sympathetic innervation of the cornea from the superior cervical ganglion. an hrp study in the cat
TRANSCRIPT

Journal of the Autonomic Nem~ousSystem, 20 (1987) 179-183 179 Elsevier
JAN00745
Short Communicat ions
Sympathetic innervation of the cornea from the superior cervical ganglion. An HRP study in the cat
Charles Morgan i Wil l iam C. D e G r o a t 2 and Peter J. J anne t t a 2
I Department of Anatomy, UCSF School of Medicine, San Francisco, CA 94143 (L(S.A.)," and ' Departments of Pharmacology and Neurosurgerv and Center for Neuroscience, Unit,ersitv of Pittsburgh, Pittsburgh, PA 15261 (U.S.A.)
(Received 20 February 1987) (Revised version received 20 May 1987)
(Accepted 26 May 1987)
Key words: Cornea; Superior cervical ganglion; Horseradish peroxidase (HRP); Sympathetic innervation
Summary
Horseradish peroxidase (HRP) was applied to the cornea in cats using a variety of methods. Small numbers of labeled neurons were observed in the superior cervical ganglion (SCG), the majority were located in the rostral half. This confirms previous histofluorescence data which indicated the SCG was the likely source of adrenergic fibers to the cornea.
Catecholamines have been identified in the cornea in a variety of species using histofluores- cence techniques and it has been suggested that the superior cervical ganglion (SCG) was their source. Adrenergic fibers were present in the cornea and other tissues of the eye of the rabbit [3,6] and have also been demonstrated in small myelinated and unmyelinated axons in the rat cornea with the electron microscope [12]. The SCG origin of these fibers is indicated indirectly in other studies. For example, using formalde- hyde-induced fluorescence, Tervo and Palkama [11] and Tervo [10] identified adrenergic nerves in the rat cornea which increased after application of a monoamine oxidase inhibitor, were eliminated following administration of reserpine, sympathetic neurectomy, or ophthalmic nerve neurectomy, but
Correspondence: C. Morgan, Department of Anatomy, Eastern Virginia Medical School, Box 1980, Norfolk, VA 23501, U.S.A.
were unaffected by ciliary ganglionectomy. As the trigeminal ganglion is considered entirely sensory, it was reasonable to conclude that the SCG was the source of these adrenergic fibers. Zander and Weddell [13] also reported the presence of a few axons remaining in the cornea of a monkey fol- lowing the destruction of the trigeminal ganglion. Again the SCG is a good candidate for the source of these axons. The present study was carried out to determine if these conclusions were justified and could be verified in the cat and also to determine the type of organization these neurons might have in the SCG. These results have been presented in abstract form [5].
Adult and juvenile cats of either sex were anesthetized with Nembutal, 30 mg/kg i.p. or with a mixture of halothane and oxygen adminis- tered through a face mask. Horseradish peroxidase (HRP) was applied to the corneas in the following ways: in most cats (Group l) a 25% solution of HRP (Sigma Type VI) dissolved in 2% dimethyl-

180
sulfoxide (DMSO) was injected with the aid of a surgical microscope into the substantia propria of the cornea in approximately 2 /,1 quantities using a 30 gauge needle on a Hamilton syringe. As the needle was withdrawn a Q-tip cotton sponge pre- vented any leakage to surrounding tissue and the defect sealed in a few minutes. A 1% solution of xylocaine was applied topically to the cornea be- fore the cats recovered from the anesthesia 1- 2 h later. In other cats (Group II), in order to test for the effects of spillage of HRP onto tissue sur- rounding the cornea, 2-5 ~1 of HRP was dropped on the cornea and allowed to spread freely to surrounding tissue. In some cases this control test was applied to the cornea opposite an injected cornea. In Group III, the cats were treated the same as those in Groups I and II except that the HRP was dissolved in water containing a small amount of lysolecithin. In one cat (Group IV), the cornea of one eye was scored superficially with a no. 11 knife blade, and 5 t~l of HRP applied under a contact lens. The opposite cornea received the same amount of HRP under a contact lens but was undamaged. In Group V as a control to test
for possible ~pread of HRP to the internal struc- tures of the eye, the ciliary ganglia were removed and examined in 3 cats that were also part of Group 1.
