structure of the carbohydrate units of iga, · pdf filestructure of the carbohydrate units of...
TRANSCRIPT

THE JOURNAL or B~oma~ca~ CHE?~I~TRY Vol. 249, No. 22, Iseue of November 25, pp. 7279-7281, 1974
Printed in U.S.A.
Structure of the Carbohydrate Units of IgA, Immunoglobulin
II. STRUCTURE OF THE 0-GLYCOSIDICALLY LINKED OLIGOSACCHARIDE UNITS*
(Received for publication, April 25, 1974)
JACQUES BAENZIGER! AND STUART KORNFELD~
From the Departments of Medicine and Biochemistry, Washington University School of Medicine, St. Louis, Missouri 63110
SUMMARY
In the preceding paper (BAENZIGER, J., AND KORNFELD, S., J. Biol. Chem. 249, 7260-7269) the structure of the two asparagine-linked oligosaccharide units of an IgAl myeloma protein was defined. In this paper we present data which establish the structure and linkage points of the five O- glycosidically linked oligosaccharide units found on the CX~ subtype heavy chain to be as shown below.
myeloma protein. We now present data establishing the struc- ture and location of the 0-glycosidically linked oligosaccharide units of IgAl (Walker) and of the hinge region glycopeptide from IgAl (Oso) (2). The structure of the oligosaccharide units present in this region has become of special interest in view of the finding by Frangione and Wolfenstein-Todel that the amino acid sequence of this region of the crl subtype heavy chain is very symmetrical and is deleted in the (~2 subtype, which also does not
Gal Gal Gal Gal
1 61,3 1
61,3 1
61,3 I
81,3
GalNAc GalNAc GalNAc GalNAc GalNAc
1 Set-- Thr- Pro -Pro - Thr- (Pro- Pro- Ser - Thr- Pro -Pro- Thr- Pro - Ser - Pro- b--N-
This structure appears to be identical with that which we have determined for the hinge region glycopeptide isolated by Frangione and Wolfenstein-Todel [FRANGIONE, B., AND WOLFENSTEIN-TODEL, C. (1972) Proc. Nai. Acad. Sci. U. S. A. 69, 3673-36761, from a different IgA, myeloma pro- tein. This region is deleted in the (YZ subtype of IgA. Aside from the presence of a terminal N-acetylgalactosamine at the NH2 terminus, the location and structure of the 5 O- glycosidically linked oligosaccharide units of the glycopep- tide preserve the symmetry observed by Frangione and Wolfenstein-Todel to be present in the amino acid sequence of the hinge region.
In the preceding paper (1) we established the sequence of the 2 asparagine-linked oligosaccharide units found on an IgAIl
* This work was supported in part by United States Public Health Service Grant ROl CA08759.
2 Recipient of United States Public Health Service Medical Scientist Training Award 5 TO5 GM 02016. This paper consti- tutes one chapter of a thesis submitted in partial fulfJlment of the requirements for the Ph-D. degree in Biological Chemistry.
$ Recipient of the United States Public Health Service Re- search Career Development Award AM 50298.
1 The abbreviations used are: IgA,, IgA1, IgA, IgG, IgM,
contain N-acetylgalactosamine (2). A comparison of the struc- tures of the oligosaccharide units found on IgG (3), IgM (4), IgE (5, 6), and IgA myeloma proteins is also presented.
EXPERIMENTAL PROCEDURE
Materials IgA glycopeptide I was prepared as described in the preceding
paper (1). The hinge region peptide from IgA, (Oso) was kindly provided by Dr. Blas Frangione, Department of Medicine, New York University School of Medicine (2).
Methods
The majority of the methods employed have been presented previously (1, 3, 5, 6) with the following additions.
Amino Acid Sequence Determination-Edman degradations were carried out, according to the procedure of Waterfield and Huber (7). Dansyl amino acids were identified on polyamide thin layer plates by a modification of the Hartley procedure (8) and sub- tractive Edman degradations were performed as described by Bradshaw et al. (9).
Performate Oxidation-Performate oxidation for cysteic acid determination was carried out according to the method of Hirs (10).
Amino Acid Hydrolyses-Amino acid hydrolyses were routinely carried out, in 6 N HCI for 48 hours at 110” in evacuated tubes. This was necessitated by t,he appearance of a poorly hydrolyzed tripeptide consisting of Thr-Pro-Pro when shorter hydrolysis times were ut.ilized.
IgE, IgD, immunoglobulins A, etc.; dansyl, 5-dimethylamino- napthalene-1-sulfonyl.
7270
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Alkaline Degradation-Alkaline degradation was carried out under several different conditions which varied with respect to temperature (445”), normality of NaOH (0.1 to 1.0 N), duration (24 to 72 hours), and the presence or absence of NaBH4. The conditions used for the various experiments reported are presented under “Results.”
Alkaline degradation in 0.5 M NatSO and 0.1 N NaOH was carried out for 48 to 72 hours at 40” as described by Spiro (11). After neutralization the samples were hydrolyzed for 48 hours in 6 N HCl at 110” in evacuated tubes. A port,ion of the hydrolysate was subjected to amino acid analysis while the remainder was loaded onto a column of Dowex AG l-X8 (200 to 400 mesh, formate form) previously equilibrated with 5 mM pyridine-formate buffer, pH 4.0. The column was washed with 10 volumes of 5 mM pyri- dine formate buffer pH 4.0 and then the acidic amino acids were eluted with 5 volumes of 4 N formic acid. After drying by lyophili- zation the samples were spotted on Whatman 1 paper and sub- jected to descending chromatography for 5 days in l-butanol- acetic acid-H20 (4:1:5). Chromatograms were developed by dipping in 0.2% ninhydrin in acetone and then heating at 100”.
Acetylation-Glycopeptide I and the product of alkaline degra- dation were acetylated by incubation in 2oj, aqueous acetic an- hydride-l M bicarbonate for 30 min at room temperature.
Periodate Consumption-Periodate consumption was measured by the spectrophotometric method, following the decrease in absorbance at 223 nm (12). The assay was standardized using sorbitol. The reaction mixtures contained a 4- to B-fold excess of sodium metaperiodate over that expected to be consumed and were carried out in HZO.
RESULTS
Composition, Partial Amino Acid Sequence, and Molecular Weight of GZycopeptide Z-Three preparations of IgAl glycopep- tides were made as described in the preceding paper (1). Each preparation of glycopeptide I had the same composition consist- ing of threonine, serine, proline, galactose, and N-acetylgalac- tosamine in the molar ratios of 4:5:8.5 :4:5, respectively (Table I). The hinge region peptide from IgAl (Oso) had a composition very similar to that of glycopeptide I (Table I). The carbo- hydrate content was identical with that of glycopeptide I and the presence of additional amino acid residues is accounted for by the fact that trypsin digestion produces a much larger peptide fragment than that produced by pronase digestion (see Fig. 1).
