substance p-containing nerve fibres in large peripheral blood vessels of the rat
TRANSCRIPT

Cell Tissue Res (1983) 229: 411422 Cell and Tissue Research �9 Springer-Verlag 1983
Substance P-containing nerve fibres in large peripheral blood vessels of the rat
F. Barja, R. Mathison, and H. Huggel Department of Animal Biology, University of Geneva, Geneva, Switzerland
Summary. Substance P-immunoreactive nerve fibres were localized by the indirect immunohistochemical method in the adventitia and the ad- ventitial-medial border of large peripheral arteries and veins of the rat. Arteries showed a richer substance P-containing innervation than veins. The superior mesenteric artery was densely innervated, whereas no sub- stance P-containing fibres were found around the carotid artery. Sub- stance P produced a vasoconstriction of the veins, but was basically without effect on arteries, although with the carotid artery a dose-depen- dent relaxation was observed. The absence of a correlation between the degree of innervation of the blood vessels and their responsiveness to exogenous substance P suggests that these nerves do not subserve a vasomotor function. The depletion of substance P immunoreactivity from nerves in arteries and veins by capsaicin suggests that substance P-containing vascular nerves are primarily sensory in nature.
Key words" Substance P - Neuropeptides - Immunohistochemistry - Vascular smooth muscle
The association of substance P (SP)-containing nerve fibres with smooth muscle has been demonstrated in intestinal (Costa et al. 1980; Jessen et al. 1980; Schultzberg et al. 1980), tracheobronchial (Nilsson et al. 1977) and urogenital tissues (Alm et al. 1978; Wharton et al. 1981a). In these tissues, SP-fibres are frequently found in close association with blood vessels, and the presence of SP-fibres in blood vessels appears to be a generalized occur- rence. SP-fibres around large blood vessels have been noted in coronary and pulmonary arteries of the guinea pig (Reinecke et al. 1980; Wharton et al. 1981b), pial arteries of the cat (Chan-Palay 1977; Edvinsson et al. 1981, 1982), portal vein of the rat (Barja and Mathison, 1982), and numerous large peripheral veins and arteries of the guinea pig (Furness et al. 1982).
Send offprint requests to: Ronald Mathison, Department of Animal Biology, Physiology Labo- ratory, 22, Bd. des Philosophes, 1205 Geneva 4, Switzerland

412 F. Barja et al.
Both vasomotor and sensory functions can be suggested to account for the presence of SP-fibres in blood vessels. SP has a vasodilator effect on coronary, pulmonary, mesenteric, renal and brain circulation (Maxwell 1968; Hallberg and Pernow 1975; Eklund et al. 1977). Vasodilation of pial arteries by SP has been demonstrated by Edvinsson et al. (1981, 1982). Vasocontraction of the hepatic portal vein of the rat (Hellstrand and J~rhult 1980; Mastrangelo et al. 1980) and the anterior mesenteric vein of the rabbit (Barub6 et al. 1978) has also been documented. A sensory function is impli- cated for SP in vascular nerves since capsaicin pretreatment eliminates SP- immunoreactivity from these nerves (Furness et al. 1982; Matthews and Cuello 1982).
In the present study we report the presence of nerve fibres containing SP-like immunoreactivity in some large peripheral blood vessels of the rat. A sensory function is suggested for the majority of these fibres since capsa- icin pretreatment depletes these nerves of SP-immunoreactivity. Further- more, a lack of correlation was found between the presence of SP-fibres and the responsiveness of the blood vessels to exogenous SP.
Materials and methods
Immunohistochemistry
Male and female Wistar rats weighing 200 250 g were used in the present study. The indirect immunofluorescence method of Coons (1958) was employed for the immunohistochemical localization of SP. The larger blood vessels (e.g. portal, renal and caval veins, the aorta, and superior mesenteric, coeliac and iliac arteries) were perfused in situ with 4% formaldehyde in 0.1 M phosphate buffer (PBS) and opened longitudinally prior to removal from the animal. Smaller vessels were removed intact and washed immediately in the same fixative. For whole- mount preparations the tissues were postfixed in 4% formaldehyde-PBS for 4 h and then washed in 5% sucrose (W/V), with a final washing in PBS for 2 x 20 rain. Arteries and veins were treated for cryostat sectioning (10 ~tm) by fixing the tissues for 24 h in 4% formaldehyde- PBS and then washed overnight in 5% sucrose (Edvinsson et al. 1981).
Antiserum against SP, raised in our laboratory, was used at a final dilution of 1:300. The antiserum has been employed in another histochemical study (Barja and Mathison 1982), and does not cross-react with physalaemin, and tri-, penta- and hepta-COOH terminal frag- ments of SP. Tissues were incubated with antiserum for 36 h at 4 ~ C. Following a rinse, tissues were incubated in FITC-conjugated anti-rabbit IgG at ambient temperature for 1.5 h at a dilution of 1:100. After a final rinse the tissues were mounted in buffered glycerol (1:3, V/V) and examined with a Leitz fluorescence microscope. Sections and whole mounts incubated with antiserum preabsorbed with an excess of exogenous antigen (SP, 10 -5 M) served as controls.
Denervation experiments
Two groups of rats were pretreated with capsaicin (Merck) and examined 5 to 20 days later for SP-immunofluorescence and catecholamine fluorescence. One group of rats was injected subcutaneously with a single dose of capsaicin (50 mg/kg), whereas the second group was treated with increasing doses of capsaicin over two days (25 mg/kg on the first day, with two treatments on the second day: one of 50 mg/kg and the subsequent with 100 mg/kg). Capsaicin was dissolved in 0.9% NaC1 with 10% ethanol and 10% Tween 80 (Merck). Another group of rats was treated with 6-hydroxydopamine (6-OHDA; 2 x 50 mg/kg intraperitoneally) dissolved in 0.9% NaCI with 1 mg/kg ascorbic acid. Animals treated with 6-OHDA were examined for SP- and catecholamine fluorescence. For catecholamine fluorescence the vessels

