sveuČiliŠte u zagrebu - digre.pmf.unizg.hrdigre.pmf.unizg.hr/4604/1/marija mulanovic...
TRANSCRIPT

Sveučilište u Zagrebu
Prirodoslovno – matematički fakultet
Biološki odsjek
Marija Mulanović
Učestalost rekombinacija između gena
glavnog sustava tkivne podudarnosti u čovjeka
Diplomski rad
Zagreb, 2015

Ovaj rad izrađen je u Kliničkoj jedinici za tipizaciju tkiva Kliničkog zavoda za
transfuzijsku medicinu i transplantacijsku biologiju, Kliničkog bolničkog centra
Zagreb, pod vodstvom prof.dr.sc. Zorane Grubić. Rad je predan na ocjenu Biološkom
odsjeku Prirodoslovno-matematičkog fakulteta Sveučilišta u Zagrebu radi stjecanja
naziva diplomirani inženjer biologije, smjer molekularna biologija.

ZAHVALE
Zahvaljujem svojoj mentorici prof.dr.sc. Zorani Grubić koja mi je svojim znanstvenim i
stručnim savjetima pomogla u izradi ovog diplomskog rada. Iskreno joj zahvaljujem
na susretljivosti, strpljenju i poklonjenom vremenu. Zahvaljujem i svim ostalim
djelatnicima Kliničke jedinice za tipizaciju tkiva na razumijevanju, te na ugodnoj i
poticajnoj radnoj atmosferi.
Posebno se zahvaljujem svom suprugu, roditeljima i prijateljicama na bezuvjetnoj
ljubavi i nesebičnoj podršci.
Ovaj rad posvećujem pok. mr.sc. Marici Kodrić-Šmit, nekadašnjoj Voditeljici Službe
za zdravstvenu ekologiju Zavoda za javno zdravstvo Sisačko-moslavačke županije.

TEMELJNA DOKUMENTACIJSKA KARTICA
Sveučilište u Zagrebu Prirodoslovno-matematički fakultet Biološki odsjek
Diplomski rad
Učestalost rekombinacija između gena
glavnog sustava tkivne podudarnosti u čovjeka
Marija Mulanović
Rooseveltov trg 6, 10000 Zagreb, Hrvatska
U radu je analizirana učestalost rekombinacija između lokusa HLA-A i -B, te HLA-B i -DRB1 unutar skupine od 2049 obitelji iz registra obitelji bolesnika mogućih primatelja krvotvornih matičnih stanica. Svi ispitanici testirani su za antigene/gene HLA-A, -B, -DR jednom od metoda: testom mikrolimfocitotoksičnosti ili metodom lančane reakcije polimerazom i početnicama specifičnim za pojedini alel ili skupinu alela (metode: PCR-SSP i PCR-SSO). Krosingover je utvrđen kod 3,61% obitelji, od toga u 3,47% obitelji krosingover je bio prisutan samo kod jednog djeteta, a u 0,15% obitelji utvrđeno je prisustvo krosingovera kod dvoje djece. Učestalost rekombinacija između lokusa HLA-A i -B iznosila je 0,61%, a između lokusa HLA-B i -DRB1 iznosila je 0,77%. Učestalost krosingovera u oogenezi bila je 1,8 puta veća od učestalosti krosingovera u spermatogenezi. Haplotipovi kod kojih je krosingover bio prisutan između lokusa HLA-A i -B najčešće su bili pozitivni za antigen/gen HLA-B27. Haplotipovi s krosingoverom između lokusa HLA-B i -DRB1 najčešće su na lokusu HLA-DRB1 imali antigen/gen HLA-DR13. Rezultati ovog istraživanja u skladu su s rezultatima istraživanja provedenim u drugim populacijama u svijetu. (49 stranica, 19 slika, 6 tablica, 48 literaturnih navoda, jezik izvornika: hrvatski) Rad je pohranjen u Središnjoj biološkoj knjižnici Ključne riječi: geni HLA, rekombinacija, krosingover Voditelj: Dr.sc. Zorana Grubić, izv.prof., KBC Zagreb Suvoditelj: Dr.sc. Ana Galov, doc., PMF Sveučilišta u Zagrebu Ocjenitelji: Dr.sc. Mirjana Kalafatić, prof., PMF Sveučilišta u Zagrebu Dr.sc. Božena Mitić, prof., PMF Sveučilišta u Zagrebu Zamjena: Dr.sc. Zlatko Liber, izv.prof., PMF Sveučilišta u Zagrebu Rad prihvaćen: 02.12.2015.

BASIC DOCUMENTATION CARD
University of Zagreb Faculty of Science Division of Biology
Graduation Thesis
Recombination frequency between genes
of the human major histocompatibility complex
Marija Mulanović
Rooseveltov trg 6, 10000 Zagreb, Hrvatska
In this study we analyzed the recombination frequency between loci HLA-A and -B, and between loci HLA-B and -DRB1 in the group of 2049 families from the registry of hematopoietic stem cells recipients. All subjects were tested by one of the methods: microlymphocytotoxicity test or by polymerase chain reaction and primers specific for a particular allele or group of alleles (methods: PCR-SSP and PCR-SSO). The presence of crossingovers has been determined in 3.61% of families, out of which in 3.47% of families the crossingover has been determined in one child, and in 0.15% families the presence of crossingovers revealed in two children. The recombination frequency between loci HLA-A and -B was 0.61%, while between loci HLA-B and -DRB1 was 0.77%. Analysis of crossingover origin in the family revealed that the recombination frequency in oogenesis was 1.8 times higher than the recombination frequency in spermatogenesis. Among haplotypes with crossingovers between loci HLA-A and -B the crossingover was most common in those carrying the antigen/gene HLA-B27. In families with crossingovers between loci HLA-B and -DRB1 the recombination usually occurred in haplotypes positive for antigen/gene HLA-DR13. Results of this study are in concordance with the results from similar investigations. (49 pages, 19 figures, 6 tables, 48 references, original in: Croatian) Thesis deposited in the Central Biological Library Key words: HLA genes, recombination, crossingover Supervisor: Dr. Zorana Grubić, Assoc.Prof., University Hospital Centre Zagreb Co-supervisor: Dr. Ana Galov, Asst.Prof., Faculty of Science, University of Zagreb Reviewers: Dr. Mirjana Kalafatić, Prof., Faculty of Science, University of Zagreb Dr. Božena Mitić, Prof., Faculty of Science, University of Zagreb Replacement: Dr. Zlatko Liber, Assoc.Prof., Faculty of Science,University of Zagreb Thesis accepted: 02.12.2015.

POPIS KRATICA
AgB – glavni sustav tkivne podudarnosti kod štakora
ChL-A – glavni sustav tkivne podudarnosti kod čimpanze
DL-A – glavni sustav tkivne podudarnosti kod psa
H-2 – glavni sustav tkivne podudarnosti kod miša
RhL-A – glavni sustav tkivne podudarnosti kod majmuna
SL-A – glavni sustav tkivne podudarnosti kod svinje
C2 – komponenta komplementa
C4A – komponenta komplementa
C4B – komponenta komplementa
CD4 – glikoprotein na površini stanica imunosnog sustava (pomagačkih limfocita T,
monocita, makrofaga i dendritičkih stanica)
CD8 – glikoprotein na površini stanica imunosnog sustava (citotoksičnih limfocita T)
C.O. – prema engl. crossing over, krosingover
DNA – deoksiribonukleinska kiselina
IHWS – Međunarodni radni sastanak o tkivnoj podudarnosti
HLA – humani leukocitni antigeni
MHC – glavni sustav tkivne podudarnosti
mRNA – glasnička ribonukleinska kiselina
NK – prirodne ubilačke stanice
TNF-α – faktor nekroze tumora alfa
TNF-β – faktor nekroze tumora beta
WHO – Svjetska zdravstvena organizacija

SADRŽAJ 1. UVOD ................................ ................................ ......................... 1
1.1. Općenito o glavnom sustavu tkivne podudarnosti (HLA) ............. 1
1.1.1. Otkriće sustava HLA, uloga, organizacija i nasljeđivanje ...... 1
1.1.2. Geni HLA ................................ ................................ ......... 4
1.1.3. Molekule HLA ................................ ................................ ... 6
1.1.4. Nazivlje sustava HLA ................................ ...................... 10
1.2. Osobine sustava HLA ................................ ............................ 12
1.2.1. Polimorf izam ................................ ................................ .. 12
1.2.2. Rekombinacija ................................ ................................ 14
1.2.3. Neravnoteža udruživanja ................................ ................. 17
1.2.4. Segregacija ................................ ................................ .... 18
1.3. Primjena istraživanja sustava HLA ................................ ......... 20
1.3.1. Transplantacija tkiva i organa ................................ .......... 20
1.3.2. Geni HLA i bolesti ................................ ........................... 21
1.3.3. Populacijska istraživanja ................................ ................ 22
2. CILJEVI ISTRAŽIVANJA ................................ ............................. 24
3. MATERIJALI I METODE ................................ .............................. 25
3.1. Ispitanici ................................ ................................ .............. 25
3.2. Metode ................................ ................................ ................ 25
3.2.1. Određivanje antigena HLA ................................ ............... 25
3.2.2. Određivanje gena HLA ................................ .................... 26
3.3. Statistička obrada podataka ................................ .................. 27
4. REZULTATI ................................ ................................ ............... 28
4.1. Zbirni prikaz ispit ivane skupine obitel j i ................................ ... 28
4.2. Analiza obitelj i s krosingoverom nastalim u oogenezi .............. 29
4.3. Analiza obitelj i s krosingoverom nastalim u spermatogenezi .... 30
4.4. Analiza obitelj i s rekombinantnim haplotipom HLA nepoznatog
pori jekla ................................ ................................ ..................... 32
4.5. Analiza obitelj i s dva krosingovera ................................ ......... 33
4.6. Analiza haplot ipova HLA u obitel j ima s krosingoverom između
lokusa HLA-A i -B................................ ................................ ........ 36

4.7. Analiza haplot ipova HLA u obitel j ima s krosingoverom između
lokusa HLA-B i -DRB1 ................................ ................................ . 38
5. RASPRAVA ................................ ................................ ............... 40
6. ZAKLJUČAK ................................ ................................ .............. 43
7. LITERATURA ................................ ................................ ............. 44
8. ŽIVOTOPIS ................................ ................................ ............... 49

1
1. UVOD
1.1. Općenito o glavnom sustavu tkivne podudarnosti (HLA)
Kod većine dosad istraživanih kralješnjaka pronađena je usko vezana skupina
gena i obitelji gena čiji produkti, odnosno molekule, imaju ključnu ulogu u
međusobnom prepoznavanju stanica i raspoznavanju vlastitog od tuđeg (Klein,
1986). Taj kompleks gena nazvan je glavni sustav gena tkivne podudarnosti,
odnosno MHC (engl. Major Histocompatibility Complex). Produkti gena ovog sustava
– antigeni tkivne podudarnosti – dolaze u različitim količinama na različitim stanicama
pojedinog organizma. Kod čovjeka je taj sustav gena najprije otkriven na leukocitima i
naziva se sustav HLA (engl. Human Leukocyte Antigens).
1.1.1. Otkriće sustava HLA, uloga, organizacija i nasljeđivanje
Sam koncept odbacivanja stranog tkiva kao rezultat imunosnog odgovora na
antigene tkivne podudarnosti, potječe od radova Petera Gorera 1930-ih godina
(Thorsby, 2009). Gorer je izvodio istraživanja na visokosrodnim sojevima
laboratorijskih miševa u cilju određivanja antigena krvnih grupa. Otkrio je četiri grupe
gena koje su nosile nasljednu uputu za antigene na krvnim stanicama. Gorer je
kasnije nastavio istraživanja 40-ih i 50-ih godina prošlog stoljeća zajedno s
Georgeom Snellom i utvrdio da upravo antigeni kodirani genima jedne od tih grupa
sudjeluju u odbacivanju transplantiranih tumora i ostalih tkiva. Snell je ovu grupu
gena nazvao H geni (engl. Histocompatibility Genes), odnosno geni tkivne
podudarnosti.
Eksperimentalne metode križanja u srodstvu i presađivanja transplantata kože koje
su se izvodile na miševima dovele su do otkrića glavnog sustava tkivne podudarnosti
kod miša, ali dokaze o postojanju takvog sustava kod čovjeka pružile su tek
alogenične transfuzije krvi i transplantacije organa. Dausset i suradnici su 1954.
godine uočili da pacijenti koji su odbacili bubreg ili su imali posttransfuzijsku reakciju
često razvijaju cirkulirajuća citotoksična protutijela. Navedena protutijela su reagirala

2
na molekule antigena s površine bijelih krvnih stanica davatelja krvi, odnosno organa,
te je serum primatelja uz prisustvo komplementa lizirao limfocite davatelja (Thorsby,
2009).
Na važnost sustava HLA u fiziologiji imunosnog odgovora ukazali su Benacerraf i
suradnici 1960-ih koji su pokazali da različiti visokosrodni sojevi zamoraca i miševa
nakon imunizacije polipeptidnim antigenima razvijaju ili ne razvijaju protutijela kao
odgovor na imunizaciju. Taj imunosni odgovor je autosomalno nasljedno svojstvo
koje se pripisuje regiji MHC, dok su geni koji ga kontroliraju nazvani Ir genima (engl.
Immune Response). Tek 1970-ih je objašnjena centralna uloga gena MHC nakon što
se pokazalo da antigen-specifični limfociti T ne prepoznaju antigene slobodne u
otopini, već prepoznaju samo dijelove, točnije peptidne fragmente koji su
nekovalentno vezani za molekule MHC. Snell, Dausset i Benacerraf dobili su
Nobelovu nagradu 1980. godine za svoj doprinos otkriću i razumijevanju glavnog
sustava tkivne podudarnosti kod miša i kod čovjeka (Thorsby, 2009).
Iako je do otkrića sustava HLA došlo tek kad su produkti gena ovog sustava
identificirani kao odgovorni za odbacivanje nepodudarnog presatka, ključna uloga
gena HLA počiva u pokretanju i regulaciji imunosnog odgovora u svrhu obrane od
mikroorganizama. Molekule HLA su membranski proteini čije izvanstanične domene
izlažu peptidne fragmente nastale preradom stranih proteinskih antigena. Postoje
dvije osnovne grupe molekula HLA koje prepoznaju dvije različite vrste limfocita T.
Citotoksični limfociti T prepoznaju strani antigen istodobno s molekulama HLA
razreda I, dok molekule HLA razreda II udružene sa stranim peptidima na tzv.
predočnim stanicama (makrofagi, limfociti B) prepoznaju pomagački limfociti T. Na taj
način sustav HLA sudjeluje u obrani organizma kao sastavni dio stečene ili specifične
imunosti, kako u humoralnoj, tako i u staničnoj imunosti. Geni HLA posreduju u
staničnim interakcijama, sudjeluju u regulaciji imunoreaktivnosti, određuju osjetljivost
na viruse, imunosnu preosjetljivost i autoimunosne bolesti (Andreis i sur., 2004).
Postoji velika homologija unutar sustava tkivne podudarnosti između različitih
vrsta, no svaka vrsta pokazuje pojedine specifičnosti. Glavni sustav tkivne
podudarnosti kod različitih vrsta nosi različite nazive: AgB kod štakora, B kod pilića,
ChL-A kod čimpanze, DL-A kod psa, H-2 kod miša, RhL-A kod rhesus majmuna, a
SL-A kod svinje. Kod čovjeka se geni HLA nalaze na kromosomu 6 (Janeway i sur.,
2005). Na slici 1 prikazana je regija HLA i raspored gena u njoj.