The cats were allowed to recover and ~urv~ve for 48 h after which they were deeply anaesthe- tized and perfused with isotontc saline and then a fixative of 1% glutaraldehyde, 1% paraformalde- hyde. The superior cervical ganglia in all cats and the ciliary ganglia in 3 cats were removed, im- mersed in buffered saline and 30% sucrose, cut into 42-56 /~m sections on a cryostat, and processed with tetramethylbenzidine [8]. Tissue sections were examined with a Leitz Dialux micro- scope using polarized darkfield illumination
Labeled cells were counted m both the rostral and caudal halves of the SCG. Relatively small numbers of cells were labeled m the SCG after injection of HRP into the cornea and after appli- cation of HRP to the damaged cornea. As shown in Table 1. in 13 ganglia in Group I an average of 122 cells per ganglion were labeled, the range being 18-450 cells. While mosl of the neurons (81%) were located in the rostral halves of the
TABLE 1
Summa~ of experiments
Number of Avg. no. cells Cells in Cells m experiments per expt. rostral end caudal end
Group I (injected cornea) 13 122 81% 19~
range 18-450 range 63-95% range 5- 37%
Group I1 (topical application) 9 1
range 0-2 NA N~,
Group I I I * (lysolecithin) Injection 2 (234, 116) * * 81% 19':7c Topical 2 (88, 120) * * 66% 34%
Group IV (contact lens) Scratched 1 (449) * * 83% t7% Undamaged 1 (159) * * 779~ 23%
Group V * (injected cornea) Ciliary ganglion 3 123 81 ~ 1 ~- ~ examined range 18-234 range 75-86% range 17-25%
* Included in Group I, * * totals, not average. NA, not available.
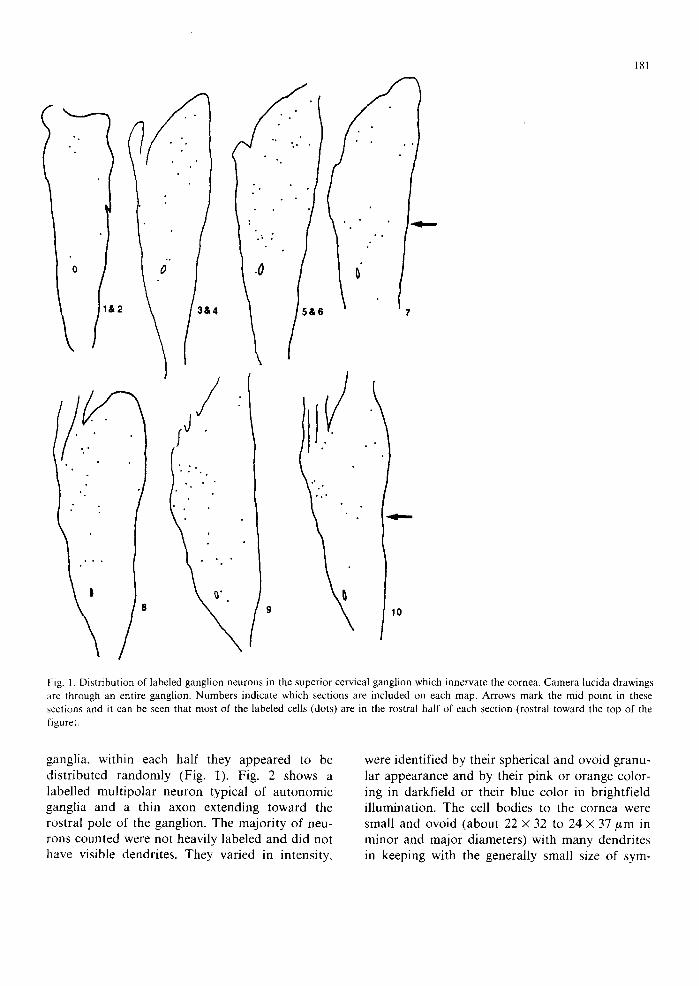
181
° °
.0
2
° ° ° .
°°
9 10
Fig. 1. Distribution of labeled ganglion neurons in the superior cervical ganglion which innervate the cornea. Camera lucida drawings are through an entire ganglion. Numbers indicate which sections are included on each map. Arrows mark the mid point in these sections and it can be seen that most of the labeled cells (dots) are in the rostral half of each section (rostral toward the top of the figure).