Vascular nerves containing substance P 413
were spread on glass slides, dried in a dessicator over phosphorus pentoxide and subsequently exposed to paraformaldehyde vapour for 2.5 h at 85 ~ prior to examination with the flluores- cence microscope. For both capsaicin and 6-OHDA experiments separate groups of rats were taken as controls and treated only with the solvent.
Vasomotor actions of SP
Hepatic portal and jugular veins, as well as superior mesenteric and carotid arteries were used in this portion of the study. Arteries prepared either as longitudinal strips or spiral strips, and veins mounted as longitudinal strips were examined for spasmogenic and relaxing effects of SP. Initial tension of 500 mg was applied for measurement of isometric contractions. For relaxation experiments an additional tonus was imposed on the blood vessels with 10-7 M noradrenaline (NA).
Preparations were mounted in a microchamber, possessing a volume of 85 ~tl (Mastrangelo and Mathison (1983) and continuously perfused at 35 ~ C with Krebs solution of the following composition: NaC1 1 2 2 m M ; KC1 4 . 7 m M ; CaCI/ 2 . 8 m M ; KH2PO 4 1 .2mM; NaHCO 3 24.8 mM; MgSO4 1.2 raM; glucose 10.1 m M; 95% 02 - 5% COz; pH 7.4 - 7.5). SP and NA were added to the superfusate to obtain the desired final concentration.
Results
Distribution of SP-fibres
The detection of SP in nerves is generally qualified by the term "SP-like immunoreactivity". For simplicity we will refer to the immunoreactive prod- uct to SP antibodies in nerves as SP, with the reserve that the product identified is "SP-like".
Use of the whole-mount procedure for screening large peripheral arteries and veins revealed that many of these blood vessels contained SP-nerve fibres (Table 1). An arbitrary scale of - t o + + +, based on visual observa-
Table 1. Relative densities of substance P-immunoreactive nerve fibres in arteries and veins of the rat
Artery Rating" Vein Rating
Common carotid - Jugular + Brachial + Brachial + Descending aorta + Posterior vena cava + Hepatic + + + Hepatic + Coeliac + + Portal + Splenic + + Splenic + Renal - Renal - Superior mesenteric + + Superior mesenteric + + Ileolumbar - Ileolumbar - Spermatic + + Saphenous + Inferior mesenteric + + + Caudal + Common iliac + Common iliac + Femoral + Femoral +
a An arbitrary scale of the density of SP nerve fibres was chosen. Most dense ( + + +) , to absence of fibres ( - )

414 F. Barja et al.
Fig. 1A-D. Whole-mount preparations of arteries showing SP-containing nerve fibres. A Hepatic, x440. B Coeliac, x440. C Aorta, • D Iliac, x 360

Vascular nerves containing substance P 415
Fig. 2A-D. Whole-mount preparations showing SP-containing nerve fibres. A Inferior vena cava, x 320. B Femoral, x 320. C Jugular, x 480. D Saphenous, x 320

