tempol reduces podocyte apoptosis via parp signaling pathway in experimental diabetes mellitus
TRANSCRIPT

Fax +41 61 306 12 34E-Mail [email protected]
Original Paper
Nephron Exp Nephrol 2012;120:e81–e90 DOI: 10.1159/000337364
Tempol Reduces Podocyte Apoptosis via PARP Signaling Pathway in Experimental Diabetes Mellitus
Elisa B.M.I. Peixoto Alexandros Papadimitriou Jacqueline M. Lopes de Faria
Jose B. Lopes de Faria
Renal Pathophysiology Laboratory, Investigation on Diabetes Complications, Division of Nephrology,Department of Internal Medicine, Faculty of Medical Sciences, State University of Campinas, Campinas , Brazil
vitro, high glucose increased caspase-3 activity and led to a higher number of apoptotic cells that were prevented by tempol and the PARP-1 inhibitor. Conclusion: In DM, tempol reduces albuminuria associated with reduction of podocyte apoptosis and decreasing oxidative stress via PARP signal-ing. Copyright © 2012 S. Karger AG, Basel
Introduction
Podocytes or glomerular visceral epithelial cells are important components of the integrity of the glomerular filtration barrier [1–3] . It has been demonstrated in pa-tients with diabetes mellitus (DM) that podocyte deple-tion is not only associated with overt diabetic nephropa-thy (DN), but it also precedes the onset of microalbumin-uria [1, 4] . Experimental studies have also shown that podocyte depletion is one of the earliest cellular lesions affecting diabetic kidney [2, 3] . The mechanisms that un-derlie the loss of podocytes in DN are under intense in-vestigation.
Several lines of evidence suggest that in DN podocyte apoptosis contributes to reduced podocyte number [2, 3, 5] . It has been suggested that increased reactive oxygen species (ROS) may be a trigger mechanism in podocyte
Key Words
Podocyte � Apoptosis � Poly(ADP-ribose) polymerase � Diabetic nephropathy � Albuminuria � Oxidative stress � Tempol
Abstract
Background/Aims: In diabetic hypertensive rats, tempol re-duces albuminuria by restoring the redox imbalance. In-creased formation of reactive oxygen species leading to ac-tivation of poly(ADP-ribose) polymerase (PARP)-1 and podo-cyte loss by apoptosis contribute to albuminuria in diabetes mellitus (DM). In the present study, we investigated the hy-pothesis that in DM tempol reduces albuminuria by inhibi-tion of PARP-induced podocyte apoptosis. Methods: DM was induced in 4-week-old spontaneously hypertensive rats by streptozotocin. Mouse and human podocyte cell lines were cultured in normal or high-glucose conditions, with or without tempol and/or a PARP-1 inhibitor, PJ34. Results: In diabetic rats, tempol treatment did not affect plasma glu-cose levels or systolic blood pressure. Albuminuria was high-er in diabetic rats, and it was reduced by tempol. DM leads to an elevation of glomerular apoptotic cells and to podo-cyte loss; both were prevented by tempol treatment. DM in-creases the expression of poly(ADP-ribose)-modified pro-teins in isolated glomeruli, and it was reduced by tempol. In
Received: November 24, 2011 Accepted: February 17, 2012 Published online: April 28, 2012
Dr. Jose B. Lopes de Faria, MD, PhD, Renal Pathophysiology LaboratoryDivision of Nephrology Department of Internal Medicine Faculty of Medical Sciences, State University of Campinas Campinas, São Paulo 13084-971 (Brazil) Tel. +55 19 3521 7499, E-Mail jblfaria @ fcm.unicamp.br
© 2012 S. Karger AG, Basel1660–2129/12/1202–0081$38.00/0
Accessible online at:www.karger.com/nee

Peixoto/Papadimitriou/Lopes de Faria/Lopes de Faria
Nephron Exp Nephrol 2012;120:e81–e90 e82
apoptosis and loss in DM [4, 6] . Upregulation of NADPH oxidases and cytochrome P450 have been implicated as major sources of ROS in the podocytes in DM [6] . In ad-dition, it has been suggested that the activation of the poly(ADP-ribose) polymerase 1 (PARP-1) is involved in hyperglycemia-induced ROS and podocyte apoptosis since PARP inhibitors blocked these later events [3] . PARP-1 inhibition also ameliorated hallmarks of DN, named albuminuria and mesangial expansion. These ob-servations lead to the suggestion that PARP inhibitors could be used as a therapeutic strategy in DN.
