a pH 11 (Abugoch, 2006). En dicho caso, la temperatura de
desnaturalización de las proteínas del amaranto fue un poco mayor a pH 7 que la de

37
A9, de alrededor de 103 ºC, mientras que a pH 9 la temperatura fue de 100 ºC, valor
casi idéntico que el del aislado de quinua A9.
5.3 Determinación de las Propiedades Funcionales
5.3.1 Solubilidad
El uso exitoso de fuentes de proteínas vegetales en la formulación de
alimentos depende de las propiedades funcionales de la materia prima. En ese
sentido, la baja solubilidad de un aislado proteico evidentemente limita su uso en
algunos tipos de productos en la industria alimentaria, pero en otros podría
favorecerlos. Sin embargo, esta propiedad puede ser modificada por la influencia de
varios factores, como el pH, la concentración de sal, la constante dieléctrica del
solvente y la temperatura (Bora y Riveiro, 2004).
En la Figura 15 se muestra la curva de tendencia de solubilidad del aislado
A9 a distintos pHs, y con una fuerza iónica constante de 0,5. En él se evidencia una
notoria diferencia de solubilidad entre los pHs más ácidos (pHs 3 y 4), con niveles
muy bajos de solubilidad, y los restantes, con valores superiores al 75%. Así, la
mínima solubilidad se encuentra a pH 3, siendo de 7,6%. Le sigue muy de cerca el
pH 4, con 11,3 %. El resto de los pHs, presentan solubilidades que van desde
76,6% a pH 5, llegando a un máximo de 94,6% a pH 11.
a a
b bcd de cd bc e
0
10
20
30
40
50
60
70
80
90
100
2 3 4 5 6 7 8 9 10 11 12
pH
% S
olub
ilida
d
Figura 15: Solubilidad del aislado proteico de quinua A9 a distintos pHs a una
fuerza iónica constante de 0,5. Letras distintas indican diferencias estadísticamente significativas, con un nivel de confianza del 95%

38
Las variaciones de la solubilidad con el pH, está relacionada con la
modificación de la carga neta de las proteínas y por lo tanto con su balance
electrostático; en la zona cercana a su pI la carga neta de las proteínas tiende a 0 y
la variación de la solubilidad es mínima debido al aumento de la atracción entre las
moléculas. Del lado alcalino al pI las proteínas tendrán una carga neta negativa y
probablemente mayor que la carga neta positiva que presenta en medio ácido, de
este modo las fuerzas repulsivas a pHs alcalinos y ácidos extremos serán más
importantes que las fuerzas de atracción aumentando la solubilidad (Abugoch,
2006).
En un estudio anterior en que se investigó la solubilidad de la harina de
quinua en función del pH, la curva obtenida describe una “U” con un mínimo de
solubilidad, de aproximadamente el 15%, a pH 6, y un máximo cercano al 50%, a pH
10, aunque a pHs ácidos también se obtuvo solubilidades medianamente altas, de
hecho, a pH 2 la solubilidad fue cercana al 40% (Ogungbenle, 2003). Ello implica
que el pI de las proteínas de la harina de quinua es el pH 6, superior al pI obtenido
para el aislado A9, el que se encontraría próximo al pH 3. Por lo tanto, la
aplicabilidad del aislado A9 no es la misma que para las proteínas de su harina,
puesto que mientras estas últimas serían apropiadas para bebidas con alta acidez,
el aislado sería más ventajoso en formulaciones de bebidas con pHs iguales o
superiores a 5.
Siguiendo con lo anterior, debe colocarse atención en el hecho de que las
solubilidades para el aislado A9, en la región que va desde el pH 5 al 7 (pHs
normales en alimentos) son muy superiores a las solubilidades obtenidas por las
proteínas de la harina de quinua, pues los resultados para el aislado prácticamente
quintuplican los de la harina. Estos resultados estarían demostrando que se puede
mejorar esta propiedad de la quinua, preparando un aislado proteico.
Asimismo, las solubilidades obtenidas son muy superiores a las exhibidas
por el aislado de quinua A11. En dicho caso la máxima solubilidad también se
produjo en la zona alcalina, sin embargo ésta fue de sólo un 41,4%, a pH 11 (Silva,
2006). El motivo de dichas disconformidades podría atribuirse a diferencias
estructurales entre las proteínas de los aislados en cuestión.
Por otra parte, los resultados son concordantes con los presentados por
aislados proteicos de amaranto, en cuanto al aumento de la solubilidad a pHs

