stress, drugs and neuroscienceuu.diva-portal.org/smash/get/diva2:1190921/fulltext01.pdfendogenous...
TRANSCRIPT

ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2018
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Pharmacy 253
Stress, Drugs and Neuroscience
Neurobiological Effects of Social Stressors and DrugExposure in Young and Adolescent Rats
LINNEA GRANHOLM
ISSN 1651-6192ISBN 978-91-513-0282-9urn:nbn:se:uu:diva-346151

Dissertation presented at Uppsala University to be publicly examined in A1:107a,Biomedicinskt centrum, Husargatan 3, Uppsala, Wednesday, 9 May 2018 at 13:00 for thedegree of Doctor of Philosophy (Faculty of Pharmacy). The examination will be conductedin English. Faculty examiner: Associate Professor Annika Thorsell (Linköpings University,Department of Clinical and Experimental Medicine).
AbstractGranholm, L. 2018. Stress, Drugs and Neuroscience. Neurobiological Effects of SocialStressors and Drug Exposure in Young and Adolescent Rats. Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty of Pharmacy 253. 79 pp. Uppsala: ActaUniversitatis Upsaliensis. ISBN 978-91-513-0282-9.
Experiences early in life or during adolescence modulate neuronal networks in the immaturebrain and consequently lay the foundation for future susceptibility or resilience towardspsychiatric disorders. The objective in this thesis is to understand, in part, how the surroundingenvironment shapes the brain of a young individual. Three types of negative life events werestudied, in an animal model, for their effects on the brain reward system (i.e., endogenousopioids and dopamine) and voluntary drug intake. These were: disruption of maternalcare, disruption of interaction with peers, and exposure to drugs. Stress, in the form ofmaternal separation, altered expression of opioid genes in the dorsal striatum and amygdala,and the response to subsequent alcohol intake on these genes was dependent on early lifeconditions. Basal levels of endogenous opioids were also dependent on how the animals werehoused in early adolescence. Short single housing (30 minutes) caused an acute stress responseas evidenced by increased serum corticosterone and nociceptin/orphanin FQ in brain areasassociated with stress. A prolonged single housing resulted in a marked decrease of Met-Enk-Arg6-Phe7 (i.e., a marker of enkephalins) in several brain areas. The endogenous opioids werealso affected by repeated exposure of ethanol during adolescence; ethanol intoxication increasedthe accumbal levels of Met-Enk-Arg6-Phe7 and decreased those of β-endorphin. Residual effectsof the adolescent ethanol exposure were found in Met-Enk-Arg6-Phe7 levels in the amygdala,ventral tegmental area, and substantia nigra. Furthermore, rats exposed to ethanol as adolescentshad alterations in the dopamine dynamics in the dorsal striatum. Both endogenous opioids anddopamine are essential in mediating rewarding properties. Alterations of these systems, causedby environmental disturbances and alcohol exposure, presented herein could explain, in part,the increased susceptibility for alcohol- and substance use disorders later in life.
Keywords: Adolescence, alcohol exposure, amphetamine, beta-endorphin, dopamine,dynorphin, early life, endogenous opioids, enkephalin, gene expression, high-speedchronoamperometry, nociceptin/orphanin FQ, radioimmunoassay, self-administration, socialstress, voluntary alcohol intake
Linnea Granholm, Department of Pharmaceutical Biosciences, Box 591, Uppsala University,SE-75124 Uppsala, Sweden.
© Linnea Granholm 2018
ISSN 1651-6192ISBN 978-91-513-0282-9urn:nbn:se:uu:diva-346151 (http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-346151)

Hörde fåglarna sjunga om vår


List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals.
I. Granholm, L., Todkar, A., Bergman, S., Nilsson, K., Comasco E., Nylander I. (2017) The expression of opioid genes in non-classical reward areas depends on early life conditions and ethanol intake. Brain Research, 1668:36–45
II. Granholm, L*., Roman, E*., Nylander, I. (2015) Single housing during adolescence causes time-, area-, and peptide-specific altera-tions in endogenous opioids of rat brain. British Journal of Pharma-cology, 172:606-614
III. Granholm, L., Segerström L., Nylander I. (2018) Episodic ethanol exposure in adolescent rats – Effects on endogenous opioid peptides. In manuscript.
IV. Granholm, L*., Rowley, S*., Ellgren, M., Segerström, L., Nylander, I. (2015) Impact of adolescent ethanol exposure and adult amphetamine self-administration on evoked striatal dopamine re-lease in male rats. Psychopharmacology, 232(24):4421-4431
*Authors contributed equally
Reprints were made with permission from the respective publishers.


Contents
Introduction ................................................................................................... 11Early life ................................................................................................... 11Adolescence .............................................................................................. 12Alcohol ..................................................................................................... 12
Consumption behavior ......................................................................... 12Adolescent drinking ............................................................................. 14
Dopamine .................................................................................................. 14Endogenous opioids .................................................................................. 15Social behavior and endogenous opioids .................................................. 17
Early life ............................................................................................... 17Adolescence ......................................................................................... 17
Central circuitries in the addiction cycle .................................................. 18Ethanol ...................................................................................................... 19
Effects on dopamine in dorsal and ventral striatum ............................. 20Effects on endogenous opioids ............................................................ 21
Amphetamine ............................................................................................ 22
Methodology ................................................................................................. 24Maternal separation – simulation of early life stress ........................... 24Experimental housing conditions – simulation of social environments .............................................................................................................. 25Alcohol exposure – simulation of adolescent binge-like drinking ...... 26Voluntary alcohol drinking – evaluation of consumption behavior .... 27Operant self-administration – evaluation of drug-taking behavior ...... 28In vivo high-speed chronoamperometry – analysis of dopamine ......... 29Real-time polymerase chain reaction – analysis of gene expression ... 31Radioimmunoassay – quantification of peptides ................................. 33
Aims .............................................................................................................. 35
Methods ......................................................................................................... 36Animals ..................................................................................................... 36Maternal separation (Paper I) ................................................................... 37Adolescent housing conditions (Paper II) ................................................ 38Adolescent ethanol exposure (Paper III-IV) ............................................. 38Adult voluntary ethanol intake (Paper I) .................................................. 38Operant self-administration (Paper IV) .................................................... 38

Operant boxes apparatus ...................................................................... 38Sucrose training ................................................................................... 39Intravenous amphetamine self-administration ..................................... 39
High speed chronoamperometry (Paper IV) ............................................. 40Quantitative polymerase chain reaction (Paper I) .................................... 41Tissue stabilization (Paper III) ................................................................. 41Radioimmunoassay (Paper II-III) ............................................................. 42Statistical analysis ..................................................................................... 42
Results and discussion ................................................................................... 44Early life stress, alcohol intake and opioid gene expression .................... 44
Amygdala ............................................................................................. 44Dorsal striatum ..................................................................................... 45
Adolescent housing conditions and opioid peptides ................................. 47Nociceptin/Orphanin FQ ...................................................................... 47Methionine-Enkephalin-Arginine6-Phenylalanine7 ............................. 49Dynorphin B ......................................................................................... 50
Adolescent alcohol exposure .................................................................... 50Effects of intoxication on opioid peptides ........................................... 51Residual effects on opioid peptides ..................................................... 52Residual effects on in vivo dopamine in the dorsal striatum ............... 53Subsequent amphetamine self-administration ..................................... 54Subsequent amphetamine self-administration and in vivo dopamine .. 56
Conclusions ................................................................................................... 57
Populärvetenskaplig sammanfattning ........................................................... 58
Acknowledgment .......................................................................................... 60
References ..................................................................................................... 62

Abbreviations
AFR Animal facility rearing ANOVA Analysis of variance cDNA Complementary DNA CRF Corticotrophin-releasing factor DAT Dopamine transporter DNA Deoxyribonucleic acid dPCR Digital polymerase chain reaction FR Fixed ratio GABA γ-aminobutyric acid HPA Hypothalamic-pituitary-adrenal MAO Monoaminooxidase MEAP Methionine-enkephalin-arginine6-phenylalanine7
mRNA Messenger ribonucleic acid MS15 Maternal separation 15 minutes MS360 Maternal separation 360 minutes N/OFQ Nociceptin/Orphanin FQ NOP Non-opioid receptor Oprd1 Opioid receptor delta type 1 Oprk1 Opioid receptor kappa type 1 Oprm1 Opioid receptor mu type 1 PCR Polymerase chain reaction PENK Proenkephalin PDYN Prodynorphin POMC Proopiomelanocortin PR Progressive ratio qPCR Quantitative real-time polymerase chain reaction RNA Ribonucleic acid VMAT2 Vesicular monoamine transporter-2 VTA Ventral tegmental area


11
Introduction
Experiences in early life and adolescence modulate neuronal networks in the immature brain and lay the foundation for future susceptibility or resilience to psychiatric disorders. The objective in this thesis was to understand, in part, how the surrounding environment shapes the developing brain. Three types of negative life events were studied, in an animal model, for their ef-fects on the brain reward system (i.e., endogenous opioids and dopamine) and voluntary drug intake. These were; disruption of maternal care, disrup-tion of interaction with peers, and exposure to drugs.
In the last decades, this area of research has gained more attention. In 2010, the European IMAGEN consortium launched their plan of recruiting high school participants, to follow them throughout adolescence. The partic-ipants undergo a battery of tests ranging from psychological to neuroimaging and genomics to develop prevention strategies and improved therapies for mental health disorders. (Schumann et al., 2010). To answer similar ques-tions in a U.S population, the Adolescent Brain Cognitive Development study was created a couple of years later (Volkow et al., 2017).
Even if these large-scale clinical studies will provide a lot of knowledge, preclinical research is crucial when studying neurobiological consequences of adverse events in young individuals. Animal experimental models provide the opportunity for creating environments where the exposure of the stimuli can be controlled, and enable detailed brain analysis that cannot be made in humans for ethical reasons.
Early life The brain undergoes extensive maturation after birth. Both gray and white matter matures with synaptic formation, axonal growth, neuronal differentia-tion, neurotransmitter modulation and myelination (Dehaene-Lambertz and Spelke, 2015). Genes and environment affects brain development; hence, it is crucial for a child to grow up in an environment with limited stressors and external stimuli of an adverse nature.
From birth and throughout childhood, the most crucial relationship a child has is to its primary caregivers, usually the mother and/or father. A safe and stable environment is important to promote a secure attachment between caregiver and child, which in turn, is important for developmental outcomes.

12
The caregivers' role is to act as a buffer from surrounding stressors. Mal-treatment in childhood, for example, parental neglect or abuse, alters trajec-tories of brain development (Teicher et al., 2016) and increases the risk of psychiatric disorders in adulthood (Carr et al., 2013). Alterations in neurobi-ology and behavior have also been found in preclinical models of early life stress, such as maternal separation (Lyons et al., 2010; Nylander and Roman, 2013).
Adolescence Adolescence is the period of transition from childhood to adulthood. Accord-ing to the World Health Organization, adolescence starts at age 10 and ends at 19 (WHO, 2014). This period also represents a time of extensive reorgani-zation and maturation of brain circuits involved in emotions, motivation, reward and cognition (Spear, 2013). These reflect the development of age-characteristic behaviors such as increased risk taking, impulsivity, and nov-elty seeking (Crews et al., 2007). These behaviors are not exclusive to hu-mans as they can be found in many species (Panksepp, 1981). Furthermore, the social roles and relationships undergo significant changes in adolescent humans (Heinrich and Gullone, 2006) as well as in rats (Spear, 2000). Al-most all mammalians develop an age-specific adolescent play behavior, which follows an inverted U-shaped pattern, i.e., the begins in early adoles-cence and declines with the approach of adulthood (Spinka et al., 2001). In rats, this behavior progressively increases between postnatal day 18-28 and peaks around 32-40, followed by a gradual decline (Panksepp, 1981). It is suggested that this type of social behavior is a part of the social and cogni-tive development as well as a preparation for forthcoming life events (Spinka et al., 2001). Preclinical research has shown that this type of behavior has reinforcing properties, and furthermore, it involves actions of many different neurotransmitters (e.g., opioids, dopamine, noradrenaline) (Trezza et al., 2014; 2010). Evidence from both human and experimental animal research shows that deprivation of social contact during adolescence can lead to se-vere consequences for normal neurobiological and behavioral maturation (Fone and Porkess, 2008; Heinrich and Gullone, 2006).
Alcohol Consumption behavior Alcohol drinking has a long history in human cultures. The word alcohol originates from the Arabic word “alkuhl” which means “something subtle”. The effects of alcohol consumption can range from the subtle to the more

13
obvious, depending on the amount ingested. Alcohol (i.e., ethanol; alcohol and ethanol are used interchangeably in this thesis) is often consumed for its euphoric and sedative effects on the central nervous system. Intoxication also affects, for example, motor- and cognitive functions.
For the majority of alcohol consumers, the goal of drinking is to experi-ence the euphoric and positive reinforcing, and rewarding effects of intoxica-tion. Many individuals can handle repeated and intermittent drinking that sometimes leads to binge levels, i.e., 0.08 g/dl in about two hours according to National institute on Alcohol Abuse and Alcoholism (NIAAA). However, for others, binge drinking (i.e., five or more alcoholic drinks in males, or four or more alcoholic drinks in females, within a two hours (Spear, 2018)) can escalate into chronic use and addiction in which the goal of drinking shifts to finding relief from the negative effects of withdrawal (e.g., dyspho-ria and stress). The desire for becoming intoxicated increases, which leads to preoccupation with, and anticipation of, obtaining the drug. Finally, the per-son loses control of their drinking habit. This pattern is called the addiction cycle (Figure 1) and is further described in “The neuronal circuitries in the addiction cycle”. In this thesis, the effect of ethanol is not evaluated in ani-mals with addiction. Instead, it comprises studies of how the brain is affected in the early stages of alcohol use.
Figure 1. The circle represents the three stages in the addiction cycle—binge/intoxication, withdrawal/negative effects, and preoccupation/anticipation. Inside the circle is a schematic representation of a rat brain and the regions analyzed in the present thesis. The color-coding of the brain areas represents which stages in the addiction cycle that are most influenced by actions in this region; green, binge/intoxication; purple, withdrawal/negative effects; yellow, preoccupa-tion/anticipation; light gray, not defined.
Dorsal striatumNucleus accumbens
Ventral tegmental area
Amygdala
Hippocampus
Medial prefrontal cortex
Hypothalamus
Frontal cortex
Substantia nigra
Cingulate cortex
Pituitary
Binge/Intoxication
Pre
occu
patio
n/a
nticip
ation
Withdrawal/Negative effects

14
Adolescent drinking Impulsivity and novelty seeking are predictors of drug use (Belin and Deroche-Gamonet, 2012; Jentsch et al., 2014). Not surprisingly, the debut of drug use usually occurs during adolescence, with alcohol being one of the most widely used drugs during this period. The consumption pattern is main-ly episodic binge drinking. Since the beginning of this century, there is a positive trend with decreasing alcohol consumption among Swedish teenag-ers, and the use is now down to historically low levels (CAN, 2017). How-ever, this annual report on adolescents drug habits in Sweden also investi-gated how adolescents experience their parents’ alcohol consumption. Ado-lescents growing up in a families with alcohol abuse use alcohol themselves more frequently and in greater quantities (CAN, 2017).
Both clinical and preclinical research shows that exposure to alcohol dur-ing adolescence affects neurobehavioral functions and brain maturation as well as intake of drugs (Guerri and Pascual, 2010; Hanson et al., 2011; Silveri et al., 2016; Spear, 2014; 2018; Squeglia et al., 2015). There is an association between the age of first alcohol use and an increased risk of sub-sequent alcohol and substance use disorders (DeWit et al., 2000; York et al., 2004). Spear (2015) argues that the increased liability with early drug use could be a result of three factors: (i) an environmental context in which drugs are frequently used; (ii) an inherited vulnerability for drugs of abuse, and (iii) a reshaping of the brain by drugs towards a susceptible state. Papers III and IV in this thesis focus on the third factor.
Dopamine In December 2000 the Swedish scientist Arvid Carlsson received the Nobel Prize for his achievements in the field of neurophysiology (Nobelprize.org) . He proved that dopamine is a neurotransmitter and not just a passive player in the synthesis of catecholamines (Carlsson et al., 1957).
A part of the synthesis of dopamine is outlined in Figure 3. The rate-limiting step is the conversion of tyrosine to dihydroxyphenylalanine (DO-PA). Dopamine is packed into synaptic vesicles by vesicular monoamine transporter-2 (VMAT2) before it is released in the synaptic cleft (Howell and Kimmel, 2008). Released dopamine acts on the dopamine receptors and thereafter it can be transported back into the presynapse by dopamine trans-porters (DAT) were it can be repacked into vesicles or metabolized by mon-oaminooxidase (MAO). Dopamine can also be metabolized outside the syn-apse by MAO or catechol-O-methyltransferase (i.e., COMT) (Meiser et al., 2013).
Dopamine receptors were discovered in 1978. They were divided into two classes, D1 or D2 receptors, on the basis of their ability to stimulate adenylyl

15
cyclase (Garau et al., 1978; Spano et al., 1978). This classification is still valid today, but with gene cloning, three additional dopamine receptor sub-types (D3, D4 and D5) have been discovered (Grandy et al., 1991; Sokoloff et al., 1990; Sunahara et al., 1991; Van Tol et al., 1991). The D5 is consid-ered a D1-like receptor because of its similar transmembrane structure as the D1 receptor and as it shares the ability to stimulate adenylyl cyclase (Missale et al., 1998). The D3 and D4 receptors are classified as D2-like receptors because of their homology with the transmembrane domains of the D2 re-ceptor and as they do not stimulate adenylyl cyclase (Missale et al., 1998).
The Swedish researchers, Annika Dahlström and Kjell Fuxe, started map-ping the dopaminergic pathways in the central nervous system in the begin-ning of the 1960s (Dahlstrom and Fuxe, 1964) and their work laid the foun-dation for what we today know about the three major midbrain dopaminergic pathways: the nigrostriatal, mesolimbic and mesocortical (Björklund and Dunnett, 2007).
The dopamine receptors are activated in a variety of functions such as lo-comotor control, attention, cognition and regulation of food intake (Beaulieu and Gainetdinov, 2011). The focus in this thesis is on dopamine action in reward and reinforcement processes within the dorsal striatum.
Endogenous opioids The first references to use of opium are from the third century B.C. in the writings of Theophrastus (van Ree et al., 1999). The use of opioids – for both medical and recreational purposes – is well-documented throughout human history (van Ree et al., 1999), but the first proof of endogenous opi-oids came in the 1970s when it was shown that the brain contained specific binding sites for opiates (Pert and Snyder, 1973; Simon et al., 1973; Terenius, 1973). The hypothesis that there were multiple types of opioid receptors, i.e., μ, δ and κ, came from Martin and his co-workers (Martin et al., 1976), and the discovery of the opioid receptor ligand, enkephalin, (Hughes et al., 1975) confirmed their hypothesis. This was followed by the identification of two other endogenous opioid receptor ligands; endorphin and dynorphin (Cox et al., 1976; Goldstein et al., 1981). A fourth member of the opioid receptor family was discovered in the 1990s. This receptor was first called orphan receptor-like 1 or LC132 receptor (Bunzow et al., 1994; Mollereau et al., 1994), but the name was later changed to non-opioid recep-tor (NOP) when its endogenous ligand, nociceptin orphanin/FQ (N/OFQ) was discovered (Meunier et al., 1995; Reinscheid et al., 1995).
The opioid receptors belong to the G-protein coupled receptor family and exert their actions through inhibition of cyclic adenosine monophosphate (i.e., cAMP) via adenylyl cyclase and consequently decrease the neuronal activity by stimulating outward K+ and inhibiting voltage-gated Ca2+ chan-

16
nels. μ-, δ- and κ-receptors are classical opioid receptors whereas the NOP receptor is a non-classical opioid receptor.
The endogenous ligands of the classical opioid receptors derive form three larger precursor proteins, proopiomelanocortin (POMC), proenkepha-lin (PENK) and prodynorphin (PDYN) that are cleaved by trypsin-like en-zymes to produce multiple bioactive peptides, see Figure 2 for details (Yaksh and Wallace, 2011). These ligands share an N-terminal amino acid sequence, Tyrosine-Glycine-Glycine-Phenylalanine-Methionine/Leucine, followed by various specific extensions of the C-terminus (Yaksh and Wallace, 2011). In contrast, N/OFQ does not share this signature sequence and it lacks affinity to the μ-, δ- and κ-receptors. As for the classical opioids, β-endorphin has strongest binding properties to μ- and δ-receptors, enkepha-lins have the highest affinity to δ-receptors but exert binding properties to μ-receptors as well, and dynorphins primarily target the κ-receptor (Yaksh and Wallace, 2011).
Figure 2. A simplified illustration of the endogenous opioid prohormones—proopiomelanocortin, proenkephalin, prodynorphin and proorphanin. Peptides in bold are those analyzed within this thesis. Abbreviations: ACTH, adrenocortico-tropic hormone; Arg, arginine; CLIP, corticotropin-like intermediate lobe peptide; Gly, glycine; Leu, leucine; Met, methionine; MSH, melanocyte-stimulating hor-mone; N/OFQ, nociceptin/orphanin FQ; Phe, phenylalanine. Adapted from (Yaksh and Wallace, 2011) and (Allen et al., 2001).
The overview of Le Merrer et al. (2009) shows the distribution patterns of receptors and precursor proteins for endorphins, dynorphins and enkephalins and reveals how extensively widespread the endogenous opioid system is in the central nervous system. This system is involved in a variety of physio-logical functions, which is easy to understand, since receptors, precursors
Proopiomelanocortin
Proenkephalin
Prodynorphin
Proorphanin
γ-MSH ACTH β-lipotropin
α-MSH CLIP γ-lipotropin β-endorphin
β-MSH
↓ ↓ ↓ ↓
↓
Met-enkephalin-Arg6Phe7
Met-enkephalin-Arg6Gly7Leu8
α-neoendorphin
β-neoendorphin
↓dynorphin 32
dynorphin A1-17 dynorphin B1-13
↓ ↓
dynorphin A1-8
↓
}}
Nocistatin N/OFQ
Met-enkephalin Leu-enkephalin