In a recent study, we demonstrated that administra-tion of antioxidant tempol (4-hydroxy-2,2,6,6-tetrame-thylpiperidine-1-oxyl), a superoxide dismutase (SOD) mimetic, to streptozotocin (STZ)-induced diabetic hy-pertensive rats reestablished the imbalance in redox sta-tus [7] . Tempol elevated the renal expression of renal an-tioxidant extracellular SOD, decreased the production of NADPH-dependent superoxide production, reduced markers of tissue oxidative stress, and reduced renal ex-pression of collagen IV and albuminuria. The mecha-nism by which tempol reduced albuminuria in the dia-betic rats remains unknown. The aim of the present study was to investigate whether tempol reduces albuminuria by diminishing podocyte apoptosis and PARP activity.
Methods
Experimental Animals and Protocol All experiments complied with the guidelines established by
The Brazilian College for Animal Experimentation, and were ap-proved by the Institutional Ethical Committee. Experiments were performed on male spontaneously hypertensive rat (SHR). DM was induced in 4-week-old SHR by injection of STZ, as previous-ly reported [7] . The diabetic rats either did or did not receive tem-pol treatment (Calbiochem, La Jolla, Calif., USA), as previously reported [7] . We have chosen to use SHR rats because others and we have shown that diabetic SHR display features of accelerated nephropathy and also because of the frequent association of hy-pertension with DM in human DN [8, 9] .
Blood Pressure Determination Systolic blood pressure was obtained by tail-cuff plethysmog-
raphy using an MK III physiograph (Narco Bio-System, Houston, Tex., USA) [7] .
Albumin Excretion Rate Albumin excretion rate (AER) was determined by single ra-
dial immunodiffusion [7] .
Glomeruli Isolation Glomeruli were isolated using established sieving technique.
Fragments from the outer renal cortex were pressed through
stainless steel screens (W. Tyler Co., Mentor, Ohio, USA), and iso-lated glomeruli were prepared for Western blot analysis [10] .
Immunohistochemistry Immunohistochemistry for Wilms’ tumor protein (WT-1;
Santa Cruz) was performed in 4% paraformaldehyde-fixed and paraffin-embedded kidney sections using avidin-biotin complex ABC Vectastain (Vector Laboratories) and 3,3-diaminobenzidine tetrahydrochloride (Sigma).
Double Immunofluorescence for WT-1 and Caspase-3 Kidney halves fixed in 4% paraformaldehyde and frozen in
Tissue-Tek O.C.T. compound embedding medium were incubat-ed with both primary antibodies to WT-1 and cleaved caspase-3 (Cell Signaling).
Immunofluorescence for Nephrin Kidney halves were prepared the same way as for double im-
munofluorescence staining assay. The semiquantitative analyses were performed using a Bio Color Scanner program, and the flu-orescence of nephrin (primary antibody provided by Dr. Hiroshi Kawachi, University Graduate School of Medical and Dental Sci-ences, Niigata, Japan) was expressed as percentage of fluorescence per glomerulus area, and the mean of glomeruli intensity of each rat was expressed as a percentage of mean of control rats. We as-sessed an average of 5–20 glomeruli per rat, and we analyzed at least 5 animals per group.
Western Blot Western blot assay [7] was performed to analyze poly(ADP-
ribose) (PAR) polymers expression in isolated glomeruli, incubat-ing the membranes with anti-PAR mouse monoclonal antibody (Trevigen) which detects the ribosylated proteins [11] .
Podocyte Culture Conditionally immortalized podocytes from mice (Peter
Mundel, Mount Sinai School of Medicine, N.Y., USA) and hu-mans (Moin A. Saleem, University of Bristol, Bristol, UK, and Peter Mundel) were kindly provided by Dr. Luigi Gnudi (King’s College London, UK). Podocytes were cultured in RPMI 1640 (Gibco) in the presence of 20 U/ml of interferon- � (Invitrogen) for mouse podocytes, or ITS (100 ! insulin, transferrin and sele-nium; Invitrogen) for human podocytes at 33 ° C (permissive con-dition).
Podocyte Treatments Podocytes were incubated with 7 m M and 30 m M D -glucose,
as in vitro models of normoglycemia and hyperglycemia, respec-tively. High glucose (HG) treatment was performed in the pres-ence or absence of tempol (100 n M ) and/or PJ34 (a specific inhib-itor of PARP-1 [12] ; 1 � M ; Sigma), with 12 h of incubation for mouse podocytes and 48 h for human podocytes.
Terminal Deoxynucleotidyl Transferase-Mediated Nick-End Labeling The terminal deoxynucleotidyl transferase-mediated nick-
end labeling (TUNEL) method was applied to the mouse podo-cyte culture using a DeadEnd Fluorometric TUNEL System de-tection kit (Promega, Madison, Wisc., USA), according to the manufacturer’s instructions. The quantitative analysis of the
Tempol Reduces Podocyte Apoptosis in Diabetes
Nephron Exp Nephrol 2012;120:e81–e90 e83
podocyte cells was done by counting the percentage of positive cells per total number of cells per square millimeter. The TUNEL method was also applied to cortex tissue from the studied group [10] . The quantitative analysis in renal cortex was done by count-ing glomeruli-positive cells per 100 glomeruli of cortex from the studied group.
