subtidal littering: indirect effects on soft substratum … files/med_mar_sci_akoumianaki_et... ·...
TRANSCRIPT

Medit. Mar. Sci., 9/2, 2008, 35-52 35
Introduction
Marine litter, also known as marinedebris, is any man-made, solid wastematerial that enters the marine environ-
ment, along shorelines, coastal waters,estuaries and oceans throughout theworld. Marine litter originates either fromland sources, e.g. beach users, municipallandfills located near the coast, sewage
Mediterranean Marine ScienceVolume 9/2, 2008, 35-52
Subtidal littering: Indirect effects on soft substratum macrofauna?
I. AKOUMIANAKI1, P. KONTOLEFAS2, S. KATSANEVAKIS3,A. NICOLAIDOU2 and G. VERRIOPOULOS2
1 Hellenic Centre for Marine Research, Institute of Oceanography,P.O. Box 712, 19013, Anavissos, Attica, Hellas2 Department of Zoology – Marine Biology, University of Athens,Panepistimiopolis 15784, Athens, Hellas3Hellenic Centre for Marine Research, Institute of Marine Biological Resources, P.O. Box 712, 19013, Anavissos, Attica, Hellas
e-mail: [email protected]
Abstract
Changes in macrofauna community structure, abundance and species richness were examined bothbefore and one year after the deployment of plastic and glass bottles at littered (litter density: 16 items /100 m2) and non-littered (control) surfaces at three unimpacted coastal areas of the western SaronikosGulf (Greece). In parallel, LOI% at the adjacent sediments and changes in the composition of feedingtypes of the megaepifauna that colonized the litter were examined across treatments. Significant changesin macrofauna community structure were demonstrated between before and after littering. At only one ofthe sites was there detected a significant difference in macrofauna community structure between controland littered plots after littering. This difference was linked with a significant increase in the abundance ofopportunistic polychaete species and LOI% levels in the sediment surface due to the entrapment ofmacrophytal debris within the littered surface. The study did not show a consistent direct response ofmacroinfauna community to litter and the associated megafauna. Unlike the megafauna attracted by lit-ter items, soft-substratum macrofauna is less responsive to the addition of novel hard substrates in adja-cent sediments. Alternatively, it could be that the impact of littering with small items triggers a macrofau-na response detectable in the long-run.
Keywords: Macrofauna; Community structure; Marine litter; Coastal zone; Saronikos Gulf; Mediter-ranean Sea.
Research Article

treatment and port installations, land run-off, or from maritime sources, e.g. ships oroffshore installations (COE & ROGERS,1997). Objects ranging from householdand industrial containers, packagingmaterial, medical wastes and discardedfishing line all qualify as marine debris.The main types of marine litter impactsinclude potential health risks to inhabi-tants of coastal areas, economic losses tofisheries and tourism (NASH, 1992;WILLIAMS et al., 1993), substantial dam-age to coral reefs (DONOHUE et al.,2001; CHIAPPONE et al., 2005) andreduced fitness and increased mortality tovertebrate pelagic wildlife through entan-glement and intestinal tract blockagewhen ingested (LAIST, 1997). Specialattention is also required to the potentialthreat posed by the dispersal of alienspecies encrusted or attached on freelydrifting debris in receiving environments(WINSTON et al., 1997; BARNES, 2002).Finally, there is rising concern about theimpacts on benthic ecosystems as theseafloor from the intertidal and shallowsublittoral to outer shelf, slope and abyssaldepths has been identified as an importantsink for marine debris (GOLDBERG,1997; BACKHURST & COLE, 2000).
Plastic litter makes up to 80 % ofseabed debris, followed by metal and glass(RYAN & MOLONEY, 1990; GABRIE-LIDES et al., 1991; KANEHIRO et al.,1995; STEFATOS et al., 1999; DERRAIK,2002). The accumulation of plastic litteron the seafloor can inhibit ventilation ofthe sediments thus resulting in hypoxia oranoxia (GOLDBERG, 1997, UNEPUTTY& EVANS, 1997). Benthic plastic debrismay also provide solid attachment forspecies that would not usually occur there(RYAN & MOLONEY, 1990; MINCHIN,1996), thus acting as novel hard substrata,
often in areas that are otherwise sandy ormuddy (CHAPMAN & CLYNICK, 2006).Indeed, waste material, such as tyres andcar bodies, has been used to build artificialreefs, often specifically to attract fish andturn a ‘sand desert’ to a ‘rich in habitats’benthic environment (DAVIS et al., 1982;BOHNSACK & SUTHERLAND, 1985;GROSSMAN et al., 1997; SVANE & PE-TERSEN, 2001). Therefore, the impactscaused by large marine litter items areexpected to bear great similarities to otherman-made structures placed at the bot-tom of the sea, including artificial reefs.These impacts include alterations of wavefield and current patterns, thus causingscour and changes in sediment grain sizeand texture (MIZUTANI et al., 2000;GUIRAL et al., 1995) and entrapment ofalgae and other organic material. These,along with the activities and deaths ofreef-associated organisms, can result inorganic enrichment (WILDING, 2006)and modification of granulometry causedby the introduction of shell fragments thatderive from fouling biota (DAVIS et al.,1982; AMBROSE & ANDERSON, 1990;BAROS et al., 2001). Finally, predatorsattracted by the structures may forage onplants and animals that live in adjacentsediments (DAVIS et al., 1982; BAROS etal., 2001, FABI et al., 2002).
Smaller items such as plastic, metaland glass containers comprise a large partof benthic litter near urban centres(STEFATOS et al., 1999; BLACKHURST& COLE, 2000; MOORE & ALLEN,2000). There is little documented infor-mation about the potential use of thisform of waste as a habitat for coastal com-munities and its potential impact on adja-cent soft bottom communities (CHAPMAN& CLYNICK, 2006). In a recent manipu-lative study on the effect of benthic litter
Medit. Mar. Sci., 9/2, 2008, 35-5236

