stofftransport im ovar von pyrrhocoris apterus l
TRANSCRIPT

Z. Zellforsch. 123, 395~410 (1972) �9 by Springer-Verlag 1972
Stofftransport im Ovar von Pyrrhocoris apterus L. A u t o r a d i o g r a p h i s c h e U n t e r s u c h u n g e n z u m S t o f f t r a n s p o r t y o n d e n N~ihrze l len
z u r O o c y t e d e r F e u e r w a n z e Pyrrhocoris apterus L. ( H e t e r o p t e r a ) * **
UDO MAYs
Zoologisches Insti tut der Universit/it Mfinster
Eingegangen am 27. Jul i 1971
A u t o r a d i o g r a p h i c S tud ie s on the T r a n s p o r t of S u b s t a n c e s
f r o m t h e N u r s e Cells to t h e O o c y t e of Pyrrhocoris apterus L. ( H e t e r o p t e r a )
Summary. Synthesis and transport of DNA, RNA, and proteins were studied in a telo- trophic ovary using autoradiography. The occurrence of endomitosis in nuclei of differentia- ting nurse cells and follicular epithelium can be confirmed for Pyrrhocoris and also the trans- port of high molecular RNA from the nurse cell nuclei into the oocytes during their phase of euplasmatic growth. There is no transport into younger oocytes. In Pyrrhocoris no contri- bution of the follicular epithelium to the RNA supply of the oocytes is found as described for a different Heteroptere; contrary to the findings reported for other Heteroptera, a rela- t ively low rate of RNA synthesis is detected in all oocyte nuclei. Protein is synthesized with different intensity in all parts of the ovariole and transported from the nurse cells into the oocytes.
I t is possible, like in the polytrophic ovary, to distinguish between two modes of RNA transport. The bulk of the I~NA seems to moore at the same low speed as the proteins, pre- sumably representing a passive movement of ribosomes in the flow of euplasma. A smaller fraction of labelled RNA is transported faster through the cytoplasm; it may consist of messenger RNA.
Key words: RNA-Transport - - Telotrophic ovary - - Oogenesis.
Zusammen]assung. Synthese und Transport yon DNS, RNS und Proteinen in einem telo- trophen Ovar wurden autoradiographisch untersucht. Das Vorkommen yon Endomitosen in Kernen sich differenzierender N~hrzellen und des Follikelepithels kSnnen ffir Pyrrhocoris besthtigt werden, desgleichen der Transport yon hochmolekularer RNS yon den N~hrzell- kernen zu den Oocyten in der Phase euplasmatischen Waehstums. Ein Transport in jfingere Oocyten erfolgt nicht. Ein Beitrag des Follikelepithels zur RNS-Versorgung der Oocyten wie er fiir eine andere Wanze beschrieben wurde, ist bei Pyrrhocoris nicht feststellbar; gleich- falls im Gegensatz zu den Befunden an anderen Wanzen finder eine relativ schwache RNS- Synthese in allen Oocytenkernen start. Protein wird mit unterschiedlicher Intensit~t in der gesamten Ovariole synthetisiert und yon den N~hrzellen in die Oocyten transportiert.
Wie im polytrophen Ovar sind zwei RNS-Transporte unterscheidbar: Die langsamere Hauptfraktion hat anscheinend die gleiche, geringe Geschwindigkeit wie der Proteintrans- port; es wird daher angenommen, dab es sich um die passive Verfrachtung von Ribosomen im Strom des Euplasma handelt. Eine geringere Menge markierter RNS wird schneller durch das Cytoplasma transportiert und als messenger-RNS diskutiert.
* Gewidmet dem Andenken an meinen verehrten Lehrer, Herrn Prof. Dr. K. Bier, unter dessen Anleitung diese Arbeit entstand.
** Die Untersuchungen wurden z. T. mit Mitteln durchgefiihrt, die die Deutsche Forschungs- gemeinschaft Herrn Prof. Bier zur Verffigung stellte.

396 U. Mays:
Einleitung
Das Wachstum der Eier im meroistischen Insektenovar unterscheidet sich prinzipiell vom Eiwachstum bei den meisten anderen im Tierreieh verbreiteten Ovartypen dadurch, dab hier ein mehr oder weniger groBer Teil der erforderliehen RNS in den Kernen von Schwesterzellen der Oocyten, die relativ sp~t aus der Keimbahn austreten, synthetisiert wird (Bier, 1964b, 1967).
I m polytroph-meroistischen Ovar yon Musca dome.~tica konnte Bier (1963) autoradiographisch zwei RNS-Fraktionen durch ihre unterschiedlichen Wande- rungs- und Umsatzgeschwindigkeiten unterscheiden. Eine geringere Menge nicht 16slicher RNS yon hoher Umsatzrate, mSglieherweise messenger-RNS, wird zur Zeit des st~rksten euplasmatischen Waehstums der Oocyte von den Ni~hrzellkernen durch das Cytoplasma des N~hrfaehs fiber Fusome in die Oocyte transportiert. Dieser Transport erfolgt gerichtet und aktiv (Bier, 1964a). Die Hauptmenge der synthetisierten RNS bildet eine stabflere Fraktion, die in einem sp~teren Ab- schhitt der Oogenese zusammen mit dem Inhal t des N~hrfachs der Ooeyte zuge- ffihrt wird; es dfirfte sich hierbei um ribosomale RNS handeln.
Ffir weitere Untersuchungen der damit verbundenen Problematik bietet sich der telotroph-meroistische Ovartyp an. Hier sind die Niihrzellen oder Trophocyten ffir alle waehsenden Oocyten einer Ovariole gemeinsam in der Endkammer zu- sammengefal~t. Trophogonien in der Spitze der Endkammer liefern mitotisch suk- zessiv Zellen, die sieh zu Trophocyten differenzieren. Die Syntheseprodukte der polyploiden N~hrzellen gelangen in einen gemeinsamen zentralen Plasmahof. Am Grunde der Endkammer schliei]t sich an die N~hrzellen ringf6rmig das Lager der jfingsten Oocyten, die noch nicht mit dem Eiwachstum begonnen haben, an. Sie werden durehsetzt und umgeben vom prAfollikul~ren Gewebe, dessen Zellen als Bildungsgewebe des Follikelepithels fungieren. Dieses Epithel wiederum um- kleidet die Oocyten w~hrend ihrer gesamten weiteren Entwicklung. Vom Beginn des euplasmatischen Wachstums an und bis zum Einsetzen der Dottereinlagerung einsehlieBlich sind die Oocyten des telotrophen Ovars fiber je einen NAhrstrang, der durch das Follikelepithel verli~uft, mit dem Plasmahof verbunden. Die L~nge dieses Transportweges fiberschreitet teilweise 1 mm. Die gfinstigen histologischen Verh~ltnisse ermSgliehen eine Untersuchung dieses Ovartyps, deren autoradio- graphisehe Ergebnisse hier vorgelegt werden (vgl. hierzu aueh Mays, 1969). In einer zweiten Arbeit soll die Ultrastruktur des Transportwegs behandelt werden.
Material und Methoden
Als Versuchstiere dienten Weibchen der Feuerwanze Pyrrhocoris apterus L. Die Haltung erfolgte bei 21~ und 50---70% rel. Luftfeuchte in Kunststoffgef~Ben, Ern~hrung mit Samen der Winterlinde, Tilia cordata Miller und Wasser. T~glich 18stfindige Belichtung verhindert die Diapause. Die Weibchen legen dann in regelm~lligen Zyklen das ganze Jahr fiber ihre Eier ab (SlAma, 1964a). Von der Adulth/iutung an wurden die Tiere in Gruppen gleichen H~utungsalters gehalten, so da[l datierte Versuchstiere zur Verffigung standen.