Immunofluorescence for Caspase-3 The staining for cleaved caspase-3 was performed in mouse
podocyte. The quantitative analysis was done by counting the percentage of positive cells per total cells per square millimeter.
Caspase-3 Activity Assay Caspase-3 activity was analyzed in mouse and human immor-
talized podocytes by colorimetric assay [13] .
Statistical Analysis Comparisons between groups were done with one-way analy-
sis of variance followed by Bonferroni test. Albuminuria was ana-lyzed by nonparametric tests Kruskal-Wallis followed by Mann-Whitney U test. A value of p ! 0.05 was considered significant.
Results
Tempol Treatment Did Not Alter Blood Pressure or Glycemia Levels Diabetic rats showed impaired body weight gain and
elevated blood glucose levels compared with control rats, and treatment with tempol did not affect these parame-ters ( table 1 ). Systolic blood pressure was altered neither by diabetes nor by treatment with tempol.
DM-Induced Urinary AER Increment and Nephrin Expression Reduction Were Improved by Tempol Treatment Untreated diabetic rats exhibited an increase in uri-
nary AER, and treatment with tempol significantly re-duced AER ( fig. 1 A). Immunofluorescence ( fig. 1 B) re-
vealed a reduction in nephrin expression in diabetic glomerulus ( fig. 1 Bb, d) when compared to the control ( fig. 1 Ba), and treatment significantly increased this ex-pression ( fig. 1 Bc, d).
Tempol Treatment Prevented Podocyte Apoptosis and Depletion Induced by DM in SHR Animals By the TUNEL method, the number of glomerular
apoptotic cells in renal cortex was higher in untreated diabetic rats, and tempol reduced this number ( fig. 2 A, B). We demonstrated that apoptotic glomeruli cells were characterized as podocytes using double immunofluo-rescence for cleaved caspase-3, a marker of apoptosis, and WT-1, a marker of podocyte [14] ( fig. 2 C). Immunohisto-chemistry for WT-1 ( fig. 3 ) showed that untreated dia-betic rats exhibited a marked reduction in the podocyte number, and treatment increased this number to the lev-els seen in control rats. Therefore, altogether, we can con-clude that most glomeruli apoptotic cells in diabetic renal cortex were podocytes, and tempol treatment decreased this DM-induced podocyte apoptosis.