17
and ligands are found in brain circuitries regulating, for example, reward, pain, and mood (Le Merrer et al., 2009; Lutz and Kieffer, 2013; Mansour et al., 1987; Yaksh and Wallace, 2011). The distribution of N/OFQ and NOP in the central nervous system includes brain areas (e.g., cortical regions, hippo-campus, amygdala, hypothalamus, locus coeruleus, nucleus accumbens, cau-date putamen) associated with mood regulation (including anxiety and stress), food intake, pain and locomotor activity (Mollereau and Mouledous, 2000; Witkin et al., 2014) .
Social behavior and endogenous opioids Early life As mentioned in “Early life”, the first bond an individual forms is to its pri-mary caregiver and the endogenous opioids have important roles in the neu-robiological underpinnings for parental bonding.
In humans and non-human primates, a variant of the gene encoding the μ-receptor influences attachment behavior and the relationship between parent and child (Barr et al., 2008; Copeland et al., 2011; Higham et al., 2011; Troisi et al., 2012). Furthermore, blocking μ-receptors in the early postpar-tum period reduces both caregiving and protective behavior in rhesus ma-caques (Martel et al., 1993; 1995). Stimulating μ-receptors with morphine increases foraging and hunting behavior in female rats at the cost of reduced maternal behavior (Bridges and Grimm, 1982; Felicio et al., 1991; Sukikara et al., 2007). Across species, administrations of μ-receptor agonists reduce infants’ distress calls when separated from their mothers, and opioid antago-nists induce or exacerbated distress behavior (Carden and Hofer, 1990a; 1990b; Herman and Panksepp, 1978; Kalin et al., 1988; Panksepp et al., 1980a). Furthermore, mice pups lacking the μ-receptor emit fewer distress calls when separated from their mother (Moles et al., 2004) In rodent models of parental neglect and loss, it has been shown that that maternal separation induces long-lasting alterations in endogenous opioids (Nylander and Roman, 2012) and alters the response to both naltrexone (Daoura and Nylander, 2011) and morphine (Kalinichev et al., 2001) in adulthood.
Adolescence Social grooming in non-human primates is fundamental for establishing long-term relationships with social support and protection (Dunbar, 2010). During this behavior, β-endorphins are released and grooming can be ma-nipulated with μ-opioid antagonists and agonists (Keverne et al., 1989; Martel et al., 1995; Meller et al., 1980; Schino and Troisi, 1992). Neuroim-aging data from humans shows that non-sexual, tactile contact stimulates the

18
μ-opioid system, implicating that endogenous opioids are also important for our social bonding (Nummenmaa et al., 2016).
Social play behavior in rats involves a lot of physical contact such as wrestling, during which rodents emits positive ultrasonic vocalizations asso-ciated with joy (Vanderschuren et al., 2016). Endogenous opioids are re-leased during play (Panksepp and Bishop, 1981; Vanderschuren et al., 1995d) and thus contribute to the rewarding effects of the behavior (Calcagnetti and Schechter, 1992; Douglas et al., 2004; Yates et al., 2013). Activation of μ-receptors in the nucleus accumbens is especially important for mediating this behavior (Manduca et al., 2016; Trezza et al., 2011). In both rats and non-human primates, pharmacological manipulations with μ-opioid agonists and antagonists increase and decrease play behavior, respec-tively (Beatty and Costello, 1982; Guard et al., 2002; Niesink and Vanree, 1989; Panksepp et al., 1980b; 1985; Trezza and Vanderschuren, 2008; Vanderschuren et al., 1995a; 1995b; 1995c). Studies have shown that social deprivation decreases opioid receptor binding (Schenk et al., 1982) and alter the response to pharmacological treatments affecting the opioid system (Palm and Nylander, 2014a; Smith et al., 2003; 2005; Wongwitdecha and Marsden, 1996).
Central circuitries in the addiction cycle A brief and simplified description of the neurobiology in the addiction cycle is given below, with the aim of putting the brain areas of interest in Papers I-IV into a theoretical framework. The stages of the addiction cycle are cov-ered comprehensively in reviews by Koob and Volkow (2016; 2010)
Different brain areas and neuronal circuitries account for the main effects in the three stages of the addiction cycle. Importantly, the three stages—binge/intoxication, withdrawal/negative effects, and preoccupation/ anticipation—are all interconnected.
In the first stage, when a drug is consumed for its rewarding effects, the mesolimbic dopamine system is activated. The pleasurable and rewarding effect of drug use is the consequence of a sharp increase of accumbal dopa-mine (Di Chiara and Imperato, 1988). Specifically, the shell, and not the core of accumbens, mediates the rewarding effects (Di Chiara et al., 2004). Stimulants, such as amphetamine and cocaine, can act directly on dopamine release whereas other drugs, alcohol for example, indirectly activate dopa-minergic projections by actions on other transmitter networks. These might be, for example, endogenous opioids that act on the opioid receptors of in-hibitory γ-aminobutyric acid (GABA) neurons in the ventral tegmental area (VTA) and that thereby disinhibit dopamine neurons (Trigo et al., 2010). Repeated binges induce alterations in dopamine dynamics in the dorsal stria-tum, an area involved in habit formation and thereby a shift towards compul-

19
sive drug use (Everitt and Robbins, 2016). In addition to these mesolimbic structures, the central nucleus of amygdala also mediates the rewarding ef-fects (Heyser et al., 1999; Hyytia and Koob, 1995; Moller et al., 1997; Robinson et al., 2014). Most drugs activate the hypothalamic-pituitary-adrenal (HPA) axis, which facilitates actions in the mesolimbic reward pathway (Koob, 2008).
In the next stage of the addiction cycle, the repeated drug-binges have de-sensitized the reward system (due to tolerance in, for example, dopamine, serotonin and endorphins). Both drug- and non-drug rewards become attenu-ated (Koob and Volkow, 2016). During withdrawal, the brains stress sys-tem/antireward system, i.e., the HPA axis and the extended amygdala (the central amygdala, bed nucleus stria terminalis, and nucleus accumbens shell) is dysregulated and sensitized (Koob, 2009). The main neurotransmitters during this stage are corticotrophin releasing factor (CRF), dynorphin, and noradrenaline. Their actions, along with the weakened reward system, result in dysphoria, stress, and unease (Koob, 2008). This will motivates the indi-vidual to continue to use the drug to relieve the negative effects of with-drawal, i.e., negative reinforcement (Koob, 2008).
In the last stage, alterations in prefrontal cortical regions are of im-portance. These structures control executive functions such as decision-making and regulation of action, emotions, and impulses (Abernathy et al., 2010); therefore they are highly involved in the decision to take a drug or not. The prefrontal cortex has excitatory (i.e., glutamatergic) neurons that project directly into many areas in the reward networks such as, the VTA, dorsal and ventral striatum and amygdala, and thereby modulates the action of these regions (Koob and Volkow, 2016). During preoccupation and antic-ipation, the glutamatergic system within the prefrontal cortex becomes hy-persensitized. Along with the dysregulation of dopamine and the increase in the brain stress system, this leads to poor self-control (Belin et al., 2009). Consequently, individuals who reach this stage of the addiction cycle have problems resisting impulses and alcohol cravings. After a period of absti-nence, the person becomes preoccupied with using the drug again.
Ethanol Ethanol is a small molecule that is both hydrophilic and lipophilic. Due to its chemical properties, ethanol is widely distributed in the body when ingested and affects almost all organs. Ethanol has no specific target protein. Instead, it acts on many of the reward-related brain areas and associated neurotrans-mitters.
As for almost all other drugs of abuse, ethanol increases dopamine release in the mesolimbic pathway (Boileau et al., 2003; Di Chiara and Imperato, 1985; Weiss et al., 1993; 1996). Since ethanol has such a complex mecha-

20
nism of action, it is not fully understood how it increases extracellular do-pamine. One of the proposed mechanisms is that it activates endogenous opioids that bind to μ- and δ-receptors on inhibitory GABA interneurons in the VTA and thereby decreases the inhibiting effects of GABA, which con-sequently facilitates release of dopamine. Furthermore, ethanol can also act directly upon the nucleus accumbens (Trigo et al., 2010).
Ligand-gated ion channels are also involved in the dopamine-releasing actions of ethanol. Ethanol-induced increase of acetylcholine and glycine activates nicotinic acetylcholine receptors in the anterior VTA and glycine receptors in the nucleus accumbens, leading to increased mesolimbic dopa-mine activity (Söderpalm and Ericson, 2013). However, ethanol exerts re-warding effects independent of accumbal dopamine, as evidenced by ani-mals continuing to self-administer ethanol after lesions in dopaminergic neurons in the nucleus accumbens (Fahlke et al., 1994; Lyness and Smith, 1992; Rassnick et al., 1993; Shoemaker et al., 2002).
In this thesis, the focus is on ethanol effects on striatal dopamine and en-dogenous opioid peptides. More detailed information about the mechanism of action of ethanol is covered in reviews by Engel and Jerlhag (2014), Söderpalm and Ericson (2013), Tabakoff and Hoffman (2013).
Effects on dopamine in dorsal and ventral striatum The first indications of a connection between catecholamines and ethanol were in the 1970s when an inhibitor of the catecholamine synthesis sup-pressed ethanol-induced locomotion in rats (Engel et al., 1974), and euphoria and social interactions in humans (Ahlenius et al., 1973). Later, microdialy-sis studies showed that injections of ethanol (Di Chiara and Imperato, 1985) as well as voluntary intake (Weiss et al., 1993), increased accumbal dopa-mine. Within the nucleus accumbens, the shell is responsible for the primary rewarding effects of ethanol, whereas the core is implicated in cue-induced ethanol seeking (Ding et al., 2015; Engleman et al., 2009). In humans, neu-roimaging shows that alcohol increases dopamine in the ventral striatum (Aalto et al., 2015; Boileau et al., 2003) and that the amount released corre-lates to self-reported feelings of intoxication (Ramchandani et al., 2011; Urban et al., 2010; Yoder et al., 2007). Furthermore, alcohol downregulates striatal dopamine D2 receptors, and alcohol craving in alcoholics is associat-ed with a low availability of D2 receptors in the ventral striatum (Heinz et al., 2004; Hietala et al., 1994; Volkow et al., 1996).
The early studies regarding acute ethanol exposure and dopaminergic ac-tivity in the dorsal striatum were inconsistent. Di Chiara and Imperato (1985) showed that moderate to high doses increase dopamine but a low dose has no significant effect. Blanchard et al. (1993) presented conflicting results with increased dopamine at low doses. The inconsistency in results could be due to interregional differences within the dorsal striatum, as the

21
ethanol-induced increase in extracellular dopamine occurs in the dorsomedi-al, and not the dorsolateral part (Vena et al., 2016). Dopaminergic activity in the dorsolateral striatum seems to be more involved in the shift to habitual alcohol seeking (Corbit et al., 2012) and influence the motivation to obtain alcohol when it requires high levels of effort (Spoelder et al., 2017). Thus, the dorsolateral part is involved when the consumption behavior becomes increasingly compulsive. In human studies, the ventral striatum in social drinkers is activated when presented with an alcohol cue, whereas the dorsal striatum becomes active in heavy drinkers (Vollstädt-Klein et al., 2010). Furthermore, alcohol-dependent patients have an imbalance between goal-directed and habitual control with a decreased activity in the ventromedial prefrontal cortex and anterior putamen (i.e., regions implicated in goal-directed learning), and increased activity in the posterior putamen (i.e., a region involved in habit learning) (Sjoerds et al., 2013).
Alcohol consumption is indeed modulated by pharmacological agents af-fecting dopamine transmission (Gilpin and Koob, 2008). However, the ad-verse effects of dopamine antagonists (e.g., anhedonia and extrapyramidal actions) have made clinical use difficult. Furthermore, dopamine agonists do not induce relief of the dysphoria caused by the low dopamine activity after excessive drinking (Engel and Jerlhag, 2014; Kosten et al., 2002). There is attempt to use more dynamic dopamine stabilizers, such as aripripazole and (-)-OSU6162, in the treatment of alcohol use disorders (Ingman et al., 2006; Steensland et al., 2012).
Effects on endogenous opioids On the Swedish market, two of the four drugs available for alcohol use dis-order targets opioid receptors. The first of these, naltrexone, is an unspecific antagonist with the highest affinity to μ-receptors and slightly higher affinity to κ- over δ-receptors (Nutt, 2014). The other drug, nalmefene, modulates the opioid system, with antagonistic properties to μ- and δ-receptors and partial agonism to κ-receptors (Bart et al., 2005). Nalmefene exhibits greater affinity to κ- over δ-receptors compared to naltrexone (Emmerson et al., 1994; Michel et al., 1985).
Opioid receptors are located in all brain areas of importance for reward mechanisms (Le Merrer et al., 2009). As mentioned in “Ethanol”, opioids modulate reward and reinforcement. The μ- and δ-receptors account for the positive effects of the ethanol intake and the κ-receptors act counterbalanc-ing (Nutt, 2014). The agonists for μ- and δ-receptors mediate dopamine re-lease in nucleus accumbens, induce place preference and are self-administered (Devine et al., 1993; Devine and Wise, 1994; Spanagel et al., 1990). The κ-receptor agonists decrease accumbal dopamine release, pro-duce aversive effects in preference tests, and are not self-administered

22
(Mucha and Herz, 1985; Mulder et al., 1984; Spanagel et al., 1990; Werling et al., 1988).
The literature describing ethanol-induced effects on opioids is not conclu-sive. However, in rodents, ethanol releases β-endorphin from the hypothala-mus, pituitary, nucleus accumbens and VTA, and increases enkephalins in the nucleus accumbens (de Waele and Gianoulakis, 1993; Gianoulakis, 1990; Jarjour et al., 2009; Lam and Gianoulakis, 2011; Olive et al., 2001). In humans, ethanol intake elevates β-endorphins in the nucleus accumbens and orbitofrontal cortex of both healthy controls and heavy drinkers (Mitchell et al., 2012). Dynorphin is also increased in the nucleus accumbens and central amygdala after acute ethanol exposure (Lam et al., 2008; Marinelli et al., 2006). In a non-dependent state, the κ-system suppresses ethanol reinforce-ment. However, after chronic ethanol use, the dynorphin/κ-system becomes upregulated and during withdrawal, it is involved in the dysphoria that leads to the negatively reinforcing effects of ethanol (Koob, 2008; Sirohi et al., 2012).
N/OFQ counteracts the CRF-induced stress response in the brain (Ciccocioppo et al., 2014; Cruz et al., 2012) and it has been hypothesized that NOP agonists could potentially attenuate the negative effects during withdrawal (Witkin et al., 2014). Preclinical studies have supported the hy-pothesis, with agonists able to reduce intake, prevent somatic effects of withdrawal, and block cue- and stress-induced alcohol reinstatement (Aziz et al., 2016; Ciccocioppo et al., 2004; 1999; Economidou et al., 2011; Martin-Fardon et al., 2000; 2010). However, a recent neuroimaging study found no differences in NOP receptor binding for alcohol-dependent individuals and healthy controls (Narendran et al., 2017).
Amphetamine Amphetamine exerts multiple actions on the dopamine synapse that leading to increased levels of extracellular dopamine (Figure 3). Amphetamine binds to DAT in the cell membrane and competes with dopamine binding (Sulzer, 2011). Furthermore, amphetamine can be transported into the presynapse where it disrupts vesicular storage of dopamine through blockade of VMAT and/or collapse of the pH gradient in the vesicular membrane, leading to vesicular leakage (Sulzer, 2011). Together, these events lead to reversed DAT transport and increased levels of dopamine in the synaptic cleft. Other mechanisms of action have also been proposed such as inhibition of MAO and increased dopamine synthesis (Sulzer, 2011).

23
Figure 3. Mechanism of action of amphetamine. Amphetamine increases the synap-tic concentration of dopamine by; reversing DAT; disrupting the storage of dopa-mine in the synaptic vesicles; and increasing the synthesis of new dopamine (Sulzer, 2011). Abbreviations; D, dopamine receptor; DAT, dopamine transporter; DOPA, dihydroxyphenylalanine; DOPAC, dihydroxyphenylacetic acid; VMAT, vesicular monoamine transporter. Modified with permission from Stina Lundberg, Dept. of Pharmaceutical Biosciences, Uppsala University.
DOPA decarboxylase
Tyrosine hydroxylase
DOPA
Tyrosine
MAOA
D2
Gαi DAT
VMAT
DAT DAT
DOPAC
-+
-+
+
Aldehyde dehydrogenase+
DD D
Dopamine Amphetamine

24
Methodology
Maternal separation – simulation of early life stress The environment of a rat pup is almost exclusively determined by the inter-action with its mother, as the pup and its littermates, live in the nest until weaning. The most common strategy to simulate an adverse environment, and induce early life stress, is to disturb the maternal care. This is usually done by separating the mother from the pup (Figure 4A). The separation can be litter-wise or individual, varying in duration from 180-360 minutes; long-er separations up to 24 hours are usually called maternal deprivation (Knop et al., 2017). The frequency and the timing of the separation can be modulat-ed. The separation can be either daily or intermittent and the timing can be during the first two weeks or until weaning (Molet et al., 2014). Another important aspect for this experimental model is the choice of control group. Litters reared under conventional animal facility standards or left non-handled (i.e., undisturbed) can control for the repeated handling and separa-tions, but it is not possible to distinguish between handling and separation effects (Nylander and Roman, 2013). To only investigate the effect of the separation, one could use groups exposed to either brief handling (i.e., 1-5 minutes of separation) or short maternal separation (i.e., 15 minutes) (Nylander and Roman, 2013). Furthermore, short separations between dam and pups provide a more naturalistic environment; in the wild, the dam leaves the nest regularly for shorter periods (Grota and Ader, 1969). Mater-nal separation models have been criticized for their lack of reproducible results, but the outcome of the experiment depends on all the above-mentioned factors, i.e., type, duration and frequency of separation as well as timing and the choice of control group. Therefore, maternal separation is not a single model of early life stress, but instead several models of separation that fall under the same umbrella.
Another way to induce early life stress, without separation, is to limit the nesting and bedding material in the cage; this stresses the dam which results in fragmented and unpredictable maternal care (Molet et al., 2014). Natural variance in the quality of maternal care is a factor difficult to control for, but important since it per se affects the offspring (Curley and Champagne, 2016). However, this factor is independent of early life stress model and can only be controlled for by scoring the dams’ maternal behavior.

25
Figure 4. Manipulations of housing conditions (Papers I-II). A) Early life stress was assessed in Paper I by 360 minutes of maternal separation. 15 minutes of maternal separation was used to control for handling effects, and animal-facility reared (i.e., left undisturbed) rats were used as a conventional control. B) Effects of housing conditions during adolescence were assessed in either group-housed or single-housed rats in Paper II.
Experimental housing conditions – simulation of social environments The laboratory rat (Rattus norvegicus) is a social species, that in the wild, lives in large colonies with defined hierarchies; if laboratory rats are placed in a semi-naturalistic environment they readily revert to their wildlife behav-ior (Berdoy, 2003). The knowledge of how sensitive rats are to environmen-tal conditions has been used in biomedical research to develop models that alter the social environment (i.e., housing conditions) (Figure 4B). By simp-ly manipulating the number of animals in the same cage, it is possible to induce stress by crowding or isolation (Miczek et al., 2008). The hierarchical system can also be manipulated to induce social stressors, such as social instability, defeat, and subordination (Miczek et al., 2008). Enrichment of the housing conditions can also be used to study the effects of a secure and stimulating environment (Simpson and Kelly, 2011). Even when the aim is not to study the consequences of the environmental setting, it is still im-
A
BAnimal facility rearing
15 minutes ofmaternal separation
360 minutes ofmaternal separation
Group housing
Single housing

26
portant to control the housing conditions throughout the entire experiment to avoid confounders. Uncontrolled for, inconsistencies in the animals’ social conditions can potentially affect the outcome of an experiment, making it difficult to reproduce results between different laboratories.
Alcohol exposure – simulation of adolescent binge-like drinking Several aspects need to be considered when exposing adolescent rats to eth-anol, depending on research question. To investigate drinking phenotypes in a voluntary intake model with high face validity (i.e., to mimic aspects of human intake of alcohol), then operant self-administration or home-cage drinking (non-operant self-administration) are the preferred methods. Etha-nol-naïve rats do not readily self-administer ethanol with the operant tech-nique and therefore extensive training or pre-exposure is necessary before the actual experiments can begin (June and Gilpin, 2010). Adolescence is a fairly short period in a rat’s life and operant self-administration is a time-consuming method; therefore, home-cage drinking is more frequently used because rats readily consume ethanol via this route of administration (Sanchis-Segura and Spanagel, 2006). However, to assess the individual intake of ethanol, this model usually involves single housing which induces stress and long-lasting neurobiological consequences in adolescent rats (Siviy and Panksepp, 2011).
Models that involve forced exposure have weaker face validity but facili-tate the study of neurobiological outcomes of ethanol exposure in a con-trolled way. Both intragastric administration (Figure 5A) and intraperitoneal injections of ethanol activate the HPA-axis (Ogilvie et al., 1997) and are therefore possibly stressful events. However, Turner et al. (2012) have shown that a daily gavage dose not negatively affect animals welfare and Hoffman (2014) showed that gavage had less impact than intraperitoneal injections on corticosterone levels, as a measure of stress. Another method to minimize stress-related handling is the use of intragastric cannulas (Ogilvie et al., 1997). However, as mentioned above, adolescence in rats only lasts a few weeks and the recovery after such an operation has a large impact on both the time and length of the ethanol exposure. Another model that can be used is ethanol exposure through vapor chambers, which requires neither single housing nor handling, but inhalation does not mimic human ingestion of alcohol.