Als Tracer wurden die folgenden tritiumsubstituierten Verbindungen des Radiochemical Centre, Amersham, England, verwandt: aH-Uridin, aH-Uridin-5'-monophosphat, 3H-Cytidin- 5'-monophosphat, aH-Thymidin und ein Gemisch der Aminos/~uren 3H-Valin, 3H-Leucin, aH-Lysin und aH-Histidin. Die spezifischen und absoluten Aktivit/iten werden bei der Versuchs- beschreibung angegeben. Die Substanzen lagen als L6sungen mit 1 Curie Aktivit~t pro Liter vor; 3--10 txl wurden je Tier in die Leibesh6hle injiziert.

Stofftransport im Ovar von Pyrrhocoris apterus L. 397
Nach Ablauf der Inkubationszeit wurden die Ovarien in modifiziertem Carnoyschem Ge- misch (Athanol/Essigsiiure/Chloroform = 8/1/1) fixiert, nach der fiblichen Paraffintechnik die Schnitte auf mit Chromalaun-Gelatine behandelte ObjekttrKger gezogen, entparaffiniert, 15 min bei +5~ in 5%iger Trichloressigs~urel6sung behandelt, ausgewaschen, mit Strip- film Kodak AR 10 versehen und nach angemessener Exposition im Kfihlschrank in luftdich- ten Beh~ltern, die mit Trockenmittel bestiickt waren, entwickelt. Die Spezifit~t des Einbaus yon Tracern in RNS wurde durch Behandlung yon Kontrollobjekttr~gern mit RNase gepriift. Fixierte, gewiisserte und lufttrockene Pr~iparate wurden in Euparal eingeschlossen. Vergl. auch Mays (1969).
Beobaehtung
1. Der Einbau von Thymidin
Jungen Pyrrhocoris-9~ (5--8 Tage nach der Imaginalh~utung) wurden je 5 ~1=5 ~c att-Thymidin (spezifische Aktivit~it 8 100 c/Mol) injiziert. Die Inku- bationszeiten erstreckten sich yon 1 Std bis zu 10 Tagen.
Nach kurzen Inkubationszeiten ergibt sich ein einheitliches Markierungsbild. Einzelne markierte Kerne, die sich w~hrend der Verffigungszeit des Tracers in S-Phase befanden, bflden ein Mosaik. Die Zellen in S-Phase verteilen sich wie folgt auf die Regionen des Ovars (Abb. 1):
1. Trophogonien (im Apex der Endkammer unter dem Ansatz des Terminal- filum gelegen), die mitotisch N~hrzellen naehliefern.
2. Die N~hrzellen insgesamt. Jedoch finden sich markierte Zellen in gr6Berer Zahl in den mehr apikal gelegenen Bereichen. Da sie nicht mitotisch akt iv sind, ist dies der direkte Nachweis fiir die Endomitosen, auf die andere Autoren aus der Zunahme yon Chromatin und Kernvolumen geschlossen haben. Damit werden auch die autoradiographisehen Befunde yon Vanderberg (1963) an Rhodnius pro- lixus (Reduviidae) best~tigt.
3. Das pr~follikul~re Gewebe; dort linden reichlich Mitosen statt . 4. Das Follikelepithel, nicht nur apikal wo Mitosen noch vorkommen, sondern
in den ganzen Bereiehen der Pr~vitellogenese und Vitellogenese; auch dies ist eine Best~tigung sehon frfiher besehriebener Endomitosen.
Mit zunehmender Inkubationszeit treten zweierlei Ver~nderungen auf: Einmal verschieben sich die markierten Kerne durch Wachstumsprozesse in der Ovariole ab- w~rts (Abb. 2) zum Pedizell hin; Trophogonien, pr~follikul~res Gewebe und Follikel- epithel der jfingsten Oocyten in Pr~vitellogenese enthalten nach 8 Tagen nur noch wenige markierte Kerne. Zweitens laufen zwei Vorg~nge nebeneinander ab, die in ihrer Deutlichkeit bei den Versuchstieren individuell variieren: Eine diffuse Markierung breitet sich fiber das ganze Objekt aus; ihr zeitlicher Beginn ist wegen des background nicht feststellbar. Zuerst f~llt sie naeh 24 Std auf, naeh 5- -8 Tagen ist sie stark und nach 10 Tagen wieder recht schwach. Daneben ver- ringert sich die Zahl der markierten Kerne im Ovar fiberhaupt. In einem Fall war nach 10 Tagen nur noeh eine sehwache allgemeine Markierung des gesamten Sehnittes vorhanden, w~hrend andererseits ein anderes Tier naeh acht Tagen noch viele markierte N~hrzellkerne besaB, die in zwei Tagen nicht altern und zugrunde gehen k6nnen - - zumal keine Radioaktivit~t in den pyknotischen Ker- nen der degenerierenden ~lteren N~hrzellen gefunden wurde. Eine spezifische Markierung von Plasmahof und Ni~hrstr~ngen t ra t naeh Inkubat ion mit 3tt- Thymidin nicht auf.
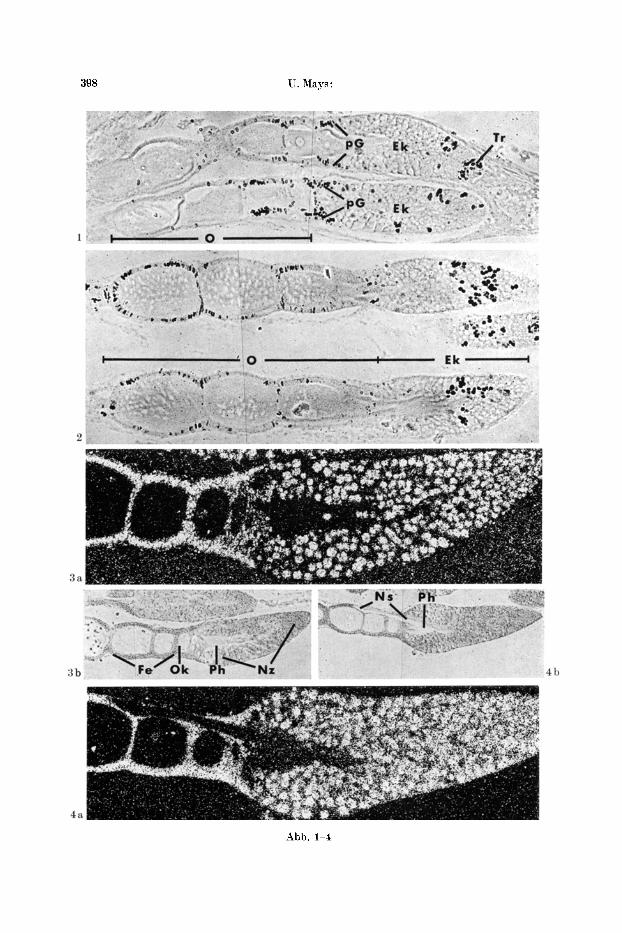
398 U. Mays:
Abb. 1-4

Stofftransport im Ovar von Pyrrhocoris apterus L. 399
2. Der Einbau von RNS-Vorstu/en Die Versuchstiere ha t t en ein Alter von 5 - -14 Tagen nach der Imaginalhgu-
tung. Sie befanden sich z.T. in der gleichen Phase des Reprodukt ionszyklus , da ihnen unmi t t e lba r nach der Eiablage Tracer injiziert wurde. Der Vorteil dieses Verfahrens liegt mehr in der histologischen Aufarbe i tung (keine Chorioneier, vielc wachsende Oocyten ohne oder mi t wenig Dot ter in der Ovariole) als in der physiologischen Gleichschaltung der Tiere, denn die pr~vitellogenetische Wachs- tumsphase der Oocyten, mi t der sich diese Unte r suchung befaBt, ist bei Pyrr- hocoris nach Sls (1964b) und Masner (1968) im Gegensatz zur Vitellogenese, die vom Juven i lho rmon gesteuert wird, kciner humora len Kontrol le unterworfen. Je Tier wurden Akt iv i t~ ten yon 3 - -10 ~c injiziert. Als Tracer d ienten aH-Uridin der spezifischen Akt iv i t~ ten 3000, 17400 und 30000 c/Mol sowie ein Gemisch yon aH-Uridin-5 ' -monophosphat (11250 c/Mol) und 3H-Cytidin-5 ' -monophosphat (14600 c/Mol) in Mengen yon 3 - - 5 ~c je Tier. Die Inkuba t ionsze i t en fiberstrichen einen Bereich yon 15 min bis zu 5 Tagen.
