staphylococcus aureus - aspects of pathogenesis and ...647005/fulltext01.pdf · linköping...
TRANSCRIPT

Linköping University Medical Dissertations No. 1371
Staphylococcus aureus - aspects of pathogenesis and molecular epidemiology
Lisa Stark
Department of Clinical Microbiology, Ryhov County Hospital, Jönköping
Division of Medical Microbiology Department of Clinical and Experimental Medicine
Faculty of Health Sciences Linköping University, Sweden
Linköping 2013

© Lisa Stark, 2013
Published articles have been reprinted with the permission of the copyright holders.
Printed in Sweden by LiU-Tryck, Linköping 2013
ISBN 978-91-7519-568-1 ISSN 0345-0082

Contents
CONTENTS
ABSTRACT……………………………………………………………………………………………….. 1
SAMMANFATTNING PÅ SVENSKA……………………………………………………… 3
LIST OF PAPERS……………………………………………………………………………………… 5
ABBREVIATIONS……………………………………………………………………………………. 7
INTRODUCTION……………………………………………………………………………………… 9
Staphylococcus aureus………………………………………………………………………….. 9
Background………………………………………………………………………………………… 9
Epidemiology……………………………………………………………………………………… 10
Antibiotic resistance…………………………………………………………………………… 11
Methicillin-resistant Staphylococcus aureus………………………………………. 12
Virulence factors and strategies…………………………………………………………. 14
Adhesion to host………………………………………………………………………………… 20
The host…………………………………………………………………………………………………… 25
Innate immunity………………………………………………………………………………… 25
Endothelial cells…………………………………………………………………………………. 28
Typing methods……………………………………………………………………………………… 30
Conventional typing................................................................................ 31
Phage typing……………………………………………………………………………………. 31
Molecular typing methods…………………………………………………………………. 32
Pulsed field gel electrophoresis………………………………………………………... 32
Multilocus sequence typing………………………………………………………………. 33
spa typing…………………………………………………………………………………......... 35

Contents
Terminal restriction fragment length polymorphism………………………… 37
Denaturing gradient gel electrophoresis…………………………………………… 38
Microarray…………………………………………………………………………………....... 40
Gene expression………………………………………………………………………………… 44
Microarray…………………………………………………………………………………....... 44
Real-time PCR…………………………………………………………………………………… 45
AIMS………………………………………………………………………………………………………… 47
RESULTS AND DISCUSSION………………………………………………………………….. 48
Isolation and cultivation of S. aureus (papers I and IV)………………………….. 48
HUVEC as a model to study interactions with S. aureus (papers I and II).………. 52
Internalization and survival of S. aureus in HUVEC (papers I and II)……. 53
S. aureus induction of apoptosis in endothelial cells (papers I and II)…. 54
Gene expression in HUVEC infected with S. aureus (papers I and II)…… 55
Prevalence and molecular epidemiology of S. aureus (paper III)………. 59
Colonization and persistent carriage of S. aureus (paper III)……………… 61
Antibiotic resistance found in S. aureus (papers II and III)…………………….. 62
Multiclonality of S. aureus (papers III and IV)…………………………………………… 63
CONCLUSIONS………………………………………………………………………………………… 65
Benefit for individuals and perspectives for the future……………….. 67
ACKNOWLEDGEMENTS………………………………………………………………………… 68
REFERENCES……………………………………………………………………………………………. 69

Abstract
1
ABSTRACT Staphylococcus aureus is a human commensal colonizing about 30 per cent of the population. Besides, it is a frequent cause of infections such as skin, wound and deep tissue infections and also more life-threatening conditions such as pneumonia, endocarditis and septicaemia. S. aureus may also cause different toxicoses. Moreover, this bacterium is one of the most common causes of nosocomial infections worldwide and an increase in antibiotic resistance, especially against methicillin, is seen. This underlines the importance to prevent and control outbreaks of S. aureus. The aims of this thesis were to increase the knowledge of S. aureus virulence and pathogenesis as well as to understand pattern of colonization and transmission. Various virulence factors operate together in the pathogenic process of S. aureus. The virulence of S. aureus was studied by the interaction with human umbilical vein endothelial cells (HUVEC) as a model. In paper I, we found that one bacterial isolate survived intracellularly and that 156 genes were differentially regulated in microarray analysis of HUVEC. The major part of these genes coded for proteins involved in innate immunity. In paper II, we wanted to explore possible differences in global gene expression patterns in HUVEC induced by invasive compared to colonizing isolates of S. aureus. We also used microarray to investigate possible differences in the presence of virulence genes between the two groups. The main finding was that virulent and commensal S. aureus did not differ in interaction with HUVEC and in the presence of virulence genes. All isolates survived intracellularly for days. Since no obvious differences in virulence between the two groups of isolates were found, we focused on epidemiology and transmission patterns. Colonization with S. aureus is an important risk factor for subsequent S. aureus infection. In paper III, we investigated S. aureus colonization and transmission among nursing home residents in three regions in the south of Sweden and used staphylococcal protein A (spa) typing as an epidemiological tool. A diverse distribution of different spa types was found and a majority of types were unique to one individual. Interestingly, we found a local accumulation of one spa type in one nursing home. Also common spa types were equally distributed in the different regions. We also noted that some individuals were colonized with two different spa types of S. aureus and in five of these cases there was one resistant and one non-resistant strain. The issue of multiclonal colonization and infection is highly important and clinical diagnostic laboratories do not routinely address this problem.

Abstract
2
Therefore, in paper IV a novel method to assess multiclonality of S. aureus was developed. It was based on denaturing gradient gel electrophoresis with the amplification of the spa gene. The method simultaneously separated eight different spa types. It also detected two spa types in an outbreak. In conclusion, we found no differences in virulence genes and in the interaction with HUVEC between commensal and invasive isolates. This indicates that any isolate of S. aureus might have a pathogenic potential. We also confirmed that some spa types are more successful colonizers with a potential to nosocomial spread. The method for detection of multiclonality of S. aureus is of importance in future epidemiological and clinical studies.

Sammanfattning på svenska
3
SAMMANFATTNING PÅ SVENSKA Staphylococcus aureus är en bakterie som koloniserar cirka 30 procent av populationen. Den är dessutom en vanlig orsak till både hud och sårinfektioner men kan även orsaka livshotande tillstånd som lunginflammation, endokardit och sepsis. Vissa stammar kan också orsaka toxinmedierade tillstånd. Därutöver är S. aureus också en av de vanligaste orsakerna till vårdrelaterade infektioner i världen och en ökning av antibiotikaresistens kan ses, särskilt mot meticillin. Detta understryker vikten av att förebygga och bekämpa utbrott av S. aureus. Syftet med denna avhandling var att öka kunskapen om virulens och patogenes hos S. aureus samt att öka förståelsen för kolonisering och smittspridningsmönster. Olika virulensfaktorer bidrar till S. aureus förmåga att orsaka infektion och det är troligt att flera olika faktorer samverkar i den sjukdomsframkallande processen. Virulensegenskaper hos S. aureus studerades i en interaktionsmodell med humana endotelceller från navelsträngar (HUVEC). I den första studien fann vi att bakterien överlevde intracellulärt och med microarray detekterades 156 olika gener som var reglerade i HUVEC. En stor del av dessa gener kodade för proteiner med anknytning till det medfödda immunförsvaret. I den andra studien ville vi undersöka om det fanns skillnader i genuttryck i HUVEC som infekterats med invasiva jämfört med koloniserande isolat av S. aureus. Vi använde också microarray för att studera eventuella skillnader i förekomsten av virulensgener mellan de två grupperna. Den viktigaste slutsatsen var att virulenta och koloniserande isolat av S. aureus inte skiljde sig i interaktionen med HUVEC och inte heller i förekomst av virulensgener. Alla isolaten överlevde intracellulärt under flera dagar.
Eftersom inga uppenbara skillnader i virulens mellan de två grupperna kunde påvisas, fokuserade vi istället på epidemiologi och smittspridningsmönster. Kolonisation med S. aureus är en viktig riskfaktor för att drabbas av S. aureus infektion. I studie tre undersökte vi hur S. aureus koloniserar och eventuellt sprids bland äldre individer på ett antal äldreboenden i tre regioner i södra Sverige. Som ett epidemiologiskt verktyg användes typning av stafylokockprotein A (spa). En stor diversitet i fördelning av olika spa typer hittades och en majoritet av typerna var unika för en enskild individ. Ett

Sammanfattning på svenska
4
intressant fynd var också en lokal ansamling av en spa typ på ett av äldreboendena. De vanligaste spa typerna påträffades oftast i mer än en av regionerna. Vi noterade också att vissa individer var koloniserade med två olika spa typer av S. aureus och fem av dessa individer var samtidigt koloniserade med en resistent och en icke-resistent stam.
Frågan om multiklonal kolonisering och infektion är mycket viktig och kliniska laboratorier är i dagsläget inte rutinmässigt medvetna om detta problem. I den fjärde studien utvecklade vi därför en ny metod för att kunna studera multiklonalitet hos S. aureus. Den baseras på denaturerande gradient gel elektrofores med amplifiering av spa genen. Metoden kunde samtidigt separera åtta olika spa typer och användes också för att studera ett lokalt MRSA utbrott med två olika stammar.
Sammanfattningsvis fann vi inga skillnader i förekomst av virulensgener eller i interaktion med HUVEC mellan koloniserande och invasiva isolat. Detta indikerar att varje isolat av S. aureus kan ha potential att orsaka sjukdom. Vi bekräftade också att vissa spa typer är mer framgångsrika vid kolonisation med potential till vårdrelaterad spridning. Metoden för detektion av multiklonalitet hos S. aureus är av betydelse i framtida epidemiologiska och kliniska studier.

List of papers
5
LIST OF PAPERS I. Andreas Matussek, Jan Strindhall, Lisa Stark, Manfred Rohde,
Robert Geffers, Jan Buer, Erik Kihlström, Per-Eric Lindgren and Sture Löfgren. Infection of human endothelial cells with Staphylococcus aureus induces transcription of genes encoding an innate immunity response. Scand J Immunol; 2005, 61: 536–544
II. Lisa Stark, Andreas Matussek, Jan Strindhall, Robert Geffers, Jan
Buer, Erik Kihlström, Stefan Monnecke, Sture Löfgren and Per-Eric Lindgren. Staphylococcus aureus isolates from blood and anterior nares induce similar innate immune responses in endothelial cells. APMIS; 2009, 117: 814–824
III. Lisa Stark, Magnus Olofsson, Sture Löfgren, Sigvard Mölstad, Per-Eric Lindgren and Andreas Matussek. Prevalence and molecular epidemiology of Staphylococcus aureus in Swedish nursing homes – as revealed in the SHADES study. Epidemiology and Infection; 2013 In press (doi:10.1017/S0950268813002033)
IV. Andreas Matussek,*Lisa Stark,* Olaf Dienus, Joakim Aronsson, Sara
Mernelius, Sture Löfgren and Per-Eric Lindgren. Analyzing multiclonality of Staphylococcus aureus in clinical diagnostics using spa-based denaturing gradient gel electrophoresis. Journal of Clinical Microbiology; 2011, 49: 3647–3648
* The authors have contributed equally to this work
All papers are reprinted with kind permission from the respective publisher
Paper I. © John Wiley and Sons Paper II. © John Wiley and Sons Paper III. © Cambridge University Press Paper IV. © American Society for Microbiology