27
Voluntary alcohol drinking – evaluation of consumption behavior In this section, only home-cage drinking (Figure 5B) will be described; op-erant self-administration (Figure 5C) will be explained in “Operant self ad-ministration – evaluation of drug taking behavior”.
Figure 5. Drug exposure paradigms (Papers I-IV). A) Orogastric exposure i.e., ga-vage (Papers III-IV), B) Voluntary ethanol drinking in a two-bottle, free-choice paradigm (Paper I), C) Intravenous self-administration in operant boxes (Paper IV).
Home-cage drinking is a convenient way of evaluating consumption be-havior as it can be easily implemented by adding bottles with ethanol into the cage (Richter and Campbell, 1940). It has both the face and construct validity of human alcohol consumption and pharmacological treatments can prevent excessive drinking, indicating a predictive validity (Sanchis-Segura and Spanagel, 2006). However, to induce addiction in outbred rats by volun-tary drinking is challenging and therefore, home cage drinking is most suita-ble for assessing acquisition and habitual drinking in these rats.
A B
Operantself-administration
Voluntary drinkingOrogastric
exposure
C

28
In most alcohol intake models, the animal has a free choice of drinking ethanol with water and intake of food ad libitum so that the alcohol is con-sumed for its pharmacological effects and not its caloric value or thirst (Bell et al., 2017). Sweetened solutions are sometimes added to increase the amount of liquid ingested, but that makes it difficult to distinguish between the preference for sweet taste and the psychopharmacologic actions of the ethanol (Sanchis-Segura and Spanagel, 2006). In low concentrations (< 4% v/v), ethanol by itself can have a sweet taste (Sanchis-Segura and Spanagel, 2006), and hence it is important to use pharmacological relevant concentra-tions, i.e., >10%. Rodents are nocturnal animals with their highest level of eating, drinking and general activity in the dark phase of their circadian cy-cle. Thus, ethanol consumption increases when administered in the dark (Crabbe et al., 2011; Rhodes et al., 2005).
The access to alcohol can be continuous or intermittent during the drink-ing period; restricting the availability to every other day increases alcohol intake and preference (Wise, 1973). In the intermittent paradigms, the access can be either unlimited or limited. Unlimited access makes it difficult to determent if the intake has led to relevant blood alcohol concentrations or if the animal sporadically sips from the ethanol bottle and hence, not reaching significant pharmacological effects (Becker, 2013). Limited access is also better when analyzing tissues, since it decreases the inter-individual differ-ences. This leads to better relation between the results (e.g., tissue levels of peptides) and the amount of ethanol intake.
Operant self-administration – evaluation of drug-taking behavior Operant self-administration (Figure 5C) is frequently used for studying vol-untary intake of psychostimulants (e.g., cocaine and amphetamine) since rats easily learn the operant technique with this type of substances. The disad-vantage with operant self-administration is that it is not as convenient as home-cage voluntary intake. It requires both training and more sophisticated equipment, i.e., operant self-administration boxes equipped for intravenous drug delivery. Self-administration boxes are also known as Skinner-boxes, named after their inventor, B.F. Skinner. He developed these boxes for oper-ant conditioning where the animal learns that a certain behavior (e.g., a lever press) is associated with a stimuli (e.g., a light signal) and positive rein-forcement (e.g., drug infusion) (Skinner, 1938). Operant self-administration facilitates observations of many behaviors, for example consumption behav-ior, reward efficacy, extinction and cue-induced reinforcement. In this thesis, consumption behavior and reward efficacy were studied.
Consumption behavior is mainly studied through fixed ratio (FR) sched-ules. In this type of trial the drug is delivered after a preset number of re-sponses (e.g., a number of lever presses). To examine oral ethanol intake with operant boxes, low FR schedules (i.e., one press on the active lever,

29
FR1) are required to maintain self-administration. In contrast, higher FR schedules (e.g., FR3) are used for drug delivery systems with an immediate drug effect (e.g., intravenous amphetamine infusions) (Sanchis-Segura and Spanagel, 2006).
Progressive ratio (PR) trials are used to investigate reward efficacy. This method was first developed by William Hodos (1961) when he tested the reinforcing properties of sweetened milk in rats. In PR trials the response requirements increase systematically for each reward and therefore give an estimation of the maximum response for maintenance of self-administration (Richardson and Roberts, 1996). The point in the escalating series of re-sponse requirements at which the animal stop responding is called the break-point. This value reflects the maximum effort the animal is willing to make in order to receive a reward (e.g., a drug infusion) (Richardson and Roberts, 1996). Different reinforcers produce different breakpoint values. For exam-ple, psychostimulants have high breakpoints values (Richardson and Roberts, 1996) whereas alcohol has a lower one (June and Gilpin, 2010).
In vivo high-speed chronoamperometry – analysis of dopamine A variety of methods (e.g., electrochemical recordings, microdialysis, and neuroimaging) can be used to measure in vivo dopamine. In this thesis, do-pamine was measured with high-speed chronoamperometry (Figure 6), an electrochemical technique. A small electrode is placed into the brain area of interest and a potential applied to the electrode causing any molecules in contact with the electrode to be oxidized (Michael and Wightman, 1999). Not all molecules in the extracellular fluid can be oxidized, therefore the application of this method depends on the analyte of interest (Gerhardt and Burmeister, 2000). However, with the proper potential, dopamine can be oxidized to form an o-quionone that can be reduced back to dopamine. This reaction results in measurable elicitations of Faradaic currents that are pro-portional to the analyte concentration (Gerhardt and Burmeister, 2000). The ratio between the oxidation and reduction currents creates an electrochemical fingerprint of the analyte (Gerhardt and Burmeister, 2000). The way of ap-plying the potential over the electrode differs depending on the electrochem-ical technique. The simplest way is to use a constant and fixed potential (re-ferred to as amperometry in the literature), which leads to continuous meas-urements of currents at the electrode and a good temporal resolution (Michael and Wightman, 1999). However, the chemical selectivity is poor. In high-speed chronoamperometry the voltage is applied to the electrode in a square-wave manner that gives the opportunity for oxidation and reduction of the molecule, and hence the possibility to identify the analyte (Michael and Wightman, 1999). Nevertheless, it should be mentioned that dopamine and other biogenic amines (e.g. noradrenaline, serotonin) could be difficult to distinguish from each other with this method. However, the chronoam-

30
perometric recordings in this thesis are performed in the dorsal striatum where dopamine is the mostly abundant and dominant biogenic amine. Mi-crodialysis, a third technique, allows collection and analysis of samples con-taining a number of different molecules in both basal state and after a phar-macological or physiological challenge (Gerhardt and Burmeister, 2000). The main advantage with high-speed chronoamperometry over microdialysis is its spatial and temporal resolution (Gerhardt and Burmeister, 2000).
Figure 6. Overview of high-speed chronoamperometry (Paper IV). A) The micro- and reference electrodes are inserted into the dorsal part of striatum through a stereo-taxic operation in anesthetized rats. B) K+ is released (I) into the brain area of inter-ested and causes a neuronal depolarization (II), and release of dopamine (III). A square-wave potential is applied to an electrode, which then causes dopamine to oxidize (IV). The current generated is proportional to the concentration of released dopamine. C) A schematic example of a trace of oxidation current from a rat receiv-ing amphetamine. D) A close-up of a peak showing how amplitude and T80 values are calculated. Figures B) and C) is modified with permission from Stina Lundberg, Dept. of Pharmaceutical Biosciences, Uppsala University (B) and Sara Palm (2014) (C).
The disadvantage is that measurements of resting or basal levels of dopa-mine are difficult to perform due to low extracellular levels (Gerhardt and Burmeister, 2000). To overcome this limitation, measurements of neuronal
A
Time (s)
[μM
]
Amplitude
T80
Time (s)
BNH2OH
OH
Dopamine
K+
K+ K+K+
K+
K+
I.
II.
III.
IV.
+2H+
2e-2e-
O
NH2O
C D
Amphetamine
References
[μM
]

31
release can be done through chemical (e.g., potassium-induced) or electrical stimulation of the neurons (Michael and Wightman, 1999) in pharmacologi-cally untreated or treated subjects. Commercial systems for high-speed chronoamperometry can easily measure and analyze the neuronal release of for example dopamine. This is displayed as a peak where the peak amplitude relates to the amount of released dopamine, and the time for the signal to decay 80% (T80), is a measurement of dopamine re-uptake (Zahniser et al., 1998) (Figure 3D).
Real-time polymerase chain reaction – analysis of gene expression The major breakthrough in gene research occurred in the 1950´s when Rosalind Franklin produced an X-ray diffraction picture of deoxyribonucleic acid, DNA (Franklin and Gosling, 1953). With the help of her data, Watson and Crick solved the double helix structure of DNA (Watson and Crick, 1953). In 1962, Watson and Crick shared the Nobel Prize with Maurice Wil-kins for this discovery (Nobelprize.org). Another groundbreaking, and Nobel Prize awarded (Nobelprize.org), discovery was that of polymerase chain reaction (PCR) (Mullis and Faloona, 1987). PCR is a process whereby spe-cific segments of DNA (e.g., genes) can be amplified into multiple copies. Briefly, the process involves several steps of increasing and decreasing tem-perature in a solution containing; DNA, nucleotides, gene-specific primers, and thermostable DNA polymerase. The reaction begins by separating the double helix of the DNA to allow the attachment of primers designed for specific parts of DNA. Thereafter, the DNA polymerase begins to synthesize new DNA strands with the primers as starting points. After synthesis, the original DNA-molecule has now been duplicated and in forthcoming cycles, the designated DNA is exponentially amplified.
In 1993, the first real-time PCR was made (Higuchi et al., 1993). Before real-time PCR instruments were available, the quantification of DNA had to be done after the PCR; with real-time PCR, the amplification of DNA occurs simultaneously and combines both amplification and detection (Wong and Medrano, 2005). Real-time PCR is also known as quantitative PCR (qPCR). Real-time measurement is made possible by incorporation of fluorescent probes (e.g., SYBR green) that bind to the copied DNA; as the number of PCR cycles increases the fluorescent signal from amplified DNA intensifies (Figure 7). Real-time PCR can be divided into four phases; a linear ground phase in which the fluorescence is below background noise; an exponential phase with doubling and accurate measurement of the PCR product at every cycle; a linear phase during which the PCR begins to slow down and gives high variability between runs; and finally, a plateau phase, in which the reac-tants are consumed and no more products can be synthesized. The strength

32
of the fluorescence during the exponential phase is turned into quantifiable numerical values (Ishmael and Stellato, 2008). Genomic DNA amplification can provide important information, but does not reveal if the selected gene is expressed in a specific tissue.
Figure 7. Reverse transcriptase real-time PCR (Paper I). A) Reverse transcriptase PCR. A RNA molecule (1) is used for the first-strand cDNA synthesis (2) that leads to a cDNA:RNA hybrid (3). The hybrid will thereafter act as the first template in the forthcoming amplification of cDNA (4). B) SYBR Green incorporates to double-stranded DNA and emits fluorescent signals, (1), when the DNA breaks during de-naturation SYBR Green is released and is no longer able to emit (2). In the extension phase, double-stranded DNA is yet again produced and fluorescence re-appears (3). When the PCR cycle is finished, the fluorescent signal is twice as intense as before (4). C) The four phases—ground, exponential, linear and plateau—in a real-time PCR run.
To find out how gene expression is affected by different conditions, it is necessary to convert and amplify messenger RNA (mRNA) to complemen-tary DNA (cDNA) with reverse transcriptase before amplification by PCR (Bank et al., 1972; Ishmael and Stellato, 2008; Temin and Mizutani, 1970; Verma et al., 1972). This type of qPCR is then called reverse transcriptase-PCR (RT-qPCR).
A disadvantage with qPCR its lack of spatial resolution, which is im-portant in the analysis of complex tissues such as brain samples. To over-come this obstacle, one could use in situ hybridization; however, quantitative measurements with this method are not as accurate as those with qPCR.
The most recent invention in PCR methodology is absolute quantitative digital PCR (dPCR), which is based on the principle that a sample is diluted into a very large number of reaction chambers in which the PCR reaction can occur. In the beginning of the reaction there should be at most only one mol-ecule in each chamber, and by counting the chamber in which a PCR reac-tion has occurred the absolute number of DNA can be calculated (Baker,
1.
2.
3.
4.
Reverse transcriptase
PCR
RTase
A
Primer
B
Nucleotides
Denaturation
Extension
Polymerization
1.
2.
3.
4.
Flou
resc
ence
Cycle number
ThresholdGround
Exponential
Line
ar
}
Plateau
C

33
2012). Besides the benefit of absolute quantification, it does not need any baseline estimation of the fluorescence, a factor that sometimes leads to am-plification bias in qPCR (Cao et al., 2017). However, dPCR is in its early days, hence, it is much more expensive than qPCR and it is not yet widely used.
Radioimmunoassay – quantification of peptides Various immunoassays can be used to measure the concentration of macro-molecules in tissues and body fluids, and in this thesis, radioimmunoassays were used to measure opioid peptides and corticosterone (Figure 8). Two pioneers of radioimmunoassays were Solomon Berson and Rosalyn Yalow. During the 1950s they developed an assay that could detect insulin levels in human plasma (Yalow and Berson, 1959). For this accomplishment, Yalow was awarded the Nobel prize in 1977 (Nobelprize.org). The method has been developed further and today, proteins and peptides from a variety of tissues can be measured with radioimmunoassay.
Figure 8. Peptide analysis (Papers II-III). A) Frozen whole brains are stabilized to avoid enzymatic degradation (Paper III). B) Peptides are extracted from brain tissues by ion-exchange chromatography; samples are placed in columns filled with Se-phadex gel and the peptides are eluted by buffers with varying ion strength. C) The purified peptides are mixed with radioactive (125I) labeled peptides and a primary antibody. The immunoreactive level of antibody-bound peptide is thereafter meas-ured in a γ-counter.
The general principle of the method is that antigens labeled with radioiso-topes (e.g., 125I) bind to specific antibodies and form a labeled antigen-antibody complex. Unlabeled antigen from the tissue of interest competes with the labeled antigen and consequently inhibits the binding of the labeled antigen (Berson and Yalow, 1968). By using solutions of known concentra-tions of unlabeled peptides, a standard curve can be obtained and used to calculate unknown concentrations in a tissue sample.
125I labeled peptideUnlabeled peptide
Primary antibody
Bound peptide
Sephadex gelSample
Extractedpeptides
A
Buffers
B C

34
The advantages with radioimmunoassays are the high specificity and sen-sitivity, the possibility to measure different opioid peptides in several brain areas in the same animal, and the ease of analyzing a large number of sam-ples in a time efficient way. The disadvantage with the method is the risk of cross reactivity with similar peptide fragments.
There are some other, general problems in analyzing tissue levels of pep-tides. When dissecting tissues, it is difficult to separate specific subregions within brain areas (e.g., the basolateral or the central part of amygdala), therefore the analysis is usually conducted on the whole region. Post-mortem degradation of peptides is another problem with dissected tissues and careful handling of the samples during the purification of the peptides is crucial. However, the post-mortem degradation can be reduced by stabilizing the tissue by conductive heat transfer using the Stabilizor T1 instrument (Dena-tor AB) (see Tissue stabilization) (Segerström et al., 2016).
Opioid peptides in the brain can also be analyzed with in vivo microdialy-sis, which allows for real time measurements of the basal levels and the ef-fect of pharmacological and physiological challenges. The disadvantage with this method is that usually a single brain structure is measured in the same animal and the method is much more time consuming than radioimmunoas-says.

35
Aims
The overall aim was to investigate the effect of stressful environments or alcohol exposure early in life on the central nervous system and/or drug tak-ing later in life.
The specific aims were to examine the effects of: • Early life stress on opioid gene expression and subsequent voluntary
ethanol intake in adult rats. • Housing conditions on endogenous opioid peptides in the brain and the
pituitary of adolescent rats. • Episodic binge-like ethanol exposure during adolescence on the endoge-
nous opioid peptides in the brain and the pituitary. • Ethanol exposure in adolescence on in vivo dopamine dynamics in dorsal
striatum and amphetamine self-administration behavior in adulthood

36
Methods
An overview of the experiments is presented in Figure 9.
Figure 9. Experimental outline for Papers I-IV. A more detailed outline is presented in the respective Papers (I-IV). Words in bold represent the measured outcome pre-sented in this thesis. Early life refers to the period before weaning, postnatal days 0-21. Adolescence represents the period between postnatal days 22-63.
Animals Pregnant Wistar (Harlan Laboratories B.V., the Netherlands (Papers I, III and IV)) and Sprague Dawley (Scanbur BK AB, Sweden (Paper II)) rats arrived at gestational day 15 (Papers I, III and IV) or 17 (Paper II). The dams were kept on a 12 hours light/dark cycle in type IV cages (59 × 38 × 20 cm) containing wood chip bedding and nesting materials. Animals born on the
Paper I
Paper II
Timeline
Paper III
Maternal separation Voluntary ethanol drinkingOpioidgenes
Opioid peptides
Episodic ethanol binge exposureOpioid
peptides
Episodic ethanol binge exposure
Dopaminerecordings
DopaminerecordingsAmphetamine
self-administrationSucrose
self-administration
Paper IV
Early life Adolescence Adulthood
Experimentalhousing conditions
Opioidpeptides

37
same day (postnatal day 0) were sexed and cross-fostered to avoid biological littermates, each litter contained the same number of males and females. In Paper I, the litters were either reared under standard animal facility condi-tions or exposed to maternal separation (see Maternal separation). In Papers II-IV, male rats were randomly assigned to the different experimental groups immediately after weaning and group-housed on a reversed light/dark cycle before the experiments. All animals were group-housed during adolescence unless otherwise stated. In adulthood, the animals were group-housed with the exception of Paper I where single housing was necessary due to the ex-perimental procedure.
All animals had access to water and food ad libitum unless otherwise stat-ed. Animal weights were regularly monitored throughout the experiments. The cages were kept in temperature- (22 ± 1°C) and humidity- (50 ± 10%) controlled cabinets and the animal rooms had a masking background noise to minimize unexpected sounds.
All experimental procedures and animal care followed the guidelines of the Swedish Legislation on Animal Experimentation (Animal Welfare Act SFS1998:56) and the European Communities Council Directive (86/609/EEC) and were approved by the Uppsala Animal Ethical Commit-tee.
Maternal separation (Paper I) Litters were randomly assigned to one of the three rearing environments: 15 or 360 minutes of maternal separation (MS15/360) or conventional animal facility rearing (AFR). The separations of litters and dam were performed during the light period. Firstly, the dam was removed from the home cage to a type II (26 × 20 × 14 cm) macrolon cage containing wood chip bedding. Secondly, the litter was moved from the home cage to a type II cage (with bedding) and transferred to a heating cabinet (30.4 ± 0.2°C, 52.1 ± 0.7 hu-midity) in an adjacent room. The dams of the MS15 groups remained in the type II cage during the 15 minutes of separation and the pups returned to the home cage before the return of the dam. The dams in the MS360 group were returned to the home cage during the 360 minutes of separation. The separa-tions were performed every day from postnatal day 0 to 21. The animals in the AFR group were left undisturbed with the exception of care taking and weighing of the litter. On postnatal day 22, all animals were weighed and weaned. The separations and care taking was always performed in the same animal rooms and by the same person

38
Adolescent housing conditions (Paper II) At postnatal day 22, the rats were randomly assigned to three experimental groups: (i) a longer period (seven days) of single housing; (ii) group-housing with 30 minutes of short single housing before decapitation; and (iii) group housing until decapitation (i.e., the controls).
Adolescent ethanol exposure (Paper III-IV) Between four and nine weeks of age, animals received orogastric (i.e., ga-vage) administration of either water or 2 g/kg (20% v/v) ethanol diluted with water. Administrations were given at 09:00 on three consecutive days fol-lowed by four days without treatment. Animals were kept group-housed during the entire ethanol exposure period.
Adult voluntary ethanol intake (Paper I) The rats were individually housed in cages containing wood chip bedding and a wooden house. For three consecutive days per week, the rats had free access to both ethanol and water. During the first week, the rats were given 24 hours ad libitum access to 5% (v/v) ethanol and two hours during the second week. Thereafter ethanol concentration was increased to 20% (v/v) and access was limited to two hours. Ethanol and water were changed every day during the three days and the bottle position was altered daily to avoid position preference. At the end of each drinking session, ethanol and water intake was measured by weighing the bottles.
Operant self-administration (Paper IV) Operant boxes apparatus Self-administration was conducted in sound-attenuated operant boxes (MED Associates Inc., VT, USA) equipped with two stimulus lights above two retractable, stainless steel levers. A white house-light placed on the wall opposite the levers was on during the entire session; a ventilating fan operat-ed throughout the sessions and served as a masking noise. Intravenous solu-tions were delivered by infusion pump; a plastic syringe placed in the pump was connected to the implanted catheter through CoEx tubing (Harvard Ap-paratus, Kent, UK) and protected by a flexible metal leash. Experiments were run and data collected with the MED-PC software. The experiments were conducted in the same operant boxes for both sucrose training and am-

39
phetamine sessions. Ethanol-treated animals and water controls were pro-cessed simultaneously throughout all phases of the self-administration pro-cedure.
Sucrose training Food restriction was initiated following the last session of ethanol exposure and was maintained throughout the sucrose-training period. Animal weights were carefully monitored and were not allowed to decrease more than 15 % after the commencement of food restriction. After 48 hours of food re-striction, the training schedule to self-administer 45 mg sucrose food pellets (5-TUL, TestDiet, MO, USA) on a fixed ratio-1 (FR1) was initiated. The criteria for fulfilled self-administration training were 100 active lever presses within 30 minutes and a specificity >0.85 for the active lever.
Intravenous amphetamine self-administration Intravenous catheters (CamCaths, Ely, UK) were implanted into the right jugular vein under isoflurane (Forene Abbott, Sweden) anaesthesia. Postop-erative analgesia (buprenorphine and carpofen) and antibiotic (amoxicillin) were administered. The catheters were flushed with a heparin solution before and after each session, and a heparinized glycerol lock solution was used over weekends. Catheter patency was tested before the start and at the end of the study with an infusion of the short-acting anaesthetic agent propofol.
After a minimum of four days of recovery from surgery, rats from both the ethanol and water groups were allowed to self-administer amphetamine (d-amphetamine sulfate, Sigma-Aldrich, MO, USA) on a daily 60 minutes FR3 schedule of reinforcement. Three responses on the active lever resulted in an infusion of amphetamine (0.1 mg/kg/infusion) and a 10 seconds time-out period was initiated during which both levers were retracted and a white stimulus light above the active lever was turned on. The lever designated as the active one was switched between sucrose training and intravenous self-administration. The maximum number of rewards during the 60 minutes baseline sessions was set to 20. A press on the inactive lever had no pro-grammed consequences, but the activity was recorded by the software for statistical analysis.
After five 60 minutes FR3 amphetamine (0.1 mg/kg/infusion) baseline sessions to familiarize the animals with the amphetamine effects, operant requirements were switched to the PR format. Under this schedule of rein-forcement, the response requirement started at one press on the active lever and escalated for each drug infusion delivered according to following scheme: 1, 2, 4, 6, 9, 12, 15, 20, 25, 32, 40, 50, 62, 77, 95, 118, 145, 178, 219, 268, 328, 402, 492, 603, 737, 901 see Richardson and Roberts (1996). The PR schedule was tested at two doses (0.1 and 0.05 mg/kg per infusion)