Der Wandel des Markierungsmusters der RNS im Laufe der Inkubation isf vielgesta|tig. Das Bild kompliziert sich durch zwei Nebeneffekte. Einmal tritt nach kurzen Inkubations- zeiten bis etwa 4 Std eine Untergrundmarkierung auf, die sich ganz fiber die Ovariolen er- streckt, zum Teil aber als noch st~rkerer Hof aul3en um das histologische Objekt herum liegt (Abb. 3) und die Konturen des Paraffinschnitts nachzeichnet. Ahnliches wird aueh bei anderen Tieren nach kurzen Inkubationszeiten mit RNS-Vorstufen beobachtet (Bier, mfindliche Mit- teilung). Vermutlich 16sen sich beim Aufziehen und Antrocknen der Schnitte aus dem Gewebe markierte niedermolekulare RNS oder die phosphorylierten Vorstufen, die aus dem betreffen- den pool der Zellen stammen. Sie werden wahrscheinlich an der Oberfl~che des mit Chroma- laun-Gelatine fiberzogenen Objekttr~gers absorbiert. Versuche, die Paraffinschnitte durch l~ngeres Schwimmenlassen auf Wasser yon diesen radioaktiven Verbindungen zu befreien, verliefen ergebnislos. Weiterhin entsteht ira Objekt selbst, und zwar nicht nur in den Kernen, eine Untergrundmarkierung, die zum Teil RNase-resistent ist. Diese Beobachtung macht man auch an anderen Objekten. Es k6nnte sich um einen biochemischen Einbau des Tritiums in andere Verbindungen handeln.
Nach kurzen Inkuba t ionsze i t en von 15--20 min ents teht - - y o n den erwi~hnten Effekten abgesehen - - eine reine Kernmarkierung . Die intensivste Markierung
Abb. 1. Autoradiogramm. DNS-Markierung, 24 Std inkubiert mit aH-Thymidin. L~ngsschnitt zweier Ovariolen. Die markierten Kerne treten dunkel hervor. ]ntensiver Einbau an Orten mitotischer Aktivitiit: Trophogonien (Tr) an der Spitze der Endkammer (Ek), in der oberen Ovariole rechts angeschnitten und priifollikulgres Gewebe (pG) am ~bergang der Endkammer zum Bereich der euplasmatisch wachsenden Oocyten (0). Unterschiedlich verstreuter Ein- bau infolge yon Endomitosen: vereinzelte N~hrzellkerne in der Endkammer und Kerne des
Follikelepithels. Phasenkontrast 100:1 Abb. 2. Wie Abb. 1. DNS-Markierung, 8 Tage inkubiert. Wachstumsbewegungen haben mar- kierte Kerne in der Ovariole abw~rts versehoben: yon den Trophogonien zu den N~ihrzellen
in der Endkammer und vom prafo|likul~ren Gewebe zum Follikelepithel Abb. 3a u. b. Autoradiogramm, RNS-Markierung, 1 Std inkubiert mit aH-Uridin-5'-mono- phosphat und 3H-Cytidin-5'-monophosphat. (}berwiegende Kernmarkierung, N~hrzellen (Nz), pr~follikul~res Gewebe und Follikelepithel (Fe) stark, Oocytenkern (Ok) schwaeh, background fiber Plasmahof (Ph) und Oocyten. a Dunkelfeld 150: l, die einzelnen Silberk6rner des Auto-
radiogramms erscheinen hell. b Phasenkontrast 60:1 Abb. 4a u. b. Wie Abb. 3. RNS-Markierung, 3 Std inkubiert. Noch fiberwiegende Kernmar- kierung, doch findct sich schon Aktivit~t im Plasmahof (Ph). Der Nithrstranganschnitt (Ns)
hat den gleichen background wie die Oocyten
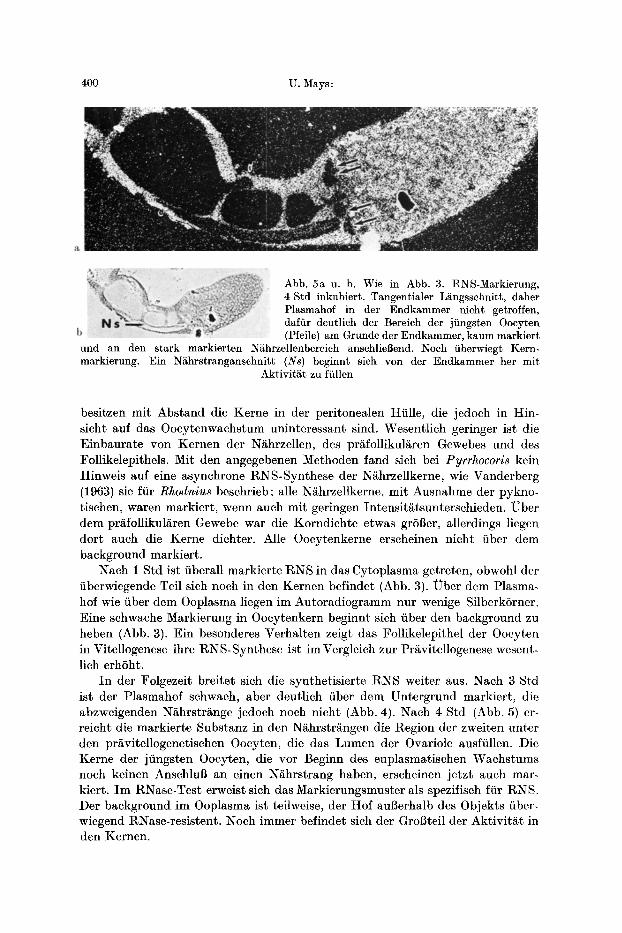
400 U. Mays :
Abb. 5a u. b. Wie in Abb. 3. RNS-Markierung, 4 Std inkubiert. Tangentialer L~ingsschnitt, daher PIasmahof in der Endkammer nicht getroffen, daffir deutlich der Bereich der jiingsten Oocyten (Pfeile) am Grunde der Endkammer, kaum markiert
und an den stark markierten N~hrzellenbereich anschlieBend. Noch iiberwiegt Kern- markierung. Ein N/ihrstranganschnitt (Ns) beginnt sich vonder Endkammer her mit
Aktivit/~t zu ffillen
besitzen mit Abstand die Kerne in der peritoncalen Hfille, die jedoch in Hin- sicht auf das Oocytenwachstum uninteressant sind. Wesentlich gcringer ist die Einbaurate yon Kernen der Ns des pr~ifollikul~ren Gewebes und des Follikelepithcls. Mit den angegcbenen Methoden land sich bei Pyrrhocoris kein Hinweis auf eine asynchrone RNS-Synthese der N~hrzellkerne, wie Vanderberg (1963) sie ffir Rhodnius beschrieb; alle N~hrzellkerne, mit Ausnahme der pykno- tischen, waren markiert, wenn auch mit geringen Intensitiitsunterschieden. l~ber dem pr~follikuli~ren Gewcbe war die Korndichte etwas grSl~er, allerdings liegen dort auch die Kerne dichter. Alle Oocytenkerne erscheinen nicht fiber dem background markiert.