on biological communities in unimpactedshallow soft bottom areas in the SaronikosGulf (Eastern Mediterranean),KATSANEVAKIS et al. (2007) demon-strated an enhancement in the abundanceand diversity of megafauna attached todiscarded bottles, either because the litterprovided refuge or reproduction sites formobile species or because hard-substra-tum sessile species had the opportunity tosettle on the provided substrata. In paral-lel to megafauna, macrofauna abundanceand species composition were collectedfrom the adjacent sediments. The aim ofthe present study was to investigate theeffect of littering on benthic macrofaunain these recently formed small litter reefsand explore several litter-associated caus-es of impact, including changes in organicmatter content of adjacent sediments aswell as changes in the presence of epifau-nal predators.
Material and Methods
The study was conducted from June2005 to June 2006 in the Saronikos Gulf,an area of 2600 km2 and a maximum depth
450 m that receives in its northeast partthe urban effluents of Athens, untreateduntil 1995. Surface water temperaturesrange from 11ÆC in the winter up to 30ÆCin the summer. The circulation of watermasses within the gulf have been reportedto depend strongly on the local wind whilethe prevailing currents are from thenortheast to the southwest, continuinganticyclonically in the deeper layer of thewest sub-basin (NCMR, 2001).
The experiment took place at threecoves, unimpacted in terms of littering andorganic pollution, in the western SaronikosGulf, i.e. Amoni (A), Frangolimano (F),and Lychnari (L) (Fig. 1). The study siteswere away from urban centres and werecharacterized by bare sandy substrates,varying from medium to very fine sand(KATSANEVAKIS et al., 2007). At eachcove, two 100 m2 plots (10 m X 10 m), 50 mapart were defined on the seafloor withnylon line, at similar depths (16-20 m). Onone of the surfaces, 16 items of litter com-prising 12 plastic bottles and 4 glass jarswere placed uniformly after the first sam-pling, while the other surface remained‘clean’ (i.e. control). Detailed information
Medit. Mar. Sci., 9/2, 2008, 35-52 37
Fig. 1: Map of the experimental sites.

on the sampling sites and the samplingdesign is given by KATSANEVAKIS et al.(2007).
At each site, the sampling designincluded two factors: (a) ‘Plot’, fixed withtwo levels: littered plots containing litterand control plots (denoted with C) devoidof litter and (b) ‘Time’ of sampling, fixedand orthogonal to Plot with two levels:before the deployment of litter items inJune 2005, hereafter denoted with 1, andafter the deployment of litter items in June2006, hereafter denoted with 2. The time-lag of a year was proved by in situ visualobservations to be appropriate for theestablishment of a litter-associatedmegafauna. Five replicate diver-heldundisturbed cores (internal diameter: 10cm, penetration depth: 20 cm, core sam-pling area: 78.5 cm2) were taken withineach plot at each site to assess macrofaunacommunity changes between before andafter littering, and between control and lit-tered surfaces at each site. In conjunctionwith macrofauna sampling, five undis-turbed sediment cores were taken for theestimation of organic matter content. Sam-ples were taken at random within the plots.
Organic matter content of the sedi-ment was determined as loss on ignition(LOI), i.e. the difference between the dryweight (60ÆC, 24h) of the sediment andthe residue left after combustion at 450ÆCfor 3h (PARKER, 1983).
The sediment from each replicate wassieved through a 0.5 mm mesh and theresidue was immediately fixed in 4%formaldehyde. The macrofauna organ-isms retained were sorted, stored in 70%ethanol and then enumerated and identi-fied to species level. The species from allmajor taxa were classified to functionalgroups according to their food acquisitionmode, be it surface (S) or subsurface (B)
deposit feeders, suspension feeders (F),omnivorous (O) and carnivorous (C)feeders. The information used in this clas-sification was based on the ecological lit-erature for families, genera or species(WOODIN, 1976; FAUCHALD &JUMARS, 1979; RUPPERT et al., 2004;KAMERMANS, 1994; LEVINTON,1982) as well as in specific systematic keysfor the Mediterranean Sea. The specieswere also classified in three groups withrespect to their time of appearance in thesuccession process, temporal and spatialpersistence as well as rates of populationgrowth and decline, following RHOADSet al. (1978) i.e (a) Group 1 or first orderopportunistic species, (b) Group 2 or sec-ond order opportunistic species and (c)Group 3 or equilibrium species. Addition-al information for the present classifica-tion was taken from the review bySIMBOURA & ZENETOS (2002) assum-ing that similar species are eliminated, orsignificantly reduced, following any majorenvironmental disturbance.
Megafauna were censused by diversbefore and after littering at the littered andthe control plots (KATSANEVAKIS et al.,2007). In the present study, the speciescensused by KATSANEVAKIS et al.(2007), were classified to major taxa, i.e.fish, bivalves, gastropods, crustacea,cephalopods and miscellanea, includingbryozoans, tunicates, sponges and cnidari-ans and feeding and motility types. Feed-ing types included predators, suspensionand deposit feeders whereas motility typescomprised partially motile species, i.e.those restricted within a plot, highly motile– schooling species, i.e. capable of movingfurther away from the plot, sedentary andcryptic species. The information given forfeeding and motility classification wastaken by WHITEHEAD et al. (1986) for
Medit. Mar. Sci., 9/2, 2008, 35-5238