DM-Induced Overactivation of PARP-1 Was Prevented by Tempol Treatment In the present study, the activation of PARP-1 was ac-
cessed through the amount of PAR-modified proteins, ribosylated proteins, by Western blotting analysis in iso-lated glomeruli [11] . PARP-1 is a nuclear enzyme respon-sible for the bulk of poly(ADP-ribosyl)ation: attachment of NAD+-derived and form long branches of ADP-ri-bose polymers (PAR) to acceptor proteins such as PARP-1 itself, histones, DNA repair proteins and transcription factors, following DNA damage. This covalent protein modification alters the physicochemical properties of the modified target proteins and thus inhibits their func-tion [15] . In our study, we could observe that the expres-sion of PAR ribosylated proteins was elevated in diabetic rats; therefore, DM induced overactivation of PARP-1. In addition, treatment with tempol markedly reduced this expression and consequently PARP-1 activity ( fig. 4 ).
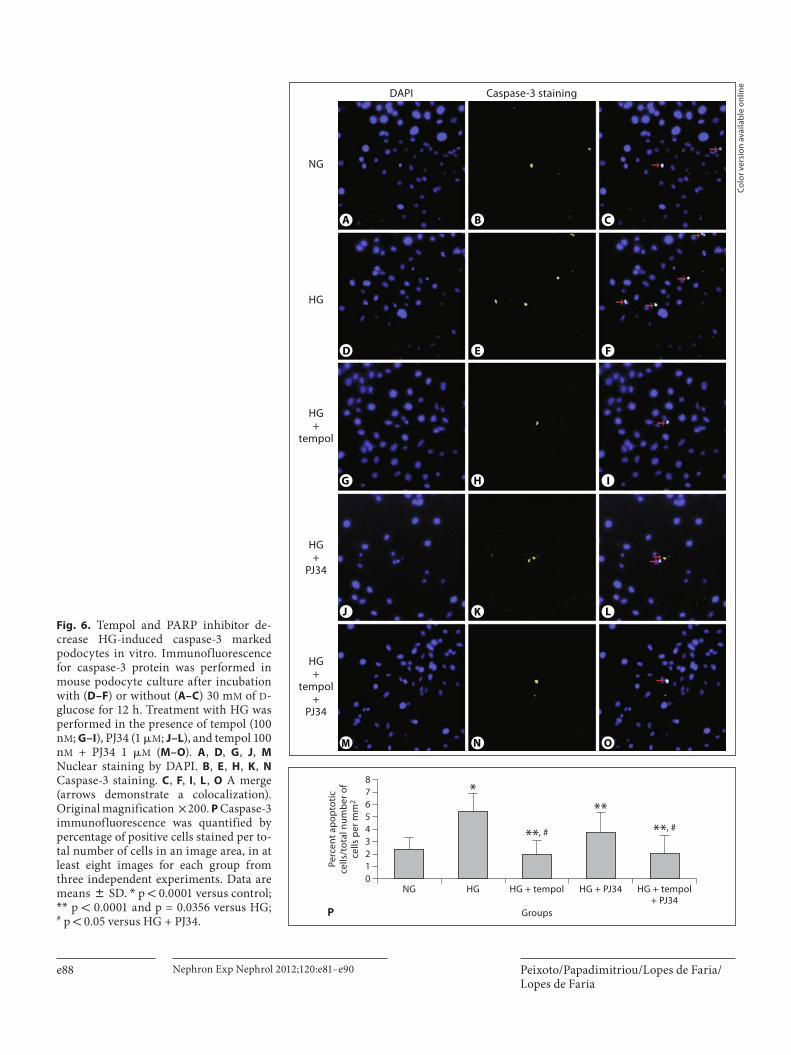
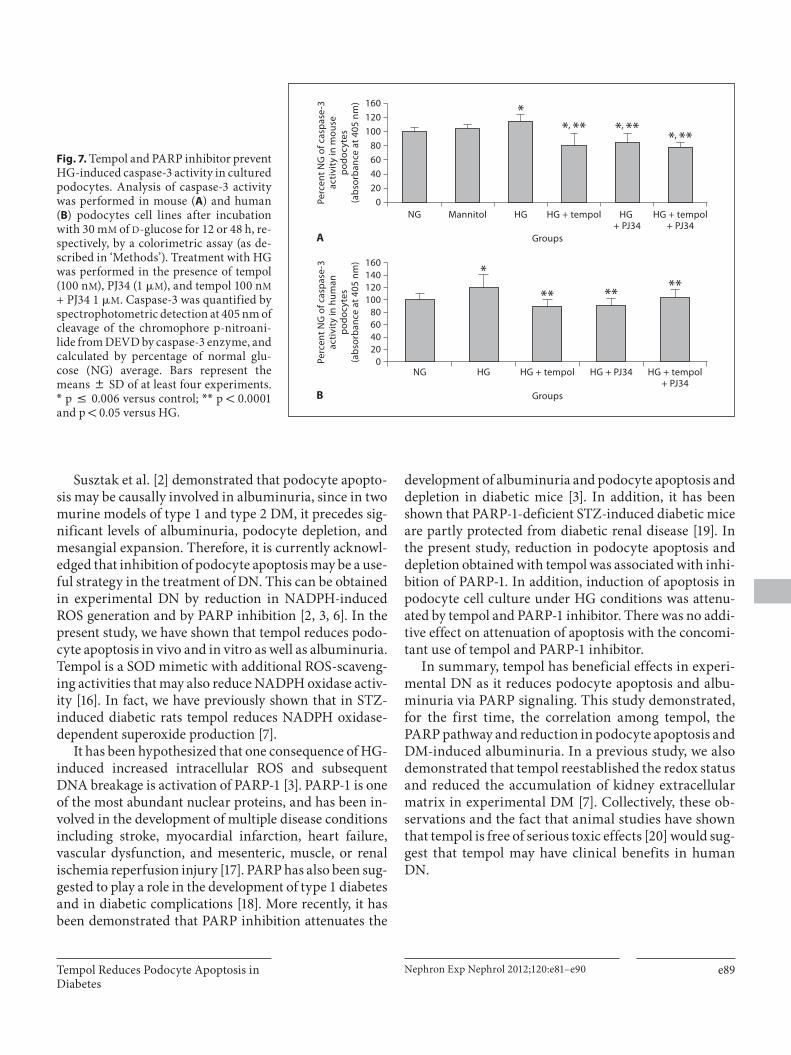
Podocyte Apoptosis Was Prevented by Tempol Treatment and PARP Inhibition in Cultures of Mouse and Human Podocytes Mouse and human podocytes were exposed to HG (30
m M ) for 12 and 48 h, respectively, with tempol and/or PJ34. The TUNEL method ( fig. 5 ) and immunofluores-cence for caspase-3 ( fig. 6 ) showed that HG markedly increased the number of apoptotic mouse podocytes ( fig. 5 D–F, P; fig. 6 D–F, P; fig. 5 A–C; fig. 6 A–C). Treat-
Table 1. P hysical and metabolic parameters in control and dia-betic rats with and without tempol treatment
Body weight, g
Systolic bloodpressuremm Hg
Plasma glucosemg/dl
SHR control (n = 15) 194815 158810 153823SHR diabetic (n = 15) 119834* 157811 547882*SHR diabetic + tempol
(n = 15) 109818* 149807 545889*
D ata were expressed as means 8 SD.* p < 0.0001 versus SHR control group.