39
alcalinos, con la diferencia de que en dicho caso los aislados obtenidos a distintos
pHs (9, 11 y 9-11) exhibieron solubilidades inferiores entre los pHs 5 a 8. En efecto,
a pH 5 los aislados tuvieron entre un 20 y 30% de solubilidad, mientras que para los
pHs 6 a 8 dicha solubilidad se encontró entre un 30 y un 60% (Abugoch, 2006). Esta
diferencia con el amaranto (considerado pseudocereal, al igual que la quinua), en la
zona de pHs ácidos, estaría señalando diferencias en los pI de sus respectivas
proteínas que los componen como aislados.
La misma tendencia en su solubilidad la han presentado proteínas de otros
alimentos. Es el caso del aislado proteico del trigo sarraceno (Tomotake y col.,
2002), el que aumenta su solubilidad a pHs alcalinos, alcanzando un máximo de
55%, a pH 10; del aislado proteico de mucuna (Adebowale y Lawal, 2003), con un
máximo de solubilidad del 96%, a pH 12; y del aislado proteico de salvado de arroz
(Wang y col., 1999), el que presentó una solubilidad máxima de 82%, a pH 10. Cabe
señalar que estos aislados preseen un punto isoeléctrico cercano al pH 4, un poco
superior al del aislado A9 (próximo al pH 3).
5.3.2 Capacidad de Retención de Agua
La capacidad de retención de agua de las proteínas, WHC, es un índice
importante en la evaluación del comportamiento de las mismas como ingredientes
en productos de panadería, carnes embutidas, salchichas y geles alimentarios. Esta
propiedad afecta no sólo las condiciones del procesamiento, sino también la calidad
final de los productos (Abugoch, 2006).
En la Figura 16 se muestran los resultados de la capacidad de retención de
agua para el aislado A9, en agua (control) y en buffers de distintos pHs. En la
gráfica se aprecia que no existen diferencias estadísticamente significativas (p>0,05)
entre los diversos pHs, de modo que los valores encontrados van de un mínimo de
2,5 mL/g aislado proteico, a pH 5, hasta un valor máximo de 3,0 mL agua/g aislado,
a pH 3.
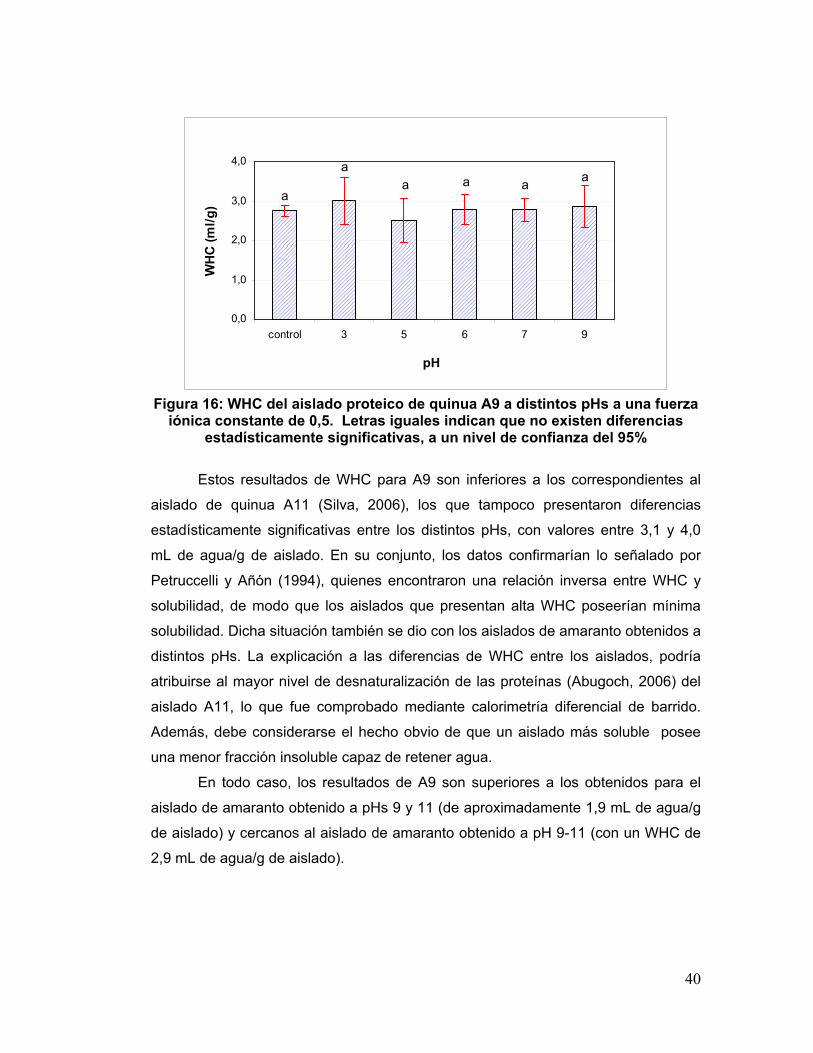
40
a
aa a a a
0,0
1,0
2,0
3,0
4,0
control 3 5 6 7 9
pH
WH
C (m
l/g)
Figura 16: WHC del aislado proteico de quinua A9 a distintos pHs a una fuerza
iónica constante de 0,5. Letras iguales indican que no existen diferencias estadísticamente significativas, a un nivel de confianza del 95%
Estos resultados de WHC para A9 son inferiores a los correspondientes al
aislado de quinua A11 (Silva, 2006), los que tampoco presentaron diferencias
estadísticamente significativas entre los distintos pHs, con valores entre 3,1 y 4,0
mL de agua/g de aislado. En su conjunto, los datos confirmarían lo señalado por
Petruccelli y Añón (1994), quienes encontraron una relación inversa entre WHC y
solubilidad, de modo que los aislados que presentan alta WHC poseerían mínima
solubilidad. Dicha situación también se dio con los aislados de amaranto obtenidos a
distintos pHs. La explicación a las diferencias de WHC entre los aislados, podría
atribuirse al mayor nivel de desnaturalización de las proteínas (Abugoch, 2006) del
aislado A11, lo que fue comprobado mediante calorimetría diferencial de barrido.
Además, debe considerarse el hecho obvio de que un aislado más soluble posee
una menor fracción insoluble capaz de retener agua.
En todo caso, los resultados de A9 son superiores a los obtenidos para el
aislado de amaranto obtenido a pHs 9 y 11 (de aproximadamente 1,9 mL de agua/g
de aislado) y cercanos al aislado de amaranto obtenido a pH 9-11 (con un WHC de
2,9 mL de agua/g de aislado).

