structural and biochemical basis for intracellular …evolve.harvard.edu/125-inhibitor.pdfarticle...
TRANSCRIPT

Article
Structural and Biochemical Basis for Intracellular
Kinase Inhibition by Src-specific PeptidicMacrocyclesGraphical Abstract
Highlights
d Peptidic macrocycles selectively inhibit Src kinase
d Compounds block ATP and peptide binding site
d Compounds inhibit activity of Src kinase in cells
d Migration of breast tumor-derived cells is inhibited
Aleem et al., 2016, Cell Chemical Biology 23, 1103–1112September 22, 2016 ª 2016 Elsevier Ltd.http://dx.doi.org/10.1016/j.chembiol.2016.07.017
Authors
Saadat U. Aleem, George Georghiou,
Ralph E. Kleiner, ..., W. Todd Miller,
David R. Liu, Markus A. Seeliger
[email protected] (D.R.L.),[email protected](M.A.S.)
In Brief
Aleem et al. show the structural basis for
the selective inhibition of c-Src kinase by
a macrocycle with activity in cells. This
demonstrates that synthetic macrocycles
can be engineered to become potent
inhibitors of intracellular enzymes.
Accession Numbers
5BMM

Please cite this article as: Aleem et al., Structural and Biochemical Basis for Intracellular Kinase Inhibition by Src-specific Peptidic Macrocycles, CellChemical Biology (2016), http://dx.doi.org/10.1016/j.chembiol.2016.07.017
Cell Chemical Biology
Article
Structural and Biochemical Basis for IntracellularKinase Inhibition by Src-specificPeptidic MacrocyclesSaadat U. Aleem,1,5 George Georghiou,2,5 Ralph E. Kleiner,3,4,5,6 Kip E. Guja,2 Barbara P. Craddock,1 Agatha Lyczek,2
Alix I. Chan,3,4 Miguel Garcia-Diaz,2 W. Todd Miller,1 David R. Liu,3,4,* and Markus A. Seeliger2,7,*1Department of Physiology and Biophysics, Stony Brook University, Stony Brook, NY 11794-8661, USA2Department of Pharmacological Sciences, Stony Brook University, Stony Brook, NY 11794-8651, USA3Department of Chemistry and Chemical Biology4Howard Hughes Medical Institute
Harvard University, Cambridge, MA 02138, USA5Co-first author6Present address: Department of Chemistry, Princeton University, Princeton, NJ 08544, USA7Lead Contact
*Correspondence: [email protected] (D.R.L.), [email protected] (M.A.S.)
http://dx.doi.org/10.1016/j.chembiol.2016.07.017
SUMMARY
Protein kinases are attractive therapeutic targetsbecause their dysregulation underlies many dis-eases, including cancer. The high conservation ofthe kinase domain and the evolution of drug resis-tance, however, pose major challenges to the devel-opment of specific kinase inhibitors. We recentlydiscovered selective Src kinase inhibitors from aDNA-templated macrocycle library. Here, we revealthe structural basis for how these inhibitors retainactivity against a disease-relevant, drug-resistantkinase mutant, while maintaining Src specificity. Wefind that these macrocycles display a degree ofmodularity: two of their three variable groups interactwith sites on the kinase that confer selectivity, whilethe third group interacts with the universally con-served catalytic lysine and thereby retains the abilityto inhibit the ‘‘gatekeeper’’ kinase mutant. We alsoshow that these macrocycles inhibit migration ofMDA-MB-231 breast tumor cells. Our findings estab-lish intracellular kinase inhibition by peptidic macro-cycles, and inform the development of potent andspecific kinase inhibitors.
INTRODUCTION
Selective inhibition of protein kinases is an effective clinical strat-
egy for the treatment of diseases caused by aberrant kinase
signaling (Cohen and Alessi, 2013; Levitzki, 2013). However,
we lack selective small-molecule inhibitors for many disease-
associated protein kinases, and the limited kinase selectivity of
available inhibitors leads to dose-limiting off-target toxicity (Da-
vis et al., 2011).
The Src family of protein tyrosine kinases (SFKs) consists of
eight non-receptor tyrosine kinases that share high sequence
Cell Chemical Biolo
homology, domain architecture, and regulation (Parsons and
Parsons, 2004). SFKs regulate fundamental cellular processes
such as cell migration, differentiation, growth, and survival (Par-
sons and Parsons, 2004). Src kinase, the prototypical SFK, is
overexpressed or constitutively activated in many solid tumors
types (Summy and Gallick, 2003; Yeatman, 2004), and inhibition
of Src decreases metastasis and tumor growth in both cellular
and animal cancer models. Therefore, Src is considered a phar-
macological target for cancer therapy (Gargalionis et al., 2014;
Krishnan et al., 2012; Nagaraj et al., 2011; Tang et al., 2011;
Tsai et al., 2013; van Oosterwijk et al., 2013; Zhang and Yu,
2012). However, selective pharmacologic inhibition of Src kinase
is challenging because the eight members of the Src kinase fam-
ily are highly conserved, and few small-molecule kinase inhibi-
tors can distinguish between them (Anastassiadis et al., 2011;
Blake et al., 2000; Brandvold et al., 2012, 2015; Georghiou
et al., 2012; Gushwa et al., 2012; Kwarcinski et al., 2012).
Achieving specificity among different Src kinase family members
is crucial because off-target inhibition can create significant clin-
ical problems, such as immunosuppression and impaired T cell
function through inhibition of the hematopoietic SFKs Lck and
Hck (Lowell, 2004; Palacios and Weiss, 2004).
Most small-molecule kinase inhibitors were discovered in
high-throughput screens and their optimization was guided by
Lipinski’s ‘‘ruleof five’’ (RO5) thatdescribes featuresof someorally
bioavailable drugs (Lipinski et al., 2001). More recently, com-
poundssuchasmacrocycles that explore chemical spacebeyond
RO5-compliant compounds have received attention as selective
enzyme inhibitors (Driggers et al., 2008; Heinis, 2014; Villar et al.,
2014). Macrocycles are typically larger and can possess more
rotatable bonds than typical RO5 compounds. This plasticity is
balanced by the conformational restriction throughmacrocycliza-
tionandallowsmacrocycles toadoptconformations thatprecisely
complement a binding site (Villar et al., 2014). In addition, func-
tional groups can be displayed from the macrocycle backbone
and engage multiple interaction sites on the receptor indepen-
dently. Macrocyclic kinase inhibitors therefore have the potential
to be highly specific by exploiting multiple small differences in
the structure and sequence of the conserved kinase domain.
gy 23, 1103–1112, September 22, 2016 ª 2016 Elsevier Ltd. 1103

Figure 1. Chemical Structures of the Sec-
ond-Generation Compounds MC4 and
MC25 Described in This Work
Please cite this article as: Aleem et al., Structural and Biochemical Basis for Intracellular Kinase Inhibition by Src-specific Peptidic Macrocycles, CellChemical Biology (2016), http://dx.doi.org/10.1016/j.chembiol.2016.07.017
Approximately 70 macrocycles are currently used in the clinic.
Most of these are either natural products or natural product de-
rivatives, such as the immunosuppressant cyclosporine A and
the antibiotic vancomycin (Heinis, 2014). The challenges associ-
ated with synthesizing macrocycles and understanding their
pharmacological properties has deterred studies of these com-
pounds, and many questions concerning the molecular basis
of macrocycle-target interactions remain (Driggers et al., 2008;
Heinis, 2014; Rezai et al., 2006; Villar et al., 2014). In particular,
surprisingly little structural information is available for macro-
cycle-protein complexes. A recent comprehensive study listed
only 22 distinct X-ray cocrystal structures of macrocycles bound
to proteins (Villar et al., 2014); in contrast, 3,000 structures of
protein kinase domains have been determined in complex with
acyclic kinase inhibitors.