Abbreviations
7
ABBREVIATIONS bp base pair
BURP the algorithm based upon repeat patterns
Cbp collagen binding protein
CC clonal complex
CFU colony forming unit
Clf clumping factor
CPS capsular polysaccharide
Ct threshold cycle
DGGE denaturing gradient gel electrophoresis
ds double-stranded
E efficiency
Eap extracellular adherence protein
ECM extracellular matrix
ECs endothelial cells
Efb extracellular fibrinogen binding protein
ELISA enzyme-linked immunosorbent assay
ET exfoliative toxin
FBP fibrinogen binding protein
FC fold change
FITC fluorescein isothiocyanate
FnBP fibronectin binding protein
Hlg gamma hemolysine
HUVEC human umbilical vein endothelial cells
ICAM intercellular adhesion molecule
Ig immunoglobulin
IL interleukin
LFA lymphocyte function-associated antigen
LPS lipopolysaccharide
Luk leucocidine
MLST multilocus sequence typing
MRSA methicillin resistant Staphylococcus aureus
MSCRAMMs microbial surface components recognizing adhesive matrix molecules

Abbreviations
8
MSSA methicillin sensitive Staphylococcus aureus
NK-cells natural killer cells
PAMPs pathogen-associated molecular patterns
PBP penicillin binding protein
PFGE pulsed field gel electrophoresis
PRRs pattern recognition receptors
PVL Panton Valentine leukocidin
RFLP restriction fragment length polymorphism
RT revers transcription
SERAMS secreted expanded repertoire adhesive molecules
Spa staphylococcal protein A
SSCP single strand conformation polymorphism
ST sequence type
TGGE temperature gradient gel electrophoresis
TLR toll-like receptor
TNF tumour necrosis factor
TNFR tumour necrosis factor receptor
tRFLP terminal restricted fragment length polymorphism
TSS toxic shock syndrome
VCAM vascular cell adhesion molecule
WGS whole genome sequencing
vWbp von Willebrand binding protein

Introduction
9
INTRODUCTION Staphylococcus aureus
Background Staphylococcus aureus belongs to the family Micrococcaceae and is part of the genus Staphylococcus, which contains more than 30 species such as S. epidermidis, S. saprophyticus and S. haemolyticus. Among the staphylococcal species, S. aureus is by far the most virulent and pathogenic for humans. S. aureus is a 1 μm, Gram-positive cell that in the laboratory may be observed as single cells, in pairs or as grape-like irregular clusters. It is characterized as coagulase- and catalase positive, non-motile, non-spore-forming and as facultative anaerobic. It grows in yellow colonies on nutrient rich media and is referred to as the yellow staphylococci (Winn Washington 2006). S. aureus was discovered in 1880 by the surgeon Sir Alexander Ogston. He observed grape-like clusters of bacteria when examining a purulent discharge from patients with post-operative wounds during microscopy. He named them staphylé, the Greek expression for a bunch of grapes. In 1884, Rosenbach succeeded in isolating yellow bacterial colonies from abscesses and named them Staphylococcus aureus, “aureus” from the Latin word for golden.
S. aureus has the ability to adapt to different environments and it may colonize the human skin, nails, nares and mucus membranes and may thereby disseminate among recipient host populations via physical contact and aerosols (Lowy, 1998). Colonization with S. aureus is an important risk factor for subsequent S. aureus infection (Wertheim et al., 2004; von Eiff et al., 2001).
S. aureus causes a wide range of infections from a variety of skin, wound and deep tissue infections to more life-threatening conditions such as pneumonia, endocarditis, septic arthritis and septicemia. This bacterium is also one of the most common species in nosocomial infections. However, little is known about the virulence factors behind all these conditions. In addition, S. aureus may also cause food poisoning, scalded-skin syndrome and toxic shock syndrome, through production of different toxins (Winn Washington 2006).

Introduction
10
Epidemiology The prevalence of S. aureus nasal carriage varies in different populations. In a general population, the mean carriage rate is 37% (range 19-55 %) (Kluytmans et al., 1997) but some subpopulations show significantly higher carriage rates, e.g. patients with insulin-dependent diabetes mellitus, patients in dialysis, intravenous drug users, individuals with human immunodeficiency virus and patients with S. aureus skin infections. For example, up to 100 % of patients with atopic dermatitis are colonized (Hoeger et al., 1992; Kluytmans et al., 1997; Monti et al., 1996). S. aureus carriage may be classified into three different groups; persistent carriers (~20 %) presumed to always carry the bacterium; intermittent carriers (~60 %), who sometimes carry the bacterium; and non-carriers (~20 %), presumed never to carry the bacterium (Kluytmans et al., 1997). When comparing populations of different age, the colonization rate in children has been reported to be significantly higher compared to adults (Armstrong-Esther, 1976; Cunliffe, 1949; Melles et al., 2004; Noble et al., 1967; Wertheim et al., 2005a), and in some studies a variation between genders has been observed, both in the rate of colonization (Mernelius et al., 2013a; Olsen et al., 2013) and in the presence of different spa types (Sangvik et al., 2011).
Differences in prevalence between various countries have also been noted. In a recent European study, great variation in the nasal carriage rates was found, the lowest in Hungary (12 %) and the highest (29 %) in Sweden (den Heijer et al., 2013). In a Norwegian study, the same rate (29 %) in Norway as in the general Swedish population has been reported (Olsen et al., 2013).
In one study from Canada, the reported incidence of invasive S. aureus infections was 28.4 cases /100 000 individuals and infections were more common in persons over 65 years and in males (Laupland et al., 2003). In the United States, 0.8 % of all hospital inpatients were diagnosed with an S. aureus infection and these patients had significantly longer stay in hospital, paid higher costs and had a higher risk of death than inpatients without S. aureus infection (Noskin et al., 2005). In Europe the level of bacteraemia caused by methicillin-sensitive S. aureus (MSSA) consistently increased between 2002-2008 (12125-15266) (de Kraker et al., 2012).
Over the years, most of the epidemiological studies on S. aureus have been focused on methicillin resistant S. aureus (MRSA) and the epidemiology of MSSA has so far been little studied.

Introduction
11
Antibiotic resistance At first, penicillin was used to treat S. aureus infections. Soon afterwards, resistance emerged when strains acquired a genetic element coding for β-lactamase production, and today over 80 % of all S. aureus strains are resistant to penicillins. The next drug to be introduced for treating infections with S. aureus was the semisynthetic, penicillinase-resistant penicillin named oxacillin or methicillin, but shortly after its introduction the first isolate with resistance was detected (see text below) (Winn Washington 2006). With the emergence of resistance to the penicillinase-resistant penicillins, the glucopeptide agent vancomycin became the treatment of choice for infections with MRSA, and in 1996 the first isolate with intermediate vancomycin resistance was detected in Japan (Winn Washington 2006). So far, this has not emerged to be a major concern, but the resistance has been detected in different parts of the world and needs to be monitored. Although resistance to methicillin is considered the most important for S. aureus, other types of resistance exist. For example, a fusidic acid-resistant impetigo clone has caused infections around Europe. The antibiotic fusidic acid is used to treat superficial skin infections caused by S. aureus, which include impetigo and atopic dermatitis (Brown and Thomas, 2002), and the substance has been in use since the early 1960s. Despite this, the resistance remained low until the 1990s (Brown and Thomas, 2002). Through the last decade an increase in prevalence of fusidic acid-resistant S. aureus has been seen in northern Europe, and this resistance has been primarily associated with strains causing impetigo bullosa (O'Neill et al., 2004; Osterlund et al., 2002; Tveten et al., 2002). The resistance is a consequence of the recruitment of the fusB gene (O'Neill and Chopra, 2006; O'Neill et al., 2004). Since fusidic acid is the primary treatment for impetigo in many countries, this is likely to be the reason for the success of this clone in causing disease. The management with antibiotic-resistant bacteria of infections suffered by the elderly living in nursing homes is something to take into consideration now and in the future. For example, MRSA has become endemic in hospitals as well as in health care settings globally (Chambers and Deleo, 2009; DeLeo and Chambers, 2009). Many nursing home residents have chronic and multiple diseases, and therefore generally require constant medical care and significant assistance with daily living. This causes the residents to be considered as unintentional vectors disseminating pathogens between hospitals and nursing homes but also the other way around (Bonomo, 2000; Chamchod and Ruan, 2012). So far this does not seem to be the case in Sweden, where the resistance in general is

Introduction
12
low and no increase in resistance has been detected for S. aureus isolated from nursing home residents (Olofsson et al., 2012; Olofsson et al., 2013). For over 10 years, Swedish laboratories have reported results on the resistance of S. aureus and other bacteria to a national database ResNet (http://www.srga.org/ResNet_sok.htm). For S. aureus, information about resistance to cefoxitin, erytromycin, clindamycin, fusidic acid, gentamicin and norfloxacin has been registered. The rates for 2010 and 2011 are shown in Figure 1. When comparing antibiotic resistance between European countries, Sweden is among those with the lowest rates and France, Belgium and Austria are those with high resistance for some of the agents (den Heijer et al., 2013).
Figure 1. Resistance of S. aureus in Sweden among isolates from skin and wound infections. The information is taken from the Swedish Institute for Communicable Disease Control. (Statistik för Staphylococcus aureus. www.smittskyddsinstitutet.se/statistik/staphylococcus-aureus/ [2013-06-19]).

Introduction
13
Methicillin-resistant Staphylococcus aureus The massive consumption of antibiotics over the past 50 years has led to the selection of drug-resistance among S. aureus strains, and by far the most important is the resistance against methicillin. In 1961, methicillin (celbenin) became available for treatment of penicillin-resistant S. aureus strains. Only six months thereafter, the first methicillin-resistant S. aureus was detected and nosocomial infections began to increase, and in Sweden efforts to combat the spread were established. In the 1980s the detection of MRSA isolates suddenly increased, and a few strains began to expand worldwide (Chen et al., 2012). MRSA is now a leading cause of nosocomial infections worldwide and has also emerged as a community-associated pathogen (Chambers and Deleo, 2009). MRSA strains are inherently cross-resistant to virtually all beta-lactam antibiotics, the most effective and widely used class of antimicrobials. Moreover, in many countries clinical strains are quite often multi-resistant, which significantly reduces the therapeutic options for treatment of staphylococcal infections (Oliveira and de Lencastre, 2011). The resistance mechanism against methicillin involves the acquisition of the mecA gene, which is a determinant of a unique penicillin binding protein, (PBP)2a, that has reduced affinity for β-lactames, including cephalosporins (Hartman and Tomasz, 1981; Song et al., 1987). The expression of PBP2a causes resistance to all β-lactam antibiotics as the protein blocks binding at the active site for β-lactams (Fuda et al., 2005a; Fuda et al., 2005b). mecA is inserted in a large heterologous chromosomal cassette, the SCCmec element (Ito et al., 1999). In the first international molecular epidemiological study of MRSA, it was discovered that only a few MRSA lineages were responsible for MRSA infections in hospitals located in Europe, the USA and the Far East (Oliveira et al., 2002). Later studies have confirmed these results (Enright et al., 2002). In a recent European study, the prevalence of MRSA, in blood stream infections varied between 0.5 and 30.2 % in the different participating countries (ECDC). This was in contrast to the low prevalence of MRSA in the general healthy population, where the rates did not exceed 2.1 % (den Heijer et al., 2013). To prevent further spread of MRSA in Sweden, a nationwide surveillance program was launched. All hospitalized patients at risk of carriage of MRSA (i.e. known carriage of MRSA, hospital care outside the Nordic countries, or hospital care in connection with an ongoing outbreak) are screened for the presence of MRSA and other multi-resistant bacteria. Confirmed carriers of MRSA must be isolated and contact tracing is performed around this individual.

