supporting information - pnas antibodies, and expression analysis. ... mogenized by mortar and...
TRANSCRIPT

Supporting InformationMoulinier Anzola et al. 10.1073/pnas.0913918107SI Materials and MethodsConstructs, Antibodies, and Expression Analysis. Floral dip was usedfor transformation of all T-DNA constructs (1). For generation ofPRZ1p::GFP:PRZ1, mGFP was cloned into the XhoI site atnucleotide +11 of genomic PRZ1 (2). 35S::myc:PRZ1 was gen-erated by transferring a myc:PRZ1 fragment from pGBP1 (2) intop235a, a derivative of pPZP211 (3). For generation of pGEX-PRZ1, we fused an EcoRI-SalI fragment ranging from nucleo-tides 859 to 1461 of the PRZ1 cDNA into pGEX4T-1. ForKRP7p::GUS, 1.8 kb immediately upstream of the KRP7 ORFwere amplified with primers 5′-GCAAACAGAGAACGCTC-AAATT and 5′-GTTTCGCTCATTCTCTCAAGCTC and thenligated into pPZP-GUS (4). For KRP7p::KRP7:GUS we usedprimers 5′-GGGGTCGACTTATCACAGTTGAAATTTAG and5′-GGGTCGACAGGTTTCAGACTAACCCACTGG for ampli-fication of the genomic KRP7 locus (promoter and coding region),which was subsequently cloned into SalI-cut pPZP-GUS (4). ForKRP7ox a KRP7 cDNA was amplified with primers 5′-GGCC-ATGGGCGAAACAAAACCCAAGAG and 5′-GGGGATCCC-TAAGGTTTCAGACTAAACC and cloned into p235a. For gen-eration of KRP antisense lines, we expressed the 3′ portion of theKRP7 coding region (nucleotides 270–588) in antisense orientationunder control of a duplicate 35S-promoter in p135a (a derivative ofpPZP112) (3).
Tissue Culture. For tissue culture experiments we used Callus-inducing medium (CIM) (5) supplemented with variable amountsof auxin: (i) CIM, containing 0.5 mg/L 2,4-D and 2 mg/L IAAand (ii) low-auxin CIM, containing 0.2 mg/L 2,4-D and 0.2 mg/LIAA. Twenty-day-old plants were scored after growth on regularCIM for 14 days.
RNAAnalysis.RNAexpression analysis was performed as described(2, 6). Poly-A+ enriched RNA was isolated with a kit fromBioNobile and first strand cDNA synthesis was performed withSuperScriptII (Invitrogen). UBQ5 and TUB9 transcript levelswere used as standards. Primer sequences are listed in Table S2.
Chromatin Analyses, ChIP.Affinity-purified anti-PRZ1 was used forprotein blots against nuclear extracts generated as described (7),with some modifications. Briefly, frozen plant material was ho-mogenized by mortar and pestle in resuspended in nuclear ex-traction buffer [2.5% (wt/vol) Ficoll 400, 5% (wt/vol) DextranT40, 0.4 M sucrose, 25 mM Tris-HCl (pH 7.4), 10 mM MgCl2,0.1% (vol/vol) β-mercaptoethanol, 1 mM PMSF, 1 μg/mL Pep-statin A, 1 μg/mL Leupeptin, 2 μg/mL; Aprotinin, 2 mM EGTA].After filtration and centrifugation the resulting pellet was re-suspended in extraction buffer and used for Western blotting.ChIP was performed as described (8, 9) with some modifi-
cations. Plant material was cross-linked in MC-buffer [10 mMKPO4 pH 7, 50 mM NaCl, 0.1% (wt/vol) sucrose, 10 mM Na-butyrate, 1% (vol/vol) formaldehyde] on ice and upon vacuumapplication for 30 min. After adding cold glycine to a final con-centration of 125 mM, nuclear proteins were extracted as de-scribed above. Sonication of nuclei was performed in 10 mMHepes/NaOH pH 7.4, 1 mM EDTA, 0.25% (wt/vol) SDS, andprotease inhibitors as in the nuclei extraction buffer (except forEGTA). After five pulses (20 s) on a sonicator (Sonic & Mate-