416 F. Barja et al.
tion of individual blood vessels, was chosen as a means to evaluate the density of reactive fibres. Arteries were generally found to be more densely innervated than veins; such differences are emphasized by comparing the arteries shown in Fig. 1 to the veins in Fig. 2.
The superior mesenteric, hepatic, coeliac and inferior mesenteric arteries exhibited the most dense innervation. Other major arteries, such as the aorta, iliac and the femoral arteries were less densely innervated, and an absence of SP-fibres was noted in the carotid, renal and ileo-lumbar arteries. Due to a much less dense innervation of veins by SP-fibres, generalizations with regard to differential innervation of the veins leaving various vascular beds cannot be made. It is pointed out, however, that the mesenteric and superior caval veins appear to be more densely innervated than other veins, and that SP-fibres were not found in the renal veins.
A general feature of the arterial innervation was a predominately circular orientation of the SP-fibres, reflecting to some extent the general orientation of the smooth muscle cells. In veins SP-fibres did not appear to follow a particular orientation. SP-fibres were frequently found in association with vasa vasorum of the veins and arteries (Fig. 3).
Upon examining whole-mount preparations of both veins and arteries it was noted that two plexuses of SP-fibres could be detected. Beginning at the exterior of the artery a plexus generally devoid of varicose fibres was observed, but as the focal plane was lowered into the blood vessel wall, a second plexus became apparent. This latter plexus contained a large number of varicose fibres (Fig. 4). The presence of these two plexuses was confirmed with cryostat sections. SP-fibres were detected in the external adventitia and at the adventitial-medial border in both arteries and veins (Fig. 5).
Denervation experiments
Pretreatment of rats with 6-OHDA eliminated the majority of catecholamine fibres from all blood vessels studied while leaving SP-fibres intact. Capsaicin pretreatment over several days resulted in a marked depletion of SP- immunofluorescence from vascular nerves. A single injection of 50 mg/kg of capsaicin, however, did not modify the number of SP-fibres found around blood vessels. Catecholamine fluorescence was not modified by capsaicin pretreatment.
Vasomotor actions of SP
Longitudinal strips of portal and jugular veins contracted in the presence of exogenous SP. For the jugular vein the threshold dose of SP was 10-6 M and the ED50 was greater than 10 -s M. The portal vein contracted with a threshold dose of 7.4x 1 0 - 8 M and the ED50 was 1.2x 10 - 6 M. The carotid artery prepared as a longitudinal strip was found to relax upon additon of SP; this relaxation was observed against the 500 mg preload. (Fig. 6). Noradrenaline also relaxed the preload imposed on the carotid.

Vascular nerves containing substance P 417
Fig. 3A, B. SP-containing nerve fibres associated with vasa vasorum. A Portal vein, x 400. B Aorta~ • 320
Fig. 4A, B. Two nerve plexuses can be observed in whole-mount preparations of the superior mesenteric artery. A non-varicose plexus is seen in the outer limits of the adventitia (A), and a varicose plexus becomes visible as the focal plane is lowered through the adventitia (B). x 480

