studies on the development of echinoidea. ii. the early larva of
TRANSCRIPT

STUDIES ON TUJ5 DEVKr.OPMKNT OF KOHINOIDEA. 29D
Studies on the Development of Echinoidea.
II. The Early Larva of Echinocardium cordatum and theResult of Crossing this Species with Echinus esculentus.
ByProf. E. W. IHiiclu i<lr, D.Sc, LX.D., F.K.S.,
Imperial College of Science and Technology, South Kensington.
With Plates 14 and 15.
THE results recorded in this paper were obtained duringtwo months' sojourn at the Biological Station of the West ofScotland Marine Biological Association at Millport lastsummer. A preliminary account of the same has alreadybeen published in the ' Proceedings of the Royn.1 Society ' (16).
I have to record my thanks to Mr. Richard Elmhirst,Director of the Station, for the whole-hearted manner inwhich he aided my endeavours, and to Dr. James F. Gemmill,Vice-President of the West of Scotland Marine BiologicalAssociation, for the assistance he rendered me in providingme with pure cultures of diatoms, which were invaluable asfood for the developing larvas. To Prof. Graham Kerr, F.R.S.,and to Dr. Agar, of Glasgow University, my best thanks arcdue for the loan of apparatus from the Zoological departmentof that University. And finally, my warmest thanks are due tomy friend, L. W. Byrne, Esq., who re-drew my figures for meso us to make them suitable for publication.
The main object of my research was to test the distributionof paternal and maternal characters in the hybrid producedby crossing two species, in whose larvae distinctly specificcharacters could be found.
A great deal of work has been done in crossing distinct

300 TKOF. E. W. HACJ3111DE.
species of Echinoidea, and most contradictory results havebeen arrived at, as a short review of the principal papers onthis subject will show.
Interest in the question of the character of the hybridlarvae produced by crossing two species of Echinoidea wasfirst awakened by two remarkable papers of Boveri (1, 2).In these papers, which record the same researches, liedescribes the effect of fertilising fragments of SphEereehinuseggs with spermatozoa derived from Echinus. He foundthat he obtained some hybrids of purely maternal type, othersof mixed character, and some very small ones of purely paternaltype. These last he attributed to the development of non-nucleated fragments of eggs which had been entered byspermatozoa of Echinus, since he had previously proved thatnon-nucleated fragments of egg's could develop into larvaewhen entered by the spermatozoa of their own species. Hetherefore concluded that the nucleus alone was the bearer ofheredity, since it could impress an Echinus-character on afragment of a SphEereehinus egg.
This conclusion was attacked by Seeliger (17, 18), who con-firmed Boveri's statement that enucleated fragments of eggscould be fertilised by sperm of the same species, but who deniedthat any hybrids figured by Boveri had been derived from .suchenucleated fragments, since, in a normal hybrid culture, larvaeof every type occur—from those showing a purely maternalcharacter to those which closely approximate to the paternaltype. Therefore he concluded that Boveri's inference thatthe nucleus alone was the bearer of the hereditary qualitieswas not sustained by his experiments. Seeliger also pointedout that Boveri had not properly described the typical formof the larva in each of the parent species, and that no validconclusion as to the character of the hybrid could be drawnuntil this had been done.
Driesch (5) then took up the subject, and pointed out thatin crosses of Sph aerechinus and S t r o n g y l o c e n t r o t u sEch inus the hybrids could be of purely maternal type inrespect of (A) rapidity of development, (B) number of mesen-

STUDIES ON" THE DEVELOPMENT OF ECHINOIDEA. 301
chyme cells, (c) general form. But the first thorough investi-gation of the characters of the larvas of the genera E c h i n u ss t r o n g y l o c e n t r o t u s and Sphaarechinus and of the hybridsproduced by crossing these species was made by Vernon(23, 24). It must be borne in mind that it is possible tofertilise the eggs of Sphserechinus with the sperm ofEch inus and S t r o n g y l o c e n t r o t u s , but that eggs ofEchinus or Strongylocentrotus1 generally refuse to developwhen fertilised by the sperm of Sphaerechinus. Vernoncame to the general conclusion that the predominance ofpaternal or maternal characteristics in the hybrid offspringwas a question of the relative sexual ripeness of the male andfemale parents.
It may be incidentally remarked that thelarvas of E c h i n u sand of S t r o n g y l o c e n t r o t u s are not separable from oneanother by any clearly defined characters, but that those ofSphaarechinus are sharply separated from the others byhaving the skeleton supporting each of the two post-or;ilarms in the form of a lattice-work consisting of severalparallel calcareous rods joined to one another by numerouscross-bars. The skeleton of the post-oral arms of the othergenera consists simply of single bars. It follows that themain featm-e relied on in determining which parent's influencepredominates in the hybi-id is the character of the skeletonof these arms.
demon's work was criticised in 1902 in a remarkably goodpaper by Stembriick (20), in which he dealt with the crossbetween S t r o n g y l o c e n t r o t u s and Sphaarechinus. Steiu-briick called attention to the fact that in their so-calleddistinctive characters the larvas of both forms are v a r i a b l e ;that in pure cultures of S t r o n g y l o c e n t r o t u s larvas areoccasionally met with which possess two calcareous rods withoccasional junctions between them in the post-oral arms, andtliat in pure cultures of Sphsere'chinu s, larvaa are some-times found in which the lattice-work is partly abolished in
1 Driesch (5) records tliat 1 per cent, of Strongylocentrotus eggsdevelop when fertilised with Sphserechinus sperm.

302 HtOl-1. E. W. MACBE1DE.
the skeleton o£ the same arms; and further, that the hybridsbetween these two species are of a very variable character,so that from them a complete chain of forms can be selededleading from larvae showing purely paternal to those showing1
purely maternal characters.Steinbriick's work has not received the attention which was
due to it. It was almost ignored by the next workers whooccupied themselves with the subject—Doncaster andHerbst.Doncaster (4) admits that variations occur in the charactersexhibited by pure cultures of Strongylocentrotus :uid Sphaer-echinus, but believes them to be relatively so infrequent thatthey may be disregarded, and he bases his estimate of therelntive intensity of paternal and materual influence exhibitedin the character of the hybrid larvae on the same criteria asthose employed by Vernon. He arrives at the conclusion,however, that the influence of either parent—the father, forinstance—does not vary with the ripeness, immaturity orstaleness of the sexual products, but with the temperatnre ofthe water, for he found that he obtained the same results inDecember as in May if he artificially warmed the water usedfor the December cultures. This he explains by stating thatwarmed water causes the stronger larva to develop rapidlyuntil they reach the point of development where further pro-gress depends on food. Then they die and the weaker andmore slowly developiug larvse survive them, and it is thesefcliiit sli o w th e pa, tern al inf luence most. In colder waterthe stronger law as develop more slowly, and hence are presenttit the time when the estimate is made (eight days afterfertilisation).
Boveri (3) returned to the subject of the characters exhib-ited by the bastards produced by crossing S fcrongy lo cen-fcrotns, Ech inus and Sphaorechinus. He maintains thatthe influence of the male parent is visible in the shape, skeleton,pigmentation, mesenchyine cells, and sometimes the size ofthe hybrid larva. To tliis Driesch replied (6), controverting allthese points except the one concerning pigmentation.
Herbst, in a most elaborate paper (10), in which he deals