fish fauna and by RUPPERT et al. (2004)for invertebrate megafauna.
All analyses were completed at thelevel of replicate unit (n=5) at each sta-tion. Macrofauna species diversity foreach sample and the associated evennesscomponent J’ were calculated applyingthe (log2) Shannon-Wiener diversityindex (H’) (SHANNON & WEAVER,1963). Total community variables, i.e.abundance / 100 cm2 and numbers ofspecies per corer and LOI, were com-pared according to the two-factor sam-pling design using ANOVA, preceded byCochran's test for homogeneity of vari-ances and followed by a posteriori Stu-dent–Newman–Keuls (SNK) tests. Abun-dance and species numbers per macrofau-na major taxon were also tested byANOVA. The Pearson product momentcorrelation coefficients were also calculat-ed between abundance and species num-bers, and the LOI levels.
The response of the macrofauna com-munity to the two-factor mensurativesampling design was examined using per-mutational analysis of variance(PERMANOVA, ANDERSON, 2001)followed by analysis of multivariate dis-persion to test for homogeneity of disper-sions among plots and times(PERMDISP, ANDERSON, 2006). Inthe present factorial design, the evidencefor an impact (littering effect) at each siteappears as a significant ‘time’ by ‘plot’interaction (GREEN, 1979). The testswere based on 9999 unrestricted randompermutations of the raw data.
Non-metric multidimensional scaling(MDS, KRUSKAL & WISH, 1978) wascarried out to visualize multivariate pat-terns in macrofauna species data amongtreatments at each site. The effect of thespecies on the observed ordination of
treatments was visualized with projectionbiplots in which the vectors represent thePearson correlation relationship betweenmacrofauna species and the MDS axes.All multivariate analyses were obtainedusing Primer 6 for Windows (CLARKE &GORLEY, 2006) and Permanova+ forPrimer (ANDERSON & GORLEY,2007) and used Bray-Curtis dissimilaritiesthat were calculated between all pairs ofrange-standardized observations.
Results
Average LOI varied from 2.29 (1AC)to 4.82% (1LC) in spring 2005 and from3.2 (2AC) to 6.67% (2L) in spring 2006.Significant interactions between Plot andTime were only detected at L site (F1, 16 =40.3) where LOI% levels at plot 2L werehigher that at plots 1L, 1LC and 2LC.Seagrass leaf detritus derived from Posi-donia oceanica or Cymodocea nodosapatches at site L, was observed duringvisual censing one year after littering onlywithin the littered plot.
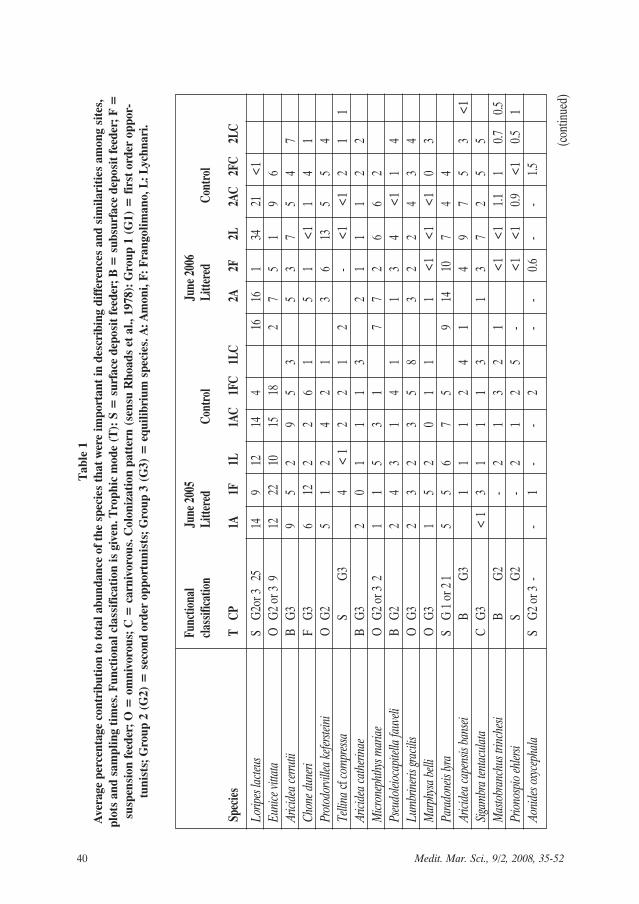
A total of 248 macrofauna species wasrecorded from the three sites, 167 werepresent at site A, 139 at site F, and 159 atsite L. Overall, 29% of the species foundin this study were common between site Aand F and F and L whereas sites A and Lshared 24% of the species. 67% of thespecies were different between before andafter littering while 41% of the specieswere common between littered and con-trol plots. At each site and plot 15 to 20%of species were represented by 1 individ-ual. The average values of total abun-dance, species richness and diversityindices at each site and plot are given inTable 1. The two most dominant speciescomprised more than 20% of total abun-dance at all treatments. A significant Plot
Medit. Mar. Sci., 9/2, 2008, 35-52 39
Medit. Mar. Sci., 9/2, 2008, 35-5240
Tab
le 1
Ave
rage
per
cent
age
cont
ribu
tion
to
tota
l abu
ndan
ce o
f the
spe
cies
tha
t w
ere
impo
rtan
t in
des
crib
ing
diff
eren
ces
and
sim
ilari
ties
am
ong
site
s,pl
ots
and
sam
plin
g ti
mes
. Fun
ctio
nal c
lass
ific
atio
n is
giv
en. T
roph
ic m
ode
(T):
S =
sur
face
dep
osit
feed
er; B
= s
ubsu
rfac
e de
posi
t fe
eder
; F =
susp
ensi
on fe
eder
; O =
om
nivo
rous
; C =
car
nivo
rous
. Col
oniz
atio
n pa
tter
n (s
ensu
Rho
ads
et a
l., 1
978)
: Gro
up 1
(G
1) =
firs
t or
der
oppo
r-tu
nist
s; G
roup
2 (
G2)
= s
econ
d or
der
oppo
rtun
ists
; Gro
up 3
(G
3) =
equ
ilibr
ium
spe
cies
. A: A
mon
i, F
: Fra
ngol
iman
o, L
: Lyc
hnar
i.
Func
tiona
lJu
ne 2
005
June
200
6cla
ssifi
catio
nLi
ttere
dCo
ntro
lLi
ttere
dCo
ntro
l
Spec
iesT
CP1A
1F1L
1AC
1FC
1LC
2A2F
2L2A
C2F
C2L
C
Lorip
es la
cteus
SG
2or 3
2514
912
144
1616
134
21<
1Eu
nice
vitta
taO
G2
or 3
912
2210
1518
27
51
96
Arici
dea
cerru
tiiB
G3
95
29
53
53
75
47
Chon
e dun
eri
FG
36
122
26
15
1<
11
41
Prot
odor
villea
kefe
rstein
iO
G2
51
24
21
36
135
54
Telli
na cf
com
pres
saS
G3
4<
12
21
2-
<1
<1
21
1Ar
icide
a ca
ther
inae
BG
32
01
11
32
11
12
2M
icron
epht
hys m
aria
eO
G2
or 3
21
15
31
77
26
62
Pseu
dolei
ocap
itella
fauv
eliB
G2
24
31
41
13
4<
11
4Lu
mbr
iner
is gr
acili
sO
G3
23
23
58
32
24
34
Mar
phys
a be
lliO
G3
15
20
11
1<
1<
1<
10
3Pa
rado
neis
lyra
SG
1 o
r 21
55
67
59
1410
74
4Ar
icide
a ca
pens
is ba
nsei
BG
31
11
24
14
97
53
<1
Siga
mbr
a ten
tacu
lata
CG
3<
13
11
13
13
72
55
Mas
tobr
anch
us tr
inch
esi
BG
2-
21
32
1<
1<
11.1
10.7
0.5Pr
iono
spio
ehler
siS
G2
-2
12
5-
<1
<1
0.9<
10.5
1Ao
nide
s oxy
ceph
ala
SG
2 or
3-
-1
--
2-
-0.6
--
1.5
(con
tinue
d)
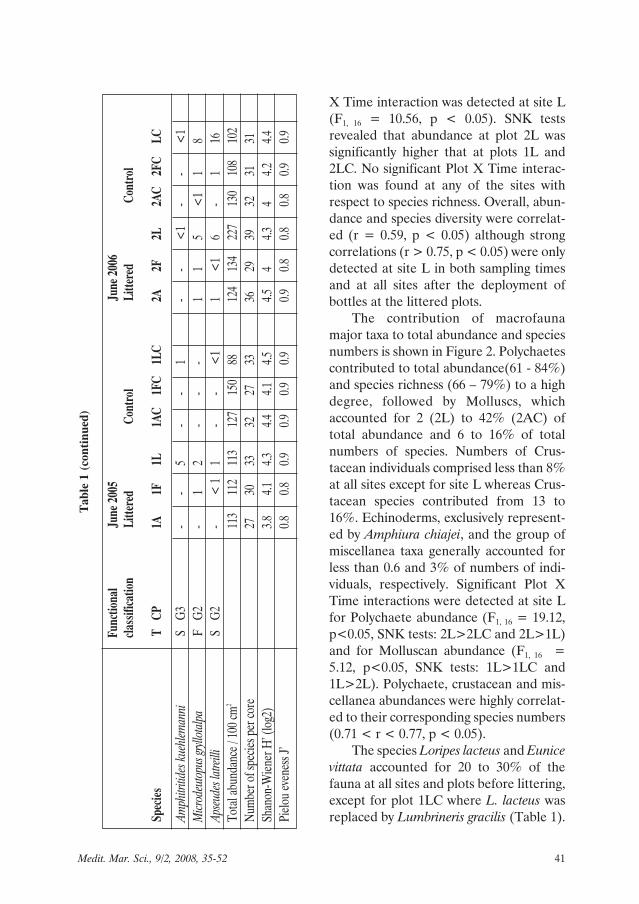
X Time interaction was detected at site L(F1, 16 = 10.56, p < 0.05). SNK testsrevealed that abundance at plot 2L wassignificantly higher that at plots 1L and2LC. No significant Plot X Time interac-tion was found at any of the sites withrespect to species richness. Overall, abun-dance and species diversity were correlat-ed (r = 0.59, p < 0.05) although strongcorrelations (r > 0.75, p < 0.05) were onlydetected at site L in both sampling timesand at all sites after the deployment ofbottles at the littered plots.
The contribution of macrofaunamajor taxa to total abundance and speciesnumbers is shown in Figure 2. Polychaetescontributed to total abundance(61 - 84%)and species richness (66 – 79%) to a highdegree, followed by Molluscs, whichaccounted for 2 (2L) to 42% (2AC) oftotal abundance and 6 to 16% of totalnumbers of species. Numbers of Crus-tacean individuals comprised less than 8%at all sites except for site L whereas Crus-tacean species contributed from 13 to16%. Echinoderms, exclusively represent-ed by Amphiura chiajei, and the group ofmiscellanea taxa generally accounted forless than 0.6 and 3% of numbers of indi-viduals, respectively. Significant Plot XTime interactions were detected at site Lfor Polychaete abundance (F1, 16 = 19.12,p<0.05, SNK tests: 2L>2LC and 2L>1L)and for Molluscan abundance (F1, 16 =5.12, p<0.05, SNK tests: 1L>1LC and1L>2L). Polychaete, crustacean and mis-cellanea abundances were highly correlat-ed to their corresponding species numbers(0.71 < r < 0.77, p < 0.05).
The species Loripes lacteus and Eunicevittata accounted for 20 to 30% of thefauna at all sites and plots before littering,except for plot 1LC where L. lacteus wasreplaced by Lumbrineris gracilis (Table 1).
Medit. Mar. Sci., 9/2, 2008, 35-52 41
Tab
le 1
(co
ntin
ued)
Func
tiona
lJu
ne 2
005
June
200
6cla
ssifi
catio
nLi
ttere
dCo
ntro
lLi
ttere
dCo
ntro
l
Spec
iesT
CP1A
1F1L
1AC
1FC
1LC
2A2F
2L2A
C2F
CLC
Amph
itriti
des k
uehl
eman
niS
G3
--
5-
-1
--
<1
--
<1
Micr
odeu
topu
s gryl
lota
lpa
FG
2-
12
--
-1
15
<1
18
Apse
udes
latre
illi
SG
2-
< 1
1-
-<
11
<1
6-
116
Tota
l abu
ndan
ce /
100
cm2
113
112
113
127
150
8812
413
422
713
010
810
2N
umbe
r of s
pecie
s per
core
2730
3332
2733
3629
3932
3131
Shan
on-W
iene
r H' (
log2
)3.8
4.14.3
4.44.1
4.54.5
44.3
44.2
4.4Pi
elou
eve
ness
J'0.8
0.80.9
0.90.9
0.90.9
0.80.8
0.80.9
0.9