rials, Inc.) RIPA buffer was added and sheared chromatin wasprecleared with protein-A Sepharose. Extracts were then in-cubated with the respective antibodies (anti-histone H3, Abcam;ab1791; anti-histone H3-acK9/K14, Upstate; 06–599; anti-Myc[9E10]; anti-PRZ1; 2 μg were used for each reaction) at 4 °C for16 h. Precipitated chromatin-antibody complexes were washedseveral times in RIPA buffer, followed by elution of DNA-protein complexes in glycine buffer (100 mM Glycine, 500 mMNaCl, 0.05% (vol/vol) Tween 20, brought to pH 2.8 with HCl),After neutralization in 500mM Tris pH 9, proteinase K (Fluka,1 μg/20μg protein) was added and left o/n at 37 °C. Cross-linkingwas reverted at 68 °C for 8 h, and DNA from phenol-extractedsamples was EtOH-precipitated in the presence of glycogen(1 mg/mL).qPCR analysis was performed as described previously (10) on
a Rotor Gene 3000A. Genomic Arabidopsis DNA was used as anexternal standard for setting up calibration curves. The absoluteamount of specific DNA fragments amplified from the immuno-precipitated sample was normalized to the amount of these frag-ments in the corresponding input DNA fraction. No prominentdifferences were detected when comparing amounts of DNAfragments precipitated with histone H3 in wild-type and prz1-1samples. For determination of relative amounts of histone mod-ifications, the normalized values obtained from anti-histone H3-acK9/K14 ChIPs were divided by the normalized values obtainedfrom ChIPs performed with nondiscriminating anti-histone H3 an-tibody. Samples were run as triplicates and at least two biologicalrepetitions were performed. Primers are listed in Table S2.The position of the genomic KRP7 fragments analyzed was as
follows: Fragment 1 = nucleotides −801 to −589; fragment 2 =nucleotides −612 to –380; fragment 3 = nucleotides −442 to –
224; fragment 4 = nucleotides −212 to +2; fragment 5 = nu-cleotides −19 to +666; fragment 6 = nucleotides +5 to +838;fragment I = nucleotides −801 to −520; fragment II = nucleo-tides −442 to −202; fragment III = nucleotides 212–2; fragmentIV = nucleotides 596–838 (relative to the KRP7-Start ATG).
KRP Mutant Identification. The krp3-1 and krp7-1 alleles have beenidentified in the GABI-kat collection (krp3-1: GK-185-C07; krp7-1:GK-841D12) (11). Plants homozygous for krp7-1 were identifiedby PCR, using primers 5′-GGCCATGGGCGAAACAAAACC-CAAGAG, 5′-GCTCTGCCTGCGTCGGTGAT and 5′-ATATT-GACCATCATACTCATTGC. Homozygous krp3-1 was identi-fied by primers 5′-CGGTTAGGAGAAGAGAACGAG and 5′-CAAATCATCGTAAATCCACGAC. Krp1-1 was identified asSALK_100189 with primers 5′-GTTTCGTCGTCTTGTAGTGGand 5′-TTGAGTTGTTTCTCAGCTTCC (12). The krp2-1 muta-tion corresponds to T-DNA insertion line SALK_068815 and wasidentified by primers 5′-CGGTTAGGAGAAGAGAACGAG and5′-CAAATCATCGTAAATCCACGAC.
Histochemistry and Microscopy. GUS-staining protocols have beendescribed elsewhere (2). Lugol staining was analyzed on a ZeissAxiovert conventional microscope. Microscopy of GFP was per-formed with alive or fixed material mounted in MTSB and ana-lyzed on a Leica TCS SP2 confocal laser scanning microscope.DAPI (1 mg/mL) was added when indicated. Images were pro-cessed by Adobe Photoshop software.