Nach 1 Std ist fiberall markierte RNS in das Cytoplasma getreten, obwohl der fiberwiegende Tell sich noch in den Kernen befindet (Abb. 3). Ober dem Plasma- hof wie fiber dem Ooplasma liegen im Autoradiogramm nur wenige SilberkSrner, Eine schwache Markierung in Oocytenkern beginnt sich fiber den background zu heben (Abb. 3). Ein besonderes Verhaltcn zeigt das Follikelepithel der Oocyten in Vitellogencse ihre RNS-Synthesc ist im Vergleich zur Pr~vitellogenese wcsent- lich erhSht.
In der Folgezeit breitet sich die synthetisicrte RNS welter aus. Nach 3 Std ist der Plasmahof schwach, aber deutlich fiber dem Untergrund markiert, die abzweigenden N~hrstri~nge ]cdoch noch nicht (Abb. 4). Nach 4 Std (Abb. 5) er- reicht die markierte Substanz in den N/~hrstriingen die Region der zweiten unter den pr~vitellogenetischen Oocyten, die das Lumen der Ovariole ausffillen. Die Kerne der jfingsten Oocyten, die vor Beginn des euplasmatischen Wachstums noch keinen Anschlul~ an einen Ni~hrstrang haben, erscheinen jetzt auch mar- kiert. I m RNasc-Test erweist sich das Markicrungsmuster als spezifisch ffir RNS. Der background im Ooplasma ist teilweise, der Hof au~erhalb des Objekts fiber- wicgend RNase-resistent. Noch immer befindet sich der Grol~teil der Aktivit~t in den Kernen.

Stofftransport im Ovar von Pyrrhocoris apterus L. 401
Nach 10 Std haben die N/~hrzellkerne soviel radioaktive RNS an das Cyto- plasma abgegeben, dab sie im Autoradiogramm sehw~cher hervortreten (Abb. 6). In den N/~hrstrs hat sich die Markierung allm/~hlich verst/~rkt und welter ausgebreitet; bei Tieren, die gerade die Vitellogenese begonnen haben, erreieht sie fiber die noch funktionierenden N/ihrstr/~nge die dottereinlagernden Oocyten. Trotzdem beginnt sich nur in der ersten Dotteroocyte ein Markierungsgradient vonder N~hrstrangmfindung her aufzubauen. Die Dotterschollen sind ausgespart, aueh in bezug auf den background.
Naeh 25 Std hat die tritiierte RNS weitgehend die Kerne verlassen, Das trifft auch ffir die Kerne pr~vitellogenetischer Oocyten zu ; sie sind im Autoradiogramm ausgespart in einem Cytoplasma, das sich mit Markierung aus dem N/ihrstrang anreichert. AuBerdem erscheint in den Oocyten ein Markierungsgradient, der yon der Einmfindung des N~hrstrangs ausgeht. Die jfingsten Ooeyten ohne N/~hr- strang bleiben relativ arm an radioaktiver RNS.
30 Std naeh der Injektion erreicht die Markierung der N~hrzellkerne ein Mini- mum, die des Plasmahofs undvor allemdie der N/ihrstr/~nge ein Maximum (Abb. 7). Die Menge der jetzt dort wandernden RNS mag eine bis zwei Gr6Benordnungen fiber derjenigen naeh 10 Std liegen. Z/ihlungen sind sehwierig, da die N/~hrstr/~nge sehmal sind und jedes Fl~ehenelement einen anderen Abstand yon der Endkammer hat. In den Oocyten die bereits mit der Dottereinlagerung begonnen haben - - so- weir vorhanden und noch yon einem N/~hrstrang versorgt - - sieht man durchweg Markierungsgradienten; auch entlang der Reihe wachsender Oocyten einer Ovariole finder sieh ein Markierungsabfall *nit zunehmender Entfernung v o n d e r End- kammer. Die Dotterschollen bleiben frei von Aktivit/it.
Nach 80 Std beginnt sieh das Markierungsmuster etwas zu verwischen, wahr- scheinlich dureh Neueinbau und biochemisehen Umbau der Basen desjenigen Tefls der tritiierten RNS, die inzwisehen abgebaut wurde (Abb. 8). Die Kerne zeigen wieder mehr Markierung, bleiben aber deutlich gegen das hoch aktive Cyto- plasma abgesetzt. AuBer den Dottersehollen ist kein Teil der Ovariole unmar- kiert. Die hSehste Konzentrat ion herrscht in den pr/~vitellogenetischen Oocyten, dort liegt der Gehalt an markierter RNS jetzt fiber dem des pr~follikul/~ren Ge- webes und des Follikelepithels. Trotzdem sieht man kein Auslaufen der Markie. rung aus N/ihrzellen, Plasmahof und N/~hrstr/ingen. Die Oocyten ohne N/~hrstrang bleiben immer noch relativ aktivit/~tsarm. Dieses Bild ist RNS-spezifiseh; nach RNase-Behandlung bleibt eine ziemlieh gleichm/~Bige - - aber entschieden abge- schw/~chte - - Markierung des ganzen Objekts, bei der sieh das Follikelepithel stiirker hervorhebt.
Die letzte untersuchte Inkubationszeit betr~gt 120 Std (keine Abb.). Sie bringt nichts wesentlich Neues. Die Nivellierung des Einbaumusters fiber dem ganzen Objekt, die bei 80 Std beginnt, setzt sieh fort. Noch immer ist der N/ihrzellbereich stark markiert; die Oocyten im Wartestand und die Ooeytenkerne bleiben etwas ausgespart, das Ooplasma in Pr~vite|lo- genese und Vitellogenese extrem stark markiert und der iiberwiegende Teil der Markierung - - besonders das spezifische Muster - - gegen RNase empfindlieh. Eine weitere Verl~ngerung der Inkubationszeiten erscheint daher nicht notwendig.