After littering, Paradoneis lyra and L. lac-teus contributed from 25 to 30 % to totalabundance at sites A and F whereasProtodorvillea kefersteini and P. lyra at plot2L and Apseudes latreilli and Microdeuto-pus gryllotalpa at plot 2LC comprised 24%of total abundance (Table 1). As shown inTable 1, despite the change in the rank ofdominance between sampling times andplots, the majority of the most abundantspecies exhibited ubiquitous distributionalong the study area, with considerabledensities even when their percentages tototal abundance was low. Among thespecies that were abundant at all sites,times and plots are Prionospio ehlersi,Mastobranchus trinchesi, Aricidea catheri-nae and L. gracilis.
A significant relationship (0.4 < r < 0.5,
p < 0.05) between abundance and densi-ties of dominant species such as P. kefer-steini, P. lyra, Pseudoleiocapitella fauveliand Aricidea cerrutii and LOI% wasdetected at site L. This is in accordancewith the observation that significantlyhigher levels of LOI% at plot 2L coincid-ed with highest abundance (Fig. 3) andhigh dominance of species known to dis-play a second order opportunistic patternof colonization (Table 1).
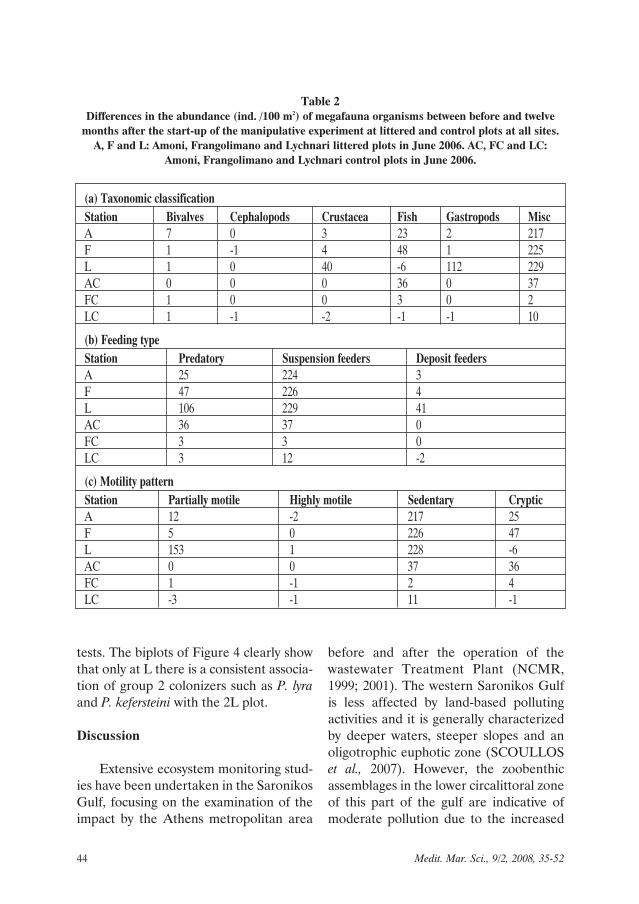
The difference in the abundance of lit-ter-associated megafauna major taxa,feeding and motility types between beforeand one year after littering is given inTable 2. The greatest changes were exhib-ited by miscellanea, mostly sedentary andfilter feeding megafauna. On the otherhand, fish and Crustacea, mostly predatory
Medit. Mar. Sci., 9/2, 2008, 35-5242
Fig. 2: Contribution of macroinfauna major taxa to (a) total abundance /100 cm-2 and (b) total speciesrichness per core sample at littered plots (1A, 1F, 1L) and control (1AC, 1FC, 1LC) plots in June 2005and at littered (2A, 2F, 2L) and control (2AC, 2FC, 2LC) plots in June 2006.