3
Slika 1. Organizacija gena glavnog sustava tkivne podudarnosti u čovjeka
(izvor: Janeway, Travers, Walport, Shlomchik, 2005)
Geni sustava HLA pokazuju veliku raznolikost, što znači da u populaciji postoji
mnogo alternativnih oblika – alela nekog gena. Geni koji su dio sustava HLA fizički su
smješteni vrlo blizu jedan drugome zbog čega većina osoba nasljeđuje alele ove
usko vezane skupine gena u setovima, po jedan set od svakog roditelja. Takav set
gena HLA naziva se haplotip HLA, a svaka jedinka nasljeđuje jedan majčin i jedan
očev haplotip HLA. U jednoj stanici vidljiva je genska ekspresija oba naslijeđena
haplotipa HLA, jer su geni HLA kodominantni (Goldsby i sur., 2003). Nasljeđivanje
haplotipova HLA od heterozigotnih roditelja je prikazano na slici 2. U slučaju da otac i
majka imaju različite haplotipove HLA, mogućnost da potomci naslijede isti majčin i
očev haplotip HLA i budu međusobno identični je 25%. Na slici 2 vidi se i primjer
rekombinantnog majčinog haplotipa koji je nastao kao posljedica krosingovera u
mejozi prilikom nastanka majčinih spolnih stanica.

4
Slika 2. Nasljeđivanje haplotipova HLA u obitelji
(A i B – očevi haplotipovi HLA, C i D – majčini haplotipovi HLA,
R – rekombinantni majčin haplotip HLA)
1.1.2. Geni HLA
Regija HLA smještena je na kraćem kraku kromosoma 6 (6p21.3). Sastoji se
od preko 220 gena i obuhvaća više od 4x106 parova baza DNA (Horton i sur., 2004).
Podijeljena je u tri subregije unutar kojih se nalaze i geni koji nisu geni HLA. Geni
HLA dijele se u dvije skupine gena:
1. geni HLA razreda I kodiraju glikoproteine prisutne na površini gotovo svih
stanica s jezgrom; glavna uloga im je predočivanje peptidnih antigena
citotoksičnim limfocitima T
2. geni HLA razreda II kodiraju glikoproteine koji se nalaze na antigen-predočnim
stanicama poput makrofaga, dendritičkih stanica i limfocita B gdje predočuju
prerađene antigenske peptide pomagačkim limfocitima T

5
Slika 3. Detaljna mapa regije HLA
(izvor: Janeway, Travers, Walport, Shlomchik, 2005)
Regija gena HLA razreda I proteže se na oko 2x106 parova baza i sadrži
otprilike 20-ak gena. Kako je istaknuto na slici 3, unutar nje se nalaze tri osnovna
gena (HLA-A, -B i -C) koji kodiraju tzv. klasične molekule HLA razreda I. Ti geni
kodiraju α lanac molekule HLA razreda I. Unutar regije HLA razreda I nalaze se i geni
koji kodiraju neklasične molekule HLA razreda I (HLA-E , -F , -G , -J i -X). Tu se
nalazi i obitelj gena MIC koja uključuje gene MIC-A, -B, -C, -D i -E koji su dobili ime
MIC (engl. MHC class I related chain) zbog toga što njihove molekule nalikuju
molekulama HLA razreda I. Geni HLA razreda I koji kodiraju neklasične molekule su
mnogo manje polimorfni od gena HLA-A, -B i -C. Neki od neklasičnih gena HLA
razreda I su pseudogeni pa prema tome ne nose nasljednu uputu za proteinski
produkt. Točna funkcija većine neklasičnih molekula HLA razreda I još nije do kraja
razjašnjena (Goldsby i sur., 2003).
Geni HLA-DR, HLA-DP i HLA-DQ nose nasljednu uputu za molekule HLA
razreda II – tzv. klasične molekule HLA razreda II. U toj regiji nalaze se još i tzv.
neklasični geni HLA razreda II (HLA-DM i HLA-DO).
Obitelj gena HLA-DR sastoji se od jednog gena koji kodira za α lanac (HLA-
DRA) i čak do 9 gena za β lanac (HLA-DRB1 do HLA-DRB9) uključujući i
pseudogene. U humanoj regiji HLA-DR postoje 4 funkcionalna gena za β lanac, ali su

6
u haplotipu prisutna maksimalno dva gena za β lanac i na taj način na nekim
stanicama mogu biti prisutne dvije različite vrste molekula HLA-DR.
Geni obitelji HLA-DP i HLA-DQ imaju svaki po jedan gen za α i β lanac, te
dodatni par pseudogena. U stanici se α lanci molekula HLA-DR, HLA-DQ i HLA-DP
povezuju prvenstveno s β lancima iste genske obitelji. To znači da će se produkti
gena HLA-DPA1 i HLA-DPB1 povezati i tvoriti molekulu HLA-DP. Slično tome će geni
HLA-DQA1 i HLA-DQB1 nositi uputu za molekule HLA-DQ.
Skupini neklasičnih gena HLA razreda II pripadaju geni HLA-DM i HLA-DO.
Produkti ovih gena su molekule HLA-DM i HLA-DO. Molekule HLA-DM olakšavaju
vezivanje stranih peptida na molekule HLA razreda II, dok molekule HLA-DO koje se
eksprimiraju samo u timusu i na zrelim limfocitima B služe kao regulatori procesiranja
stranih antigena.
Centralna regija se sastoji od vrlo raznolike skupine gena. Produkti gena
centralne regije su ili na neki način povezani s funkcijama molekula HLA ili su geni
centralne regije pod nadzorom sličnih kontrolnih mehanizama kao i geni HLA (Yung i
sur., 2000). Geni ove regije kodiraju neke komponente komplementa (C2, C4A, C4B i
faktor B), proteine toplinskoga šoka i citokine TNF-α i TNF-β.
1.1.3. Molekule HLA
Molekule (antigeni) HLA su membranski glikoproteini, sastavni dijelovi
staničnih membrana. Slično kao i geni HLA, molekule HLA se prema svojim
funkcijama dijele na molekule HLA razreda I i molekule HLA razreda II. Oba tipa
molekula HLA funkcioniraju kao visoko specijalizirane molekule koje formiraju
stabilne komplekse s peptidnim fragmentima stranih antigena i predočuju ih na
površini stanice kako bi ih prepoznali limfociti T. Molekule HLA razreda I pronalazimo
na gotovo svim stanicama s jezgrom. Molekule HLA razreda II nalaze se samo na
određenim stanicama imunosnog sustava i to na limfocitima B, makrofagima,
dendritičkim i endotelijalnim stanicama (Andreis i sur., 2004).
Kako bi limfociti T prepoznali strani antigen, on mora biti pocijepan na male
peptidne fragmente koji će formirati kompleks s molekulama HLA razreda I ili
molekulama HLA razreda II (Germain, 1994). Konverzija stranog antigena u peptidne
fragmente vezane na molekulu HLA naziva se procesiranje i predočivanje antigena.

7
Strani antigen može biti predočen u kompleksu s molekulom HLA razreda I ili
molekulom HLA razreda II, ovisno o načinu njegovog ulaska u stanicu. Egzogeni
antigeni su oni koji nastaju izvan stanice domaćina, te endocitozom ili fagocitozom
ulaze u antigen-predočne stanice (makrofagi, dendritičke stanice i limfociti B).
Antigen-predočne stanice zatim razgrađuju progutani egzogeni antigen u peptidne
fragmente endocitoznim procesiranjem. Peptidni fragmenti stranog antigena bit će
potom vezani na molekulu HLA razreda II. Taj kompleks prepoznaju receptori
limfocita T na čijoj se površini nalaze CD4 glikoproteini i općenito funkcioniraju kao
pomagački limfociti T. Endogeni antigeni nastaju unutar stanice domaćina, to su npr.
virusni proteini koji nastaju unutar stanice zaražene virusom ili jedinstveni proteini
koje stvaraju tumorske stanice. Endogeni antigeni se cijepaju na peptidne fragmente
koji se zatim vežu na molekule HLA razreda I. Nastali kompleks peptida s molekulom
HLA razreda I transportira se do stanične membrane. Citotoksični limfociti T s CD8
glikoproteinima prepoznaju strani antigen na površini stanica koje ih izražavaju u
kompleksu s molekulom HLA razreda I, te ih zatim napadaju i ubijaju (Janeway i sur.,
2005).
1.1.3.1. Molekule HLA razreda I
Kao što je prikazano na slici 4, molekula HLA razreda I sastoji se od α lanca
veličine 45 kDa koji je nekovalentno povezan s β2 mikroglobulinom veličine 12 kDa.
Nasljednu uputu za β2 mikroglobulinski lanac nosi visoko konzervirani gen koji se
nalazi na kromosomu 15. Dok je α lanac usidren u staničnoj membrani hidrofobnim
transmembranskim dijelom i hidrofilnim citoplazmatskim dijelom, β2 mikroglobulinski
lanac nalazi se i slobodan u serumu. Za ekspresiju molekula HLA razreda I
neophodno je nekovalentno vezivanje β2 mikroglobulina uz α lanac (Natarajan i sur.,
1999).

8
Slika 4. Shematski prikaz molekule HLA razreda I
(izvor: Goldsby, Kindt, Osborne, Kuby, 2003)
Strukturne analize α lanca pokazale su da se on sastoji od 3 izvanstanične
domene (α1, α2 i α3) od kojih se svaka sastoji od otprilike 90 aminokiselina,
transmembranski dio sastoji se od oko 25 hidrofobnih aminokiselina, a citoplazmatski
dio sastoji se od 30-ak aminokiselina (Goldsby i sur., 2003). Domene α1 i α2 tvore
platformu sastavljenu od 8 antiparalelnih β-nabora koju sa svake strane obuhvaća po
jedna α-uzvojnica tako da formiraju duboku pukotinu ili utor dovoljno velik da veže
peptid sastavljen od 8-10 aminokiselina (Madden, 1995). Ta pukotina je najvažniji dio
molekule jer služi kao vezno mjesto za prerađene strane peptide. Veličinom i
rasporedom aminokiselina β2 mikroglobulin je najsličniji domeni α3, samo što ne
posjeduje transmembranski dio i nekovalentno je vezan na molekulu HLA razreda I.
Zbog sličnosti u aminokiselinskom slijedu s imunoglobulinskim konstantnim regijama,
kompleks α lanca molekule HLA razreda I i β2 mikroglobulina ubrajamo u
imunoglobulinsku superporodicu (slika 6). Domena α3 je visoko konzervirana kod
svih molekula HLA razreda I. Unutar nje je slijed koji stupa u interakciju s CD8
molekulom na membrani citotoksičnih limfocita T (Goldsby i sur., 2003).

9
1.1.3.2. Molekule HLA razreda II
Slika 5. Shematski prikaz molekule HLA razreda II
(izvor: Goldsby, Kindt, Osborne, Kuby, 2003)
Molekule HLA razreda II svojom cjelokupnom trodimenzionalnom strukturom
polipeptidnih lanaca vrlo su slične molekulama HLA razreda I kao što je vidljivo na
slici 5. Po svojoj strukturi su heterodimeri i sastoje se od dva različita polipeptidna
lanca povezana nekovalentnim interakcijama. Molekularna težina lanca α je između
30-34 kDa, dok je lanac β veličine 26-29 kDa, ovisno o genskom lokusu. Poput α
lanca molekula razreda I, molekule razreda II su također glikoproteini koji imaju
izvanstaničnu domenu, transmembranski dio i citoplazmatski dio (Goldsby i sur.,
2003). Oba polipeptidna lanca molekula HLA razreda II imaju po dvije ekstracelularne
domene: α1 i α2 na jednom lancu te domene β1 i β2 na drugom lancu. Domene α2 i
β2 pokazuju strukturnu sličnost imunoglobulinskom slijedu, stoga i molekule HLA
razreda II ubrajamo u imunoglobulinsku superporodicu (slika 6). Domene α1 i β1 koje
su udaljenije od membrane formiraju antigen-veznu pukotinu. Za razliku od pukotine
kod molekule HLA razreda I koja je zatvorena na oba kraja, antigen-vezna pukotina
molekule HLA razreda II je otvorena i zbog toga se u nju mogu smjestiti nešto duži
peptidi duljine 13-18 aminokiselina (Madden, 1995). Domena β2 sadrži vezno mjesto
za molekulu CD4 pomoću kojeg molekule HLA razreda II na antigen-predočnim