1. Clough SJ, Bent AF (1998) Floral dip: A simplified method for Agro-bacterium-mediated transformation of Arabidopsis thaliana. Plant J 16:735–743.
2. Sieberer T, Hauser MT, Seifert GJ, Luschnig C (2003) PROPORZ1, a putative Arabidopsistranscriptional adaptor protein, mediates auxin and cytokinin signals in the control ofcell proliferation. Curr Biol 13:837–842.
Moulinier Anzola et al. www.pnas.org/cgi/content/short/0913918107 1 of 8

3. Hajdukiewicz P, Svab Z,Maliga P (1994) The small, versatile pPZP family ofAgrobacteriumbinary vectors for plant transformation. Plant Mol Biol 25:989–994.
4. Diener AC, et al. (2000) Sterol methyltransferase 1 controls the level of cholesterol inplants. Plant Cell 12:853–870.
5. Mathur J, Koncz C (1998) Methods in Molecular Biology, eds Martinez-Zapater J,Salinas J (Humana Press Inc., Totowa, NJ), Vol 82, pp 31–34.
6. Mladek C, Guger K, Hauser MT (2003) Identification and characterization of theARIADNE gene family in Arabidopsis. A group of putative E3 ligases. Plant Physiol 131:27–40.
7. Xia Y, Nikolau BJ, Schnable PS (1997) Developmental and hormonal regulation of thearabidopsis CER2 gene that codes for a nuclear-localized protein required for thenormal accumulation of cuticular waxes. Plant Physiol 115:925–937.
8. Turck F, Zhou A, Somssich IE (2004) Stimulus-dependent, promoter-specific binding oftranscription factor WRKY1 to its native promoter and the defense-related genePcPR1-1 in Parsley. Plant Cell 16:2573–2585.
9. Wang H, Tang W, Zhu C, Perry SE (2002) A chromatin immunoprecipitation (ChIP)approach to isolate genes regulated by AGL15, a MADS domain protein thatpreferentially accumulates in embryos. Plant J 32:831–843.
10. Karsai A, Müller S, Platz S, Hauser MT (2002) Evaluation of a homemade SYBR green Ireaction mixture for real-time PCR quantification of gene expression. Biotechniques32:790–792, 794–796.
11. Rosso MG, et al. (2003) An Arabidopsis thaliana T-DNA mutagenized population (GABI-Kat) for flanking sequence tag-based reverse genetics. Plant Mol Biol 53:247–259.
12. Alonso JM, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana.Science 301:653–657.
Fig. S1. Expression analysis of PRZ1 and KRP7 translational reporter lines. (A) PRZ1:GFP is expressed in the entire root meristem. (B) KRP7:YFP (greenpseudocoloration) is abundant in and proximal to the QC. (C) KRP7:YFP accumulating in nuclei of adjacent root epidermis cells (arrowheads). For generation ofKRP7p::KRP7:YFP, YFP was fused in frame to the 3′ end of genomic KRP7 and transformed into Col-O. (D and E) Comparison of the expression patterns ofCYCB1;1-GUS (D) and KRP7p::KRP7:GUS (E) in primary root meristems of wild-type seedlings. (F and G) Comparison of KRP7p::KRP7:GUS activity in emergingfirst true leaves of wild type (F) and prz1-1 (G). (H–K) KRP7p::KRP7:GUS activity in wild type (H and J) and in prz1-1 (I and K) on regular medium (H and I) andafter treatment with 50 μM MG132 for 16 h (J and K). (Scale bars, A, B, D, E, H–K = 100 μm; C = 10 μm; F, G = 200 μm.)