41
5.3.3 Capacidad de Absorción de Agua
La capacidad para absorber agua es considerada una propiedad funcional de
las proteínas, fundamental en alimentos viscosos tales como sopas, salsas, masas y
alimentos horneados, productos donde se requiere una buena interacción proteína-
agua (Granito y col., 2004).
En la Figura 17 se observa la curva de absorción de agua en función del
tiempo para el aislado A9. A partir de dicha gráfica se obtuvo la capacidad de
absorción de agua (WIC), la que no es más que el volumen máximo de agua
absorbido por gramo de aislado. La WIC para A9 fue de 1,8 ± 0,3 g agua/g aislado.
El tiempo para alcanzar la WIC o el equilibrio -esto es, donde el aislado es incapaz
de seguir captando agua espontáneamente- se denomina tiempo de estabilización
(te). El te para A9 fue de 3,9 min. Un tercer parámetro cinético es la velocidad inicial
(vi) con que el aislado absorbe agua. Dicho valor se obtiene calculando la pendiente
del gráfico, en los primeros instantes en que aumenta el volumen de agua. Para A9
la vi fue de 1,34 g agua/ g aislado × min.
Figura 17: Agua absorbida por el aislado A9 en función del tiempo
En pruebas de WIC a aislados proteicos de amaranto los resultados
mostraron que cuánto más desnaturalizadas se encontraban las proteínas, mayor

42
era la capacidad de imbibición de agua. Es así como la WIC registrada para el
aislado de amaranto obtenido a pH 9 fue de 1,7 mL de agua/g aislado, valor muy
cercano al del aislado de quinua A9, e inferior a los aislados de amaranto extraídos
a pHs 11 y 9-11, que fueron de 2,5 mL de agua/g aislado en ambos casos
(Abugoch, 2006). Este efecto estaría dado por el incremento de la accesibilidad a
las proteínas y en consecuencia a los aminoácidos polares, los cuales tienen una
gran afinidad por el agua, produciéndose un incremento en la capacidad para
absorber agua (Granito y col., 2004).
La WIC expuesta por el aislado de quinua A11, el que presentó una mayor
desnaturalización en su estructura que el aislado A9, fue mayor a la de A9,
alcanzando los 2,8 mL de agua/g aislado (Silva, 2006) lo que ratificaría lo expuesto
en el párrafo anterior.
La velocidad de absorción de agua para el aislado A9 fue bastante mayor
que para el amaranto al mismo pH, el que alcanzó sólo 0,5 mL agua/g aislado × min.
También sobrepasó al aislado de amaranto a pH 9-11 (0,4 mL agua/g aislado × min),
acercándose al aislado de amaranto de pH 9-11, el que tuvo una vi de 1,1 mL
agua/g aislado × min. Fue inferior sí al aislado de quinua A11, el que alcanzó una vi
de 3,5 mL agua/g aislado × min.
En lo relativo a los tiempos de estabilización, el tiempo determinado para A9
fue mucho mayor que para los aislados de amaranto extraídos a pHs 9 y 11 (0,9 y
0,6 respectivamente), pero inferior al aislado de amaranto obtenido a pH 9-11, que
fue de 5 min. El tiempo requerido para alcanzar el equilibrio por A11, por su parte,
fue de apenas 0,8 min.

43
VI. CONCLUSIONES
A partir del proceso de extracción de las proteínas de la harina de quinua a
pH 9, se obtuvo un aislado proteico con un contenido de proteínas del 77,2%
y un perfil de aminoácidos balanceado, muy similar al de la harina de quinua,
lo cual podría motivar su empleo como ingrediente en la formulación de
alimentos destinados al consumo humano y, en especial, de alimentos
funcionales altamente proteicos.
Los estudios de los perfiles polipeptídicos, a través de la técnica PAGE, en
condiciones nativas y desnaturantes, reveló la presencia de proteínas del tipo
albúminas 2S y globulinas 11S, las cuales están presentes en la quinua.
A través de los estudios espectroscópicos de fluorescencia y UV se
evidenció el efecto del pH sobre la estructura de las proteínas del aislado,
constatándose un efecto mayor a pHs alcalinos que los cambios sufridos a
pHs ácidos, con diferencias leves entre pHs correspondientes a un mismo
grupo (ácidos o alcalinos).
El aislado presentó una solubilidad bastante alta, la que aumentó con el
incremento del pH, registrándose valores bastante mayores que para la
harina de quinua y semejantes o superiores a los reportados en la literatura
para otros aislados proteicos. Esta mayor solubilidad del aislado respecto a
su harina, estaría dada por la desnaturalización de las proteínas durante el
proceso de obtención del aislado. La alta solubilidad del aislado lo hace
propicio para su utilización en la formulación de sopas y bebidas,
enriquecidas en proteínas.
No se encontró una correlación entre la capacidad de retención de agua
(WHC) y el pH, y por tanto, con el grado de desnaturalización de las
proteínas. De todos modos, los valores obtenidos son apropiados para la
elaboración de productos de panadería, embutidos y geles alimentarios,
entre otros.