STUDIES ON THE DEVELOPMENT OF ECHINOIDEA. 303
with the result of fertilising the eggs of Sphaarechinu s withthe sperm of S t rongyloceu fcrotus and of Echinus , arrivesat much the same results as Doncaster, but he admits thatthere is also a factor independent of temjaerature whichdetermines the greater or less predominance of paternal char-acters, and this he finds in the varying character of the eggs.In a subsequent paper (11) he records the results of the experi-ment of initiating parthenogenesis in the eggs of Sphae-rechinus by treating them for a short period with valerianicacid and then fertilising them with the sperm of S t rongy lo-cen t ro tu s . Under these circumstances he claims that heobtained a displacement of the development in the maternaldirection, and he maintains that in a few cases he even obtaineda Inrva maternal on the one side of the body and paternal onthe other—a circumstance which he accounts for by supposingthat the spermatozoon had entered the egg after the egg-nucleus had divided and had uuited with one of the twodaughter-nuclei so formed.
Fischel (7) a little later dealt with the hybrids producedby crossing A r b a c i a and Sphserechinus and alsoA r b a c i a and S t r o n g y l o c e n t r o t u s . (It is to be notedthat he persistently and erroneously uses the term E c h i n u sbre v ispinosus for Sphserechinu s g r a n u l a r i s ) . Hearrived at results of generally the same character as thosegained by Driesch : but he tries to show that the effect offoreign sperm entering the egg is to interfere with the normaldistribution of pigment in the egg. He asserts that thespermatozoon can influence the rapidity of development, theform and size of the larva, the development of pigment, theskeleton and the histology of the cells.
Tenuent (22) tried the experiment of crossing the Americanforms Toxopneustes and Hipponoe. The larva? of Toxop-neus tes are like those of S t r o n g y l o c e n t r o t u s . Those ofHipponoe, on the contrary, resemble those of Sphaarechinus.The cross could'be made by .using Hipponoe as male or asfemale parent, but in either case the larvse showed the influenceof Hipponoe as evidenced bythe "lattice-work" in theskcleton

304 PROP. JE. W. MACBRIDE.
of the post-oral arms. Tennenfc's most remarkable conclusionwas that the Hipponoe influence in the hybrid could be de-creased and the Toxopneustes' influence correspondinglyincreased by decreasing the alkalinity of the sea-water by theaddition of a few drops of very dilute acid.
Hagedoorn (9) crossed two species of the same genus, viz.Strongylocentrotus purpura tus and Strongylocen-trotus f ranciscanus, and arrived at the conclusion thatthe hybrid was in every case purely maternal in its character,bub in a paper published a little while after (12) by Loeb,Redman King and Moore recording the results of hybridisa-tion experiments between the same two species, theseauthors state that they are unable to confirm Hagedoorn'sresults, but arrive at the curious conclusion that certain char-acters appear in the hybrid whichever way the cross is made—are, in fact, a dominant over correlative characters; so forinstance the clavate aboral ends of body rods are dominantover the pointed form, the spherical form of larva over thepyramidal, the presence of the recurrent rod in the skeletonover its absence and so on.
The reader will gain from the preceding review animpression of results of a most unsatisfactory and contra-dictory character as the fruit of the work of all these ex-perimenters. Having had considerable experience in rearingthe normal larvas of Echinoderms, it struck me that one prin-cipal cause of such discordant results was the great variabilityof the characters relied on as distinctive of the differentspecies of larvae.
I therefore looked about for " species " to experiment onwhose larvae were distinguished from one another by clearand unmistakable characters. Two possible cases presentedthemselves to mind: first the case of the species Echinusesculentus and Echinus miliaris, and second, the case ofEchinus esculentus and Echinocardium cordatum.
With regard to the first case, it should be noted that thedifferences between the larvae of the two species do notbecome clear until the larvae are about a month old and have

STUDIES ON THE DEVELOPMENT OF E0HIN01DEA. 305
developed all eight arms. These differences concern thearrangement of the ciliated epaulettes, and the appearance ofa green pigment in the larvae of Ech inus mi l i a r i s ; they weredescribed by me in 1899 (IS). IE anyone unacquainted withthe details of the normal development of Echinoderm larvaewere to read through the papers of all the workers ori hybridi-sation he would never suspect that the Echinopluteuslarva ever developed more than four arms; and in this cir-cumstance alone a strong instance is afforded of the necessityof knowing the ordinary development of a species before wemake it the subject of " experimental embryology." In hisfirst paper Boveri actually described the larva of Sphfer-echinus as possessing only two arms ! Whilst I was seekingan opportunity to begin work the questions of the result ofhybridising Echinus esculentus and Echinus mi l ia r i swas taken up by Shearer, De Morgan and Fuchs, who pub-lished a preliminary note of the results of their work (19).In this note they maintain that che hybrid between these twospecies is of a purely maternal character with respect towhatever character be selected for examination; and theymaintain that this maternal character is nob altered bychanging the acidity or alkalinity of the sea-water employed.This certainly is a remarkable result to obtain by hybridisingtwo species of the same genus, and experiments which have beenmade at the Imperial College, the results of which will shortlybe published, do not bear out the view of these authors.
The second case forms the subject of the present paper.Vernon (25) had already recorded the results of crossing theeggs of two species of Ech inoca rd ium with the sperm ofArbacia , Ech inus , Sphaarechinus and S t rongylocen-trotus,and he found that the hybrid larvas were all of a purelymaternal type. When the sperm of Echiuocardiurn wasused to fertilise the eggs of the other genera no result wasobtained except in one instance with the eggs of Echinus. Inthis case one third of the eggs developed and produced larvae ofa purely maternal type. The great distinguishing feature of thelarvaa of Echinocard ium, which it shares with the larvas
VOL. 5 8 , PART 2. NEW SEEIE3. 20