Recently, we discovered two families of Src-specific kinase by
in vitro selection of a 13,824-membered DNA-templated macro-
cyclic peptide library (Kleiner et al., 2010). The two families of
Src-specific kinase inhibitors are classified as nitrophenylala-
nine- or pyrazine-based macrocycles depending on the first
building block incorporated during synthesis (Figure 1). Mem-
bers of these families inhibit Src kinase with nanomolar potency
and more than 80-fold selectivity over the closely related Hck
and Abl kinase (Kleiner et al., 2010). This selectivity is remark-
able, as no clinically approved kinase inhibitor is selective for
Src over other SFKs or Abl kinase (Davis et al., 2011).
We previously reported the X-ray crystal structures of two pyr-
azine macrocycles, MC1 and MC4, bound to the Src kinase
domain (Georghiou et al., 2012). Our studies revealed that these
macrocycles bind to the Src/CDK-like inactive conformation of
the kinase, and occupy the ATP binding pocket while simulta-
neously disrupting substrate peptide binding. We showed that
three amino acid substitutions between Src and Hck determine
the specificity of macrocycles for Src over Hck. These structures
provide important insights into the binding mode of the pyrazine
macrocycles and how they achieve specificity for Src kinase.
However, the structural basis of the nitrophenylalanine macro-
cycles’ ability to inhibit Src kinase as well as the activating ‘‘gate-
keeper’’ mutant (Thr338Ile) of Src (Azam et al., 2008; Foda et al.,
2015) while maintaining selectivity has remained unknown. Phar-
macological inhibition of the Src gatekeeper mutant is of partic-
1104 Cell Chemical Biology 23, 1103–1112, September 22, 2016
ular interest, since the analogous muta-
tion in other kinases frequently underlies
drug resistance in the clinic, and few ki-
nase inhibitors are active against this mu-
tation. Moreover, inhibitors that are active
against the gatekeeper mutation typically
show reduced kinase selectivity (Kwar-
cinski et al., 2016; Zhao et al., 2014).
Here we present the X-ray cocrystal
structure of a nitrophenylalanine-based
macrocycle with Src kinase. This struc-
ture reveals the molecular basis of Src
kinase inhibition by the nitrophenylalanine macrocycles, and
identifies a novel interaction between the nitrophenyl group
and the catalytically essential and universally conserved Lys
295, thereby demonstrating how these compounds retain po-
tency against Src containing the gatekeeper T338I mutation.
We also demonstrate that themacrocycles retain activity against
disease-related inhibitor-resistance mutations in the activation
loop of the kinase. Finally, we show that the macrocycles inhibit
Src-dependent cell migration in a breast tumor-derived cell line.
Taken together, our findings establish that macrocycles can be
effective and highly selective kinase inhibitors in mammalian
cells and reveal the molecular basis of their activity against
wild-type and mutant forms of Src kinase. Our findings inform
the future development of macrocycles as research tools that
selectively modulate Src-dependent cell signaling without inhib-
iting hematopoietic SFKs.
RESULTS
The Nitrophenylalanine Group of MC25b Forms UniqueInteractions in the ATP Binding Pocket of Src KinaseTo elucidate the structural basis for the differences in potency,
selectivity, and response to kinase mutations observed between
the pyrazine (MC4) and nitrophenylalanine series of macrocycles
(MC25) (Figure 1), we determined the X-ray crystal structure of
MC25b bound to Src (Figures 2A and S1) and compared it
with our previously reported crystal structure of the Src,MC4b
complex (Georghiou et al., 2012). MC25 consists of a 19-atom
macrocycle, an ornithine scaffold and a trans-olefin, while MC4
consists of an 18-atom macrocycle, a diaminobutyric acid scaf-
fold, and cis-olefin (Figure 1). The site of DNA attachment in the
DNA-templated library is a carboxamide in MC4a/MC25a and a
carboxylate in MC4b/MC25b (Figure 1).
Despite their different A building blocks and different macro-
cyclic scaffolds, MC25b and MC4 bind to Src in similar confor-
mations and occupy the same three binding pockets in the active
site of Src (Figure 2C). The A, B, and C building blocks occupy
the adenine binding pocket, a hydrophobic pocket underneath
the b3-aC loop, and an amphipathic pocket near the DFG motif,
respectively (Figures 2A and 2B). The peptidic macrocycle back-
bones form almost identical hydrogen bonding patterns with the
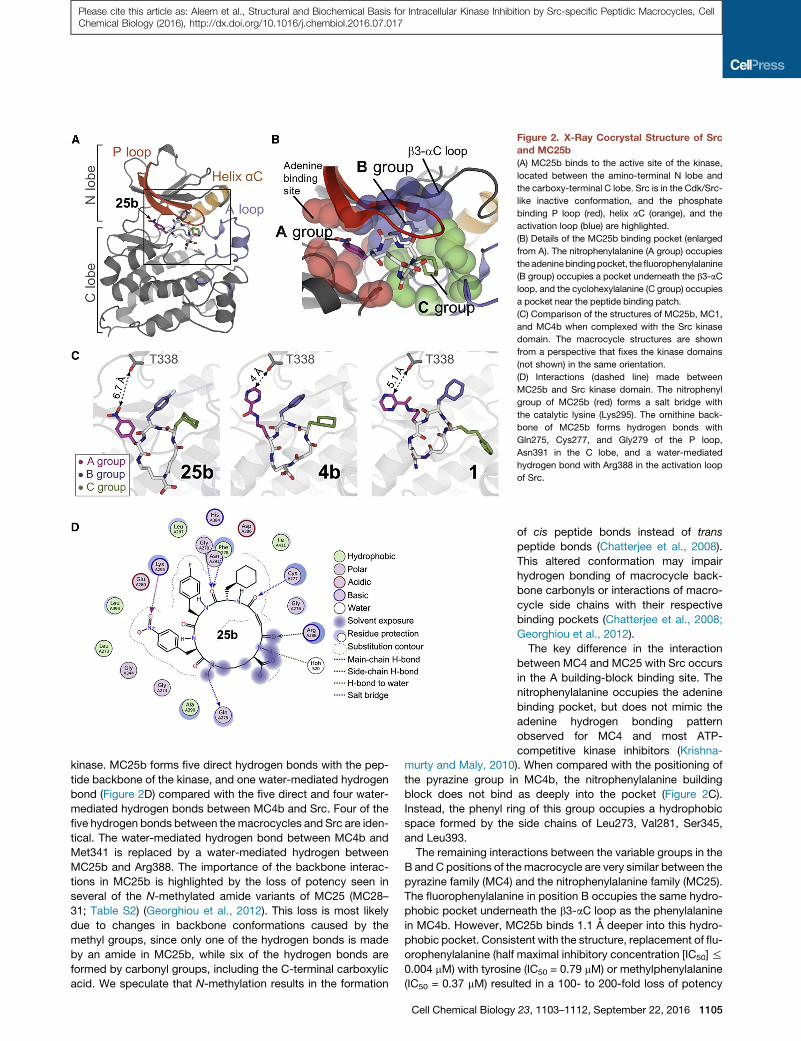
Figure 2. X-Ray Cocrystal Structure of Src
and MC25b
(A) MC25b binds to the active site of the kinase,
located between the amino-terminal N lobe and
the carboxy-terminal C lobe. Src is in the Cdk/Src-
like inactive conformation, and the phosphate
binding P loop (red), helix aC (orange), and the
activation loop (blue) are highlighted.
(B) Details of the MC25b binding pocket (enlarged
from A). The nitrophenylalanine (A group) occupies
the adeninebindingpocket, the fluorophenylalanine
(B group) occupies a pocket underneath the b3-aC
loop, and the cyclohexylalanine (C group) occupies
a pocket near the peptide binding patch.
(C) Comparison of the structures of MC25b, MC1,
and MC4b when complexed with the Src kinase
domain. The macrocycle structures are shown
from a perspective that fixes the kinase domains
(not shown) in the same orientation.
(D) Interactions (dashed line) made between
MC25b and Src kinase domain. The nitrophenyl
group of MC25b (red) forms a salt bridge with
the catalytic lysine (Lys295). The ornithine back-
bone of MC25b forms hydrogen bonds with
Gln275, Cys277, and Gly279 of the P loop,
Asn391 in the C lobe, and a water-mediated
hydrogen bond with Arg388 in the activation loop
of Src.