ganglia, within each half they appeared to be distributed randomly (Fig. 1). Fig. 2 shows a labelled multipolar neuron typical of autonomic ganglia and a thin axon extending toward the rostral pole of the ganglion. The majority of neu- rons counted were not heavily labeled and did not have visible dendrites. They varied in intensity,
were identified by their spherical and ovoid granu- lar appearance and by their pink or orange color- ing in darkfield or their blue color in brightfield illumination. The cell bodies to the cornea were small and ovoid (about 22 × 32 to 24 x 37 ~m in minor and major diameters) with many dendrites in keeping with the generally small size of sym-
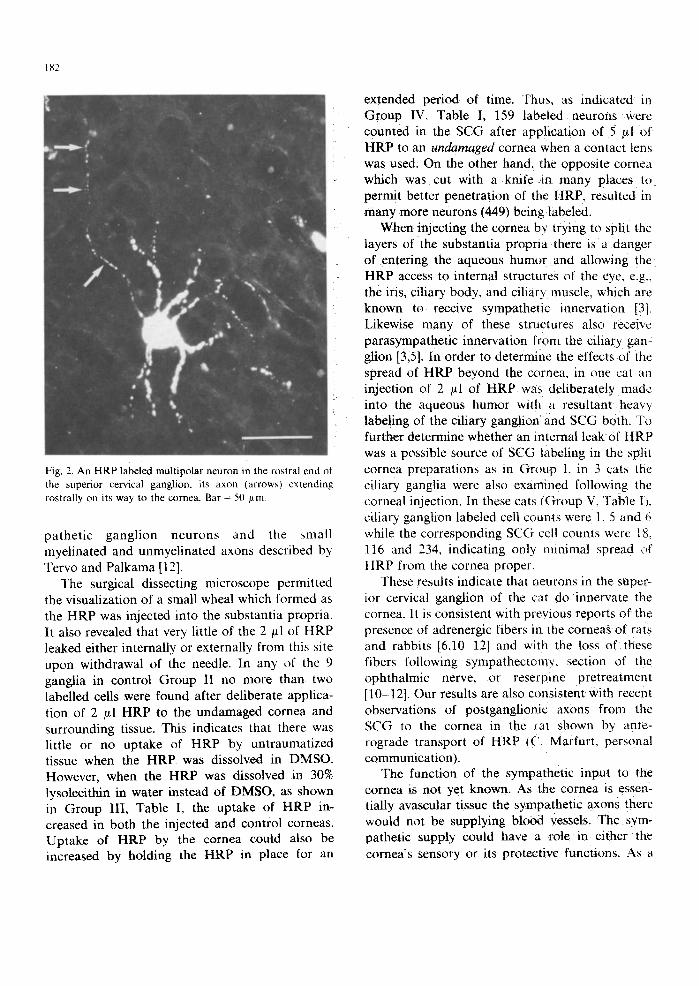
182
Fig. 2. An HRP labeled multipolar neuron in the rostral end of the superior cervical ganglion, its axon qarrows~ extending rostrally on its way to the cornea. Bar = 50 ~tm
pathetic ganglion neurons and the ~mall myelinated and unmyelinated axons described by Tervo and Palkama [12].
The surgical dissecting microscope permitted the visualization of a small wheal which formed as the HRP was injected into the substantia propria. It also revealed that very little of the 2 ~tl of HRP leaked either internally or externally from this site upon withdrawal of the needle. In any of the 9 ganglia in control Group II no more than two labelled cells were found after deliberate applica- tion of 2 /zl HRP to the undamaged cornea and surrounding tissue. This indicates that there was little or no uptake of HRP by untraumatized tissue when the HRP was dissolved in DMSO. However, when the HRP was dissolved in 30% lysolecithin in water instead of DMSO. as shown in Group III, Table I, the uptake of HRP in- creased in both the injected and control corneas. Uptake of HRP by the cornea could also be increased by holding the HRP in place for an
extended period of time. Thus. as indicated in Group IV. Table I, t59 labeled neurons were counted in the SCG after applicauon of 5 /~1 of HRP to an undamaged cornea when a contact lens was used. On the other hand. the opposite cornea which was cut with a knife in many places to permit better penetration of the HRP. resulted in .many more neurons (449) being labeled.
When injecting the cornea by trying to split the layers of the substantia propna there is a danger of entering the aqueous humor and allowing the HRP access to internal structures of the eye, e.g,, the iris. ciliary body, and ciliary muscle, which are known to receive sympathetic innervation [3]. Likewise many of these structures also receive parasympathetic innervation from the ciliary gan- glion [3,5]. In order to determine the effects of the spread of HRP beyond the cornea, in one cal an injection of 2 ,td of HRP was deliberately made into the aqueous humor with ~ resultant heavy labeling of the ciliary ganglion and SCG both. To further determine whether an internal leak of HRP was a possible source of SCG labeling in the split cornea preparations as in Group I. in 3 cats the ciliary ganglia were also examined following the corneal injection. In these cats (Group V. '-Fable I t. ciliary ganglion labeled cell counts were I. 5 and 6 while the corresponding SCG cell counts were 1 & 116 and 234. indicating only minimal spread t,f HRP from the cornea proper.