3. Der Einbau von Aminosguren
Weibchen gleichen Alters wie bei den Versuchen zur RNS-Synthese erhielten Injektionen yon je 5 ~c eines Gemisehes der Aminos/iuren 3H-Histidin (500 c/Mol),

Abb. 6 b
Abb. 6 a Abb. 7 b
Abb. 6 u. 7
Abb. 7 a
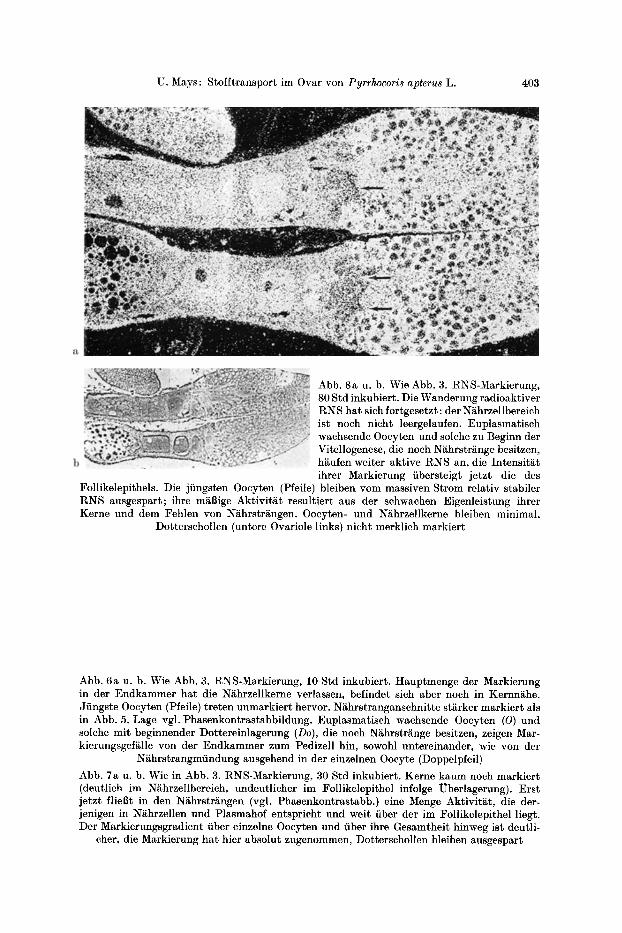
U. Mays: Stofftransport im Ovar von Pyrrhocoris apterus L. 403
Abb. 8a u. b. Wie Abb. 3. RNS-Markierung, 80 Std inkubiert. Die Wanderung radioaktiver RN S hat sich fortgesetzt: der N~hrzellbereich ist noch nicht leergelaufen. Euplasmatisch wachsende Oocyten und solche zu Beginn der Vitellogenese, die noch N~hrstr~inge besitzen, h~ufen weiter aktive RNS an, die Intensit~t ihrer Markierung fibersteigt jetzt die des
Follikelepithels. Die jfingsten Oocyten (Pfeile) bleiben vom massiven Strom relativ stabiler RNS ausgespart; ihre m~Bige Aktivit~t resultiert aus der schwachen Eigenleistung ihrer Kerne und dem Fehlen yon Ni~hrstr~ngen. Oocyten- und N~hrzellkerne bleiben minimal,
Dottersehollen (untere Ovariole links) nicht merklich markiert
Abb. 6a u. b. Wie Abb. 3. RNS-Markierung, 10 Std inkubiert. Hauptmenge der Markierung in der Endkammer hat die N~hrzellkerne verlassen, befindet sich aber noch in Kernn~he. Jfingste Oocyten (Pfeile) treten unmarkiert hervor. N~hrstranganschnitte starker markiert als in Abb. 5, Lage vgl. Phasenkontrastabbildung. Euplasmatisch wachsende Oocyten (O) und solche mit beginnender Dottereinlagerung (Do), die noch N~hrstr~nge besitzen, zeigen Mar- kierungsgef~lle yon der Endkammer zum Pedizell hin, sowohl untereinander, wie yon der
N~hrstrangmfindung ausgehend in der einzelnen Oocyte (Doppelpfeil)
Abb. 7a u. b. Wie in Abb. 3. RNS-Markierung, 30 Std inkubiert. Kerne kaum noch markiert (deutlich im N~hrzellbereich, undeutlicher im Follikelepithel infolge Uberlagerung). Erst jetzt flie6t in den ~ h r s t r ~ n g e n (vgl. Phasenkontrastabb.) eine Menge Aktivit~t, die der- jenigen in N~hrzellen und Plasmahof entspricht und weir fiber der im Follikelepithel liegt. Der Markierungsgradient fiber einzelne Oocyten und fiber ihre Gesamtheit hinweg ist deutli-
cher, die Markierung hat hier absolut zugenommen, Dotterschollen bleiben ausgespart

404 U. Mays:
Abb. 9a u. b. Autoradiogramm, Protein- markierung, 18 min inkubiert mit einem Gemisch yon aH-Aminos~uren. Auffhlliger M~rkierungsgradient : N~hrzellregion - - Plasmahof - - N~hrstr~inge - - Oocyten. 9a Dunkelfeld 150:1. b Phasenkontrast
60:1
aH-Lysin (108 c/Mol), aH-Valin (480 c/Mol) und aH-Leucin (1400 c/Mol) zu glei- chen Aktiviti~ten. Es wurde yon 18 min bis 120 Std inkubiert.
Im Kurzzeitautoradiogramm nach 18 min (Abb. 9) ist jeder Teil der Ovariole, allerdings unterschiedlich stark, markiert, wie auch Vanderberg (1963) bei Rhod- nius bemerkt. Die intensivste Proteinsynthese findet im Follikelepithel statt. Die N~hrzellen stehen dem wenig nach. Der Hof yon Cytoplasma im Germarium zeigt einen geringeren Einbau als die Ni~hrzellen. Die Markierungsintensit~t sinkt von den N~hrzellen fiber Plasmahof und N~hrstr~nge zu den Oocyten und bei diesen mit zunehmendem Alter nach unten deutlich ab.
Nach 5 Std verst~rkt sich im wesentlichen das Kurzzeitbild der Aminosiiure- inkorporation durch Proteinsynthese an Ort und Stelle. Schnelle Verschiebungen yon Markierung sind nicht erkennbar. Lediglich Kerne der Oocyten in Pr~vitello- genese zeigen zwischenzeitlich eine Proteinmarkierung, die zuvor nicht sichtbar war und sich spiiter (s. u.) verwischt.
Nach 20 Std wird eine langsame Verschiebung yon Markierung die Niihrstr/inge hinab deutlich. Sie setzt sich in der Folge fort (vgl. Abb. 10, nach 40 Std). Diese Markierung ist kri~ftig, sie wfirde sonst nicht auffallen, da fiberall (s.o.) Protein synthetisiert wird. Nach sehr langen Inkubationszeiten von 60--120 Std kommt es wie im RNS-Autoradiogramm zu einer Verwischung. Das Bild wird zunehmend gleichfSrmiger. Auch hier ist kein Auslaufen der Markierung aus dem Germarinm zu beobachten.