Peixoto/Papadimitriou/Lopes de Faria/Lopes de Faria
Nephron Exp Nephrol 2012;120:e81–e90 e84
ment with tempol ( fig. 5 G–I, P; fig. 6 G–I, P) and with PJ34 ( fig. 5 J–L, P; fig. 6 J–L, P) significantly reduced the number of podocytes in apoptosis. We observed a reduc-tion in caspase-3 activity induced by HG in the culture of mouse and human podocytes treated with tempol and with PJ34 ( fig. 7 ). There was no effect of PJ34 on tempol-
induced reduction of podocyte apoptosis with concomi-tant use of tempol and PJ34 ( fig. 5 M–P; fig. 6 M–P; fig. 7 ). These findings suggest that tempol treatment prevented HG-induced podocyte apoptosis at least partly by PARP-1 inhibition.
24-h
our u
rin
ary
AER
(μg
)
SHR control0
200
400
600
800
1,000
1,200
SHR diabetic
Groups
SHR diabetic+ tempol
#
*
Fig. 1. Tempol treatment reduced AER ( A ) and im-peded diabetes-induced decrease in nephrin expres-sion ( B ) in diabetic SHR. A AER was expressed as geometric mean 8 SD. * p = 0.004 versus SHR control group; # p = 0.006 versus SHR diabetic untreated group. Expression of nephrin was estimated by im-munofluorescence assay in control ( Ba ), diabetic ( Bb ) and treated ( Bc ) SHR groups. Glomerular nephrin in-tensity was evaluated by a semiquantitative method, expressed as the percentage of fluorescence per glo-merulus area, and the mean of glomeruli intensity of each rat was expressed as a percentage of the mean of control rats ( Bd ) from at least four independent ex-periments (5 animals in each group). Original magni-fication ! 400. Bd Data are means 8 SD. * p ! 0.0001 versus SHR control group; # p = 0.0380 versus SHR diabetic untreated group.
d
a b
c
Perc
ent
con
trol
mea
n n
eph
rin
inte
nsi
ty (%
fluo
resc
ence
/g
lom
erul
us a
rea)
SHR control0
20
40
60
80
100
120
140
SHR diabetic
Groups
SHR diabetic+ tempol
#
*
A
B
Co
lor v
ersi
on
avai
lab
le o
nlin
e

Tempol Reduces Podocyte Apoptosis in Diabetes
Nephron Exp Nephrol 2012;120:e81–e90 e85
Discussion
Herein, we demonstrated that in diabetic hypertensive rats podocyte apoptosis and loss are associated with gen-eration of ROS and activation of PARP-1. These changes, which were accompanied by nephrin loss and albumin-uria, were prevented by a SOD mimetic, tempol. Attenu-
ation of ROS-induced podocyte apoptosis by tempol was confirmed in mouse and human podocyte cell culture exposed to HG, and this anti-apoptotic effect was also observed with a PARP-1 inhibitor. Collectively, these ob-servations suggest for the first time that in experimental diabetes tempol reduces podocyte apoptosis and albu-minuria through PARP signaling.
TUN
EL s
tain
ing
-pos
itiv
ece
lls/1
00 g
lom
erul
i
SHR control0
0.5
1.0
1.5
2.0
3.0
2.5
3.5
4.0
4.5
SHR diabetic
Groups
SHR diabetic+ tempol
#
*
Merge
WT-1 Caspase-3
Fig. 2. Tempol treatment blocks podocyte apoptosis in diabetic SHR. Glomeruli apoptotic cells were identi-fied by the TUNEL method (arrow, A ) and quantified as described in the ‘Methods’ ( B ). C Apoptotic cells within glomeruli were identified as podocytes by label-ing these cells for a specific podocyte marker (WT-1) and apoptosis marker (caspase-3; arrows). Data are means 8 SD. Original magnification ! 400. * p = 0.0014 versus SHR control group; # p = 0.01 versus SHR diabetic untreated group.
Co
lor v
ersi
on
avai
lab
le o
nlin
e
A B
C

Peixoto/Papadimitriou/Lopes de Faria/Lopes de Faria
Nephron Exp Nephrol 2012;120:e81–e90 e86
WT-
1-p
osit
ive
cells
/glo
mer
ulus
hig
h-p
ower
fiel
d
SHR control0
5
10
15
20
25
SHR diabetic
GroupsD
SHR diabetic+ tempol
#
*
A B
C
Co
lor v
ersi
on
avai
lab
le o
nlin
e
Fig. 3. Tempol treatment blocks podocyte depletion in diabetic SHR. Positive immunostaining for WT-1, a spe-cific podocyte marker, was determined by brown-colored nuclei in control ( A ), diabetic ( B ), and tempol-treated diabetic ( C ) SHR (arrows). Original magnification ! 1,000. D The quantification of WT-1 expression was as-sessed in four independent experiments. Data are means 8 SD. * p = 0.014 versus SHR control group; # p = 0.036 versus SHR diabetic untreated group.