and cryptic, increased after littering at sitesA and F, although not exclusively at lit-tered plots. At site L there was a substan-tial increase in crustaceans and gas-tropods, mainly represented by predatory,cryptic and partially motile species. Sum-marising, predatory and partially motilegastropods and crustaceans mainlyincreased at site L whereas fish and crypticfauna mainly increased at sites A and F.
Permanova detected significant Plot XTime interaction only at site L (Table 3).Permdisp did not reveal any heterogeneityin multivariate dispersion among the
treatments. However, when dispersionsacross times and plots were tested at eachsite, there was detected significantly high-er dispersion before littering at site A andat the control plots at site F.
The MDS ordination plots of stationssupported the results of permanova byrevealing a great overlap in macrofaunacommunity structure among treatments ateach site A and F (Fig. 4). At site L therewas observed an effect of littering onmacrofauna community, with plot 2Lreplicates clustering separately from plot2LC, in agreement with the Permanova
Medit. Mar. Sci., 9/2, 2008, 35-52 43
Fig. 3: Relationship of (a) total number of individuals /100 cm-2 and (b) total number of species per coresample with LOI% at littered plots (1A, 1F, 1L) and control (1AC, 1FC, 1LC) plots in June 2005 andat littered (2A, 2F, 2L) and control (2AC, 2FC, 2LC) plots in June 2006.
tests. The biplots of Figure 4 clearly showthat only at L there is a consistent associa-tion of group 2 colonizers such as P. lyraand P. kefersteini with the 2L plot.
Discussion
Extensive ecosystem monitoring stud-ies have been undertaken in the SaronikosGulf, focusing on the examination of theimpact by the Athens metropolitan area
before and after the operation of thewastewater Treatment Plant (NCMR,1999; 2001). The western Saronikos Gulfis less affected by land-based pollutingactivities and it is generally characterizedby deeper waters, steeper slopes and anoligotrophic euphotic zone (SCOULLOSet al., 2007). However, the zoobenthicassemblages in the lower circalittoral zoneof this part of the gulf are indicative ofmoderate pollution due to the increased
Medit. Mar. Sci., 9/2, 2008, 35-5244
Table 2Differences in the abundance (ind. /100 m2) of megafauna organisms between before and twelve
months after the start-up of the manipulative experiment at littered and control plots at all sites.A, F and L: Amoni, Frangolimano and Lychnari littered plots in June 2006. AC, FC and LC:
Amoni, Frangolimano and Lychnari control plots in June 2006.
(a) Taxonomic classificationStation Bivalves Cephalopods Crustacea Fish Gastropods MiscA 7 0 3 23 2 217F 1 -1 4 48 1 225L 1 0 40 -6 112 229AC 0 0 0 36 0 37FC 1 0 0 3 0 2LC 1 -1 -2 -1 -1 10
(b) Feeding typeStation Predatory Suspension feeders Deposit feedersA 25 224 3F 47 226 4L 106 229 41AC 36 37 0FC 3 3 0LC 3 12 -2
(c) Motility patternStation Partially motile Highly motile Sedentary CrypticA 12 -2 217 25F 5 0 226 47L 153 1 228 -6AC 0 0 37 36FC 1 -1 2 4LC -3 -1 11 -1

organic load and nutrients transported bythe prevailing westward currents from theinner Saronikos Gulf (NCMR, 1999).
On the other hand, the sandy infralit-toral benthic communities of the westernpart were examined for the first time dur-ing the course of this study. The speciescomposition at the three study sites, A, Fand L, indicated the co-existence of taxafound at a variety of infralittoral and cir-calittoral biocoenoses as described byPERES (1967). These biocoenosesinclude the ‘coastal detritic’ community(DC) and the communities of sands ormuddy sands in shallow areas protectedagainst wave action and currents (SRPVand SVMC, respectively), and other sub-littoral communities described at unpol-luted heterogeneous sediments in coastalareas of the western and central Mediter-ranean Sea (DRAGO & ALBERTELLI,1978; SIMONINI et al., 2004; COSE-
NTINO & GIACOBBE, 2006). Thisresemblance is consistent with the obser-vation that the sediments across the studyarea varied from fine to medium sandsand remained highly heterogeneous, asshown by the poor sorting coefficient ofgrain size (KATSANEVAKIS et al.,2007), thus allowing for a great variety ofspecies, with no specific preference ingrain size, to settle and establish sizeablepopulations. This finding is in agreementwith the high temporal and spatial disper-sion at sites A and F.
Marine litter densities on the sea floorof the Saronikos Gulf range from 0.4 to 25items / 100 m2, higher along the northeastcoastline (KATSANEVAKIS & KATSA-ROU, 2004). This is the first study withrespect to the impact of littering on theinfauna in the Saronikos Gulf. Thehypothesis of indirect impact of seafloorlittering upon macrofauna community
Medit. Mar. Sci., 9/2, 2008, 35-52 45
Table 3Testing the effect of plots and sampling time and their interactions on macrofauna communities
at the three study sites using permutational analysis of variance (permanova) and dispersion (permdisp). df: degrees of freedom, perm: permutation, ns: non significant,
*: p < 0.05, **: p < 0.01, ***: p < 0.001. BRAY – CURTIS distance measure was used.
(a) PERMANOVA tests
Amoni Frangolimano LychnariSource of variation df Pseudo-F P(perm) Pseudo-F P(perm) Pseudo-F P(perm)
Time (Ti) 1 3.334 ** 4.304 *** 7.904 ***Plot (Pl) 1 1.492 ns 0.976 ns 2.127 *TixPl 1 1.812 ns 1.206 ns 2.257 *Resisual 16Total 19
(b) PERMDISP tests
Amoni Frangolimano LychnariGroup factor F1, 18 P(perm) F1, 18 P(perm) F1, 18 P(perm)
Time (Ti) 7.777 * 0.784 ns 0.296 nsPlot (Pl) 1.838 ns 10.345 ** 0.002 ns

structure due to attraction of predatoryorganisms, previously absent or at signifi-cantly lower densities, stated byKATSANEVAKIS et al. (2007), predicts
that macroinfauna abundance and speciescomposition will be substantially limitedwithin the littered plots. By contrast, anincrease from June 2005 to June 2006 by
Medit. Mar. Sci., 9/2, 2008, 35-5246
Fig. 4: Euclidean MDS biplot showing the Pearson correlation coefficients of macrofauna species den-sities with MDS axes at each site. Non-metric multidimensional scaling was applied on the range stan-dardized densities of macrofauna species using BRAY-CURTIS similarities. At each biplot prefixes 1and 2 for each sample code denote June 2005 and June 2006, respectively, while the suffix C denotescontrol plot. A: Amoni, F: Frangolimano, L: Lychnari.