306 PKOl'. E. W. MACBRIDK.
of other Spatangoidea, is the possession of an abora lprocess of the body suppor ted by a special skele ton.This, according to Vernon, is only developed on the fifth day,but according to him it appears in all the hybrid larvas,although in them it is shorter than in the normal larva.Tennent (21), in a short preliminary notice read before theInternational Congress of Zoology held in Boston in 1907,had already announced that he had crossed the eggs ofMoira, a Spatangid, with the sperm of S t rongy locen-t ro tu s . He gives no description of the hybrids, however,but in a later paper gives more details (23). He succeeded infertilising the eggs of Moira with the sperm of Toxo-pneus tes . He kept the larvas alive for seven days. Noneof them developed the aboral spine. Nevertheless, lie saysthat they were of the " maternal intermediate type." He alsomade the reciprocal cross and obtained similar "inter-mediate " larvas ! Some were of the purely maternal typeand developed for twenty days, but the possibility is notexcluded that these were developed from eggs fertilised bychance spermatozoa of their own species.
Since the results which I obtained differ markedly from thoserecorded by Vernon, and are in many respects different fromthose obtained by Tenuent, it may be worth while to give someaccount of the methods employed. At Millport large num-bers of both Ech inus escu len tus and of Ech inocard iumcorda tum were available. The former species could bepicked off the rocks a stone's throw from the laboratory atordinary low tides; the latter species could be obtained bythe bucket-full at low spring tides by digging in a sandybeach about half a mile from the laboratory. The tank-waterin the laboratory was unpolluted by drainage of any kind,for the laboratory is situated over a mile and a half from thecentre of the little town[of Millport, and the full tidal currentof the Firth of Clyde sweeps past the rocks on which it isbuilt. In the water [in the tanks the adults of both specieslive comfortably, but it is, nevertheless, inimical to thelai-vse. The reason of this is to be found in the metallic

STUDIES ON THE DEVELOPMENT OF ECHINOIPEA. 307
pipes through which it is poured into the tanks; it is onlynecessary to dip up the water from the Firth in an earthen-ware or glass vessel in order to provide the hirvas with amedium in which they can grow and flourish. The eggs ofboth species when shaken out of the ovaries are providedwith a glassy chorion which makes fertilisation difficult, but ifthe eggs are allowed to stand in clean sea-water for an hourtwo before being fertilised this membrane disappears. Astested by controls in which the eggs of each species werefertilised by its own sperm, it was evident that the materialused was of the healthiest kind. The eggs of Bchino-cardium fertilised with their own sperm ran t h r o u g hthei r complete l a rva l deve lopment and metamor-phosed into young u rch ins in g r e a t numbers ,accomplishing the whole cycle in three weeks. A full accountof the normal development of this species is reserved foranother paper. In the case of Ech inus escu len tus thelarvae lived for three weeks, developed cilated epaulettes andall eig'ht arms, but as I had previously given a very fullaccounts of the normal development of this species I gave nolurther heed to the larvte. My success in rearing thelarvse I attribute to the cultures of diatoms provided byDr. G-emmill.
When the eggs of Bch inoca rd ium were treated with thesperm of Ech inus a moderate number of hybrids were pro-duced, although possibly not more than one egg in a thousanddeveloped. These hybrids lived for eight days, but then, inspite of being surrounded by abundance of food, they died.When the eggs of Ech inus were treated with the sperm ofEchinocard ium about an equal number (one in a thousaud)developed, and the resulting larvae were of a purely maternaltype. When, however, the sea-water in which thefe r t i l i s a t ion was accompl ished was previously s t e r i -lised by be ing hea ted to 70° C. no single Ech inus eggdeveloped, and when it was found that a certain number ofEch inus eggs would develop if allowed to stand in cleansea-water without the addition of any sperm at all, it became

308 PROF. E. W. MACBBLDE.
obvious that; the supposed hybrids were really normallyfertilised eggs whose fertilisation had been brought aboutby the accidental and unsuspected presence of spermatozoaof Echinus in the water employed. One cannot help wonder-ing whether some of the results recorded in the literature, cf.that of Driesch that 1 per cent, of Strongylocentrotus eggsdevelop when fertilised with tlie sperm of Sphasrechinus, maynot be due to a similar source of error.
Once this source of error had been detected all furtherexperiments in cross-fertilisation were made in sterilised sea-water, and every adult urchin before being opened wascarefully washed in fresh water in order to destroy auyspermatozoa which might be adherent to the outside of thetest. The instruments employed were also carefully steri-lised after each urchin had been opened before another wastaken in hand. New experiments under these conditionsgave exactly the same results when Echinocardiurn eggswere fertilised with the sperm of Echinus, and I conse-quently feel complete confidence in the accuracy of theresults so obtained, but when the eggs of Echinus werefertilised with the sperm of Echinocardium no single eggdeveloped.
Several authors, notably Loeb (12) and Godlewski (8),have succeeded in fertilising the eggs of sea urchins withthe sperm of animals belonging to distinct classes, even todistiuct phyla of the animal kingdom of Crinoidea, Mollusca,etc. In the case of Mollusca it has been proved that thesperm nucleus does not unite with the egg nucleus, i.e. thechromatin which the former brings into the egg is notincluded in the first karyokinetic spindle. In the case ofOrinoids, however, Godlewski asserts that sperm and eggnucleus do unite, and that the chromatin of both is includedin the formation of the first spindle. In all such cases ofheterogeneous fertilisation the larvee are of a purely maternaltype and show no trace whatever of the paternal influence,and therefore this kind of development is termed " Entwick-lungs-erregung," on the supposition that it is due to a

STUDIES ON THE DEVELOPMENT OP ECHINO1DEA. 309
chemical influence exercised by the entering spermatozoonon the egg analogous to the action of salts in producing arti-ficial parthenogenesis and having no relation to the ordinaryhereditary action of the spermatozoon. The method employedto bring about this anomalous development is to add a smallquantity of dilute alkali to the water in which the hetero-geneous fertilisation is effected. G-odlewski found that the
addition of 2-5 c.c. of -^-R solution of NaOH was most effective
in producing results. I therefore tried the effect of ferti-lising the eggs of Echinus with the sperm of Echinocardiumin vessels of sea-water, to which "5 c.c, l'O c.c, 1'5 c.c,
n
Tobeen previously added to every 100 c.c. of sea-water employed.
In the mixture of 2 c.c. of ^n NaOH to 100 c.c. of sea-watera few unhealthy granular blastulse were observed. In noneof the other mixtures did a single egg develop.
Now Loeb has shown (13) that it is possible to get theeggs of sea urchins to develop if they are treated for a verybrief period with a weak solution of butyric acid, thenwashed in sea-water, and then placed for an hour or so in sea-water rendered hypertonic by the addition of a few cubic
n10
The effect of the butyric acid is to cause the eggs to formmembranes closely similar to those formed by eggs whennormally fertilised. Development then begins, but if theeggs are not subsequently placed in hypertonic sea-waterthey break up into spheres which resolve themselves intosmaller spheres and the whole egg is thus reduced to a heapof granules. Now if the actual formation of a vitellinemembrane be carefully watched it will be found that thefirst step in this pi'ocess is the formation at the surface ofthe egg of a large number of minute spherules whose outerwalls coalesce to form the membrane. Hence Loeb puts