Moulinier Anzola et al. www.pnas.org/cgi/content/short/0913918107 2 of 8

Fig. S2. Quantitative analysis of transcript levels of auxin-inducible genes in 6-day-old wild type and prz1-1 after treatment with 1 μM NAA for 0, 4 or 18 h.Standard deviations are indicated (n = 3). UBQ5 and TUB9were used for normalization. (B) Schematic drawing of portions of theWOX5 and the PLT2 loci. Boxesindicate predicted exons. Gray lines correspond to the DNA fragments tested by ChIP with histone antibodies (a = −1535 to −1762; b = −125 to −426 upstream oftheWOX5 Start ATG; c = 1448 to−1210; d =−1109 to−848 upstreamof the predicted PLT2 Start ATG). (C) HistoneH3-acK9/K14 levels inwild type and prz1-1 upongrowth on regular medium. Amounts of acetylated histone H3 were determined after normalization to control ChIPs performed with nondiscriminating anti-histoneH3 (= 100%). Standard deviations are indicated as bars (n= 3). (D) Relative amounts of histoneH3K9/K14 inwild type andprz1-1 after treatmentwith 1 μMNAA for 18 h. Histone acetylation is expressed as fold changes after normalization to corresponding control samples that were not treated with auxin (= 1). Inthe wild type, an increase in the amounts of H3 K9/K14 levels was observed for promoter fragments of WOX5 (b) and of PLT2 (c and d). In prz1-1, no increasewas observedwhen testing fragments b and d,whereas analysis of fragment c indicated persistent auxin responsiveness. Standard deviations are indicated as bars(n = 3). Experiments shown are representative of at least two independent assays.
B
chrom.-PRZ1
-+
++
+-
Col prz1-1kDa
170130
100
70
55
35
A
-Colmyc:PRZ1
Colmyc:PRZ1kDa
86
47
34
26
Fig. S3. (A Upper) Western analysis of nuclear protein extracts derived from 35S::myc:PRZ1 and wild-type seedlings. Anti-myc recognizes myc:PRZ1 in nuclearextracts of 35S::myc:PRZ1 seedlings (arrowhead). (Lower) Anti-myc IP performed with chromatin preparations from Col-0 and 35S::myc:PRZ1, subsequentlyprobed with anti-PRZ1. A myc:PRZ1-specific signal is indicated (arrowhead). (B) Western analysis of nuclear protein extracts derived from wild-type and prz1-1seedlings probed with affinity-purified anti-PRZ1. Arrowhead indicates PRZ1-specific signal (an unspecific signal occurs at approximately 35 kDa. (Lower) IPperformed with affinity-purified anti-PRZ1 recognizes a specific signal in Col-0 chromatin preparations (arrowhead). No corresponding signal was observedwhen either chromatin or antibody was excluded from the IP.
Moulinier Anzola et al. www.pnas.org/cgi/content/short/0913918107 3 of 8

AKRP3KRP2
- +inp. - +inp.
PRZ1control
fold
enr
ichm
ent
10
5
15
K3K2
B
rDNAinput
ChIPw/o ab. +ab.
PRZ1control
fold
enr
ichm
ent
2
1
PRZ1::GFP:PRZ1
C
H3 acK9/K14
H3
Col-O prz1-1
C NAA C NAA
D
Fig. S4. PRZ1 is enriched in the promoter region of KRP2 and KRP3 but does not associate with rDNA (A) ChIP performed with affinity-purified anti-PRZ1 onchromatin preparations from 10-day-old Col-0 grown on PNS. Before IP the input fraction was put aside. The remaining sample was divided into aliquots, whichwere used for IP either with anti-PRZ1 (+) or by omitting the antibody (−) and subsequently used for PCR with locus-specific primers. PRZ1 interacts withpromoter fragments of KRP2 (nucleotides −491 to −15) and KRP3 (nucleotides −265 to −23). (Right) Quantification of signal intensities to determine therelative enrichment (arbitrary units) of the genomic DNA fragments. (B) ChIP and signal quantification performed as in A but tested for a fragment of the rDNAintergenic region (C) Localization of GFP:PRZ1 in fixed root meristem cells (Left), DAPI-staining (Center) and merged images (Right). Arrowhead indicatesnucleolus. (Scale bar, 5 μm.) (D) Quantification of histone H3-acK9/K14 levels in proteins extracts of wild type and prz1-1, treated with 1 μM NAA for 18 h.Chromatin was normalized with anti-histone H3 and subsequently probed with an antibody recognizing histone H3-acK9/K14. C corresponds to controls thatwere not treated with auxin. NAA corresponds to samples derived from auxin-treated seedlings.