A
Control DM
PAR-modifiedproteins
Actin
Treated
PAR-
mod
ified
pro
tein
s/ac
tin
(arb
itra
ry d
ensi
tom
etri
c un
its)
SHR control0
2
4
3
1
5
6
7
8
9
SHR diabetic
GroupsB
SHR diabetic+ tempol
#
*
Fig. 4. Tempol blocks the diabetes-induced elevation in PAR-mod-ified protein expression in renal cortex. A Representative Western blot analysis of PAR polymer expression in control, diabetic and tempol-treated diabetic rats. B Densitometric analysis of PAR-modified protein/actin expression in different groups. The smear
of bands higher than 130 kDa represents several PAR ribosylated proteins. Actin was used as a control of protein loading. Bars rep-resent means 8 SD of band densities of two independent experi-ments (mean of 5 rats in each group). * p = 0.0379 versus SHR con-trol group; # p = 0.0262 versus SHR diabetic untreated group.

Tempol Reduces Podocyte Apoptosis in Diabetes
Nephron Exp Nephrol 2012;120:e81–e90 e87
DAPI
NG
HG
HG+
tempol
HG+
tempol+
PJ34
HG+
PJ34
TUNEL staining
A B C
D E F
G H I
J K L
M N O
Perc
ent
apop
toti
cce
lls/t
otal
num
ber
of
cells
per
mm
2
NG0
2
6
4
8
10
12
HG
GroupsP
HG + tempol
*
HG + PJ34 HG + tempol+ PJ34
**
****
Fig. 5. Tempol and PARP inhibitor prevent HG-induced podocyte apoptosis in vitro by TUNEL analysis. The TUNEL method was performed in mouse podocyte culture after incubation with ( D–F ) or without( A–C ) 30 m M of D -glucose for 12 h. The treatment with HG was performed in the presence of tempol (100 n M ; G–I ), PJ34, a PARP-1 inhibitor (1 � M ; J–L ), and tempol 100 n M + PJ34 1 � M ( M–O ). A , D , G , J , M Nuclear staining by DAPI. B , E , H , K , N TUNEL staining. C , F , I , L , O A merge(arrows demonstrate the colocalization). Original magnification ! 200. P TUNEL assay was quantified by percentage of pos-itive cells stained per total number of cells in an image area, in at least 16 images for each group from two independent experi-ments. Data are means 8 SD. * p = 0.0002 versus control; * * p ! 0.0001 and p = 0.0005 versus HG. NG = Normal glucose.
Co
lor v
ersi
on
avai
lab
le o
nlin
e
Peixoto/Papadimitriou/Lopes de Faria/Lopes de Faria
Nephron Exp Nephrol 2012;120:e81–e90 e88
Perc
ent
apop
toti
cce
lls/t
otal
num
ber
of
cells
per
mm
2
NG0
2
43
1
5678
HG
GroupsP
HG + tempol
*
HG + PJ34 HG + tempol+ PJ34
**, #**
, #**
DAPI
NG
HG
HG+
tempol
HG+
tempol+
PJ34
HG+
PJ34
Caspase-3 staining
A B C
D E F
G H I
J K L
M N O
Fig. 6. Tempol and PARP inhibitor de-crease HG-induced caspase-3 marked podocytes in vitro. Immunofluorescence for caspase-3 protein was performed in mouse podocyte culture after incu bation with ( D–F ) or without ( A–C ) 30 m M of D -glucose for 12 h. Treatment with HG was performed in the presence of tempol (100 n M ; G–I ), PJ34 (1 � M ; J–L ), and tempol 100 n M + PJ34 1 � M ( M–O ). A , D , G , J , M Nuclear staining by DAPI. B , E , H , K , N Caspase-3 staining. C , F , I , L , O A merge (arrows demonstrate a colocalization). Original magnification ! 200. P Caspase-3 immunofluorescence was quantified by percentage of positive cells stained per to-tal number of cells in an image area, in at least eight images for each group from three independent experiments. Data are means 8 SD. * p ! 0.0001 versus control; * * p ! 0.0001 and p = 0.0356 versus HG; # p ! 0.05 versus HG + PJ34.