418 F. Barja et al.
Fig. 5A-D. Cryostat sections of various arteries and veins showing SP-containing nerve fibres (arrows) in the adventitia and at the adventitial-medial border. A Superior mesenteric artery, x 480. B Coeliac artery, x 400. C Vena cava, x400. D Portal vein, x 400; a adventitia;
m media
Vascular nerves containing substance P 419
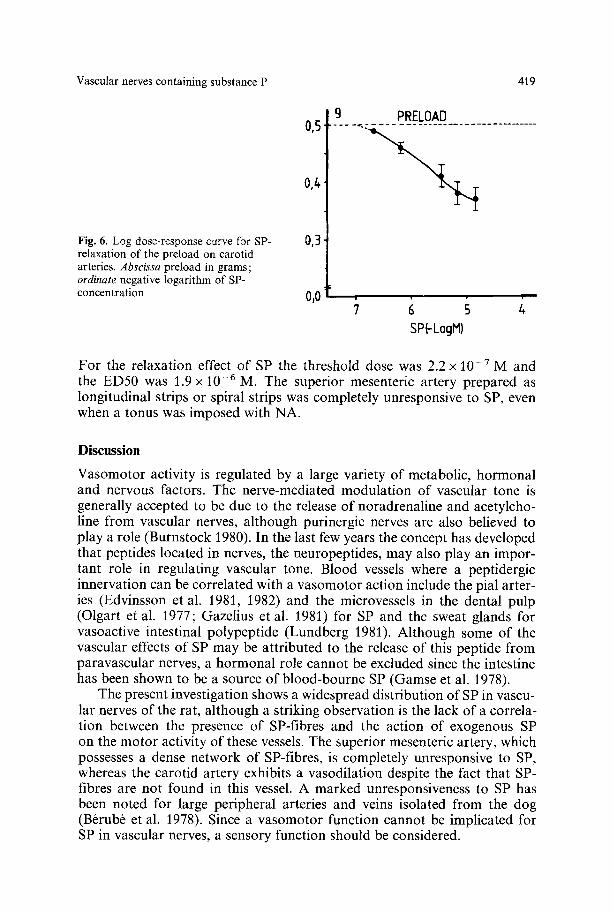
Fig. 6. Log dose-response curve for SP- relaxation of the preload on carotid arteries. Abscissa preload in grams; ordinate negative logarithm of SP- concentration
0,5 ~
0,/+
0,3 1
%0
9 PRELOAD
§ 6 s 4 SP(-LogM)
For the relaxation effect of SP the threshold dose was 2.2 x 10 -7 M and the ED50 was 1.9 x 10 - 6 M. The superior mesenteric artery prepared as longitudinal strips or spiral strips was completely unresponsive to SP, even when a tonus was imposed with NA.
Discussion
Vasomotor activity is regulated by a large variety of metabolic, hormonal and nervous factors. The nerve-mediated modulation of vascular tone is generally accepted to be due to the release of noradrenaline and acetylcho- line from vascular nerves, although purinergic nerves are also believed to play a role (Burnstock 1980). In the last few years the concept has developed that peptides located in nerves, the neuropeptides, may also play an impor- tant role in regulating vascular tone. Blood vessels where a peptidergic innervation can be correlated with a vasomotor action include the pial arter- ies (Edvinsson et al. 1981, 1982) and the microvessels in the dental pulp (Olgart et al. 1977; Gazelius et al. 1981) for SP and the sweat glands for vasoactive intestinal polypeptide (Lundberg 1981). Although some of the vascular effects of SP may be attributed to the release of this peptide from paravascular nerves, a hormonal role cannot be excluded since the intestine has been shown to be a source of blood-bourne SP (Gamse et al. 1978).
The present investigation shows a widespread distribution of SP in vascu- lar nerves of the rat, although a striking observation is the lack of a correla- tion between the presence of SP-fibres and the action of exogenous SP on the motor activity of these vessels. The superior mesenteric artery, which possesses a dense network of SP-fibres, is completely unresponsive to SP, whereas the carotid artery exhibits a vasodilation despite the fact that SP- fibres are not found in this vessel. A marked unresponsiveness to SP has been noted for large peripheral arteries and veins isolated from the dog (Brrub6 et al. 1978). Since a vasomotor function cannot be implicated for SP in vascular nerves, a sensory function should be considered.

420 F. Barja et al.
SP-nerves can be divided into two groups, each group arising from two distinct populations of neurons: capsaicin-sensitive and capsaicin-insensi- tive. The latter group of neurons is present in the enteric nervous system, the ventral spinal cord and various regions of the brain (Jessen et al. 1980; Gamse et al. 1981 a; Cuello et al. 1981). The capsaicin-sensitive nerve fibres have their cell bodies located in sensory ganglia, and a variety of immunolog- ical, anatomical and physiological studies (H6kfelt et al. 1975; Gamse et al. 1981 a, b; Nagy et al. 1981) indicate that SP is a neurotransmitter in primary sensory neurons. The SP-fibres found in association with blood vessels are probably sensory in nature since the SP-immunoreactivity is depleted by capsaicin, but not by 6-OHDA. Similar conclusions have been reached for vascular nerves of the guinea pig that also contain SP (Furness et al. 1982).
In many respects the distribution of SP-fibres in peripheral blood vessels of the rat is similar to that of the guinea pig. In both species arteries possess a somewhat denser innervation than veins, and the superior mesenteric arter- ies, their branches, and the inferior mesenteric arteries are among the most densely innervated. A high degree of parallelism in the relative distribution of SP-fibres to other blood vessels in these two species is also noted. How- ever, the dense innervation of aortic vessels and the carotid artery of the guinea pig is not duplicated in the rat. This difference may reflect a species difference and is not a reflection of the fact that the guinea pig possesses larger quantities of SP-immunoreactive material than the rat (Gamse et al. 1979; Buck et al. 1981). Furness et al. (1982) have suggested a sparser supply of SP-fibres in blood vessels of the rat and rabbit than in the guinea pig, and similar species differences have been noted for the SP-innervation of the urinary and genital organs (Alm et al. 1978). If SP-vascular nerves are monitoring blood vessels for as yet unidentified parameters, the aorta and the carotid artery appear to be two vessels that would be closely monitored. SP, which has been suggested to be the afferent neurotransmitter from baro- and chemoreceptors (Gillis et al. 1980), is localized to the carotid sinus (Helke et al. 1980) and is thus a serious candidate for the transmission of information from the cardiovascular system. The absence of a SP-innerva- tion in the carotid artery and a relatively weak innervation of some other major vessels by this peptide may indicate that another neurotransmitter may be important in relaying sensory information from blood vessels to the central nervous system.
Acknowledgements. This study was supported by Swiss National Fund No. 3.800.80. The tech- nical assistance of D. Solomos and valuable discussions with E. Kolodziejczyk are gratefully acknowledged.
References
Aim P, Alumets J, Brodin E, Hakanson R, Nilsson G, Sj6berg N-O, Sundler F (1978) Pepti- dergic (substance P) nerves in the genito-urinary tract. Neuroscience 3:419-425
Barja F, Mathison R (1982) The adrenergic and peptidergic (substance P and vasoactive intesti- nal polypeptide) innervation of the rat portal vein. Blood Vessels. 19:263 272