These results indicate that neurons in the super- ior cervical ganglion of the cat do innervate the cornea. It is consistent with previous reports of the presence of adrenergic fibers in the corneas of rats and rabbits [6,10 12] and with the loss of these fibers following sympathect(~m>, secuon of the ophthalmic nerve, or reserpine pretreatment [10-12]. Our results are also consistent with recent observations of postganglionic axons from the SCG to the cornea in the ~-at shown by ante- rograde transport of HRP ((" Marfurt. personal communication ).
The function of the sympathetic input to the cornea is not yet known. As the cornea is essen- tially avascular tissue the sympathetic axons there would not be supplying blood vessels. The sym- pathetic supply could have a role in either the cornea's sensory or ~ts protective functions. As a

sensory structure the cornea receives primary afferent axons from the trigeminal ganglion and it is possible that the sympathetic axons from the SCG might have some modulatory role toward these sensory axons. This could be a control over the sensitivity level of these axons by influencing the release of substance P or leucine enkephalin which are known to reach the cornea in trigeminal primary afferents [2,7].
With regard to a protective function, the cornea itself provides a clear and tough shield for the delicate tissues of the eye. It is possible that trophic actions of the sensory nerves from the trigeminal ganglion interact with the sympathetic nerves to repair the cornea following minor in- juries. A number of studies now suggest such a relationship. For example, it has been reported that in some systems up to 90% of substance P produced in a sensory neuron is delivered to the peripheral axon and can be released from that axon [1,9]. Substance P is thought to be released during injury and to be partly responsible for the flare of the wheal and flare formation when skin is scratched [4]. Perhaps then the sympathetic input controls the release of substance P which in turn is involved in the healing response of the cornea to minor scratches or abrasions.
The authors would like to thank Mary Houston and David Blaise for their technical assistance. This work has been supported by grants from the National Institutes of Health (1F32 NS 07057-01; AM 37241; AM 317888), the National Science Foundation (BNS-8507113) and the Western Pennsylvania Heart Association.
183
1 Brimijoin, S., Lundberg, J.M., Brodin, E., H~kfelt, T. and Nilsson, G., Axonal transport of substance P in the vagus and sciatic nerves of the guinea pig, Brain Res., 191 (1980) 443 457.
2 deGroat, W.C., Lowe, I.P., Kawatani, M., Morgan, C.W., Kuo, D., Roppolo, J.R. and Nagel, J., Identification of
enkephalin immunoreactivity in sensory ganglion cells, J. Auton. Nerv. Syst., Suppl. (1986) $361 368.
3 Ehinger, B., Ocular and orbital vegetative nerves, Acta Physiol. Scand., 67, Suppl. (1966) 3-35.
4 Foreman, J. and Jordan, C., Histamine release and vascular changes induced by neuropeptides, Agents Actions, 13 (1983) 105.
5 Frioni, C., Morgan, C., Nadelhaft, I., deGroat, W.C. and Jannetta, P.J., Identification of neurons in the superior
ganglion innervating the cornea of the cat, Neurosci. Abstr., 8 (1982) 553.
6 Laties, A. and Jacobowitz, D., A histochemical study of the
adrenergic and cholinergic innervation of the anterior seg- ment of the eye, Invest. Ophthalmol., 3 (1964) 592-600.
7 Morgan, C., deGroat, W.C. and Jannetta, P.J., Leucine enkephalin and substance-P identified in trigeminal gangl- ion neurons innervating the cornea in the cat, Neurosci. Abstr., 8 (1982) 474.
8 Morgan, C,, deGroat, W.C. and Nadelhaft, l., The spinal distribution of sympathetic preganglionic and visceral primary afferent neurons which send axons into the hypo- gastric nerves of the cat, J. Comp. Neurol., 243 (1986) 23-40.
9 0 l g a r t , L., Gazelius, B., Brodin, E. and Nilsson, G., Release
of substance P-like immunoreactivity from the dental pulp, Acta Physiol. Scand., 101 (1977) 510-512.
10 Tervo, T., Consecutive demonstration of nerves containing catecholamine and acetylcholinesterase in the rat cornea, Histochemistry, 50 (1977) 291-299.
11 Tervo, T. and Palkama, A., Ultrastructure of the corneal
nerves after fixation with potassium permanganate, Anat. Rec., 190 (1978) 851 860.
12 Tervo, T. and Palkama, A., Sympathetic nerves to the rat cornea, Acta Ophthalmol., 54 (1976) 75-84.
13 Zander, E. and Weddell, G., Reaction of corneal nerves to injury, Brit. J. Ophthalol., 35 (1951) 6188.