Co
lor v
ersi
on
avai
lab
le o
nlin
e
Tempol Reduces Podocyte Apoptosis in Diabetes
Nephron Exp Nephrol 2012;120:e81–e90 e89
Susztak et al. [2] demonstrated that podocyte apopto-sis may be causally involved in albuminuria, since in two murine models of type 1 and type 2 DM, it precedes sig-nificant levels of albuminuria, podocyte depletion, and mesangial expansion. Therefore, it is currently acknowl-edged that inhibition of podocyte apoptosis may be a use-ful strategy in the treatment of DN. This can be obtained in experimental DN by reduction in NADPH-induced ROS generation and by PARP inhibition [2, 3, 6] . In the present study, we have shown that tempol reduces podo-cyte apoptosis in vivo and in vitro as well as albuminuria. Tempol is a SOD mimetic with additional ROS-scaveng-ing activities that may also reduce NADPH oxidase activ-ity [16] . In fact, we have previously shown that in STZ-induced diabetic rats tempol reduces NADPH oxidase-dependent superoxide production [7] .
It has been hypothesized that one consequence of HG-induced increased intracellular ROS and subsequent DNA breakage is activation of PARP-1 [3] . PARP-1 is one of the most abundant nuclear proteins, and has been in-volved in the development of multiple disease conditions including stroke, myocardial infarction, heart failure, vascular dysfunction, and mesenteric, muscle, or renal ischemia reperfusion injury [17] . PARP has also been sug-gested to play a role in the development of type 1 diabetes and in diabetic complications [18] . More recently, it has been demonstrated that PARP inhibition attenuates the
development of albuminuria and podocyte apoptosis and depletion in diabetic mice [3] . In addition, it has been shown that PARP-1-deficient STZ-induced diabetic mice are partly protected from diabetic renal disease [19] . In the present study, reduction in podocyte apoptosis and depletion obtained with tempol was associated with inhi-bition of PARP-1. In addition, induction of apoptosis in podocyte cell culture under HG conditions was attenu-ated by tempol and PARP-1 inhibitor. There was no addi-tive effect on attenuation of apoptosis with the concomi-tant use of tempol and PARP-1 inhibitor.
In summary, tempol has beneficial effects in experi-mental DN as it reduces podocyte apoptosis and albu-minuria via PARP signaling. This study demonstrated, for the first time, the correlation among tempol, the PARP pathway and reduction in podocyte apoptosis and DM-induced albuminuria. In a previous study, we also demonstrated that tempol reestablished the redox status and reduced the accumulation of kidney extracellular matrix in experimental DM [7] . Collectively, these ob-servations and the fact that animal studies have shown that tempol is free of serious toxic effects [20] would sug-gest that tempol may have clinical benefits in human DN.
Perc
ent
NG
of c
asp
ase-
3ac
tivi
ty in
hum
anp
odoc
ytes
(ab
sorb
ance
at
405
nm
)
NG0
40
8060
20
100120140160
HG
GroupsB
HG + tempol
*
HG + PJ34 HG + tempol+ PJ34
** ****
Perc
ent
NG
of c
asp
ase-
3ac
tivi
ty in
mou
sep
odoc
ytes
(ab
sorb
ance
at
405
nm
)
NG0
40
80
60
20
100
120
160
HGMannitol
GroupsA
HG + tempol
*
HG + PJ34
HG + tempol+ PJ34
,* **,* **,* **
Fig. 7. Tempol and PARP inhibitor prevent HG-induced caspase-3 activity in cultured podocytes. Analysis of caspase-3 activity was performed in mouse ( A ) and human ( B ) podocytes cell lines after incubation with 30 m M of D -glucose for 12 or 48 h, re-spectively, by a colorimetric assay (as de-scribed in ‘Methods’). Treatment with HG was performed in the presence of tempol (100 n M ), PJ34 (1 � M ), and tempol 100 n M + PJ34 1 � M . Caspase-3 was quantified by spectrophotometric detection at 405 nm of cleavage of the chromophore p-nitroani-lide from DEVD by caspase-3 enzyme, and calculated by percentage of normal glu-cose (NG) average. Bars represent the means 8 SD of at least four experiments. * p ̂ 0.006 versus control; * * p ! 0.0001 and p ! 0.05 versus HG.

Peixoto/Papadimitriou/Lopes de Faria/Lopes de Faria
Nephron Exp Nephrol 2012;120:e81–e90 e90
References
1 Wolf G, Chen S, Ziyadeh FN: From the pe-riphery of glomerular capillary wall toward the center of disease: podocyte injury comes of age in diabetic nephropathy. Diabetes 2005; 54: 1626–1634.
2 Susztak K, Raff AC, Schiffer M, Böttinger EP: Glucose-induced reactive oxygen species cause apoptosis of podocytes and podocyte depletion at the onset of diabetic nephropa-thy. Diabetes 2006; 55: 225–233.
3 Szabó C, Biser A, Benko R, Böttinger E, Susz-tak K: Poly(ADP-ribose) polymerase inhibi-tors ameliorate nephropathy of type 2 dia-betic Leprdb/db mice. Diabetes 2006; 55: 3004–3012.