40
for two consecutive days each. The sessions ended when one hour had passed since the last reward or after a maximum session time of four hours. The breakpoint was defined as the total number of infusions during the ses-sion. Additionally, to test the dose-response function on an FR schedule the unit dose of amphetamine (0, 0.025, 0.05 or 0.1 mg/kg/infusion) was varied and each dose was tested for three consecutive 90 minutes FR3 sessions. The first session of each dose in PR and dose-response trials was considered an acclimatization one and these data collected were not used in the statistical analysis.
High speed chronoamperometry (Paper IV) Dopamine recordings were conducted using carbon fibre microelectrodes (Quanteon, KY, USA). A high-speed chronoamperometric protocol was used (550mV, 1 Hz sampling rate, 200 ms total) via a FAST16-mkII recording system (Fast Analytical Sensing Technology, KY, USA). Electrode-pipette assemblies were prepared and calibrated immediately before in vivo record-ings, as previously described (Gerhardt and Hoffman, 2001; Littrell et al., 2012). Briefly, electrodes were coated with Nafion and calibration were conducted with cumulative additions of ascorbic acid and dopamine. Elec-trodes had a detection limit of 0.0237±0.0037 μM and a selectivity of 3864.35±881.16 for dopamine over ascorbic acid. Responses to dopamine were linear, with an average correlation coefficient (R2) of 0.827±0.025. The average reduction/oxidation ratio was 0.628±0.011, which indicates specific dopamine detection (Gerhardt and Hoffman, 2001). After calibration, a mi-cropipette filled with isotonic potassium chloride solution was affixed with the tip 150-200 μm from the recording site of the electrode.
Animals were anaesthetized with Inactin and body temperature main-tained with a thermostatic heating pad. The electrode-pipette assembly was stereotaxically placed in the dorsolateral striatum (Anterior-Posterior: +1.0, Medial-Lateral: +3.0, Dorsal-Ventral: -4.2 mm) and an Ag/AgCl reference electrode placed in the brain contralaterally and remotely from the electrode-recording site.
After one hour post-surgical stabilization of the electrode and surrounding tissue, 100 nl of potassium chloride solution was locally ejected using pres-sure ejection (PicoSpritzer III, Parker Hannifin Corporation, NJ, USA, ejec-tion pressure <22 psi, ejection time < two seconds) and the subsequent do-pamine release was detected by the electrode as a peak of rising dopamine concentration. Ejections were repeated every 10 minutes until three succes-sive consistent dopamine releases were recorded for use as baseline refer-ence peaks. Five minutes after the third reference peak was evoked, a single 2.0 mg/kg dose of amphetamine was injected via the tail vein. Five minutes post-injection of amphetamine, dopamine release was evoked again and this

41
was then repeated every 10 minutes until 55 minutes post-injection. After the measurements, the animals were killed and the brains removed and frozen for subsequent histological identification of electrode location.
Quantitative polymerase chain reaction (Paper I) To purify and isolate RNA from brain tissues, All Prep DNA/RNA /miRNA Universal Kit (Qiagen, Sweden) was used according to the manufacturer’s instructions. Isolated RNA was quantified using a Nanodrop ND 1000 spec-trophotometer. RNA was converted to cDNA by using the QuantiTect Re-verse Transcription Kit (Qiagen AB, Sweden) including a genomic wipeout reaction. The synthesis of cDNA was performed at 42°C for 30 minutes and inactivated at 95°C. Newly synthetized cDNA was diluted and stored at -20°C. The diluted cDNA was used for qPCR (CFX96, Bio-Rad, Sweden) to assess the expression of the genes of interest – Pomc, Penk, Pdyn, Oprm1, Oprd1 and Oprk1 – as well as the housekeeping genes Actb, Gapdh and RpI19. Primers were designed using Primer 3 (http://frodo.wi.me.edu/) and crosschecked using Primer Map (http://www.bioinformatics.ord/sms2/ pri-mer_map.html). The final reaction mixture contained SYBR® Green Super-mix (Bio-Rad), forward and reverse primer (Thermo Fisher Scientific, Ger-many), cDNA template and MilliQ-water. Each sample was run in tripli-cates. To control for genomic DNA contamination, an on-column DNase treatment was performed during extraction. Before the cDNA synthesis, a genomic DNA wipeout extraction was used; and the primers were designed across two adjacent exons to avoid unspecific amplification of genomic DNA. Each real time PCR plate contained positive internal controls.
Tissue stabilization (Paper III) Deep frozen (-80°C) whole brains were placed in -20°C the day before stabi-lization to reduce a steep temperature gradient. The brains and pituitaries (separated from the brain before freezing) were stabilized with the bench-top Stabilizor T1 (Denator AB, Sweden) according to the manufacturer´s manual (http://www.denator.com). The stabilization process involves a combination of conductive heat transfer and pressure under vacuum to avoid enzymatic degradation (e.g., of peptides) that might occur during freeze-thawing pro-cesses (Segerström et al., 2016). Whole brains were placed in a Maintainor Tissue card (Denator AB, Sweden) and stabilized in “frozen structure pre-serve” mode and thereafter in “fresh structure preserve” mode to ensure suf-ficient treatment. The stabilized brains were thereafter dissected on ice ac-cording to the rat brain atlas of Paxinos and Watson (1997) and stored at -80°C.

42
Radioimmunoassay (Paper II-III) For corticosterone measurements trunk blood was collected. Blood samples were centrifuged at room temperature after which the serum was stored at -20°C. Analysis of corticosterone was performed using the commercial Im-munChemTM Double Antibody Corticosterone 125I radioimmunoassay kit for rats and mice (ICN Biomedicals Inc., CA, USA).
The dissected tissues were heated in acetic acid in a 95° water bath for five minutes and thereafter homogenized with a sonicator. After centrifuga-tion, the supernatants were run on ion exchange columns with conditioned with Sephadex gel. The peptides were eluted with pyridine and formic acid containing buffers with varying ion strength. For more information about tissue extraction and purification see Christensson-Nylander et al. (1985). Radioimmunoassays for and dynorphin B (Nylander et al., 1997), MEAP (Nylander et al., 1995) and N/OFQ (Ploj et al., 2000) have preciously been described. Antiserum for the respective peptide was generated in rabbits. Antibody-bound peptides in the dynorphin B and N/OFQ assays were sepa-rated from free peptides by adding goat-anti-rabbit-IgG and normal rabbit serum. For the MEAP assay, separation was performed by adding charcoal suspension. For β-endorphin measurements, commercial β-endorphin radio-immunoassay kits from Phoenix Pharmaceuticals (CA, USA) were used according to manufacturer’s protocol.
In this thesis, the immunoreactive levels measured by radioimmunoassay will only be named levels from here on.
Statistical analysis Statistical analyses were performed using Statistica (Statsoft Inc., OK, USA) version 12 in Papers II and IV and version 13 in Papers I and III and SPSS (IBM, New York, USA) version 22 in Paper I. For the multivariate data analysis (Paper II), SIMCA-P+ 12.0 (Umetrics AB, Umeå, Sweden) was used. The statistically significant level was set to p < 0.05.
Parametrical statistics Factorial analysis of variance (ANOVA) was used to analyze differences in opioid peptide and corticosterone levels (Papers II and III). A repeated measures ANOVA was used to analyze time-related differences in dopamine and the dose-response function of amphetamine self-administration in Paper IV. Post hoc analyses were performed with Bonferroni (Paper II) and Tuk-ey’s HSD (Papers III and IV). To test differences in the PR trials in Paper IV, Student’s t-test was applied.

43
Non-parametrical statistics Kruskal Wallis, together with the Mann-Whitney post hoc test, was used to analyze effects of rearing and ethanol intake, on gene expression in Paper I. To investigate possible interactions between rearing and intake a two-factor analysis was tested using the Univariate General Linear Model test, two-way ANOVA with type III sum of squares. Correlation analyses in paper I were performed using Spearman correlations.
Multivariate analysis In Paper II, the multivariate method partial least square discriminant analysis was used to complemented conventional statistics to illustrate patterns among individual rats and the peptide and corticosterone levels of im-portance for creating such patterns.
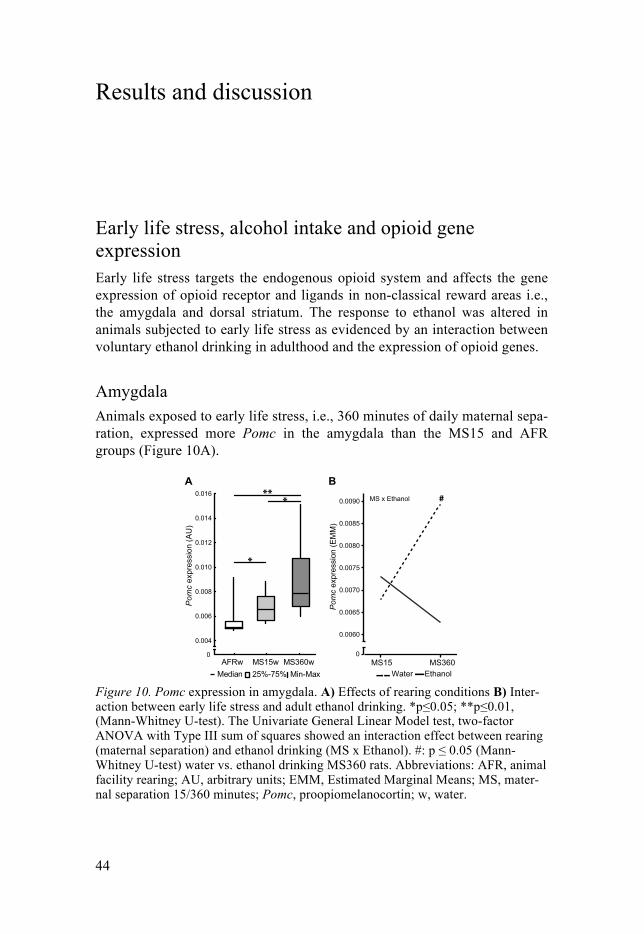
44
Results and discussion
Early life stress, alcohol intake and opioid gene expression Early life stress targets the endogenous opioid system and affects the gene expression of opioid receptor and ligands in non-classical reward areas i.e., the amygdala and dorsal striatum. The response to ethanol was altered in animals subjected to early life stress as evidenced by an interaction between voluntary ethanol drinking in adulthood and the expression of opioid genes.
Amygdala Animals exposed to early life stress, i.e., 360 minutes of daily maternal sepa-ration, expressed more Pomc in the amygdala than the MS15 and AFR groups (Figure 10A).
Figure 10. Pomc expression in amygdala. A) Effects of rearing conditions B) Inter-action between early life stress and adult ethanol drinking. *p≤0.05; **p≤0.01, (Mann-Whitney U-test). The Univariate General Linear Model test, two-factor ANOVA with Type III sum of squares showed an interaction effect between rearing (maternal separation) and ethanol drinking (MS x Ethanol). #: p ≤ 0.05 (Mann-Whitney U-test) water vs. ethanol drinking MS360 rats. Abbreviations: AFR, animal facility rearing; AU, arbitrary units; EMM, Estimated Marginal Means; MS, mater-nal separation 15/360 minutes; Pomc, proopiomelanocortin; w, water.
AFRw MS15w MS360w
0.004
0.006
0.008
0.010
0.012
0.014
0.016
Pom
c ex
pres
sion
(AU
)
Median 25%-75% Min-Max
*
***
0MS360MS15
Pom
c ex
pres
sion
(EM
M)
0.0090
0.0085
0.0080
0.0075
0.0070
0.0065
0.0060
0
MS x Ethanol #
EthanolWater
A B

45
Furthermore, ethanol affected expression solely in the MS360 group; the expression of Pomc was lower than the water-drinking MS360 rats and clos-er to the MS15 rats, whose expression of Pomc was unaffected by ethanol drinking (Figure 10B). These results indicate a normalizing effect after etha-nol drinking, i.e., a reduction of the initially high, Pomc expression levels in the MS360 rats.
The product from Pomc expression is proopiomelanocortin, which is cleaved enzymatically into several biologically active peptides (see Endoge-nous opioids and Figure 2); hence, it is difficult to interpret the outcome of the alterations in Pomc expression. The literature regarding ethanol, β-endorphins and the amygdala is scarce but there are some reports that β-endorphin increases in the central amygdala after acute exposure to ethanol (Lam and Gianoulakis, 2011; Lam et al., 2008). The higher levels of Pomc expression could also be due to increased adrenocorticotropic hormone (ACTH) and/or melanocyte-stimulating hormone (MSH). A functional study by Brunson et al. (2001) shows that ACTH can act directly on neurons in the amygdala and downregulate the expression of CRF, which is highly involved in the response to ethanol (Koob, 2008). Furthermore, ethanol exposure dur-ing adolescence and adulthood reduces alpha-MSH in the central amygdala (Lerma-Cabrera et al., 2013; Navarro et al., 2008), an effect that resembles the interaction effect found herein.
Dorsal striatum In the dorsal striatum, the expression of opioid receptors was dependent on rearing conditions (Figure 11).
Figure 11. Effects of rearing conditions on A) Oprd1, B) Oprk1 and C) Oprm1 gene expression in the dorsal striatum. *p≤0.05; **p≤0.01 (Mann-Whitney U-test). Ab-breviations: AFR, animal facility rearing; AU, arbitrary units; MS, maternal separa-tion 15/360 minutes; w, water drinking animals; Oprd1, Opioid receptor delta 1; Oprk1, opioid receptor kappa 1; Oprm1, opioid receptor mu 1.
AFRw MS15w MS360w0
0.0005
0.0010
0.0015
0.0020
0.0025
0.0030
0.0035
0.0040
0.0045
Opr
d1 e
xpre
ssio
n (A
U)
***
Median 25%-75% Min-Max
AFRw MS15w MS360w0
0.002
0.004
0.006
0.008
0.010
0.012
Opr
k1 e
pres
sion
(AU
)
****
AFRw MS15w MS360w0
0.002
0.004
0.006
0.008
0.010
0.012
Opr
m1
expr
essi
on (A
U)
*A B C

46
Rats exposed to 360 minutes of maternal separation had higher expression of Oprd1 (Figure 11A) and Oprk1 (Figure 11B) than the AFR and MS15 rats, and higher Oprm1 expression than the AFR (Figure 11C).
Previous studies show that rats exposed to MS360 have lower immunore-active levels of MEAP in the dorsal striatum than MS15 rats (Gustafsson et al., 2008; 2007). Decreased MEAP could be either a cause or an effect of upregulated δ-opioid receptors, i.e., increased expression of Oprd1. The exact role of opioid receptors in the dorsal striatum is not fully understood, but they are regulators of other neurotransmitters such as dopamine, GABA and glutamate (Atwood et al., 2014; Banghart et al., 2015; Christensson-Nylander et al., 1986; Herrera-Marschitz et al., 1986; Steiner and Gerfen, 1998).
Evidence that ethanol affects endogenous opioids in the dorsal striatum can be seen in a series of experiments by Mendez et al., in which they stud-ied the acute impact of forced administration of ethanol. They show that a single administration of ethanol (2.5 g/kg) increased both binding of a δ-receptor ligand and Penk expression 1–2 hours later (Mendez et al., 2004; 2008). Furthermore, increased binding was not found when using a μ-receptor ligand (Mendez et al., 2003). Voluntary drinking shows that high intake increases δ-receptor activity and that intra-striatal injection of naltrin-dole (a selective δ-receptor antagonist) decreases ethanol drinking (Nielsen et al., 2012). However, the literature is inconclusive regarding pharmacolog-ical treatments with δ-receptor antagonists and agonists and the effects on ethanol intake; studies show a decrease, increase or no effect on intake after δ-receptor modulation (Ciccocioppo et al., 2002; Franck et al., 1998; Hyytia and Kiianmaa, 2001; Ingman et al., 2003; Krishnan-Sarin et al., 1995; Margolis et al., 2008).
In Paper I, an interaction between early life stress and ethanol intake was found in the expression of Oprd1 (Figure 12A) and Oprk1 (Figure 12B). The expression of both genes was reduced in ethanol-drinking MS360 rats com-pared to the water controls, whereas ethanol-drinking MS15 rats had similar expression levels as the controls. These data agree with previous findings showing that voluntary drinking results in higher striatal MEAP levels in MS360 rats whereas the MEAP were unaffected by ethanol in the MS15 rats (Gustafsson et al., 2007). Furthermore, MS360 and MS15 rats respond dif-ferently to naltrexone in a voluntary ethanol intake paradigm (Daoura and Nylander, 2011). The lower expression of δ-receptors seen in our study and the previous findings of higher MEAP after ethanol, could contribute to dif-ferences in treatment response.

47
Figure 12. Interaction and main effects for early life stress and adult ethanol drink-ing on A) Oprd1 and B) Oprk1 expression in the dorsal striatum. The Univariate General Linear Model test, two-factor ANOVA with Type III sum of squares shows an interaction effect between rearing (maternal separation) and ethanol drinking (MS x Ethanol) and a main effect of ethanol (Ethanol). *p<0.05, ***p<0.001 MS360 water vs. MS 360 ethanol (Mann-Whitney U-test). Abbreviations: EMM, estimated marginal means; MS, maternal separation 15/360; Oprd1, opioid receptor delta 1; Oprk1, opioid receptor kappa 1.
Adolescent housing conditions and opioid peptides Short and prolonged single housing in early adolescence resulted in time-, area- and peptide specific changes in endogenous opioids.
Nociceptin/Orphanin FQ A short period of single housing (30 minutes) increased N/OFQ in the amygdala, nucleus accumbens and medial prefrontal cortex (Figure 13). Furthermore, a trend (p=0.07) of higher levels was seen in the hypothalamus. The animals housed singly for a short period had higher levels in the medial prefrontal cortex and hypothalamus than the rats exposed to a prolonged period of single housing (Figure 13). N/OFQ were unaffected by seven days of single housing (Figure 13). Increased serum corticosterone was seen only after the short-term single housing.
Corticosterone levels are unaffected by a centrally administered NOP re-ceptor antagonist (UFP-101), but the antagonist attenuates the effect of in-tracerebroventricular N/OFQ, which suggests that N/OFQ is not tonically active (Leggett et al., 2006). An acute stress challenge increases Oprl1 (i.e., the gene encoding N/OFQ) expression and CRF in central and basomedial amygdala, and acute stress also increases Oprl1 expression in the para-ventricular hypothalamus whereas chronic stress does not (Ciccocioppo et al., 2014; Green and Devine, 2009). Furthermore, N/OFQ blocks CRF-
MS360MS15
Opr
d1 e
xpre
ssio
n (E
MM
)
0.0030
0.0028
0.0025
0.0023
0.0020
0.0018
EthanolWaterMS360MS15
Opr
k1 e
xpre
ssio
n (E
MM
)
0.009
0.008
0.007
0.006
0.005
0.004
A B
0 0
MS x EthanolEthanol
MS x EthanolEthanol
******

48
induced inhibitory postsynaptic potentials in central and basolateral amygda-la; this inhibition is even greater in restrained rats (Ciccocioppo et al., 2014). Behavioral data regarding the effects of NOP activation are conflicting, as anxiolytic effects are reported for both agonists, and antagonists (Duzzioni et al., 2011; Fernandez et al., 2004; Gavioli et al., 2003; Green et al., 2007; Jenck et al., 2000; Koster et al., 1999; Varty et al., 2005; 2008; Vitale et al., 2009).
Figure 13. The effects of short and prolonged single housing on N/OFQ in different brain areas. The values are presented as a percentage of the levels in the control group. *p<0.05; **p<0.01, compared to group housed controls; #p<0.05: ##p<0.01, compared to single housed rats (one-factor ANOVA followed by Bonferroni post hoc test). Abbreviations: AMY, amygdala; FCx, frontal cortex; HC, hippocampus; HT, hypothalamus; LSH, long single housing; MPFCx, medial prefrontal cortex; NAc, nucleus accumbens; N/OFQ, nociceptin/orphanin FQ; PAG, periaqueductal gray; SSH, short single housing; Sn, substantia nigra; dStr, dorsal striatum; VTA; ventral tegmental area.
A hypothesis in Witkin et al. (2014), suggests that N/OFQ is recruited after acute stress to facilitate adaption to the new environmental condition. When the stressful stimuli are protracted, stimulation of N/OFQ activity at extrahy-pothalamic areas may serve to attenuate the pathological consequences asso-ciated with chronic stress. This hypothesis could explain why the acute stress (i.e., short single housing) increased N/OFQ and circulating corticosterone, but the levels were similar to those of group-housed non-stressed, rats during prolonged single housing.
When stress-induced effects on the N/OFQ system are investigated, the methods used are usually: restraint stress, forced swim test, defeat, chronic unpredictable stress, open field, or elevated plus maze. In Paper II, single housing, a mild, common, and sometimes necessary procedure in experi-mental research, was also able to induce a significant stress response in ado-lescent animals.