44
El nivel de capacidad de absorción de agua (WIC) del aislado es apropiado
para el desarrollo de productos deshidratados y alimentos viscosos en
general.
A partir de los resultados de solubilidad se puede presumir una alta
capacidad espumante en el aislado, debido a que ambas propiedades serían
directamente proporcionales. Dicha capacidad se evidenció a través del
manejo del aislado, pero deberá ser evaluada en estudios posteriores.
Asimismo, se vislumbra la necesidad de estudiar otras propiedades
funcionales, como las propiedades texturales, y de investigaciones centradas
en el aspecto tecnológico de la obtención de aislados proteicos de quinua a
nivel industrial, que permitan la obtención del aislado con mayores niveles de
rendimiento.

45
VII. REFERENCIAS BIBLIOGRÁFICAS
Abugoch James, Lilian Elizabeth. Relación Estructura-Funcionalidad de
Glutelinas y Aislados Proteicos de Amaranto (Amaranthus hypochondiacus).
Tesis doctoral, Buenos Aires, Argentina. Universidad Nacional de La Plata,
Facultad de Ciencias Exactas, 2006.
Alaiz, M.; Navarro, J.; Girón, J. y Vioque, E. Amino Acid Analisis by High-
Performance Liquid Chromatography after Derivatization with Diethyl
Ethoxymethylenemalonate, J. of Chromatogr. 591:181-186, 1992.
Adebowale, K. y Lawal, O. Foaming, Gelation and Electrophoretic
Characteristics of Mucuna Bean (Mucuna prueriens) Protein Concentrates.
Food Chem. 83: 237-246, 2003.
Albarrán, R. Estudio de Algunos Componentes Químicos, Caracteres
Morfoanatómicos y Patrones Proteicos en Semillas de dos Ecotipos de
Quinoa (Chenopodium Quinoa Willd). Tesis (Ingeniero Agrónomo). Chillán,
Chile. Universidad de Concepción, Facultad de Agronomía. 1993.
Añón, M. C.; Genovese, M.; Jovanovich, G.; Lajolo, F.; Martínez, E. y Pilosof,
A. Caracterización Funcional y Estructural de Proteínas, Editorial
Universitaria de Buenos Aires, Buenos Aires, Argentina, 2000.
Añón, M. C.; Sorgentini, D. y Wagner, J. Relationships between Different
Hydration Properties of Comercial and Laboratory Soybean Isolates. J. Agric.
Food. Chem. 49: 4852-4858, 2001.
AOAC - Association of Official Analytical Chemists Inc. Official Methods of
Analysis, 16th ed; Ed.Williams, S.; Arlington, VA., 1995.
Araneda Godoy, Giovanna. Obtención, Caracterización y Estudio de Vida Útil
de la Harina Integral de Quinua. Memoria (Ingeniero en Alimentos). Santiago,

46
Chile. Universidad de Chile, Facultad de Ciencias Químicas y Farmacéuticas,
2004.
Avanza, M. y Añón, M.C. Structural Characterization of Amaranth Protein
Gels. J. Food Sci. 70(3):223-229, 2005.
Azcui, M. El tesoro Dorado de Quechuas y Aymaras. Quinua: Grano de Oro.
[en línea]
<http://revista.serindigena.cl/props/public_htmL/?module=displaystory&story_
id=761&format=htm> [consulta: 10 de septiembre 2006].
Beldarraín, A. Aplicaciones de la Calorimetría Diferencial de barrido al
Estudio de la Estabilidad de las Proteínas. Biotecnología Aplicada 18:10-16,
2001.
Berti, C.; Ballabio, C.; Restan, P.; Porrini, M.; Bonomi, F. y Lametti, S.
Immunochemical and Molecular Properties of Proteins in Chenopodium
Quinoa. Cereal Chem. 81 (2):275-277, 2004.
Bradford, M. M. A. Rapid and Sensitive Method for the Quantitation of
Micrograms Quantities of Protein Utilizing the Principle of Protein-Dye
Binding. Anal. Biochem. 72: 248 – 254, 1976.
Braverman, S. Introducción a la Bioquímica de Alimentos. El Manual
Moderno, México. 1980.
Brinegar, C.; Sine, B. y Nwokocha, L. High-cysteine 2S Seed Storage
Proteins from Quinoa (Chenopodium Quinoa). J. Agric. Food Chem. 44(7):
1621–1623, julio 1996.
Brinegar, C. y Goundan, S. Isolation and Characterization of Chenopin, the
11S Seed Storage Protein of Quinoa (Chenopium Quinoa). J. Agric. Food
Chem. 41 (2): 182 – 185, 1993.