10
stanicama stupaju u interakciju s limfocitima T, analogno interakciji molekula HLA
razreda I s molekulom CD8 (Brown 1993).
Slika 6. Neki članovi imunoglobulinske superporodice
(imunoglobulinske konzervirane domene osjenčane su sivom bojom, dok su antigen-
vezne domene osjenčane plavom bojom)
(izvor: Alberts, Johnson, Lewis, Raff, Roberts, Walter, 2002)
1.1.4. Nazivlje sustava HLA
Ubrzo nakon otkrića antigena HLA i nakon održavanja 1. Međunarodnog
radnog sastanka o tkivnoj podudarnosti (1st International Histocompatibility
Workshop – IHWS) 1965. godine ukazala se potreba za standardiziranim nazivljem
serološki definiranih antigena. Prvi istraživači sustava HLA su neovisno jedan od
drugog dodjeljivali vlastite oznake, tako da su različiti laboratoriji imali različite nazive
za iste antigene HLA (Thorsby, 2009).
Odbor za nazivlje HLA pod pokroviteljstvom Svjetske zdravstvene organizacije
(WHO Nomenclature Comitee for Factors of the HLA System) zadužen je za
imenovanje novih gena i alela HLA. Naziv svakog pojedinog alela HLA ima svoj
jedinstveni broj. Taj broj može se sastojati od najviše 4 seta brojeva međusobno
odijeljenih dvotočkom. Svi aleli dobivaju oznaku od najmanje četiri broja, što
odgovara duljini od prva dva seta brojeva, a dulji nazivi se dodjeljuju ukoliko je to
potrebno. Brojevi prije prve dvotočke opisuju tip alela koji u najvećem broju slučajeva
11
odgovara određenoj serološkoj specifičnosti. Nakon prve dvotočke slijedi set brojeva
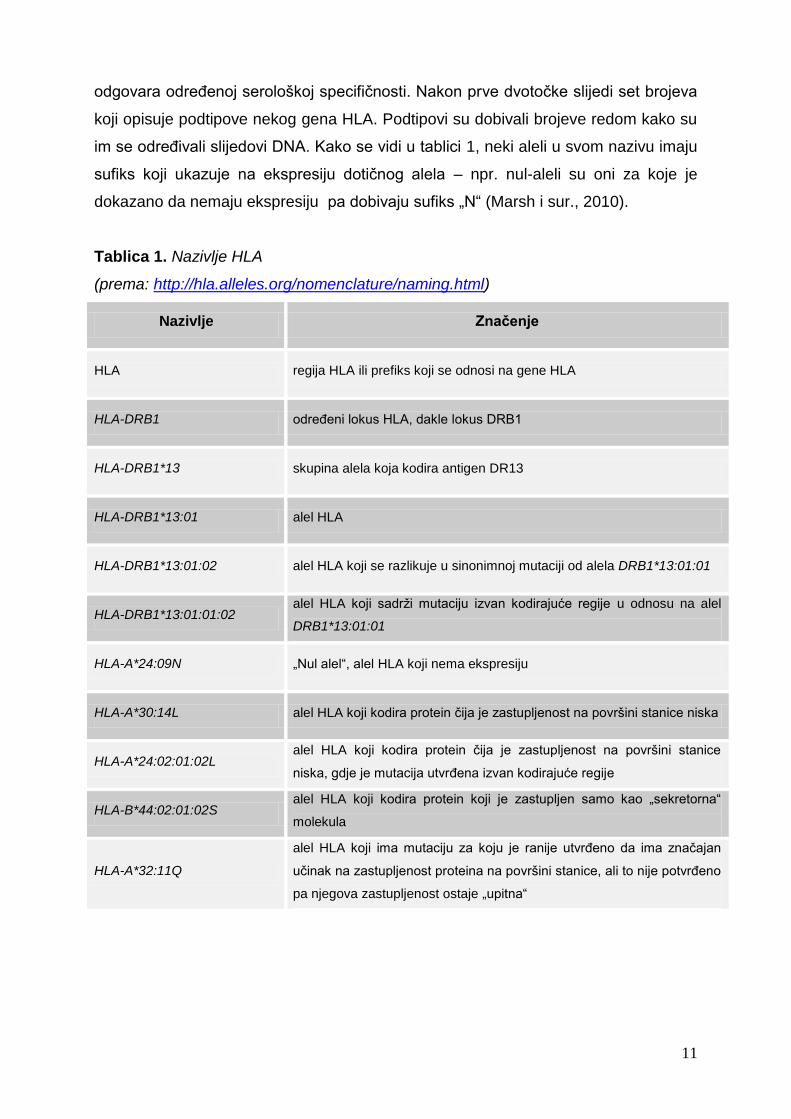
koji opisuje podtipove nekog gena HLA. Podtipovi su dobivali brojeve redom kako su
im se određivali slijedovi DNA. Kako se vidi u tablici 1, neki aleli u svom nazivu imaju
sufiks koji ukazuje na ekspresiju dotičnog alela – npr. nul-aleli su oni za koje je
dokazano da nemaju ekspresiju pa dobivaju sufiks „N“ (Marsh i sur., 2010).
Tablica 1. Nazivlje HLA
(prema: http://hla.alleles.org/nomenclature/naming.html)
Nazivlje Značenje
HLA regija HLA ili prefiks koji se odnosi na gene HLA
HLA-DRB1 određeni lokus HLA, dakle lokus DRB1
HLA-DRB1*13 skupina alela koja kodira antigen DR13
HLA-DRB1*13:01 alel HLA
HLA-DRB1*13:01:02 alel HLA koji se razlikuje u sinonimnoj mutaciji od alela DRB1*13:01:01
HLA-DRB1*13:01:01:02 alel HLA koji sadrži mutaciju izvan kodirajuće regije u odnosu na alel
DRB1*13:01:01
HLA-A*24:09N „Nul alel“, alel HLA koji nema ekspresiju
HLA-A*30:14L alel HLA koji kodira protein čija je zastupljenost na površini stanice niska
HLA-A*24:02:01:02L alel HLA koji kodira protein čija je zastupljenost na površini stanice
niska, gdje je mutacija utvrđena izvan kodirajuće regije
HLA-B*44:02:01:02S alel HLA koji kodira protein koji je zastupljen samo kao „sekretorna“
molekula
HLA-A*32:11Q
alel HLA koji ima mutaciju za koju je ranije utvrđeno da ima značajan
učinak na zastupljenost proteina na površini stanice, ali to nije potvrđeno
pa njegova zastupljenost ostaje „upitna“

12
1.2. Osobine sustava HLA
Osnovna uloga molekula HLA je vezivanje peptidnih fragmenata proizašlih od
patogena i njihovo predočivanje na staničnoj membrani kako bi ih prepoznali
odgovarajući limfociti T. To gotovo uvijek rezultira uništavanjem patogena tako da
stanice zaražene virusom budu ubijene, makrofagi se aktiviraju i ubijaju bakterije koje
eventualno žive u unutarstaničnim vezikulama, a limfociti B se aktiviraju tako što
proizvode antitijela koja eliminiraju ili neutraliziraju izvanstanične patogene. Prema
tome, postoji jak selektivni pritisak u korist bilo kojeg patogena da mutira na takav
način koji će mu omogućiti da izbjegne prezentaciju od strane molekule HLA (Franco
i sur., 1995). No, postoje dvije vrlo važne osobine sustava HLA koje onemogućuju
patogenima da izbjegnu imunosni odgovor. U prvom redu zbog velikog broja gena
HLA, a potom zbog velikog broja alela na svakom od tih gena. Prema tome svaka
jedinka posjeduje set molekula HLA koje mogu vezati veliki broj različitih peptida
(Potts i Slev, 1995).
1.2.1. Polimorfizam
Nakon što su se metode serološke tipizacije upotpunile direktnim
sekvenciranjem gena HLA, pronađen je ogroman broj novih alela. Ovi aleli razlikuju
se u DNA slijedovima između dvije različite jedinke od 5-10%. Broj aminokiselina u
kojima se proteinski produkti različitih alela HLA međusobno razlikuju može iznositi
čak do 20 aminokiselinskih ostataka. Broj novootkrivenih alela HLA razreda I u
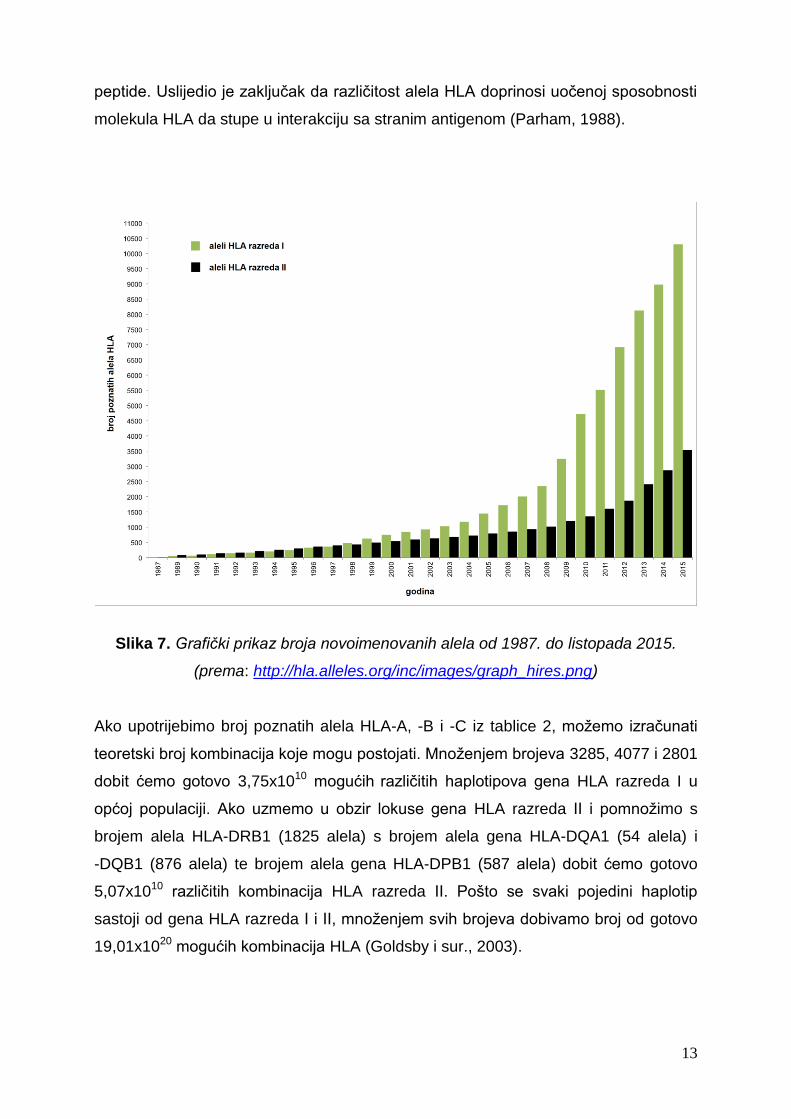
posljednjih nekoliko godina pokazuje eksponencijalni rast (slika 7). Zanimljivo je da
razlike u slijedovima parova baza između različitih alela HLA nisu nasumično
raspoređene uzduž čitavog polipeptidnog lanca, već su uglavnom smještene u onim
dijelovima gena koji kodiraju domene α1 i α2 molekula HLA razreda I, odnosno kod
gena HLA razreda II u onom dijelu odgovornom za kodiranje domena α1 i β2
molekule. Primjenom rendgenske kristalografije utvrdilo se da je od 17 aminokiselina
koje su pokazivale značajan polimorfizam unutar molekule HLA-A2, čak njih 15
smješteno unutar procjepa u molekuli koji ima ulogu veznog mjesta za strane
13
peptide. Uslijedio je zaključak da različitost alela HLA doprinosi uočenoj sposobnosti
molekula HLA da stupe u interakciju sa stranim antigenom (Parham, 1988).
Slika 7. Grafički prikaz broja novoimenovanih alela od 1987. do listopada 2015.
(prema: http://hla.alleles.org/inc/images/graph_hires.png)
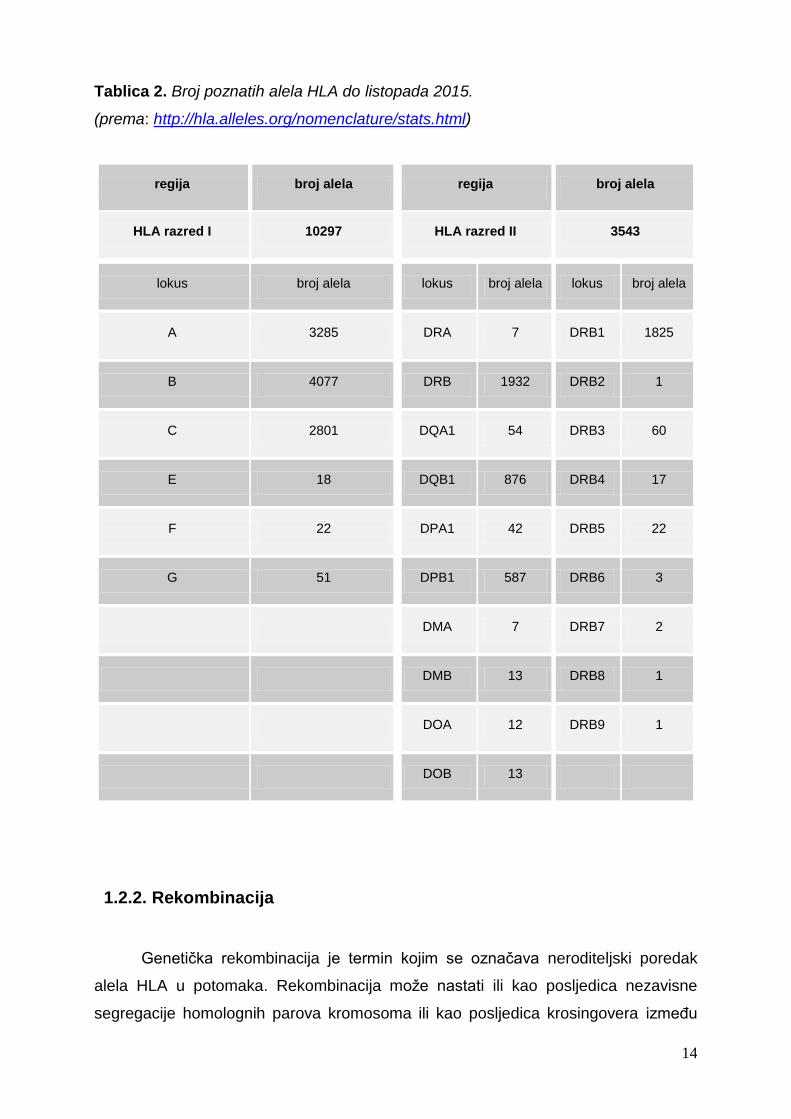
Ako upotrijebimo broj poznatih alela HLA-A, -B i -C iz tablice 2, možemo izračunati
teoretski broj kombinacija koje mogu postojati. Množenjem brojeva 3285, 4077 i 2801
dobit ćemo gotovo 3,75x1010 mogućih različitih haplotipova gena HLA razreda I u
općoj populaciji. Ako uzmemo u obzir lokuse gena HLA razreda II i pomnožimo s
brojem alela HLA-DRB1 (1825 alela) s brojem alela gena HLA-DQA1 (54 alela) i
-DQB1 (876 alela) te brojem alela gena HLA-DPB1 (587 alela) dobit ćemo gotovo
5,07x1010 različitih kombinacija HLA razreda II. Pošto se svaki pojedini haplotip
sastoji od gena HLA razreda I i II, množenjem svih brojeva dobivamo broj od gotovo
19,01x1020 mogućih kombinacija HLA (Goldsby i sur., 2003).
14
Tablica 2. Broj poznatih alela HLA do listopada 2015.
(prema: http://hla.alleles.org/nomenclature/stats.html)
regija broj alela regija broj alela
HLA razred I 10297 HLA razred II 3543
lokus broj alela lokus broj alela lokus broj alela
A 3285 DRA 7 DRB1 1825
B 4077 DRB 1932 DRB2 1
C 2801 DQA1 54 DRB3 60
E 18 DQB1 876 DRB4 17
F 22 DPA1 42 DRB5 22
G 51 DPB1 587 DRB6 3
DMA 7 DRB7 2
DMB 13 DRB8 1
DOA 12 DRB9 1
DOB 13
1.2.2. Rekombinacija
Genetička rekombinacija je termin kojim se označava neroditeljski poredak
alela HLA u potomaka. Rekombinacija može nastati ili kao posljedica nezavisne
segregacije homolognih parova kromosoma ili kao posljedica krosingovera između