49
Methionine-Enkephalin-Arginine6-Phenylalanine7
Rats subjected to seven days of single housing in early adolescence had low-er MEAP levels than group-housed animals in the hypothalamus, frontal cortex, medial prefrontal cortex, hippocampus, amygdala, substantia nigra and periaqueductal gray (Figure 14). In addition, reduced hippocampal levels of MEAP were found in short- compared to prolonged single housed rats (Figure 14). Short single housing had no effect on MEAP levels (Figure 14).
Figure 14. The effects of short and prolonged single housing on MEAP in different brain areas. The values are presented as a percentage of the levels in the control group. *p<0.05; **p<0.01, compared to group housed controls; #p<0.05: ##p<0.01, compared to single housed rats (one-factor ANOVA followed by Bonferroni post hoc test). Abbreviations: AMY, amygdala; FCx, frontal cortex; HC, hippocampus; HT, hypothalamus; LSH, long single housing; MEAP, Methionine-enkephalin-Arginine6-Phenylalanine7, MPFCx, medial prefrontal cortex; NAc, nucleus accum-bens; PAG, periaqueductal gray; SSH, short single housing; Sn, substantia nigra; dStr, dorsal striatum; VTA; ventral tegmental area.
The enkephalin system continues to mature after birth and during the first weeks of life δ-receptors and enkephalin levels increase (Bayon et al., 1979; Kornblum et al., 1987; Spain et al., 1985); therefore, this system might be extra sensitive to environmental disturbances. As mentioned in the introduc-tion, social contacts during adolescence are important for normal develop-ment and endogenous opioids are highly involved in mediating the reward-ing effect of this behavior. Deprivation of social contact during this period could result in an increased state of anxiety as reflected by reduced levels of enkephalins.
Behavioral tests with Oprd1 and Penk knockout mice show increased anxiety-like behavior in a variety of tests (Filliol et al., 2000; Konig et al., 1996; Ragnauth et al., 2001). Pharmacological studies with systemic admin-

50
istrations and local injections of δ-receptor agonists into the amygdala, cin-gulate cortex, and hippocampus decrease the anxious behavior (Perrine et al., 2006; Randall-Thompson et al., 2010; Saitoh et al., 2004; 2018; Vergura et al., 2008). Likewise, the administration of antagonists increases the anxie-ty-like behaviors (Narita et al., 2006; Perrine et al., 2006; Saitoh et al., 2004; 2005). Surprisingly however, specific Oprd1 gene inactivation in forebrain GABAergic neurons results in a low-anxiety phenotype (Chu Sin Chung et al., 2015), indicating that there might be a more complex and regions-specific effects of enkephalin signaling and anxiety. Furthermore, it has been shown that enkephalin expression in the basolateral amygdala is linked to stress coping (Berube et al., 2013; 2014) and enkephalins may be implicated in both vulnerability and resilience to stress (Henry et al., 2017). Palm and Nylander (2014a) investigated the effects of 24 hours or six weeks of single housing and showed that MEAP increases in the hippocampus, medial pre-frontal cortex and in the pituitary gland at 24 hours, but a residual increase is only found in the cingulate cortex. The results from Paper II, together with those in Palm and Nylander (2014a) suggest a time-dependent change in the MEAP system after exposure to single housing, with an initial increase after 24 hours. This is followed by declining levels and, after a period of time, the levels reset.
Dynorphin B Dynorphin B was unaffected by either short or prolonged single housing. This was also demonstrated by Palm and Nylander (2014a) where dynorphin B levels remained unchanged 24 hours and six weeks after isolation, with an exception of reduced levels in the amygdala. The maturation of the dy-norphin system occurs at an earlier stage than the enkephalin one, (Kornblum et al., 1987; McDowell and Kitchen, 1987); furthermore, κ-receptors and dynorphin signaling are not as involved as β-endorphin and enkephalins in the age-specific play behavior (Niesink and Vanree, 1989; Vanderschuren et al., 1995b). Therefore, single housing may not affect the dynorphins to the same extent as enkephalins.
Adolescent alcohol exposure Intermittent ethanol exposure during adolescence induced alterations in en-dogenous opioid peptides and dopamine dynamics in brain areas involved in stress regulation, reward, and reinforcement.
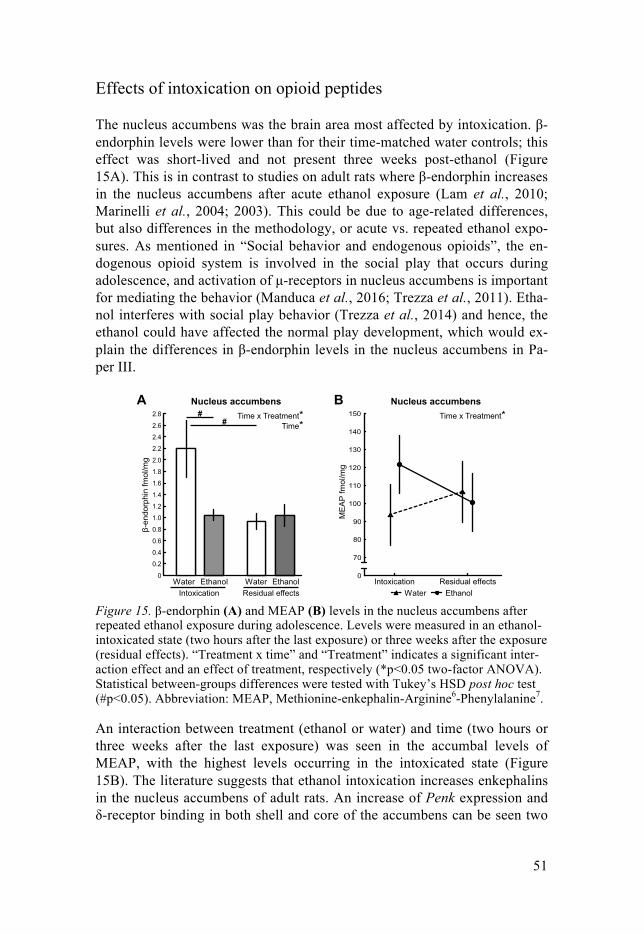
51
Effects of intoxication on opioid peptides
The nucleus accumbens was the brain area most affected by intoxication. β-endorphin levels were lower than for their time-matched water controls; this effect was short-lived and not present three weeks post-ethanol (Figure 15A). This is in contrast to studies on adult rats where β-endorphin increases in the nucleus accumbens after acute ethanol exposure (Lam et al., 2010; Marinelli et al., 2004; 2003). This could be due to age-related differences, but also differences in the methodology, or acute vs. repeated ethanol expo-sures. As mentioned in “Social behavior and endogenous opioids”, the en-dogenous opioid system is involved in the social play that occurs during adolescence, and activation of μ-receptors in nucleus accumbens is important for mediating the behavior (Manduca et al., 2016; Trezza et al., 2011). Etha-nol interferes with social play behavior (Trezza et al., 2014) and hence, the ethanol could have affected the normal play development, which would ex-plain the differences in β-endorphin levels in the nucleus accumbens in Pa-per III.
Figure 15. β-endorphin (A) and MEAP (B) levels in the nucleus accumbens after repeated ethanol exposure during adolescence. Levels were measured in an ethanol-intoxicated state (two hours after the last exposure) or three weeks after the exposure (residual effects). “Treatment x time” and “Treatment” indicates a significant inter-action effect and an effect of treatment, respectively (*p<0.05 two-factor ANOVA). Statistical between-groups differences were tested with Tukey’s HSD post hoc test (#p<0.05). Abbreviation: MEAP, Methionine-enkephalin-Arginine6-Phenylalanine7.
An interaction between treatment (ethanol or water) and time (two hours or three weeks after the last exposure) was seen in the accumbal levels of MEAP, with the highest levels occurring in the intoxicated state (Figure 15B). The literature suggests that ethanol intoxication increases enkephalins in the nucleus accumbens of adult rats. An increase of Penk expression and δ-receptor binding in both shell and core of the accumbens can be seen two
0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
2.0
2.2
2.4
2.6
2.8
EthanolWaterEthanolWaterIntoxication Residual effects
Time*Time x Treatment*#
#
0
70
80
90
100
110
120
130
140
150
A B
ME
AP
fmol
/mg
β-en
dorp
hin
fmol
/mg
Residual effectsIntoxication
Time x Treatment*
EthanolWater
Nucleus accumbens Nucleus accumbens

52
hours after ethanol administration (Mendez and Morales-Mulia, 2006; Oliva et al., 2008). Awake rats have increased accumbal levels of Methionine-enkephalin when injected with 1.6 g/kg ethanol. However, higher and lower doses than 1.6 g/kg have no effect on Methionine-enkephalin (Marinelli et al. 2005). In anesthetized rats, 2.5 g/kg leads to a peak of Methionine-enkephalin at 30 minutes whereas lower doses delay the peak (Mendez et al. 2010).
Residual effects on opioid peptides In several brain areas, the effects of ethanol exposure on MEAP persisted three weeks post ethanol (Figure 16).
Figure 16. MEAP levels in (A) amygdala, (B) ventral tegmental area and (C) sub-stantia nigra after repeated ethanol exposure in adolescence. MEAP were measured in an ethanol-intoxicated state (two hours after last exposure) or three weeks after the last exposure (residual effects). “Treatment” indicates a significant effect of treatment, (*p<0.05, **p<0.01 two-factor ANOVA). Abbreviations: EtOH, ethanol; MEAP, Methionine-enkephalin-Arginine6-Phenylalanine7.
MEAP were lower in the amygdala after ethanol exposure (Figure 16A). As mentioned in “Adolescent housing conditions and opioid peptides”, low levels of enkephalins are associated with increased anxiety and stress. Im-portantly, the amygdala is a key region for mediating these emotional states (Janak and Tye, 2015; Tye et al., 2011). In the present thesis, behavioral manifestations after adolescent ethanol exposure were not studied, but other studies have reported increased anxiety-like behaviors (Van Skike et al., 2015; Varlinskaya et al., 2016), which could be a consequence of low enkephalin levels. δ-receptor knockout mice have an increased consumption of ethanol (Roberts et al., 2001) and their elevated ethanol intake may be a way to reduce an innately high anxiety level (Chu Sin Chung and Kieffer, 2013). The low enkephalin after adolescent ethanol exposure may therefore constitute a risk factor for elevated intake of ethanol later in life.
Amygdala
0
20
40
60
80
100
120
MEA
P fm
ol/m
g
EtOHWaterEtOHWaterIntoxication Residual effects
Treatment*Substantia nigra
0
2
4
6
8
10
12
14
16
18
MEA
P frn
ol/m
g
Ventral tegmental area
0
5
10
15
20
25
30
MEA
P fm
ol/m
g
Treatment* Treatment**
EtOHWaterEtOHWaterIntoxication Residual effects
EtOHWaterEtOHWaterIntoxication Residual effects
A B C

53
In the VTA and substantia nigra, MEAP were higher in ethanol-exposed rats (Figure 16B and 16C). In the intoxicated state (two hours), the dy-norphin B levels in the substantia nigra were the same for the ethanol-treated group and water controls, but higher levels were found in the ethanol group at three weeks after the last exposure. The substantia nigra and VTA contain dopaminergic neurons that extend into the striatal, limbic and cortical areas (Björklund and Dunnett, 2007). Importantly, endogenous opioids are highly involved in regulating dopamine output (Christensson-Nylander et al., 1986; Fields and Margolis, 2015; Herrera-Marschitz et al., 1986).
Residual effects on in vivo dopamine in the dorsal striatum Repeated episodic binges of ethanol during adolescence did affect dopamine dynamics in dorsal striatum.
A single amphetamine challenge significantly increased the peak area re-sponse in water controls at 5, 15, 25 and 35 minutes. In the ethanol-exposed animals, the increase was only seen at the 5-minute time point. The peak area involves two parameters—peak amplitude (the amount dopamine re-leased) and T80 (the time of dopamine reuptake). For more information see “In vivo high-speed chronoamperometry – analysis of dopamine”. The dif-ference in peak area was mainly driven by an interaction effect between time and treatment in peak amplitude whereas the T80 was comparable for the two groups (see Figure 17 A and B). Hence, these results indicate that ado-lescent ethanol exposure suppresses dopamine release rather than inducing a more efficient dopamine removal.
Palm and Nylander (2014b) investigated the dopamine response in dorsal striatum after voluntary ethanol intake and showed that ethanol drinking in adolescence reduces basal (i.e., pre-amphetamine challenge) potassium-induced peak amplitudes in adulthood, but the response to a single amphet-amine challenge is unaffected. The discrepancy with our results could be due to methodological differences; Paper IV describes the effects of ethanol binges during adolescence independent of phenotypes, whereas Palm and Nylander (2014b) describes the effects in a drinking phenotype.
The effect of adolescent ethanol exposure in the ventral part of striatum is more studied than the dorsal part. Shnitko and colleagues (2016) did not find any alterations in baseline dopamine release or uptake after electric stimula-tion of VTA in ethanol-exposed rats. Studies using microdialysis show that repeated ethanol exposure increases the basal levels of extracellular dopa-mine in adolescent rats, but these effects do not persist into adulthood (Pascual et al., 2009). An ethanol challenge after a history of adolescent ethanol results in decreased accumbal dopamine release 7 and 14 days after treatment, but at 28 days the effect is diminished (Zandy et al., 2014). In contrast, it has also been reported that adolescent exposure increases extra-cellular dopamine levels after an ethanol challenge (Badanich et al., 2007;

54
Pascual et al., 2009; Philpot et al., 2009). Furthermore, adolescent alcohol blunts the acute ethanol effects on evoked dopamine release in nucleus ac-cumbens (Shnitko et al., 2016). Beside the striatum, effects of adolescent ethanol exposure on dopamine dynamics have also been reported in the VTA (Avegno et al., 2016; Schindler et al., 2016) and cortical regions (Trantham-Davidson et al., 2017)
Figure 17. In vivo chronoamperometric recordings in the dorsal striatum. Ampli-tudes (A and C) and T80 (B and D) measurements over time after different drug exposures. The graphs show the potassium-induced dopamine responses after an intravenous amphetamine challenge (2 mg/kg intravenously) as percent (mean± SEM) of reference values after adolescent ethanol or water exposure (A and B) and after adolescent ethanol exposure followed by amphetamine self-administration (C and D). **p<0.01 compared to the time point (−5) before the amphetamine chal-lenge, repeated measures ANOVA followed by Tukey’s HSD post hoc test. Abbre-viations; Amph, amphetamine; SA, self-administration.
Subsequent amphetamine self-administration Although a history of adolescent ethanol exposure increases subsequent vol-untary intake of ethanol in rats (Alaux-Cantin et al., 2013; Amodeo et al., 2017; Criado and Ehlers, 2013; Pascual et al., 2009), the literature is incon-clusive; several studies report only subtle or no effects on subsequent intake
-25 -15 -5 5 15 25 35 45 55minutes
80
100
120
140
160
180
% o
f ref
eren
ce a
mpl
itude
0
-25 -15 -5 5 15 25 35 45 55minutes
80
100
120
140
160
180
200
220
% o
f ref
eren
ce T
80
0
-25 -15 -5 5 15 25 35 45 55minutes
% o
f ref
eren
ce T
80
0
**** ** **
Ethanol + SA Amph Water + SA Amph
-25 -15 -5 5 15 25 35 45 55minutes
80
100
120
140
160
180
% o
f ref
eren
ce a
mpl
itude
0
Amphetamine challenge
80
100
120
140
160
180
200
220
Ethanol Water
Ethanol Water
A B
C D Ethanol + SA Amph Water + SA Amph
Amphetamine challenge
Amphetamine challenge
Amphetamine challenge

55
(Gilpin et al., 2012; Siegmund et al., 2005; Slawecki and Betancourt, 2002; Vetter et al., 2007). Surprisingly, little work has investigated how adolescent ethanol exposure influences intake of drugs other than ethanol. With regard to the growing evidence that adolescent ethanol exposure affects dopamine transmission, drugs that specifically target this system are of interest. Etha-nol exposure during early/mid adolescence sensitizes the rewarding effects of cocaine and attenuates the aversive effects; it also altered gross locomotor activity (Hutchinson and Riley, 2012; Hutchison et al., 2010). However, adolescent ethanol drinking does not alter cocaine self-administration (0.5 and 0.75 mg/kg) in mice on a FR1 schedule of reinforcement (Esteve-Arenys et al., 2017).
As mentioned in “Neuronal circuitries in the addiction cycle”, the dorsal striatum is involved in habitual drug consumption and transition to compul-sive drug use (Everitt and Robbins, 2013). On the basis of the results from the chronoamperometric recordings (see Adolescent ethanol exposure: Re-sidual effects on in vivo dopamine), we hypothesized that there might be a difference in the rats’ motivational drive to consume amphetamine. Howev-er, there were no differences between the water- and ethanol-exposed rats in the PR trials (Figure 18A). A dose-response function was used to explore self-administration behavior on a FR schedule, but the groups displayed similar intake behavior at all doses tested (Figure 18B).
Figure 18. Amphetamine self-administration. A) Progressive ratio trial. Breakpoint values and corresponding response requirements for the last earned reward. The bars represent mean + SEM. B) Dose-response function. The values represent mean (± SEM) number of amphetamine infusions earned on a fixed-ratio three schedule across different unit doses.
Hence, differences in peak area—driven by reduced release of dopamine after amphetamine challenge in the ethanol-exposed group—had no influ-ence on drug-taking behavior during the relatively short period of self-administration tested herein. A different setup with prolonged self-
Water Ethanol
0.0 0.025 0.05 0.1mg/kg/infusion
0
5
10
15
20
25
30
Infu
sion
s
mg/kg/infusion
0
2
4
6
8
10
12
14
16
WaterEthanol
Bre
akpo
int
0.1 0.05
2
6
12
20
32
50
77
118
Res
pons
e re
quire
men
t
A B

56
administration or a shift toward lower doses could possibly reveal more sub-tle alterations in self-administration behavior.
Subsequent amphetamine self-administration and in vivo dopamine In vivo dopamine dynamics were analyzed in the dorsal striatum two weeks after the last amphetamine self-administration session in rats with and with-out a history of adolescent ethanol exposure. The response to this ampheta-mine challenge was dependent on adolescent history. Ethanol-exposed rats did not increase their peak area after the amphetamine challenge, but the water-exposed animals had an increased peak area at 5, 15, 35 and 45 min. The reduced response was driven by an abolished effect on T80 (Figure 17D) and not due to differences in peak amplitude (Figure 17C) indicative of differences in dopamine transporter function. Even though no differences were noted in amphetamine self-administration behavior, the response to the drug in the brain was altered in ethanol-exposed animals. These results indi-cate a potentially synergistic effect on dopaminergic response to ampheta-mine after adolescent ethanol and adult amphetamine exposure. It would have been interesting to investigate how these rats responded if reintroduced to amphetamine self-administration to for example, another PR trial, lower unit doses of amphetamine or extended repeated periods of self-administration.

57
Conclusions
The present thesis provides further evidence that early life stress affects the endogenous opioid system. The expression of four opioid genes—Oprd1, Oprk1, Oprm1, in the dorsal striatum and Pomc in the amygdala—is affected by 360 minutes of maternal separation. Interestingly, changes in gene ex-pression after voluntary ethanol intake depend on early life conditions.
Levels of endogenous opioid peptides are also affected by single housing in early adolescence. A short period of single housing activates the HPA axis and increases N/OFQ in the amygdala, hypothalamus and medial prefrontal cortex. Prolonged single housing causes lower basal levels of MEAP in the amygdala, substantia nigra, periaqueductal gray, hippocampus, hypothala-mus and in frontal- and medial prefrontal cortex. Shorter periods of single housing are common during behavioral measurements, pharmacological treatments, or when terminating an experiment. When measuring individual consumption behavior such as ethanol intake, the animals are often single-housed for longer periods. The knowledge of how endogenous opioid pep-tides are affected by housing conditions has to be taken into consideration when designing an experiment; otherwise, the interpretation of the results would be incorrect.
Repeated ethanol binges during adolescence also affect the endogenous opioid system. The levels of MEAP and β-endorphin in the nucleus accum-bens change during intoxication and residual effects of ethanol exposure are found in MEAP levels in the amygdala, substantia nigra and VTA. The amygdala is involved in the regulation of stress and anxiety, while the sub-stantia nigra and VTA are important for dopaminergic projections in the reward circuitry, which is regulated by endogenous opioids. Furthermore, adolescent ethanol exposure causes alterations in dopamine dynamics in the dorsal striatum; a single amphetamine challenge reduces dopamine release although the reuptake of dopamine remains unaffected. Ethanol-exposed rats and controls display similar amphetamine self-administration behavior, but our analysis shows a more efficient striatal removal of dopamine in ethanol-exposed rats.
It has been postulated that changes in the brain stress system and in do-paminergic activity increase susceptibility for alcohol dependence. The al-terations in endogenous opioids and dopamine dynamics presented in this thesis could partly explain why stress early in life and exposure to ethanol in adolescence increases the risk of elevated alcohol consumption later in life.

58
Populärvetenskaplig sammanfattning
Från att du föds, genom hela din ungdomstid och fram till att du befinner dig i mitten av 20-årsåldern så utvecklas och mognar din hjärna. Under den här tiden är hjärnan även extra känslig för olika former av yttre påverkan. Två exempel på faktorer som kan ha negativ effekt på hjärnan är stress och expo-nering av droger. Om du växer upp i en otrygg och stressfylld miljö eller utsätts för upprepad exponering av droger i ung ålder så ökar exempelvis risken att du drabbas av en psykiatrisk sjukdom senare i livet.
Den här avhandlingen har undersökt hur två av hjärnans signalsubstanser, dopamin och endogena opioider (kroppsegna morfinliknande ämnen), på-verkas när en ung individ utsätts för stress eller upprepat intag av alkohol. Dopamin och endogena opioider utgör en viktig del av hjärnans belönings-system och hjälper till att förmedla de positiva känslor som du upplever när du till exempel äter en god middag, har sex eller uppnår en framgång. Dessa positiva känslor kallas för naturlig belöning. Men det är inte bara vid naturlig belöning som aktiviteten av dopamin och endogena opioider ökar, även alla beroendeframkallande droger har den här egenskapen.
För att undersöka hur den unga hjärnan påverkas av droger eller stress an-vänds ofta djurmodeller. Anledningen är att dessa frågeställningar är svåra att besvara i människa på grund av etiska principer. I den här avhandlingen har därför djurmodeller använts för att simulera olika typer av negativa livs-händelser som förknippas med en otrygg uppväxtmiljö. I en av avhandling-ens studier stördes kontakten mellan mamman och ungarna genom tidsbe-gränsade separationer av nyfödda råttungar och i en annan av studierna pla-cerades unga råttor tidvis ensamma för att efterlikna en miljö med störd social kontakt. Alkohol gavs även till unga råttor för att undersöka hur alko-hol påverkar den unga hjärnan.
Resultaten från studierna visade bland annat att en otrygg uppväxtmiljö påverkade endogena opioider i dorsala striatum - en del av hjärnan som bland annat är kopplad till bildandet av vanor. En otrygg uppväxt förändrade även hur de endogena opioiderna svarade på alkoholintag i vuxen ålder. De endogena opioiderna påverkades även av en störd social miljö under ung-domstiden då minskade nivåer sågs i många av hjärnans olika regioner. Kvarvarande förändringar i både endogena opioider och dopamin syntes efter att alkohol intagits under ungdomstiden. Intaget av alkohol i ung ålder hade ingen effekt på viljan att självadministrera amfetamin, däremot skiljde

59
sig dopaminaktiviteten åt i dorsala striatum efter en injektion av amfetamin i vuxen ålder.
Otrygghet, stress och alkohol formar med andra ord den unga hjärnan och förändrar dess belöningssystem. Detta kan, till viss del, förklara varför en otrygg uppväxtmiljö och upprepat intag av droger i ung ålder ökar risken för psykiatrisk sjukdom senare i livet.