Moulinier Anzola et al. www.pnas.org/cgi/content/short/0913918107 4 of 8
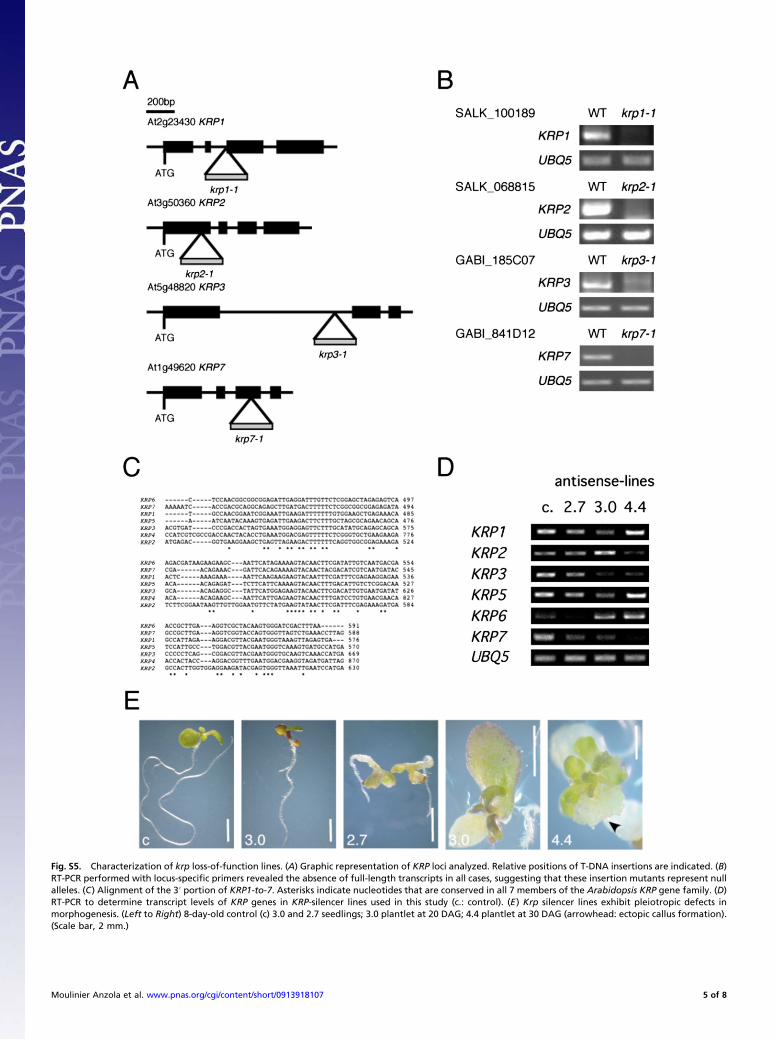
Fig. S5. Characterization of krp loss-of-function lines. (A) Graphic representation of KRP loci analyzed. Relative positions of T-DNA insertions are indicated. (B)RT-PCR performed with locus-specific primers revealed the absence of full-length transcripts in all cases, suggesting that these insertion mutants represent nullalleles. (C) Alignment of the 3′ portion of KRP1-to-7. Asterisks indicate nucleotides that are conserved in all 7 members of the Arabidopsis KRP gene family. (D)RT-PCR to determine transcript levels of KRP genes in KRP-silencer lines used in this study (c.: control). (E) Krp silencer lines exhibit pleiotropic defects inmorphogenesis. (Left to Right) 8-day-old control (c) 3.0 and 2.7 seedlings; 3.0 plantlet at 20 DAG; 4.4 plantlet at 30 DAG (arrowhead: ectopic callus formation).(Scale bar, 2 mm.)