Please cite this article as: Aleem et al., Structural and Biochemical Basis for Intracellular Kinase Inhibition by Src-specific Peptidic Macrocycles, CellChemical Biology (2016), http://dx.doi.org/10.1016/j.chembiol.2016.07.017
kinase. MC25b forms five direct hydrogen bonds with the pep-
tide backbone of the kinase, and one water-mediated hydrogen
bond (Figure 2D) compared with the five direct and four water-
mediated hydrogen bonds between MC4b and Src. Four of the
five hydrogen bonds between themacrocycles and Src are iden-
tical. The water-mediated hydrogen bond between MC4b and
Met341 is replaced by a water-mediated hydrogen between
MC25b and Arg388. The importance of the backbone interac-
tions in MC25b is highlighted by the loss of potency seen in
several of the N-methylated amide variants of MC25 (MC28–
31; Table S2) (Georghiou et al., 2012). This loss is most likely
due to changes in backbone conformations caused by the
methyl groups, since only one of the hydrogen bonds is made
by an amide in MC25b, while six of the hydrogen bonds are
formed by carbonyl groups, including the C-terminal carboxylic
acid. We speculate that N-methylation results in the formation
Cell Chemical Biology
of cis peptide bonds instead of trans
peptide bonds (Chatterjee et al., 2008).
This altered conformation may impair
hydrogen bonding of macrocycle back-
bone carbonyls or interactions of macro-
cycle side chains with their respective
binding pockets (Chatterjee et al., 2008;
Georghiou et al., 2012).
The key difference in the interaction
between MC4 and MC25 with Src occurs
in the A building-block binding site. The
nitrophenylalanine occupies the adenine
binding pocket, but does not mimic the
adenine hydrogen bonding pattern
observed for MC4 and most ATP-
competitive kinase inhibitors (Krishna-
murty and Maly, 2010). When compared with the positioning of
the pyrazine group in MC4b, the nitrophenylalanine building
block does not bind as deeply into the pocket (Figure 2C).
Instead, the phenyl ring of this group occupies a hydrophobic
space formed by the side chains of Leu273, Val281, Ser345,
and Leu393.
The remaining interactions between the variable groups in the
B and C positions of themacrocycle are very similar between the
pyrazine family (MC4) and the nitrophenylalanine family (MC25).
The fluorophenylalanine in position B occupies the same hydro-
phobic pocket underneath the b3-aC loop as the phenylalanine
in MC4b. However, MC25b binds 1.1 A deeper into this hydro-
phobic pocket. Consistent with the structure, replacement of flu-
orophenylalanine (half maximal inhibitory concentration [IC50]%
0.004 mM) with tyrosine (IC50 = 0.79 mM) or methylphenylalanine
(IC50 = 0.37 mM) resulted in a 100- to 200-fold loss of potency
23, 1103–1112, September 22, 2016 1105

Figure 3. The Effect of Ionic Strength on the
Binding of Nitrophenylalanine and Pyrazine
Macrocycles to Src Kinase
(A) Fluorescence anisotropy was used to measure
the binding of the purified forms of the indicated
Src kinase domains with a fluorescein-labeled
macrocycle containing either a pyrazine (fluores-
cein-MC2) or nitrophenylalanine (fluorescein-
MC9) group at the A position at 0 mM NaCl (low
salt) or 500 mMNaCl (high salt). Experiments were
performed in triplicate. Error bars represent mean
values ± SD.
(B) KD values from (A) are plotted. Error bars
represent mean values ± SD.
Please cite this article as: Aleem et al., Structural and Biochemical Basis for Intracellular Kinase Inhibition by Src-specific Peptidic Macrocycles, CellChemical Biology (2016), http://dx.doi.org/10.1016/j.chembiol.2016.07.017
(Table S2). The cyclohexylalanine at position C occupies an
amphipathic pocket in both structures of MC4 and MC25, which
is generated by the outward rotation of helix aC into the Cdk/Src-
like inactive conformation.
The Nitrophenylalanine Interacts with the UniversallyConserved Catalytic LysineThe crystal structure of MC25b suggests that the catalytic lysine
(Lys295) of Src is a key residue for MC25 binding, contributing
favorable van der Waals interactions and longer-range electro-
static interactions across 4.6 A with the nitrophenylalanine of
MC25 (Figures 2D and S2). These electrostatic interactions be-
tween the negatively charged oxygen of the zwitterionic nitro
group and Lys295 would not be expected if the nitrophenyl
group was replaced with pyrazine, as in MC4. Furthermore, the
importance of this interaction is supported by the observed
loss in potency when the nitrophenylalanine group was
substituted with other functionalities (in MC17–MC23) (Table
S2). Only the cyanophenylalanine containing MC21 retained its
potency for Src, possibly because it is capable of forming an
interaction between a negative partial charge at the cyano nitro-
gen and the positive charge of Lys295.
To probe the putative electrostatic interaction between the ni-
trophenyl group found inMC25b and the catalytic lysine (Lys295)
of Src, we used twomacrocycles that were fluorescein labeled at
the C terminus for direct binding studies: fluorescein-MC2, a
pyrazine macrocycle, and fluorescein-MC9, a nitrophenyl mac-
rocycle (Table S2). We anticipate that attaching the fluorophore
at this position does not interfere with macrocycle binding to
Src, since the C terminus was linked to DNA during the macro-
cycle library selection and our macrocycle Src structures indi-
cate that the C terminus is solvent exposed. We measured
how the ionic strength of the solvent andmutation of the catalytic
Lys295 affected binding affinity using fluorescence anisotropy.
We found that high salt concentrations lowered the affinity of
the nitrophenylalanine compound (fluorescein-MC9) to Src by
1106 Cell Chemical Biology 23, 1103–1112, September 22, 2016
4-fold (Figures 3A and 3B). When the cat-
alytic lysine was mutated to methionine
(K295M), the binding affinity for the
nitrophenylalanine compound decreased
more than 100-fold and became indepen-
dent of ionic strength (Figures 3A and 3B).
In contrast, there were no significant dif-
ferences in the binding affinities of the
pyrazine compound (fluorescein-MC2) toward Src, or the
K295M mutant, at low or high salt concentrations (Figures 3A
and 3B). Taken together, these results suggest that the nitro-
phenyl group of MC25 forms an ionic interaction with the cata-
lytic lysine (Lys295) of Src kinase, whereas the pyrazine-based
MC4 does not. Since Lys295 is essential for kinase activity, resis-
tance mutations at this position that disrupt this ionic interaction
are unlikely.
MC25 and MC4 Have Differential Activity againstDrug-Resistance MutationsTyrosine kinases (such as Abl kinase) commonly present muta-
tions in the clinic that confer drug resistance. The gatekeeper
residue (Abl Thr315, Src Thr338) is the most common clinical im-
atinib-resistance mutation, followed by mutations in the P loop
and the activation loop (Bikker et al., 2009). Replacement of
the gatekeeper residue with a hydrophobic residue (Abl T315I,
Src T338I/M, EGFR T790M) activates the kinase (Azam et al.,
2008; Foda et al., 2015). A single nucleotide exchange in the hu-
man gene for Src kinase results in the T338Mmutation, while the
T338I mutation is present in approximately half of all chicken
Rous sarcoma strains of viral Src kinase. Since Src is currently
not a primary target of clinical kinase inhibitors, no clinical resis-
tance mutations have emerged.
The crystal structure of Src,MC25b presented here demon-
strates why the nitrophenylalanine family of macrocycles is
active against the T338I mutation (inhibition constant Ki =
0.016 mM) while the pyrazine family is not (Table S3) (Georghiou
et al., 2012). The distance between pyrazine and the Thr338 side
chain is only 4 A (Figure 2C). Replacement of threonine with the
larger isoleucine side chain would clash with the pyrazine (Geor-
ghiou et al., 2012). In contrast, the distance between the nitro-
phenyl of MC25b and Thr338 is 6.7 A (Figure 2C), and the
T338I mutation is not predicted to cause steric clashes (Fig-
ure S3). Introduction of the T338M mutation causes resistance
to MC25b (Ki = 5.3 mM) (Table S3), and the mutation is predicted

Figure 4. A Bcr-Abl Activation-Loop Inhibitor-Resistance Mutation
Modeled in Src(R419P) Results in a Loss of Potency for MC25b
(A) Inhibitory potency of macrocycles in biochemical kinase assay. Experi-
ments were performed in triplicate, and data represent mean values ± SD.