60
Acknowledgment
Now this period of my life is finished, without my main supervisor Ingrid Nylander it would not have been possible. Thank you for believing in me! To know that you have trust in what I do creates a stimulating environment to work in. Even if you have tons of work to do, you always try to find some time to help. I´m very happy to have had you as my main supervisor! My co-supervisors. Anne-Lie Svensson, you are the gate-way to a PhD. Without your fantastic supervision during SOFOSKO projects, many theses would not have been written. You are one of the most positive persons I know! Maria Ellgren, I have missed you a lot since you left the academia, you were a wonderful support when you worked here. Erika Roman, for all the knowledge that you willingly share about animals, without that, this the-sis would be full of caveats. To all the members of the NAB-group, thanks for being great colleagues. Lova, you are a fantastic person, who does so much for others. I don’t know a single person in the department who haven´t been helped by you. You de-serve the best. Stina and Nikita, it was good to have some familiar PhD-faces when coming back to the lab, keep up the good work. Victoria and Lena, for help and support with teaching and students. Åsa, for being an energy injec-tion! Marita, for all the help with animal work. Carin, for taking care of the everyday maintenance of lab stuff. Sam, a lot of fun left the lab when you headed back to England, you ́re missed! Sara and Loudin, thanks for intro-ducing and welcoming me as a new PhD student and for all the travelling to conferences! Shima, for being my light and laughter in the office for the first years. The distance between Norrland and Middle East has never been so small. To all the students who has helped me in my projects. The Comasco-group, Erika, Aniruddah, Megha and Maria, for your hard work and fruitful discussions! The staff in department of Pharmaceutical Biosciences for helping out with everyday problems. Especially to all PhD students, the past ones (Alfhild, Anna, Anneli, Erika, Helen, Inga, Jenny, Jonathan, Mats, Oskar, Patrik) for showing it is possible and to the present ones (Lisa (You go!), Erik, Sofia)

61
for doing it. Lena (O ́Raili) for being a reliable source of knowledge, the solid rocks in the department. One “Blaj” a day keeps the doctor away! Shima, thanks for introducing me to the fantastic group of friends you have. That you having fantastic friends is off course no surprise! Emelie, you are wise, fun and helpful, what else can one ask for? Srebrenka, why didn’t we hang out more during the phar-macy program? I’m glad we had a second chance, I love the way you ap-proach life! Sahar, the sweetest and toughest person I know. Sofie, the calm-ness and the (positive) nerdiness of your personality are a perfect match. Maria, your ability of being both superfun an superserious is amazing! Jenny, Alexandra and Mikka, we are having a 12 (!) years anniversary this year. I hope our 2017-babies will share the same friendship. To all the babies of the next generation (Alma, Abbe, Astrid, Evy, Iris, Greta and Sigge) that decided to be born around the time I was on maternal leave. I had so many great parents to hang out with because of you. I hope I can fol-low you through your lives. An extra thank to Abbe, it has been great to have your mom (Elin) to run beside in the last miles of the PhD-marathon. To my family for being the foundation of my wellbeing, you are the best! I could not have asked for a better environment to grew up in, I really hope I can pass it forward. My PhD period would not have gone this smoothly without being raised of the master of organization and the lifestyle without procrastination, i.e., my mom. I have also tried to apply the calmness that my dad always brings to any situation. To know that the two of you always have my back makes it easier to go through life. My sister Erika for always (or at least before kids came into the picture…) having time to talk and analyzing things and for being inspiring in the way that you really loves your job, if every teacher were as you are there would not be any crisis in the Swedish school system! Per, you have been in our family for such a long time that I almost cannot recall the time before, I´m happy you are a part of “the Granholms”. Bastian, the families little whirlwind, your energy will take you far! My grandmother, the most inspiring 92-year old I know, if I will be half as you are in your age I will be happy. I love you all! My own little family that I love so much. Tobias, for always letting me be me, for spoiling me with great food every day, letting me whine as much as I want, for always remembering me of the funny things in life and making me forget work as soon as I come home. Ebbe, my funniest person in the world, for you being you is the best thing I know. To see you grow up is a privilege, but please can we slow down the time for a while now!

62
References
Aalto S, Ingman K, Alakurtti K, Kaasinen V, Virkkala J, Nagren K, et al. (2015). Intravenous ethanol increases dopamine release in the ventral striatum in humans: PET study using bolus-plus-infusion administration of [(11)C]raclopride. J Cereb Blood Flow Metab 35(3): 424-431.
Abernathy K, Chandler LJ, Woodward JJ (2010). Alcohol and the prefrontal cortex. Int Rev Neurobiol 91: 289-320.
Ahlenius S, Carlsson A, Engel J, Svensson T, Sodersten P (1973). Antagonism by alpha methyltyrosine of the ethanol-induced stimulation and euphoria in man. Clin Pharmacol Ther 14(4): 586-591.
Alaux-Cantin S, Warnault V, Legastelois R, Botia B, Pierrefiche O, Vilpoux C, et al. (2013). Alcohol intoxications during adolescence increase motivation for alcohol in adult rats and induce neuroadaptations in the nucleus accumbens. Neuropharmacology 67: 521-531.
Allen RG, Peng B, Pellegrino MJ, Miller ED, Grandy DK, Lundblad JR, et al. (2001). Altered processing of pro-orphanin FQ/nociceptin and pro-opiomelanocortin-derived peptides in the brains of mice expressing defective prohormone convertase 2. J Neurosci 21(16): 5864-5870.
Amodeo LR, Kneiber D, Wills DN, Ehlers CL (2017). Alcohol drinking during adolescence increases consumptive responses to alcohol in adulthood in Wistar rats. Alcohol 59: 43-51.
Atwood BK, Kupferschmidt DA, Lovinger DM (2014). Opioids induce dissociable forms of long-term depression of excitatory inputs to the dorsal striatum. Nat Neurosci 17(4): 540-548.
Avegno EM, Salling MC, Borgkvist A, Mrejeru A, Whitebirch AC, Margolis EB, et al. (2016). Voluntary adolescent drinking enhances excitation by low levels of alcohol in a subset of dopaminergic neurons in the ventral tegmental area. Neuropharmacology 110(Pt A): 386-395.
Aziz AM, Brothers S, Sartor G, Holm L, Heilig M, Wahlestedt C, et al. (2016). The nociceptin/orphanin FQ receptor agonist SR-8993 as a candidate therapeutic for alcohol use disorders: validation in rat models. Psychopharmacology (Berl) 233(19-20): 3553-3563.
Badanich KA, Maldonado AM, Kirstein CL (2007). Chronic ethanol exposure during adolescence increases basal dopamine in the nucleus accumbens septi during adulthood. Alcohol Clin Exp Res 31(5): 895-900.
Baker M (2012). Digital PCR hits its stride. Nature Methods 9(6): 541-544. Banghart MR, Neufeld SQ, Wong NC, Sabatini BL (2015). Enkephalin Disinhibits
Mu Opioid Receptor-Rich Striatal Patches via Delta Opioid Receptors. Neuron 88(6): 1227-1239.
Bank A, Terada M, Metafora S, Dow L, Marks PA (1972). In vitro synthesis of DNA components of human genes for globins. Nat New Biol 235(58): 167-169.
Barr CS, Schwandt ML, Lindell SG, Higley JD, Maestripieri D, Goldman D, et al. (2008). Variation at the mu-opioid receptor gene (OPRM1) influences

63
attachment behavior in infant primates. Proc Natl Acad Sci U S A 105(13): 5277-5281.
Bart G, Schluger JH, Borg L, Ho A, Bidlack JM, Kreek MJ (2005). Nalmefene induced elevation in serum prolactin in normal human volunteers: partial kappa opioid agonist activity? Neuropsychopharmacology 30(12): 2254-2262.
Bayon A, Shoemaker WJ, Bloom FE, Mauss A, Guillemin R (1979). Perinatal development of the endorphin- and enkephalin-containing systems in the rat brain. Brain Res 179(1): 93-101.
Beatty WW, Costello KB (1982). Naloxone and play fighting in juvenile rats. Pharmacol Biochem Behav 17(5): 905-907.
Beaulieu JM, Gainetdinov RR (2011). The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol Rev 63(1): 182-217.
Becker HC (2013). Animal models of excessive alcohol consumption in rodents. Curr Top Behav Neurosci 13: 355-377.
Belin D, Deroche-Gamonet V (2012). Responses to novelty and vulnerability to cocaine addiction: contribution of a multi-symptomatic animal model. Cold Spring Harb Perspect Med 2(11).
Belin D, Jonkman S, Dickinson A, Robbins TW, Everitt BJ (2009). Parallel and interactive learning processes within the basal ganglia: relevance for the understanding of addiction. Behav Brain Res 199(1): 89-102.
Bell RL, Hauser SR, Liang T, Sari Y, Maldonado-Devincci A, Rodd ZA (2017). Rat animal models for screening medications to treat alcohol use disorders. Neuropharmacology 122: 201-243.
Berdoy M (2003). The Laboratory Rat: A Natural History: http://www.ratlife.com.
Berson SA, Yalow RS (1968). General principles of radioimmunoassay. Clin Chim Acta 22(1): 51-69.
Berube P, Laforest S, Bhatnagar S, Drolet G (2013). Enkephalin and dynorphin mRNA expression are associated with resilience or vulnerability to chronic social defeat stress. Physiol Behav 122: 237-245.
Berube P, Poulin JF, Laforest S, Drolet G (2014). Enkephalin knockdown in the basolateral amygdala reproduces vulnerable anxiety-like responses to chronic unpredictable stress. Neuropsychopharmacology 39(5): 1159-1168.
Björklund A, Dunnett SB (2007). Dopamine neuron systems in the brain: an update. Trends Neurosci 30(5): 194-202.
Blanchard BA, Steindorf S, Wang S, Glick SD (1993). Sex differences in ethanol-induced dopamine release in nucleus accumbens and in ethanol consumption in rats. Alcohol Clin Exp Res 17(5): 968-973.
Boileau I, Assaad JM, Pihl RO, Benkelfat C, Leyton M, Diksic M, et al. (2003). Alcohol promotes dopamine release in the human nucleus accumbens. Synapse 49(4): 226-231.
Bridges RS, Grimm CT (1982). Reversal of morphine disruption of maternal behavior by concurrent treatment with the opiate antagonist naloxone. Science 218(4568): 166-168.
Brunson KL, Khan N, Eghbal-Ahmadi M, Baram TZ (2001). Corticotropin (ACTH) acts directly on amygdala neurons to down-regulate corticotropin-releasing hormone gene expression. Ann Neurol 49(3): 304-312.
Bunzow JR, Saez C, Mortrud M, Bouvier C, Williams JT, Low M, et al. (1994). Molecular cloning and tissue distribution of a putative member of the rat opioid receptor gene family that is not a mu, delta or kappa opioid receptor type. FEBS Lett 347(2-3): 284-288.

64
Calcagnetti DJ, Schechter MD (1992). Place conditioning reveals the rewarding aspect of social interaction in juvenile rats. Physiol Behav 51(4): 667-672.
CAN (2017). Skolelevers drogvanor 2017 - CAN rapport 170. Stockholm: Centralförbundet för alkohol- och narkotikaupplysning (CAN).
Cao L, Cui X, Hu J, Li Z, Choi JR, Yang Q, et al. (2017). Advances in digital polymerase chain reaction (dPCR) and its emerging biomedical applications. Biosens Bioelectron 90: 459-474.
Carden SE, Hofer MA (1990a). Independence of benzodiazepine and opiate action in the suppression of isolation distress in rat pups. Behav Neurosci 104(1): 160-166.
Carden SE, Hofer MA (1990b). Socially mediated reduction of isolation distress in rat pups is blocked by naltrexone but not by Ro 15-1788. Behav Neurosci 104(3): 457-463.
Carlsson A, Lindqvist M, Magnusson T (1957). 3,4-Dihydroxyphenylalanine and 5-hydroxytryptophan as reserpine antagonists. Nature 180(4596): 1200.
Carr CP, Martins CMS, Stingel AM, Lemgruber VB, Juruena MF (2013). The Role of Early Life Stress in Adult Psychiatric Disorders A Systematic Review According to Childhood Trauma Subtypes. Journal of Nervous and Mental Disease 201(12): 1007-1020.
Christensson-Nylander I, Herrera-Marschitz M, Staines W, Hokfelt T, Terenius L, Ungerstedt U, et al. (1986). Striato-nigral dynorphin and substance P pathways in the rat. I. Biochemical and immunohistochemical studies. Exp Brain Res 64(1): 169-192.
Christensson-Nylander I, Nyberg F, Ragnarsson U, Terenius L (1985). A general procedure for analysis of proenkephalin B derived opioid peptides. Regul Pept 11(1): 65-76.
Chu Sin Chung P, Keyworth HL, Martin-Garcia E, Charbogne P, Darcq E, Bailey A, et al. (2015). A novel anxiogenic role for the delta opioid receptor expressed in GABAergic forebrain neurons. Biol Psychiatry 77(4): 404-415.
Chu Sin Chung P, Kieffer BL (2013). Delta opioid receptors in brain function and diseases. Pharmacol Ther 140(1): 112-120.
Ciccocioppo R, de Guglielmo G, Hansson AC, Ubaldi M, Kallupi M, Cruz MT, et al. (2014). Restraint stress alters nociceptin/orphanin FQ and CRF systems in the rat central amygdala: significance for anxiety-like behaviors. J Neurosci 34(2): 363-372.
Ciccocioppo R, Economidou D, Fedeli A, Angeletti S, Weiss F, Heilig M, et al. (2004). Attenuation of ethanol self-administration and of conditioned reinstatement of alcohol-seeking behaviour by the antiopioid peptide nociceptin/orphanin FQ in alcohol-preferring rats. Psychopharmacology (Berl) 172(2): 170-178.
Ciccocioppo R, Martin-Fardon R, Weiss F (2002). Effect of selective blockade of mu(1) or delta opioid receptors on reinstatement of alcohol-seeking behavior by drug-associated stimuli in rats. Neuropsychopharmacology 27(3): 391-399.
Ciccocioppo R, Panocka I, Polidori C, Regoli D, Massi M (1999). Effect of nociceptin on alcohol intake in alcohol-preferring rats. Psychopharmacology (Berl) 141(2): 220-224.
Copeland WE, Sun H, Costello EJ, Angold A, Heilig MA, Barr CS (2011). Child mu-opioid receptor gene variant influences parent-child relations. Neuropsychopharmacology 36(6): 1165-1170.
Corbit LH, Nie H, Janak PH (2012). Habitual alcohol seeking: time course and the contribution of subregions of the dorsal striatum. Biol Psychiatry 72(5): 389-395.

65
Cox BM, Goldstein A, Hi CH (1976). Opioid activity of a peptide, beta-lipotropin-(61-91), derived from beta-lipotropin. Proc Natl Acad Sci U S A 73(6): 1821-1823.
Crabbe JC, Harris RA, Koob GF (2011). Preclinical studies of alcohol binge drinking. Ann N Y Acad Sci 1216: 24-40.
Criado JR, Ehlers CL (2013). Effects of adolescent onset voluntary drinking followed by ethanol vapor exposure on subsequent ethanol consumption during protracted withdrawal in adult Wistar rats. Pharmacol Biochem Behav 103(3): 622-630.
Cruz MT, Herman MA, Kallupi M, Roberto M (2012). Nociceptin/orphanin FQ blockade of corticotropin-releasing factor-induced gamma-aminobutyric acid release in central amygdala is enhanced after chronic ethanol exposure. Biol Psychiatry 71(8): 666-676.
Curley JP, Champagne FA (2016). Influence of maternal care on the developing brain: Mechanisms, temporal dynamics and sensitive periods. Front Neuroendocrinol 40: 52-66.
Dahlstrom A, Fuxe K (1964). Localization of monoamines in the lower brain stem. Experientia 20(7): 398-399.
Daoura L, Nylander I (2011). The response to naltrexone in ethanol-drinking rats depends on early environmental experiences. Pharmacol Biochem Behav 99(4): 626-633.
de Waele JP, Gianoulakis C (1993). Effects of single and repeated exposures to ethanol on hypothalamic beta-endorphin and CRH release by the C57BL/6 and DBA/2 strains of mice. Neuroendocrinology 57(4): 700-709.
Dehaene-Lambertz G, Spelke ES (2015). The Infancy of the Human Brain. Neuron 88(1): 93-109.
Devine DP, Leone P, Pocock D, Wise RA (1993). Differential involvement of ventral tegmental mu, delta and kappa opioid receptors in modulation of basal mesolimbic dopamine release: in vivo microdialysis studies. J Pharmacol Exp Ther 266(3): 1236-1246.
Devine DP, Wise RA (1994). Self-administration of morphine, DAMGO, and DPDPE into the ventral tegmental area of rats. J Neurosci 14(4): 1978-1984.
DeWit DJ, Adlaf EM, Offord DR, Ogborne AC (2000). Age at first alcohol use: a risk factor for the development of alcohol disorders. Am J Psychiatry 157(5): 745-750.
Di Chiara G, Bassareo V, Fenu S, De Luca MA, Spina L, Cadoni C, et al. (2004). Dopamine and drug addiction: the nucleus accumbens shell connection. Neuropharmacology 47 Suppl 1: 227-241.
Di Chiara G, Imperato A (1988). Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci U S A 85(14): 5274-5278.
Di Chiara G, Imperato A (1985). Ethanol preferentially stimulates dopamine release in the nucleus accumbens of freely moving rats. Eur J Pharmacol 115(1): 131-132.
Ding ZM, Ingraham CM, Rodd ZA, McBride WJ (2015). The reinforcing effects of ethanol within the nucleus accumbens shell involve activation of local GABA and serotonin receptors. J Psychopharmacol 29(6): 725-733.
Douglas LA, Varlinskaya EI, Spear LP (2004). Rewarding properties of social interactions in adolescent and adult male and female rats: impact of social versus isolate housing of subjects and partners. Dev Psychobiol 45(3): 153-162.

66
Dunbar RI (2010). The social role of touch in humans and primates: behavioural function and neurobiological mechanisms. Neurosci Biobehav Rev 34(2): 260-268.
Duzzioni M, Duarte FS, Leme LR, Gavioli EC, De Lima TC (2011). Anxiolytic-like effect of central administration of NOP receptor antagonist UFP-101 in rats submitted to the elevated T-maze. Behav Brain Res 222(1): 206-211.
Economidou D, Cippitelli A, Stopponi S, Braconi S, Clementi S, Ubaldi M, et al. (2011). Activation of brain NOP receptors attenuates acute and protracted alcohol withdrawal symptoms in the rat. Alcoholism-Clinical and Experimental Research 35(4): 747-755.
Emmerson PJ, Liu MR, Woods JH, Medzihradsky F (1994). Binding affinity and selectivity of opioids at mu, delta and kappa receptors in monkey brain membranes. J Pharmacol Exp Ther 271(3): 1630-1637.
Engel J, Strombom U, Svensson TH, Waldeck B (1974). Suppression by alpha-methyltyrosine of ethanol-induced locomotor stimulation: partial reversal by L-dopa. Psychopharmacologia 37(3): 275-279.
Engel JA, Jerlhag E (2014). Alcohol: mechanisms along the mesolimbic dopamine system. Dopamine 211: 201-233.
Engleman EA, Ding ZM, Oster SM, Toalston JE, Bell RL, Murphy JM, et al. (2009). Ethanol is self-administered into the nucleus accumbens shell, but not the core: evidence of genetic sensitivity. Alcoholism-Clinical and Experimental Research 33(12): 2162-2171.
Esteve-Arenys A, Gracia-Rubio I, Cantacorps L, Pozo OJ, Marcos J, Rodriguez-Arias M, et al. (2017). Binge ethanol drinking during adolescence modifies cocaine responses in mice. J Psychopharmacol 31(1): 86-95.
Everitt BJ, Robbins TW (2016). Drug Addiction: Updating Actions to Habits to Compulsions Ten Years On. Annu Rev Psychol 67: 23-50.
Everitt BJ, Robbins TW (2013). From the ventral to the dorsal striatum: Devolving views of their roles in drug addiction. Neuroscience and Biobehavioral Reviews 37(9): 1946-1954.
Fahlke C, Hansen S, Engel JA, Hard E (1994). Effects of ventral striatal 6-OHDA lesions or amphetamine sensitization on ethanol consumption in the rat. Pharmacol Biochem Behav 47(2): 345-349.
Felicio LF, Mann PE, Bridges RS (1991). Intracerebroventricular cholecystokinin infusions block beta-endorphin-induced disruption of maternal behavior. Pharmacol Biochem Behav 39(1): 201-204.
Fernandez F, Misilmeri MA, Felger JC, Devine DP (2004). Nociceptin/orphanin FQ increases anxiety-related behavior and circulating levels of corticosterone during neophobic tests of anxiety. Neuropsychopharmacology 29(1): 59-71.
Fields HL, Margolis EB (2015). Understanding opioid reward. Trends Neurosci 38(4): 217-225.
Filliol D, Ghozland S, Chluba J, Martin M, Matthes HW, Simonin F, et al. (2000). Mice deficient for delta- and mu-opioid receptors exhibit opposing alterations of emotional responses. Nat Genet 25(2): 195-200.
Fone KC, Porkess MV (2008). Behavioural and neurochemical effects of post-weaning social isolation in rodents-relevance to developmental neuropsychiatric disorders. Neurosci Biobehav Rev 32(6): 1087-1102.
Franck J, Lindholm S, Raaschou P (1998). Modulation of volitional ethanol intake in the rat by central delta-opioid receptors. Alcoholism-Clinical and Experimental Research 22(6): 1185-1189.
Franklin RE, Gosling RG (1953). Molecular configuration in sodium thymonucleate. Nature 171(4356): 740-741.

67
Garau L, Govoni S, Stefanini E, Trabucchi M, Spano PF (1978). Dopamine receptors: pharmacological and anatomical evidences indicate that two distinct dopamine receptor populations are present in rat striatum. Life Sci 23(17-18): 1745-1750.
Gavioli EC, Marzola G, Guerrini R, Bertorelli R, Zucchini S, De Lima TC, et al. (2003). Blockade of nociceptin/orphanin FQ-NOP receptor signalling produces antidepressant-like effects: pharmacological and genetic evidences from the mouse forced swimming test. Eur J Neurosci 17(9): 1987-1990.
Gerhardt GA, Burmeister JJ (2000). Voltammetry In Vivo for Chemical Analysis of the Nervous System. In: A. MR (ed)^(eds). Encyclopedia of Analytical Chemistry, edn. Chichester: John Wiley & Sons Ltd. p^pp.
Gerhardt GA, Hoffman AF (2001). Effects of recording media composition on the responses of Nafion-coated carbon fiber microelectrodes measured using high-speed chronoamperometry. J Neurosci Methods 109(1): 13-21.
Gianoulakis C (1990). Characterization of the effects of acute ethanol administration on the release of beta-endorphin peptides by the rat hypothalamus. Eur J Pharmacol 180(1): 21-29.
Gilpin NW, Karanikas CA, Richardson HN (2012). Adolescent binge drinking leads to changes in alcohol drinking, anxiety, and amygdalar corticotropin releasing factor cells in adulthood in male rats. PLoS One 7(2): e31466.
Gilpin NW, Koob GF (2008). Neurobiology of alcohol dependence: focus on motivational mechanisms. Alcohol Res Health 31(3): 185-195.
Goldstein A, Fischli W, Lowney LI, Hunkapiller M, Hood L (1981). Porcine pituitary dynorphin: complete amino acid sequence of the biologically active heptadecapeptide. Proc Natl Acad Sci U S A 78(11): 7219-7223.
Grandy DK, Zhang YA, Bouvier C, Zhou QY, Johnson RA, Allen L, et al. (1991). Multiple human D5 dopamine receptor genes: a functional receptor and two pseudogenes. Proc Natl Acad Sci U S A 88(20): 9175-9179.
Green MK, Barbieri EV, Brown BD, Chen KW, Devine DP (2007). Roles of the bed nucleus of stria terminalis and of the amygdala in N/OFQ-mediated anxiety and HPA axis activation. Neuropeptides 41(6): 399-410.
Green MK, Devine DP (2009). Nociceptin/orphanin FQ and NOP receptor gene regulation after acute or repeated social defeat stress. Neuropeptides 43(6): 507-514.
Grota LJ, Ader R (1969). Continuous Recording of Maternal Behaviour in Rattus-Norvegicus. Animal Behaviour 17: 722-&.
Guard HJ, Newman JD, Roberts RL (2002). Morphine administration selectively facilitates social play in common marmosets. Dev Psychobiol 41(1): 37-49.
Guerri C, Pascual M (2010). Mechanisms involved in the neurotoxic, cognitive, and neurobehavioral effects of alcohol consumption during adolescence. Alcohol 44(1): 15-26.
Gustafsson L, Oreland S, Hoffmann P, Nylander I (2008). The impact of postnatal environment on opioid peptides in young and adult male Wistar rats. Neuropeptides 42(2): 177-191.
Gustafsson L, Zhou Q, Nylander I (2007). Ethanol-induced effects on opioid peptides in adult male Wistar rats are dependent on early environmental factors. Neuroscience 146(3): 1137-1149.
Hanson KL, Medina KL, Padula CB, Tapert SF, Brown SA (2011). Impact of Adolescent Alcohol and Drug Use on Neuropsychological Functioning in Young Adulthood: 10-Year Outcomes. J Child Adolesc Subst Abuse 20(2): 135-154.