15
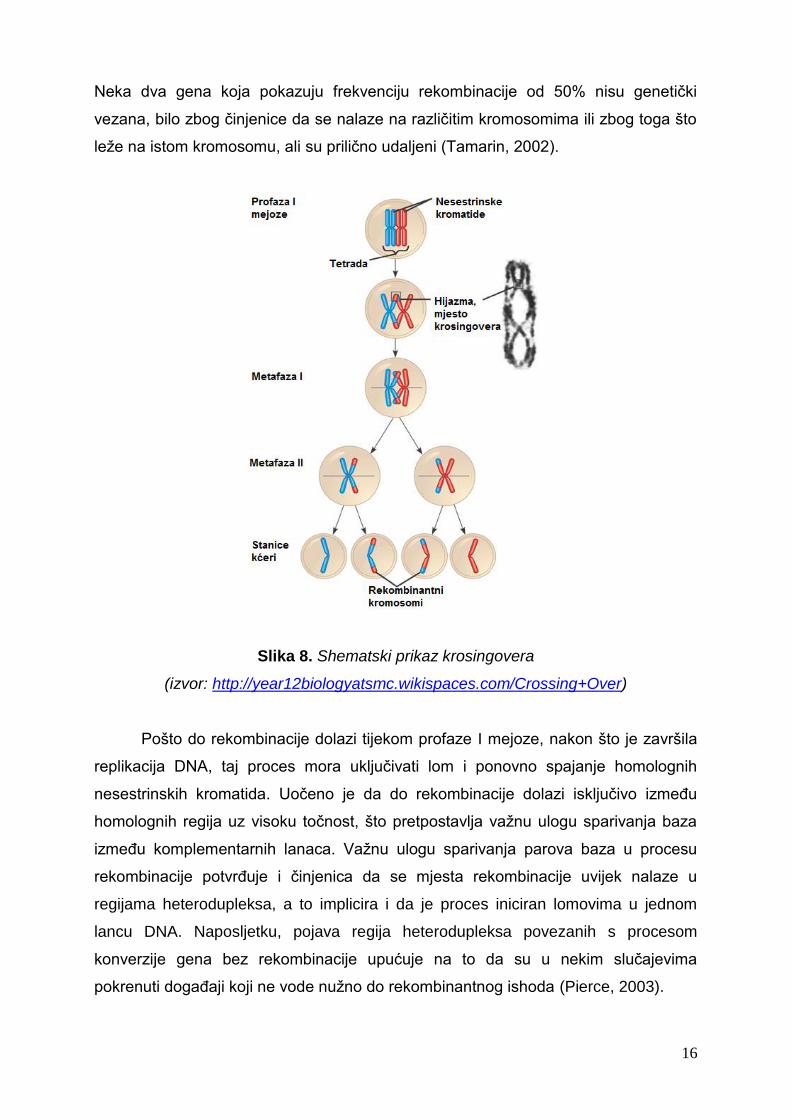
homolognih dijelova DNA. Krosingover (engl. crossing over) je postupak u kojem
dolazi do izmjene dijelova nesestrinskih kromatida homolognih kromosoma u profazi I
mejoze.
Svaki par kromosoma koji ulazi u proces mejoze uslijed prethodnog
udvostručenja sastoji se od 4 kromatide. Takvi kromosomi nazivaju se bivalenti ili
tetrade. Kromatide dva homologna kromosoma su međusobno spojene pomoću
proteinske strukture, tzv. sinaptonemskog kompleksa. U profazi I mejoze dolazi do
sparivanja homolognih kromosoma na karakterističan način što se naziva sinapsa, pri
čemu se određeno područje na jednom kromosomu poklapa s istim područjem na
drugom kromosomu. Tako nastaje ključni trenutak mejoze u kojem se stvara
prekriženje između dvije nesetrinske kromatide homolognih kromosoma. Na
mjestima prekriženja dolazi do izmjene genskog materijala između nesestrinskih
kromatida – dolazi do krosingovera. Nakon rekombinacije genskog materijala niti
jedna kromatida nije više identična drugoj. Krosingoverom nastaju dvije vrste
rekombinanata u jednakom broju zbog načina na koje se na određenom mjestu u
bivalentu prekidaju, izmjenjuju i ponovno spajaju dvije nesestrinske kromatide. Nakon
mejoze nastat će dva produkta koji su rekombinantni i dva koji su roditeljski. Ukoliko
se ne dogodi krosingover, svi produkti mejoze bit će roditeljski. Značaj procesa
krosingovera je da on znatno može povećati genetičku raznolikost u gametama
povezujući alele koji prije njega nisu bili skupa (Tamarin, 2002).
Ako uspoređujemo parove gena koji su smješteni blizu jedan drugome na
istom kromosomu i time ne poštuju Mendelov zakon nezavisne segregacije, kažemo
da su to vezani geni, jer su manje ili više usko povezani u procesu prenošenja na
sljedeću generaciju u kojoj će prevladavati određeni roditeljski haplotip. Upravo
saznanje o postojanju vezanih gena i rekombinaciji najviše je doprinijelo genetičkim
istraživanjima kojima se određivao relativan položaj gena na kromosomu –
uspoređivanjem frekvencija rekombinacije mnogo genskih parova (Pierce, 2003).
Vidljivi fizički znakovi nastanka krosingovera su hijazme (slika 8). Ukoliko
pretpostavimo da se hijazme mogu pojaviti bilo gdje uzduž kromosoma može se
zaključiti da će se što su neka dva gena udaljenija, hijazme formirati većom
učestalošću. To je razlog zašto veća frekvencija rekombinacije nekog para gena
odražava veću udaljenost između njih. Općenito, frekvencija rekombinacije varira od
0-50%, s tim da statistički značajne vrijednosti veće od 50% ukazuju na činjenicu da
su dva gena koja promatramo vezana i da se moraju nalaziti na istom kromosomu.
16
Neka dva gena koja pokazuju frekvenciju rekombinacije od 50% nisu genetički
vezana, bilo zbog činjenice da se nalaze na različitim kromosomima ili zbog toga što
leže na istom kromosomu, ali su prilično udaljeni (Tamarin, 2002).
Slika 8. Shematski prikaz krosingovera
(izvor: http://year12biologyatsmc.wikispaces.com/Crossing+Over)
Pošto do rekombinacije dolazi tijekom profaze I mejoze, nakon što je završila
replikacija DNA, taj proces mora uključivati lom i ponovno spajanje homolognih
nesestrinskih kromatida. Uočeno je da do rekombinacije dolazi isključivo između
homolognih regija uz visoku točnost, što pretpostavlja važnu ulogu sparivanja baza
između komplementarnih lanaca. Važnu ulogu sparivanja parova baza u procesu
rekombinacije potvrđuje i činjenica da se mjesta rekombinacije uvijek nalaze u
regijama heterodupleksa, a to implicira i da je proces iniciran lomovima u jednom
lancu DNA. Naposljetku, pojava regija heterodupleksa povezanih s procesom
konverzije gena bez rekombinacije upućuje na to da su u nekim slučajevima
pokrenuti događaji koji ne vode nužno do rekombinantnog ishoda (Pierce, 2003).

17
Prema jednom molekularnom modelu krosingovera, rekombinacija je vjerojatno
uvijek pokrenuta zarezivanjem lanca DNA kojeg za vrijeme mejoze cijepaju specifični
enzimi na točno određenim mjestima. Ta mjesta su tzv. preferirana mjesta
rekombinacije i kod sisavaca se nazivaju „hot spots“ (eng. hot spot = vruće mjesto)
Na molekularnom nivou, rekombinacija se sastoji od loma, invazije, migracije i
razdruživanja komplementarnih lanaca DNA koji potječu od različitih homolognih
kromosoma. U tom procesu važnu ulogu imaju sparivanje baza i popravak DNA
(Carrington, 1999).
Otkriće krosingovera između pojedinih gena HLA bilo je najjači dokaz da
unutar sustava HLA postoji nekoliko segregantnih skupina alelomorfnih gena.
Zahvaljujući tom otkriću bilo je moguće na kromosomu 6 točno odrediti položaj
pojedinih lokusa HLA. Premda je učestalost rekombinacije unutar sustava HLA koja
je nastala kao posljedica krosingovera vrlo niska i iznosi približno 1% na 1x106
parova baza DNA po mejozi, značajno doprinosi raznolikosti HLA. Genska
rekombinacija razlog je nastanka novih kombinacija alela, tj. haplotipova HLA.
Učestalost krosingovera između lokusa HLA-A i -B iznosi oko 1%, između lokusa
HLA-B i -C oko 0,2%, te između lokusa HLA-B i -DRB1 oko 0,5%, koliko iznosi i
učestalost krosingovera između lokusa HLA-A i –C (Kaštelan, 1983).
1.2.3. Neravnoteža udruživanja
Teoretski izračun raznolikosti opisan u poglavlju o polimorfizmu pretpostavlja
činjenicu da su kombinacije alela potpuno nasumične. U stvarnosti je raznolikost
manja zbog toga što se određeni aleli u haplotipovima HLA pojavljuju češće nego što
se to može predvidjeti nasumičnim kombiniranjem i ta pojava se naziva neravnoteža
udruživanja (eng. linkage disequilibrium). Ukratko, neravnoteža udruživanja je razlika
između učestalosti koja je uočena za određenu kombinaciju alela i one frekvencije
koja je bila očekivana s obzirom na frekvenciju pojedinačnih alela. Očekivana
frekvencija kombinacije neka dva alela može se izračunati množenjem pojedinačnih
frekvencija tih alela. Npr. ako se HLA-A1 pojavljuje u 16% osoba u nekoj populaciji
(dakle frekvencijom koja iznosi 0,16%), a HLA-B8 se pojavljuje kod 9% osoba u toj
skupini (dakle frekvencijom od 0,09%), može se očekivati da će otprilike 1,4% jedinki
te skupine imati oba alela HLA. Podaci pokazuju da se kombinacija alela HLA-A1 i

18
HLA-B8 pronalazi kod 4,5% pojedinaca u populacijama europskog porijekla. Ova
razlika je mjera neravnoteže udruživanja između ova dva navedena alela HLA
razreda I. Navedeni aleli su sastavni dio haplotipa HLA-A*01, -B*08, -DRB*0301, -
DQB1*0201 koji je primjer jedne od najjače neravnoteže udruživanja među
populacijama europskog porijekla (Shankarkumar, 2004).
Visokoj učestalosti određenih kombinacija alela HLA prema jednoj teoriji mogli
su doprinijeti učinci selekcije jer su npr. određene kombinacije omogućile otpornost
na neke bolesti, ili su naprotiv, neki drugi haplotipovi HLA mogli proizvesti štetne
učinke poput osjetljivosti na autoimune bolesti (Abbas, 2004).
1.2.4. Segregacija
Prema Mendelovom prvom zakonu nasljeđivanja princip segregacije navodi da
svaki pojedini diploidni organizam posjeduje dva alela za neko nasljedno svojstvo.
Prilikom formiranja gameta ova dva alela se odvajaju – segregiraju, tako da po jedan
alel odlazi u svaku od gameta. Prema Suttonovoj kromosomalnoj teoriji nasljeđivanja
diploidne stanice posjeduju dva seta kromosoma, s tim da svaki kromosom posjeduje
odgovarajući homologni kromosom s kojim se sparuje. Jedan kromosom iz svakog
homolognog para nasljeđen je od majke, a drugi od oca. Dakle, diploidne stanice u
kojima postoje dvije kopije svakog kromosoma posjeduju po dva alela za svaki lokus
koji čine genotip. Pošto u diploidnom organizmu svaki gen postoji u dvije kopije,
moguće su varijacije u fenotipu – izraženim osobinama tog gena. Ukoliko su obje
kopije gena identične, dakle neki organizam ima dva ista alela, taj je organizam
homozigotan za navedeni gen, dok je organizam koji ima dva različita alela
heterozigotan. Princip segregacije navodi da se od svakog roditelja nasljeđuje po
jedan alel genotipa. Tijekom S-faze mejotičke interfaze svaki kromosom se
udvostručuje producirajući dvije kopije svakog alela, po jednu na svakoj sestrinskoj
kromatidi. Tijekom anafaze I mejoze homologni kromosomi se odvajaju i to
predstavlja temelj principa segregacije (Tamarin, 2002). Ukoliko se za vrijeme
profaze I mejoze dogodio krosingover, tada dvije kromatide koje potječu od svakog
udvostručenog kromosoma nisu više identične i nastupa segregacija različitih alela
tijekom anafaze I i anafaze II.

19
Upravo metoda klasične genske analize segregacije gena bila je jedna od
polaznih metoda za utvrđivanje položaja sustava HLA unutar genoma, te za
određivanje udaljenosti između pojedinih lokusa HLA (Hartwell i sur., 2000).
Analiza segregacije nekog lokusa HLA provodi se na uzorku od 100 ili više
obitelji s minimalno 2 djece u kojima su roditelji heterozigoti na tom lokusu. Kroz više
generacija promatrano je da li je segregacija gena HLA pravilna. Pošto je
ustanovljeno da je iz generacije u generaciju očuvana uvijek ista količina nekog
gena, mnogim istraživanjima dokazano je da je segregacija gena HLA pravilna.

20
1.3. Primjena istraživanja sustava HLA
Istraživanje antigena i gena HLA ujedno je i glavno područje rada
imunogenetike upravo zbog činjenice da poznavanje i proučavanje tkivnih antigena
ima praktičnu primjenu u:
transplantacijskoj medicini
dijagnostici brojnih bolesti povezanih s genima HLA
populacijskim istraživanjima
1.3.1. Transplantacija tkiva i organa
Transplantacija je postupak prenošenja tkiva ili organa s jedne jedinke na
drugu ili s jednog mjesta na drugo unutar iste jedinke.
Transplantacija tkiva i organa u svrhu zamjene oboljelih organa u današnje
vrijeme je važna grana medicine. Najveća zapreka uspješnoj transplantaciji u većini
slučajeva je odgovor mehanizma stečene imunosti na presađeno tkivo, pošto su
glavni cilj imunosnog odgovora primatelja upravo molekule HLA eksprimirane na
površini stanica davatelja (Mahdi, 2013).
Nakon otkrića da je raspoznavanje vlastitog od tuđeg glavna odrednica za
odbacivanje transplantata, ulagalo se mnogo napora u određivanje podudarnosti
između davatelja i primatelja. Podudarnost u alelima HLA znatno poboljšava rezultate
transplantacije organa (Andreis i sur. 2004).
Kod čovjeka se od nelimfnih organa presađuje najčešće bubreg, zatim srce,
pluća, jetra, gušterača, koža i rožnica. Pri presađivanju se nastoje pronaći HLA što
podudarniji davatelj i primatelj. Kako oni gotovo nikada nisu potpuno HLA podudarni,
najčešće se razvija reakcija odbacivanja koju se nastoji potisnuti imunosupresijom
primatelja. Kod većine transplantiranih organa jednogodišnje preživljavanje
transplantata je 80-90% (Barshes i sur., 2004).
Uspjeh presađivanja bubrega poklapa se s podudarnosti davatelja i primatelja
u genima HLA. Kao uvjet za presađivanje bubrega uzima se potpuna podudarnost u
genima HLA razreda II uz što je moguće veću podudarnost u genima HLA razreda I,
a posebice se to odnosi na lokus HLA-B (Andreis i sur., 2004).