(B) Destabilization of Src/Cdk-like inactive conformation results in loss of
macrocycle potency. Src autophosphorylation (pTyr-416) or the L407G mu-
tation cause a loss of potency for both MC4b and MC25b. Experiments were
performed in triplicate, and data represent mean values ± SD.
Please cite this article as: Aleem et al., Structural and Biochemical Basis for Intracellular Kinase Inhibition by Src-specific Peptidic Macrocycles, CellChemical Biology (2016), http://dx.doi.org/10.1016/j.chembiol.2016.07.017
to decrease the minimum distance to 4.3 A (Figure S3). Interest-
ingly, MC25b retains its selectivity for Src kinase (Ki = 0.010 mM)
and shows low activity against Hck (Ki = 1.4 mM), Fyn (Ki =
1.12 mM), and Lck (Ki = 1.0 mM) as well as moderate activity
against Yes kinase (Ki = 0.12 mM) (Table S3).
Since the backbone of MC25 extended further toward the
C-terminal end of the activation loop than MC4 (Figure S4), we
compared the potency of activation-loop mutations on macro-
cycle potency. We chose the most common imatinib-resistance
mutation found in the activation loop of BCR-Abl (H396P) (Corbin
et al., 2003; Young et al., 2006) and generated the analogous Src
mutant (R419P) to assess macrocycle potency (Seeliger et al.,
2007). Based on the crystal structure of the homologous muta-
tion in Abl (H396P), we hypothesized that substituting a proline
at this position altered the conformation of the activation loop
in the Src/Cdk-like inactive conformation, and would potentially
result in the loss of a hydrogen bond for MC25, but not for MC4
(Young et al., 2006). Indeed, MC4 was able to largely retain po-
tency (2.6-fold higher IC50 than wild-type Src) against this muta-
tion, while MC25 inhibited this mutant 26-fold less potently than
wild-type Src (Figure 4A). In the crystal structures of MC4 and
MC25, Arg419 faces away from the active site. A carbonyl group
from the backbone of MC25 forms a hydrogen bond with
Arg388, and the perturbation of the activation loop by R419P
would likely shift Arg388 too far to retain this hydrogen bond.
Collectively, these results show how the modularity in the
macrocycle architecture results in differential sensitivity to muta-
tions: the size and nature of the A group determines the sensi-
tivity to the gatekeeper mutation, while the size and conforma-
tion of the macrocycle backbone determines interaction with
the activation loop of the kinase.
Both Classes of Macrocycles Bind Selectively to theSrc-like Inactive ConformationThe crystal structures of MC4 and MC25 both demonstrate that
the macrocyclic inhibitors bind to the Src/Cdk-like inactive
conformation of Src. In particular, the binding pocket for the B
group underneath the b3-aC loop is enlarged by the outward
rotation of helix aC in the Src/Cdk-like inactive conformation.
To verify that the macrocycles are selective for the inactive
conformation, we tested their potency against the autophos-
phorylated form of Src that is locked in the active conformation.
We found that neither MC4b nor MC25b could inhibit Src when
the kinase was autophosphorylated (IC50 values >20 mM) (Fig-
ure 4B).We also tested the potency ofMC4b andMC25b against
a mutant form of Src (L407G) that is proposed to destabilize the
Src/Cdk-like inactive conformation (Seeliger et al., 2007). The
L407G mutation decreased the potency of MC4 and MC25 by
200–fold and 2,000-fold, respectively (Figure 4B).
These results emphasize that the macrocycle Src inhibitors
have a strong preference for a specific inactive kinase conforma-
tion, and that mutations or posttranslational modifications that
destabilize this conformation reduce the activity of the inhibitor.
MC25 Inhibits Migration of Breast Cancer CellsOur structural data explain how the macrocycles achieve po-
tency, kinase selectivity, and activity against certain mutations
while recognizing the Src/Cdk-like inactive conformation. We
next investigated how the in vitro inhibition data would translate
to the cellular context. We previously showed that high micro-
molar concentrations of MC25a decreased global tyrosine phos-
phorylation in NIH 3T3 Src�/� cells overexpressing a constitu-
tively activated form of Src (Y529F) (Georghiou et al., 2012).
While the modest cellular potency of MC25a against the consti-
tutively active Src Y529F mutant may be a result of limited mem-
brane permeability and cellular uptake, as has been observed for
other macrocycles (Bockus et al., 2015), we also speculated that
the Y529F mutation increases intracellular autophosphorylation
of Src on Tyr416 (chicken c-Src numbering) (Hunter, 1987), lock-
ing the kinase in the active conformation and disfavoring macro-
cycle binding. Indeed, our biochemical data indicate that MC4
and MC25a were more than 1,000-fold less potent against auto-
phosphorylated Src kinase (Figure 4B). We therefore tested
compound potency against endogenous levels of Src in breast
tumor-derived MDA-MB-231 cells. To quantify the effect of in-
hibitor of cellular kinase activity, we treated cells with inhibitor
or vehicle control for 24 hr, washed the cells, and immunopuri-
fied Src from cells. Kinase activity of the Src,MC25 complexes
was quantified in radioactive kinase assays. MC25a decreased
the activity of Src from MDA-MB-231 in a dose-dependent
manner with an IC50 of 0.06 mM (Figure 5A). Dasatinib as a control
at 1 mM inhibited Src kinase activity completely. The levels of im-
munopurified Src were unaffected by inhibitor concentration
(Figure 5B).
MDA-MB-231 cells depend upon Src activity for migration,
which is necessary for metastasis (Sanchez-Bailon et al., 2012,
Cell Chemical Biology 23, 1103–1112, September 22, 2016 1107

Figure 5. MC25a Inhibits Cell Migration in a Metastatic Breast Cancer Cell Line(A) Dose-dependent inhibition of MDA-MB-231 cells by MC25a. MDA-MB-231 cells were treated for 24 hr with MC25a or DMSO. Following cell lysis and
immunoprecipitation with Src antibody, radioactive kinase activity assays were performed, yielding an IC50 of 0.06 mM. Data represent mean values of four
experiments ±SEM.
(B) Western blot of immunoprecipitated Src from MC25a-treated cells as input control for radioactive kinase assays in (A).
(C) Wound-closure assay of MDA-MB-231 cells. Following serum starvation for 12 hr, cells were treated with either 20 mM of MC25a, or DMSO vehicle control.
(D) Following introduction of a sterile wound, wound closure was quantified by measuring the free space unoccupied by cells at the indicated time points.
Data represent mean values ±SEM. *p < 0.05.
Please cite this article as: Aleem et al., Structural and Biochemical Basis for Intracellular Kinase Inhibition by Src-specific Peptidic Macrocycles, CellChemical Biology (2016), http://dx.doi.org/10.1016/j.chembiol.2016.07.017
2015; Tang et al., 2011; Tsai et al., 2013). Treatment of cells with
20 mMMC25 decreased the rate of cell migration and the closure
of an artificial wound. After 17.5 hr of treatment with MC25 (or
45 hr, when cells had been serum starved), the remaining wound
size doubled compared with vehicle control (Figures 5C and 5D).
Proliferation of MDA-MB-231 cells was not affected by MC25 as
observed previously for other Src inhibitors (Tsai et al., 2013).
These results show that the macrocycle Src inhibitors are active
in cells and can be used to inhibit a metastasis-related pheno-
type in a breast tumor-derived cell line.