Diskussion
Die autoradiographischen Befunde nach Kurzzeitinkubationen mit aH-Thymi- din stimmen bei Pyrrhocoris mit den Untersuchungen yon Vanderberg (1963) an Rhodnius fiberein, was die Verteilung von Kernen in S-Phase und den Nachweis yon Endomitosen angeht. Vanderberg wandte keine l~ngeren Inkubationszeiten an; daher entging ihm wahrscheinlich die Verschiebung yon Kernen, die in ihrer S-Phase markiert wurden, in Richtung auf das Pedicell zu. Diese Lageveriinderung
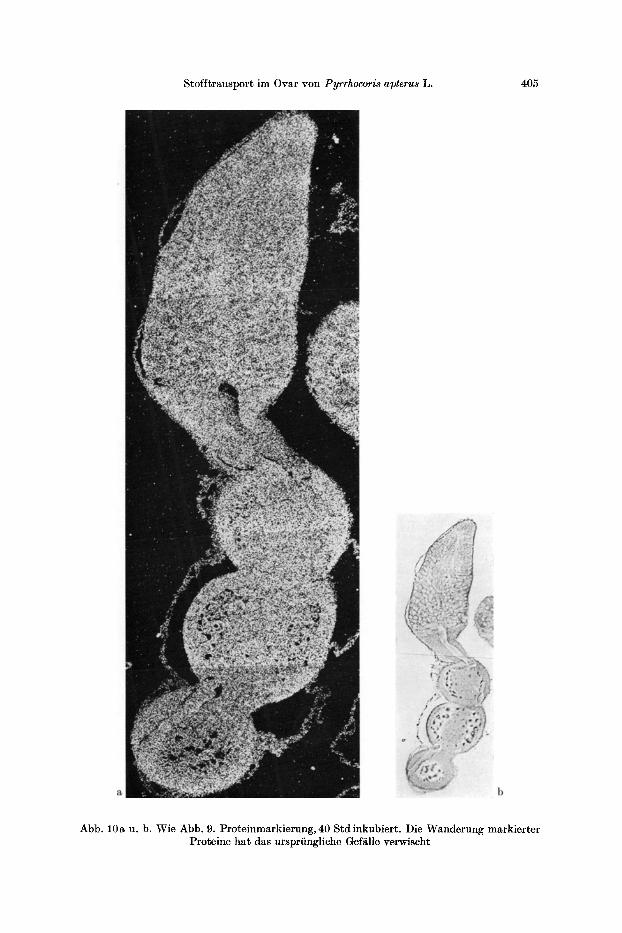
Stofftransport im Ovar von Pyrrhocoris apterus L. 405
Abb. 10a u. b. Wie Abb. 9. Proteinmarkierung, 40 Std inkubiert . Die Wanderung markierter Proteine ha t das ursprfingliche Gef~lle verwischt

406 U. Mays:
ist zwanglos als Folge von Wachstumsprozessen zu deuten, die aus den beiden Zentren mitotischer Aktivit/it in der Ovariole fortw/ihrend Zellen zur Differen- zierung nachliefern, und zwar N/~hrzellen aus dem Bereich der Trophogonien und Zellen des Follikelepithels aus dem Bereich des pr~follikul/~ren Gewebes.
Auch das verz6gerte Auftreten einer cytoplasmatischen Markierung korrespon- diert mit entsprechenden Befunden Vanderbergs. Sie soll bei Rhodnius schon nach 6 Std auftreten, gleichfalls - - wie er ausdrficklich schreibt - - nicht in einer klaren Progression yon den Kernen zum Cytoplasma, sondern als ein allm/~hliches Aussiekern. Er interpretiert seine Beobachtungen als Abbau der Kern-DNS und Transport von mehr oder weniger depolymerisierten Nukleins/iuren in die Oocyte. Bei dieser Deutung stfitzt sich Vanderberg aul~er auf seine eigenen auch auf an- dere histochemische Untersuchung (Schrader und Leuchtenberger, 1952, Acantho- cephala; Bonhag, 1955, Oncopeltus), bei denen sich nie eine positive Feulgen-Reak- tion in den N/ihrstr/~ngen land. Seine Hypothese erscheint plausibel, da einerseits laufend N/ihrzellkerne unter dem Bilde der Pyknose degenerieren, andererseits im Ei ffir die Embryonalentwicklung DNS-Vorstufen angeh/iuft werden mfissen; es ist abet zweifelhaft, ob die autoradiographischen Befunde in dieser Hinsicht Be- weiskraft haben. Erstens ist es wahrscheinlich, dal~ kleine DNS-Bruchstficke oder gar Nukleotide unter den angewandten Priiparationsbedingungen herausgewa- schen werden, zweitens erscheint die diffuse Markierung gleichzeitig fiber dem ganzen Objekt, kann also in den Mitochondrien lokalisiert oder durch Umbau des Tracers unspezifisch sein. Vielleicht muI~ man diese Cytoplasmamarkierung mit dem eigentfimlichen Verschwinden radioaktiv markierter Kerne nach mehrt~giger Inkubationszeit im Zusammenhang sehen. MSglicherweise sind Kerne, die einen erheblichen Tell ihrer DNS aus radioaktiven Vorstufen synthesiert haben, bei langen Inkubationszeiten durch Strahlenschi~den derart im Stoffwechsel gest6rt, dab sie vorzeitig zugrunde gehen und dabei ihre radioaktiven Substanzen frei- setzen, die dann wieder in die Stoffwechselprozesse anderer Zellen eingeschleust werden. Ein Vergleich mit Tieren, denen geringe Aktivit/itsmengen appliziert werden und deren Schnitte entsprechend liinger zu exponieren w/iren, mfiBte darfiber Auskunft geben.
Im telotroph-meroistisehen Ovar entsprechen N/ihrzellen, Plasmahof, N~hr- strKnge und die damit verbundenen Oocyten einer jeden Ovariole in ihrem funk- tionellen Zusammenhang dem Ein/s tines einzelnen Follikels im poly- troph-meriostischen Ovar. Anhand der Autoradiogramme ergibt sich folgender Weg ffir die RNS: Die Synthese erfolgt in den Ni~hrzellkernen der Endkammer, und zwar gleichm/i$ig intensiv in allen Regionen (Abb. 3). Der Transport fiber den Plamahof und die N/~hrstr/inge in die betreffenden Oocyten ist deutlich (Abb. 4--8). Dies stimmt fiberein mit Befunden yon Vanderberg (1963) bei Rhod- nius und Bier (1964b) bei Oncopeltus. Darfiber hinaus gelang es, durch die lan- gen Transportwege begfinstigt, in Analogie zu den Bierschen Untersuchungen (1963, 1964 a), zwei getrennt wandernde RN S-Fraktionen deutlicher als bei Musca darzustellen. Da die Synthese im ganzen Niihrzellbereich gleichzeitig und etwa gleich stark erfolgt und die Wanderwege yon dort zu einem gemeinsamen Be- zugspunkt - - z.B. der Abzweigung der N/ihrstr~nge vom Plasmahof - - unter- schiedlich welt sind, kommt es nicht zu einer Ausbildung klarer Fronten. Z/~h- lungen der Korndichte im Autoradiogramm zur quantitativen Auswertung muB-

Stofftransport im Ovar yon Pyrrhocoris apterus L. 407
ten aus diesen Grfinden unterbleiben und die maximalen Geschwindigkeiten im NKhrstrang lassen sich nur grSBenordnungmKBig mit etwa 200 ~zm pro Stunde ffir die schnell wandernde und 30 ~m/Std fiir die langsamere Fraktion angeben.
Eine relativ geringe Menge markierter RNS l~uft vorweg, die t tauptmenge str6mt erheblich langsamer, innerhalb sinnvoller Inkubationszeiten offenbar nicht wesentlich abebbend, nach. Der Vorlauf ffillt yon der ersten (Abb. 3) zur 3. Std den Plasmahof (Abb. 4), in der 4. (Abb. 5) ist er fiber die zweite Oocyte hinweg die N~hrstr~nge hinabgewandert, hat in der 10. (Abb. 6) die dritte vitellogene- tische Oocyte erreicht und sich weiter apikal in den N~hrstr~ngen verst~irkt. Zu dieser Zeit liegt die Hauptmenge markierter I%NS noch im N~hrzellplasma und der zur/ickgelegte Weg yon der N~hrstrangabzweigung bis zur dritten Oocyte in Vitellogenese ist fast doppelt so groB wie die Distanz der entferntesten N~hr- zellkerne zur Abzweigung. Erst naeh 30 Std hat ein Tell der dichten Markierung (zweite Welle) diese Stecke zurfickgelegt (Abb. 7) und auch nach 80 Std (Abb. 8) ebbt der Nachstrom nicht deutlich ab.