21
Presađivanje limfatičkog tkiva specifično je po tome što i sam transplantat
može reagirati protiv različitih molekula HLA jednako kao što i primatelj reagira na
različite molekule HLA davatelja. Od limfnih tkiva najčešće se presađuju krvotvorne
matične stanice. Postoje tri vrste transplantata u odnosu na razloge mogućeg
presađivanja, a to su alogenična koštana srž, autologna koštana srž ili matične
stanice izdvojene iz periferne krvi. Alogenična koštana srž presađuje se samo ukoliko
su davatelj i primatelj podudarni u genima HLA, jer razlike u genima HLA mogu
dovesti do odbacivanja transplantata, kao i do reakcije transplantata protiv primatelja
(engl. Graft versus host desease, GVHD). Optimalni davatelj krvotvornih matičnih
stanica je brat ili sestra koji je s njim HLA identičan. Za primatelje koji nemaju
davatelja u obitelji, potencijalne davatelje podudarne u alelima HLA moguće je naći i
među nesrodnim osobama zahvaljujući postojanju registra dobrovoljnih nesrodnih
davatelja. Izbor nesrodnog davatelja temelji se na podudarnosti za gene HLA lokusa
HLA-A, -B, -C, -DRB1, -DQB1 – takozvana podudarnost 10/10 (Nowak, 2008).
1.3.2. Geni HLA i bolesti
Regiju HLA povezuju s puno više bolesti nego bilo koju drugu regiju unutar
humanog genoma. Većinom se ta povezanost odnosi na autoimunosne bolesti, ali i
neke druge bolesti (tablica 3). Tako na primjer osobe koje imaju gen HLA-B*27 imaju
90 puta veću vjerojatnost za razvoj ankilozantnog spondilitisa, nego što ju imaju
osobe koje nemaju taj gen. Dakle, relativni rizik nosioca gena HLA-B*27 da oboli od
ove bolesti iznosi 90. No, postojanje povezanosti između nekih gena HLA i određenih
bolesti ne smije se interpretirati na način da je ekspresija dotičnog gena HLA
uzrokovala nastanak bolesti. Veza između gena HLA i bolesti je složena. Postoje dva
općenita objašnjenja veze između HLA i nekih bolesti (McDevitt, 1985). Prvo je
mogućnost postojanja neravnoteže udruživanja između alela točno određenog lokusa
odgovornog za nastanak bolesti i pojedinog gena HLA povezanog s tom bolesti.
Drugo moguće objašnjenje je da molekula HLA sama po sebi ima neku ulogu u
nastanku bolesti i to na način sličan jednom od predloženih modela. Prvi model
pretpostavlja da molekula HLA slabo predočuje određeni virusni ili bakterijski antigen,
dok drugi opisuje dotičnu molekulu HLA na staničnoj površini kao vezno mjesto za
virusni ili bakterijski uzročnik bolesti. Prema trećem modelu neka molekula HLA

22
omogućuje transport virusa u stanicu dok prema četvrtom modelu molekularna
podudarnost molekule HLA s patogenom dovodi do neuspješnog prepoznavanja
stranog patogena od strane domaćina. Iz svega navedenog se nameće pretpostavka
da su najvjerojatnije uključeni svi predloženi mehanizmi, ali u različitim razmjerima
kod različitih bolesti (Thorsby, 1997).
Tablica 3. Povezanost molekula HLA i određenih bolesti
(prema: Andreis i sur., 2004)
Bolest Podložna molekula HLA Relativni rizik
Ankilozantni spondilitis B27 90-100
Akutni prednji uveitis B27 10,04
Goodpastureov sindrom DR2 15,9
Multipla skleroza DR2 4,8
Autoimunosna hipertireoza DR3 3,7
Miastenija gravis DR3 2,5
Sistemni eritematozni lupus DR3 5,8
Šećerna bolest ovisna o inzulinu DR3,DR4,
heterozigoti DR3/DR4 5-25
Reumatoidni artritis DR4 4,2
Pemphigus vulgaris DR4 14,4
Hashimotoov tireoiditis DR5 3,2
1.3.3. Populacijska istraživanja
Određivanjem raspodjele antigena/gena, tj. alela HLA na reprezentativnom
uzorku nesrodnih osoba provode se populacijska istraživanja. Dosadašnja
populacijska istraživanja ukazuju na različitu zastupljenost pojedinih gena HLA u
različitim populacijama i etničkim skupinama. Pojedini aleli HLA karakteristični su za
određene populacije, a neki drugi aleli HLA prisutni su u gotovo svim do sada
istraživanim populacijama. Upravo analizom raspodjele alela HLA došlo se do
zaključka da se aleli HLA mogu podijeliti u tri skupine. Prva skupina obuhvaća alele
HLA s relativno visokom zastupljenošću unutar svih populacija kao što je npr. HLA-

23
A*02:01. Druga skupina uključuje alele koji su prisutni kod većine populacija, ali s
različitom učestalošću kao npr. HLA-A*24:02. Npr. gen HLA-A*34 prisutan je
učestalošću od 78% kod australskih Aboridžina, dok je njegova frekvencija manja od
1% kod australskih bijelaca. Treću skupinu sačinjavaju aleli HLA karakteristični samo
za pojedine populacije kakav je npr. HLA-B*42:01 karakterističan za populacije
afričkog porijekla (Shankarkumar, 2004).
Na temelju dosadašnjih istraživanja koje obuhvaćaju populacije afričkog,
europskog i azijskog porijekla, brojne populacije, izolirane populacije kao i etničke
skupine mogu se izvesti važni antropološki zaključci o smjerovima seoba
stanovništva i interakcijama između različitih populacija. Populacije vrlo sličnih
učestalosti pojedinih alela HLA imaju zajedničko porijeklo (Pierce, 2003).
Populacijskim istraživanjima sustava HLA otkriveno je da je broj HLA
homozigotnih osoba znatno manji od broja heterozigotnih osoba. Ta činjenica je
pokazatelj da je na gene HLA djelovala selekcija u korist polimorfizma i
heterozigotnosti. Proučavanje raspodjele alela HLA među različitim populacijama
dovelo je do spoznaje koje kombinacije alela HLA najčešće dolaze vezane u
haplotipove. Na osnovu toga se mogla odrediti i evolucijska stabilnost pojedinih
haplotipova.

24
2. CILJEVI ISTRAŽIVANJA
1. Analizirati rodoslovlje obitelji ispitanika i odrediti haplotipove HLA-A, -B, -DRB1
2. Istražiti učestalost krosingovera između lokusa HLA u obiteljima s dvoje i više
djece
3. Analizirati porijeklo krosingovera unutar obitelji
4. Utvrditi postoji li povezanost pojedinih haplotipova HLA s određenim
krosingoverom
5. Usporediti dobivene rezultate s podacima iz literature

25
3. MATERIJALI I METODE
3.1. Ispitanici
Za potrebe izrade ovog diplomskog rada korišteni su podaci iz registra obitelji
bolesnika mogućih primatelja krvotvornih matičnih stanica koji su u Zavod za
tipizaciju tkiva Kliničkog bolničkog centra Zagreb došli u okviru obavljanja pretrage
određivanja antigena/gena HLA.
Ukupan broj analiziranih obitelji bio je 2049, a ukupan broj djece 5936.
3.2. Metode
3.2.1. Određivanje antigena HLA
3.2.1.1. Test mikrolimfocitotoksičnosti
Dovesti u kontakt antigene HLA prisutne na membrani limfocita s protutijelima
HLA koji se nalaze u serumu je osnovni princip testa mikrolimfocitotoksičnosti
(MLCT) (Kaštelan, 1983). Od osobe kojoj ispitujemo antigene HLA uzima se mala
količina periferne krvi s heparinom i zatim se na gradijentu gustoće izdvoje limfociti.
Suspenzija limfocita T i limfocita B se pročišćava uzastopnim ispiranjem i
centrifugiranjem, te joj se podesi koncentracija. Sam test izvodi se tako da se u
svaku jažicu Terasakijeve pločice dodaje suspenzija limfocita osobe kojoj
određujemo antigene HLA. Terasakijeva pločica je prethodno pripremljena na način
da se na dno svake jažice nanose različiti antiserumi s protutijelima HLA specifičnim
za različite antigene HLA prema točno utvrđenom planu. Nakon toga slijedi inkubacija
za vrijeme koje dolazi do reakcije antigen-protutijelo u onim jažicama na pločici u
kojima su upravo oni antiserumi HLA koji sadrže protutijela HLA specifična za
antigene HLA osobe koju tipiziramo. Zatim se u svaku jažicu dodaje kunićji serum i
slijedi inkubacija. Za vrijeme druge inkubacije se na kompleks antigen-antitijelo, koji

26
je u prvoj inkubaciji nastao na membrani stanica, veže komplement iz seruma kunića
i time dolazi do lize stanica. Kako bismo ih mogli promatrati pod mikroskopom,
potrebno ih je obojiti. To se čini dodavanjem tripanskog modrila koji će mrtve lizirane
stanice obojiti plavo, dok se žive stanice neće obojiti i pod mikroskopom će biti
vidljive kao blijedožute. Iz odnosa broja živih i mrtvih stanica dobiva se rezultat testa.
Na osnovi dobivenih reakcija prema planu baterije za tipizaciju odredimo antigene
HLA osobe koju smo tipizirali. Dobiveni rezultat predstavlja fenotip HLA dotične
osobe. Ukoliko se tipiziraju i članovi obitelji, tada na temelju dobivenih tipizacija
dedukcijom možemo odrediti i genotip HLA, tj. haplotipove HLA.
3.2.2. Određivanje gena HLA
3.2.2.1. Metoda PCR-SSO
Metoda PCR-SSO (engl. Polymerase Chain Reaction – Sequence Specific
Oligonucleotides) razvijena je u cilju poboljšanja postojećih tehnika za analizu
proteina i nukleinskih kiselina. Temelji se na umnožavanju specifičnog dijela DNA uz
biotinom obilježene dimere koji se za vrijeme hibridizacije inkubiraju sa suspenzijom
mikrosfera. Mikrosfere su polistirenske kuglice veličine 5,6 mikrona čija je
unutrašnjost ispunjena crvenim ili infracrvenim fluorokromom, a na površini sadrže
vezane ili proteine ili oligonukleotidne sekvence, već prema tome za što se
primjenjuju. U našem slučaju za određivanje gena HLA na površini mikrosfera nalaze
se oligonukleotidne sekvence. Kad završi hibridizacija, reakcijskoj smjesi se dodaje
fluorescentna boja SAPE (engl. Streptovidin Phycoeritrin) koja se veže za biotinom
označeni dio DNA. Nakon hibridizacije ovako označeni dio DNA ostao je specifično
vezan s probama na određenim mikrosferama. U Luminex aparatu provodi se
očitavanje rezultata na način da aparat detektira fluorescenciju pomoću dva lasera i
očitava koje su se probe vezale na uzorak DNA. Analiza rezultata i određivanje gena
HLA u ispitivanom uzorku DNA provodi se analitičkim programom Quicktype for
Lifecodes 3.0 (IMMUCOR®).

27
3.2.2.2. Metoda PCR-SSP
Temelj metode PCR-SSP (engl. Polymerase Chain Reaction – Sequence
Specific Primers) je da se za pokretanje reakcije PCR koriste oligonukleotidne
početnice koje imaju komplementaran slijed onome koji je karakterističan za određeni
alel HLA. Tipizacijski set PCR-SSP sastoji se od određenog broja reakcija za svaki
pojedini lokus, što znači za lokuse HLA-A i -DRB1 po 24 različite reakcije, a za lokus
HLA-B 48 različitih reakcija. Svaka reakcija zasebno predstavlja jednu reakciju PCR
kojom se ispituje prisustvo jednog ili skupine alela HLA, a u svakoj reakciji su
prisutne: dvije početnice koje su specifične za pojedini alel ili za grupu alela, kontrolni
par početnica koje će nam pokazati da li smo u reakciju stavili sve potrebno, smjesa
deoksiribonukleotida (dATP, dCTP, dGTP, dTTP), komercijalni pufer, termostabilna
Taq polimeraza i genomska DNA koncentracije 50 μg/ml. Očitavanje pozitivnih
reakcija tipizacije metodom PCR-SSP izvodi se elektroforezom na agaroznom gelu
(1,5%). Naziv programa za interpretaciju rezultata je Helmberg Score SSP.
3.3. Statistička obrada podataka
Učestalost krosingovera određena je izravnim brojanjem. Upotrebom χ2 testa
utvrđene su razlike između pojedinih čimbenika. U slučaju kad je broj bio manji od 5,
korišten je Fischer-ov test. U slučaju da je vrijednost „p“ bila manja od 0,05
usporedba dva čimbenika smatrana je statistički značajnom.
Učestalost krosingovera u ispitivanoj skupini izračunali smo prema niže
navedenoj formuli:

28
4. REZULTATI
4.1. Zbirni prikaz ispitivane skupine obitelji
Ovim radom obuhvaćeno je 2049 obitelji s ukupno 5936 djece (2998 kćeri i
2938 sinova). Prosječaj broj djece po obitelji iznosio je 2,9, a omjer spolova bio je
podjednak, tj. 1,02 : 1,00 u korist kćeri (tablica 4).
Svim ispitanicima određeni su antigeni/geni HLA jednom od navedenih
metoda. Provedena je analiza rodoslovlja obitelji ispitanika i određeni su haplotipovi
HLA-A, -B i -DRB1 u obiteljima s dvoje ili više djece.
Unutar ispitivane skupine od 2049 obitelji kod njih 74 (3,61%) ustanovljeno je
prisustvo krosingovera. Kod 3 (0,15%) obitelji pronađen je krosingover kod 2 djece. U
tablici 4 također su prikazani i podaci o porijeklu rekombinantnog haplotipa.
Potrebno je naglasiti da je za istraživanja krosingovera HLA u obiteljima
neophodno da obitelji imaju najmanje dvoje djece, a tek unutar obitelji s minalno troje
djece možemo utvrditi kod koga je došlo do krosingovera, tj. koji je haplotip
rekombinantan.
Tablica 4. Zbirni prikaz podataka za ispitivanu skupinu
N %
Ukupan broj obitelji 2049
Ukupan broj djece 5936
Broj ♀ djece 2998 50,51
Broj ♂ djece 2938 49,49
Obitelji kod kojih je ustanovljen krosingover 74 3,61
Obitelji s jednim krosingoverom 71 3,47
Obitelji s dva krosingovera 3 0,15
Krosingover u oogenezi 44 57,14
Krosingover u spermatogenezi 25 32,47
Nepoznato porijeklo rekombinantnog haplotipa 8 10,39
Krosingover između lokusa HLA-A i HLA-B 37 48,05
Krosingover između lokusa HLA-B i HLA-DRB1 40 51,95
Legenda: ♀ - ženski spol, ♂- muški spol, N – broj

29
4.2. Analiza obitelji s krosingoverom nastalim u oogenezi
Unutar ispitivane skupine od 71 obitelji, kod njih 39 (54,93%) utvrđeno je
postojanje rekombinantnog haplotipa HLA naslijeđenog od majke. Od toga je kod 18
(46,15%) obitelji utvrđen krosingover između lokusa HLA-A i -B, te je kod 21
(53,85%) obitelji utvrđen krosingover između lokusa HLA-B i HLA-DRB1. Na slici 9 je
prikazano nasljeđivanje rekombinantnog majčinog haplotipa HLA između lokusa
HLA-A i -B unutar obitelji broj 29. Iz slike 9 se vidi da je kod kćeri br. 5 došlo do
rekombinacije i nastanka haplotipa HLA-A25-B5-DRB1*15.
Slika 9. Primjer rodoslovlja HLA obitelji s krosingoverom u oogenezi između lokusa
HLA-A i -B (obitelj broj 29)
Legenda: C.O. – krosingover
U obitelji broj 55, prikazanoj na slici 10, došlo je do krosingovera između
lokusa HLA-B i -DRB1. Obitelj je imala čak sedmero djece te je dedukcijom bilo
moguće odrediti kod kojeg djeteta (broj 6) je došlo do krosingovera.