Introduction
14
All new cases of MRSA are reported to the Department for Control of Communicable Diseases in the county as well as to the Swedish Institute for Communicable Disease Control (since 2000) where the newly discovered isolates must be sent for national epidemiological surveillance.
Virulence factors and strategies Various virulence factors contribute to the ability of S. aureus to cause infection (Figure 2); enzymes (Table 1), toxins (Table 2), adhesion proteins, cell-surface proteins, factors that help the bacteria to evade the innate immune defense, and antibiotic resistance mediate survival of the bacteria and tissue invasion at the site of infection (Zecconi and Scali, 2013). Moreover, certain toxins cause specific disease entities.
Figure 2. A selection of Staphylococcus aureus virulence factors.

Introduction
15
In the case of severe S. aureus disease, the infection may not be explained by the action of a single virulence factor, and it is likely that a number of different factors operating together in the pathogenic process. This assumption is supported by studies in animal models where the infection caused by a mutant isolate, deficient in a single virulence determinant, is compared with the infection caused by the wild type strain. These studies have indicated a decrease in severity of the infection (Hienz et al., 1996; Moreillon et al., 1995). The survival of S. aureus in the host is important for pathogenesis. The bacteria may be protected by a polysaccharide capsule that inhibits opsonization by complement and thereby escapes phagocytosis (O'Riordan and Lee, 2004). It may also secrete cytolytic toxins and tissue-cleaving enzymes (Dinges et al., 2000). Moreover, S. aureus may expresse a multitude of adhesion factors that mediate interactions with host cells and extracellular matrix (ECM), allowing efficient colonization (Chavakis et al., 2005; Foster and Hook, 1998). S. aureus has developed strategies against the antimicrobial peptides, the complement system, and the recruitment and actions of phagocytes (Chavakis et al., 2007) all of which are strategies against the innate immune response of the host (Foster, 2005; Rooijakkers et al., 2005).

Tabl
e 1.
Sel
ectio
n of
com
mon
enz
ymes
rega
rded
as S
. aur
eus v
irule
nce
fact
ors.
Viru
lenc
e fa
ctor
En
zym
atic
func
tion
Effe
ct a
s viru
lenc
e fa
ctor
in h
ost
Refe
renc
e
Cata
lase
De
activ
ates
free
hyd
roge
n pe
roxi
de
Ha
s bee
n sh
own
to b
e es
sent
ial f
or n
asal
co
loni
zatio
n (C
hava
kis e
t al.,
20
07; C
osgr
ove
et
al.,
2007
) Co
agul
ase
Bi
nds t
o pr
otro
mbi
n an
d th
ereb
y be
com
es
enzy
mat
ical
ly a
ctiv
e Ca
taly
zes t
he c
onve
rsio
n of
fibr
inog
en to
fibr
in
Coat
ing
the
bact
eria
with
fibr
in a
nd m
akes
them
re
sista
nt to
ops
oniza
tion
and
phag
ocyt
osis
(Kaw
abat
a et
al.,
19
86)
Hyal
uron
idas
e De
grad
es h
yalu
roni
c ac
id in
con
nect
ive
tissu
e Hy
drol
yzes
the
intr
acel
lula
r mat
rix o
f aci
d m
ucop
olys
acch
arid
es in
tiss
ue a
nd, t
hus m
ay
act t
o sp
read
the
orga
nism
s to
adja
cent
are
as
in ti
ssue
May
con
vert
loca
l tiss
ue in
to n
utrie
nts r
equi
red
for
bact
eria
l gro
wth
(D
inge
s et a
l., 2
000;
W
inn
Was
hing
ton
2006
)
Nuc
leas
e Ex
onuc
leas
e an
d en
donu
clea
se a
ctiv
ity
Cont
ribut
es to
eva
sion
of n
eutr
ophi
l ext
race
llula
r tr
aps
May
deg
rade
hos
t tiss
ue in
to n
utrie
nts r
equi
red
for
bact
eria
l gro
wth
(Ber
ends
et a
l.,
2010
; Che
ung
et a
l.,
2004
; Din
ges e
t al.,
20
00)
Prot
ease
De
grad
es h
uman
fibr
onec
tin, f
ibrin
ogen
and
ki
nino
gen
Clev
es h
uman
α1-
prot
ease
inhi
bito
r, th
e he
avy
chai
n of
all
hum
an im
mun
oglo
bulin
cl
asse
s and
ela
stin
May
con
trib
ute
to th
e ab
ility
of S
. aur
eus t
o di
ssem
inat
e in
hos
t Ai
ds in
tiss
ue in
vasio
n
(Imam
ura
et a
l.,
2005
; Mas
simi e
t al.,
20
02; P
otem
pa e
t al
., 19
86; P
roke
sova
et
al.,
199
2)
16

Sele
ctio
n of
com
mon
enz
ymes
rega
rded
as
S. a
ureu
s viru
lenc
e fa
ctor
s (co
ntin
ued)
.
Stap
hylo
kina
se
Plas
min
ogen
act
ivat
or th
at c
onve
rts
plas
min
ogen
to a
serin
e pr
otea
se, p
lasm
in
Mor
e th
an 6
7% o
f S. a
ureu
s str
ains
exp
ress
th
e ge
ne fo
r sta
phyl
okin
ase
Neu
tral
izes t
he b
acte
ricid
al e
ffect
by
form
ing
com
plex
with
α-d
efen
sin.
May
cle
ave
com
plem
ent f
acto
r C3
Co
ntro
ls fib
rinol
ysis
Th
e ba
cter
ia e
xplo
it th
e pr
oteo
lytic
act
ivity
of
plas
min
to d
egra
de c
o mpo
nent
s of E
CM a
s wel
l as
fibrin
ogen
for d
issem
inat
ion
in th
e ho
st
(Cha
vaki
s et a
l.,
2007
; Koc
h et
al.,
20
12; L
ahte
enm
aki
et a
l., 2
000 ;
Ro
oija
kker
s et a
l.,
2005
; Zec
coni
and
Sc
ali,
2013
)
17

Tabl
e 2.
Sel
ectio
n of
exo
toxi
ns re
gard
ed a
s viru
lenc
e fa
ctor
s of S
. aur
eus.
Viru
lenc
e fa
ctor
Fu
nctio
n Vi
rule
nce
effe
ct o
n ho
st
Refe
renc
es
Exfo
liativ
e to
xins
Glut
amat
e-sp
ecifi
c se
rine
prot
ease
s tha
t di
gest
des
mog
lein
1, a
ker
atin
ocyt
e ce
ll-ce
ll ad
hesio
n m
olec
ule.
Ex
folia
tive
toxi
ns (E
Ts) a
ct a
s “m
olec
ular
sc
issor
s” fa
cilit
atin
g ba
cter
ial s
kin
inva
sion
Prev
alen
ce o
f eta
and
/or e
tb ra
nge
from
0.5
-3
% in
MSS
A bu
t 10
% o
f MRS
A st
rain
s hav
e be
en fo
und
to b
e et
a po
sitiv
e
The
ETA
and
ETB
are
the
two
mos
t im
port
ant
isofo
rms a
nd th
ey a
re a
ssoc
iate
d w
ith
stap
hylo
cocc
al b
ullo
us im
pert
igo
and
stap
hylo
cocc
al
scal
ded
skin
synd
rom
e
ETA
ETB
ETC
(not
ass
ocia
ted
with
hum
an d
iseas
e)
and
ETD
Med
iate
supe
rant
igen
act
ivity
(Bec
ker e
t al.,
200
3;
Kato
et a
l., 2
011;
Pe
acoc
k et
al.,
200
2;
Sila
et a
l., 2
009;
Ze
ccon
i and
Sca
li,
2013
)
Hem
olys
ins
Pore
form
ing
toxi
n w
ith c
ytol
ytic
effe
ct o
n er
ythr
ocyt
es a
nd m
onoc
ytes
(α–t
oxin
) Cy
toly
tic a
ctiv
ity o
n cy
toki
ne c
onta
inin
g ce
lls
(β –
hem
olys
in a
lso k
now
n as
sp
hing
omye
linas
e C)
N
eutr
ophi
l and
mon
ocyt
e bi
ndin
g
(δ –
hem
olys
in)
The
vast
maj
ority
of t
he h
emol
ysin
s are
hem
olyt
ic
α-to
xin
has p
ro-in
flam
mat
ory
prop
ertie
s on
host
(Cha
vaki
s et a
l.,
2007
; Zec
coni
and
Sc
ali,
2013
)
18
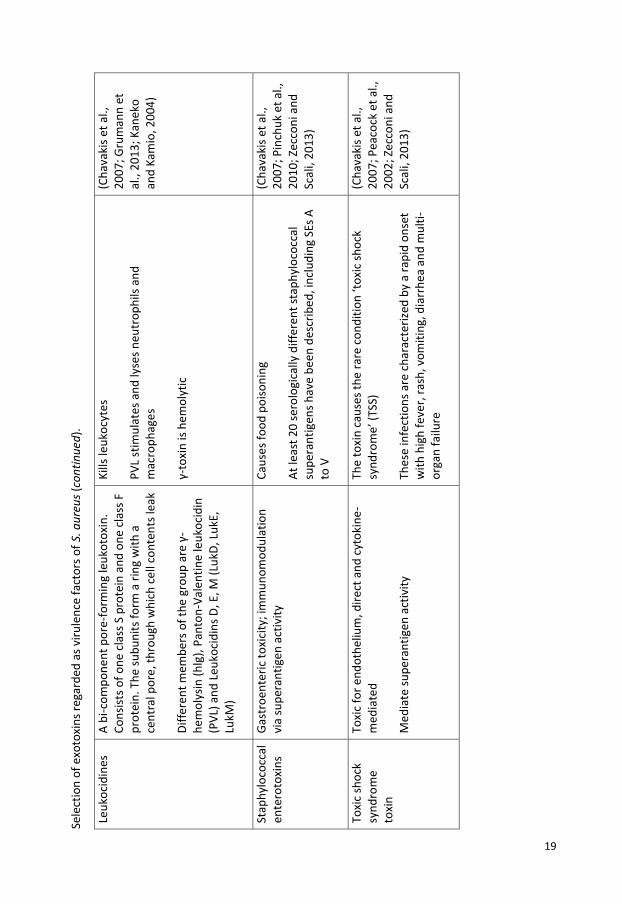
Sele
ctio
n of
exo
toxi
ns re
gard
ed a
s viru
lenc
e fa
ctor
s of S
. aur
eus (
cont
inue
d).
Leuk
ocid
ines
A bi
-com
pone
nt p
ore-
form
ing
leuk
otox
in.
Cons
ists o
f one
cla
ss S
pro
tein
and
one
cla
ss F
pr
otei
n. T
he su
buni
ts fo
rm a
ring
with
a
cent
ral p
ore,
thro
ugh
whi
ch c
ell c
onte
nts l
eak
Di
ffere
nt m
embe
rs o
f the
gro
up a
re γ
-he
mol
ysin
(hlg
), Pa
nton
-Val
entin
e le
ukoc
idin
(P
VL) a
nd L
euko
cidi
ns D
, E, M
(Luk
D, L
ukE,
Lu
kM)
Kills
leuk
ocyt
es
PVL
stim
ulat
es a
nd ly
ses n
eutr
ophi
ls an
d m
acro
phag
es
γ-to
xin
is he
mol
ytic
(Cha
vaki
s et a
l.,
2007
; Gru
man
n et
al
., 20
13; K
anek
o an
d Ka
mio
, 200
4)
Stap
hylo
cocc
al
ente
roto
xins
Ga
stro
ente
ric to
xici
ty; i
mm
unom
odul
atio
n vi
a su
pera
ntig
en a
ctiv
ity
Caus
es fo
od p
oiso
ning
At
leas
t 20
sero
logi
cally
diff
eren
t sta
phyl
ococ
cal
supe
rant
igen
s hav
e be
en d
escr
ibed
, inc
ludi
ng S
Es A
to
V
(Cha
vaki
s et a
l.,
2007
; Pin
chuk
et a
l.,
2010
; Zec
coni
and
Sc
ali,
2013
)
Toxi
c sh
ock
synd
rom
e to
xin
Toxi
c fo
r end
othe
lium
, dire
ct a
nd c
ytok
ine-
med
iate
d M
edia
te su
pera
ntig
en a
ctiv
ity
The
toxi
n ca
uses
the
rare
con
ditio
n ‘to
xic
shoc
k sy
ndro
me ’
(TSS
) Th
ese
infe
ctio
ns a
re c
hara
cter
ized
by a
rapi
d on
set
with
hig
h fe
ver,
rash
, vom
iting
, dia
rrhe
a an
d m
ulti-
orga
n fa
ilure
(Cha
vaki
s et a
l.,
2007
; Pea
cock
et a
l.,
2002
; Zec
coni
and
Sc
ali,
2013
)
19