310 PROF. E. W. MAOBBIDE.
forward the view that the chemical action of the sperma-tozoon in provoking development consists of two parts;first a process of cytolysis is set up in virtue of whicha vitelline membrane is formed, but this process would leadto the destruction of the egg if it were not checked ; accord-ingly a second chemical action sets in in virtue of whichthe first action is arrested. In artificial parthenogenesisthe checking of the process of cytolysis is effected by thehypertonic sea-water.
I was curious to see what actually had happened to theEchinus egg when treated with the sperm of Bchino-cardiura . Microscopical examination revealed the factthat the eggs had formed fertilisation membranes, showingthat the spermatozoa had entered them, but that they hadthen undergone cytolysis. One such egg is shown in fig. 1,PI. 14. The spermatozoa of Echinocardium were thereforeable to produce a cytolytic action, but incompetent to checkit when it went too far.
Here, then, the cause of the sterility of the cross has beenunmasked.
Turning now to the experiments in which Echinocard iumeggswere fertilised with Echinus sperm Ishall divide what1 have to say into three parts. First Ishall describe carefullythe normal development of Echinus esculentus up till thesixth day; then I shall describe that of Echinocardiumcorda tum up till the fourth day; and then I shall describethe development of the hybrids, which, although they livedfor eight days, did not attain a greater degree of developmentthan that reached by the normal larvae in about five days.
(1) THE DEVELOPMENT OV ECHINUS ESCULENTUS.
On the morning following the fertilisation of the eggs,which had been accomplished in the previous afternoon, clearspherical blastulaa (PL 14, fig. 2) were seen swimming at thesurface of the water. The vegetative pole was already markedby the appearance of the first mesenchyme cells, but they hadnot yet migrated into the blastocele. Soon the blastula

STUDIES ON THE DEVELOPMENT OF ECHINOIDEA. o i l
became flattened on the vegetative pole and the meseuchymecells wandered inwards. The majority of them formed a ringround the periphery of the flattened surface, but somewandered up the sides of the blastula and even reached theanimal pole. At this pole there is a tuft of specially longcilia. The invagination which forms the archenteron began atthe vegetative pole, and the ring of mesenchyrae cellsbecame thickened at two spots diametrically opposite to oneanother, and in these thickenings the rudiments of the larvalskeleton appeared as two trifid " s t a r s . " Each arm of eachstar grew out, the growth being caused by the deposition ofcalcareous matter by the mesenchyrne cells which cling tothe arm. One arm of each " s t a r " grows upwards andoutwards—this is the rudiment of the pos t -ora l rod of theskeleton j one arm grows backwards towards the opening ofthe invagination or " blastopore " and is the rudiment of thefuture "body r o d " of the skeleton, and one grows horizon-tally across and forms the " hor izon ta l rod " of the skeleton.
The invagination now deepens and forms the archenteronand the larva is now termed a g a s t r a l a. From the apex ofthe archenteron a single transversely elongated vesicle is cutoff, which is the rudiment of the coeloru. This stage,reached in two and a half days, is represented iu PI. 14,fig. 3. At the animal pole of the embryo a tuft of speciallylong cilia {all.) is to be seen.
In the course of the next day the stomodseum makes itsappearance as a shallow pit on one side of the anterior end ofthe larva. The side of the larva on which it appears becomesconcave and constitutes the future v e n t r a l surface. Thepit rapidly deepens and reaches the apex of the archeuteron,with which it unites, and thus the alimentary canal is com-plete. The coelomic sac has just previously divided into rightand left halves. From the " s t a r " on each side anotherbranch is given off which extends upwards at the sides of thestomodaeum and here causes a slight protrusion of the ecto-derm, so that the outline of the anterior part of the larvabecomes quadrate instead of being rounded as it was pre-

312 PROF. E. W. MACBKIDE.
vionsly. This extra rod is the " an te ro- la te ra l rod" ofthe larval skeleton and the protrusion is the rudiment of theuntero- lateral arm. Meanwhile the post-oral rod haspushed out a posterior protrusion of the ectoderm on eachside just above the blastopore, now become the anus. Thisprotrusion is the rudiment of the post-oral arm (culled"anal arm" by Driesch and Herbst). Up till now the larvahas been uniformly ciliated all over, with a tufc of speciallylong cilia at the anterior pole, but now the cilia become re-stricted to a ridge forming the edge of the concave ventralsurface. This is the longi tudinal ciliated baud of thelarva, and it runs along the edges of the protrusions which arethe rudiments of the larval arms.
The tuft of specially long cilia becomes incorporated in theanterior border of this band.
A dorsal view of a larva in this stage is given in PI. 14,fig. 4. As soon as the alimentary canal is complete spots oforange-red pigment appear just under the ectoderm allover the larva. The pigment is carried by wanderingmesenchyme cells, which can be seen to migrate into theectoderm and to discharge their pigment (which is probablyof an excretory nature) to the exterior in the form of granules.During the next day the antero-lateral and post-oral armsgrow longer and an adoral band of cilia (ad., fig. 5) becomeswell defined. This consists of two ridges of thickened epithe-lium lying in the ventro-lateral walls of the larval oesophagusbelonging partly to the ectodermal and partly to the endodermalregion of this, which carry long cilia. There is reasonto believe tha t these cilia produce an outwardlydirected cur ren t and that their function is to re-move excess of food matter from the region of themouth. At least if a living larva be watched, particles sus-pended in the water shoot out violently from the ventral side ofthe mouth. On the left side the madrepoj-ic pore-canal and poreare formed by the union of a dorsally directed up-growth fromthe left ccelomic sac and a slight in-pitting of the ectoderm.Constrictor muscles, which cause the oesophagus to exe-