30
Slika 10. Primjer rodoslovlja HLA obitelji s krosingoverom u oogenezi
između lokusa HLA-B i -DRB1 (obitelj broj 55)
Legenda: C.O. – krosingover
4.3. Analiza obitelji s krosingoverom nastalim u spermatogenezi
U ovoj skupini obitelji (N= 24, 33,8%) rekombinacija između lokusa HLA
dogodila se na haplotipovima HLA koje su djeca naslijedila od svojih očeva. Kod 11
obitelji (45,83%) krosingover se dogodio između lokusa HLA-A i -B, a kod njih 13
(54,17%) krosingover je bio između lokusa HLA-B i HLA-DRB1.
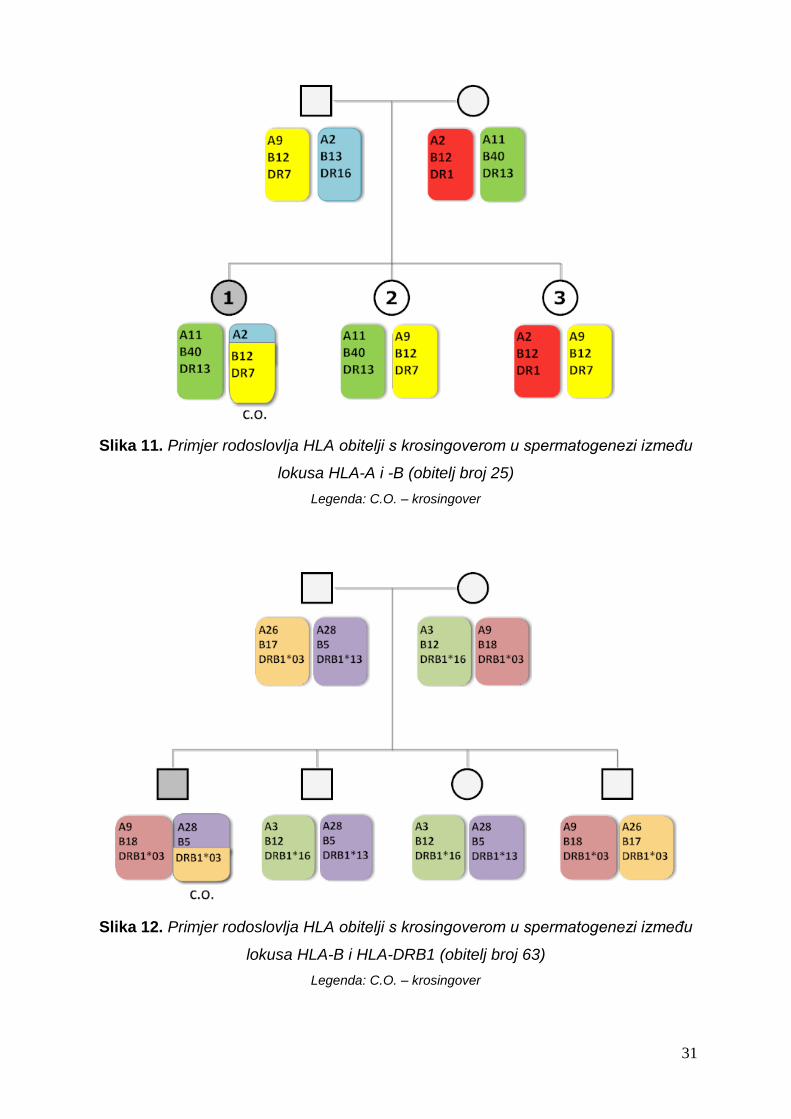
Na slici 11 prikazana je obitelj br. 25 koja je imala troje djece te smo mogli
utvrditi da se krosingover dogodio tijekom spermatogeneze i to između lokusa HLA-A
i -B. Iz slike se vidi da je dijete „1“ naslijedilo rekombinantni haplotip HLA-A2-B27-
DR7. Primjer obitelji s krosingoverom između lokusa HLA-B i -DRB1 nastao tijekom
spermatogeneze prikazan je na slici 12.
31
Slika 11. Primjer rodoslovlja HLA obitelji s krosingoverom u spermatogenezi između
lokusa HLA-A i -B (obitelj broj 25)
Legenda: C.O. – krosingover
Slika 12. Primjer rodoslovlja HLA obitelji s krosingoverom u spermatogenezi između
lokusa HLA-B i HLA-DRB1 (obitelj broj 63)
Legenda: C.O. – krosingover

32
4.4. Analiza obitelji s rekombinantnim haplotipom HLA nepoznatog
porijekla
Ovim istraživanjem obuhvaćena je i grupa od 8 (11,27%) obitelji s
krosingoverom ali kod kojih nismo mogli utvrditi porijeklo rekombinantnog haplotipa
HLA, tj. kada je nastao krosingover (tijekom oogeneze ili spermatogeneze). Naime,
riječ je bila o obiteljima bez živih roditelja gdje smo tipizirali samo braću i sestre. Kod
5 obitelji radilo se o krosingoveru između lokusa HLA-A i -B, a kod 3 obitelji je bio
utvrđen krosingover između lokusa HLA-B i -DRB1.
Primjer rodoslovlja obitelji s rekombinantnim haplotipom HLA nepoznatog
porijekla između lokusa HLA-A i -B prikazan je na slici 13, dok je primjer rodoslovlja
obitelji s rekombinantnim haplotipom HLA (krosingover između lokusa HLA-B i HLA-
DRB1) čije porijeklo nismo mogli utvrditi prikazan na slici 14.
Slika 13. Primjer rodoslovlja HLA obitelji s krosingoverom nepoznatog porijekla
između lokusa HLA-A i -B (obitelj broj 05)
Legenda: C.O. – krosingover
33
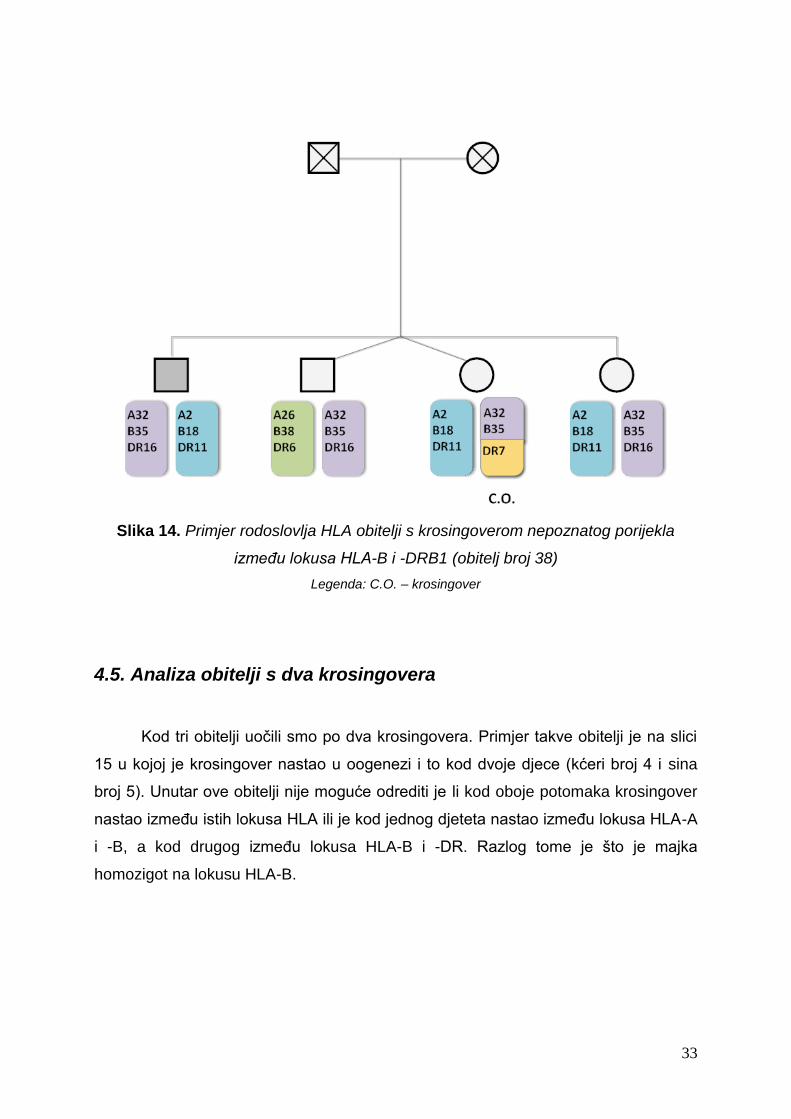
Slika 14. Primjer rodoslovlja HLA obitelji s krosingoverom nepoznatog porijekla
između lokusa HLA-B i -DRB1 (obitelj broj 38)
Legenda: C.O. – krosingover
4.5. Analiza obitelji s dva krosingovera
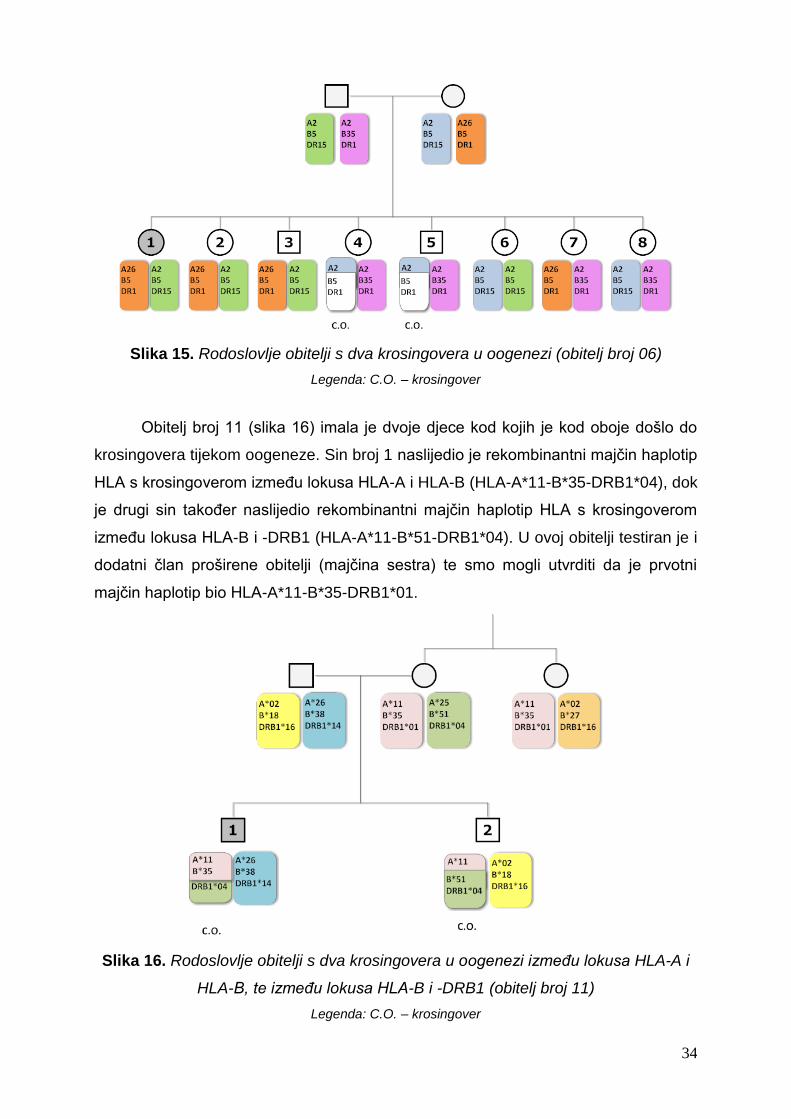
Kod tri obitelji uočili smo po dva krosingovera. Primjer takve obitelji je na slici
15 u kojoj je krosingover nastao u oogenezi i to kod dvoje djece (kćeri broj 4 i sina
broj 5). Unutar ove obitelji nije moguće odrediti je li kod oboje potomaka krosingover
nastao između istih lokusa HLA ili je kod jednog djeteta nastao između lokusa HLA-A
i -B, a kod drugog između lokusa HLA-B i -DR. Razlog tome je što je majka
homozigot na lokusu HLA-B.
34
Slika 15. Rodoslovlje obitelji s dva krosingovera u oogenezi (obitelj broj 06)
Legenda: C.O. – krosingover
Obitelj broj 11 (slika 16) imala je dvoje djece kod kojih je kod oboje došlo do
krosingovera tijekom oogeneze. Sin broj 1 naslijedio je rekombinantni majčin haplotip
HLA s krosingoverom između lokusa HLA-A i HLA-B (HLA-A*11-B*35-DRB1*04), dok
je drugi sin također naslijedio rekombinantni majčin haplotip HLA s krosingoverom
između lokusa HLA-B i -DRB1 (HLA-A*11-B*51-DRB1*04). U ovoj obitelji testiran je i
dodatni član proširene obitelji (majčina sestra) te smo mogli utvrditi da je prvotni
majčin haplotip bio HLA-A*11-B*35-DRB1*01.
Slika 16. Rodoslovlje obitelji s dva krosingovera u oogenezi između lokusa HLA-A i
HLA-B, te između lokusa HLA-B i -DRB1 (obitelj broj 11)
Legenda: C.O. – krosingover
35
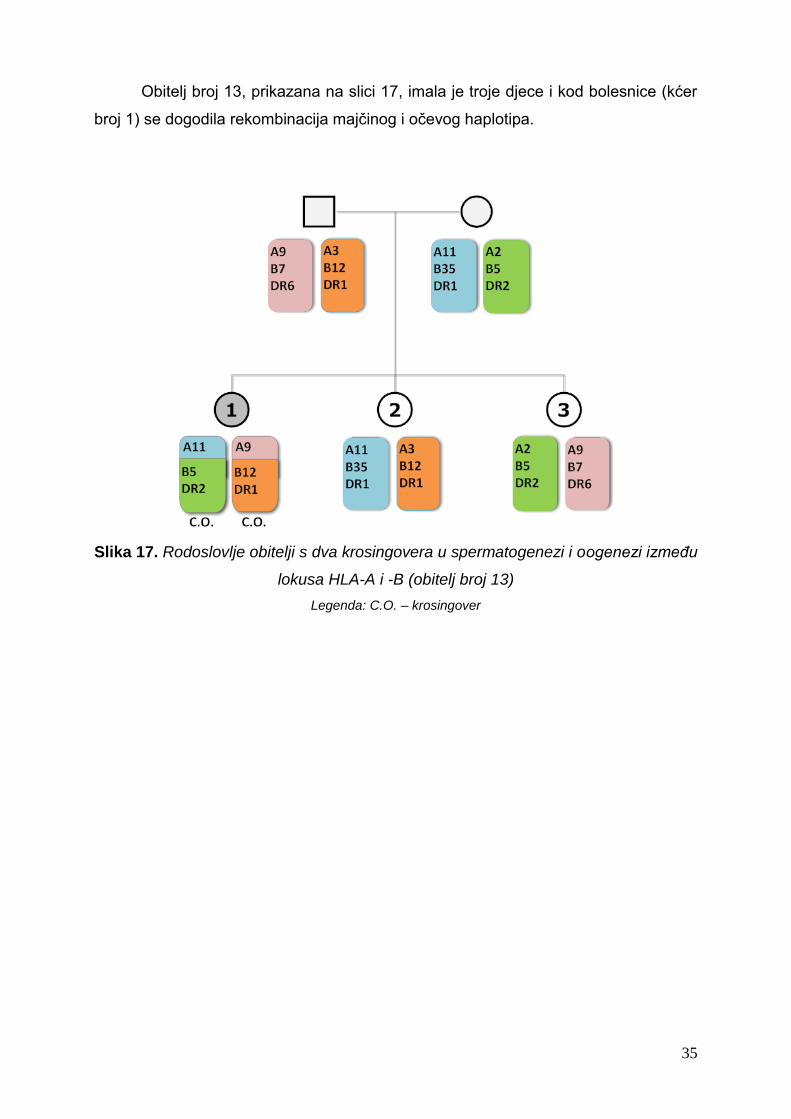
Obitelj broj 13, prikazana na slici 17, imala je troje djece i kod bolesnice (kćer
broj 1) se dogodila rekombinacija majčinog i očevog haplotipa.
Slika 17. Rodoslovlje obitelji s dva krosingovera u spermatogenezi i oogenezi između
lokusa HLA-A i -B (obitelj broj 13)
Legenda: C.O. – krosingover
36
4.6. Analiza haplotipova HLA u obiteljima s krosingoverom između
lokusa HLA-A i -B
U tablici 5 prikazani su haplotipovi HLA u 23 obitelji s krosingoverom između
lokusa HLA-A i -B kod kojih smo mogli utvrditi haplotipove HLA (obitelji s troje i više
djece). Od toga je 15 (65,22%) krosingovera bilo u oogenezi, a 8 (34,78%) u
spermatogenezi.
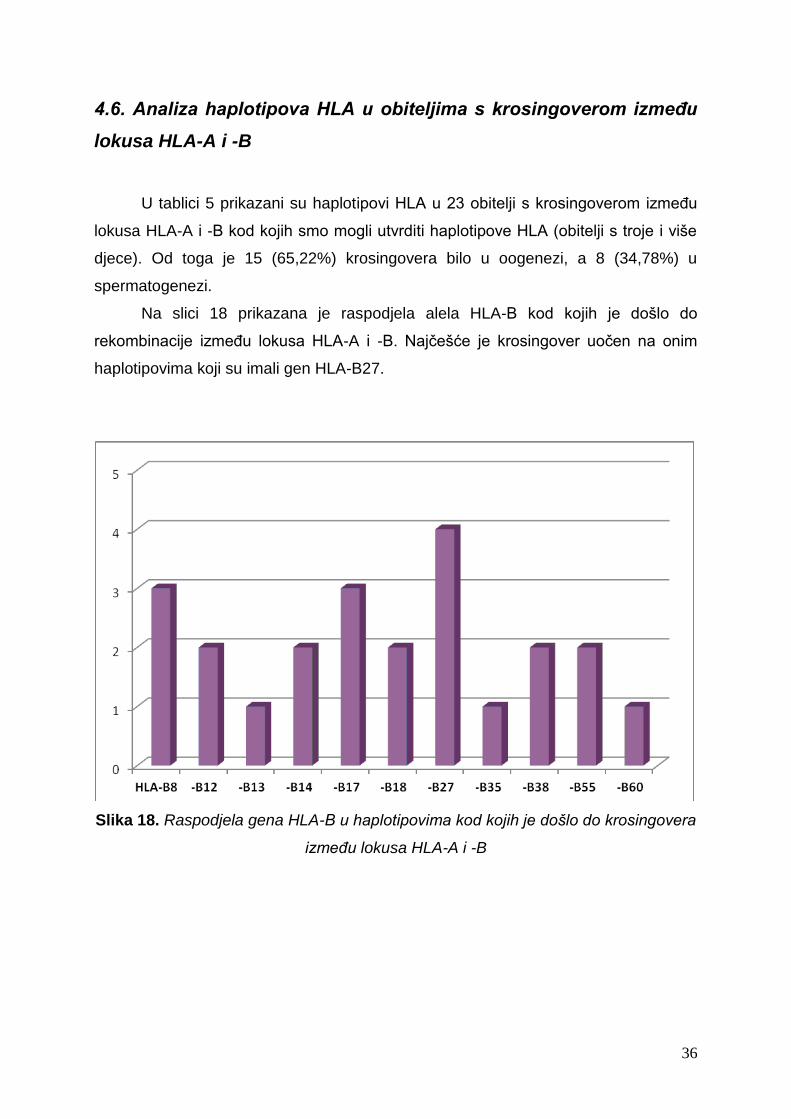
Na slici 18 prikazana je raspodjela alela HLA-B kod kojih je došlo do
rekombinacije između lokusa HLA-A i -B. Najčešće je krosingover uočen na onim
haplotipovima koji su imali gen HLA-B27.
Slika 18. Raspodjela gena HLA-B u haplotipovima kod kojih je došlo do krosingovera
između lokusa HLA-A i -B
37
Tablica 5. Prikaz haplotipova HLA u obiteljima s krosingoverom između lokusa
HLA-A i -B
Obitelj Porijeklo
rekombinantnog haplotipa HLA
1. haplotip HLA 2. haplotip HLA Rekombinantni haplotip HLA
01 majka A1-B17-DR4 A2-B35-DR11 A1-B35-DR11
03 majka A3-B17-DR7 A32-B39-DR16 A3-B39-DR16
15 majka A9-B55-DR8 A30-B35-DR2 A9-B35-DR2
16 majka A25-B27 A3-B18 A25-B18
19 majka A2-B18 A11-B38 A2-B38
20 majka A2-B55-DR2 A11-B35-DR2 A2-B35-DR2
25 otac A2-B13-DR16 A9-B12-DR7 A2-B12-DR7
27 majka A2-B12-DR16 A19-B7-DR1 A2-B7-DR1
29 majka A25-B12-DRB1*14 A2-B51-DRB1*15 A25-B51-DRB1*15
31 otac A28-B8-DRB1*03 A25-B18-DRB1*15 A28-B18-DRB1*15
33 majka A11-B18-DRB1*11 A28-B53-DRB1*14 A11-B53-DRB1*14
34 otac A24-B14-DRB1*01 A2-B35-DRB1*11 A24-B35-DRB1*11
39 otac A29-B17-DR7 A2-B27-DR15 A29-B27-DR15
41 otac A1-B8-DR3 A3-B51-DR13 A1-B51-DR13
42 majka A28-B35-DRB1*14 A2-B15-DRB1*01 A28-B15-DRB1*01
44 majka A2-B38-DR13 A26-B38-DR4 A2-B38-DR4
54 otac A2-B27-DR7 A3-B7-DR15 A2-B7-DR15
60 majka A9-B60-DRB1*13 A2-B50-DRB1*03 A9-B50-DRB1*03
61 majka A2-B38-DRB1*15 A9-B35-DRB1*13 A2-B35-DRB1*13
62 otac A26-B27-DR16 A2-B12-DR16 A26-B12-DR16
67 majka A*33-B*14-DRB1*01 A*02-B*08-DRB1*03 A*33-B*08-DRB1*03
69 otac A*31-B*08-DRB1*03 A*24-B*35-DRB1*14 A*31-B*35-DRB1*14
70 majka A*11-B*27-DRB1*07 A*03-B*27-DRB1*16 A*11-B*27-DRB1*16
38
4.7. Analiza haplotipova HLA u obiteljima s krosingoverom između
lokusa HLA-B i -DRB1
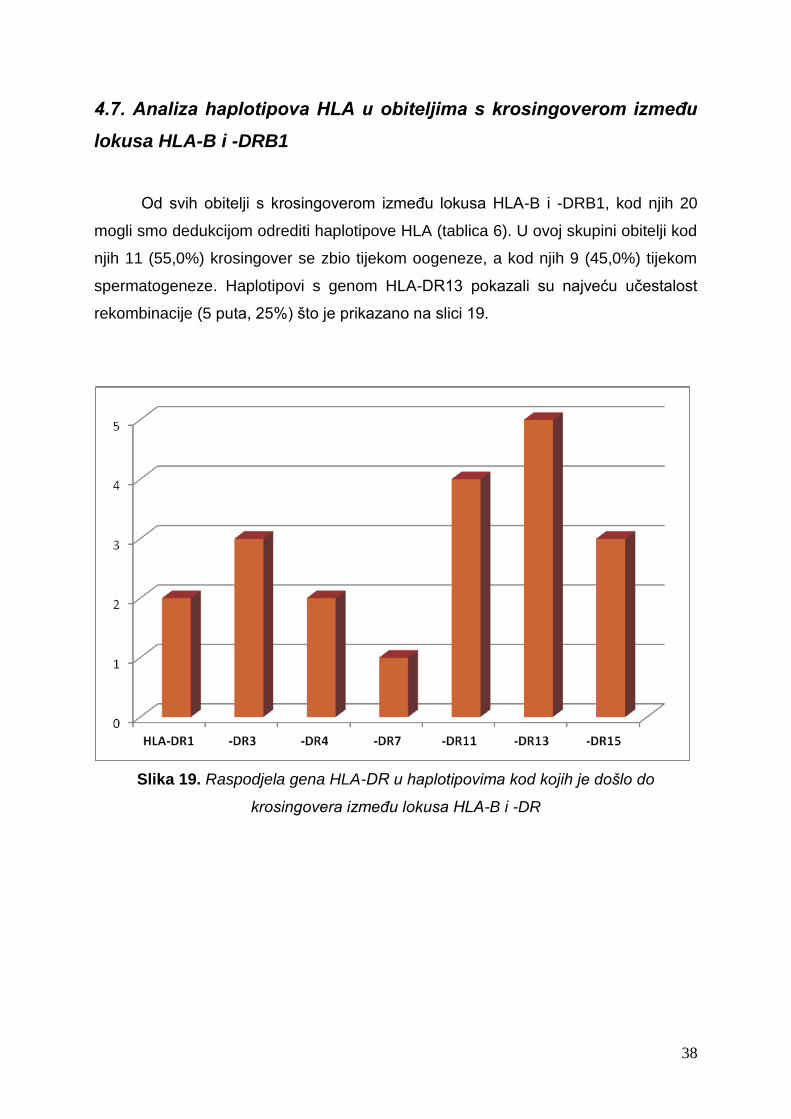
Od svih obitelji s krosingoverom između lokusa HLA-B i -DRB1, kod njih 20
mogli smo dedukcijom odrediti haplotipove HLA (tablica 6). U ovoj skupini obitelji kod
njih 11 (55,0%) krosingover se zbio tijekom oogeneze, a kod njih 9 (45,0%) tijekom
spermatogeneze. Haplotipovi s genom HLA-DR13 pokazali su najveću učestalost
rekombinacije (5 puta, 25%) što je prikazano na slici 19.
Slika 19. Raspodjela gena HLA-DR u haplotipovima kod kojih je došlo do
krosingovera između lokusa HLA-B i -DR
39
Tablica 6. Prikaz haplotipova HLA u obiteljima s krosingoverom između lokusa
HLA-B i -DRB1
Obitelj Porijeklo
rekombinantnog haplotipa HLA
1. haplotip HLA 2. haplotip HLA Rekombinantni haplotip HLA
02 majka A2-B35-DR4 A1-B22-DR2 A2-B35-DR2
09 majka A25-B12-DRB1*11:01 A3-B51-DRB1*11:04 A25-B12-DRB1*11:04
14 otac A28-B51-DRB1*13 A2-B35-DRB1*01 A28-B51-DRB1*01
18 majka A3-B21-DR3 A1-B18-DR11 A3-B21-DR11
22 otac A1-B8-DR3 A25-B8-DR2 A1-B8-DR2
30 majka A1-B8-DRB1*07 A2-B62-DRB1*04 A1-B8-DRB1*04
35 otac A1-B18-DRB1*11 A1-B57-DRB1*07 A1-B57-DRB1*11
36 otac A32-B51-DR15 A2-B35-DR13 A32-B51-DR13
40 majka A25-B18-DR15 A33-B14-DR1 A25-B18-DR1
45 majka A19.2-B35-DR11 A3-B27-DR1 A19.2-B35-DR1
46 majka A2-B21-DR1 A2-B51-DR7 A2-B21-DR7
47 otac A1-B8-DR13 A3-B7-DR15 A1-B8-DR15
50 otac A31-B51-DRB1*13 A11-B60-DRB1*04 A31-B51-DRB1*04
51 otac A1-B8-DRB1*03 A9-B18-DRB1*07 A1-B8-DRB1*07
55 majka A1-B35-DR4 A3-B27-DR16 A1-B35-DR16
56 otac A11-B35-DR1 A9-B12-DR15 A11-B35-DR15
63 otac A28-B51-DRB1*13 A26-B17-DRB1*03 A28-B51-DRB1*03
68 majka A*33-B*18-DRB1*15 A*33-B*14-DRB1*01 A*33-B*18-DRB1*01
71 majka A2-B18-DRB1*11 A -B39-DRB1*12 A*02-B*18-DRB1*12
72 majka A*02-B*15-DRB1*13 A*02-B*07-DRB1*03 A*02-B*15-DRB1*03