47
Castellani, O. Caracterización Fisicoquímica y Estructural de Globulina P de
Amaranto. Tesis Doctoral, Buenos Aires, Argentina. Universidad Nacional de
La Plata, 2001.
Ccbolgroup. Quinoa-Quinua: Antecedentes. [en línea].
<http://ccbolgroup.com/quinoaTodo.htmL#ANTECEDENTES> [consulta: 25
de agosto de 2006].
Chauhan, G.; Eskin, N. Y Tkachuk, R. Nutrients and Antinutrients in Quinoa
Seed. Cereal Chem. 69 (1):85-88, 1992.
Cheftel, J.; Cud, J. y Lorient, D. Proteínas Alimentarias. Editorial Acribia.
España. 1989.
CIED - Centro de Investigación, Educación y Desarrollo. Quinua:
Chenopodium Quinoa Willdenow [en línea]
<http://www.ciedperu.org/productos/quinua.htm> [consulta: 26 de septiembre
de 2006].
Curare. Péptidos de Girasol: Antecedente a los Hidrolizados Proteicos. [en
línea] <http://www.curare.com/Proteina%20Vegetal.htm> [consulta: 24 de
marzo de 2006].
Diario Pyme – Diario de la Pequeña y Mediana Empresa. Acuerdo Busca
Introducir Quinoa en Europa. (2003) [en línea]
<http://www.diariopyme.cl/newtenberg/1550/article-56781.htmL> [consulta:
10 de octubre de 2006].
Drzewiecki, J.; Delgado-Licon, E.; Haruenkit, R.; Pawelzik, E.; Martin-Belloso,
O.; Park Y.; Jung, S.; Trakhtenberg, S. y Gorinstein, S. Identification and
Differences of Total Proteins and Their Soluble Fractions in Some
Pseudocereals Based on Electrophoretic Patterns. J. Agric. Food Chem. 51
(26): 7798 – 7804, 2003.

48
FAO - Organización de las Naciones Unidas para la Agricultura y la
Alimentación. Contenido en Aminoácidos de los Alimentos y Datos Biológicos
sobre las Proteínas”. Roma, Italia. 1970. Colección Alimentación y Nutrición.
Fontúrbel, F. Problemática de la Producción y Comercialización de
Chenopodium Quinoa W. (Chenopodiaceae), debida a la Presencia de las
Saponinas”. 2003. [en línea]
<http://cabierta.uchile.cl/revista/21/articulos/pdf/paper6.pdf> [consulta: 25 de
septiembre de 2006].
Gajardo Repetto, Pilar Inés. Caracterización y Determinación de la
Estabilidad Durante el Almacenamiento de las Proteínas de Harina de
Quinua Orgánica sin Pulir y Pulida Proveniente de la VI Región de Chile.
Santiago, Chile. Memoria (Ingeniero en Alimentos). Universidad de Chile,
Facultad de Ciencias Químicas y Farmacéuticas, 2005.
Giese, J. Proteins as Ingredients: Types, Functions, Applications. Food Tech.
50-60, octubre, 1994.
Granito, M.; Guerra, M.; Torres, A. y Guinand, J. (2004) Efecto del
Procesamiento sobre las Propiedades Funcionales de Vigna Sinensis. [en
línea] <http://www.scielo.org.ve/scielo.php?script=sci_arttext&pid=S0378-
18442004000900009&lng=es&nrm=iso> [consulta: 20 de octubre de 2006].
IESN Chile – Instituto de Estudios de Salud Natural de Chile. Todo Sobre la
Quinoa. [en línea] <http://www.geocities.com/iesnchile/quinoa.htmL> (2001)
[consulta: 30 de octubre de 2006].
Junge, I. y Cerda, Patricia. Estado Actual de las Investigaciones sobre
Quioa para la Alimentación Humana. Alimentos 3 (1):19-25, 1978.
LaemmLi U. K. Cleavage of Structural Proteins During the Assembly of
Bacteriophage T4. Nature 227: 680 – 685, 1970.

49
Ma, C. y Karwalkar, V. Study of Termal Denaturation of Oat Globulin by
Ultraviolet and Fluorescence Spectrophotometry. J. Agric. Food Chem. 36:
155-160, 1988.
Martínez E., Nora. Caracterización de Proteínas de Amaranto, Tesis
Doctoral. Buenos Aires, Argentina. Universidad Nacional de La Plata. 1997
Martínez, N. y Añón, M.C. Composition and Structural Characterization of
Amaranth Protein Isolates. An Electrophoretic and Calorimetric Study. J.
Agric. Food Chem. 44: 2523-2530, 1996.
Mathews, C.; Van Holde, K.E. y Ahern, K. Bioquímica. 3ª ed. Madrid, España.
Editorial Pearson Educación S.A. 2002.
Ogungbenle, H. Nutritional Evaluation and Functional Properties of Quinua
(Chenopodium quinoa) Flour. Int. J. Food Sci. Nutr. 54:153-158, 2003.
Osborne, D. R. y Voogt, P. Análisis de los Nutrientes de los Alimentos.
Editorial Acribia S.A. Zaragoza, España. 1986.
Pearson, D. Chemical Analysis of Foods. 6th edition. London Churchill. New
York, U.S.A. 1970.
Pennacchiotti, I. Las Proteínas: Generalidades y su Importancia en Nutrición
y en la Industria de Alimentos. Santiago, Chile. Impresos Universitaria S.A.
1998.
Permyakov, E. Luminescent Spectroscopy of Proteins. USA, CRC Press Inc,
1993.
Primo, E. Química Agrícola III Alimentos. España. Editorial Alambra S.A.
1979.