STUDIES ON THE DEVELOPMENT OF ECEINOIDEA. 313
cute peristaltic swallowing movements, are developed from theinner walls of the ccelomic sacs, where they rest against theoesophagus; and d i l a t o r muscles are represented by proto-plasmic strings [dil., PI. 14, fig. 5) which join the antero-lateral rods to the outer walls of the ccelomic sacs. A larvafour days old viewed from the dorsal side is shown in fig. 5.When the larva has attained the a.ge of six days it hasincreased greatly in size, and rudiments of the remainingfour arms, viz. the two prse-oral and the two pos t e ro -dorsal , are visible as very slight protrusions of the ciliatedband. Underneath the rudiments of the postero-dorsal armsis seen an accumulation of mesenchyme cells, in the centre ofwhich a bigh power of the microscope reveals a minute cal-careous spicule—the rudiment of the skeleton of the arm.
No snch accumulation is seen beneath the rudiments of theprse-oral arms ; their skeleton arises in a later stage, as amedian dorsal spicule, situated above the oesophagus farfrom the arms, and the actual prse-oral rods are subsequentoutgrowths from this spicule. This is important, because itproves that the outgrowth of the arm is not directly due to amechanical push exercised by the growing arm rod, but mustrather be due to a chemical influence emanating from thespicule. The aboral ends of the body rods become bentinwards in a crook-like form (cr., fig. 6), often far bettershown than in the specimen figured.
(2) THE DEVELOPMENT OF ECHINOCARDIUH CORDATUM.
The egg of Ech inoca rd ium corda tum is not more thantwo thirds the diameter of the egg of Echinus esoulentus, andthe blastula which develops from it and rises to the top abouteighteen hours after fertilisation is correspondingly small.Moreover, it is not spher ica l , but is e longa ted along oneaxis more than along the other. It is not, however, regularlyoval, but would be more correctly described as being ofcylindrical shape, rounded at the ends. In fig. 7 (PL 14) oneof these blastulse is shown. It is a little older than the blastulaof Echinus shown in fig. 2, and the mesenchyme is more

314 PROF. B. W. MACBR1DE.
developed. The blastula in course of the nest day becomesa gastrula, and this stage is shown in fig. 8. In the gastrularepresented in this figure we see that the mesenchyme isarranged in a circle ; it is termed the primary mesenchyme,because it is given off as in the Ech inus embryo from thevegetative pole of the blastula before the invagination whichforms the archenteron has begun. At two opposite points inthis circle a special aggregation of mesenchyme is to be seen,and inside each aggregation there is already clearly to bemade out the four-armed calcareous " s t a r " which is therudimeut of the larval skeleton. Of the four arms, one isdirected upwards towards the anterior pole of the larva; thisbecomes, as in Echinus, the a n t e r o - l a t e r a l rod. One isdirected backwards towards the blastopore ; this becomes the" b o d y rod." One is directed horizontally and forms thehorizontal rod; whilst the fourth is directed outwards intoa very slight elevation as ectoderm. This elevation of theectoderm is the first trace of the post-oral arm, and thearm of the star corresponding to it is the rudiment of thepos t -ora l rod. This " r o d " is double, i. e.it is representedby two rods parallel to each other. At the anterior pole ofthe larva is to be seen the group of specially long cellscarrying specially powerful cilia (dl.) similar to what wasdescribed in the case of the larva of Echinus. Thesecondary mesenchyme, which is budded from the apexof the archenteron and becomes the loose connective tissue olithe larva, is well seen in the figure. The gastrula rapidlydevelops into an Echinopluteus larva by the same stages asthose described in the case of Echinus . A concave ventralsurface becomes defined, and to the edge?-of this surface thecilia become confined. The anterior tuft of cilia becomesincorporated in the anterior border of this surface, and thepos t -o ra l arms also arise from its border. The stoinodaaumarises as a pit on the ventral surface, and in the larva showuin fig. 9 this is just touching the anterior apes of the archen-teron, although the two cavities are not yet open into eachother. The ccelom arises, as in Echinus, as a transversely

STUDIES ON THE DEVELOPMENT OP ECHINOIDEA. 315
elongated outgrowth from the archenteron, which becomesnipped off and then divided into right and left vesicles. Spotsof da rk red p i g m e n t have been developed. But it is theskeleton which especially arrests our attention. The post-oral arms are already developed, and each is supported by twoparallel rods connected with each other by numerous cross-bars. The antero-lateral arms have not yet appeared, but theantero-lateral rods are already formed—each a single cal-careous rod. From each of these rods a branch (r. r.) pro-jects backwards, this is the rudiment of the r e c u r r e n t rod.The body rods are well-developed single bars. Between theiraboral ends an accumulation of mesenchyme cells can be seen,in the centre of which a tiny calcareous star (ab) can beseen. This accumula t ion of cells r e p r e s e n t s theformat ive mat r ix of the skeleton of the abora lspike—a structure characteristic of the Spatangoid larva andnot found in tlie larva of any regular Echinoid. During thecourse of the next day the antero-lateral arms sprout out andso does the club-shaped aboral spike. The recurreut rod hasgrown backwards parallel to the body rod which it now equalsin length; this rod is usually vestigial in Echinus escu-lentus , but is shown on one side in the larva represented infig. 6. But in Echinocard ium cordatum the recurrentrods extend to the aboral pole and here fork ; the dorsal forkunites with its fellow, whilst the ventral forks unite the endsof the body rods, and in this way a terminal ring is formed inevery way comparable to the so-called " frame" at the aboralend of the Sphasrechinus larva. The skeleton of the aboralspike (ab) consists of three slightly diverging rods connectedwith each other by cross-burs. At their anterior ends theyjoin the terminal ring, which has just been described. Oneof these rods is dorsal and median, the other two are lateral.It follows that the aboral spike possesses a most complicatedskeleton. The apex of the aboral spike is covered with acrest of long-ciliated cells (cil,, figs. 10 and 11). Duringthe course of the next day the rudiments of the postero-dorsalarms make their appearance and in each there is a tiny star,

316 PBOF. E. W. MACBRIDE.
the rudiment of its skeleton (p.d.). The coelomic sacs, whichiu the previous stage lay at the sides of the oesophagus, nowbegin to grow back along the sides of the stomach and oneach side a madrepor ic pore is formed. Whether thisduplicity of the madreporic pore is constant or not I have notyet been able to determine; it is, at any rate, very frequent.A larva in this stage of development is represented in fig. 11.Further than this stage it is not my intention to pursue thehistory of the development. The larva of Echinocardiumhasin four days reached the same stage of development as thatattained by the larva of Echinus esculentus in six days.
Let us now review the differences between the two types oflarvse. Leaving out the question of the duplicity of theniadreporic pore, which may turn out to be a most importantcharacter but which cannot be used at present—we find thatthe larva of Echinocardium differs from that of Echinus —
(1) In possessing an aboral spike supported by a corn-plicated skeleton, whilst at the aboral pole of the Echinusnothing of this kind is to be seen.
(2) In possessing "latticed" bars instead of single rodsas supports for the post-oral arms. (Exceptionally inEchinus a second rod can appear in the post-oral arm. Atrace of such rod is represented in fig. 4. I have never seencross-connections between the two, but I am informed thatthese also can appear as a rare exception.)
(3) In possessing well-developed r e c u r r e n t rods insteadof only vestiges of such rods.
(4) In possessing brownish-red instead of orange-redpigment.
(5) In its more rapid development.We shall now see how far these characters are represented
in the hybrids.
DEVELOPMENT OF EGGS OP ECHINOCAKDIUM CORDATUM FERTILISED
WITH THE SPERM OF ECHINUS ESCULENTUS.
The earlier course of the development of Echino-cardium eggs fertilised with the sperm of Ech inus