40
5. RASPRAVA
U ovom radu istraživana je učestalost krosingovera u uzorku od 2049 obitelji iz
registra obitelji bolesnika mogućih primatelja krvotvornih matičnih stanica. U
obiteljima je bilo ukupno 5936 djece s podjednakim omjerom spolova. Veličina
uzorka je bila dovoljna za dobivanje točnih rezultata o frekvenciji rekombinacije
unutar neke populacije što nam potvrđuju i primjeri iz literature (Thomsen i sur., 1989;
Thomsen i sur., 1994). Naime, navedeni autori su analizirali skupinu od 1332 obitelji i
utvrdili da je uzorak dovoljan za praćenje rekombinacija unutar gena HLA. Ukupan
broj rekombinacija u našem uzorku bio je 3.76% što je u skladu sa zaključkom kojeg
je naveo Carrington o učestalosti rekombinacije (<5.0%) unutar regije HLA
(Carrington, 1999). Istovremeno autor ukazuje na nužnost ovakvih istraživanja s
velikim brojem informativnih mejoza, na temelju kojih bi mogli dobiti uvid o učestalosti
rekombinacija u ovoj regiji. Iz tog razloga treba istaknuti važnost ovog istraživanja
provedenog u našoj populaciji, kojim smo po prvi put na ovako velikom uzorku obitelji
dobili podatak o rekombinacijama između lokusa HLA.
Jedan od ciljeva istraživanja bio je i analizirati rodoslovlje obitelji ispitanika.
Rodoslovlja obitelji ispitanika su analizirana na način da su u obiteljima s dvoje ili više
djece, s roditeljima ili bez njih, nakon tipizacije HLA određeni haplotipovi HLA-A, -B i
-DRB1. Za istraživanje su „vrijednije“ obitelji u kojima smo imali roditelje jer smo u
njima mogli odrediti porijeklo rekombinantnog haplotipa. Prisustvo krosingovera
utvrđeno je kod 3,61% obitelji, od toga je kod 3,47% obitelji utvrđen krosingover kod
jednog djeteta, a kod 0,15% obitelji utvrđeno je prisustvo krosingovera kod dvoje
djece. Pojava dva krosingovera unutar iste obitelji je vrlo rijetka pojava i obično ne
dolazi do krosingovera na istom mjestu unutar regije HLA. U ovom istraživanju je
unutar obitelji 06 (slika 15) utvrđeno prisustvo dva rekombinantna haplotipa HLA
porijeklom od majke s krosingoverom između lokusa HLA-B i -DRB1. S obzirom da
su ova dva lokusa HLA udaljena postavlja se pitanje je li do rekombinacije došlo na
istom mjestu tzv. „hot spot“ mjestu na kromosomu 6 ili je pak do rekombinacije došlo
na različitim mjestima. Konačan zaključak nije moguće donijeti već bi trebalo testirati
druge genske markere (npr. lokuse STR-Short Tandem Repeats ili SNP-Single
Nucleotide Polymorphism) unutar ove regije. S druge strane unutar obitelji broj 11
(slika 16) kod oboje djece je uočen krosingover ali između različitih lokusa HLA što je