7271
Using the dansyl-Edman degradation procedure the sequence Ser of the first 5 residues of glycopeptide I was found to be -
Thr Pro Pro -7 I - Thr (Table II). Although no de- crease in galactosamine was observed at any of the steps of the dansyl-Edman degradation, this cannot be held as significant since no attempt to repurify the glycopeptide was made between steps. Having determined the sequence of the first 5 residues of glycopeptide I, it was possible to align this glycopeptide with the hinge region peptides sequenced by Frangione and Wolfen- stein-Todel (2) as shown in Fig. 1. The proposed locations for the 0-glycosidically linked oligosaccharides found in glyco- peptide I all coincide with regions which are deleted from the IgAz subtype.
The estimation of the molecular weight of glycopeptide I by exclusion chromatography on Sephadex G-50-80 was described in Table II and Fig. 3 of the preceding paper (1). The calcu- lated molecular weight of 3278 is somewhat greater than the observed value of 2720; however, this is not unexpected since glycopeptide I has a large amount of peptide which accounts for 50% of the molecular weight (1645 of the 3278) but would not be expected to contribute its full calculated molecular weight on a column calibrated with oligosaccharides (13).
Methylation-Glycopeptide I was methylated using the Hako- mori method as described in the previous paper (1). The results of the methylation shown in Table III and Fig. 2 indicate that all 4 residues of galactose and 1 residue of N-acetylgalactosamine are terminal. At least three and probably all 4 of the remaining N-acetylgalactosamine residues are substituted at position 3. This agrees quite well with the results obtained by periodate oxidation of glycopeptide I, which show destruction of all 4 resi- dues of galactose and of only 1 of the 5 residues of N-acetyl- galactosamine (Table I).
Although the extremely basic conditions utilized in the Hako- mori method of methylation might have been expected to result in destruction of a large proportion of the 0-glycosidically linked N-acetylgalactosamine by a /3 elimination reaction, it was quite apparent from other data that this was not the case. In fact,
TABJ~E I
Effect of alkaline treatment and periodate oxidation on glycopeptide I and hinge region glycopeptide of IgAl (0.~0)
Galactose. Galactitol. N-Acetylgalactosamine N-Acetylgalactosaminitol Cysteic acid or
a-amino-&sulfonylbutyric acid Sulfonylgalactosamine Threonine................. Serine........................... Proline...................... Glycine........... . . ..__.... Alanine. . Valine Histidine.............. ,... Arginine S-Cys (Cm)
-
-
--
-
Composition of glycopeptide I Composition of hinge region glyco- peptide from IgAl (030)
Native 0.1 N NaOH, 0.4 M 0.1 N NaOH, 0.5 M NaIO4 NaBH,, 66 hrs, 45’ N&02, 48 hrs, 40’ oxidation Native 0.1 N NaOH, 0.4 N
NaBHa, 36 hrs, 45”
4.1 f 0.2 0
5.0 f 0.2 0
3.4 0.7 2.1 1.8
4.3 f 0.2 1.4 4.9 f 0.2 2.0 8.6 f 0.5 8.7 0.4 0.9
0 0.9 0 0 0 0 0 0
moles/vwlc g1ycopepGdc
2.0
3.1 2.7 2.1 2.2 6.8 0.4 0 0 0 0
0
3.9
3.8 3.5 0 0.6
5.0 2.2 0 1.6
4.8 2.1 5.0 2.0
12.5 12.5
0 3.0 0.8 1.0 2.1
2.9 2.2
a Following performate oxidation as described under “Methods.”
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7272
TABLE II
Amino acid analysis of Edman degradation steps of glycopeptide I
The dansyl Edman gave single spots for the first 4 residues of serine, threonine, proline and proline, respectively.
Amino acid
Initial 1st
Threonine 4.30 3.96 Serine............ 5.00 4.03 Proline. . 8.25 8.25 Galactosamine. 4.57 5.12
Edman step
2nd I I
3rd 4th
moles/molt @#tide
3.44 3.34 3.32 4.00 3.98 3.98 8.25 7.45 6.90 5.15 4.65 4.91
- I - 5th
2.74 3.98 6.85 4.71
TABLE III
Relative proportions of methylated sugars in glycopeplide I
The sum of 2,3,4,6-tetramethylgalactose and 3,4,6-trimethyl- IV-acetylgalactosamine was set to 5.0.
Galactose (2,3,4,6-Tetramethyl)
4.1
N-Methyl-N-acetylgalactosamine
3.4,6-Trimethyl 4.6-Dimethyl
0.9 2.8
only 1 residue of IV-acetylgalactosamine, substituted at position 3 (Z-N-methyl-N-acetyl-4,6-dimethylgalactosamine), appeared to have been destroyed out of a total of 4 such residues. This was especially surprising since N-acetylgalactosamine substi- tuted at position 3 is considered more labile to destruction during p elimination than is either terminal N-acetylgalactosamine or N-acetylgalactosamine substituted at positions 4 or 6 (14). It may be that the duration of the reaction is short enough to pre- clude extensive /3 elimination or that the conditions are less de- structive than those more commonly used.
Demonstration of Alkalin&zbile Linkages in Glycopeptide I-
Treatment of glycopeptide I with NaOH under various conditions established that the majority of N-acetylgalactosamine present on glycopeptide I was alkali labile indicating that it was bound to the peptide by 0-glycosidic linkages to serine or threonine (or both). A large number of different conditions were explored in an attempt to prevent destruction of sugars by the so-called “peel reaction” (14, 15) as well as nonspecific destruction of amino acids. Although sugar destruction could in large part be avoided, what appeared to be nonspecific destruction of threo- nine could not, be prevented.
A time course for the treatment of glycopeptide I at, 45” with 0.1 N NaOH, 0.4 M NaB(*H)4 is shown in Fig. 3. Within 24 hours 3 of the 5 N-acetylgalactosamine residues had disappeared. Two of these N-acetylgalactosamine residues were converted to N-acetylgalactosaminitol while the third was destroyed. As the incubation continued beyond 24 hours the amount of galactose slowly dropped from 4.1 to 3.4 residues and 0.7 residue of galacti- to1 was found, probably the result of a peeling reaction where the underlying N-acetylgalactosamine was destroyed. The incorpo- ration of aH continued for a full 24 hours after the destruction of N-acetylgalactosamine had stopped. This could reflect such processes as the reduction of dehydroalanine to alanine and aminocrotonic acid to aminobutyric acid which may occur at a much slower rate.