106 individuals / 100 m2 in predatorymegafauna at site L coincided with a sig-nificant density increase in macrofauna,especially polychaetes and crustacea.These taxa are included in the prey ofmany infralittoral schooling and crypticfish and decapods (CASTRIOTA et al.,2005; RELLINI et al., 2002) as well as ofthe predatory muricid gastropodHexaplex trunculus (MORTON et al.,2007), which was one of the most abun-dant predators at the littered plots(KATSANEVAKIS et al., 2007). Con-versely, there was observed a significantreduction of macrofaunal molluscs at thissite, which may indicate that bivalves mayhave belonged among the preferred preyof the predatory colonizers of litter. How-ever, molluscs simultaneously increased,not only at the corresponding control plot(2LC) but also at the littered and controlplots at sites A and F, indicating thatchanges in the density of molluscs cannotbe always associated with predatorsattracted by litter. Finally, the noteworthyfilter feeding community that was estab-lished using the litter items as substrates isunlikely to have negatively interacted withmacroinfauna in the adjacent sediments,even though a lot of soft-bottom speciesfound, including those that dominated theinfauna, follow pelagic larval development(RUPPERT et al., 2004).
Regarding organic matter (OM) lev-els at the surface sediments of the studysites, these were elevated for sandy bot-toms. OM values after littering were onaverage 1.2 times higher than before atboth littered and clean surfaces. However,only at site L did these levels exceed 5% atthe littered plots. As pointed out byPEARSON & ROSENBERG (1978) dis-turbances associated with OM enrichmentand the resulting hypoxic conditions in the
sediment leave carnivorous and generallypredatory macrofauna species unaffected,while they affect tolerant subsurface andsurface deposit feeders positively. Con-versely, suspension feeders and speciestypical of later stages of succession in soft-bottom communities are gradually elimi-nated, thus resulting in low diversity andhigh dominance values. At the study areafilter feeding, omnivory-carnivory and sur-face and subsurface deposit feeding werewell represented among the most abun-dant species, implying high trophic com-plexity. Therefore, in terms of trophicstructure, the macrofauna community inthe study area resembled other sandy sub-littoral communities of the MediterraneanSea not directly influenced by eutrophica-tion, exhibiting a wide range of trophicethological habits (GAMBI & GIAN-GRANDE, 1985; CARDELL et al., 1999;SIMONINI et al., 2004).
However, species not affected byalterations and species known to takeadvantage under conditions of environ-mental stress by means of having a shortlife-span, rapid growth and many genera-tions throughout the year, were also pres-ent. These belonged mainly to polychaetesand crustaceans, and exhibited, as expect-ed, considerable variations. Despite thehigh dominance levels of the two mostabundant species (10 to 25% before and15 to 35% after littering), these comprisedgroup 3 colonizers before littering. On theother hand, after littering the contributionand the ranking of the group 2 colonizerssuch as P. kefersteini, A. latreilli, P. lyra andM. gryllotalpa increased at the litteredplots and especially at site L. Simultane-ously, L. lacteus, a species typical of laterstages of succession, remained the mostdominant species at sites A and F whileother bivalves that reach high abundances
Medit. Mar. Sci., 9/2, 2008, 35-52 47

in disturbed sediments, such Corbulagibba, Thyasira flexuosa, Mysella bidentatahad a very low density throughout thestudy. Although the group 2 colonizerswere represented by elevated numbers atlittered and control plots in both samplingtimes, their enhancement after littering atlittered plots, especially at site L, raisesquestions as to whether this is an indica-tion of stress triggered by littering.
Since organic matter levels at sites Aand F were not indicative of enrichmentbut significantly higher LOI% levels atsite L after littering coincided with densemacrophyte detrital cover and the pre-dominance of second order opportunisticspecies, it is reasonable to assume that themacrofauna community was impacted bya reduction in sediment oxygenationcaused by the entrapment and subsequentaccumulation of seagrass detritus. Never-theless, the accumulation started takingplace only two months before the end ofthe manipulative experiment, i.e. June2006, thus indicating that changes detect-ed in this study could be the first stages ofa gradual organic enrichment process. It isnoteworthy that comparable LOI% valueswere recorded in the sediments adjacentto artificial reefs that trapped macroalgalphytodetritus at their periphery (WILDING,2006). The accumulation of phytodetritalmaterial for longer periods is expected tocause effects similar to that of localizedorganic enrichment (GRAY et al., 2002).As the phytodetrital mats significantlyreduce water movement over the sedi-ment, the bottom water in the mats isoften stagnant, resulting in a permanentlyanoxic sediment surface. It is possible thatthe marked increase of P. kefersteini at Llittered plots, where there was a pro-nounced accumulation of seagrass debris,is an indication of the hypoxia caused by
the slow decomposition rates of seagrassleaves. Apart from the nature of thedeposited material, the extent of theimpact of the accumulation of phytodetri-tus is proportional to a number of factors,including decreases in current velocityaround the litter (WILDING, 2006).Given the small volume of the litter itemsin the present study, impacts related tochanges of hydrodynamic regime aroundthe marine litter may take longer than ayear (i.e. the duration of the manipulativeexperiment of the present study) to bedetectable. The entrapment of phyto-detritus and subsequent changes in sedi-ment oxygenation around artificial reefs isa side-effect of littering on macrofauna ofadjacent sediments, as the entrapment ofphytodetritus was facilitated by the deploy-ment of litter and requires a longer moni-toring for an adequate evaluation of theimpact on soft-bottom macrozoobenthos.
It has been estimated that up to 70%of the marine litter that enters the seaends up on the sea bed (UNEP, 2005).The studies indicating that discardedwaste material, in the form of pieces ofmetal, glass, plastic, tyre and wood doesprovide usable habitat for subtidal inver-tebrates and fish either in areas wherenatural habitat is lost or even in non-pol-luted areas are gradually increasing(CHAPMAN & CLYNICK, 2006; KA-TSANEVAKIS et al., 2007). The lack ofsignificant differences between litteredand control plots at sites A and F, pre-clude any generalization about the effectof littering on macrofauna. Logisticrestrictions did not allow for several sam-pling times before and several samplingtimes after littering and adequate withinsite replication of plots, so as to estimatethe range of natural fluctuations of macro-fauna populations in the area and elimi-
Medit. Mar. Sci., 9/2, 2008, 35-5248