Moulinier Anzola et al. www.pnas.org/cgi/content/short/0913918107 5 of 8

Fig. S6. Consequences of KRP7 overexpression. (A) Semiquantitative RT performed with wild-type and two independent KRP7ox lines. UBQ5 was used asa normalization control. (B) Lateral root formation in wild type, KRP7ox 3–7, and KRP7ox 3–10 at 16 DAG. The number of lateral roots was scored on 40 to 50individuals for each genotype. Standard deviations are indicated. (C) Shoot development in wild type and KRP7ox. (Top) After germination (5 DAG) KRP7oxseedlings are delayed in development and form cotyledons with enlarged cells. (Bottom) At later developmental stages (18 DAG). (D) Col-0 and KRP7ox at 36DAG. [Scale bars, C = 2.5 mm (seedlings), 5 mm (vegetative stage); D = 20 mm.]
Moulinier Anzola et al. www.pnas.org/cgi/content/short/0913918107 6 of 8

Fig. S7. Analysis of KRP7 overexpression lines on callus inducing medium. (A) Phenotypes of 20-day-old Col-0, KRP7ox, prz1-1, and prz1-1 KRP7ox after in-cubation on CIM (supplemented with 0.5 mg/L 2,4-D and 2 mg/L IAA) for 14 days. Note the less pronounced callus formation phenotype of KRP7ox and prz1-1KRP7ox. (B) Phenotypes of 20-day-old Col-0, KRP7ox, prz1-1, and prz1-1 KRP7ox after incubation on low-auxin CIM (supplemented with 0.2 mg/L 2,4-D and0.2 mg/L IAA) for 14 days. Only prz1-1 develops callus tissue (arrowheads).
Table S1. Analysis of KRP overexpression and silencer lines
Terminal differentiation of primary root meristems*
no NAA 1 μM NAA 2.5 μM NAA 5 μM NAA
PRZ1 0/87 0/73 0/60 1/44KRP7ox 0/79 1/54 7/56 12/44prz1-1 0/39 0/70 0/38 0/38prz1-1 KRP7ox 0/45 0/42 11/48 20/52
Frequency of spontaneous callus formation in krp silencer lines†
line 2.7 3.0 4.4 -Callus formingindividuals/totalnumber of plants
34/155 19/238 6/160 -
Primary root dedifferentiation in response to auxin treatment‡
no NAA 2.5 μM NAA 5 μM NAAPRZ1 0/43 - 0/40 0/38KRP7ox 0/31 - 0/45 0/42prz1-1 0/28 - 12/39 21/41prz1-1 KRP7ox 0/38 - 1/47 3/49
*Seedlings were grown on PNS and after 5 days transferred onto NAA medium for another 7 days. Meristem differentiation wasscored on fixed, Lugol stained material.†Seedlings were grown on PNS and scored for callus formation after 22 days (line 2.7) or 30 days (line 3.0 and 4.4). Callus formation wasnever observed on isogenic control plants that were not transformed with the silencer construct.‡Seeds were germinated on hormone or control medium and scored for root callus formation after 12 days. Seedlings, in which theentire root pole has dedifferentiated, were scored as positive.