DISCUSSION
Previously we reported two potent and unusually selective
macrocyclic inhibitors of Src kinase, MC4 and MC25, and eluci-
dated the structural and biochemical basis of Src-specific
inhibition by MC4 (Georghiou et al., 2012). While these two
macrocycles share structural similarities, MC25 demonstrated
several unique features over MC4 including cellular efficacy,
insensitivity to the activating and drug-resistant T338I gate-
keepermutation, and a higher degree of selectivity for Src kinase.
Here we have characterized the structural basis of Src kinase in-
hibition byMC25.Our structural data indicate that this compound
makes unique interactions in the adenine binding pocket that
enable retention of activity against T338I Src without compro-
mising inhibitor specificity. Furthermore, we demonstrate that
MC25 inhibits Src-dependent cell migration in a metastatic
breast tumor-derived cell line. These studies will be useful for
developing the next generation of macrocycle kinase inhibitors.
Comparison of the structures of pyrazine-based MC4 and ni-
trophenylalanine-based MC25 bound to Src kinase allowed us
1108 Cell Chemical Biology 23, 1103–1112, September 22, 2016
to rationalize their Src selectivity and differential activity against
drug-resistant kinase mutations. We hypothesize that the excel-
lent kinase selectivity of these inhibitors is tied to their preference
for the inactive kinase conformation as well as the engagement
ofmultiple binding sites. Kinase inhibitors that target a distinct ki-
nase conformation are often more selective, since the relevant
conformational states may be more accessible to the target ki-
nase or may reveal target-specific structural features exploited
by the inhibitor (Muller et al., 2015; Tong and Seeliger, 2015;
Wang et al., 2014). For example, the highly selective kinase inhib-
itors imatinib and lapatinib bind to their targets in distinct inactive
kinase conformations. However, inhibition of specific kinase
conformations can also represent an Achilles heel for inhibitors,
as destabilization of the targeted conformation can render the in-
hibitor inactive. The macrocycles presented here and the Abl ki-
nase inhibitor, imatinib, target specific inactive conformations
that are less accessible upon phosphorylation of the kinase acti-
vation loop (Schindler et al., 2000). Consequently, phosphoryla-
tion of Abl kinase domain increases the dissociation constant
(KD) for imatinib 20-fold (Davis et al., 2011) and the enzymatic
Ki 100-fold (Seeliger et al., 2007). Despite this limitation, imatinib
is highly successful in the clinic, likely because activation-loop
phosphorylation is in a dynamic equilibrium mediated by phos-
phatases. Other examples of clinically successful kinase inhibi-
tors that lose potency against their activated kinase target
include the vascular endothelial growth factor receptor inhibitor
axitinib (50-fold increase in Ki) (Solowiej et al., 2009) and the
c-Kit inhibitor sunitinib (300-fold increase in IC50) (DiNitto et al.,
2010). The clinical success of imatinib, axitinib, and sunitinib in-
dicates that resistance of activation-loop phosphorylated ki-
nases to inhibitors can be tolerated and should not diminish

Please cite this article as: Aleem et al., Structural and Biochemical Basis for Intracellular Kinase Inhibition by Src-specific Peptidic Macrocycles, CellChemical Biology (2016), http://dx.doi.org/10.1016/j.chembiol.2016.07.017
the potential of the macrocycles from being useful kinase
inhibitors.
MC25b buries almost 70% more solvent-accessible surface
area on the kinase (537 A2) than dasatinib (316 A2). Thus, the
larger size of MC25b compared with dasatinib may explain the
ability to connect multiple distinct binding sites and to spread
the binding energy over multiple and potentially lower-affinity
interactions.
Our macrocycle Src structures show that the main differences
between MC25 and MC4 interactions with Src occur in the
adenine binding site and the kinase activation loop. These two
regions are highly conserved among tyrosine kinases and are
of particular interest in the clinic as they are frequent sites of
drug resistance to small-molecule inhibitors. We find that while
MC25 extends toward the activation loop, the smaller scaffold
of MC4 does not extend into this region and MC4 binding is
therefore insensitive to the R419P mutation, which is analogous
to the clinically relevant H396P mutation in c-Abl that confers
resistance to imatinib in chronic myeloid leukemia patients.
Conversely, while MC4 inserts deep into the adenine binding
pocket and forms an adenine-like hydrogen bond with Thr338,
MC25 does not. Rather, our structural and biochemical data indi-
cate that MC25 resides 6.7 A from Thr338 and that its affinity for
Src derives from additional electrostatic interaction between the
nitrophenyl group and catalytic Lys295, thereby explaining how
MC25 binding is not perturbed by the T338I mutation. This
shallow positioning in the nucleotide binding site and interaction
with Lys295 are unusual, as most small-molecule kinase inhibi-
tors make a characteristic set of hydrophobic and hydrogen
bond interactions that mimic the adenine ring of ATP (Krishna-
murty and Maly, 2010). Exceptions are inhibitors such as the
50-fluorosulfonylbenzoate derivatives of the pan-SFK inhibitor
PP1 that covalently attach to the catalytic lysine (Gushwa
et al., 2012).
Notably, first- and second-generation small-molecule inhibi-
tors of c-Abl that are used in the clinic, such as imatinib and da-
satinib, respectively, form hydrogen bonds with Thr315 (the
analogous gatekeeper residue in c-Abl) and are inactive against
c-Abl containing a T315I mutation, the most common drug-
resistance mutation occurring after imatinib treatment (Soverini
et al., 2006). While c-Abl inhibitors that can inhibit this mutant
protein (e.g., ponatinib) have been described, these com-
pounds typically have compromised target specificity (Liu
et al., 2012). In this regard, MC25may serve as a promising tem-
plate for the development of future Abl/Src kinase inhibitors,
since it is both highly selective and insensitive to mutation of
the gatekeeper residue. Moreover, we predict that interactions
with Lys295 (or the analogous Lys290 in c-Abl), a residue
essential for catalysis, are less likely to be perturbed by resis-
tance mutations. Overall, the structural basis for the inhibition
of wild-type Src kinase and c-Abl-related drug-resistant mu-
tants by these two families of macrocycles provides a strategy
to develop specific and potent macrocyclic kinase inhibitors
that would be insensitive to mutations known to arise in the
clinic. Such an inhibitor would benefit from an aromatic group
with polar functionality in the A position to form long-distance
interactions with Lys295 while retaining distance from Thr338,
and a smaller MC4-based macrocycle backbone that would
be insensitive to mutations in the activation loop. We speculate
that such a compound would retain selectivity due to the B- and
C-position building blocks.
Finally, we show that MC25 inhibits endogenous Src activity in
the MDA-MB-231 breast tumor cell line (Tang et al., 2011; Tsai
et al., 2013) and that Src inhibition in MDA-MB-231 cells corre-
lates with a reduction of cell migration, a disease-relevant
phenotype of this metastatic breast tumor-derived cell line that
is known to depend on Src activity. The observation that MC25
inhibits migration of MDA-MB-231 cells, which depends on Src
activity but does not inhibit their proliferation, which depends
on Abl activity (Srinivasan and Plattner, 2006; Srinivasan et al.,
2008), can be attributed to MC25’s excellent selectivity for Src
over Abl kinase.
Metastasis is a marker of advanced stage disease clinically,
and represents an important target for drug inhibition Notably,
MC25 inhibited wound healing at concentrations not expected
to inhibit the hematopoietic kinases Hck and Lck. At present,
therapeutic strategies to inhibit Src are limited by off-target ef-
fects that cause immunosuppression and inhibitor toxicity (Lev-
itzki, 2013). Therefore, the biological potency and selectivity of
MC25 may provide a tool to help evaluate Src-dependent
signaling in tumor cell lines.
SIGNIFICANCE
Protein kinases are involved in virtually all cellular signaling
pathways, and their dysregulation underlies many diseases.
Kinases are therefore attractive drug targets if specific and
potent inhibition can be achieved. The high level of conser-
vation of structure and sequence within the catalytic kinase
domain complicates the development of specific kinase in-
hibitors. In addition, pharmacologic kinase inhibition is
made challenging by drug-resistant mutations in the target
that arise after treatment. All small-molecule kinase inhibi-
tors currently in clinical use bind to the conserved ATP bind-
ing site in the protein kinase domain. We have previously
identified a series of peptidicmacrocycles that are unusually
specific for Src kinase over other related Src family kinases,
and interfere both with ATP and peptide substrate binding.