nate effects of short-term temporal varia-tion. However, the present study demon-strated the increase of second orderopportunistic macrofauna species adja-cent to marine litter sediments in thepresence of seagrasses trapped within thelittered plots. Further manipulative stud-ies, with extensive temporal replication ata variety of receiving environments with avariety of waste materials are required soas to evaluate the full array of impacts ofmarine litter at sublittoral areas.
Acknowledgements
The project was co-funded by theEuropean Social fund and NationalResources (Hellenic Ministry of NationalEducation and Religious Affairs:EPEAEK II: Pythagoras II). We wouldlike to thank E. Kyriakou for her help inmacrofauna sorting.
References
AMBROSE, R.F. & ANDERSON, T.W.,1990. Influence of an artificial reef onthe surrounding infaunal community.Marine Biology 107: 41-52.
ANDERSON, M.J., 2001. A new methodfor non-parametric multivariateanalysis of variance. Austral Ecology26: 32-46.
ANDERSON, M.J., 2006. Distance-basedtests for homogeneity of multivariatedispersions. Biometrics 62: 245-253.
ANDERSON, M.J. & GORLEY, R.N.,2007. Permanova+ for Primer: Guideto statistical methods. Primer-E, Ply-mouth, UK.
ANDRADY, A., 1987. Research on theuse of degradable fishing gear andpackaging materials. NWAFC Pro-cessed Rep. 87-03, National Marine
and Fisheries Service, NOAA, 49 p. BACKHURST, M.K. & COLE, R.G.,
2000. Subtidal benthic marine litter atKawau Island, north-eastern NewZealand. Journal of EnvironmentalManagement 6: 227-237.
BARNES, D.K.A., 2002. Invasions bymarine life on plastic debris. Nature416: 808-809.
BOHNSACK, J.A. & SUTHERLAND,D.L., 1985. Artificial reef research: areview with recommendations forfuture priorities. Bulletin of MarineScience 3: 11-39.
CARDELL, M.J., SARDA R. & ROME-RO, J., 1999. Spatial changes in sublit-toral soft bottom polychaete assem-blages due to river inputs and sewagedischarges. Acta Oecologia 20: 343-351.
CASTRIOTA, L., SCARABELLO, M.P.,FINOIA M.G., SINIPOLI, M. &ANDALORO, F., 2005. Food andfeeding habits of pearly razorfish,Xyrichthys novacula (Linnaeus, 1758)in the southern Tyrrhenian Sea: varia-tion by sex and size. Environmentalbiology of Fish 72: 123-133.
CHAPMAN, M.G. & CLYNICK, B.G.,2006. Experiments testing the use ofwaste material in estuaries as habitatfor subtidal organisms. Journal ofExperimental Marine Biology and Ecol-ogy 338: 164-178.
CHIAPPONE, M., DIENS, H.,SWANSON, D.W. & MILLER, S.L.,2005. Impacts of lost fishing gear oncoral reef sessile invertebrates in theFlorida Keys National Marine Sanctua-ry. Biological Conservation 121: 221-230.
CLARKE, K.R. & GORLEY, R.N., 2006.Primer v6. Primer-E, Plymouth, UK.
COE, J.M. & ROGERS, D.B. (eds.),1997. Marine Debris: Sources,Impacts and Solutions. New York:
Medit. Mar. Sci., 9/2, 2008, 35-52 49

Springer-Verlag.COSENTINO, A. & GIACOBBE, S.,
2006. A case study of mollusc andpolychaete soft-bottom assemblagessubmitted to sedimentary instability inthe Mediterranean Sea. Marine Ecol-ogy 27: 170-183.
DAVIS, N., VAN BLARICOM, G.R. &DAYTON, P.K., 1982. Man-madestructures on marine sediments:effects on adjacent benthic communi-ties. Marine Biology 70, 295-303.
DERRAIK, J.G.B., 2002. The pollution ofthe marine environment by plasticdebris: a review. Marine Pollution Bul-letin 44: 842-852.
DONOHUE, M.J., BOLAND, R.C., SRA-MEK, C.M. & ANTONELIS, G., 2001.Derelict fishing gear in the Northwest-ern Hawaiian Islands: diving surveysand debris removal in 1999 confirmthreat to coral reef ecosystems. MarinePollution Bulletin 42: 1301-1312.
DRAGO, N. & ALBERTELLI, G., 1978.Etude faunistique et bionomique dulittoral de Cogoleto (golfe de Gênes).Tethys 8: 203-212.
FABI, G., LUCARINI, F., PANFILI, M.,SOLUSTRI, C. & SPANGOLO, A.,2002. Effects of an artificial reef onthe surrounding soft-bottom commu-nity (central Adriatic Sea). ICES Jour-nal of Marine Science 59: 343-349.
FAUCHALD, K. & JUMARS, P.A.,1979. The diet of worms: a study ofpolychaete feeding guilds. Oceanogra-phy and Marine Biology: an annualreview 17: 193-284.
GABRIELIDES, G.P., GOLIK, A.,LOIZIDES, L., MARINO, M.G.,BINGEL, F. & TORREGROSSA,M.V., 1991. Man made garbage pollu-tion on the Mediterranean coastline.Marine Pollution Bulletin 23: 437-441.
GAMBI, M.C. & GIANGRANDE, A.,1985. Caratterizzazione e dis-tribuzione delle categorie trofiche deiPolicheti nei fondi mobile del golfo diSalerno. Oebalia 11: 223-240.
GOLDBERG, E.D., 1997. Plasticizing thesea-floor: an overview. EnvironmentalTechnology 18: 195-202.
GRAY, J.S., WU, R.S.S. & OR, Y.Y.,2002. Effects of hypoxia and organicenrichment on the coastal environ-ment. Marine Ecology Progress Series238: 249-279.
GREEN, R.H., 1979. Sampling designand statistical methods for environ-mental biologists. Wiley, New York.
GROSSMAN, G.D., JONES, G.P.,SEAMAN, W.J.J., 1997. Do artificialreefs increase regional fish produc-tion? A review of existing data. Fish-eries 22: 17-27.
GUIRAL, D., GOURBAULT, N.,HELLEOUET, M.N., 1995. Sedimentnature and meiobenthos of an artificialreef (Acadja) used for extensive aqua-culture. Oceanologica Acta 18: 543-555.
KAMERMANS, P., 1994. Similarity infood source and timing of feeding indeposit – and suspension feedingbivalves. Marine Ecology ProgressSeries 104: 63-75.
KANEHIRO, H., TOKAI, T.,MATUDA, K., 1995. Marine littercomposition and distribution on theseabed of Tokyo Bay. Fisheries Engi-neering 31: 195-199.
KATSANEVAKIS, S., KATSAROU, A.2004. Influences on the distribution ofmarine debris on the seafloor of shal-low coastal areas in Greece (EasternMediterranean). Water, Air and SoilPollution 159: 325-337.
KATSANEVAKIS, S., VERRIO-POULOS, G., NICOLAIDOU, A. &
Medit. Mar. Sci., 9/2, 2008, 35-5250