Vascular nerves containing substance P 421
Brrub6 A, Marceau F, Drouin JN, Rioux F, Regoli D (1978) The rabbit mesenteric vein: a specific bioassay for substance. P. Can J Physiol Pharmacol 56: 603-609
Buck SH, Deshmukh PP, Yamamura HI, Burks TF (1981) Differences between rats and guinea pigs in gastrointestinal and nervous system substance P levels. Neuropeptides 1 : 383-389
Burnstock G (1980) Cholinergic and purinergic regulation of blood vessels. In: Bohr DF, Somlyo AP, Sparks HV (eds) Handbook of physiology - The cardiovascular system vol. 2 Vascular smooth muscle. Am Physiol Soc, Bethesda, p 567-612
Chan-Palay V (1977) Innervation of cerebral blood vessels by norepinephrine, indoleamine, substance P and the leptomeningeal indoleamine axons: their role in vasomotor activity and local alterations of brain blood composition. In: Owman Ch, Edvinsson L (eds) Neu- rogenic control of brain circulation. Pergamon Press, Oxford, p 39-53
Coons AH (1958) Fluorescent antibody methods. In: Danielli (ed) General cytochemical meth- ods. Academic Press, New York, p 399-422
Costa M, Cuello AC, Furness JB, Franco R (1980) Distribution of enteric neurons showing immunoreactivity for substance P in the guinea-pig ileum. Neuroscience 5:323-331
Cuello AC, Gamse R, Holzer P, Lembeck F (1981) Substance P immunoreactive neurons following neonatal administration of capsaicin. Arch Pharmacol 315:185-194
Edvinsson L, McCulloch J, Uddman R (1981) Substance P: Immunohistochemical localization and effect upon cat pial arteries in vitro and in situ. J Physiol (Lond) 318:251-258
Edvinsson L, McCulloch J, Uddman R (1982) Feline cerebral veins and arteries: Comparison of autonomic innervation and vasomotor responses. J Physiol (Lond) 325:161-173
Eklund B, Jogestrand T, Pernow B (1977) Effect of substance P on resistance and capacitance vessels in the human forearm. In: von Euler US, Pernow B (eds) Substance P, Raven Press, New York, p 275-285
Furness JB, Papka RE, Della NG, Costa M, Eskay RL (1982) Substance P-like immunoreactivi- ty in nerves associated with the cardiovascular system of guinea-pigs. Neuroscience 7: 447-459
Gamse R, Mroz E, Leeman S, Lembeck F (1978) The intestine as source of immunoreactive substance P in plasma of the cat. Arch Pharmacol 305:17-21
Gamse R, Lembeck F, Cuello AC (1979) Substance P in the vagus nerve, immunochemical and immunohistochemical evidence for axoplasmic transport. Arch Pharmacol 306:37-44
Gamse R, Wax A, Zigmond RE, Leeman SE (1981 a) Immunoreactive substance P in sympa- thetic ganglia: Distribution and sensitivity towards capsaicin. Neuroscience 6:437-441
Gamse R, Lackner D, Gamse G, Leeman S (1981b) Effect of capsaicin pretreatment on capsaicin-evoked release of immunoreactive somatostatin and substance P from primary sensory neurons. Arch Pharmacol 316:38-41
Gazelius B, Brodin E, Olgart L, Panopoulous P (1981) Evidence that substance P is a mediator of antidromic vasodilation using somatostatin as a release inhibitor. Acta Physiol Scand 113:155-159
Gillis RA, Helke CJ, Hamilton BL, Norman WP, Jacobowitz DM (1980) Evidence that sub- stance P is a neurotransmitter of baro- and chemoreceptor afferents in nucleus tractus solitarius. Brain Res 181:476~81
Hallberg D, Pernow B (1975) Effect of substance P on various vascular beds in the dog. Acta Physiol Scand 93 : 277-285
Helke C J, O'Donohue TL, Jacobowitz DM (1980) Substance P as a baro- and chemoreceptor afferent neurotransmitter: Immunocytochemical and neurochemical evidence in the rat. Peptides 1 : 1-9
Hellstrand P, J/irhult J (1980) Effects of nine different gastro-intestinal polypeptides on vascular smooth muscle in vitro. Acta Physiol Scand 110:89-94
Hrkfelt T, Kellerth JO, Nilsson G, Pernow B (1975) Substance P: Localization in the central nervous system and in some primary sensory neurons. Science 190:889-890
Jessen KR, Saffrey M J, van Noorden S, Bloom SR, Polak JM, Burnstock G (1980) Immunohis- tochemical studies of the enteric nervous system in tissue culture and in situ: Localization of vasoactive intestinal polypeptide (VIP), substance-P and enkephalin immunoreactive nerves in the guinea-pig gut. Neuroscience 5:1717-1735
Lundberg JM (1981) Evidence for coexistence of vasoactive intestinal polypeptide (VIP) and