4 Meyer TW, Bennett PH, Nelson RG: Podo-cyte number predicts long-term urinary al-bumin excretion in Pima Indians with type II diabetes and microalbuminuria. Diabeto-logia 1999; 42: 1341–1344.
5 Schiffer M, Bitzer M, Roberts IS, Kopp JB, ten Dijke P, Mundel P, Böttinger EP: Apop-tosis in podocytes induced by TGF-beta and Smad7. Clin Invest 2001; 10: 807–816.
6 Eid AA, Gorin EY, Fagg BM, Maalouf R, Barnes JL, Block K, Abboud HE: Mechanism of podocyte injury in diabetes: role of cyto-chrome P450 and NADPH oxidases. Diabe-tes 2009; 58: 1201–1211.
7 Peixoto EBMI, Pessoa BS, Biswas SK, Lopes de Faria JB: Antioxidant SOD mimetic pre-vents NADPH oxidase-induced oxidative stress and renal damage in the early stage of experimental diabetes and hypertension. Am J Nephrol 2009; 29: 309–318.
8 Cooper ME, Allen TJ, O’Brien RC, Macmil-lan PA, Clarke B, Jerums G, Doyle AE: Ef-fects of genetic hypertension on diabetic ne-phropathy in the rat – functional and struc-tural characteristics. J Hypertens 1988; 6: 1009–1016.
9 Lopes de Faria JB, Silva KC, Lopes de Faria JM: The contribution of hypertension to dia-betic nephropathy and retinopathy: the role of inflammation and oxidative stress. Hy-pertens Res 2011; 34: 413–422.
10 Silveira LA, Bacchi CE, Pinto GA, Lopes de Faria JB: The genetics of hypertension mod-ifies the renal cell replication response in-duced by experimental diabetes. Diabetes 2002; 51: 1529–1534.
11 Rosales MA, Silva KC, Lopes de Faria JB, Lopes de Faria JM: Exogenous SOD mimetic tempol ameliorates the early retinal changes reestablishing the redox status in diabetic hypertensive rats. Invest Ophthalmol Vis Sci 2010; 51: 4327–4336.
12 Yam-Canul P, Chirino YI, Sánchez-González DJ, Martinez-Martínez CM, Cruz C, Pedra-za-Chaverri J: PJ34, a poly adenosine diphos-phate-ribose polymerase inhibitor, attenu-ates chromate-induced nephrotoxicity. Basic Clin Pharmacol Toxicol 2008; 102: 483–488.
13 Papadimitriou A, King AJ, Jones PM, Per-saud SJ: Anti-apoptotic effects of arachidon-ic acid and prostaglandin E2 in pancreatic beta-cells. Cell Physiol Biochem 2007; 20: 607–616.
14 Menini S, Iacobini C, Oddi G, Ricci C, Simo-nelli P, Fallucca S, Grattarola M, Pugliese F, Pesce C, Pugliese G: Increased glomerular cell (podocyte) apoptosis in rats with strep-tozotocin-induced diabetes mellitus: role in the development of diabetic glomerular dis-ease. Diabetologia 2007; 50: 2591–2599.
15 Erdélyi K, Bakondi E, Gergely P, Szabó C, Virág L: Pathophysiologic role of oxidative stress-induced poly(ADP-ribose)polymer-ase-1 activation: focus on cell death and transcriptional regulation. Cell Mol Life Sci 2005; 62: 751–759.
16 Wilcox CS, Pearlman A: Chemistry and an-tihypertensive effects of tempol and other nitroxides. Pharmacol Rev 2008; 60: 418–469.
17 Pacher P, Szabo O: Role of the peroxyni-trite-poly(ADP-ribose) polymerase path-way in human disease. Am J Pathol 2008; 173: 2–13.
18 Szabó C, Ischiropoulos H, Radi R: Peroxyni-trite: biochemistry, pathophysiology and de-velopment of therapeutics. Nat Rev Drug Discov 2007; 6: 662–680.
19 Shevalye H, Maksimchyk Y, Watcho P, Ob-rosova IG: Poly(ADP-ribose) polymerase-1 (PARP-1) gene deficiency alleviates diabetic kidney disease. Biochim Biophys Acta 2010; 1802: 1020–1027.
20 Burney BO, Kalaitzidis RG, Bakris GL: Nov-el therapies of diabetic nephropathy. Curr Opin Nephrol Hypertens 2009; 18: 107–111.
Acknowledgments
This work was supported by grants from Fundação de Amparo a Pesquisa do Estado de São Paulo (grant number 07/51137-5), Con-selho Nacional de Desenvolvimento Científico e Tecnológico, and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior.
We thank Rosana Aparecida Trevisan Pereira for her technical support, and Dr. Cecile Dessapt and Dr. Anthea Hayward (King’s College London, UK) for their technical assistance with podocyte cell culture.
Disclosure Statement
There are no potential conflicts of interest relevant to this ar-ticle.