Introduction
20
Adhesion to host S. aureus expresses several adhesion factors that mediate interactions with host cells and ECM, and that allow efficient colonization (Figure 3) (Chavakis et al., 2005; Foster and Hook, 1998). More than one staphylococcal adhesin usually recognizes a single host ECM component. These adhesins can be classified into two groups: “microbial surface components recognizing adhesive matrix molecules” (MSCRAMMs) (Table 3), and “secreted expanded repertoire adhesive molecules” (SERAMS) (Table 4). The major components of MSCRAMMs, a family constituted of more than 20 members, are fibronectin-binding proteins (FnBPs), clumping factors (Clfs), staphylococcal protein A (SpA) and collagen-binding protein (Cbp) (Chavakis et al., 2007; Chavakis et al., 2005; Foster and Hook, 1998; Mazmanian et al., 2000). SERAMs are a group of five structurally unrelated secreted adhesins, including the fibrinogen-binding protein (FBP) A, the coagulase (Coa), the extracellular fibrinogen-binding protein (Efb), the ECM-binding protein and the extracellular adherence protein (Eap) (Chavakis et al., 2005), which interact with a broad array of host ligands, thereby mediating bacteria adhesion but also interfering with host defense mechanisms (Chavakis et al., 2005).
Figure 3. Example of factors involved in the interaction between S. aureus and the host cells.

Tabl
e 3.
Sel
ectio
n of
S. a
ureu
s viru
lenc
e fa
ctor
s inv
olve
d in
adh
esio
n to
hos
t cel
ls, M
SCRA
MM
s.
Viru
lenc
e fa
ctor
Fu
nctio
n Ef
fect
on
host
Re
fere
nces
Clum
ping
- fa
ctor
Plat
elet
adh
esio
n (fi
brin
-med
iate
d)
ClfA
is a
fibr
inog
en-b
indi
ng su
rfac
e pr
otei
n It
may
act
ivat
e se
rum
com
plem
ent
regu
lato
r fac
tor I
and
con
vert
C3b
to iC
3b
and
C3d
ClfA
bin
ds c
ompl
emen
t reg
ulat
or fa
ctor
I
ClfB
bin
ds to
cyt
oker
atin
10
Viru
lenc
e is
likel
y to
be
incr
ease
d by
bac
teria
l cel
ls be
com
ing
coat
ed w
ith fi
brin
ogen
whi
ch in
hibi
ts d
epos
ition
of o
r acc
ess t
o op
soni
ns
Inac
tivat
ion
of c
ompl
emen
t fac
tors
also
resu
lting
in th
e lo
ss o
f th
e ef
fect
of t
he o
pson
in.
ClfA
mut
ants
are
sign
ifica
ntly
att
enua
ted
in m
urin
e m
odel
s for
se
psis
and
arth
ritis
and
in a
rat e
ndoc
ardi
tis m
odel
(Jose
fsso
n et
al
., 20
08;
Mor
eillo
n et
al
., 19
95)
Colla
gen-
bind
ing
prot
ein
Adhe
sin fo
r col
lage
n ty
pes I
and
IV
Cont
ribut
es to
the
deve
lopm
ent o
f sep
tic a
rthr
itis
It ha
s bee
n sh
own
that
col
lage
n is
impo
rtan
t in
endo
card
itis,
no
t in
the
initi
al p
roce
ss b
ut in
the
late
r sta
te
(Cha
vaki
s et
al.,
2007
; Pe
acoc
k et
al.,
20
02)
Fibr
onec
tin-
bind
ing
prot
ein
Adhe
sins f
or fi
brin
ogen
(FnB
PA o
nly)
, fib
rone
ctin
and
ela
stin
Ad
hesin
for n
on-p
rofe
ssio
nal p
hago
cyte
s suc
h as
epi
thel
ial a
nd
endo
thel
ial c
ells,
fibr
obla
sts,
ost
eobl
asts
and
ker
atin
ocyt
es
(Cha
vaki
s et
al.,
2007
; Flo
ck
et a
l., 1
987;
Pe
acoc
k et
al.,
20
02)
21

22
Se
lect
ion
of S
. aur
eus v
irule
nce
fact
ors i
nvol
ved
in a
dhes
ion
to h
ost c
ells,
MSC
RAM
Ms
(con
tinue
d).
Stap
hylo
cocc
al
prot
ein
A
Bind
s the
Fc-γ
regi
on o
f all
hum
an
imm
unog
lobu
lin (I
g)G
subc
lass
es e
xcep
t Ig
G3
Activ
ates
pla
tele
t agg
rega
tion
via
its
bind
ing
to v
on W
illeb
rand
fact
or
Bind
s com
plem
ent p
rote
in C
3 Bi
nds t
o tu
mou
r nec
rosis
fact
or re
cept
or
(TN
FR)1
, the
rece
ptor
for t
umou
r nec
rosis
fa
ctor
(TN
F)-α
Inhi
bits
pha
gocy
tosis
Bi
nds c
ompl
emen
t pro
tein
C3
and
prom
otes
C3-
C3b
conv
ersio
n
Spa
has b
een
show
n to
initi
ate
stap
hylo
cocc
al p
neum
onia
and
sp
a de
ficie
nt m
utan
ts se
em u
nabl
e to
form
abs
cess
es in
mic
e
Redu
ces T
NF -α
pro-
infla
mm
ator
y sig
nalin
g
(Goo
dyea
r and
Si
lver
man
, 20
03; P
eaco
ck
et a
l., 2
002;
Ro
ben
et a
l.,
1995
; Sm
ith e
t al
., 20
11;
Zecc
oni a
nd
Scal
i, 20
13)

Tabl
e 4.
Sel
ectio
n of
S. a
ureu
s viru
lenc
e fa
ctor
s in
the
grou
p of
secr
eted
exp
ande
d re
pert
oire
adh
esiv
e m
olec
ules
(S
ERAM
s).
Viru
lenc
e fa
ctor
Fu
nctio
n Ef
fect
on
host
Re
fere
nces
Ca
psul
ar
poly
sacc
harid
es
Alte
r C3
(cap
sula
r pol
ysac
char
ides
(C
PS)5
and
8) o
r C3b
(CPS
1) d
epos
ition
M
ask
C3 (C
PS5
and
8) a
nd C
3b (C
PS1)
de
posit
ion
Mic
roor
gani
sms t
hat c
ause
inva
sive
dise
ase
com
mon
ly p
rodu
ce e
xtra
cellu
lar
caps
ular
pol
ysac
char
ides
Caps
ules
enh
ance
mic
robi
al v
irule
nce
by re
nder
ing
the
bact
eriu
m re
sista
nt to
pha
gocy
tosis
and
ther
eby
cont
ribut
e to
th
e ba
cter
ial p
ersis
tenc
e in
the
bloo
dstr
eam
of i
nfec
ted
host
s Fe
wer
com
plem
ent m
olec
ules
are
abl
e to
bin
d to
cap
sula
ted
bact
eria
, whi
ch h
as a
n an
tipha
gocy
tic e
ffect
Th
e S.
aur
eus c
apsu
les a
lso p
rom
otes
abs
cess
(Cha
vaki
s et
al.,
2007
; O
'Rio
rdan
and
Le
e, 2
004)
Coag
ulas
e
von
Will
ebra
nd
fact
or b
indi
ng
prot
ein
Bind
s and
act
ivat
es p
roth
rom
bin
whi
ch
prom
otes
con
vers
ion
of fi
brin
ogen
to
fibrin
Th
e st
aphy
loco
agul
ase-
prot
rom
bin
com
plex
cle
aves
fibr
inog
en
Prom
ote
coag
ulat
ion,
thro
ugh
bind
ing
and
activ
atio
n of
pr
othr
ombi
n Th
e ge
nera
tion
of fi
brin
pep
tides
pro
mot
es c
lot f
orm
atio
n. T
he
fibrin
clo
t inh
ibit
phag
ocyt
osis
by p
reve
ntin
g th
e pe
netr
atio
n of
pha
gocy
tic c
ells
into
the
site
of in
fect
ion
Co
agul
ase
is an
impo
rtan
t fac
tor i
n th
e fo
rmat
ion
of a
bsce
sses
. Ab
sces
s for
mat
ion
and
bloo
d co
agul
atio
n ca
nnot
occ
ur in
m
utan
t mic
e la
ckin
g ge
nes f
or b
oth
Coa
and
von
Will
ebra
nd
bind
ing
prot
ein
(vW
bp)
(Cha
vaki
s et
al.,
2007
; Ch
eng
et a
l.,
2011
; M
cAdo
w e
t al
., 20
12)
23

Sele
ctio
n of
S. a
ureu
s viru
lenc
e fa
ctor
s in
the
grou
p of
secr
eted
exp
ande
d re
pert
oire
adh
esiv
e m
olec
ules
(SER
AMs)
(con
tinue
d).
Extr
acel
lula
r ad
hesio
n pr
otei
n
Bind
s to
plas
ma
prot
eins
such
fibr
inog
en,
fibro
nect
in a
nd p
rotr
ombi
n St
rong
inte
ract
ion
betw
een
Eap
and
the
inte
rcel
lula
r adh
esio
n m
olec
ule
(ICAM
)-1
in v
itro
Impa
irs a
ngio
gene
sis a
nd w
ound
hea
ling
M
ajor
hist
ocom
patib
ility
com
plex
II a
nalo
g pr
otei
n; a
dhes
ion
to S
. aur
eus c
ells
and
host
cel
ls; in
volv
ed in
bio
film
form
atio
n Th
e va
st m
ajor
ity o
f S. a
ureu
s str
ains
seem
to
secr
ete
Eap
but n
o ot
her s
taph
yloc
occi
Bi
nds t
o EC
M m
olec
ules
Bi
nds t
o IC
AM-1
Bl
ocks
the
lym
phoc
yte
func
tion-
asso
ciat
ed
antig
en (L
FA)-1
and
ICAM
-1 in
tera
ctio
n
Enha
nces
the
bind
ing
and
inte
rnal
izatio
n of
S. a
ureu
s int
o eu
kary
otic
cel
ls M
ay in
hibi
t the
bin
ding
of l
euko
cyte
s (T-
cells
) to
activ
ated
en
doth
elia
l cel
ls an
d th
ereb
y in
hibi
t the
ext
rava
satio
n of
le
ucoc
ytes
(mon
ocyt
es a
nd T
-cel
ls) fr
om th
e bl
oods
trea
m in
to
the
site
of in
fect
ion
The
bind
ing
of E
ap to
ICAM
-1 tr
igge
rs th
e re
leas
e of
the
pro-
infla
mm
ator
y cy
toki
nes T
NF-α
and
inte
rleuk
in (I
L)-6
Bl
ocks
neu
trop
hils
and
T ce
ll re
crui
tmen
t In
hibi
ts M
AP k
inas
e ph
osph
oryl
atio
n In
hibi
ts d
elay
ed-t
ype
hype
rsen
sitiv
ity
(Cha
vaki
s et
al.,
2007
; Ed
war
ds e
t al.,
20
12; H
agga
r et
al.,
200
4;
Huss
ain
et a
l.,
2008
; Scr
iba
et
al.,
2008
)
24