STUDIES ON THE DEVELOPMENT OF EOHINOIDEA. 317
e s c u l e n t u s is very similar to that undergone by these eggswhen fertilised with the sperm of their own species. Fig. 12shows the appearance of the free-swimming blastula twenty-four hours fertilisation. In this specimen the vegetative end israther broader than the animal end, whereas in the blastula ofEchinocnrdiam represented in fig. 7, the reverse is the case,but this difference between the hybrid and normal blastulais not constant. By the end of the second day the blastulais converted into a gastrula such as is shown in fig. 13. Therate of development of the hybrid, however, varies with thespecimens used as parents. In fig. 14 a hybrid larva ofthe same age is represented, which, however, belonged to adifferent culture, and we can see that it has attained the stagewhere the calcareous stars have been formed and where theccelom is already grooved off from the archenteron. If fromthe same specimen eggs are taken some of which are fertilisedwith sperm of its own species and some with the sperm ofE c h i n u s e scu len tus , then the hybrids will always developmore slowly than the normal Iarv83. As development pro-ceeds the hybrids fall more and more behind the normallarvae. In fig. 15 a hybrid four days old is represented. Inthis specimen the post-oral arms are well developed but thestomodseurn has not pined the oesophagus. The bars sup-porting these arms are "latticed," bat the cross bars arecomparatively few. The antero-lateral bars are there but theantero-lateral arms are not yet developed. As shown in fig.16, however, hybrids four days old may be more advanced indevelopment. In the specimen shown in this figure thealimentary canal is complete and the post-oral arms are longerthan in the specimen shown in fig". 15. But the s u p p o r t i n gbars of these arms are single rods for the greaterpart of their length, although a second short rod accom-panies the first at its base, as it does as a variation inEchinus esculentus. Both larvas agree in the totalabsence of any indication of the aboral spike or ofits skeleton.
When we turn our attention to the hybrid larvae five days

318 PROF. E. W. ilACBEIDE.
old which are shown in figs. 17 and 18 the same totalabsence of an aboral spike or any indication of its skeletonstrikes us. The antero-lateral arms are now developed. Inthe larva represented in fig. 17 the skeleton of one post-oralarm consists at its base of no less than four parallel rods;more distally a curious vestige of latticing is seen in the formof short rods accompanying the main rod and each connectedwith it by a transverse bar. The ends of the. body rods arein-bent in a crook-like fashion such as occurs in the normallarva of Ech inus esculen tus . The larvse represented infig. 18 lias an almost normal Bchinocardium skeleton in eachof the post-oral arms, but it is absolutely devoid of the aboralspike. In fig. 19 a hybrid six days old is represented; itshows much the same features as those shown by the five-dayhybrid represented in fig. 18, only the arms are betterdeveloped and the crook-like in-bending of the aboral ends ofthe body rods is very marked. We notice also that arecurrent rod is well developed. In the seven-day larvashown in fig. 20 one of the antero-lateral arms has beenabsorbed—aphenomenon which often occurs with normal larvfflwhich are not quite healthy. The skeleton of this larvashows hardly any trace of the maternal influence ; it is almostpurely of the paternal type.
None of the hybrids lived longer than eight days althoughthey were supplied with abundant food. Three of these eight-day larvas are represented in figs. 21, 22 and 23. Thatshown in fig. 21 has a skeleton almost purely paternal incharacter; the only hint of maternal influence is to be seen inthe thorns besetting the single rods which constitute theskeleton of the post-oral arms. One striking maternal characteris, however, shown in the dupl ic i ty of the madrepor icpor e. The larva shown in fig. 22 has a double rod in each post-oral arm, but on one side the two rods are fused into one forthe middle of their length, whilst on the other they are widelyseparated from one another and connected by a few cross-bars. Finally in the remarkable larva shown in fig. 23 thereis a skeleton of a purely maternal type, and the aboral spike

STUDIES ON THE DEVELOPMENT OF ECH1NOIDEA, 319
is typically developed. The larva is distorted, however,since the oral lobe containing stomodEeum is, as it were,twisted round on the body, and the result of this twist on thedisposition of the skeletal rods is at first not a little puzzling.Care, however, enables us to recognise all the constituentparts of the maternal skeleton.
To sum up : The hybrid produced by fertilising the eggsof B c h i n o c a r d i u m c o r d a t u m with the sperm of E c h i n u sescu len tus follows the mother in the character and distribu-tion of the pigment: it is much smaller than larvse of eitherthe paternal or maternal species; it almost always followsthe father in the total absence of the aboral spike and of itssupporting skeleton, since in only one hybrid out of thehundreds examined was the aboral spike formed. Inthe skeleton of the post-oral ai*ms the hybrid may be ofthe paternal type, of the maternal type, or of an intermediatecharacter. In the inbending of the aboral ends of the bodyrods the hybrid follows the father.
The most important of these results is undoubtedly thetotal inhibition in the vast majority of cases of the formationof the aboral spike in a larva developed from a Spatangidegg, and the formation of a larva with a rounded aboral endand in-bent body rods, recalling in these features the E c h i n u slarva. When we recollect that according to Shearer,De Morgan and Fuchs the crossing of two species of thegenus Ech inus results in the production of larvae of thematernal type, uo matter what feature is considered, it is nota little remarkable to find the paternal influence so strong ina cross between two species belonging not only to differentgenera but to different orders, species which must havediverged from a common ancestor at the beginning of thesecondary epoch many millions of years ago.
If the attempt be made to explain the absence of theaboral spike as a mere concomitant oi the feeble develop-ment of the hybrid, a glance at the figures of normalEchinocardium larvas will be sufficient to refute this sugges-tion. We see there that theaboral skeleton and its formative