41
češća pojava u literaturi (Ikeda i sur., 2015). Obitelj broj 13 (slika 17) je vrlo zanimljiva
jer je kod jednog djeteta došlo do dva krosingovera tj. i majčinog i očevog haplotipa
što je iz literaturnih navoda vrlo rijedak slučaj (Ikeda i sur., 2015).
Analiza porijekla rekombinantog haplotipa (majka ili otac) koju smo mogli
pratiti u 2041 obitelji, jer kod osam obitelji nismo imali niti jednog roditelja, pokazala je
veću učestalost rekombinacija tijekom oogeneze nego tijekom spermatogeneze
(44:25; 1,76) što je u skadu s očekivanjima i rezultatima iz literature (Strachan i
Read, 1996, Thomsen i sur. 1994). Navedeni autori su utvrdili da se krosingover
javlja 1,8 puta češće u oogenezi nego u spermatogenezi.
Naredni cilj našeg rada bio je utvrditi učestalost krosingovera između praćenih
lokusa HLA. S obzirom na međusobnu udaljenost lokusa HLA-A i -B, odnosno HLA-B
i -DRB1, na kraćem kraku kromosoma 6, bilo je za očekivati da će učestalost biti
podjednaka što je i rezultat pokazao i što je u skladu s podacima iz literature
(Carrington, 1999; Pierce 2003).
Istraživanje je također obuhvatilo i analizu haplotipova HLA na kojima je došlo
do krosingovera, odnosno postoji li povezanost određenih haplotipova HLA s
učestalošću i mjestom nastanka krosingovera. U skupini od 23 obitelji s troje ili više
djece u kojima smo uočili krosingover između lokusa HLA-A i -B kod najvećeg broja
obitelji riječ je bila o haplotipu koji je nosio antigen/gen HLA-B27. Istovremeno je
unutar skupine od 20 obitelji s krosingoverom između lokusa HLA-B i -DRB1
najčešće bio slučaj rekombinacije na haplotipovima pozitivnim za antigen/gen HLA-
DR13. Istraživanje Thomsena i sur. (1994) navodi da rekombinantni haplotipovi
između lokusa HLA-B i -DR često nose HLA-DR3 i -DR11, a HLA-DR2 (specifičnosti
HLA-DR15 i -DR16) i -DR7 su puno rjeđe prisutni kod rekombinantnih haplotipova
HLA (Thomsen i sur., 1994). Naši podaci su djelomično u skladu s navedenim radom
jer je kod nas bio prisutan krosingover na haplotipovima pozitivnim za antigen/gen
HLA-DR15 dok niti jedan haplotip pozitivan za HLA-DR16 nije bio rekombinantan.
Važno je istaknuti da je antigen/gen HLA-DR16 jedan od najčešćih u našoj populaciji
(10.4%) što ukazuje da su takvi haplotipovi HLA manje podložni rekombinaciji
(Grubic i sur., 2014). Na temelju svojih rezultata Thomsen i sur. (1994) pretpostavlja
da bi razlike u učestalosti rekombinacije ovisno o određenim alelima ili haplotipovima
HLA mogle donekle objasniti konzerviranost nekih haplotipova u europskim
populacijama (Thomsen i sur., 1994). Ostali primjeri iz literature donose informacije
da unatoč sugestijama o tome da bi učestalost krosingovera mogla varirati ovisno o

42
određenim haplotipovima kod ljudi, takav zaključak se ne može donijeti zbog velikog
broja haplotipova i zbog prirode križanja u ljudskoj vrsti koja se nikako ne može
kontrolirati (Carrington, 1999). Prema jednom istraživanju u kojem se 266 haplotipova
povezivalo s 133 krosingovera između lokusa HLA-A i HLA-B zaključeno je da
učestalost krosingovera nije ovisna o određenom haplotipu (Bender, 1982).
Kako bismo naše istraživanje mogli usporediti s ostalim istraživanjima
učestalosti krosingovera u sustavu HLA morali smo izračunati učestalost
rekombinacije između lokusa HLA-A i HLA-B, te učestalost rekombinacije između
lokusa HLA-B i HLA-DRB1 tako da podijelimo broj rekombinantnih potomaka s
ukupnim brojem potomaka. Rezultat koji smo dobili u ovom istraživanju za učestalost
krosingovera između lokusa HLA-A i HLA-B iznosi 0,61% a za učestalost
krosingovera između lokusa HLA-B i HLA-DRB1 iznosi 0,77% što se uklapa u
standard od 1% na 1x106 parova baza DNA po mejozi (Pierce, 2003). Prema
podacima iz literature (Milner i sur., 2000) frekvencija rekombinacije između lokusa
HLA-A i HLA-B je ipak nešto niža od očekivane za odsječak DNA od 1,4x106 parova
baza DNA i iznosi 0,31%. Prema istom autoru učestalost rekombinacije između
lokusa HLA-B i HLA-DRB1 iznosi 0,94%. Veća učestalost krosingovera između
lokusa HLA-A i HLA-B u našem istraživanju od one navedene u literaturi vjerojatno je
i posljedica toga što određivanje antigena/gena HLA razreda I ima dužu povijest od
određivanja antigena/gena HLA razreda II. Predmetom rasprave mogla bi biti i
priroda samog uzorka koji nije nasumično odabran iz opće populacije, već se radi o
registru obitelji mogućih primatelja krvotvornih matičnih stanica.
I na kraju možemo reći da je za donošenje konačnog zaključka ovakva
istraživanja nužno nastaviti, kako u našoj populaciji, tako i u drugim populacijama u
svijetu, te uključiti i druge lokuse HLA (HLA-C, -DQA1, -DQB1 i -DPB1).

43
6. ZAKLJUČAK
1. U svim ispitivanim obiteljima (N=2049) s dvoje ili više djece mogli smo odrediti
haplotipove HLA-A-B-DRB1.
2. Prisustvo krosingovera utvrđeno je u 74 (3,61%) obitelji, od toga je kod 71
(3,47%) obitelji utvrđen krosingover kod jednog djeteta, a kod 3 (0,15%)
obitelji je krosingover otkriven kod dvoje djece.
3. Učestalost rekombinacije između lokusa HLA-A i -B iznosila je 0,6%, a između
lokusa HLA-B i -DRB1 iznosila je 0,8% što je u skladu s očekivanom.
4. Krosingover na majčinim haplotipovima HLA uočen je u 57,14% slučajeva, na
očevim haplotipovima HLA u 32,47% slučajeva, dok za 10,39% krosingovera
nismo mogli utvrditi porijeklo.
5. Učestalost krosingovera bila je 1,8 puta češća tijekom oogeneze nego tijekom
spermatogeneze.
6. Povezanost haplotipova HLA s učestalošću i točnim mjestom nastanka
krosingovera pokazala je najveću učestalost krosingovera između lokusa
HLA-A i -B na HLA-B27 pozitivnim haplotipovima.
7. Haplotipovi s krosingoverom između lokusa HLA-B i -DRB1 najčešće su bili
HLA-DR13 pozitivni.
8. Rezultati ovog istraživanja su u skladu s rezultatima sličnih istraživanja
provedenih u drugim populacijama u svijetu.

44
7. LITERATURA
Abbas A. K., Lichtman A.H. (2004): Basic immunology. Funcions and disorders of the
immune system. W.B. Saunders, Philadelphia
Abbas A. K., Lichtman A.H. (2003): Cellular and molecular immunology. W.B.
Saunders, Philadelphia
Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walter P. (2002): Molecular
Biology of the Cell. Garland Science, New York
Ambriović Ristov A. (2007): Metode u molekularnoj biologiji. Institut Ruđer Bošković,
Zagreb
Andreis I., Batinić D., Čulo F., Grčević D., Marušić M., Taradi M., Višnjić D. (2004):
Imunologija. Medicinska naklada, Zagreb
Barshes N. R., Goodpastor S. E., Goss J. A. (2004): Pharmacologic
immunosuppression. Front Biosci 9: 411-420
Bender K. (1982): The HLA-A: HLA-B crossovers and their contribution in analyzing
possible haplotype-specific recombination rates. Hum Genet, 61: 127-134
Brown J. H. Jardetzky T. S., Gorga J. C., Stern L. J., Urban R. G., Strominger J. L.,
Wiley D. C. (1993): Three-dimensional structure of the human class II
histocompatibility antigen HLA-DR1. Nature, 364: 33-39
Carrington M. (1999): Recombination within the human MHC. Immunological
Reviews, 167: 245-256
Germain R. N. (1994): MHC-dependent antigen processing and peptide presentation:
providing ligands for T lymphocyte activation. Cell, 76: 287-299

45
Griffiths A. J. F. (2002): Modern genetic analysis: integrating genes and genomes, W.
H. Freeman and Company, New York
Grubic Z., Burek Kamenaric M., Mikulic M. i sur. (2014): HLA-A, HLA-B and HLA-
DRB1 allele and haplotype diversity among volunteer bone marrow donors from
Croatia. International Journal of Immunogenetics, 41 (3): 211-221
Goldsby R. A., Kindt T. J., Osborne B. A., Kuby J. (2003): Immunology. Freeman,
New York
Hartwell L., Hood L., Goldberg M. L., Reynolds A. E., Silver L. M., Veres R. C.
(2000): Genetics: from genes to genomes. Mc Graw-Hill, New York
Horton R., Wilming L., Rand V., Lovering R. C., Bruford E. A., Khodiyar V. K., Lush
M. J., Povey S., Talbot C. C. Jr., Wright M. W. et al. (2004): Gene map of the
extended human MHC. Nat Rev Genet, 5, 889–899
Ikeda N., Kojima H., Nishikawa M., Hayashi K., Futagami T., Tsujino T., Kusunoki
Y., Fujii N., Suegami S., Miyazaki Y., Middleton D., Tanaka H., Saji H. (2015):
Determination of HLA-A, -C, -B, -DRB1 allele and haplotype frequency in Japanese
population based on family study. Tissue Antigens, 85: 252-259
Janeway Ch. A. Jr., Travers P., Walport M., Shlomchik M. J. (2005): Immunobiology.
Garland Science Publishing, New York
Kaštelan A. (1983): Imunogenetika. U: Zergollern-Čupak, LJ. (ur.) Humana Genetika.
Zagreb, Jugoslavenska medicinska naklada, str. 173-224
Klein J. (1986): Natural History of the Major Histocompatibility Complex. U:
Srivastava R., Ram B. P., Tyle P. (ur.) Immunogenetics of the major
histocompatibility complex. John Wiley, New York
Kumnovics A., Takada T., Lindahl K. F. (2003): Genomic organization of the
mammalian MHC. Annu. Rev. Immunol., 21: 629-657

46
Lefranc M. P. (2003): IMGT, the international ImMunoGeneTics database. Nucleic
Acids Res., 31: 307-310
Madden D. R. (1995): The three-dimensional structure of peptide-MHC complexes.
Annu. Rev. Immunol., 13: 587-622
Mahdi B. M. (2013): A glow of HLA typing in organ transplantation. Clinical and
Translational Medicine, 2: 1-6
Marsh S. G. E., Albert E. D., Bodmer W. F., Bontrop R. E., Dupont B., Erlich H. A.,
Fernández-Vina M., Geraghty D. E., Holdsworth R., Hurley C. K., Lau M., Lee K. W.,
Mach B., Mayr W. R., Maiers M., Müller C. R., Parham P., Petersdorf E. W., Sasazuki
T., Strominger J. L., Svejgaard A., Terasaki P. I., Tiercy J. M., Trowsdale J. (2010):
Nomenclature for factors of the HLA system. Tissue Antigens, 75: 291-455
McDevitt H. O. (1985): The HLA system and its relation to disease. Hospital Practice,
20: 57-72
Middleton D. (2005): HLA typing from serology to sequencing era. Iranian Journal of
Allergy, Asthma and Immunology, 4: 53-66
Milner C. M., Campbell R. D., Trowsdale J. (2000): Molecular Genetics of the Human
MHC. U: Warrens A., Lechler R. (ur.) HLA in Health and Disease. Academic Press,
London
Natarajan K., Li H., Mariuzza R. A., Margulies D. H. (1999): MHC class I molecules,
structure and function. Rev. Immunogenet, 1: 32-46
Nowak J. (2008): Role of HLA in hematopoietic SCT. Bone Marrow Transplantation,
42: S71-S76
Parham P. (1988): Function and polymorphism of human leukocyte antigen-A,B,C
molecules. Am J Med, 85: 2-5

47
Pierce B. A. (2003): Genetics – a conceptual approach. W. H. Freeman and
Company, New York
Potts W. K., Slev P. R. (1995): Pathogen-based models favouring MHC genetic
diversity. Immunol Rev, 143: 181-197
Powis S. H., Trowsdale J. (1994): Human major histocompatibility complex genes.
Behring Inst Mitt 94: 17-25
Robinson J., Mistry K., McWilliam H., Lopez R., Parham P., Marsh S. G. E. (2011):
The IMGT/HLA database. Nucleic Acids Research, 39 Suppl 1: D1171-D1176
Robinson J., Halliwell J. A., Hayhurst J. H., Flicek P., Parham P., Marsh S. G. E.
(2015): The IPD and IMGT/HLA database: allele variant databases. Nucleic Acid
Research, 43: D423-431
Roitt I., Brostoff J., Male D. (2001): Immunology. Mosby, London
Shankarkumar U. (2004): The Human Leukocyte Antigen (HLA) System. Int J Hum
Genet, 4: 91-103
Strachan T., Read A. P. (1996): Human Molecular Genetics. Wiley-Liss, New York
Tamarin R. H. (2002): Principles of genetics. McGraw Hill, New York
Thomsen M., Abbal M., Neugebauer M., Cambon-Thomsen A. (1989):
Recombinations in the HLA system. Tissue Antigens, 33: 38-40
Thomsen M., Neugebauer M., Arnaud J., Borot N., Sevin A. (1994): Recombination
fractions in the HLA system based on the data set „Provinces Francaises“:
indications of haplotype-specific recombination rates. Eur J Immunogenet, 21: 33-43
Thorsby E. (2009): A short history of HLA. Tissue Antigens, 74: 101-116

48
Trowsdale J. (2001): Genetic and Functional Relationships between MHC and NK
Receptor Genes. Immunity, 15: 363-374
Yung Y. C., Yang Z., Blanchong C. A., Miller W. (2000): The human and mouse class
III region: a parade of 21 genes at the centromeric segment. Immunol Today, 21:
320-328
ELEKTRONSKI IZVORI PODATAKA:
http://hla.alleles.org/nomenclature/naming.html (23.10.2015.)
http://hla.alleles.org/inc/images/graph_hires.png (04.11.2015.)
http://hla.alleles.org/nomenclature/stats.html (04.11.2015.)
http://year12biologyatsmc.wikispaces.com/Crossing+Over (12.09.2015.)

49
8. ŽIVOTOPIS
Osnovni osobni podaci
Datum i mjesto rođenja: 6. ožujka 1975., Sisak, Republika Hrvatska
Obrazovanje
Završila sam Osnovnu školu 22. lipnja u Sisku 1989. godine.
Od 1989. do 1993. godine pohađala sam Gimnaziju Sisak, smjer jezična gimnazija.
Upisala sam studij biologije na Prirodoslovno-matematičkom fakultetu, smjer
molekularna biologija 1993. godine. Diplomirala sam 14. prosinca 2015. obranivši
diplomski rad: „Učestalost rekombinacija između gena glavnog sustava tkivne
podudarnosti u čovjeka“, pod mentorstvom prof. dr. sc. Zorane Grubić.
Zaposlenja
Kao apsolvent bila sam zaposlena 2005. godine u Osnovnoj školi Dragutina
Tadijanovića u Petrinji kao nastavnik biologije i kemije.
Od 2006. do 2015. godine bila sam zaposlena na poslovima Programa mjerenja
koncentracije peludi alergogenih biljaka u Zavodu za javno zdravstvo Sisačko-
moslavačke županije u Sisku.
Ostala znanja i vještine
Dobro poznajem češki, engleski i njemački jezik.
Posjedujem uvjerenje o usavršavanju za poslove ECDL specijaliste za uredsku
primjenu računala.
Komunikativna sam, kreativna, odgovorna.
Interesi
Aktivno sudjelujem u radu pjevačke, folklorne i kazališne skupine manjinske udruge u
kulturi Češke besede Sisak. Zastupam prava češke nacionalne manjine kao
Predstavnica češke nacionalne manjine grada Siska u drugom mandatu.