Here we characterize the structural and biochemical mech-
anisms of Src kinase inhibition by one such compound,
MC25, which retains activity against a common disease-
relevant inhibitor-resistance mutation at the so-called gate-
keeper residue, T338I. Our study reveals a unique mode of
Src kinase inhibition and provides a mechanistic rational
for MC25’s unusual kinase selectivity as well as activity
against T338I Src. In addition, we show that this compound
is active in cells. Many natural product-derived drugs are
macrocycles, suggesting that the macrocyclic structure
imbues these molecules, which lie outside conventional
drug-like chemical space, with favorable properties such
as high specificity and bioavailability. However, the design
of synthetic macrocycles has been hampered by a paucity
of structural data and lack of macrocycles with cellular ac-
tivity. MC25 could serve as a tool compound informing the
future development of kinase inhibitors active against Src
kinase containing mutations at the gatekeeper residue,
including the T338Mmutation. Furthermore, the compounds
Cell Chemical Biology 23, 1103–1112, September 22, 2016 1109

Please cite this article as: Aleem et al., Structural and Biochemical Basis for Intracellular Kinase Inhibition by Src-specific Peptidic Macrocycles, CellChemical Biology (2016), http://dx.doi.org/10.1016/j.chembiol.2016.07.017
presented here could motivate future systematic studies to
derive empirical rules for bioactive macrocycles.
EXPERIMENTAL PROCEDURES
Constructs and Mutagenesis
Kinase domain constructs of chicken c-Src (residues 251–533) were purified
as previously described (Seeliger et al., 2005, 2007). Mutations were intro-
duced into Src kinase domain (K295M, L407G, and R419P) by site-directed
mutagenesis and were verified by DNA sequencing.
Protein Crystallization
The complex betweenMC25b (500 mM) and c-Src kinase domain (200 mM)was
formed in 20 mM Tris (pH 8.0), 125 mMNaCl, 2.5% DMSO, and 2.5% glycerol.
Src,MC25b crystallized in a hanging-drop format at room temperature when
mixed in a 1:1 ratio with mother liquor consisting of 14% polyethylene glycol
5000 monomethyl ether and 0.3 M NaH2PO4. Crystals were cryoprotected in
mother liquor supplemented with 20% glycerol, cryocooled, and stored in
liquid nitrogen. See Supplemental Experimental Procedures for details.
Data Collection and Processing
X-ray diffraction data were collected on beamline X29 at Brookhaven National
Laboratory at 100 K using a wavelength of 1.075 A. Phases were obtained by
molecular replacement using the kinase domain of c-Src (PDB: 3U4W) (Geor-
ghiou et al., 2012) in Phaser (McCoy et al., 2007). The diffraction data were
strongly anisotropic, and an anisotropic correction was carried out using the
Anisotropy Diffraction Server (Strong et al., 2006). These anisotropically scaled
data were then refined in Phenix (Adams et al., 2002). The model was built in
Coot. See Supplemental Experimental Procedures for further details and
refinement statistics.
Fluorescence Anisotropy
The change in fluorescence anisotropy of fluorescein-labeledMC2 andMC9 at
518 nm upon excitation at 492 nm was monitored with a Jobin Yvon Fluoro-
Max-4 spectrofluorimeter (Horiba). Src kinase domain (residues 251–533)
was titrated to 0.5 mM of the fluorescein-labeled macrocycle, in 100 mM Tris
(pH 8.0), 10mMMgCl2 at 25�C. For the salt titrations, 500mMNaCl was added
to the buffer prior to the addition of kinase. After equilibration, the increase in
anisotropy of the fluorescently labeled ligandwas recorded and fitted against a
quadratic binding equation utilizing the GraphPad Prism software to yield
the KD.
In Vitro Autophosphorylation of Src Kinase Domain
Src autophosphorylation experiments were performed by incubating 10 mM
Src kinase domain with 1 mM ATP in buffer containing 20 mM Tris (pH 8.0)
and 10 mM MgCl2 for 2 hr at 25�C.
Kinase Assays
For Src inhibition assays (Barker et al., 1995), 300 mM of an Src-optimal sub-
strate peptide (AEEEIYGEFAKKK) (Songyang et al., 1995) was combined
with 250 mM ATP. The concentrations of kinase used in these assays were
as follows: 0.0125 mM Src kinase domain, 0.025 mM R419P, 0.025 L407G. Ti-
trations of MC4b and MC25b (ranging from 0 to 83.3 mM) were performed at
30�Cas described previously for imatinib to determine the IC50 and the Ki (See-
liger et al., 2007).
Immunoprecipitation and Radioactive Kinase Assay
MDA-MB-231 cells were grown to 90% confluence and treated with dasatinib,
MC25a, or DMSO vehicle. Cells were harvested 24 hr following drug treatment,
washed once with PBS, and lysed with RIPA lysis buffer. Lysates were cleared
by centrifuging at 15,000 3 g for 10 min and then immunoprecipitated for Src
protein (32G6 antibody [Cell Signaling Technology]) overnight at 4�C. Thebeads were washed four times with ice-cold PBS supplemented with 1 mM
vanadate. The beads were divided into three tubes, two of which were used
in the radioactive kinase assay with a Src substrate peptide and [g-32P]ATP.
The remaining tube was analyzed by SDS-PAGE, transferred onto Hydro-
phobic Polyvinylidene Fluoride (PVDF) membrane (Millipore), and analyzed
1110 Cell Chemical Biology 23, 1103–1112, September 22, 2016
by western blotting for total Src (32G6). See Supplemental Experimental Pro-
cedures for details.
Wound-Healing Assay
MDA-MB-231 cells were grown to confluence in 12-well plates in DMEM/
Ham’s F12 (50:50) supplemented with 10% fetal bovine serum at 37�C in
5% CO2. In some experiments, the cells were then serum starved for 12 hr
as indicated. Multiple wounds were generated in each well by scratching
with a sterile P200 pipette tip. The cells were then washed twice with PBS,
and treated with complete media supplemented with DMSO vehicle control
or 20 mM MC25a. Each condition was repeated in triplicate. Photographs
were taken per well at four different wound locations and at different time
points. The percentage of wound closure was calculated with ImageJ by
measuring the area unoccupied by cells at each time point, and normalizing
to the zero time point.
ACCESSION NUMBERS
Atomic coordinates and structure factors for Src,MC25b have been deposited
in the PDB with accession code PDB: 5BMM.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures,
four figures, and three tables and can be found with this article online at
http://dx.doi.org/10.1016/j.chembiol.2016.07.017.
AUTHOR CONTRIBUTIONS
B.P.C., S.U.A., and W.T.M. performed all the cellular experiments. S.U.A. per-
formed the fluorescence anisotropy binding experiments. G.G., K.E.G., and
S.U.A. determined the structure of the Src,MC25b complex. G.G. performed
the in vitro kinase assays. R.E.K., A.I.C., and D.R.L. developed and synthe-
sized all the macrocyclic inhibitors utilized in this work. M.A.S. contributed
to the design of all the experiments and directed the study. All authors contrib-
uted to the writing of the manuscript.
ACKNOWLEDGMENTS
This work was supported by NIHGM119437 (M.A.S.), NIHCA058530 (W.T.M.),
NIH ES022930 (K.E.G.), NIH GM100021 (M.G.-D.), NIH GM118062 (D.R.L.),
NIH 2T32GM008444 (S.A., K.E.G., and A.L.), and HHMI (D.R.L.). Use of beam-
line X29 at the National Synchrotron Light Source of Brookhaven National Lab-
oratory (Upton, NY) was supported by the U.S. Department of Energy, Office of
Science, Office of Basic Energy Sciences under contract no. DE-AC02-
98CH10886. D.R.L. is a consultant for Ensemble Therapeutics, a company
that uses DNA-templated synthesis in drug discovery and development.