Introduction
25
The host Without a functional immune system, we could not resist infections for a long period of time. Many microbes are pathogenic, and we are constantly in danger of infections and diseases caused by them. The immunity may be innate or acquired, and the innate immune recognition has been designed to discriminate non-self from self with a limited number of recognition molecules. In this way, virtually all bacteria, viruses, fungi and protozoa are recognized. To become a pathogen, a bacterium has to be able to survive within a host. This can be achieved by the acquisition of specific adhesion factors, adaption to intracellular survival, or the development of a thick capsule.
Innate immunity The innate immune system, also known as the non-specific immune system or the first line of defense, is based on mechanisms and cells that defend the host from infection caused by different microorganisms. The system acts in an immunologically non-specific way. The innate immune system is available to rapidly protect the host from foreign invaders in contrast to the adaptive immune system, which requires days to weeks to become active for the first time of recognition. The function of the innate immunity includes several factors and mechanisms such as physical and chemical barriers to infectious agents represented by skin and mucosal membranes, as well as recruitment of inflammatory cells to the site of infection through the production of cytokines. Furthermore, other included mechanisms are activation of the complement cascade, the activation of cells for phagocytosis, which clears cell debris and antigen-antibody complexes, and the activation of the adaptive immune system through antigen presentation by macrophages and dendritic cells (Coico Richard 2009; Doan Thao 2013; Peter, 2006). The physical barriers are the epithelial surfaces of the skin, the epithelium of the mucosa membranes of, for example, the gastrointestinal and respiratory tracts where cilia help remove infectious agents. Also, mucus at different sites of the body is a protection against infectious agents. There are also chemical and environmental barriers such as the acidic pH of the skin, stomach and vagina. Molecules such as defensins, RNases, DNases and lysozyme, secreted by various cell types, also serve as environmental barriers. Another kind of barrier is the biological barrier consisting of commensal microbes. Commensal microbes are those that exist in a symbiotic relationship with the body on the skin and in the gastrointestinal tract. Microbes in these tracts defend their

Introduction
26
territory and inhibit the establishment of potentially pathogenic microbes (Doan Thao 2013). The complement system, also a part of the innate immune system, is a biochemical cascade that supports clearing of pathogens. The cascade is composed of a series of pro-enzymes and related factors that sequentially activate each other, resulting in the production of a variety of biologically active proteins. The proteins trigger the recruitment of inflammatory cells, and "tag" pathogens for destruction by other cells by opsonizing, or coating the surface of the pathogen. Some of the proteins also increase vascular permeability. Cells belonging to the innate immune system include: natural killer (NK) cells, mast cells, eosinophils, basophils and phagocytic cells, such as neutrophils, macrophages and dendritic cells. These cells collectively perform two important functions; they are able to recognize the presence of an unknown invader, for example, a bacterial infection, and they provide an immediate cellular response to the presence of an infectious agent. The nature of this cellular response differs according to the nature of the cell and the way in which it is stimulated (Peter, 2006). The innate immune system recognizes the presence of pathogens by different receptors on cells belonging to this defense system, and these cell surface receptors are called pattern recognition receptors (PRRs). These receptors recognize molecules that are mostly shared by pathogens but distinguishable from host molecules, collectively referred to as pathogen-associated molecular patterns (PAMPs). One important family of PRRs is the toll-like receptors (TLR), which recognize a wide variety of pathogen-associated molecules. Different TLRs are present on various immunological cells and they identify microbial products such as double-stranded RNA found in viral infections, lipopolysaccharide (LPS) present in Gram-negative bacteria, bacterial lipoproteins, unmethylated DNA, and flagellin. After the recognition, a response to eliminate or limit the spread of the pathogen within the host is started with activation of phagocytosis and cytokine production to stimulate other cells (Doan Thao 2013). At the beginning of an infection, one of the PRRs recognizes a PAMP which leads to the activation of the cell, thereby releasing inflammatory mediators responsible for the clinical signs of inflammation such as pain, swelling, redness and heat (Coico Richard 2009; Peter, 2006). This process is stimulated by chemical factors released by affected cells. The aim of the inflammatory response is to recruit cells which release factors into the bloodstream from surrounding tissue to assist in the remove of pathogens, dead cells or damaged

Introduction
27
tissue. Four important events occur during an inflammatory response to promote this aim; vasodilation, activation of endothelial cells, increased vascular permeability and production of chemotactic factors (Coico Richard 2009; Peter, 2006). Vasodilation of the blood vessels increases the blood flow at the inflammatory site, increasing the supply of cells and other factors to the area. Activation of endothelial cells leads to increased expression of cell adhesion molecules, promoting the migration of leucocytes from blood to tissue. Increased vascular permeability makes it easier for cells and proteins to pass through the blood vessel wall and enter the tissue. Chemotactic factors are produced that attract cells into the tissue from the blood stream (Coico Richard 2009; Peter, 2006). During inflammation, mast cells release chemical factors such as histamine, bradykinin, serotonin, leukotrienes, and prostaglandins. These factors are responsible for sensitizing pain receptors, cause prolonged vasodilation of the blood vessels, and attract phagocytes, especially neutrophils. Neutrophils will then trigger other parts of the immune system by releasing factors that recruit other leukocytes. Cytokines are produced by macrophages and other cells of the innate immune system and mediate the inflammatory response (Figure 4) (Lotze and Tracey, 2005).

Introduction
28
Figure 4. Overview of the innate immune response associated with a bacterial invasion. (Modified picture from TRENDS in Immunology)
Endothelial cells The endothelium consists of a thin layer of cells that lines the inner surface of blood vessels and lymphatic vessels. It is an interface between circulating blood or lymph in the lumen and the rest of the vessel wall. The cells that form the endothelium are called endothelial cells (ECs). ECs in direct contact with blood are called vascular ECs, whereas those in direct contact with lymph are known as lymphatic ECs. The EC surface in an adult human is composed of approximately 1 x 1013 to 6 x 1013 cells and it covers a surface of approximately 1 to 7 m2. ECs line vessels in every organ system and regulate the flow of nutrients, biologically active molecules and all kinds of different blood cells. The endothelium acts through the presence of membrane-bound receptors for different kinds of molecules including proteins, as well as through specific junctional proteins and receptors that control cell-cell and cell-matrix interactions (Cines et al., 1998).

Introduction
29
ECs are involved in many aspects of vascular biology. They act as a semi-selective barrier between the vessel lumen and surrounding tissue, they control the passage of different kinds of chemical factors, and they are central in the extravasation of white blood cells from the bloodstream. Furthermore, the endothelium has a function in coagulation. ECs normally provide a non-thrombogenic surface because they contain, for example, heparan-sulfate which acts as a cofactor for activating antithrombin, a protease that inactivates several factors in the coagulation cascade. Another important function of the ECs is their role in cell migration from the blood stream to the surrounding tissue. This is a strictly controlled process so that the cells only go where they are required. One important factor for controlling this migration is adhesion molecules, present on leucocytes and ECs, and it is the interaction between these cells that enables the migration of leucocytes across the endothelium. There are many different adhesion molecules which are represented by four families; selectins (leucocytes and ECs), integrins (leucocytes), mucin-like vascular addressins (leucocytes and some ECs), and members of the immunoglobulin superfamily (expressed on ECs). Different adhesion molecules bind to each other in a specific manner, which leads to a cell-cell interaction between leucocytes and ECs. Adhesion molecules are expressed on different cell types; some are expressed constantly on the cell surface and others are induced by cell activation, primarily by cytokines. The process by which cells leave the bloodstream and cross the endothelium to enter into various tissues is called extravasation. Extravasation can be divided into three stages; rolling, activation and adhesion. In case of an inflammation, vasodilatation occurs, leading to slower blood flow, and thereby facilitates the leucocyte migration. Inflammatory mediators such as TNF-α activate ECs and stimulate the expression of E-selectin and P-selectin on the surface of ECs. These molecules bind to sialyl-Lewisx on neutrophils and contribute to the rolling. When the leucocytes have started to roll they have become ready to adhere strongly to the ECs by release of IL-8 from ECs. This procedure is performed with the binding between LFA-1 on the surface of neutrophils and ICAM-1 on the ECs. When the leucocyte is firmly attached to the endothelium it squeezes between the ECs, making contact with the basement membrane underneath. This process is poorly understood but involves additional adhesion molecules. In inflamed tissue there will be a gradient of molecules from both microorganisms and immune cells with maximum levels at the center of the infection. The leucocyte will travel against these gradients moving towards the increasing concentrations of molecules such as LPS, complement factor C5a and IL-8 (Peter, 2006).

Introduction
30
Typing methods To prevent and control outbreaks of S. aureus, and for epidemiological investigations, appropriate typing methods are needed. Numerous methods, both phenotypic and genotypic, have been used for the typing of S. aureus. In the past, typing methods were based on phenotype characteristics, such as serotype, biotype, phage-type or antibiogram, of which phage-typing was the most common. Today these methods have been replaced with molecular genotyping. The genotyping methods can be divided into sequence-based (multilocus sequence typing [MLST] and staphylococcal protein A [spa] typing) and non-sequence based (pulsed field gel electrophoresis [PFGE], restriction fragment length polymorphism [RFLP]) (Mulligan and Arbeit, 1991). The various methods differ in requirements of equipment and expertise and are also different in cost-effectiveness. The most used methods for typing of S. aureus are PFGE, regarded as gold standard, MLST and spa typing. The choice of method depends on the problem to be solved and the epidemiological situation in which the method is going to be used, as well as the time and geographical extent of its use (David et al., 2013). Methods based on DNA sequencing are objective and suitable for database analysis. Non-sequence-based methods often involve subjectiv interpretations with comparison of gel patterns and fingerprint images. Those image-based methods do not provide reproducible biological criteria to evaluate the relatedness between different strains, and it is difficult to maintain an international database and retrieval of patterns for comparison between different laboratories (Sabat et al., 2013). International harmonization of typing techniques is important to establish surveillance networks for global epidemiological studies such as for MRSA control. The development of next generation sequencing will, in the near future, enable whole genome sequencing (WGS) even in smaller research and clinical laboratories, and may be used in outbreaks and in epidemiological investigations (Pallen et al., 2010; Sabat et al., 2013). Molecular fingerprinting is the basis of a group of molecular methods aimed at generating a fingerprint of an unknown microbial community or an individual strain and in the past 20 years, several methods based on the direct amplification and analysis of the 16S ribosomal RNA gene have been developed. Methods included in this group are denaturing gradient gel electrophoresis (DGGE), temperature gradient gel electrophoresis (TGGE), single-strand conformation polymorphism (SSCP), and terminal RFLP (tRFLP) (Su et al., 2012). We have adapted one of these methods, DGGE, to distinguish

Introduction
31
different clones of S. aureus based on the spa gene since the 16S rRNA gene is not sufficiently discriminatory. One issue connected to typing of different bacterial isolates is that the concept of a “strain” or a “clone” is not fully defined. (Dijkshoorn et al., 2000; Tenover et al., 1995). The problem is that the definition of a clone or strain depends on the discriminatory power of the test and/or on the number of different tests applied (Monecke et al., 2011). If “type” specified in connection with the method used to define the relatedness between the isolates is used (e.g spa type, MLST type etc.), it will perhaps be less confusing.
Conventional typing method
Phage typing
Until the beginning of the 1990s, phage typing was the main method used for typing of S. aureus to discriminate among strains related to outbreaks. This method is based on virus infecting bacteria called bacteriophages that are able to infect S. aureus, and some of them can only infect a single strain of that species (Bannerman et al., 1995). The bacterial isolate is grown on an agar plate. Inoculation of different phages on the agar surface is then performed. After incubation the susceptible phage regions will show a clearing where the bacteria have been lysed and this is used to differentiate the staphylococcal isolates from each other (Blair and Williams, 1961). This method had several weaknesses such as: characterization of phenotypic markers has poor reproducibility, it is not possible to type all isolates, and due to the large number of phages stocks needed, only a few reference laboratories could use the method (Bannerman et al., 1995). When PFGE, the first molecular method to be used for epidemiological investigations of S. aureus, was compared with phage typing, PFGE was found to be better than phage typing at distinguishing epidemiologically related strains from unrelated strains. In addition, it was found that all strains were typable with PFGE (Bannerman et al., 1995).