In order to characterize the products of the alkaline degrada- tion reaction, the incubation mixture was neutralized with glacial acetic acid, evaporated, and the boric acid in the residue was
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from
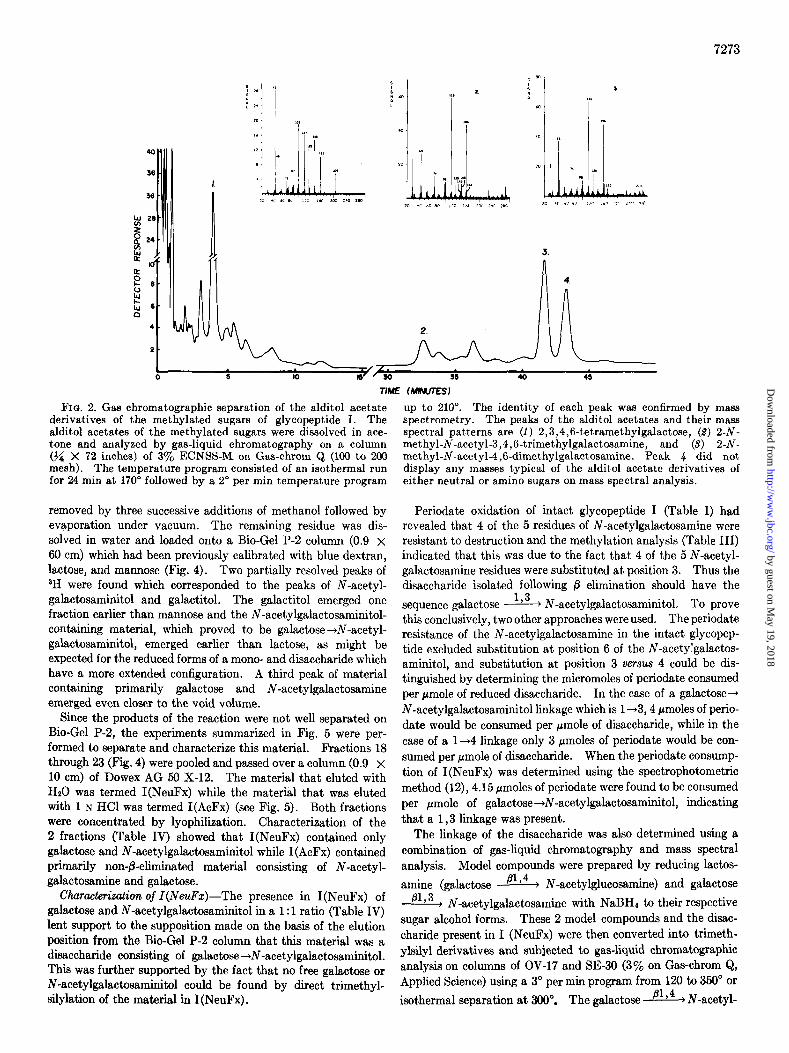
7273
FIQ. 2. Gas chromatographic separation of the alditol acetate derivatives of the methylated sugars of glycopeptide I. The alditol acetates of the methylated sugars were dissolved in ace- tone and analyzed by gas-liquid chromatography on a column (?/a X 72 inches) of 3% ECNSS-M. on Gas-chrom Q (100 to 200 mesh). The temperature program consisted of an isotherma. run for 24 min at 170” followed by a 2” per min temperature program
removed by three successive additions of methanol followed by evaporation under vacuum. The remaining residue was dis- solved in water and loaded onto a Bio-Gel P-2 column (0.9 x 60 cm) which had been previously calibrated with blue dextran, lactose, and mannose (Fig. 4). Two partially resolved peaks of aH were found which corresponded to the peaks of N-acetyl- galactosaminitol and galactitol. The galactitol emerged one fraction earlier than mannose and the N-acetylgalactosaminitol- containing material, which proved to be galactose+N-acetyl- galactosaminitol, emerged earlier than lactose, as might be expected for the reduced forms of a mono- and disaccharide which have a more extended configuration. A third peak of material containing primarily galactose and N-acetylgalactosamine emerged even closer to the void volume.
Since the products of the reaction were not well separated on Bio-Gel P-2, the experiments summarized in Fig. 5 were per- formed to separate and characterize this material. Fractions 18 through 23 (Fig. 4) were pooled and passed over a column (0.9 x 10 cm) of Dowex AG 50 X-12. The material that eluted with Hz0 was termed I(NeuFx) while the material that was eluted with 1 N HCl was termed I(AcFx) (see Fig. 5). Both fractions were concentrated by lyophilization. Characterization of the 2 fractions (Table IV) showed that I(NeuFx) contained only galactose and N-acetylgalactosaminitol while I(AcFx) contained primarily non-&eliminated material consisting of N-acetyl- galactosamine and galactose.
Characterization of I(NeuFz)-The presence in I(NeuFx) of galactose and N-acetylgalactosaminitol in a 1: 1 ratio (Table IV) lent support to the supposition made on the basis of the elution position from the Bio-Gel P-2 column that this material was a disaccharide consisting of galactose+N-acetylgalactosaminitol. This was further supported by the fact that no free galactose or N-acetylgalactosaminitol could be found by direct trimethyl- silylation of the material in I(NeuFx) .
up to 210”. The identity of each peak was confirmed by mass spectrometry. The peaks of the alditol acetates and their mass spectral patterns are (1) 2,3,4,6-tetramethylgalactose, (9) 2-N- methyl-N-acetyl-3,4,6-trimethylgalactosamine, and (3) 2-N- methyl-N-acetyl-4,6-dimethylgalactosamine. Peak 4 did not display any masses typical of the alditol acetate derivatives of either neutral or amino sugars on mass spectral analysis.
Periodate oxidation of intact glycopeptide I (Table I) had revealed that 4 of the 5 residues of N-acetylgalactosamine were resistant to destruction and the methylation analysis (Table III) indicated that this was due to the fact that 4 of the 5 N-acetyl- galactosamine residues were substituted at position 3. Thus the disaccharide isolated following /3 elimination should have the
sequence galactose 1,3 , N-acetylgalactosaminitol. To prove this conclusively, two other approaches were used. The periodate resistance of the N-acetylgalactosamine in the intact glycopep- tide excluded substitution at position 6 of the N-acetylgalactos- aminitol, and substitution at position 3 versus 4 could be dis- tinguished by determining the micromoles of periodate consumed per pmole of reduced disaccharide. In the case of a galactose-, N-acetylgalactosaminitol linkage which is l-+3,4 pmoles of perio- date would be consumed per pmole of disaccharide, while in the case of a l-+4 linkage only 3 pmoles of periodate would be con- sumed per pmole of disaccharide. When the periodate consump- tion of I(NeuFx) was determined using the spectrophotometric method (12), 4.15 pmoles of periodate were found to be consumed per pmole of galactose+N-acetylgalactosaminitol, indicating that a 1,3 linkage was present.
The linkage of the disaccharide was also determined using a combination of gas-liquid chromatography and mass spectral analysis. Model compounds were prepared by reducing hctos-
amine (galactose @lp4 > N-acetylglucosamine) and galactose 81,3 > N-acetylgalactosamine with NaBHd to their respective
sugar alcohol forms. These 2 model compounds and the disac- charide present in I (NeuFx) were then converted into trimeth- ylsilyl derivatives and subjected to gas-liquid chromatographic analysis on columns of OV-17 and SE-30 (3% on Gas-chrom Q, Applied Science) using a 3” per min program from 120 to 350” or isothermal separation at 300”. The galactose ” ) 4 + N-acetyl-
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7274
IO 20 30 40 50 60 HOURS
FIQ. 3. Alkaline treatment of glycopeptide I. Glycopeptide I (2.5 rmoles) was incubated in 0.1 N NaOH, 0.4 M NaB(3H)g at 45”. Aliquots for scintillation counting and sugar analysis were taken at the times indicated and were acidified with glacial acetic acid followed by lyophilieation. This was repeated three times with methanol-acetic acid (l:l, v/v) to remove free 3H. The samples were then counted for 3H incorporation (O-O) and assayed by gas-liquid chromatography for galactose (A-A), N-acetylgalactosamine (+H), galactitol (A-A), and N- acetylgalactosaminitol (cl----O).
glucosaminitol, the galactose ” ’ 3 ) N-acetylgalactosaminitol, and the disaccharide from glycopeptide I all had identical exit times under both sets of conditions on both SE-30 and OV-17. All three compounds were then subjected to mass spectral anal-
ysis. Both model compounds were predicted to have a base
peak at M 946, the galactose ” j4 b N-acetylglucosaminitol
a unique mass at M 858 and the galactose p1’3 + N-acetyl- galactosaminitol a unique mass at M 756 as shown in Fig. 6. The two unique masses were found to be mutually exclusive for the model compounds (Table V), and the P-eliminated disac- charide produced by treatment of glycopeptide I with alkali had
a pattern identical with that of galactose ” ‘3 + N-acetylga- lactosaminitol, in agreement with the periodate consumption and methylation results presented above.