50
Robinson, D. Bioquímica. Valor Nutritivo de los Alimentos. España. Editorial
Acribia. 1991.
Royer, C. Approaches to Teaching Fluorescence Spectroscopy. Biophys. J.
68:1191-1192, 1995.
Ruales y Nair. Nutricional Quality of the Protein in Quinoa (Chenopodium
Quinoa willd) Seeds. Plant Foods Hum Nutr. 42(1): 1-11, Jan 1992.
Schmidt-Hebbel, H. Avances en Ciencia y Tecnología de los Alimentos.
Santiago, Chile.1981.
Schmidt-Hebbel, H., Pennachiotti, I.; Masson, L. y Mella, M.A. Tabla de
Composición Química de Alimentos Chilenos, 8ª edición. Santiago, Chile.
1992.
Scilingo, Adriana. Propiedades Estructurales y Funcionales de Aislados
Proteicos de Soja Modificados. Efecto de la Presencia de Calcio. Tesis
Doctoral. Buenos Aires, Argentina. Universidad Nacional de la Plata,
Facultad de Ciencias Exactas, 2000.
Sepúlveda, J., Thomet, M.; Palazuelos, P., Mujica, M. La Kinwa Mapuche.
Recuperación de un Cultivo para la Alimentación. Temuco, Chile. 2004.
Silva Manzo, Jorge Andrés. Obtención, Caracterización y Relación
Estructura-Funcionalidad de un Aislado Proteico de Quinua (Chenopodium
Quinoa) Orgánica Proveniente de la VI Región de Chile. Memoria (Ingeniero
en Alimentos). Santiago, Chile. Universidad de Chile, Facultad de Ciencias
Químicas y Farmacéuticas, 2006.
Tapia, M.; Gandarillas, H.; Alandia, S.; Cardozo, A.; Mujica, A.; Ortiz, R.;
Otazu, V.; Rea, J.; Salas, B. y Sanabria, E. La Quinua y La Kañiwa: Cultivos
Andinos. Bogotá, Colombia. Editorial IICA. 1979.

51
Tomotake, H.; Shimaoka, I.; Kayashita, J.; Nakajoh, M. y Kato, N.
Physicochemical and Functional Properties of Buckwheat Protein Product. J.
Agric. Food Chem. 50:2125-2129, 2002.
UB–Universidad de Barcelona. Métodos en Biología Celular: Electroforesis
en Gel de Poliacrilamida. [en línea]
<http://www.ub.es/biocel/wbc/tecnicas/page.htm> [consulta: 20 de octubre de
2006].
Valenzuela, P. Evaluación del Comportamiento de Harina de Cinco Ecotipos
de Quinoa (Chenopodim quinoa Willd) en la Elaboración de Galletas”. Tesis
(Ingeniero Agrónomo mención Agroindustria y Tecnología de los Alimentos).
Santiago, Chile. Universidad de Chile, Facultad de Ciencias Agrarias y
Forestales, 1997.
Vilches, C.; Gely, M.; y Santana, E. Physical Properties of Quinoa Seeds.
Biosystems Engineering 86 (1): 59-65, 2003.
Villarroel Véliz, Andrea Carolina. Obtención de Harina de Quinua Proveniente
de dos Ecotipos de Semillas, Caracterización Bioquímica y Funcional y
Determinación de la Estabilidad de las Proteínas durante el Almacenamiento
a Temperaturas Disímiles. Memoria (Ingeniero en Alimentos). Santiago,
Chile. Universidad de Chile, Facultad de Ciencias Químicas y Farmacéuticas,
2005.
Wahli, C. Quinua: Hacia su Cultivo Comercial. Quito, Ecuador. Latinreco S.A.
1990.
Wang, G.; Kuan, S.; Francis, O.; Ware, G. y Carman, A. A Simplified HPLC
Method for the Determination of Phytoestrogens in Soybean and Its
Processed Products. J. Agric. Food Chem. 38 (1): 185-190, 1990.

52
Wang, M.; Hettiarachchy, N. S.; Burks, M. Q.; W. y Siebenmorgen, T.
Preparation and Functional Properties of Rice Bran Protein Isolate. J. Agric.
Food Chem. 47: 411- 416, 1999.
Zamudio, T. (2003). Quinua. Programa Panamericano de Defensa y
Desarrollo de la Diversidad Biológica Cultural y Social – ProDiversitas [en
línea] <http://www.prodiversitas.bioetica.org/quinua.htm> [consulta: 27 de
marzo de 2006].

53
ANEXOS

54
ANEXO 1
Métodos Electroforéticos para la Caracterización de los Perfiles Polipeptídicos
Reactivos:
Tampón de carga (pH 8,3): disolver 3,0 g de Tris, 14,4 g de glicina y 1,0 g de
SDS en un litro de agua destilada. En la preparación del buffer nativo no
incorporado SDS.
Buffer del gel de concentración (pH 6,8): disolver 6,0 g de Tris y 0,4 g de
SDS en 40 mL de agua destilada. Ajustar el pH con HCl 4 N, y completar el
volumen a 100 mL. En la preparación del buffer nativo no incorporar SDS.
Buffer del gel de separación (pH 8,8): disolver 18,2 g Tris y 0,4 g de SDS en
40 mL de agua destilada. Ajustar con HCl 4 N, y completar el volumen a 100
mL. En la preparación del buffer nativo no incorporar SDS.
Solución de persulfato de amonio: disolver 200 mg de persulfato de amonio
en 2 mL de agua destilada, eliminar el gas de la solución y congelar en
pequeñas alícuotas. Esta solución permanece estable por aproximadamente
cuatro años.
Solución colorante: disolver 1,25 g de Coomassie Blue R en 227 mL de
metanol y 46 mL de ácido acético glacial. Completar con agua destilada a
500 mL.
Solución decolorante: mezclar 70 mL de ácido acético glacial, 300 mL de
etanol y completar el volumen a 1 litro con agua destilada.
Solución acrilamida / Bis-acrilamida: disolver 30 g de acrilamida y 0,8 g de
bis-acrilamida en 100 mL de agua destilada. Almacenar en frasco oscuro a 4
ºC.
Buffer de muestra desnaturante con β-mercapto: mezclar 2,5 mL buffer de
gel de concentración (pH 6,8), 1,2 g de SDS, 4 mL de glicerol, 2 mL de β-
mercapto y punta de espátula de azul de bromofenol. Completar a 10 mL.
Buffer de muestra desnaturante: mezclar 2,5 mL buffer de gel de
concentración (pH 6,8), 1 g de SDS, 0,4 g de sacarosa y punta de espátula
de azul de bromofenol. Completar a 10 mL.