Introduction
32
Molecular typing methods
Pulsed field gel electrophoresis
PFGE has so far been regarded as the gold standard for molecular typing of S. aureus. In the 1990s, this method replaced phage typing as a tool in outbreaks and epidemiological investigations and has since become one of the most used methods for typing of different bacteria (Sabat et al., 2013). In PFGE, highly purified genomic DNA from bacterial isolates is digested with rare-cutting restriction enzymes that cut in given sites within the genomic sequence, resulting in fragments of different sizes (50 000-250 000 base pairs [bp]). The restriction fragments are comparable large (hundreds of thousands of bp), which may be separated on an agarose gel by pulse field electrophoresis, in which the orientation of the electric field across the gel is changed periodically. The separated fragments are then visualized as bands on the agarose gel after staining with a DNA binding dye. The bands form a specific band pattern, which may be compared to other patterns from different isolates (Buckingham Lela 2007). PFGE has a high discriminatory power and a wide typeability (high epidemiological concordance) since S. aureus isolates and nearly all strains are typable by this method (Bannerman et al., 1995; Mehndiratta and Bhalla, 2012). This method also addresses a large portion of an investigated genome (90 %) (Buckingham Lela 2007; Sabat et al., 2013). Another advantage is that large fragments are separated, resulting in relatively few bands in the gel, which facilitates the interpretation of the results (Sabat et al., 2013). Criteria for PFGE pattern interpretation which can be used to facilitate the assessment of the results have been produced (Tenover et al., 1997; Tenover et al., 1995). The disadvantages of PFGE are that it is technically demanding, labor-intensive and time-consuming. It may require three to four days to complete one analysis (Bannerman et al., 1995). It is also difficult to distinguish bands of nearly identical size. The evaluation depends on the judgment of the observer and there are difficulties comparing results between different laboratories (Buckingham Lela 2007; Olive and Bean, 1999; Sabat et al., 2013). In inter-laboratory studies the results have shown widely discrepant results (Cookson et al., 1996; Mulvey et al., 2001; van Belkum et al., 1998). PFGE may also overestimate the genetic differences, and it has been reported that PFGE results have insufficient stability in long-term epidemiological surveillance, due to the long evolutionary history of pandemic clones (Hallin et al., 2007; Melles et al., 2007).

Introduction
33
The technique has been compared to other techniques in various studies and has been found useful in the characterization of outbreak strains (Bannerman et al., 1995; Cookson et al., 1996; Hallin et al., 2007; Melles et al., 2007).
Multilocus sequence typing
MLST characterizes bacterial isolates after amplification of different housekeeping genes. It is based on the principles of phenotypic multilocus enzyme electrophoresis (Selander et al., 1986), which depends on the difference in electrophoretic mobility of enzymes present in a bacterium. The first developed MLST method was for typing of Neisseria meningitides (Maiden et al., 1998) and has since then become a popular tool for epidemiological studies of many different bacterial species and for studies of the molecular evolution of different pathogens (Enright et al., 2000; Jones et al., 2003; Maiden et al., 1998; Sabat et al., 2013). The bacterial housekeeping genes are essential and therefore present in the genome of all S. aureus strains. The housekeeping genes provide reliable information about the relationship between isolates, since they are not subjected to selective forces, and thereby change slowly. Mostly, seven housekeeping genes are sequenced over approximately 450-500 bases and the sequences are assigned as alleles. An isolate type, called the sequence type (ST), is a collection of all seven alleles (Figure 5), the profile of the alleles and an MLST clone are defined as a set of isolates with identical sequences at all seven loci (Buckingham Lela 2007; Enright et al., 2000).

Introduction
34
Figure 5. Description of the performance of an MLST analysis.
The advantage of this sequence-based method is that there is a standardized nomenclature and the allele sequences and ST profiles are available in large global databases, facilitating the communication of results between different laboratories. Also, reproducibility is high and primer sequences and protocols may be standardized and are electronically available. The disadvantage is that the technique is expensive, labor-intensive and time consuming, due to include a number of various gene targets (Birtles et al., 2005; Rakeman et al., 2005; Sabat et al., 2013). MLST may be used to investigate evolutionary relationships among bacteria and it provides good discriminatory power to differentiate isolates (Mehndiratta and Bhalla, 2012). The technique has also been used to identify livestock-associated MRSA strains and their transmission between humans and animal species (Cuny et al., 2010). When comparing MLST with other techniques like PFGE and spa typing, a good agreement between results obtained by the different methods has been reported (Cookson et al., 1996; Hallin et al., 2007; Melles et al., 2007; Strommenger et al., 2006).

Introduction
35
spa typing
spa typing represents the group of methods designated single-locus sequence typing, which means that sequence variations of a single target gene are compared. The genes selected are usually of short sequence repeat regions that are sufficiently polymorphic to provide useful resolution (Mehndiratta and Bhalla, 2012). The spa gene of S. aureus harbors a variable number of tandem repeats in the 3´coding polymorphic X region of the protein A gene (Figure 6).
Figure 6. Description of the staphylococcal protein A gene (modified from Current Opinion in Microbiology (Kim et al., 2012)).
The X region consists of repeated units, 21 or 24 bp long. The sequence of these repeated units varies in at least one bp (Table 5a) and to this date, 601 different repeats have been described (http://www.spaserver.ridom.de). The variable part of the X region may have 2-16 repeats. The spa type is deduced from the order of specific repeats and is associated with a figure (Table 5b) (Buckingham Lela 2007; Sabat et al., 2013; Shopsin et al., 1999).

Introduction
36
Table 5a. Sequences of different repeats. Repeat Sequence
r16 AAAGAAGACGGCAACAAACCTGGT
r17 AAAGAAGACGGCAACAAGCCTGGT
r20 AAAGAAGACAACAACAAACCTGGC
r23 AAAGAAGACGGCAACAAACCTGGC
r26 GAGGAAGACAACAAAAAACCTGGT
r34 AAAGAAGACAACAAAAAACCTGGT
Table 5b. Repeat succession of different spa types. spa types Repeat succession
t002 26-23-17-34-17-20-17-12-17-16
t008 11-19-12-21-17-34-24-34-22-25
t015 08-16-02-16-34-13-17-34-16-34
t069 08-16-02-16-34-13-17-34-16-16-34
t097 07-23-12-12-23
spa typing is a cost effective method compared to other molecular typing methods, and it has a rapid turn-around time. It is stable, easy to use and has high reproducibility. There is also a standardized international nomenclature which facilitates communication of results. The sequence results are reported to an international database from which you receive the spa type. In this database you may learn if a certain spa type has been identified in other hospitals or countries and how common it is. The level of the discriminatory power makes it suitable for local short-term outbreaks. A disadvantage of this method is that it is based on single-locus typing and therefore may misclassify particular types due to recombination and/or homoplasy (Sabat et al., 2013). MRSA isolates lacking the spa gene have been described and spa typing could not be used for these strains (Baum et al., 2009).

Introduction
37
The advantages of this method make it currently the most useful for characterizing S. aureus isolates at the local, national and international levels (Friedrich et al., 2008; Grundmann et al., 2010; Hallin et al., 2007; Harmsen et al., 2003; Melin et al., 2009). Strommenger et al. reported the spa typing technique to be an excellent tool for national and international surveillance as well as for short-term local epidemiology. However, additional markers may be needed to overcome its limitations as a single-locus typing method (Hallin et al., 2007; Strommenger et al., 2006).
Terminal restriction fragment length polymorphism
tRFLP is based on PCR amplification of a target gene, and the amplification is performed with one or both of the primers having a fluorescent molecule labeled at their 5’ end. In case both primers are labeled, different fluorescent dyes are required. The mixture of amplified PCR-products is then digested with restriction enzymes. The restricted fragments are separated by the use of sequencing equipment. The terminal fragments are determined in a fluorescence detector. As only one part of the digested PCR fragments are labeled, only the terminal fragments are determined while all other fragments are ignored. Thus, tRFLP is different from RFLP in which all digested fragments are visualized. The result of a tRFLP is often visualized as an intensity plot called an electropherogram. In an electropherogram the X-axis marks the sizes of the fragments while the Y-axis marks the fluorescence intensity of each fragment. tRFLP has commonly been used to study the diversity of different bacteria in a variety of communities but it has also been modified to distinguish different isolates of S. aureus based on amplification of the coa or the spa gene (Mehndiratta and Bhalla, 2012; Mehndiratta et al., 2009). Since RFLP mostly has been used in studies of different microbial communities there is very few if any comparative studies with other typing methods for S. aureus. One disadvantage of tRFLP is that different bacteria might give indistinguishable patterns in the electropherogram due to the presence of a restriction site in the same position as the PCR product. This may be overcome by digestion in parallel by several enzymes or by labeling of both the primers with different fluorochromes (Buckingham Lela 2007; Liu et al., 1997). RFLP is also technically demanding and great efforts must be put into the design of the assay, as rare polymorphisms have been reported to confound RFLP results (Gold et al., 2001).

Introduction
38
Denaturing gradient gel electrophoresis
DGGE (Figure 7) was originally performed on restriction fragments to detect gene mutations (Buckingham Lela 2007). This method was then further developed to study the genetic diversity of complex microbial populations by separation of amplified products from the 16S rRNA gene (Muyzer et al., 1993).
Figure 7. Flow scheme of the procedure for the DGGE-analysis.
The method is based on the possibility of detecting differences in the denaturation of molecules with different sequences and the technique allows the detection of differences as small as one nucleotide. The difference in binding power between bp on the DNA strand affects the denaturation properties of double-stranded (ds) DNA. For DGGE, dsDNA fragments, 200-700 bp in length, are prepared by PCR amplification or by restriction digestion. The products are then separated on polyacrylamide gels containing a denaturing gradient of urea and formamid (Figure 8). As the dsDNA fragments move through the gel, the denaturing conditions increase with higher concentrations of urea and formamid. At a given concentration, a sequence reaches its

Introduction
39
denaturing point and the strands begin to separate (melt). Domains of different sequences, with different melting characteristics, separate at different points in the gradient. This results in different lengths of migration in the gel. Complete strand separation is prevented by naturally occurring or artificially placed GC-rich sequences (GC-clamps). These may be conveniently placed at the ends of PCR products by using primers tailed on the 5’ end with a 40 bp 5’ GC tail (Buckingham Lela 2007; Fischer and Lerman, 1983). Like tRFLP, DGGE could also be adapted to separate isolates of S. aureus based on amplification of the spa gene (Matussek et al., 2011).
Figure 8. The principle of DGGE. A: Description of the gradient gel. B: Description of melted DNA. Large molecules migrate slower in the gel than small molecules.