320 PBOF. E. W. MAOBBIDE.
mass of mesenchyme can be detected in the normal larvabefore the autero-lateral arms have developed at all, andin many of the hybrids the antero-lateral arms are welldeveloped. The absence of the aboral spike is thereforenot a mere consequence of stunted growth, but is due topaternal influence. Moreover, it is not possible to reconcilethe facts just recorded with any theory of dominance such asthat put forward by Loeb, Redman, Kiug and Moore.
If any clear meaning be attached to the word "dominance/'it must signify that there is a certain factor "which may bepresent or absent in a germ-cell, but which, when it ispresent, produces in the resulting embryo a certain character.JSTOW if we take the question of the skeletal rods supportingthe aboral arms, these are normally single in Echinus andlatticed in Echinocardium. When these two species arecrossed we do not always find either single or latticed bars,but we find, as a matter of fact, every intermediate condi-tion of affairs. To alter the significance of the word" dominance " in such away as to make it include phenomenalike these is, in my opiniou, to empty it of all its meaning.
ZOOLOGICAL LABORATORY,IMPERIAL COLLEGE or SCIENCE AND TECHNOLOGY;
June 25th, 1912.
LIST OP WORKS .REFERRED TO IN THIS PAPEH.
1. Boveri.—"Ein geschleclitlich erzengter Organismus ohne niiitter-liche Eigenscliaften," 'Ber. G-es. Morph. u. Phys.,' Munich, 1889.
2. "Ueb.er die Befruchtungs- und Entwioklungsfiihigkeitkernloser Seeigeleier und iiber die Moglichkeit ihrer Bastardie-rung," 'Arch. Ent. inech.,' vol. ii, 1895.
3. "Uebev denEinfluss der Samenzelle auf dieLarvencharaktereder Echiniden," ibid, vol. xvi, 1903.
4. Doncaster.—" Experiments in Hybridisation, with Special Referenceto the Effect of Conditions on Dominance," 'Phil. Trans. Roy.Soc. (Lond.),' series B, vol. cxcvi, 1903.
o. Driesch.—" Ueber rein miitterliche Charaktere an Bastard-larvenvon Echiniden," ' Arch. Ent. mech.,' vol. vii, 1898.

STUDIES ON THE DEVELOPMENT OF ECH1NOIDEA. 321
6. Driesch.—"Ueber Seeigelbastarde," ibid., vol. xvi, 1903.7. Fischel.—"Ueber Bastardierungs versuche bei Echinodernien,"
ibid., vol. xxii, 1906.8. Godlewski.—" Untersuchungen iiber die Bastardierung dei1 Echi-
niden und Crinoiden-familie," ibid., vol. xx, 1906.9. Hagedoorn.—" On the purely Motherly Character of the Hybrids
produced from the Eggs of S t rongy looen t ro tus , " ibid.,vol. xxvi, 1909.
10. Herbst.—'Vererbungs-studien,' i-iii; ibid., vol. xxi, 1906.11. ' Vererbungs-studien,' iv and v; ibid'., vol. xxiv, 1907.
12. Loeb. — " Ueber die Natur der Bastardlarve zwischen demEchinodermenei (S t rongylocent ro t t i s f ranciscanus) undMolluskensamen (Chlorostoina funebra)," ibid., vol. xxvi,1908.
13. 'Die chemische Entwickelung des tierischen Eies.,' Jena,1909.
14. Redman King and Moore.—" Ueber Dominanzerscheinungenbei den hybriden Pluteen des Seeigels," 'Arch. Ent. mech.,' vol.xxix, 1910.
15. MacBride.—"Studies in the Development of Echinoidea. I. TheLarva of E c h i n u s escu len tus and of E c h i n u s nii l iaris ,"' Quart. Journ. Micr. Soc.,' vol. 42, 1899.
16. " Studies in Heredity. I. The Effect of Crossing the Sea-urchins, E c h i n u s escu len tus and Ech inoca rd ium cor-datum," ' Proc. Roy. Soc. (Lond.),' series B, vol. lxxxiv, 1911.
17. Seeliger.—" Giebt es gesclilechtlich erzengter Organismen ohnemiitterliche Eigenschaften," ? 'Arch. Ent. mech.,' vol. i, 1894.
18. " Bemerkungen iiber Bastardlarven der Seeigel," ibid.,vol. v, 1896.
19. Shearer, De Morgan and Fuchs.—"Preliminary Notice on theExperimental Hybridisation of Echinoids," ' Journ. Mai-. Biol.Ass.,' vol. ix, 1911.
20. Steinbriick.—"Ueber die Bastardbildung bei Strongylocen-t r o t u s l ividus und Sphoerechinus granular is ," 'Arch.Ent. mech.,' vol. xiv, 1902.
21. Tennent.—" Hybrid Echinodevm Larvse," ' Proc. of Seventh Inter-national Zoological Congress, Boston,' 1907.
22. "The Dominance of Maternal or Paternal Chai'acters inEchinoderm Hybrids," 'Arch. Ent. niech.,' vol. xxix, 1910.
23. " Echinoderm Hybridisation," ' Publications of CarnegieInstitution,' No. 132, 1910.
VOL. 58, PAET 2.—NEW SEEIKS. 21

322 PROF. B. W. MACBRIDB.
24. Vernon.—" The Relations between the Hybrid and Parent Formsof Ecbinoid Larvae," ' Phil. Trans. Eoy. Soc. (Lond.),' series B,vol. cxc, 1898.
25. "Cross-fertilisation amongst Echinoids," 'Arch. Ent. meoh.,'vol. ix, 1900.
EXPLANATION OF PLATES 14 AND 15,
Illustrating1 Prof. E. W. MacBride's. paper entitled " Studieson the Development of Echinoidea, Part II ."
LIST OF ABBREVIATIONS EMPLOYED.
ab. Skeleton of aboral spike of Eehinocardium larva, ad. Adoralciliated band. a. I. Skeleton of antero-lateral arm of larva, an. Anns.6. r. Body-rod of skeleton of larva, cole. First rudiment of skeleton oflarva, oil. Anterior tuft of long cilia and posterior tuft of long ciliain Echinocardium larva, coe. Ooslomic sac. or. In-bent crook of body-rod in skeleton of larva of hybrid and of Echinus, dil. Dilator musclesof ccelomic sac. / . Fertilisation membrane, h.r. Horizontal rod oflarval skeleton, int. Intestine. I.p. c. Left posterior ccelom. mad.Normal left madreporic pore, mad.1 Right madreporic pore of Echino-cardium larva, ines. Primary mesenchyme. ces. (Esophagus, p. d.Rudiment of skeleton of postero-dorsal arm. pr. o. Rudiment of prte-oral arm. p.o. Skeleton of post-oral arm. r.p.c. Right posteriorC(elom. st. Stomach, stom. Stoniodoeum'.
[All the figures were drawn from living larvse with the help of thecamera lucida, and all the lai-vse figured are represented with a uniformmagnification of 200 diameters so that their relative sizes can be seen.]
PLATE 14.
Fig. 1.—Egg of Echinus esculentus treated with the sperm ofEchinocardium cor datum and examined twenty-four hours after-wards. / . Fertilisation membrane.
Fig. 2.—Blastula of Echinus esculentus twenty hours old. ines.Primary mesenchyme in the act of being formed.
Fig. 3.—Late gastrula of Echinus esculentus two and a half