68
Heinrich LM, Gullone E (2006). The clinical significance of loneliness: a literature review. Clin Psychol Rev 26(6): 695-718.
Heinz A, Siessmeier T, Wrase J, Hermann D, Klein S, Grusser SM, et al. (2004). Correlation between dopamine D(2) receptors in the ventral striatum and central processing of alcohol cues and craving. Am J Psychiatry 161(10): 1783-1789.
Henry MS, Gendron L, Tremblay ME, Drolet G (2017). Enkephalins: Endogenous Analgesics with an Emerging Role in Stress Resilience. Neural Plast 2017: 1546125.
Herman BH, Panksepp J (1978). Effects of morphine and naloxone on separation distress and approach attachment: evidence for opiate mediation of social affect. Pharmacol Biochem Behav 9(2): 213-220.
Herrera-Marschitz M, Christensson-Nylander I, Sharp T, Staines W, Reid M, Hokfelt T, et al. (1986). Striato-nigral dynorphin and substance P pathways in the rat. II. Functional analysis. Exp Brain Res 64(1): 193-207.
Heyser CJ, Roberts AJ, Schulteis G, Koob GF (1999). Central administration of an opiate antagonist decreases oral ethanol self-administration in rats. Alcohol Clin Exp Res 23(9): 1468-1476.
Hietala J, West C, Syvalahti E, Nagren K, Lehikoinen P, Sonninen P, et al. (1994). Striatal D2 dopamine receptor binding characteristics in vivo in patients with alcohol dependence. Psychopharmacology (Berl) 116(3): 285-290.
Higham JP, Barr CS, Hoffman CL, Mandalaywala TM, Parker KJ, Maestripieri D (2011). Mu-opioid receptor (OPRM1) variation, oxytocin levels and maternal attachment in free-ranging rhesus macaques Macaca mulatta. Behav Neurosci 125(2): 131-136.
Higuchi R, Fockler C, Dollinger G, Watson R (1993). Kinetic PCR analysis: real-time monitoring of DNA amplification reactions. Biotechnology (N Y) 11(9): 1026-1030.
Hodos W (1961). Progressive ratio as a measure of reward strength. Science 134(3483): 943-944.
Hoffman J, Philpot, R., Kirstein C. (2014). Comparsion of stress response to oral gavage and intraperitoneal injection. In: Research Society on Alcoholism Vol. 38, pp 1A-292A. Bellevue, Washington: Alcoholism: Clinical and Experimental Research.
Howell LL, Kimmel HL (2008). Monoamine transporters and psychostimulant addiction. Biochem Pharmacol 75(1): 196-217.
Hughes J, Smith T, Morgan B, Fothergill L (1975). Purification and properties of enkephalin - the possible endogenous ligand for the morphine receptor. Life Sci 16(12): 1753-1758.
Hutchinson MA, Riley AL (2012). Ethanol exposure during either adolescence or adulthood alters the rewarding effects of cocaine in adult rats. Pharmacology, Biochemistry and Behaviour 101: 458-464.
Hutchison MA, Albaugh DL, Riley AL (2010). Exposure to lcohol during adolescence alters the aversive and locomotor-activating effects of cocaine in adult rats. Pharmacology, Biochemistry and Behaviour 97: 370-376.
Hyytia P, Kiianmaa K (2001). Suppression of ethanol responding by centrally administered CTOP and naltrindole in AA and Wistar rats. Alcoholism-Clinical and Experimental Research 25(1): 25-33.
Hyytia P, Koob GF (1995). GABAA receptor antagonism in the extended amygdala decreases ethanol self-administration in rats. Eur J Pharmacol 283(1-3): 151-159.

69
Ingman K, Kupila J, Hyytia P, Korpi ER (2006). Effects of aripiprazole on alcohol intake in an animal model of high-alcohol drinking. Alcohol Alcohol 41(4): 391-398.
Ingman K, Salvadori S, Lazarus L, Korpi ER, Honkanen A (2003). Selective delta-opioid receptor antagonist N,N(CH3)2-Dmt-Tic-OH does not reduce ethanol intake in alcohol-preferring AA rats. Addict Biol 8(2): 173-179.
Ishmael FT, Stellato C (2008). Principles and applications of polymerase chain reaction: basic science for the practicing physician. Ann Allergy Asthma Immunol 101(4): 437-443.
Janak PH, Tye KM (2015). From circuits to behaviour in the amygdala. Nature 517: 284.
Jarjour S, Bai L, Gianoulakis C (2009). Effect of acute ethanol administration on the release of opioid peptides from the midbrain including the ventral tegmental area. Alcoholism-Clinical and Experimental Research 33(6): 1033-1043.
Jenck F, Wichmann J, Dautzenberg FM, Moreau JL, Ouagazzal AM, Martin JR, et al. (2000). A synthetic agonist at the orphanin FQ/nociceptin receptor ORL1: anxiolytic profile in the rat. Proc Natl Acad Sci U S A 97(9): 4938-4943.
Jentsch JD, Ashenhurst JR, Cervantes MC, Groman SM, James AS, Pennington ZT (2014). Dissecting impulsivity and its relationships to drug addictions. Ann N Y Acad Sci 1327: 1-26.
June HL, Gilpin NW (2010). Operant self-administration models for testing the neuropharmacological basis of ethanol consumption in rats. Curr Protoc Neurosci Chapter 9: Unit 9 12 11-26.
Kalin NH, Shelton SE, Barksdale CM (1988). Opiate modulation of separation-induced distress in non-human primates. Brain Res 440(2): 285-292.
Kalinichev M, Easterling KW, Holtzman SG (2001). Repeated neonatal maternal separation alters morphine-induced antinociception in male rats. Brain Res Bull 54(6): 649-654.
Keverne EB, Martensz ND, Tuite B (1989). Beta-endorphin concentrations in cerebrospinal fluid of monkeys are influenced by grooming relationships. Psychoneuroendocrinology 14(1-2): 155-161.
Knop J, Joels M, van der Veen R (2017). The added value of rodent models in studying parental influence on offspring development: opportunities, limitations and future perspectives. Curr Opin Psychol 15: 174-181.
Konig M, Zimmer AM, Steiner H, Holmes PV, Crawley JN, Brownstein MJ, et al. (1996). Pain responses, anxiety and aggression in mice deficient in pre-proenkephalin. Nature 383(6600): 535-538.
Koob GF (2009). Brain stress systems in the amygdala and addiction. Brain Res 1293: 61-75.
Koob GF (2008). A role for brain stress systems in addiction. Neuron 59(1): 11-34. Koob GF, Volkow ND (2016). Neurobiology of addiction: a neurocircuitry analysis.
Lancet Psychiatry 3(8): 760-773. Koob GF, Volkow ND (2010). Neurocircuitry of addiction.
Neuropsychopharmacology 35(1): 217-238. Kornblum HI, Hurlbut DE, Leslie FM (1987). Postnatal development of multiple
opioid receptors in rat brain. Brain Res 465(1-2): 21-41. Kosten TR, George TP, Kosten TA (2002). The potential of dopamine agonists in
drug addiction. Expert Opin Investig Drugs 11(4): 491-499. Koster A, Montkowski A, Schulz S, Stube EM, Knaudt K, Jenck F, et al. (1999).
Targeted disruption of the orphanin FQ/nociceptin gene increases stress susceptibility and impairs stress adaptation in mice. Proc Natl Acad Sci U S A 96(18): 10444-10449.

70
Krishnan-Sarin S, Jing SL, Kurtz DL, Zweifel M, Portoghese PS, Li TK, et al. (1995). The delta opioid receptor antagonist naltrindole attenuates both alcohol and saccharin intake in rats selectively bred for alcohol preference. Psychopharmacology (Berl) 120(2): 177-185.
Lam MP, Gianoulakis C (2011). Effects of acute ethanol on corticotropin-releasing hormone and beta-endorphin systems at the level of the rat central amygdala. Psychopharmacology (Berl) 218(1): 229-239.
Lam MP, Marinelli PW, Bai L, Gianoulakis C (2008). Effects of acute ethanol on opioid peptide release in the central amygdala: an in vivo microdialysis study. Psychopharmacology (Berl) 201(2): 261-271.
Lam MP, Nurmi H, Rouvinen N, Kiianmaa K, Gianoulakis C (2010). Effects of acute ethanol on beta-endorphin release in the nucleus accumbens of selectively bred lines of alcohol-preferring AA and alcohol-avoiding ANA rats. Psychopharmacology (Berl) 208(1): 121-130.
Le Merrer J, Becker JA, Befort K, Kieffer BL (2009). Reward processing by the opioid system in the brain. Physiol Rev 89(4): 1379-1412.
Leggett JD, Harbuz MS, Jessop DS, Fulford AJ (2006). The nociceptin receptor antagonist [Nphe1,Arg14,Lys15]nociceptin/orphanin FQ-NH2 blocks the stimulatory effects of nociceptin/orphanin FQ on the HPA axis in rats. Neuroscience 141(4): 2051-2057.
Lerma-Cabrera JM, Carvajal F, Alcaraz-Iborra M, de la Fuente L, Navarro M, Thiele TE, et al. (2013). Adolescent binge-like ethanol exposure reduces basal alpha-MSH expression in the hypothalamus and the amygdala of adult rats. Pharmacol Biochem Behav 110: 66-74.
Littrell OM, Pomerleau F, Huettl P, Surgener S, McGinty JF, Middaugh LD, et al. (2012). Enhanced dopamine transporter activity in middle-aged Gdnf heterozygous mice. Neurobiol Aging 33(2): 427 e421-414.
Lutz PE, Kieffer BL (2013). Opioid receptors: distinct roles in mood disorders. Trends Neurosci 36(3): 195-206.
Lyness WH, Smith FL (1992). Influence of dopaminergic and serotonergic neurons on intravenous ethanol self-administration in the rat. Pharmacol Biochem Behav 42(1): 187-192.
Lyons DM, Parker KJ, Schatzberg AF (2010). Animal models of early life stress: Implications for understanding resilience. Dev Psychobiol 52(5): 402-410.
Manduca A, Lassalle O, Sepers M, Campolongo P, Cuomo V, Marsicano G, et al. (2016). Interacting Cannabinoid and Opioid Receptors in the Nucleus Accumbens Core Control Adolescent Social Play. Front Behav Neurosci 10: 211.
Mansour A, Khachaturian H, Lewis ME, Akil H, Watson SJ (1987). Autoradiographic differentiation of mu, delta, and kappa opioid receptors in the rat forebrain and midbrain. J Neurosci 7(8): 2445-2464.
Margolis EB, Fields HL, Hjelmstad GO, Mitchell JM (2008). Delta-opioid receptor expression in the ventral tegmental area protects against elevated alcohol consumption. J Neurosci 28(48): 12672-12681.
Marinelli PW, Lam M, Bai L, Quirion R, Gianoulakis C (2006). A microdialysis profile of dynorphin A(1-8) release in the rat nucleus accumbens following alcohol administration. Alcoholism-Clinical and Experimental Research 30(6): 982-990.
Marinelli PW, Quirion R, Gianoulakis C (2004). An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following exposure to stress or alcohol. Neuroscience 127(3): 777-784.

71
Marinelli PW, Quirion R, Gianoulakis C (2003). A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcohol administration. Psychopharmacology (Berl) 169(1): 60-67.
Martel FL, Nevison CM, Rayment FD, Simpson MJ, Keverne EB (1993). Opioid receptor blockade reduces maternal affect and social grooming in rhesus monkeys. Psychoneuroendocrinology 18(4): 307-321.
Martel FL, Nevison CM, Simpson MJ, Keverne EB (1995). Effects of opioid receptor blockade on the social behavior of rhesus monkeys living in large family groups. Dev Psychobiol 28(2): 71-84.
Martin WR, Eades CG, Thompson JA, Huppler RE, Gilbert PE (1976). The effects of morphine- and nalorphine- like drugs in the nondependent and morphine-dependent chronic spinal dog. J Pharmacol Exp Ther 197(3): 517-532.
Martin-Fardon R, Ciccocioppo R, Massi M, Weiss F (2000). Nociceptin prevents stress-induced ethanol- but not cocaine-seeking behavior in rats. Neuroreport 11(9): 1939-1943.
Martin-Fardon R, Zorrilla EP, Ciccocioppo R, Weiss F (2010). Role of innate and drug-induced dysregulation of brain stress and arousal systems in addiction: Focus on corticotropin-releasing factor, nociceptin/orphanin FQ, and orexin/hypocretin. Brain Res 1314: 145-161.
McDowell J, Kitchen I (1987). Development of opioid systems: peptides, receptors and pharmacology. Brain Res 434(4): 397-421.
Meiser J, Weindl D, Hiller K (2013). Complexity of dopamine metabolism. Cell Commun Signal 11(1): 34.
Meller RE, Keverne EB, Herbert J (1980). Behavioural and endocrine effects of naltrexone in male talapoin monkeys. Pharmacol Biochem Behav 13(5): 663-672.
Mendez M, Leriche M, Carlos Calva J (2003). Acute ethanol administration transiently decreases [3H]-DAMGO binding to mu opioid receptors in the rat substantia nigra pars reticulata but not in the caudate-putamen. Neurosci Res 47(2): 153-160.
Mendez M, Morales-Mulia M (2006). Ethanol exposure differentially alters pro-enkephalin mRNA expression in regions of the mesocorticolimbic system. Psychopharmacology (Berl) 189(1): 117-124.
Mendez M, Morales-Mulia M, Leriche M (2004). [3H]DPDPE binding to delta opioid receptors in the rat mesocorticolimbic and nigrostriatal pathways is transiently increased by acute ethanol administration. Brain Res 1028(2): 180-190.
Mendez M, Morales-Mulia M, Perez-Luna JM (2008). Ethanol-induced changes in proenkephalin mRNA expression in the rat nigrostriatal pathway. J Mol Neurosci 34(3): 225-234.
Meunier JC, Mollereau C, Toll L, Suaudeau C, Moisand C, Alvinerie P, et al. (1995). Isolation and structure of the endogenous agonist of opioid receptor-like ORL1 receptor. Nature 377(6549): 532-535.
Michael DJ, Wightman RM (1999). Electrochemical monitoring of biogenic amine neurotransmission in real time. J Pharm Biomed Anal 19(1-2): 33-46.
Michel ME, Bolger G, Weissman BA (1985). Binding of a new opiate antagonist, nalmefene, to rat brain membranes. Methods Find Exp Clin Pharmacol 7(4): 175-177.
Miczek KA, Yap JJ, Covington HE, 3rd (2008). Social stress, therapeutics and drug abuse: preclinical models of escalated and depressed intake. Pharmacol Ther 120(2): 102-128.

72
Missale C, Nash SR, Robinson SW, Jaber M, Caron MG (1998). Dopamine receptors: from structure to function. Physiol Rev 78(1): 189-225.
Mitchell JM, O'Neil JP, Janabi M, Marks SM, Jagust WJ, Fields HL (2012). Alcohol consumption induces endogenous opioid release in the human orbitofrontal cortex and nucleus accumbens. Sci Transl Med 4(116): 116ra116.
Moles A, Kieffer BL, D'Amato FR (2004). Deficit in attachment behavior in mice lacking the mu-opioid receptor gene. Science 304(5679): 1983-1986.
Molet J, Maras PM, Avishai-Eliner S, Baram TZ (2014). Naturalistic rodent models of chronic early-life stress. Dev Psychobiol 56(8): 1675-1688.
Moller C, Wiklund L, Sommer W, Thorsell A, Heilig M (1997). Decreased experimental anxiety and voluntary ethanol consumption in rats following central but not basolateral amygdala lesions. Brain Res 760(1-2): 94-101.
Mollereau C, Mouledous L (2000). Tissue distribution of the opioid receptor-like (ORL1) receptor. Peptides 21(7): 907-917.
Mollereau C, Parmentier M, Mailleux P, Butour JL, Moisand C, Chalon P, et al. (1994). ORL1, a novel member of the opioid receptor family. Cloning, functional expression and localization. FEBS Lett 341(1): 33-38.
Mucha RF, Herz A (1985). Motivational properties of kappa and mu opioid receptor agonists studied with place and taste preference conditioning. Psychopharmacology (Berl) 86(3): 274-280.
Mulder AH, Wardeh G, Hogenboom F, Frankhuyzen AL (1984). κ and δ-opioid receptor agonists differentially inhibit striatal dopamine and acetylcholine release. Nature 308: 278.
Mullis KB, Faloona FA (1987). Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol 155: 335-350.
Narendran R, Ciccocioppo R, Lopresti B, Paris J, Himes ML, Mason NS (2017). Nociceptin Receptors in Alcohol Use Disorders: A Positron Emission Tomography Study Using [(11)C]NOP-1A. Biol Psychiatry.
Narita M, Kuzumaki N, Narita M, Kaneko C, Hareyama N, Miyatake M, et al. (2006). Chronic pain-induced emotional dysfunction is associated with astrogliosis due to cortical delta-opioid receptor dysfunction. J Neurochem 97(5): 1369-1378.
Navarro M, Cubero I, Knapp DJ, Breese GR, Thiele TE (2008). Decreased immunoreactivity of the melanocortin neuropeptide alpha-melanocyte-stimulating hormone (alpha-MSH) after chronic ethanol exposure in Sprague-Dawley rats. Alcohol Clin Exp Res 32(2): 266-276.
NIAAA Drinking Levels Defined, NIAAA (ed). http://www.niaaa.nih.gov/alcohol-health/overview-alcohol-consumption/moderate-binge-drinking: National Institute on Alcohol Abuse and Alcoholism.
Nielsen CK, Simms JA, Li R, Mill D, Yi H, Feduccia AA, et al. (2012). delta-opioid receptor function in the dorsal striatum plays a role in high levels of ethanol consumption in rats. J Neurosci 32(13): 4540-4552.
Niesink RJM, Vanree JM (1989). Involvement of Opioid and Dopaminergic Systems in Isolation-Induced Pinning and Social Grooming of Young-Rats. Neuropharmacology 28(4): 411-418.
Nobelprize.org The Nobel Prize in Chemistry 1993: Nobel Media AB. Nobelprize.org The Nobel Prize in Medicine or Physiology 1962: Nobel Media AB. Nobelprize.org The Nobel Prize in Medicine or Physiology 1977: Nobel Media AB. Nobelprize.org The Nobel Prize in Physiology or Medicine 2000: Nobel Media AB.

73
Nummenmaa L, Tuominen L, Dunbar R, Hirvonen J, Manninen S, Arponen E, et al. (2016). Social touch modulates endogenous mu-opioid system activity in humans. Neuroimage 138: 242-247.
Nutt DJ (2014). The role of the opioid system in alcohol dependence. J Psychopharmacol 28(1): 8-22.
Nylander I, Roman E (2013). Is the rodent maternal separation model a valid and effective model for studies on the early-life impact on ethanol consumption? Psychopharmacology (Berl) 229(4): 555-569.
Nylander I, Roman E (2012). Neuropeptides as mediators of the early-life impact on the brain; implications for alcohol use disorders. Front Mol Neurosci 5: 77.
Nylander I, Stenfors C, Tan-No K, Mathe AA, Terenius L (1997). A comparison between microwave irradiation and decapitation: basal levels of dynorphin and enkephalin and the effect of chronic morphine treatment on dynorphin peptides. Neuropeptides 31(4): 357-365.
Nylander I, Vlaskovska M, Terenius L (1995). Brain dynorphin and enkephalin systems in Fischer and Lewis rats: effects of morphine tolerance and withdrawal. Brain Res 683(1): 25-35.
Ogilvie K, Lee S, Rivier C (1997). Effect of three different modes of alcohol administration on the activity of the rat hypothalamic-pituitary-adrenal axis. Alcohol Clin Exp Res 21(3): 467-476.
Oliva JM, Ortiz S, Perez-Rial S, Manzanares J (2008). Time dependent alterations on tyrosine hydroxylase, opioid and cannabinoid CB1 receptor gene expressions after acute ethanol administration in the rat brain. Eur Neuropsychopharmacol 18(5): 373-382.
Olive MF, Koenig HN, Nannini MA, Hodge CW (2001). Stimulation of endorphin neurotransmission in the nucleus accumbens by ethanol, cocaine, and amphetamine. J Neurosci 21(23): RC184.
Palm S Early Environment, Adolescent Alcohol Drinking and Neurobiological Responses to Drugs. PhD, Uppsala University, Acta Universitatis Upsaliensis Uppsala, 2014.
Palm S, Nylander I (2014a). Alcohol-induced changes in opioid Peptide levels in adolescent rats are dependent on housing conditions. Alcoholism-Clinical and Experimental Research 38(12): 2978-2987.
Palm S, Nylander I (2014b). Dopamine release dynamics change during adolescence and after voluntary alcohol intake. PLoS One 9(5): e96337.
Panksepp J (1981). The ontogeny of play in rats. Dev Psychobiol 14(4): 327-332. Panksepp J, Bean NJ, Bishop P, Vilberg T, Sahley TL (1980a). Opioid blockade and
social comfort in chicks. Pharmacol Biochem Behav 13(5): 673-683. Panksepp J, Bishop P (1981). An autoradiographic map of (3H)diprenorphine
binding in rat brain: effects of social interaction. Brain Res Bull 7(4): 405-410. Panksepp J, Herman BH, Vilberg T, Bishop P, DeEskinazi FG (1980b). Endogenous
opioids and social behavior. Neurosci Biobehav Rev 4(4): 473-487. Panksepp J, Jalowiec J, DeEskinazi FG, Bishop P (1985). Opiates and play
dominance in juvenile rats. Behav Neurosci 99(3): 441-453. Pascual M, Boix J, Felipo V, Guerri C (2009). Repeated alcohol administration
during adolescence causes changes in the mesolimbic dopaminergic and glutamatergic systems and promotes alcohol intake in the adult rat. J Neurochem 108(4): 920-931.
Paxinos G, Watson C (1997). The rat brain in stereotaxic coordinates. Compact 3rd edn. Academic Press: San Diego.
Perrine SA, Hoshaw BA, Unterwald EM (2006). Delta opioid receptor ligands modulate anxiety-like behaviors in the rat. Br J Pharmacol 147(8): 864-872.