Moulinier Anzola et al. www.pnas.org/cgi/content/short/0913918107 7 of 8

Table S2. Oligonucleotides used for PCR and ChIP
Primer ID Locus ID 5′-Sequence-3′
KRP2d At3g50630 CGGCTCGAGGAGAACCACAAACACGCKRP2u At3g50630 CGAAACTAGTTAATTACCTCAAGGAGG1184 At3g50630 CGGTTAGGAGAAGAGAACGAG1185 At3g50630 CAAATCATCGTAAATCCACGACKRP3d At5g48820 GATCCCGGGCGATATCAGCGTCATGGKRP3u At5g48820 GATCCCGGGTTAGTCTGTTAACTCCKRP3d2 At5g48820 CCCTCTTCGCTGATTGAACCKRP3u2 At5g48820 CCTCTGTTGCTGCTGCTCKRP7d At1g49620 GGCCATGGGCGAAACAAAACCCAAGAGKRP7Pu At1g49620 TCCAGATCTGAAAATGGAAGACKRP7u2 At1g49620 GCTCTGCCTGCGTCGGTGATWOX5cf At3g11260 AATCATAAGGCTAGGGAGAGGWOX5cr At3g11260 TGATGAGTATGGAGAAAACGACPLT2f At1g51190 CCCTCTAAGCATTCAACAACCPLT2r At1g51190 ACTCTCCACTATTCCATCCTCIAA2-5 At3g23030 ATGGCGTACGAGAAAGTCAIAA2-32 At3g23030 CCGTCCATACTCACTTTCACIAA19D At3g15540 ATGGAGAAGGAAGACTCGIAA19U2 At3g15540 TTCACATACCCTAACCCCACUBQ5d At3g62250 ACCCCTTGAGGTTGAATCATCUBQ5u At3g62250 GTCCTTCTTTCTGGTAAACGTTUB9f At4g20890 GTACCTTGAAGCTTGCTAATCCTATUB9r At4g20890 GTTCTGGACGTTCATCATCTGTTCK7d2n At1g49620 GGTCAATTATTGCTCATTATTTCACTGK7d2un At1g49620 GGATATAACTAGGCACATAGATTAGAGKU4 At1g49620 AACTTCTTAAGGATATAACTAGGCACAK7d3n At1g49620 GTATTTATAGTTCTGTCTCCATGTGK7u3n At1g49620 GCTGTAGAAATGTGAGTTGTGACAGK7d4 At1g49620 CGTGTAAAAGAATATAAGTTTAAGAAATK7u4 At1g49620 GAGTGGGGTCCTTTAAACTGAGTK7d5 At1g49620 GGACCCCACTCAAAACCGGCACK7u5 At1g49620 GCTCATTTCTCTCAAGCTCATTATAKRP7Pu1 At1g49620 CATTTCTCTCAAGCTCATTATAGKRP7Pd At1g49620 GGACCCCACTCAAAACCGGKRP7Pu At1g49620 TCCAGATCTGAAAATGGAAGACKRP7d At1g49620 GGCCATGGGCGAAACAAAACCCAAGAGKRP7u2 At1g49620 GCTCTGCCTGCGTCGGTGATKRP7d4 At1g49620 ACGGAGATGAGAGATCAGAGAAAGACKRP7u5 At1g49620 CGACCTTCAAGCGGCGTATCATTGACWOX5g1f At3g11260 GCGAGTCACCACAAATTAAAGWOX5g1r At3g11260 TGATGAAATGTCGGGAGGAGW5-15f At3g11260 TTCCAAGTAACATAAGCCACGW5-15r At3g11260 TAGATGAGACATGCACGGGPLT2g17f At1g51190 CGTACACACATGCAATGAACPLT2g17r At1g51190 CACACTCCGCAAATCATCTCPLT2g21f At1g51190 TGGCTCCTAGCACAATACGPLT2g21r At1g51190 AGTGAGACGGAAGAAAAAAGGSalboxd rDNA IGR AGGAGTGGAAATCGTCGAGSalboxu rDNA IGR CATCGCCCATTCATCGTCKRP2pd At3g50630 TTTCATTCTCGTTGATTTATTGGKRP2pu At3g50630 GCCATAATCTCCTACGACGTKRP3utrd At5g48820 GAAAAGTGAAAACACAACGGKRP3utru At5g48820 TGAGAGAGGGAAAAGAAGC
Moulinier Anzola et al. www.pnas.org/cgi/content/short/0913918107 8 of 8