Eacheric@ coli /3-galactosidase released 72% of the galactose under the assay conditions while a-galactosidase isolated from Ficin was completely inactive against the disaccharide. The finding that the galactose is linked fil,3 to the N-acetylgalactos- amine explains the almost complete resistance of the intact glycopeptide to release of galactose by fl-galactosidase isolated from jack bean meal since this enzyme shows very little activity toward /31,3 as opposed to /31,4 and pl,6 linkages (16).
In summary, alkaline treatment of glycopeptide I, using the con- ditions described in Fig. 3, releases 2.3 moles per mole of glycopep-
tide I of a disaccharide with the sequence galactose b1,3 , N-
12 14 16 16 20 22 24 26 26 30
.6
.2
FRACTION
FIN. 4. Separation on Bio-Gel P-2 of products resulting from alkaline treatment of glycopeptide I. Glycopeptide I (2.5 pmoles) was incubated for 66 hours at 45” in 0.1 N NaOH and 0.4 M NaB(‘H)d as described in Fig. 3. The reaction containing the /3 elimination products was then neutralized with glacial acetic acid, dried, and washed and dried 3 times with methanol-acetic acid (l:l, v/v). The sample was then taken up in 1 ml of H20 and loaded onto a column of Bio-Gel P-2 (0.9 X 60 cm) and eluted with HzO. All fractions (1.2 ml each) were assayed for the presence of aH (O-O). Fractions 12 to f8 were assayed for phenol-sulfuric acid reactive material and found to contain none. Fractions 19 to 8’ were assayed by gas-liquid chromatography for galactose (A-A), N-acetylgalactosamine (*u), galactitol (A-A), and N-acetylgalactosaminitol (O-O).
acetylgalactosamine. In addition, approximately 0.7 mole per mole of glycopeptide I of disaccharide is destroyed by the peel reaction, resulting in the appearance of 0.7 mole of galactitol.
Identification of Amino Acid Residues Involved in 0-Glycosidic
Linkage with Alkali-labile Gal g1 ’ 3 ) N-Acetylgalactosamine Units-Alkaline treatment of glycopeptide I in the pres- ence of 0.4 M NaBHI resulted in the destruction of 3 residues of serine and 2 to 3 residues of threonine (Ta- ble I). Since only 3 disaccharide units were fl elimi- nated, this represented more destruction of amino acids than could be accounted for by j3 elimination alone. The appearance of 0.9 residue of alanine subsequent to alkaline degra- dation suggested that at least 1 of the P-eliminated oligosaccha- ride units was linked to a serine. The failure to detect any cr-aminobutyric. acid following /3 elimination could result from either a lack of 0-glycosidic bonds to threonine or the inability of the NaBH4 to effectively reduce the a-aminocrotonic acid. Poor reduction of a-aminocrotonic acid to a-aminobutyric acid has been observed and has resulted in the use of paladium chlo- ride to improve reduction (17). We attempted to circumvent the problem of nonspecific destruction by carrying out the ahca- line degradation in the presence of NaGSO as described by Spiro (11). Two of the 5 residues of N-acetylgalactosamine remained intact while 2.7 residues appeared in the form of sul- fonylgalactosamine on amino acid analysis. The appearance of almost 3 residues of sulfonylgalactosamine is in good agreement with Winzler’s observation that 0-glycosidically linked N-acetyl-
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7275
FIG. 5. Scheme for the isolation of alkaline degradation prod- ucts of glycopeptide I. Although only 0.9 of an expected 3.0 residues of alanine was obtained upon alkaline-borohydride treat- ment of glycopeptide I (see Table I), all of the serine residues in- volved in 0-glycosidic linkage are shown as being converted to alanine. Since the destruction of threonine did not appear to be due to involvement in 0-glycosidic linkage, no indication of de- struction of threonine is made in the figure following alkaline treatment. NAcGalmol, N-acetylgalactosaminitol.
TABLE IV
Composition of isolated products of alkaline treatment of glycopeptide I
Fractions 18 to 23 from the Bio-Gel P-2 column (Fig. 4) were loaded on a Dowex AG 50 column (0.9 X 10 cm). The material eluted with 10 volumes of HsO was termed I(NeuFx) and the material eluted with 5 volumes of 1 N HCl was termed I(AcFx).
Component I(NeuFx) I(AcFx)
moles/molt glycoPePlide I
Galactose...................... 2.3 1.7 Galactitol....................... 0 0 N-Acetylgalactosamine 0 2.0 N-Acetylgalactosaminitol. . 2.3 0.4
galactosamine substituted at position 3 gives a high yield of a sulfonyl derivative on /I elimination in the presence of NaaSOr, while N-acetylgalactosamine substituted at position 6 yields 15% and at position 4 no sulfonylgalactosamine derivative upon ,!I elimination (18). In addition, 3.1 residues of cysteic acid or cY-amino-&sulfonylbutyric acid (or both) emerged just before the sulfonylgalactosamine on amino acid analysis. This con- firmed that only 3 amino acids were involved in 0-glycosidic linkages subject to /3 elimination, but did not offer information as to the identity of the amino acids since cysteic acid and (Y- amino-/?-sulfonylbutyric acid do not separate on the long column of the amino acid analyzer under the conditions used (18). How- ever, paper chromatography in 1-butanol-acetic acid-Hz0 (4 : 1: 5) of the acidic amino acids isolated by passage over Dowex AG l-X8 as described under “Methods” revealed a single spot which
Hz-C - 0 - si w3
Si(Me13-0 - C - H
o-----C - H
FIG. 6. Unique masses expected from trimethylsilyl derivative of 1,3- (upper) as opposed to 1,4- (lower) linked galactose-+N- acetylgalactosaminitol.
TABLE V
Determination of galactose+N-acetylgalactosaminitol linkage by mass SpeCtTOSCOpy
Trimethylsilyl derivatization was carried out on the two-model disaccharides and I(NeuFx) using pyridine-trimethylchloro- silane-hexamethyldisilazane (5:2: 1). The samples were sub- jected to gas-liquid chromatography and mass spectral analysis as described in the text. The primary ion peak at m/e 946 (M - 15) was set equal to 1.00 and the relative proportions of the unique peaks (see Fig. 6) at m/e 756 and 858 were determined.