THESSALOU-LEGAKI, M. 2007.Effect of marine litter on the benthicmegafauna of coastal soft bottoms: Amanipulative experiment. Marine Pol-lution Bulletin 54: 771-778.
KRUSKAL, J.B. & WISH, M., 1978. Mul-tidimensional scaling. Sage Publica-tions, Beverley Hills.
LAIST, D.W., 1997. Impact of marinedebris: entanglement of marine life inmarine debris including a comprehen-sive list with entanglement and inges-tion records. In: Coe, J.M., Rogers,D.B. (Eds.), Marine debris – sources,impacts and solutions. New York,NY: Springer; pp. 99–139.
LEVINTON, J.S., 1982. The diversity offeeding adaptations of benthic ani-mals. In: Caughey, E.W. (Ed.),Marine Ecology. Prentice Hall, Inc.,Englewood Cliffs, New Jersey.
MINCHIN, D., 1996. Tar pellets and plas-tics as attachment surfaces for Lepa-did cirripedes in the North AtlanticOcean. Marine Pollution Bulletin 32:855-859.
MIZUTANI, N., IWATA, K.& JINNO,N., 2000. Velocity field and localseabed scour around submergedstructure under wave action. Proceed-ings of the Coastal Structures ’99,Volumes 1 & 2, Rotterdam, 755-762.
MOORE, S.L. & ALLEN, M.J., 2000.Distribution of anthropogenic andnatural debris on the mainland shelfof the Southern California Bight.Marine Pollution Bulletin 40, 83-88.
MORTON, B., PEHARTA, M. &HARPER, E.M., 2007. Drilling andchipping patterns of bivalve prey shellpenetration by Hexaplex trunculus(Mollusca: Gastropoda: Muricidae).Journal of the Marine Biological Asso-ciation of the U.K. 87: 933-940.
NASH, A.D., 1992. Impacts of marinedebris on subsistence fisherman. Anexploratory study. Marine PollutionBulletin 3: 150–156.
NCMR (NATIONAL CENTRE FORMARINE RESEARCH), 1999. Mon-itoring of the Saronikos Gulf ecosys-tem affected by the Psittalia sea out-falls. I. Siokou (Ed.), Final Report,338 pp. Athens.
NCMR (NATIONAL CENTRE FORMARINE RESEARCH), 2001. Mon-itoring of the Saronikos Gulf ecosys-tem affected by the Psittalia sea out-falls 2000 – 2004. I. Siokou (Ed.),Final Report, 206 pp. Athens 2001.
PARKER, J.G., 1983. A comparison ofmethods used for the measurement oforganic matter in sediments. ChemicalEcology 1: 201-210.
PEARSON, T.H. & ROSENBERG, R.,1978. Macrobenthic succession inrelation to organic enrichment andpollution of the marine environment.Oceanography and Marine Biology:and annual review 16: 229-311.
PERES, J.M., 1967. The MediterraneanBenthos. Oceanography and MarineBiology: an annual review 5: 449-533.
RELINI, G., RELINI, M., TORCHIA, G.& DE ANGELIS, G., 2002. Trophicrelationships between fish and an arti-ficial reef. ICES Journal of MarineScience 59: S36-S42.
RHOADS, D.C., MCCALL, P.T. &YINGST, J., 1978. Disturbance andproduction on the estuarine seafloor.American Scientist 66: 577-586.
RUPPERT E.E., FOX R.S. & BARNESR.B., 2004. Invertebrate Zoology: AFunctional Evolutionary Approach.7th ed. Brooks Cole Thomson, Bel-mont CA. 963 p.
RYAN, P.G. & MOLONEY, C.L., 1990.
Medit. Mar. Sci., 9/2, 2008, 35-52 51

Medit. Mar. Sci., 9/2, 2008, 35-5252
Plastic and other artefacts on SouthAfrican beaches: temporal trends inabundance and composition. SouthAfrican Journal of Science 86: 450-452.
SCOULLOS, M.J., SAKELLARI, A.,GIANOPOULOU, K., PARASKEVO-POULOU, V. & DASSENAKIS, M.,2007. Dissolved and particulate tracemetal levels in the Saronikos Gulf,Greece, in 2004. The impact of the pri-mary Wastewater Treatment Plant ofPsittalia. Desalination 210: 98-109.
SHANNON, C. E. & WEAVER, W.,1963. The Mathematical Theory ofCommunication. University of IllinoisPress, Urbana/Chicago, IL.
SIMBOURA, N. & ZENETOS, A., 2002.Benthic indicators to use in ecologicalquality classification of Mediterraneansoft bottom marine ecosystems,including a new Biotic Index. Mediter-ranean Marine Science 3: 77-11.
SIMONINI, R., ANSALONI, I.,BONVICINI PAGLIAI, A.M. &PREVEDELLI, D., 2004. Organicenrichment and structure of themacrozoobenthic community in theNorthern Adriatic in an area facingthe Adige and Po mouths. ICES Jour-nal of Marine Science 61: 871-881.
STEFATOS, A., CHARALAMPAKIS,M., PAPATHEODOROU, G. &FERENTINOS, G., 1999. Marinedebris on the seafloor of the Mediter-ranean Sea: examples from twoenclosed gulfs in Western Greece.Marine Pollution Bulletin 36: 389–393.
SVANE, I. & PETERSEN, J.K., 2001. Onthe problems of epibioses, fouling and
artificial reefs; a review. Marine Ecol-ogy 22: 169-188.
UNEP, 2005. UNEP Regional Seas Pro-gramme, marine litter and abandonedfishing gear. Report to the Division ofOcean Affairs and the Law of the Sea,Office of Legal Affairs, UNHQ.Regional Seas Coordinating Office,UNEP, Nairobi.
UNEPUTTY, P. & EVANS, S.M., 1997.The impact of plastic debris on thebiota of tidal flats in Ambon Bay(Eastern Indonesia). Marine Environ-mental Research 44: 233-242.
WHITEHEAD, P.J.P., BAUCHOT,M.L., HUREAU, J.C., NIELSEN, J.& TORTONESE, E., 1986. Fishin theNorth-eastern Atlantic and Mediter-ranean, Volume III, Unesco, Paris.
WILDING, T.A., 2006. The benthicimpacts of the Loch Linnhe artificialreef. Hydrobiologia 555: 345-353.
WILLIAMS, A.T., SIMMONS, S.L. &FRICKER, A., 1993. Off-shore sinksof marine litter: a new problem.Marine Pollution Bulletin 26: 404-405.
WINSTON, J.E., GREGORY, M.R. &STEVENS, L.M., 1997. Encrusters,epibionts and other biota associatedwith pelagic plastics: a review of bio-geographical, environmental and con-servation issues. In: J.M. Coe & D.B.Rogers (Eds.) Marine Debris: Sources,Impacts and Solutions. Springer.
WOODIN, S.A., 1976. Adult-larval inter-actions in dense infaunal assemblages:patterns and abundance. Journal ofMarine Research 34: 25-41.
Submitted: June 2008Accepted: December 2008
Published on line: December 2008