74
Pert CB, Snyder SH (1973). Opiate receptor: demonstration in nervous tissue. Science 179(4077): 1011-1014.
Philpot RM, Wecker L, Kirstein CL (2009). Repeated ethanol exposure during adolescence alters the developmental trajectory of dopaminergic output from the nucleus accumbens septi. Int J Dev Neurosci 27(8): 805-815.
Ploj K, Roman E, Gustavsson L, Nylander I (2000). Basal levels and alcohol-induced changes in nociceptin/orphanin FQ, dynorphin, and enkephalin levels in C57BL/6J mice. Brain Res Bull 53(2): 219-226.
Ragnauth A, Schuller A, Morgan M, Chan J, Ogawa S, Pintar J, et al. (2001). Female preproenkephalin-knockout mice display altered emotional responses. Proc Natl Acad Sci U S A 98(4): 1958-1963.
Ramchandani VA, Umhau J, Pavon FJ, Ruiz-Velasco V, Margas W, Sun H, et al. (2011). A genetic determinant of the striatal dopamine response to alcohol in men. Mol Psychiatry 16(8): 809-817.
Randall-Thompson JF, Pescatore KA, Unterwald EM (2010). A role for delta opioid receptors in the central nucleus of the amygdala in anxiety-like behaviors. Psychopharmacology (Berl) 212(4): 585-595.
Rassnick S, Stinus L, Koob GF (1993). The effects of 6-hydroxydopamine lesions of the nucleus accumbens and the mesolimbic dopamine system on oral self-administration of ethanol in the rat. Brain Res 623(1): 16-24.
Reinscheid RK, Nothacker HP, Bourson A, Ardati A, Henningsen RA, Bunzow JR, et al. (1995). Orphanin FQ: a neuropeptide that activates an opioidlike G protein-coupled receptor. Science 270(5237): 792-794.
Rhodes JS, Best K, Belknap JK, Finn DA, Crabbe JC (2005). Evaluation of a simple model of ethanol drinking to intoxication in C57BL/6J mice. Physiol Behav 84(1): 53-63.
Richardson NR, Roberts DC (1996). Progressive ratio schedules in drug self-administration studies in rats: a method to evaluate reinforcing efficacy. J Neurosci Methods 66(1): 1-11.
Richter CP, Campbell KH (1940). Alcohol taste thresholds and concentrations of solution preferred by rats. Science 91: 507-508.
Roberts AJ, Gold LH, Polis I, McDonald JS, Filliol D, Kieffer BL, et al. (2001). Increased ethanol self-administration in delta-opioid receptor knockout mice. Alcoholism-Clinical and Experimental Research 25(9): 1249-1256.
Robinson MJ, Warlow SM, Berridge KC (2014). Optogenetic excitation of central amygdala amplifies and narrows incentive motivation to pursue one reward above another. J Neurosci 34(50): 16567-16580.
Saitoh A, Kimura Y, Suzuki T, Kawai K, Nagase H, Kamei J (2004). Potential anxiolytic and antidepressant-like activities of SNC80, a selective delta-opioid agonist, in behavioral models in rodents. J Pharmacol Sci 95(3): 374-380.
Saitoh A, Suzuki S, Soda A, Ohashi M, Yamada M, Oka JI, et al. (2018). The delta opioid receptor agonist KNT-127 in the prelimbic medial prefrontal cortex attenuates veratrine-induced anxiety-like behaviors in mice. Behav Brain Res 336: 77-84.
Saitoh A, Yoshikawa Y, Onodera K, Kamei J (2005). Role of delta-opioid receptor subtypes in anxiety-related behaviors in the elevated plus-maze in rats. Psychopharmacology (Berl) 182(3): 327-334.
Sanchis-Segura C, Spanagel R (2006). Behavioural assessment of drug reinforcement and addictive features in rodents: an overview. Addict Biol 11(1): 2-38.
Schenk S, Britt MD, Atalay J, Charleson S (1982). Isolation rearing decreases opiate receptor binding in rat brain. Pharmacol Biochem Behav 16(5): 841-842.

75
Schindler AG, Soden ME, Zweifel LS, Clark JJ (2016). Reversal of Alcohol-Induced Dysregulation in Dopamine Network Dynamics May Rescue Maladaptive Decision-making. J Neurosci 36(13): 3698-3708.
Schino G, Troisi A (1992). Opiate receptor blockade in juvenile macaques: effect on affiliative interactions with their mothers and group companions. Brain Res 576(1): 125-130.
Schumann G, Loth E, Banaschewski T, Barbot A, Barker G, Buchel C, et al. (2010). The IMAGEN study: reinforcement-related behaviour in normal brain function and psychopathology. Mol Psychiatry 15(12): 1128-1139.
Segerström L, Gustavsson J, Nylander I (2016). Minimizing Postsampling Degradation of Peptides by a Thermal Benchtop Tissue Stabilization Method. Biopreserv Biobank 14(2): 172-179.
Shnitko TA, Spear LP, Robinson DL (2016). Adolescent binge-like alcohol alters sensitivity to acute alcohol effects on dopamine release in the nucleus accumbens of adult rats. Psychopharmacology (Berl) 233(3): 361-371.
Shoemaker WJ, Vavrousek-Jakuba E, Arons CD, Kwok FC (2002). The acquisition and maintenance of voluntary ethanol drinking in the rat: effects of dopaminergic lesions and naloxone. Behav Brain Res 137(1-2): 139-148.
Siegmund S, Vengeliene V, Singer MV, Spanagel R (2005). Influence of age at drinking onset on long-term ethanol self-administration with deprivation and stress phases. Alcohol Clin Exp Res 29(7): 1139-1145.
Silveri MM, Dager AD, Cohen-Gilbert JE, Sneider JT (2016). Neurobiological signatures associated with alcohol and drug use in the human adolescent brain. Neurosci Biobehav Rev 70: 244-259.
Simon EJ, Hiller JM, Edelman I (1973). Stereospecific binding of the potent narcotic analgesic (3H) Etorphine to rat-brain homogenate. Proc Natl Acad Sci U S A 70(7): 1947-1949.
Simpson J, Kelly JP (2011). The impact of environmental enrichment in laboratory rats--behavioural and neurochemical aspects. Behav Brain Res 222(1): 246-264.
Sirohi S, Bakalkin G, Walker BM (2012). Alcohol-induced plasticity in the dynorphin/kappa-opioid receptor system. Front Mol Neurosci 5: 95.
Siviy SM, Panksepp J (2011). In search of the neurobiological substrates for social playfulness in mammalian brains. Neurosci Biobehav Rev 35(9): 1821-1830.
Sjoerds Z, de Wit S, van den Brink W, Robbins TW, Beekman AT, Penninx BW, et al. (2013). Behavioral and neuroimaging evidence for overreliance on habit learning in alcohol-dependent patients. Transl Psychiatry 3: e337.
Skinner BF (1938). The Behaviour of Organisms: An Experimental Analysis. edn. D. Appelton-Century Company, Inc: USA.
Slawecki CJ, Betancourt M (2002). Effects of adolescent ethanol exposure on ethanol consumption in adult rats. Alcohol 26(1): 23-30.
Smith MA, Bryant PA, McClean JM (2003). Social and environmental enrichment enhances sensitivity to the effects of kappa opioids: studies on antinociception, diuresis and conditioned place preference. Pharmacology Biochemistry and Behavior 76(1): 93-101.
Smith MA, Chisholm KA, Bryant PA, Greene JL, McClean JM, Stoops WW, et al. (2005). Social and environmental influences on opioid sensitivity in rats: importance of an opioid's relative efficacy at the mu-receptor. Psychopharmacology (Berl) 181(1): 27-37.
Söderpalm B, Ericson M (2013). Neurocircuitry involved in the development of alcohol addiction: the dopamine system and its access points. Curr Top Behav Neurosci 13: 127-161.

76
Sokoloff P, Giros B, Martres MP, Bouthenet ML, Schwartz JC (1990). Molecular cloning and characterization of a novel dopamine receptor (D3) as a target for neuroleptics. Nature 347(6289): 146-151.
Spain JW, Roth BL, Coscia CJ (1985). Differential ontogeny of multiple opioid receptors (mu, delta, and kappa). J Neurosci 5(3): 584-588.
Spanagel R, Herz A, Shippenberg TS (1990). The Effects of Opioid Peptides on Dopamine Release in the Nucleus Accumbens: An In Vivo Microdialysis Study. J Neurochem 55(5): 1734-1740.
Spano PF, Govoni S, Trabucchi M (1978). Studies on the pharmacological properties of dopamine receptors in various areas of the central nervous system. Adv Biochem Psychopharmacol 19: 155-165.
Spear LP (2015). Adolescent alcohol exposure: Are there separable vulnerable periods within adolescence? Physiol Behav 148: 122-130.
Spear LP (2000). The adolescent brain and age-related behavioral manifestations. Neurosci Biobehav Rev 24(4): 417-463.
Spear LP (2013). Adolescent neurodevelopment. J Adolesc Health 52(2 Suppl 2): S7-13.
Spear LP (2014). Adolescents and alcohol: acute sensitivities, enhanced intake, and later consequences. Neurotoxicol Teratol 41: 51-59.
Spear LP (2018). Effects of adolescent alcohol consumption on the brain and behaviour. Nat Rev Neurosci 19(4): 197-214.
Spinka M, Newberry RC, Bekoff M (2001). Mammalian play: training for the unexpected. Q Rev Biol 76(2): 141-168.
Spoelder M, Hesseling P, Styles M, Baars AM, Lozeman-van 't Klooster JG, Lesscher HM, et al. (2017). Dopaminergic neurotransmission in ventral and dorsal striatum differentially modulates alcohol reinforcement. Eur J Neurosci 45(1): 147-158.
Squeglia LM, Tapert SF, Sullivan EV, Jacobus J, Meloy MJ, Rohlfing T, et al. (2015). Brain development in heavy-drinking adolescents. Am J Psychiatry 172(6): 531-542.
Steensland P, Fredriksson I, Holst S, Feltmann K, Franck J, Schilstrom B, et al. (2012). The monoamine stabilizer (-)-OSU6162 attenuates voluntary ethanol intake and ethanol-induced dopamine output in nucleus accumbens. Biol Psychiatry 72(10): 823-831.
Steiner H, Gerfen CR (1998). Role of dynorphin and enkephalin in the regulation of striatal output pathways and behavior. Exp Brain Res 123(1-2): 60-76.
Sukikara MH, Platero MD, Canteras NS, Felicio LF (2007). Opiate regulation of behavioral selection during lactation. Pharmacol Biochem Behav 87(3): 315-320.
Sulzer D (2011). How addictive drugs disrupt presynaptic dopamine neurotransmission. Neuron 69(4): 628-649.
Sunahara RK, Guan HC, O'Dowd BF, Seeman P, Laurier LG, Ng G, et al. (1991). Cloning of the gene for a human dopamine D5 receptor with higher affinity for dopamine than D1. Nature 350(6319): 614-619.
Tabakoff B, Hoffman PL (2013). The neurobiology of alcohol consumption and alcoholism: an integrative history. Pharmacol Biochem Behav 113: 20-37.
Teicher MH, Samson JA, Anderson CM, Ohashi K (2016). The effects of childhood maltreatment on brain structure, function and connectivity. Nat Rev Neurosci 17(10): 652-666.
Temin HM, Mizutani S (1970). RNA-dependent DNA polymerase in virions of Rous sarcoma virus. Nature 226(5252): 1211-1213.

77
Terenius L (1973). Stereospecific interaction between narcotic analgesics and a synaptic plasm a membrane fraction of rat cerebral cortex. Acta Pharmacol Toxicol (Copenh) 32(3): 317-320.
Trantham-Davidson H, Centanni SW, Garr SC, New NN, Mulholland PJ, Gass JT, et al. (2017). Binge-Like Alcohol Exposure During Adolescence Disrupts Dopaminergic Neurotransmission in the Adult Prelimbic Cortex. Neuropsychopharmacology 42(5): 1024-1036.
Trezza V, Baarendse PJ, Vanderschuren LJ (2014). On the interaction between drugs of abuse and adolescent social behavior. Psychopharmacology (Berl) 231(8): 1715-1729.
Trezza V, Baarendse PJ, Vanderschuren LJ (2010). The pleasures of play: pharmacological insights into social reward mechanisms. Trends Pharmacol Sci 31(10): 463-469.
Trezza V, Damsteegt R, Achterberg EJ, Vanderschuren LJ (2011). Nucleus accumbens mu-opioid receptors mediate social reward. J Neurosci 31(17): 6362-6370.
Trezza V, Vanderschuren LJ (2008). Cannabinoid and opioid modulation of social play behavior in adolescent rats: differential behavioral mechanisms. Eur Neuropsychopharmacol 18(7): 519-530.
Trigo JM, Martin-Garcia E, Berrendero F, Robledo P, Maldonado R (2010). The endogenous opioid system: a common substrate in drug addiction. Drug Alcohol Depend 108(3): 183-194.
Troisi A, Frazzetto G, Carola V, Di Lorenzo G, Coviello M, Siracusano A, et al. (2012). Variation in the mu-opioid receptor gene (OPRM1) moderates the influence of early maternal care on fearful attachment. Soc Cogn Affect Neurosci 7(5): 542-547.
Turner PV, Vaughn E, Sunohara-Neilson J, Ovari J, Leri F (2012). Oral gavage in rats: animal welfare evaluation. J Am Assoc Lab Anim Sci 51(1): 25-30.
Tye KM, Prakash R, Kim S-Y, Fenno LE, Grosenick L, Zarabi H, et al. (2011). Amygdala circuitry mediating reversible and bidirectional control of anxiety. Nature 471: 358.
Urban NB, Kegeles LS, Slifstein M, Xu X, Martinez D, Sakr E, et al. (2010). Sex differences in striatal dopamine release in young adults after oral alcohol challenge: a positron emission tomography imaging study with [(1)(1)C]raclopride. Biol Psychiatry 68(8): 689-696.
van Ree JM, Gerrits MA, Vanderschuren LJ (1999). Opioids, reward and addiction: An encounter of biology, psychology, and medicine. Pharmacol Rev 51(2): 341-396.
Van Skike CE, Diaz-Granados JL, Matthews DB (2015). Chronic intermittent ethanol exposure produces persistent anxiety in adolescent and adult rats. Alcoholism-Clinical and Experimental Research 39(2): 262-271.
Van Tol HH, Bunzow JR, Guan HC, Sunahara RK, Seeman P, Niznik HB, et al. (1991). Cloning of the gene for a human dopamine D4 receptor with high affinity for the antipsychotic clozapine. Nature 350(6319): 610-614.
Vanderschuren LJ, Achterberg EJ, Trezza V (2016). The neurobiology of social play and its rewarding value in rats. Neurosci Biobehav Rev 70: 86-105.
Vanderschuren LJ, Niesink RJ, Spruijt BM, Van Ree JM (1995a). Effects of morphine on different aspects of social play in juvenile rats. Psychopharmacology (Berl) 117(2): 225-231.
Vanderschuren LJ, Niesink RJ, Spruijt BM, Van Ree JM (1995b). Mu- and kappa-opioid receptor-mediated opioid effects on social play in juvenile rats. Eur J Pharmacol 276(3): 257-266.

78
Vanderschuren LJ, Spruijt BM, Hol T, Niesink RJ, Van Ree JM (1995c). Sequential analysis of social play behavior in juvenile rats: effects of morphine. Behav Brain Res 72(1-2): 89-95.
Vanderschuren LJ, Stein EA, Wiegant VM, Van Ree JM (1995d). Social play alters regional brain opioid receptor binding in juvenile rats. Brain Res 680(1-2): 148-156.
Varlinskaya EI, Kim EU, Spear LP (2016). Chronic intermittent ethanol exposure during adolescence: Effects on stress-induced social alterations and social drinking in adulthood. Brain Res.
Varty GB, Hyde LA, Hodgson RA, Lu SX, McCool MF, Kazdoba TM, et al. (2005). Characterization of the nociceptin receptor (ORL-1) agonist, Ro64-6198, in tests of anxiety across multiple species. Psychopharmacology (Berl) 182(1): 132-143.
Varty GB, Lu SX, Morgan CA, Cohen-Williams ME, Hodgson RA, Smith-Torhan A, et al. (2008). The anxiolytic-like effects of the novel, orally active nociceptin opioid receptor agonist 8-[bis(2-methylphenyl)methyl]-3-phenyl-8-azabicyclo[3.2.1]octan-3-ol (SCH 221510). J Pharmacol Exp Ther 326(2): 672-682.
Vena AA, Mangieri R, Gonzales RA (2016). Regional Analysis of the Pharmacological Effects of Acute Ethanol on Extracellular Striatal Dopamine Activity. Alcohol Clin Exp Res 40(12): 2528-2536.
Vergura R, Balboni G, Spagnolo B, Gavioli E, Lambert DG, McDonald J, et al. (2008). Anxiolytic- and antidepressant-like activities of H-Dmt-Tic-NH-CH(CH2-COOH)-Bid (UFP-512), a novel selective delta opioid receptor agonist. Peptides 29(1): 93-103.
Verma IM, Temple GF, Fan H, Baltimore D (1972). In vitro synthesis of DNA complementary to rabbit reticulocyte 10S RNA. Nat New Biol 235(58): 163-167.
Vetter CS, Doremus-Fitzwater TL, Spear LP (2007). Time course of elevated ethanol intake in adolescent relative to adult rats under continuous, voluntary-access conditions. Alcohol Clin Exp Res 31(7): 1159-1168.
Vitale G, Ruggieri V, Filaferro M, Frigeri C, Alboni S, Tascedda F, et al. (2009). Chronic treatment with the selective NOP receptor antagonist [Nphe 1, Arg 14, Lys 15]N/OFQ-NH 2 (UFP-101) reverses the behavioural and biochemical effects of unpredictable chronic mild stress in rats. Psychopharmacology (Berl) 207(2): 173-189.
Volkow ND, Ding YS, Fowler JS, Wang GJ, Logan J, Gatley SJ, et al. (1996). Dopamine transporters decrease with age. J Nucl Med 37(4): 554-559.
Volkow ND, Koob GF, Croyle RT, Bianchi DW, Gordon JA, Koroshetz WJ, et al. (2017). The conception of the ABCD study: From substance use to a broad NIH collaboration. Dev Cogn Neurosci.
Vollstädt-Klein S, Wichert S, Rabinstein J, Buhler M, Klein O, Ende G, et al. (2010). Initial, habitual and compulsive alcohol use is characterized by a shift of cue processing from ventral to dorsal striatum. Addiction 105(10): 1741-1749.
Watson JD, Crick FH (1953). Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid. Nature 171(4356): 737-738.
Weiss F, Lorang MT, Bloom FE, Koob GF (1993). Oral alcohol self-administration stimulates dopamine release in the rat nucleus accumbens: genetic and motivational determinants. J Pharmacol Exp Ther 267(1): 250-258.
Weiss F, Parsons LH, Schulteis G, Hyytia P, Lorang MT, Bloom FE, et al. (1996). Ethanol self-administration restores withdrawal-associated deficiencies in

79
accumbal dopamine and 5-hydroxytryptamine release in dependent rats. J Neurosci 16(10): 3474-3485.
Werling LL, Frattali A, Portoghese PS, Takemori AE, Cox BM (1988). Kappa receptor regulation of dopamine release from striatum and cortex of rats and guinea pigs. Journal of Pharmacology and Experimental Therapeutics 246(1): 282-286.
Wise RA (1973). Voluntary Ethanol Intake in Rats Following Exposure to Ethanol on Various Schedules. Psychopharmacologia 29(3): 203-210.
Witkin JM, Statnick MA, Rorick-Kehn LM, Pintar JE, Ansonoff M, Chen Y, et al. (2014). The biology of Nociceptin/Orphanin FQ (N/OFQ) related to obesity, stress, anxiety, mood, and drug dependence. Pharmacol Ther 141(3): 283-299.
Wong ML, Medrano JF (2005). Real-time PCR for mRNA quantitation. Biotechniques 39(1): 75-85.
Wongwitdecha N, Marsden CA (1996). Effect of social isolation on the reinforcing properties of morphine in the conditioned place preference test. Pharmacol Biochem Behav 53(3): 531-534.
Yaksh T, Wallace M (2011). Opioids, Analgesia, and Pain Management. In: Goodman and Gillman's The Pharmacological Basis of Therapeutics, Brunton LL CB, Knollman BC (ed) Vol. 12, pp 481-526. New York: McGraw-Hill.
Yalow RS, Berson SA (1959). Assay of plasma insulin in human subjects by immunological methods. Nature 184 (Suppl 21): 1648-1649.
Yates JR, Beckmann JS, Meyer AC, Bardo MT (2013). Concurrent choice for social interaction and amphetamine using conditioned place preference in rats: effects of age and housing condition. Drug Alcohol Depend 129(3): 240-246.
Yoder KK, Constantinescu CC, Kareken DA, Normandin MD, Cheng TE, O'Connor SJ, et al. (2007). Heterogeneous effects of alcohol on dopamine release in the striatum: a PET study. Alcoholism-Clinical and Experimental Research 31(6): 965-973.
York JL, Welte J, Hirsch J, Hoffman JH, Barnes G (2004). Association of age at first drink with current alcohol drinking variables in a national general population sample. Alcoholism-Clinical and Experimental Research 28(9): 1379-1387.
Zahniser NR, Dickinson SD, Gerhardt GA (1998). High-speed chronoamperometric electrochemical measurements of dopamine clearance. Methods Enzymol 296: 708-719.
Zandy SL, Matthews DB, Tokunaga S, Miller AD, Blaha CD, Mittleman G (2014). Reduced dopamine release in the nucleus accumbens core of adult rats following adolescent binge alcohol exposure: age and dose-dependent analysis. Psychopharmacology (Berl).

Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Pharmacy 253
Editor: The Dean of the Faculty of Pharmacy
A doctoral dissertation from the Faculty of Pharmacy, UppsalaUniversity, is usually a summary of a number of papers. A fewcopies of the complete dissertation are kept at major Swedishresearch libraries, while the summary alone is distributedinternationally through the series Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty ofPharmacy. (Prior to January, 2005, the series was publishedunder the title “Comprehensive Summaries of UppsalaDissertations from the Faculty of Pharmacy”.)
Distribution: publications.uu.seurn:nbn:se:uu:diva-346151
ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2018