55
Buffer de muestra nativa: mezclar 4 mL buffer de gel de concentración (pH
6,8) (nativo), 4 mL de glicerol y punta de espátula de azul de bromofenol.
Completar a 10 mL.
TEMED
Agua destilada
Butanol
Estándar de peso molecular
Alcohol
Procedimientos: Preparación de las Muestras para Electroforesis (PAGE)
Pesar 20 mg de muestra en un tubo ependorf
Agregar 200 ul de solución nativa, desnaturante o desnaturante con β-
mercapto, según corresponda.
Agitar breve e intensamente en vortex a intervalos de 5 minutos, durante 15
minutos.
Centrifugar a temperatura ambiente por 15 minutos, a 10.000 rpm.
Recolectar el sobrenadante en otro tubo ependorf. Almacenar los tubos en
congelador hasta llevar a cabo la electroforesis.
Cuando se desee ocupar las muestras, calentarlas durante 1 minuto en agua
hirviendo (esto se hace sólo con las muestra desnaturantes), insertando los
tubos en plumavit para que floten.
Procedimiento para Electroforesis Nativa (PAGE-nativa) Los geles se preparan entre dos placas de vidrio perfectamente limpias. Para
ello, las placas deben lavarse con detergente, enjuagarse con abundante
agua destilada y por último ponerles alcohol y secarlos con toalla nova.
Colocar los vidrios en el soporte del equipo Bio-Rad, procurando que queden
derechos y fijos al soporte.
Preparar gel al 5 %, mezclando: 2.653 µl de agua destilada, 1.750 µl de
acrilamida/bisacrilamida (A/BA), 1.820 µl de buffer a pH 8,8, 70 µl de PSA al
10% y 7 µl de TEMED, con agitación.

56
Colocar la mezcla en seguida entre los vidrios, con la ayuda de una
micropipeta (hasta que rebalse).
Usando guantes colocar el peine, procurando que no queden burbujas.
Esperar hasta que gelifique por completo.
Retirar los vidrios del soporte y colocarlos en la cubeta de corrida del equipo
Bio-Rad, con el peine hacia adentro.
Retirar el peine y cargar cuidadosamente las muestras con la ayuda de una
micropipeta.
Llenar la cubeta con el tampón de carga (pH 8,3).
Tapar la cuba de corrida y conectarla a la fuente de poder. Programarla a
200 voltios por aproximadamente 40 minutos, o hasta que la muestra haya
alcanzado el borde inferior del gel.
Apagar el equipo y remover los vidrios de la cuba de corrida.
Retirar el gel cuidadosamente con la ayuda de una espátula o con las
manos, usando guantes, y colocarlo estirado en un recipiente que contenga
solución colorante, idealmente de un día para otro.
Retirar la solución colorante y adicionar solución decolorante. Cambiar la
solución a medida que se va tornando azulada. Esta solución puede ser
reaprovechada adicionándole carbón activado y filtrándola.
Procedimiento para Electroforesis Desnaturante (PAGE-SDS)
Los geles se preparan entre dos placas de vidrio perfectamente limpias.
Colocar los vidrios en el soporte del equipo Bio-Rad, procurando que queden
derechos y fijos al soporte.
Preparar el gel separador al 12 %, mezclando: 1.091 µl de agua destilada,
2.004 µl de A/BA, 1.300 µl de buffer a pH 8,8, 50 µl de SDS al 10%, 50 µl de
PSA al 10% y 5 µl de TEMED, con agitación.
Colocar en seguida 3,5 mL del gel separador entre los vidrios, con la ayuda
de una micropipeta. Cubrir la superficie con agua saturada con butanol, la
que debe ser adicionada lentamente.
Esperar a que la solución gelifique por completo, lo cual se evidencia por la
formación de una línea divisoria nítida entre el gel y el agua con butanol.

57
Sacar los vidrios del soporte y retirar la capa de agua saturada con butanol,
lavando con abundante agua destilada.
Secar entre los vidrios con papel absorbente.
Preparar gel de concentración al 5 %, mezclando: 1.050 µl de agua
destilada, 250 µl de A/BA, 190 µl de buffer a pH 6,8, 15 µl de SDS al 10%,
15 µl de PSA al 10% y 1,5 µl de TEMED, con agitación.
Colocar en seguida 1,5 mL (o hasta que rebalse) gel concentrador entre los
vidrios (sobre el gel separador), con la ayuda de una micropipeta.
Usando guantes colocar el peine, procurando que no queden burbujas.
Esperar a que el gel concentrador gelifique por completo.
Retirar los vidrios del soporte y colocarlos en la cubeta de corrida del equipo
Bio-Rad, con el peine hacia adentro. Continuar el procedimiento de igual
forma que para electroforesis nativa.