STUDIES ON THE DEVJiLOI'MENT OK ECH1NOIDEA. 323
days old. cm. Coelomic sac just separating from archenteron. 6. r.Body-rod, h. r. Horizontal rod, and p. o. post-oral rod as three branchesof trifid calcareous star, the rudiment of the larval skeleton.
Fig. 4.—Larva of E c h i n u s esculentus three days old viewed fromthe dorsal side. a. I. antero-lateral rod of skeleton, cce. Ccelomic sacs,ces. Endodermal part of oesophagus, stom. Stomodaeum. p. o. Post-oral rod of skeleton (notice the supplementary rod on right side).
Fig. 5.—Larva of E chin vis escu len tus four days old viewedfromthedorsal surface, ad. Thickening' of epithelium carrying adoral ciliatedband. dil. Dilafor muscles of coelomic sac and of oesophagus attachingthese structures to the antero-lateral rod of the larval skeleton, mod.Madreporic pore.
Fig. 6.—Larva of Ech inus escu len tus six days old viewed fromthe dorsal surface, cr. Inbent crook at aboral end of body-rod, p. d.Accumulation of mesenchyme cells, the formative tissue of skeletonof postero-dorsal arm. p. r. o. Rudiment of pra-oral arm. r. r. Vestige ofrecurrent rod of skeleton.
Fig. 7.—Blastula of Ech inoca rd ium corda tum twenty-fourhours after fertilisation, mes. Primary mesenchyme being formed.
Fig. 8.—Gastrula of Ech inoca rd ium co rda tum one and a halfdays old. calc. Rudiment of skeleton- dl. Anterior tuft of long cilia,uies. Secondary mesenchyme being budded from the apex of the archen-teron.
Fig. 9.—Larva of Ech inocard ium cordatvun two days old viewedfrom the dorsal surface, ab. Calcareous star, the rudiment of the skele-ton of the aboral spike embedded in a mass of formative mesenchyme.p. o. Latticed skeleton of post-oral arm. ces. Endodermal oesophaguswhich is in contact with, but which has not yet opened into, stotn.the stomodseum. r. r. Recurrent rod of the skeleton.
Fig 10.—Larva of Ech inoca rd ium corda tum three days oldviewed from the dorsal surface. The antero-lateral arms have grownout and the stomodseum has opened into the oesophagus. The aboralspike and its skeleton are fully formed, dl. Posterior tuft of ciliaat the apex of the aboral spike.
Fig. 11.—Larva of E c h i n o c a r d i u m corda tum four days old,viewed from the dorsal surface. I. p. c. Backward growth of the coelomon the left side of the stomach, which will be cut off as the left posteriorcoelom. r.p.c. Similar growth of the crelom ou the right side of thestomach which will be cut off as the right posterior coelom. mad.Left madreporic pore, mad.1 Right madreporic pore. p.d. Rudimentof the skeleton of the postero-dorsal arm.

324 PROF. E. W. MAOBRIDE.
' PLATE 15.
[All the following figures represent larvae developing from the eggsof Eohinocard ium corda tum which have been fertilised with thesperm of Echinus 'esculentus . ]
Fig. 12.—Hybrid blastula twenty-four hours old. mes. Primatry-mesenchyme.
Fig. 13.—Hybrid gastrula two days old.. . cil. Anterior, tuft ofcilia.
Fig. 14.—Hybrid gastrula (from another culture) two days old.cos. Rudiment of coeloni. cede. Rudiment of skeleton, mes. Secondarymesenchyme.
Fig. 15.—Hybrid larva four days old viewed from the dorsal side.a. I. Antero-lateral rod of skeleton. 6. r. Body-rod, cil. Anterior tuftof cilia now incorporated with longitudinal ciliated band. ces. Larvaloesophagus not yet joined to stomodseum. r. r. Vestigial recurrentrod.
Fig. 16.—Hybrid larva four days old, more advanced in developmentthan that shown in fig. 15 ; viewed from the dorsal side.
Fig. IV.—Hybrid larva five days old viewed from the dorsal surface.c.r. In-bent crook at aboral end of body-rod.
Fig. 18.—Hybrid larva five days old viewed from the left side. ad.Ridge of thickened epithelium in the cesophagus carrying the aboralciliated band. an. anus. h. r. Horizontal branch of the skeleton.
Fig. 19.—Hybrid larva six days old viewed from the dorsal surface.r. r. Recurrent rod.
Fig. 20.—Hybrid larva seven days old viewed from the dorsal surface.Notice that one antero-lateral arm has been absorbed.
Fig. 21.—Hybrid larva eight days old viewed from the dorsal surface.•mad., mad.1 The two inadreporic pores.
Fig. 22.—Hybrid larva eight days old viewed from the ventral surface.an. Anus. h. r. Horizontal branch of larval skeleton.
Fig. 23.—Hybrid larva eight days old viewed from the left side.This is the only hybrid in which the aboral spike and its skeleton havebeen developed. Notice the distortion of the larva. It is as if the leftantero-lateral ann and the left side of the oral lobe had been forciblytwisted away from the spectator, b. r. The right and left body-rodscrossing each other, cil. Posterior tuft of cilia.

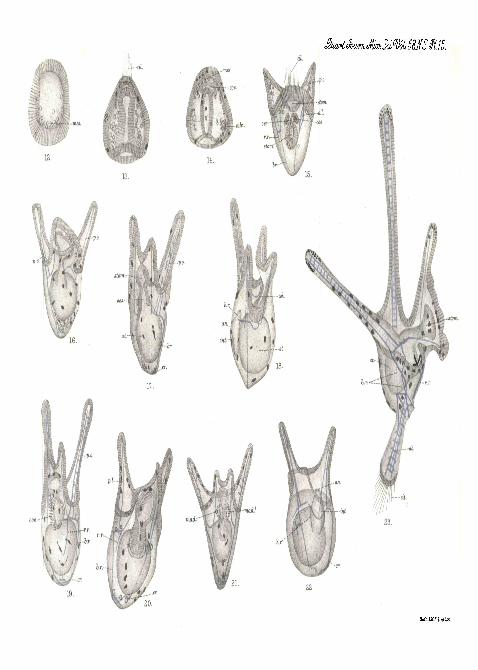
ah.
cd.
22.