Trimethylsilyl derivative of
Relative proportions of masac at m/c
756
Galactose ‘lT3 + N-acetylgalactosaminitol 3.33
858 946 --
0.00 1.00
Galactose 81,4 p N-acetylglucosaminitol. . 0.00 2.17 1.00
Galactose - N-acetylgalactosaminitol from glycopeptide I. 3.64
I 0.25 1.00
co-migrated with cysteic acid. Therefore, all 3 residues of N- acetylgalactosamine which can be /? eliminated are O-glycosidi- tally linked to glycopeptide I at serine residues. The reason for the destruction of 2 residues of threonine is not apparent. Since no cu-amino+-sulfonylbutyric acid was obtained with alkaline sulfate treatment and no a-aminobutyric acid was ob- tained (data not shown) following reduction with palladium chloride as described by Downs et a2. (17), it seems very unlikely that any of the threonine was involved in 0-glycosidic linkage to N-acetylgalactosamine.
Characterization of Non-&eliminated carbohydrate Material
Found in I(AcFz)-The absence of appreciable amounts of N- acetylgalactosaminitol in I(AcFx) eluted from Dowex AG 50 with 1 N HCl (Table IV and Fig. 5) indicated that the 2 N-acetyl- galactosamine residues present were still linked to the peptide. This raised two questions: (a) which amino acid(s) was involved
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7276
TABLE VI Effect of alkaline treatment and periodate oxidation on Z(AcFx) and Acetylated Z(AcFx)
I(AcFx) was acetylated as described under “Methods.”
Component
Composition of I(AcFx) Composition of acetylated I(AcFx)
Native NaIO4 oxidation 0.1 N NaOH, 0.4 P 0.1 N NaOH, 0.4 P NaBH,, 48 hrs, 45’ NaBH4, then NaIO, Control’ 0.1 N NaOH, 0.5 Y
NaaSOa, 24 hrs, 40’
Galactose N-Acetylgalactosamine N-Acetylgalactosaminitol Cysteic acid. Sulfonylgalactosamine Threonine Serine..................... Proline. Glycine Alanine
.
.
1.7 2.0 0.4
0 0
1.2 2.0 8.6 0.9 1.4
-
0 1.0 0
1.3 2.0
0 0
1.0 1.9 8.4 2.1 1.3
1.0 1.0
1.3 0
0.6 0.9 8.3 2.0 1.2
Q The control differed from the 0.1 N NaOH-0.5 M NazS03 incubation only in that it was immediately acidified with 6 N HCl instead of being incubated for 24 hours at 40”.
in the linkage to the peptide and (b) what was the nature of the linkage(s) which resulted in resistance to /3 elimination?
The amino acid composition of I(AcFx) (Table VI) was vir- tually identical with that of the product of alkaline degradation before fractionation on Bio-Gel P-2 and Dowex AG-50 (Table I), indicating that the entire peptide was present. Treatment of glycopeptide I (AcFx) with sodium metaperiodate resulted in destruction of all of the galactose, and 1 of the 2 residues of N- acetylgalactosamine (Table VI). This suggested that the residue of terminal N-acetylgalactosamine observed in the intact glyco- peptide was completely resistant to ,f3 elimination. If we sub- tract an amount of galactose equal to the small amount of con- taminating N-acetylgalactosaminitol in I (AcFx) (assuming that
all the N-acetylgalactosaminitol arises from galactose a,3 ,
N-acetylgalactosaminitol) there remains only 1.3 residues of galactose associated with the peptide, presumably linked to posi- tion 3 of the periodate-resistant N-acetylgalactosamine of I(AcFx). This was confirmed by methylation of I(AcFx) which yielded 1 residue of terminal N-acetylgalactosamine and 0.8 resi- due of N-acetylgalactosamine substituted at position 3. Since E. coli fl-galactosidase was able to release the galactose residue present in I(AcFx), there must be two O-glycosldically linked units in I(AcFx) with the structures N-acetylgalactosamine +
peptide and galactose ” j3 + N-acetylgalactosamine -+ pep- tide.
The question then remained as to why these 2 oligosaccharide units were resistant to /3 elimination. The most likely explana- tion is suggested by the sequence of glycopeptide I shown in Fig. 1, which would place a serine residue at both the COOH and NH2 termini of the glycopeptide. Since it has been estab- lished that fi elimination from serine and threonine will not occur if either the COOH or NH2 functions are free, it would be ex- pected that any 0-glycosidically linked units on either terminal serine would be resistant to 0 elimination (19).
To confirm this postulate, I(AcFx) was acetylated in 2% aque- ous acetic anhydride in 1 M sodium bicarbonate and was reiso- lated by passage over Dowex AG 50 (Fig. 5). As with intact glycopeptide I, acetylation of I(AcFx) resulted in elution of the glycopeptide in the Hz0 wash rather than the acid wash, indi- cating that acetylation had indeed taken place. Treatment of the acetylated I(AcFx) with 0.1 N NaOH and 0.4 M NaBH, led
to a decrease in the amount of N-acetylgalactosamine from 2.0 to 1.3 residues (Table VI). The expected increase in N-acetyl- galactosaminitol could not be quantitated due to an interfering peak on gas-liquid chromatographic analysis. Periodate oxida- tion of the product of this alkali treatment led to the destruction of only 0.3 of the 1.3 remaining residues of N-acetylgalactosamine (Table VI) indicating that it was the terminal N-acetylgalactos- amine which was located on the NHt-terminal serine. Further support for this location for the terminal N-acetylgalactosamine was obtained by treatment of another preparation of acetylated I(AcFx) with 0.1 N NaOH and 0.5 M Na2S03 at 40” for 24 hours (Table VI). Under these conditions 1 residue of N-acetylgalac- tosamine was destroyed as was 1 residue of serine and 0.4 residue of threonine (nonspecific destruction in the case of threonine). Although 1.3 residues of cysteic acid appeared on amino acid analysis, no sulfonylgalactosamine was detected, suggesting that it was the terminal N-acetylgalactosamine which had been @
eliminated since galactose ” ’ 3 tN-acetylgalactosamine would have yielded some sulfonylgalactosamine upon /3 elimination.
Several attempts to add a residue of glycine to the COOH terminus using 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide and glycine methylester were unsuccessful, which may reflect the presence of the GalL+GalNAc-+ on the COOH terminus. In any case the presence of 0.9 residue of serine and only 0.6 residue of threonine in the product of alkaline treatment of acetylated I(AcFx) (Table VI) suggests that it is the serine which is in- volved in the 0-glycosidic linkage since there is less than a full residue of threonine.
Experiments with ZgA (Oso)-Due to a very limited amount of the hinge region glycopeptide from IgAi (Oso) only a few ex- periments could be performed. Carbohydrate analysis revealed that this glycopeptide also contained 4 residues of galactose and 5 residues of N-acetylgalactosamine (Table I). Treatment with alkali (Table I) indicated that at least three of the 5 N-acetyl- galactosamine residues were 0-glycosidically linked to the pep- tide. Of the recovered 1.6 residues of N-acetylgalactosaminitol, 0.5 residue was in the form of free N-acetylgalactosaminitol and
the other 1.1 residues were found as galactose ’ ’ 3 ) N-acetylga- lactosaminitol as determined by direct trimethylsilylation and mass spectral analysis. In addition, the appearance of three resi- dues of alanine following @ elimination suggested that the three @
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7277
eliminated oligosaccharides were linked to serine and that the destruction of threonine was again nonspecific. The incom- plete p elimination may reflect the fact that the alkaline treat- ment was not continued for an adequate amount of time. (The small amount of peptide prevented us from doing a time course experiment.) The presence of 0.5 residue of free N-acetylgalac- tosaminitol suggests that if the same serine is involved in carrying the terminal N-acetylgalactosamine in IgA(Oso) as in IgA (Walker), /3 elimination is now possible because the serine in this tryptic peptide is not terminal.