58
ANEXO 2
Método de Bradford
Reactivos: Solución estándar de proteína (BSA): Pesar en balanza analítica 10 mg
exactos de albúmina de bovino, completar con 990 µl de agua destilada y
disolver completamente. Luego, colocar 100 µl de esta solución y colocarla
en un tubo ependorf. Diluir agregando 900 µl de agua destilada. Agitar en
vortex, etiquetar y congelar hasta el momento de su utilización.
Reativo de Bradford: disolver 100 mg de azul brillante de coomasie G-250 en
50 mL de etanol al 95 %. Agregar 100 mL de ácido fosforico al 85% (p/v),
diluir a un volumen final de 1 l. Agitar al menos 2 horas tapado y cubierto de
la luz. Filtrar con papel Whatman Nº1, usando plegado múltiple. Almacenar
en frasco ámbar protegido de la luz.
Agua destilada o buffer del pH que corresponda
Procedimiento: Curva de Calibración
Comprobar la concentración del estándar de proteína (BSA), leyendo a 280
nm, y usando como blanco agua destilada. El valor obtenido se divide por
0,63 y este resultado da la concentración exacta del estándar, en mg/mL.
En cada tubo agregar las cantidades de estándar de albúmina y agua
destilada de acuerdo a la tabla que se presenta a continuación. Dejar
reposar por 5 minutos y agregar el reactivo de Bradford. Luego, agitar en
vortex y esperar 10 minutos antes de leer las muestras en el
espectrofotómetro. La lectura debe realizarse a 595 nm.
Tubo Estádar BSA (µl) Agua destilada o buffer (µl) Reactivo de Bradford (µl)1 0 50 1500 2 10 40 1500 3 20 30 1500 4 30 20 1500 5 40 10 1500 6 50 0 1500

59
Dibujar la curva de calibración, con la cantidad de BSA en el eje de las
abscisas y la absorbancia, en el eje de las ordenadas.
Preparación de las Muestras
Estimar el porcentaje de proteínas aproximado que se espera en el
sobrenadante de la muestra, para caer en el rango de la curva del estándar
(entre 1 y 40 µl de proteína).
Tomar entre muestra y solvente 50 µl, agregando en tubos ependorf el
solvente que corresponda según el análisis a efectuar (agua destilada o
buffer al pH que corresponda).
Agitar y dejar reposar 5 min.
Agregar 1.500 ul de reactivo de Bradford y dejar reposar durante 10 min.
Leer las muestras en el espectrofotómetro a una longitud de onda de 595
nm, usando para ello cubetas plásticas.
Para conocer la cantidad de proteína en la muestra interpolar en la curva de
calibración, la que debe realizarse en paralelo con la medición de la
muestra.

60
ANEXO 3
Preparación de los Buffers
Buffer pH 3,0: 4,2028 g de ácido cítrico monohidratado más 0,4 g de hidróxido de
sodio y 2,34g de cloruro de sodio, disueltos en 40 mL de agua destilada. Ajustar el
pH y aforar a 100 mL.
Buffer pH 4,0: 4,2028 g de ácido cítrico monohidratado más 0,92 g de hidróxido de
sodio y 1,4 g de cloruro de sodio, disueltos en 40 mL de agua destilada. Ajustar el
pH y aforar a 100 mL.
Buffer pH 5,0: 4,2028 g de ácido cítrico monohidratado más 1,3 g de hidróxido de
sodio y 0,29 g de cloruro de sodio, disueltos en 40 mL de agua destilada. Ajustar el
pH y aforar a 100 mL.
Buffer pH 6,0: 50 mL de fosfato dihidrógeno de potasio 0,1M más 5,6 mL de
hidróxido de sodio 0,1 M y 1,3 g de cloruro de sodio.
Buffer pH 7,0: 50 mL de fosfato dihidrógeno de potasio 0,1 M más 29,1 mL de
hidróxido de sodio 0,1 M y 1,85 g de cloruro de sodio.
Buffer pH 8,0: 50 mL de fosfato dihidrógeno de potasio 0,1 M más 46,7 mL de
hidróxido de sodio 0,1 M y 1,88 g de cloruro de sodio.
Buffer pH 9,0: 50 mL de bórax 0,025 M más 4,6 mL de ácido clorhídrico 0,1 M y
2,26 g de cloruro de sodio.
Buffer pH 10,0: 50 mL de bicarbonato de sodio 0,05 M más 10,7 mL de hidróxido
de sodio 0,1 M y 1,57 mL de cloruro de sodio.
Buffer pH 11,0: 50 mL de bicarbonato de sodio 0,05 M más 22,7 mL de hidróxido
de sodio 0,1 M y 1,85 g de cloruro de sodio.
Se adicionó cloruro de sodio a los buffer para que todos tuvieran una fuerza iónica
idéntica, de 0,5.

61
Anexo 4
Gradientes de Concentración de Fase Móvil para HPLC
Perfil de Aminoácidos:
Tiempo (min)
% Acetato de Sodio(pH 6) % Acetonitrilo
0,0 92 8 3,0 88 12 6,0 86 14 22 79 21 30 69 31 40 92 8
Con un flujo de 0,9 mL/min
Determinación de Fitoestrógenos:
Tiempo(min)
% Agua % Metanol
0,0 60 40 20 25 75 21 60 40 35 60 40 Con un flujo de 0,7 mL/min