Received: July 22, 2015
Revised: July 4, 2016
Accepted: July 14, 2016
Published: September 1, 2016
REFERENCES
Adams, P.D., Grosse-Kunstleve, R.W., Hung, L.W., Ioerger, T.R., McCoy, A.J.,
Moriarty, N.W., Read, R.J., Sacchettini, J.C., Sauter, N.K., and Terwilliger, T.C.
(2002). PHENIX: building new software for automated crystallographic struc-
ture determination. Acta Crystallogr. D Biol. Crystallogr. 58, 1948–1954.
Anastassiadis, T., Deacon, S.W., Devarajan, K., Ma, H., and Peterson, J.R.
(2011). Comprehensive assay of kinase catalytic activity reveals features of ki-
nase inhibitor selectivity. Nat. Biotechnol. 29, 1039–1045.
Azam, M., Seeliger, M.A., Gray, N.S., Kuriyan, J., and Daley, G.Q. (2008).
Activation of tyrosine kinases by mutation of the gatekeeper threonine. Nat.
Struct. Mol. Biol. 15, 1109–1118.
Barker, S.C., Kassel, D.B., Weigl, D., Huang, X., Luther, M.A., and Knight, W.B.
(1995). Characterization of pp60c-src tyrosine kinase activities using a

Please cite this article as: Aleem et al., Structural and Biochemical Basis for Intracellular Kinase Inhibition by Src-specific Peptidic Macrocycles, CellChemical Biology (2016), http://dx.doi.org/10.1016/j.chembiol.2016.07.017
continuous assay: autoactivation of the enzyme is an intermolecular autophos-
phorylation process. Biochemistry 34, 14843–14851.
Bikker, J.A., Brooijmans, N., Wissner, A., and Mansour, T.S. (2009). Kinase
domain mutations in cancer: implications for small molecule drug design stra-
tegies. J. Med. Chem. 52, 1493–1509.
Blake, R.A., Broome, M.A., Liu, X., Wu, J., Gishizky, M., Sun, L., and
Courtneidge, S.A. (2000). SU6656, a selective Src family kinase inhibitor,
used to probe growth factor signaling. Mol. Cell Biol. 20, 9018–9027.
Bockus, A.T., Lexa, K.W., Pye, C.R., Kalgutkar, A.S., Gardner, J.W., Hund,
K.C., Hewitt, W.M., Schwochert, J.A., Glassey, E., Price, D.A., et al. (2015).
Probing the physicochemical boundaries of cell permeability and oral bioavail-
ability in lipophilic macrocycles inspired by natural products. J. Med. Chem.
58, 4581–4589.
Brandvold, K.R., Steffey, M.E., Fox, C.C., and Soellner, M.B. (2012).
Development of a highly selective c-Src kinase inhibitor. ACS Chem. Biol. 7,
1393–1398.
Brandvold, K.R., Santos, S.M., Breen, M.E., Lachacz, E.J., Steffey, M.E., and
Soellner, M.B. (2015). Exquisitely specific bisubstrate inhibitors of c-Src ki-
nase. ACS Chem. Biol. 10, 1387–1391.
Chatterjee, J., Gilon, C., Hoffman, A., and Kessler, H. (2008). N-methylation of
peptides: a new perspective in medicinal chemistry. Acc. Chem. Res. 41,
1331–1342.
Cohen, P., and Alessi, D.R. (2013). Kinase drug discovery - what’s next in the
field? ACS Chem. Biol. 8, 96–104.
Corbin, A.S., La Rosee, P., Stoffregen, E.P., Druker, B.J., and Deininger, M.W.
(2003). Several Bcr-Abl kinase domainmutants associatedwith imatinibmesy-
late resistance remain sensitive to imatinib. Blood 101, 4611–4614.
Davis, M.I., Hunt, J.P., Herrgard, S., Ciceri, P., Wodicka, L.M., Pallares, G.,
Hocker, M., Treiber, D.K., and Zarrinkar, P.P. (2011). Comprehensive analysis
of kinase inhibitor selectivity. Nat. Biotechnol. 29, 1046–1051.
DiNitto, J.P., Deshmukh, G.D., Zhang, Y., Jacques, S.L., Coli, R., Worrall, J.W.,
Diehl, W., English, J.M., and Wu, J.C. (2010). Function of activation loop tyro-
sine phosphorylation in themechanism of c-Kit auto-activation and its implica-
tion in sunitinib resistance. J. Biochem. 147, 601–609.
Driggers, E.M., Hale, S.P., Lee, J., and Terrett, N.K. (2008). The exploration of
macrocycles for drug discovery—an underexploited structural class. Nat. Rev.
Drug Discov. 7, 608–624.
Foda, Z.H., Shan, Y., Kim, E.T., Shaw, D.E., and Seeliger, M.A. (2015). A
dynamically coupled allosteric network underlies binding cooperativity in Src
kinase. Nat. Commun. 6, 5939.
Gargalionis, A.N., Karamouzis, M.V., and Papavassiliou, A.G. (2014). The mo-
lecular rationale of Src inhibition in colorectal carcinomas. Int. J. Cancer 134,
2019–2029.
Georghiou, G., Kleiner, R.E., Pulkoski-Gross, M., Liu, D.R., and Seeliger, M.A.
(2012). Highly specific, bisubstrate-competitive Src inhibitors from DNA-tem-
plated macrocycles. Nat. Chem. Biol. 8, 366–374.
Gushwa, N.N., Kang, S., Chen, J., and Taunton, J. (2012). Selective targeting
of distinct active site nucleophiles by irreversible SRC-family kinase inhibitors.
J. Am. Chem. Soc. 134, 20214–20217.
Heinis, C. (2014). Drug discovery: tools and rules for macrocycles. Nat. Chem.
Biol. 10, 696–698.
Hunter, T. (1987). A tail of two Src’s: mutatis mutandis. Cell 49, 1–4.
Kleiner, R.E., Dumelin, C.E., Tiu, G.C., Sakurai, K., and Liu, D.R. (2010). In vitro
selection of a DNA-templated small-molecule library reveals a class of macro-
cyclic kinase inhibitors. J. Am. Chem. Soc. 132, 11779–11791.
Krishnamurty, R., and Maly, D.J. (2010). Biochemical mechanisms of resis-
tance to small-molecule protein kinase inhibitors. ACSChem. Biol. 5, 121–138.
Krishnan, H., Miller, W.T., and Goldberg, G.S. (2012). SRC points the way to
biomarkers and chemotherapeutic targets. Genes Cancer 3, 426–435.
Kwarcinski, F.E., Fox, C.C., Steffey, M.E., and Soellner, M.B. (2012).
Irreversible inhibitors of c-Src kinase that target a nonconserved cysteine.
ACS Chem. Biol. 7, 1910–1917.
Kwarcinski, F.E., Brandvold, K.R., Phadke, S., Beleh, O.M., Johnson, T.K.,
Meagher, J.L., Seeliger, M.A., Stuckey, J.A., and Soellner, M.B. (2016).
Conformation-selective analogs of dasatinib reveal insight into kinase inhibitor
binding and selectivity. ACS Chem. Biol. 11, 1296–1304.
Levitzki, A. (2013). Tyrosine kinase inhibitors: views of selectivity, sensitivity,
and clinical performance. Annu. Rev. Pharmacol. Toxicol. 53, 161–185.
Lipinski, C.A., Lombardo, F., Dominy, B.W., and Feeney, P.J. (2001).
Experimental and computational approaches to estimate solubility and perme-
ability in drug discovery and development settings. Adv. Drug Deliv. Rev. 46,
3–26.
Liu, Q., Kirubakaran, S., Hur, W., Niepel, M., Westover, K., Thoreen, C.C.,
Wang, J., Ni, J., Patricelli, M.P., Vogel, K., et al. (2012). Kinome-wide selectivity
profiling of ATP-competitivemammalian target of rapamycin (mTOR) inhibitors
and characterization of their binding kinetics. J. Biol. Chem. 287, 9742–9752.
Lowell, C.A. (2004). Src-family kinases: rheostats of immune cell signaling.