DISCUSSION
Of the 5 immunoglobulin classes known to be present in man, only IgD2 (20) and IgAi (20, 21) have both N-acetylgalactos- amine and N-acetylglucosamine-containing oligosaccharide units. The presence of N-acetylgalactosamine was suggested by Abel and Grey (22) to be characteristic of the hinge region of IgAi as opposed to the hinge region of IgA2. The amino acid sequences reported by Frangione and Wolfenstein-Todel (2) for the hinge regions of IgAl and IgAz (see Fig. 1) showed that the IgA2 hinge peptide differed from that of IgAl by a gap of 12 to 13 residues which appeared to be the region in which the carbohydrate of IgAi was located. The sequence of this region is very unusual in that it consists of 2 identical stretches of 7 to 8 residues. The unusual amino acid composition of glycopeptide I and the pres- ence of N-acetylgalactosamine indicated that it was a part of this same region. Determination of the sequence of the first 5 resi- dues of this glycopeptide allowed us to align it with the sequence reported by Frangione and Wolfenstein-Todel for IgAi (see Fig. 1). The location of the oligosaccharide units of glycopeptide I clearly shows that IgAz could not contain any of these units be- cause all of the serines involved in 0-glycosidic linkages to this region are absent in IgAs. In keeping with the highly unusual symmetry of the amino acid sequence of this region the carbo- hydrate is also symmetrically distributed except for the terminal N-acetylgalactosamine present on the amino terminal serine of glycopeptide I. Since the hinge region of the IgA (Oso) glyco- peptide also contains 5 N-acetylgalactosamine and 4 galactose residues, it may be that this terminal N-acetylgalactosamine is a fixed feature of this region and not due to incompletion. Other reports on the N-acetylgalactosamine content of isolated IgAi hinge region glycopeptides are in good agreement with our fmd- ings. Abel and Grey (22) found 4 residues of N-acetylgalactos- amine to be present while Moore and Putnam (23) reported finding 7 and 5 residues of N-acetylgalactosamine per leucine in two diierent preparations of hinge region peptide. Thus it appears that all IgAi hinge regions may contain 0-glycosidically linked oligosaccharide units on each of the 5 serine residues pres- ent. It is not clear why analyses of the carbohydrate composi- tion of other IgAl myeloma proteins (20, 24) have been reported to give considerably lower values for N-acetylgalactosamine content per heavy chain than those just cited for the isolated hinge region peptides. These other IgAi myelomas may differ from those just discussed or there may be some difficulty inherent in determining the carbohydrate content of whole IgAi mye- lomas.
In comparing our findings with those of Dawson and Clamp (25), it is of interest to note that their N-acetylgalactosamine- containing glycopeptide also appeared larger than their N-acetyl- glucosamine-containing glycopeptides on gel filtration over Sephadex G-25. The molecular weight of 2750 obtained by sedimentation equilibrium centrifugation by these authors is
* J. Baenziger and S. Kornfeld, unpublished observation.
very similar to the value of 2720 found by us using exclusion chromatography. On the other hand, the value of 2350 ob- tained by freezing point depression osmometry is very low and led them to suggest a composition of galactose-N-acetylgalac- tosamine-serine-threonine-proline of 3 : 3 :4 :4 : 5. The remarkable resemblance of the oligosaccharide units of the IgAi hinge region to those reported by DeVries (26) for the freezing point depress- ing glycoproteins of fish (up to 500 times more potent than NaCl on a molar basis) may indicate the reason for this low value since an increase of only 50% in the freezing point-depressing activity of this glycopeptide over that expected would lead to a value of 2350 rather than the value of 3280 calculated from the composi- tion of glycopeptide I.
The glycopeptide isolated by Dawson and Clamp appears to be very similar to glycopeptide I with regard to its periodate sensitivity in that all the galactose and only 39% of the N-acetyl- galactosamine were destroyed by periodate. These authors also isolated a glycopeptide which contained sialic acid in addition to galactose and N-acetylgalactosamine, and in this case 28% of the galactose was resistant to periodate oxidation (25). We did not detect a sialic acid-containing form of glycopeptide I in IgAi (Walker).
The 0-glycosidically linked oligosaccharides of the IgAl hinge region are very similar in structure to the 0-glycosidically linked oligosaccharides found on the freezing point-depressing glyco- proteins of Antarctic fish (26), human gonadotropin (27,28), and the trypsin released glycopeptides of human erythrocytes (29) aside from the presence of sialic acid in the oligosaccharides of the latter 2 proteins.
In Fig. 7 we have summarized our findings regarding the oligo- saccharide units found on IgG, IgM, IgAi, and IgE myeloma proteins. IgAi is unique among these immunoglobulins in that it has 0-glycosidically linked oligosaccharide units. IgD mye- loma proteins have been observed by us2 and others (20) to con- tain a similar amount of N-acetylgalactosamine as IgAl, and it will be of interest to determine if the N-acetylgalactosamine found in IgD is also located in the hinge region and if the se- quence of the hinge region is homologous to that of IgAi.
Turning to the N-glycosidically linked oligosaccharides of the various immunoglobulins, several observations can be made. Only IgM and IgE appear&o have what we have termed high mannose oligosaccharides. There are 2 high mannose oligo- saccharides present on each heavy chain of IgM while in the case of IgE there is only 1 per heavy chain. The structure of the high mannose units of IgM differ in several respects from the one found on IgE. They display microheterogeneity resulting in the 3 structures shown in Fig. 7 and have a core consisting of Glc- NAc +GlcNAc AAsn. The high-mannose oligosaccharide found on IgE, on the other hand, displays no detectable microheterogene-
ity and has a core with the sequence: Man m Glc,Ac a
Man @& G~cNAc % Asn. This sequence is extremely unusual in having an a-linked N-acetylglucosamine. Since one of the high-mannose chains of IgM has been localized to an asparagine, 14 residues from the COOH terminus of the ~1 chain (Asn& (30) and the heavy chain of IgE is only 558 residues in length (31, 32), this high mannose chain cannot be analogous to the one found in IgE. Neither the high mannose chain of IgE nor the other high mannose chain of IgM has been localized, SO that no further conclusion can be made regarding any analogy in the location of these oligosaccharides. In this regard it is of interest to note that in the case of IgAi which has a heavy chain of the same length as IgM there is an asparagine at residue 563 which
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from


1 2
""k"0.
" ""k
"0."
8-l
E-2.
E-3
5.1.
02.6
1 G
1 Il.4
1
Gal
61.4
GlcN
Ac
( Gl
cNAc
6 1.2
1
Yan
1 81
.2
\/
Ma"
a 1.3
Ill.