422 F. Barja et al.
acetylcholine in neurons of cat exocrine glands, morphological, biochemical and functional studies. Acta Physiol Scand Suppl 496:1-56
Mastrangelo D, Mathison R (1983) Everted portal vein: A sensitive model for studies of vasoactive compounds. J Cardiovascular Pharmacol 5:98-101
Mastrangelo D, Solomos D, Mathison R, Huggel H (1980) Pharmacological characterization of substance P receptors in the isolated rat portal vein. Experientia 37 : 678
Matthews MR, Cuello AC (1982) Substance P-immunoreactive peripheral branches of sensory neurons innervate guinea pig sympathetic neurons. Proc Natl Acad Sci USA 79:1668-1672
Maxwell GM (1968) Action of substance P on general pulmonary and coronary hemodynamics and metabolism of intact dogs. Br J Pharmacol 32:514-522
Nagy GI, Hunt SP, Iversen LL, Emson PC (1981) Biochemical and anatomical observations on the degeneration of peptide-containing primary afferent neurons after neonatal capsa- icin. Neuroscience 6:1923-1934
Nilsson G, Dahlberg K, Brodin E, Sundler F, Strandberg K (1977) Distribution and constrictor effect of substance P in guinea pig tracheobronchial tissue. In: von Euler US, Pernow B (eds) Substance P. Raven Press, New York, p 75-81
Olgart L, H6kfelt, T, Nilsson G, Pernow B (1977) Localization of substance P-like immunoreac- tivity in nerves in the tooth pulp. Pain 4:153-159
Reinecke M, Weihe E, Forssmann WG (1980) Substance P-immunoreactive nerve fibres in the heart. Neurosci Lett 20:265-269
Schultzberg M, H6kfelt T, Nilsson G, Terenius L, Rehfeld JF, Brown M, Elde R, Goldstein M, Said S (1980) Distribution of peptide- and catecholamine-containing neurons in gastro- intestinal tract of rat and guinea pig: Immunohistochemical studies with antisera to sub- stance P, vasoactive intestinal polypeptide, enkephalins, somatostatin, gastrin/cholecysto- kinin, neurotensin and dopamine fl-hydroxylase. Neuroscience 5:689-744
Wharton J, Polak JM, Probert L, de Mey J, McGregor GP, Bryant MG, Bloom SR (1981 a) Peptide containing nerves in the ureter of the guinea-pig and cat. Neuroscience 6:969-982
Wharton J, Polak JM, McGregor GP, Bishop AE, Bloom SR (198~ b) The distribution of substance P-like immunoreactive nerves in the guinea pig heart. Neuroscience 6:2193-2204
Accepted November 10, 1982