Welter beobachtete man erst nach 10 Std (Abb. 6) in den apikal gelegenen Oocyten eine Markierung, die sich nahe der N~hrstrangmfindung anzureichern beginnt und bei li~ngeren Inkubationszeiten immer starker hervortritt . Diese Region wurde schon nach wenig mehr als 4 Std vom Vorlauf erreicht und dieser ist inzwischen mindestens zwei, eventuell drei Oocyten weiter abw~rts gewan- dert. Das Ausbleiben einer Ansammlung der frfihen Markierung steht im Gegen- satz zum Bild bei Langzeitinkubationen: die dichte, nachlaufende tritiierte RNS sammelt sich anscheinend unbegrenzt in den mit N~hrstrang versehenen Oocyten an (Abb. 8, nach 80 Std). M6glicherweise drfickt sich darin eine verh~ltnism~ig hohe Umsatzrate des Vorlaufs und relative StabflitKt der nachrfickenden Haupt- menge der RNS aus. Dieses Ergebnis stimmt gut mit den Untersuchungen yon Bier (1963, 1964a) an Musca iiberein.
Neuerdings berichten MacGregor und Stebbings (1970) ebenfalls fiber einen Transport yon RNS aus den N~hrzellkernen zu den Oocyten bei Notonecta glauca. Zum gleichen Resultat kommt Brining (1971) an Bruchidius obtectus, was insofern interessant ist, als die polyphagen K~fer telotrophe Ovariolen besitzen, die sieh deutlich yon denen der Hemipteren unterscheiden (s.d.). Die genannten Autoren erwi~hnen keine unterschiedlichen Transportgeschwindigkeiten.
Die Rolle des Follikelepithels bleibt weiterhin rgtselhaft. Vanderbergs Anga- ben fiber einen Beitrag dieses Gewebes zu RNS-Versorgung der Oocyten konnten ffir Pyrrhocoris keineswegs bestiitigt werden. Nur in wenigen Ausnahmen kam es nach mehrstfindiger Inkubation zu einer Markierung der Dotterschollen an der Oberfl~che der Oocyten; dabei handelte es sich sehr wahrscheinlich um eine Folge- erscheinung des Einbaus yon Tritium aus dem Tracer in Aminos~uren, die dann mit den Dotterproteinen eingelagert wurden. Solche unspezffischen Dottermar- kierungen nach Gabe tritiierter RNS-Vorstufen wurden auch bei Gryllus und Panorpa beobachtet (Bier, 1970). Die enorme Steigerung der RNS-Synthese im Follikelepithel nach Einsetzen der ViteUogenese spricht ffir eine entscheidende Beteiligung dieser Zellen bei der Einschleusung yon Dotterproteinen aus der tI~mo- lymphe.
Die RNS-Syntheseaktivit~t der Oocytenkerne beim Eiwachstum in der Prg- vitellogenese und frfihen Vitellogenese ist nicht bedeutend. Daffir spricht ihre
27 Z. Zellforsch., Bd. 123

408 U. Mays:
geringe Einbaurate und die Kondensation der Chromosomen zu einer Karyosph/ire. Pyrrhocoris ffigt sich damit harmonisch in die Reihe der fibrigen Meroisten (Bier, Kunz und Ribb crt, 1967). Bei den yon Bier ( 1964 b) und Vanderberg (1963) unter- suchten Heteropteren wurde kein Uridineinbau im Oocytenkern festgestellt, dies kann jedoch eine Folge der niedrigen spezifischen Aktivit/~t der Tracer sein, die ihnen seinerzeit zur Verffigung standen. Bei Pyrrhocoris ist der Einbau nachweis- bar, doch nm Gr6Benordnungen geringer als in allen anderen Geweben der Ovariole (Abb. 3, 4). Nach Ausweis des RNase-Tests handelt es sich um spezifischen Einbau in RNS. Das Fehlen yon Markierung nach 15--20 rain Inkubation wird dutch die geringe Einbaurate und den relativ hohen background hinreichend crkl/irt. Die Vorstellung eines v611ig inaktiven Oocytenkerns und seiner Markierung durch eine hypothetische einwandernde RNS ist wesentlich weniger wahrscheinlich, zumal auch der Oocytenkern seine Markierung wieder an das Cytoplasma abgibt.
Die Oocyten im Wartestand vor AnschluB an einen N/ihrstrang besitzen gleich- falls schwach aktive Kerne. Erst nach vier Stunden sind sie fiber dem background marMert. Man darf annchmen, dab ihre Synthese nur dem Erhaltungsstoffweehsel der Zellen dient. Bei langen Inkubationszeiten bleiben diese Oocyten yon der nachrfickenden starken Markierung, die die /~lteren Oocyten ffillt, ausgespart (Abb. 8). Dies ist ein wesentlich znverl/tssigerer Nachweis ffir das Fehlen jeder Verbindung zum Plasmahof als die Tatsache, dab morphologisch keine solche gefunden wurde (Mays, 1969).
Die Inkubationszeit bis zum Absinken der Markierungsdichte fiber den Kernen unter das Niveau des umgebenden Cytoplasmas gibt einen Anhaltspunkt ffir die Verffigungszeit Ireien Tracers ; sie ist allerdings kein genanes Mag daffir, da neben der RNS-Syntheserate auch die pool-Gr6Be der Vorstufen und die Geschwindig- keit des Transports der RNS dutch die Kernmembran eine Rolle spielen. Diese Zeit lag bei Inkubationen mit unterschiedlicher Gesamt- und spezifischer Aktivi- ti~t in allen Fallen bei etwa 10 Std. Daraus liiBt sich folgern, dab die applizierten absoluten Tracermengen zumindest nicht sehr groB im Verh/~ltnis zum zelleigenen Vorrat an unmarkierten RNS-Vorstufen waren.
Nach kurzen Inkubationszeiten mit aH-Aminos/~uren zeigt das Autoradio- gramm ein Intensitiitsgef/~lle der Proteinsynthese von den N/ihrzellen zum Plasma- hof und yon dort fiber die Ni~hrstr/~nge hinab zu den Oocyten. Die euplasmatisch wachsenden Oocyten selbst sind vom Germarium abwiirts, ihrer Reihenfolge in der Ovariole entsprechend, ebenfalls zunehmend schwiicher markiert. Das ist mSglicherweise die Folge eines cntsprechenden Konzentrationsgef/illes der ffir die Synthese erforderlichen messenger-RNS (an den beiden anderen RNS-Arten kann kaum Mangel sein). Das Gef~lle kommt vielleicht durch einen relativ raschen Abbau dieser RNS-Fraktion zustande; im vorigen Abschnitt wurden autoradio- graphische tIinweise daffir besprochen. Daraus ergibt sich ein verh/iltnism~Big geringeres euplasmatisches Wachstum der Oocyten mit zunehmendem Alter und Abstand yon der Endkammer, so dab schlieBlich mehrere gleich groBe Oocyten hintereinander liegen sollten. Das ist bei Pyrrhocoris in der Tat regelm/il3ig vor Beginn der Vitellogenese der Fall, undes erscheint auch biologisch sinnvoll, weft in jedem Ovarialzyklus pro Ovariole mehrere Oocyten ziemlich gleichzeitig mit der Dottereinlagerung beginnen und sp/tter als fertige Eier gemeinsam abgelegt werden. Da die Niihrstr/inge nach Vitellogenesebeginn nur kurze Zeit persistieren,

Stofftransport im Owr yon Pyrrhocoris apterus L. 409
h/itten - - wenn die Verh/~ltnisse anders 1/~gen - - die j/ingeren dieser 0ocyten keine Gelegenheit, ihr Defizit an Euplasma gegenfiber den iflteren aufzuholen. Ver- gleicht man die Syntheseaktivit/it in N/ihrzellen, Plasmahof, N/ihrstrang und Oocyten miteinander und setzt sie in Relation zum Volumen der betreffenden Regionen, so ergibt sich, dal~ ein betr/ichtlicher Anteil der Proteine ffir das euplas- matische Eiwachstum in den Oocyten selbst hergestellt wird. Mit dem vorliegenden Material 1/s sich diesc Aussage allerdings nicht quantifizieren. Bei der Mar- kierung, die nach 5 Std vorfibergehend in den Kernen ausschliel]lich der Oocyten in Pr/ivitellogenese auftritt, handelt es sich m6glicherweise um jene Proteine, die schnell zwischen Kern und Cytoplasma wandern (vgl. dazu Goldstein und Prescott, 1967).