6
GlcN
AcmM
an
' G1
~N,"4
1 61
.4 Gl
CNAC
1 *
AS" 1
Cons
tant
Fteg,o
n
27.R
esidu
e Pe
ptide
IIA
S.A.
S.
A.
0 2.6
1 Gil
I a
2.6
Gal
8 1.4
1
B 1.4
GlcN
Ac
I Gl
cNAc
.91,2
1 Ma
n )
61.2
\/
Man
a 1.3
a
1,6
Man I
81,4
GdNA
c 1 81
.4 Fu
ca~G
lcNAc
1
Unkn
own
C-l
0-G
LYC
OSI
DIC
Ga
lNAc
-O-&
r
Thr
Pro
Pro
Thr
Pro
Gal~G
alNAc
-O-S
er
Pr0
Gal)l'
JGAl
NAc-O
-SW
Thr
Pro
Pro
Thr
PT.3
G~I~,
JG~N
Ac-cI
-SU
PT.3
G=I~G
I~NAc
-o-S~
~
Hing
e Re
gion
of
a 1
Subty
pe
I
Fro.
7.
Sum
mar
y of
ol
igos
acch
arid
e st
ruct
ures
pr
esen
t on
IgG
(3
), IgM
(4
), Ig
Al
(l),
and
IgE
(5,
6)
mye
loma
prot
eins
.
by guest on May 19, 2018http://www.jbc.org/Downloaded from

7280
carries an oligosaccharide unit (32), but in IgAl the oligosac- charide is probably of the complex type (IIB and IIC) which contains both galactose and mannose (32). This indicates that carbohydrate structures located on homologous asparagines need not themselves be homologous in structure. This is a particu- larly interesting case since both IgM and IgAl have the triplet sequence of Asn-Val-Ser at this location and the plasma cells synthesizing the IgM protein clearly have the capacity for addi- tion of either a complex or a high mannose oligosaccharide unit. Obviously some more subtle aspect of the tertiary structure of the heavy chain must be involved in dictating which type of oligosaccharide will be synthesized at this location. This is further emphasized by the finding that the complex oligosaccha- ride present at Asnbe3 of IgAl may differ significantly from the one which is found on the 27-residue peptide which arises from some other location in the constant region of the heavy chain (32). One might speculate that since the high mannose chain of IgM and the complex type chain of IgAl are both located so close to the cysteine at the COOH terminus, which has been sug- gested to be involved in the binding of at least one of the IgA heavy chains to the J chain (33, 34), they may play some role in preventing the formation of hybrids of IgM and IgA monomers when they are condensed with J chains (35).
The complex type of N-glycosidically linked oligosaccharide units are found on all four types of myeloma proteins studied by US. They all appear to be very closely related in structure but do display important differences. In the case of IgG, additional experiments have allowed us to establish the more complete structures shown in Fig. 7. Most of the IgG myelomas which we studied had only 1 oligosaccharide unit per heavy chain (3). The majority of these were deficient in sialic acid and had only 1 residue of galactose. Those IgG myelomas which had 2 oligosaccharide units per heavy chain (Hum and Mul) also had additional complex type oligosaccharides which contained 2 residues of galactose and 1 or 2 residues of sialic acid. In the case of Hum B2 and Hum B3, both of which had 2 residues of sialic acid, there was a trisubstituted mannose in the core and terminal N-acetylglucosamine present in equal amounts. Thus Hum B2 and Hum B3 may be identical in structure with glyco- peptide IIA of IgAl except that they have a full residue of fucose and 2 residues of sialic acid.
Therefore the complex oligosaccharide units of IgG, IgA, and IgE are very closely related. Differences reside in the presence or absence of fucose in the core, the presence or absence of a third nonreducing terminus consisting of an N-acetylglucosamine linked to the core mannose, and the number of galactose and sialic acid residues present. As has been discussed previously (6) these differences may not reflect microheterogeneity in the sense of incompletion so much as glycopeptides which have arisen from different loci along the respective heavy chains. The fact that the monosialylated oligosaccharides of IgAl and IgE (IIA and B-l, respectively) only have sialic acid present on the nonreducing terminus arising from position 3 of the core mannose and the fact that IIA of IgAl is completely devoid of fucose but has a full residue of N-acetylglucosamine arising from the core mannose certainly lend support to this view.
The complex oligosaccharides of IgM, on the other hand, ap- pear to differ from the structures just discussed. The most obvious difference is in the presence of peripheral N-acetylglucos- amine residues which are substituted at position 6 rather than position 4. In addition, the complex oligosaccharide thought to arise from the Fd portion of the p chain (4) has a terminal fourth residue of mannose arising from the core.
The finding that the complex oligosaccharides of IgG, IgAl, and IgE and probably both the complex and high mannose oligosac- charides of IgM have the core structure:
Man
I l31,4 GlcNAc
1 Bl,4 GlyNAc
t B Asn
is very compatible with the present belief that an oligosaccharide- lipid intermediate may be involved in the synthesis of the core portions of protein-bound oligosaccharides (36, 37). If this is indeed the case, it would suggest that the differences observed among these structures is a function of the transferases thought to transfer sugar from the nucleotide sugars to the core unit after it has been transferred to the peptide (38). In addition, the transferase involved in transfer of the core from the oligo- saccharide-lipid to the asparagine would not need to be specific; in other words, the same enzyme could transfer the same core to various locations along the peptide backbone. The further addition of mannose for high mannose chains or varying amounts of N-acetylglucosamine, galactose, fucose, and sialic acid for complex oligosaccharides would then have to be a function of the individual glycosyl transferases whose specificity would be affected by the local environment of the core molecule. This mechanism could explain the presence of a high mannose chain at Asnst3 in IgM and a complex type chain at the same location in IgAl. The major exception to this synthetic pathway would be the high mannose oligosaccharide of IgE C-l since it does not have the typical chitobiose core structure. In this case a differ- ent oligosaccharide-lipid intermediate would have to be synthe- sized and a different transferase would have to transfer this core unit to the correct location along the peptide backbone. Alter- natively the individual sugars could be added directly to the peptide from nucleotide sugars without the benefit of an oligo- saccharide-lipid intermediate for synthesis of the core.
It is of interest that one of the oligosaccharides which is ac- cumulated in large amounts in the liver of patients with G,l- gangliosidosis, Type I (39) has a structure virtually identical to that of the complex oligosaccharides of immunoglobulins aside from the absence of sialic acid and fucose. This may be an indication that the liver is a site of immunoglobulin metabolism and that immunoglobulins may contribute a major portion of the oligosaccharide material present in this disease.
Our studies therefore establish that the oligosaccharide units found on IgG, IgM, IgAl, and IgE differ as regards the type of structures present (0-glycosidic, N-glycosidic, complex, and high mannose), the number of each type found per heavy chain, and in the actual structures of closely related oligosaccharide units. We have also presented evidence suggesting that complex type oligosaccharides arising from different loci on a given heavy chain show differences in their structures. All of these differ- ences suggest that the carbohydrate moieties may play an as yet
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7281
undefined role in the biologic functioning of each immunoglobu- lin class.