Introduction
40
Microarray
Microarray for bacterial typing can be carried out on a chip with a collection of DNA probes attached to a solid surface in a particular pattern. These probes detect complementary nucleotide sequences in different bacterial isolates. The array is constructed to detect genes that serve as markers for specific bacterial strains, or to detect allelic variants of a gene that is present in all strains of a particular species. The probes in the array may be cDNA, PCR products (>200 bp) or oligonucleotides (up to 70 bp). A common procedure is that total DNA is extracted from the bacterium of interest. This target DNA is then labeled, either chemically or by an enzymatic reaction, and is then hybridized to the array. Unbound target DNA is removed during the following washing steps with buffers of different concentrations. The signal from a successful hybridization between the labeled target DNA and an immobilized probe is measured automatically. The data produced are then analyzed using dedicated software (Buckingham Lela 2007; Sabat et al., 2013). DNA microarrays appear to be very well suited for bacterial typing, and microarrays are currently widely used to analyze genomic mutations, such as single-nucleotide polymorphisms. In addition, microarray technology is an efficient tool for the detection of extra-genomic elements (McCarthy et al., 2012; McCarthy and Lindsay, 2012). One advantage of microarrays is that pathogens may be simultaneously genotyped and profiled to determine their antimicrobial resistance and virulence potential (Buckingham Lela 2007; Sabat et al., 2013). The whole genome microarray approach is a useful alternative to whole genome sequencing and saves time, effort and expense, and it may be used in real-time outbreak investigations (Sabat et al., 2013). One disadvantage is that microarray analysis does not allow the identification of sequences which are not included in the array (Sabat et al., 2013).

Tabl
e 6.
Com
paris
on b
etw
een
diffe
rent
typi
ng m
etho
ds fo
r S. a
ureu
s.
Met
hod
Use
d/Su
itabl
e fo
r Ad
vant
ages
Di
sadv
anta
ges
Com
men
ts
De
natu
ring
grad
ient
gel
el
ectr
opho
resis
Dete
ctio
n of
sim
ulta
neou
s co
loni
zatio
n w
ith
diffe
rent
stra
ins
Can
sepa
rate
sequ
ence
s of
the
sam
e le
ngth
Ti
me
cons
umin
g Lo
w th
roug
hput
Has n
ot y
et fu
lly b
een
eval
uate
d in
clin
ical
sit
uatio
ns
(Mat
usse
k et
al.,
20
11)
Mic
roar
ray
May
be
used
in re
al-
time
outb
reak
in
vest
igat
ions
Dist
ingu
ishes
bet
wee
n ve
ry si
mila
r but
non
- id
entic
al se
quen
ces
Nee
ds sp
ecia
lized
eq
uipm
ent
High
cos
t of m
ater
ials
and
inst
rum
ents
Com
mer
cial
test
is n
ow
avai
labl
e bu
t the
re
prod
ucib
ility
with
in
and
betw
een
diffe
rent
la
bora
torie
s nee
ds to
be
esta
blish
ed
(Sab
at e
t al.,
20
13)
Mul
tiloc
us
sequ
ence
ty
ping
Long
-ter
m
epid
emio
logy
Ev
olut
iona
ry st
udie
s
Defin
es c
ore
gene
tic
popu
latio
ns
High
ly re
prod
ucib
le w
ithin
an
d be
twee
n la
bora
torie
s St
anda
rd n
omen
clat
ure
The
met
hod
may
be
used
fo
r oth
er p
atho
gens
as
wel
l
Tim
e co
nsum
ing
Low
thro
ughp
ut
High
cos
t Li
mite
d di
scrim
inat
ory
pow
er
For s
hort
-ter
m o
utbr
eak
situa
tions
MLS
T ha
s onl
y m
oder
ate
disc
rimin
ator
y po
wer
(Sab
at e
t al.,
20
13; S
tefa
ni e
t al
., 20
12;
Stru
elen
s et a
l.,
2009
) En
right
200
2,
Robi
nson
200
3
41

Com
paris
on b
etw
een
diffe
rent
typi
ng m
etho
ds fo
r S. a
ureu
s (co
ntin
ued)
.
Pulse
d fie
ld g
el
elec
trop
hore
sis
Has b
een
cons
ider
ed a
s “go
ld
stan
dard
” fo
r typ
ing
S. a
ureu
s(M
RSA)
in
outb
reak
situ
atio
ns
Loca
l out
brea
k in
vest
igat
ions
and
as
par
t of t
ypin
g sy
stem
for l
ong-
term
surv
eilla
nce
at
regi
onal
and
na
tiona
l lev
el
High
ly d
iscrim
inat
ory
May
be
used
for o
ther
pa
thog
ens a
s wel
l
Tim
e co
nsum
ing
Tech
nica
lly d
eman
ding
Lo
w-t
hrou
ghpu
t M
ultip
le n
omen
clat
ure
Misc
lass
ifica
tion
of so
me
linea
ges
Repr
oduc
ible
with
in th
e la
bora
tory
but
not
be
twee
n la
bora
torie
s ST
398
non-
typa
ble
by
stan
dard
pro
toco
l
(Sab
at e
t al.,
20
13; S
true
lens
et
al.,
2009
)
Stap
hylo
cocc
al
prot
ein
A
typi
ng
Firs
t lin
e ou
tbre
ak
situa
tions
Sh
ort t
erm
loca
l ep
idem
iolo
gy
Loca
l sur
veill
ance
an
d in
fect
ion
cont
rol p
urpo
ses
Nat
iona
l su
rvei
llanc
e
Tim
e ef
fect
ive
in
tran
smiss
ion
stud
ies
High
thro
ughp
ut
Stan
dard
nom
encl
atur
e Ea
sy to
shar
e an
d co
mpa
re se
quen
ce re
sults
Hi
ghly
repr
oduc
ible
w
ithin
and
bet
wee
n la
bora
torie
s At
trib
utio
ns o
f MLS
T ST
s by
BU
RP a
lgor
ithm
Misc
lass
ifica
tion
of a
smal
l nu
mbe
r of l
inea
ges
May
requ
ire
com
plem
enta
ry ty
ping
of
othe
r met
hods
O
nly
used
for t
ypin
g of
S.
aur
eus
Has b
ecom
e th
e pr
imar
y ty
ping
met
hod
for
regi
onal
and
nat
iona
l M
RSA
surv
eilla
nce
sche
mes
N
ot fo
r glo
bally
di
strib
uted
spa
type
s
(Hal
lin e
t al.,
20
07; S
abat
et a
l.,
2013
; Ste
fani
et
al.,
2012
; St
rom
men
ger e
t al
., 20
06;
Stru
elen
s et a
l.,
2009
)
42

Com
paris
on b
etw
een
diffe
rent
typi
ng m
etho
ds fo
r S. a
ureu
s (co
ntin
ued)
.
Term
inal
re
stric
tion
frag
men
t le
ngth
po
lym
orph
ism
Loca
l inv
estig
atio
n of
isol
ates
N
o in
form
atio
n N
o in
form
atio
n N
ot c
omm
only
use
d fo
r ty
ping
of S
. aur
eus a
nd
ther
efor
e no
t com
pare
d w
ith o
ther
men
tione
d m
etho
ds
(Meh
ndira
tta
et
al.,
2009
)
Who
le g
enom
e se
quen
cing
W
ill b
ecom
e a
pow
erfu
l too
l for
ou
tbre
ak
inve
stig
atio
ns a
nd
surv
eilla
nce
sche
mes
in ro
utin
e cl
inic
al p
ract
ice
Evol
utio
nary
stud
ies
of p
atho
gens
in th
e sh
ort t
erm
and
ove
r lo
nger
tim
e sc
ales
Com
pare
s diff
eren
t ge
nom
es w
ith a
sing
le-
nucl
eotid
e re
solu
tion
Port
able
Ea
sy to
shar
e an
d co
mpa
re se
quen
ce re
sults
M
ay b
e us
ed fo
r all
kind
s of
pat
hoge
ns
Spec
ializ
ed e
quip
men
t Hi
gh c
ost
The
chal
leng
e w
ill b
e to
ha
ndle
and
ana
lyze
all
the
info
rmat
ion
and
ther
e is
a ne
ed fo
r use
r-fr
iend
ly in
form
atic
s pl
atfo
rms
Stud
ies o
f the
mod
e an
d te
mpo
of g
enom
ic
evol
utio
n in
diff
eren
t ba
cter
ia li
neag
es m
ust
be p
erfo
rmed
In
vest
igat
ion
of h
ow th
is m
etho
d w
orks
whe
n ap
plie
d to
“re
al w
orld
” pr
oble
ms
(Pal
len
et a
l.,
2010
; Sab
at e
t al.,
20
13)
43

Introduction
44
Gene expression
Microarray
Microarrays for studies of gene expression are like arrays for genotyping, and are also based on DNA/RNA-hybridization (Figure 9). cDNA (250-5000 bp) or oligonucleotides (25-75 bp) corresponding to genes of interest, are attached to a solid surface, usually glass or a silica surface. The RNA from the sample of interest (cell cultures, tissue or blood) is reverse transcribed (RT) into cDNA, and then possibly transcribed to cRNA with incorporation of a fluorescent dye. The labeled cDNA/cRNA is then hybridized to the array. After stringent washing the array is scanned in a confocal microscopy scanner. This kind of analysis allows for simultaneous study of thousands of genes in a single assay, e.g. all open reading frames in the human genome may be detected. However, the complexity of such wide-ranging analysis makes microarray more of a screening method. To verify differentially expressed genes, quantitative real-time PCR may be used.
Figure 9. Example of a possible procedure to perform gene expression analysis by microarray.

Introduction
45
Real-time PCR
Since the introduction of PCR in 1988 (Mullis et al., 1986), its performance has not changed significantly, even though it has been modified and improved over the years. The greatest change has probably been the introduction of real-time PCR in 1993 (Higuchi et al., 1993). In conventional PCR, you only evaluate the end product of the PCR-reaction. In real-time PCR a fluorescent dye binds to the amplified DNA at each cycle. The fluorescent signal is continuously registered and is directly proportional to the amplified amount of DNA, which allows interpretation of the reaction in “real time”. By the incorporation of a standard with a known concentration of a target, it is possible to determine the initial amount of a sample (Buckingham Lela 2007). Real-time PCR has several advantages over traditional PCR. With real-time PCR it is possible to detect smaller variations in target copies compared to conventional PCR. Also, the quantitative detection range is much broader compared to conventional PCR, where only the endpoint product is analyzed. Nucleic acid amplification and detection are carried out in the same closed vessel, thereby minimizing the risk of cross-contamination between analyses. With continuous data collection, less time is required to obtain the result compared to conventional PCR where agarose gel electrophoresis must be performed to complete the analysis (Bustin, 2000). Real-time PCR can also be used to investigate gene expression. As for microarray, mRNA is used as starting material which requires a step of reverse transcription before the PCR analysis. To study changes in gene expression in, for example, stimulated cells, the expression are related to unstimulated cells. In order to correct for variations in starting amount of sample, a reference gene, a so-called, housekeeping gene is used for normalization. This gene should be expressed at a constant level in all cells. However, this perfect gene does not exist, so it is important to validate the reference gene to be used for the experiment. There are different methods to quantify alterations in mRNA levels using real-time PCR; the standard curve method (Larionov et al., 2005), the Pfaffl method (Pfaffl, 2001) or the delta-delta ct method (Livak and Schmittgen, 2001). The standard curve method includes a set of standards, with known concentrations, for the mRNA from both the target gene and the reference gene. By use of the standards, the amount of target and reference mRNA can be calculated. With this information it is possible to use the equation below (Figure 10). The fold change (FC) (stimulated versus unstimulated cells) for the target gene as well as for the reference gene is calculated and by dividing these values the ratio for target gene expression is given.