Ein langsamer Proteintransport ist im Langzeitautoradiogramm aus der rela- riven Zunahme der Markierung yon N/~hrstr/ingen gegenfiber dem N/~hrzellbereich zu erschliel~en. Leider kann es hier noch weniger zur Ausbildung yon Markie- rungsfronten kommen als im RNS-Autoradiogramm, da Protein im gesamten cytoplasmatischen Raum yon den N/~hrzellen bis zu den Oocyten synthetisiert wird. Vanderberg (1963), der Rhodnius prolixus untersuchte, erw/~hnt diesen Proteintransport nicht, start dessen meint er, einen ~)bertritt markierter Proteine aus den Follikelepithelzellcn in die Oocyte zu sehen. Er miBdeutet offensicht- lich die Einlagerung von Dotterproteinen aus der Hs Diese verl~uft bei Pyrrhocoris nicht anders als in der Oogenese der meisten bisher untersuchten Insekten (vgl. z.B. Bier, 1967).
Die geschilderten Befunde an Pyrrhocoris lassen die Deutung zu, da~ auch im telotrophen Ovar der Hemipteren die Hauptmenge der von den N/ihrzellen pro- duzierten RNS den Oocyten zusammen mit einem Strom yon Euplasma zuge- fiihrt wird, whhrend eine geringe Menge makromolekularer RNS von mSglicher- weise hSherer Umsatzrate mit gr6Berer Geschwindigkeit gerichtet durch das Plasma hindurch zur Oocyte transportiert wird. Es ist eine naheliegende, aber noch zu erh/irtende Hypothese, die schnelle Fraktion fiir messenger-RNS zu halten und die langsame, stabile als ribosomale RNS anzusehen. Zumindest die letztere An- nahme ist durch den Ribosomenreichtum von N~hrzellen, Plasmahof, N~hrstr~n- gen und Oocyten gut gestiitzt (vgl. Mays, 1969). Auch das Auftreten eines Schubs yon Proteinmarkierung, der anscheinend mit gleicher Gesehwindigkeit wie die lang- sam wandernde RNS zu den Ooeyten zieht, wird als Indiz fiir einen Transport yon Proteinen und Ribosomen in einem Strom von Euplasma gcwertet.
Hansen-Delkeskamp (1969) hat in der Oogenese von Gryllus neben der Bil- dung einer groBen Menge ribosomaler RNS und wenig transfer-RNS die Synthese einer heterogenen RNS-Fraktion nachgewiesen, bei der es sich wahrscheinlich um messenger-RNS handelt. Zur sicheren Charakterisierung der transportierten RNS in telotrophen Ovarien, insbesondere der Heteropteren, sind biochemische Unter- suchungen dieses Ovartypes notwendig.
Literatur
Bier, K. : Synthese, interzellul/irer Transpor t und Abbau yon Ribonukleins/iure im Ovar der Stubenfliege Musca domestica. J . Cell Biol. 16, 436--440 (1963).
- - Gerichfeter Ribonukleins/ iuretransport durch das Cytoplasma. Naturwissenschaften 51, 418 (1964a).
27*

410 U. Mays: Stofftransport im Ovar yon Pyrrhocoris apterus L.
Bier, K.: Die Kern-Plasma-Relation und das Riesenwachstum der Eizellen. Zool. Anz., Suppl.-Bd. 28, 84--91 (1964b).
- - Oogenese, das Wachstum von Riesenzellen. Naturwissenschaften 54, 189--195 (1967). - - Oogenesetypen bei Insekten und Vertebraten, ihre Bedeutung for die Embryogenese und
Phylogenese. Zool. Anz., Suppl.-Bd. 83, 7--29 (1970). - - Kunz, W., l~ibbert, D. : Struktur und Funktion der Oocytenchromosomen und Nukleolen
sowie der Extra-DNS w~hrend der Oogense panoistischer und meroistischer Insekten. Chromosoma (Berl.) 28, 214--254 (1967).
Bonhag, P. F. : Histochemical studies of the ovarian nurse tissues and oocytcs of the Milk- weed Bug, Oncopeltus /asciatus (Dallas). I. Cytology, nucleic acids, and carbohydrates. J. Morph. 9 6 , 381--439 (1955).
BOning, J. : Untersuchungen am Ovar des Speisebohnenk~fers Bruchidius obtects Say (Polyph. Col.) zur Kli~rung des Oocytenwachstums in der Pr/~vitellogenese. Diss. Math.-Naturwiss. Fak. MOnster (Westf.) (1971).
Goldstein, L., Prescott, D. M. : Proteins in nucleocytoplasmic interactions. I. The fundamental characteristics of the rapidly migrating proteins and the slow turnover proteins of the Amoeba proteus nucleus. J. Cell Biol. 83, 637--644 (1967).
Hansen-Delkeskamp, E. : Synthese yon R~NS und Protein wKhrend der Oogenese und frfihen Embryogenese yon Acheta domestica. Wilhelm Roux' Archly 162, 114--120 (1969).
MacGregor, H. C., Stebbings, H. : A massive system of microtubules associated with cytoplas- mic movement in telotrophic ovariolcs. J. Cell Sci. 6, 431--449 (1970).
Masner, P. : The inductors of differentiation of prefollicular tissue and the follicular epithelium in ovarioles of Pyrrhocoris apterus (Heteroptera). J. Embryol. exp. Morph. 20, 1--13 (1968).
Mays, U. : Der Stofftransport von den N~hrzellen zur Oozyte in einem telotrophen Insekten- ovar und seine strukturellen Grundlagen; Untersuchungen an der Feuerwanze, Pyrrhocoris apterus L. Diss. Math.-~aturwiss. Fak. Miinster (Westf.) (1969).
Schrader, F., Leuchtenberger, C. : The origin of certain nutritive substances in the eggs of Hemiptera. Exp. Cell Res. 3, 136--146 (1952).
Sl~ma, K. : Hormonal control of respiratory metabolism during growth, reproduction and diapause in female adults of Pyrrhocoris apterus L. (Hemiptera). J. Insect Physiol. 1O, 283--303 (1964a).
- - Hormonal control of haemolymph protein concentration in the adults of Pyrrhocoris ap- terue L. (Hemiptera). J. Insect Physiol. 10, 773--782 (1964b).
Vanderberg, J. P. : Synthesis and transfer of DNA, RNA, and Protein during Vitellogenesis in Rhodnius prolixus (Hemiptera). Biol. Bull. 12~, 556--575 (1963).
Dr. Udo Mays Zoologisches Insti tut der Universitat D-4400 MOnster, Badestr. 9 Deutschland