Acknowledgments-We would like to thank Dr. R. W. Brad- shaw for allowing one of us (J.B.) to carry out the amino acid sequence analysis of glycopeptide I in his laboratory. We would also like to thank Dr. Luis Glaser for his help in interpreting the mass spectral patterns shown in Table V and Fig. 6.
Note Ad&d-Recently Bahr-Lindstrom and Bennich (40) have succeeded in localizing the high manose oligosaccharide of IgE and Putnam et al. (41) have now localized both high mannose oligosaccharides of IgM. It is of interest to note that one of the high mannose units of IgM is located on an asparaaine which is
21.
22.
23.
24.
25.
homologous to the asparagine to which the high IgE is linked.
REFERENCES
mannose unit of 26.
27. 28.
1. BAENZIGER, J., AND KORNFELD, S. (1974) J. 7260-7269
2. FRANGIONE. B.. AND WOLFENSTEIN-TODEL. Nat. Acad. SC>. U. S. A. 69, 3673-3676 ’
3. KORNFELD, R., KELLER, J., BAENZIGER, J., S. (1971) J. Biol. Chem. 246, 3259-3268
4. HICKMAN. S.. KORNFELD, R., OSTERLAND, C. K., AND KORN-
18.
19.
20.
Biol. Chem. 249, 29.
C. (1972) Proc. 30.
AND KORNFELD, 31.
FELD, s. (i972) J. Bioi. Chem. 24’7, 21562163 5. BAENZIGER, J., KORNFELD, S., AND KOCHWA, S. (1974) J.
Biol. Chem. 249, 1889-1896 6. BAENZIGER, J., KORNFELD, S., AND KOCHWA, S. (1974) J.
Biol. Chem. 249, 1897-1903 7. WATERFIELD, M., AND HABER, E. (1970) Biochmistry 9, 832 8. KRETSINGER, R. H., NOCKOLD, C. E., COFFEE, C. J., AND
BRADSHAW, R. A. (1971) Cold Spring Harbor Symp. Quant. Biol. 36, 217
9. BRADSHAW, R. A., BABIN, D. R., NOMOTO, M., SRINIVASIN, N. G., ERICSSON, L. H., WALSH, K. A., AND NEURATH, H. (1969) Biochemistry 8, 3859
10. HIRS, C. W. H. (1967) Methods Enzymol. 11, 198 11. SPIRO, R. G. (1972) Methods Enzymol. 28, 35-37 12. DIXON, J. S., AND LIPKIN, D. (1954) Anal. Chem. 26, 1092 13. BHATTI, T., AND CLAMP, J. R. (1968) Biochim. Biophys. Acta
170, 266-208 14. LLOYD, K. O., AND KABAT, E. A. (1969) Curbohydr. Res. 9, 41-
48 15. BALLOU, C. E. (1954) Advan. Carbohydr. Chem. 9, 91 16. LI, Y. T., AND LI, S. C. (1972) Methods Enzymol. 28, 713 17. DOWNS, F., HERP, A., MOSCHERA, J., AND PIGMAN, W. (1973)
Biochim. Biophys. Acta 328, 182-192
32.
33.
34.
35.
36.
37.
38. 39.
40.
41.
WEBER, P., AND WINZLER, R. J. (1969) Arch. Biochem. Biophys. 129. 534638
DEREVITSKAYA, V. A., VAFINA, M. G., AND KOCHETKOV, N. K. (1962) Carbohydr. Res. 3, 377
CLAMP, J. R., AND JOHNSON, I. (1972) in Glycoproteins (GOTT- SCHALK, A., ed) 2nd Ed, Part A, pp. 636631, Elsevier, Amsterdam
GREY, H. M., ABEL, C. A., AND ZIMMERMAN, B. (1971) Ann. N. Y. Acad. Sci. 190, 32-48
ABEL, C. A., AND GREY, H. M. (1971) Nature New Biol. 288, 29-31
MOORE, V., AND PUTMAN, F. W. (1973) Biochemistry 12, 2361- 2370
TOMANA, M., NIEDERMEIER, W., MESTECKY, J., AND HAMMACK, W. J. (1972) Immunochemistru 9. 933-940
DAW~ON; G.,‘AND CLAMP, J. g. (1968) Biochem. J. 107, 341- 352
LIN, Y., DUMAN, J. G., AND DEVRIES, A. L. (1972) Biochem. Biophys. Res. Commun. 46, 87-92
BAHL, 0. P. (1969) J. Biol. Chem. 244,575-583 CARLSEN, R. B., BAHL, 0. P., AND SWAMINATHAN, N. (1973)
J. Biol. Chem. 248, 6810-6827 WINZLER, R. J. (1969) in Red Cell Membrane: Structure and
Function (JAMIESON, G. A., AND GREENWALT, T. J., eds) p. 157, J. B. Lippincott Co., Philadelphia
SHIMIZU, A., PUTNAM, F. W., PAUL, C., CLAMP, J. R., AND JOHNSON, I. (1971) Nature New Biol. 231, 73-76
BENNICH, H., MILSTEIN, C., AND SECHER, P. S. (1973) Fed. Eur. Biochem. Sot. Lett. 33, 49-53
PUTNAM, F. W., Low, T., LIU, V., HUSER, H., RAFF, E., WONG, F. C., AND CLAMP. J. R. (1973) in International Sym- posium on the Immunoglobulin A System (KRAUS, F. W., ed) Birmingham, Alabama
MENDEZ, E., PRELLI, F., FRANGIONE, B., AND FRANKLIN, E. C. (1973) Bioehem. Biophys. Res. Commun. 56, 1291-1297
MESTECKY, J., SCHROHENLOHER, R. E., KULHAVY, R., WRIGHT, G. P., AND TOMANA, M. (1974) Proc. Nat. Acad. Sci. U. S. A. 71, 544-548
DELLA CORTE, E., AND PARKHOUSE, R. M. E. (:973) Biochem. J. 136, 597-606
BAYNES, J. W., Hsu, A.-F., HEATH, E. C. (1973) J. Biol. Chem. 248, 5693-5704
BEHRENS, N. H., CARMINATTI, H., STANELONI, J., LELOIR, L., AND CANTARELLA, A. I. (1973) Proc. Natl. Acad. Sci. U. S. A. 70, 3390-3394
ROSEMAN, S. (1970) Chem. Phys. Lipids 6, 270 WOLFE, L. S., SENIOR, R. G., AND NG YING KIN, N. M. K.
(1974) J. Biol. Chem. 249, 1828-1838 BAHR-LINDSTR~M, H., AND BENNICH, H. (1974) Fed. Eur.
Biochem. Sot. Lett. 40, 57-61 PUTNAM, F. W., FLORENT, G., PAUL, C., SHINODA, T., AND
SHIMIZU, A. (1973) Science 182, 287
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Jacques Baenziger and Stuart KornfeldOF THE O-GLYCOSIDICALLY LINKED OLIGOSACCHARIDE UNITS
Immunoglobulin : II. STRUCTURE1Structure of the Carbohydrate Units of IgA
1974, 249:7270-7281.J. Biol. Chem.
http://www.jbc.org/content/249/22/7270Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/249/22/7270.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from