Mol. Immunol. 41, 631–643.
McCoy, A.J., Grosse-Kunstleve, R.W., Adams, P.D., Winn, M.D., Storoni, L.C.,
and Read, R.J. (2007). Phaser crystallographic software. J. Appl. Crystallogr.
40, 658–674.
Muller, S., Chaikuad, A., Gray, N.S., and Knapp, S. (2015). The ins and outs of
selective kinase inhibitor development. Nat. Chem. Biol. 11, 818–821.
Nagaraj, N.S., Washington, M.K., and Merchant, N.B. (2011). Combined
blockade of Src kinase and epidermal growth factor receptor with gemcitabine
overcomes STAT3-mediated resistance of inhibition of pancreatic tumor
growth. Clin. Cancer Res. 17, 483–493.
Palacios, E.H., and Weiss, A. (2004). Function of the Src-family kinases, Lck
and Fyn, in T-cell development and activation. Oncogene 23, 7990–8000.
Parsons, S.J., and Parsons, J.T. (2004). Src family kinases, key regulators of
signal transduction. Oncogene 23, 7906–7909.
Rezai, T., Bock, J.E., Zhou, M.V., Kalyanaraman, C., Lokey, R.S., and
Jacobson, M.P. (2006). Conformational flexibility, internal hydrogen bonding,
and passive membrane permeability: successful in silico prediction of the rela-
tive permeabilities of cyclic peptides. J. Am. Chem. Soc. 128, 14073–14080.
Sanchez-Bailon, M.P., Calcabrini, A., Gomez-Dominguez, D., Morte, B.,
Martin-Forero, E., Gomez-Lopez, G., Molinari, A., Wagner, K.U., and Martin-
Perez, J. (2012). Src kinases catalytic activity regulates proliferation, migration
and invasiveness of MDA-MB-231 breast cancer cells. Cell Signal. 24, 1276–
1286.
Sanchez-Bailon, M.P., Calcabrini, A., Mayoral-Varo, V., Molinari, A., Wagner,
K.U., Losada, J.P., Ciordia, S., Albar, J.P., and Martin-Perez, J. (2015).
Cyr61 as mediator of Src signaling in triple negative breast cancer cells.
Oncotarget 6, 13520–13538.
Schindler, T., Bornmann, W., Pellicena, P., Miller, W.T., Clarkson, B., and
Kuriyan, J. (2000). Structural mechanism for STI-571 inhibition of abelson tyro-
sine kinase. Science 289, 1938–1942.
Seeliger, M.A., Young, M., Henderson, M.N., Pellicena, P., King, D.S., Falick,
A.M., and Kuriyan, J. (2005). High yield bacterial expression of active c-Abl
and c-Src tyrosine kinases. Protein Sci. 14, 3135–3139.
Seeliger, M.A., Nagar, B., Frank, F., Cao, X., Henderson, M.N., and Kuriyan, J.
(2007). c-Src binds to the cancer drug imatinib with an inactive Abl/c-Kit
conformation and a distributed thermodynamic penalty. Structure 15,
299–311.
Solowiej, J., Bergqvist, S., McTigue, M.A., Marrone, T., Quenzer, T., Cobbs,
M., Ryan, K., Kania, R.S., Diehl, W., and Murray, B.W. (2009). Characterizing
the effects of the juxtamembrane domain on vascular endothelial growth factor
receptor-2 enzymatic activity, autophosphorylation, and inhibition by axitinib.
Biochemistry 48, 7019–7031.
Songyang, Z., Carraway, K.L., Eck, M.J., Harrison, S.C., Feldman, R.A.,
Mohammadi, M., Schlessinger, J., Hubbard, S.R., Smith, D.P., Eng, C., et al.
(1995). Catalytic specificity of protein-tyrosine kinases is critical for selective
signalling. Nature 373, 536–539.
Soverini, S., Colarossi, S., Gnani, A., Rosti, G., Castagnetti, F., Poerio, A.,
Iacobucci, I., Amabile, M., Abruzzese, E., Orlandi, E., et al. (2006). Contribution
of ABL kinase domain mutations to imatinib resistance in different subsets of
Cell Chemical Biology 23, 1103–1112, September 22, 2016 1111

Please cite this article as: Aleem et al., Structural and Biochemical Basis for Intracellular Kinase Inhibition by Src-specific Peptidic Macrocycles, CellChemical Biology (2016), http://dx.doi.org/10.1016/j.chembiol.2016.07.017
Philadelphia-positive patients: by the GIMEMA Working Party on Chronic
Myeloid Leukemia. Clin. Cancer Res. 12, 7374–7379.
Srinivasan, D., and Plattner, R. (2006). Activation of Abl tyrosine kinases pro-
motes invasion of aggressive breast cancer cells. Cancer Res. 66, 5648–5655.
Srinivasan, D., Sims, J.T., and Plattner, R. (2008). Aggressive breast cancer
cells are dependent on activated Abl kinases for proliferation, anchorage-inde-
pendent growth and survival. Oncogene 27, 1095–1105.
Strong, M., Sawaya, M.R., Wang, S., Phillips, M., Cascio, D., and Eisenberg, D.
(2006). Toward the structural genomics of complexes: crystal structure of a
PE/PPE protein complex from Mycobacterium tuberculosis. Proc. Natl.
Acad. Sci. USA 103, 8060–8065.
Summy, J.M., and Gallick, G.E. (2003). Src family kinases in tumor progression
and metastasis. Cancer Metastasis Rev. 22, 337–358.
Tang, Z.N., Zhang, F., Tang, P., Qi, X.W., and Jiang, J. (2011). RANKL-induced
migration of MDA-MB-231 human breast cancer cells via Src and MAPK acti-
vation. Oncol. Rep. 26, 1243–1250.
Tong, M., and Seeliger, M.A. (2015). Targeting conformational plasticity of pro-
tein kinases. ACS Chem. Biol. 10, 190–200.
Tsai, P.C., Chu, C.L., Chiu, C.C., Chang, L.S., and Lin, S.R. (2013). Inhibition of
Src activation with cardiotoxin III blocks migration and invasion of MDA-MB-
231 cells. Toxicon 74, 56–67.
1112 Cell Chemical Biology 23, 1103–1112, September 22, 2016
van Oosterwijk, J.G., van Ruler, M.A., Briaire-de Bruijn, I.H., Herpers, B.,
Gelderblom, H., van deWater, B., and Bovee, J.V. (2013). Src kinases in chon-
drosarcoma chemoresistance and migration: dasatinib sensitises to doxoru-
bicin in TP53 mutant cells. Br. J. Cancer 109, 1214–1222.
Villar, E.A., Beglov, D., Chennamadhavuni, S., Porco, J.A., Jr., Kozakov, D.,
Vajda, S., and Whitty, A. (2014). How proteins bind macrocycles. Nat. Chem.
Biol. 10, 723–731.
Wang, Q., Zorn, J.A., and Kuriyan, J. (2014). A structural atlas of kinases in-
hibited by clinically approved drugs. Methods Enzymol. 548, 23–67.
Yeatman, T.J. (2004). A renaissance for SRC. Nat. Rev. Cancer 4, 470–480.
Young, M.A., Shah, N.P., Chao, L.H., Seeliger, M., Milanov, Z.V., Biggs, W.H.,
3rd, Treiber, D.K., Patel, H.K., Zarrinkar, P.P., Lockhart, D.J., et al. (2006).
Structure of the kinase domain of an imatinib-resistant Abl mutant in complex
with the Aurora kinase inhibitor VX-680. Cancer Res. 66, 1007–1014.
Zhang, S., and Yu, D. (2012). Targeting Src family kinases in anti-cancer ther-
apies: turning promise into triumph. Trends Pharmacol. Sci. 33, 122–128.
Zhao, Z., Wu, H., Wang, L., Liu, Y., Knapp, S., Liu, Q., and Gray, N.S. (2014).
Exploration of type II binding mode: a privileged approach for kinase inhibitor
focused drug discovery? ACS Chem. Biol. 9, 1230–1241.