Introduction
46
With the Pfaffl method, the relative expression ratio is calculated only from the real-time PCR efficiencies (E) and the threshold cycle (ct) deviation of, for example, stimulated cells versus unstimulated cells (Figure 10). A standard curve is only needed to determine an average efficiency of the assay both for the target and the reference gene. The delta-delta ct method was one of the first methods to be used for relative quantification in combination with real-time PCR. However, optimal and identical real-time amplification efficiencies of target and reference gene are presumed in this method. The delta ct value between target and reference gene of stimulated cells is calculated. The same calculation is performed for the unstimulated cells. Finally the delta-delta ct value between stimulated and unstimulated cells is calculated (Figure 10).
Figure 10. Different equations used to calculate relative quantities in gene expression analysis.

Aims
47
AIMS The general aim of this thesis is to increase the knowledge of S. aureus pathogenesis and to elucidate the pattern of colonization. The virulence of S. aureus is studied by the interaction with human umbilical vein endothelial cells (HUVEC) as a model. To contribute to better knowledge of S. aureus colonization and spread in the community the spa type distribution, antibiotic susceptibility, prevalence, and carriage, among nursing home residents is studied and a method to assess multiclonality is developed.
The specific aims are:
To evaluate the induction of gene expression in HUVEC by S. aureus infection (Papers I and II).
To explore possible differences in virulence between S. aureus isolates from patients with septiceamia compared to commensal isolates. This is done by:
a) exploring a possible difference in the ability of internalization, intracellular survival and induction of apoptosis of S. aureus in HUVEC (Papers I and II).
b) studying global gene expression patterns in HUVEC (Paper II).
To investigate the presence of virulence genes in the two groups of S. aureus isolates, by the use of DNA-based genotyping (Paper II).
To study the epidemiology of S. aureus by molecular typing to explore possible variations in its ability to colonize and spread. Swedish nursing homes are used as a model, and the prevalence, carriage rate and issue of multiclonality as well as the spatial and temporal spa type distribution are evaluated (Paper III).
To develop a denaturing gradient gel electrophoresis assay for S. aureus utilizing spa to further characterize multiclonal colonization and infection (Paper IV).

Results and discussion
48
RESULTS AND DISCUSSION Isolation and cultivation of S. aureus (papers I-IV)
In the first paper a clinical isolate of S. aureus, characterized in previous studies, was used (Strindhall et al., 2002). This isolate was selected due to its ability to induce high up-regulation of cell adhesion molecules and to highly stimulate the production of cytokines in HUVEC without being cytotoxic. These properties were important since we wanted to ensure we obtained a genetic answer. When performing microarray analysis, large amounts of good quality RNA are required, therefore a non-cytotoxic isolate was chosen. In paper II, non-virulent isolates of S. aureus were compared with clinical virulent isolates. Five randomly selected isolates from a collection of S. aureus samples obtained from healthy male nasal carriers in a previous study (Matussek et al., 2007), represented the non-virulent group. S. aureus isolates obtained from blood cultures from five individuals without prior signs of disease or infection (diabetes, endocarditis, drug addicts and persons who had undergone surgery within the last three years were excluded) were included in the virulent group. These isolates were selected from a study of personal records. spa typing was performed to ensure that all isolates belonged to different strains. All ten strains were also genotyped with a DNA-based microarray to determine the presence of genes encoding virulence determinants. Prior to the array analysis, the bacterial isolates were lysed and treated with proteinase K before purification of bacterial DNA. A multiplex PCR-reaction was used for simultaneous amplification of all target genes, and PCR-products were labeled by incorporation of biotin-16-dUTP. After denaturation, the products were hybridized to the array. After washing and addition of a blocking reagent, horseradish-peroxidase-streptavidin conjugate was added. This was followed by incubation and washing. The array tube was placed in a reading device and Seramun Green precipitating dye was added. An image of the array was recorded and analyzed. Analysis of hybridization patterns indicated the presence or absence of individual genes as well as the assignment of isolates to clonal complexes or sequence types. There was no difference in the presence of the selected virulence genes between the two groups of colonizing and invasive strains (Table 7 and 8). Thus, it seems that there is no difference in virulence between colonizing and invasive strains. This is in line with studies showing that most of the isolates

Results and discussion
49
causing invasive infections are recruited from the colonizing flora of the anterior nares (Wertheim et al., 2005b; Wertheim et al., 2004; von Eiff et al., 2001). This indicates that the risk of future infection cannot be predicted only by characterizing a nasal isolate. The result of the interaction between bacteria and the host is also dependent on host factors (van den Akker et al., 2006). In paper III, samples from the anterior nares, the throat, the groin and from active skin lesions (if any) were collected from individuals participating in a study called SHADES (The Study of Health and Drugs among the Elderly in Swedish Nursing Homes) (Ernsth Bravell et al., 2011). If possible, samples were collected every six months during the study period. After the start of the study it was discovered that throat sampling was very demanding and difficult to perform in this group of elderly individuals and therefore it was problematic to get throat samples on many of the occasions. All samples were cultured in an enrichment broth for S. aureus before culture on blood agar plates or agar plates selective for S. aureus. In paper IV, clinical enrichment broth samples associated with an MRSA outbreak were used together with isolates from a strain collection bank at the Department of Microbiology at Ryhov County Hospital, Jönköping.

Ta
ble
7. P
rese
nce
of e
nter
otox
in g
enes
.
Isol
ate
Tst1
se
A se
A 32
0E
seA
N31
5 se
B se
C se
D se
E se
G se
H se
I se
J se
K se
L se
M
seN
se
O
seQ
se
R se
U/Y
ORF
CM
14
E´to
xin
hom
olog
CI
CI
I
CI
II
CI
V
CV
BI
BI
I
BI
II
BI
V
BV
*3
08
gen
e no
t pre
sent
,
g
ene
pres
ent,
am
biva
lent
resu
lt. Is
olat
es d
esig
nate
d “B
” be
long
to th
e gr
oup
of in
vasiv
e iso
late
s and
isol
ates
de
signa
ted
“C”
belo
ng to
the
grou
p of
col
onizi
ng is
olat
es. *
Isol
ate
used
in p
aper
I.
50

Tabl
e 8.
Pre
senc
e of
diff
eren
t gen
es re
pres
entin
g ot
her v
irule
nce
dete
rmin
ants
.
Le
ucoc
idin
es
Bovi
ne P
VL
Leuc
ocid
ine
lukD
/E
Leuc
ocid
ine
hom
olog
es
Hem
olys
ine
and
stap
hylo
kina
se
Isol
ate
lukF
lu
kS
hlgA
lu
kF-
PV
lukS
-PV
lu
kF-
P83
lukM
lu
kD
lukE
lu
kX
lukY
lu
kY
MRS
A252
hl
hl
a hl
b sa
k hl
d Hl
_III
CI
CII
CIII
CIV
CV
BI
BII
BIII
BIV
BV
308*
Exfo
liativ
eTox
in
Prot
ease
Ep
ider
mal
cel
l diff
eren
tiatio
n in
hibi
tors
Is
olat
e et
A et
B et
D sp
lA
aplB
ed
inA
edin
B ed
inC
CI
CII
CIII
CIV
CV
BI
BII
BIII
BIV
BV
308*
g
ene
not p
rese
nt,
g
ene
pres
ent,
am
biva
lent
resu
lt. Is
olat
es d
esig
nate
d “B
” be
long
to th
e gr
oup
of in
vasiv
e iso
late
s and
isol
ates
de
signa
ted
“C”
belo
ng to
the
grou
p of
col
onizi
ng is
olat
es.*
Isol
ate
used
in p
aper
I.
51

Results and discussion
52
HUVEC as a model to study interactions with S. aureus (papers I and II)
ECs located on the inner surface of blood vessels, are key effector cells and activate the innate immune response. Innate immunity has an important role in the defense against S. aureus infections through identifying the bacteria by pattern recognition molecules, activating complement and phagocytosis (Medzhitov and Janeway, 1998). In papers I and II, HUVEC was used as a model to study the interaction of different S. aureus isolates with ECs. Human umbilical cords were collected at the maternity ward at Ryhov County Hospital, Jönköping. The umbilical cords were collected according to a modified protocol from Jaffe et al. (Jaffe et al., 1973) in PBS pH 7.4 with ampicillin and streptomycin and ECs were isolated within 72 hours. The vein was flushed with PBS pH 7.4 before collagenase was inserted, and the cord was incubated at 37 ⁰C. After incubation, the cord was gently squeezed to loosen as many endothelial cells as possible in the vein. The collagenase was then collected in a tube and subsequently centrifuged. The cells were washed once with PBS before being seeded into 25 mL culture flasks with cell culture medium. The cells were incubated at 37 ⁰C in 5 % CO2. Within 2-3 days, the endothelial cells became confluent with growth that formed a cobblestone-monolayer. The passage of the cells was performed by trypsin/EDTA and the cells were used for experiments in passage 2-3. HUVEC was cultured in 6-, 12- or 24-well plates prior to the experiments and grown to confluence. S. aureus isolates were cultured on blood agar plates and the bacterial cells were suspended in PBS pH 7.4 to 0.5 McFarland and incubated with HUVEC for 1h. Extracellular bacteria were then killed with lysostaphin, and cell culture medium was added for indicated periods of times. In both papers I and II an “unphysiologically” high number of bacteria (108 CFU/mL) was used to infect HUVEC. This concentration of bacteria was, in a previous work (Strindhall et al., 2002), found to be optimal for activation of HUVEC in this infection model. This can be compared with the number of bacteria present in blood of patients with bacteremia which is often less than 10 CFU/mL (Reimer et al., 1997; Yagupsky and Nolte, 1990). In papers I and II, the conditions for infection of HUVEC with S. aureus were the same. After the infection period, HUVEC with internalized bacteria was incubated for different periods of time. In paper I, the experiment was performed after an incubation of 18 h, and in paper II the incubation time was 4 h only. The short period of incubation in paper II was chosen due to a cytotoxic effect observed for some of the isolates after 6 h of interaction with

Results and discussion
53
HUVEC. Cytotoxic damage may lead to destruction of RNA and hamper microarray analysis. In paper I, a non-cytotoxic strain was used and for this reason a longer incubation time could be used.
Internalization and survival of S. aureus in HUVEC (papers I and II)
In paper I, the adhesion and internalization of S. aureus to HUVEC was examined using electron microscopy, the adherence was evaluated with field emission electron microscopy, and internalization of bacteria was investigated with transmission electron microscopy. The results showed that S. aureus adhered to and was internalized into HUVEC. The bacteria cells were found in endosomes and after 18 h of incubation, dividing bacteria were observed (Figure 1 in paper I). The same result was observed in a study similar to ours (Grundmeier et al., 2010). This indicates that S. aureus is an facultative intracellular pathogen (Fraunholz and Sinha, 2012). In paper II the isolate’s ability to adhere to ECs was examined with acridine orange staining. The ECs were cultured on glass slides before infection with S. aureus. After washing, adherent cells were stained with the DNA-binding dye and the results were observed under fluorescence microscope. In both papers I and II, the intracellular survival of internalized S. aureus was quantified with viable count. The experiment was performed in triplicate wells in 24 well plates. The cells were infected for 24, 48 and 72 h, respectively. After incubation, the cell culture medium was cultured on blood agar plates. Adherent ECs were lysed with a buffer that did not affect the bacteria, and then cultured on blood agar plates after dilution. After overnight incubation for 24 h, the bacteria colonies were counted. Three independent experiments were performed for each strain. These experiments showed that all bacteria isolates used in papers I and II survived for at least 72 h. S. aureus has been described as an extracellular pathogen, although an intracellular phase in S. aureus infections has been shown by the ability to invade various cell types including ECs (Figure 11) (Grundmeier et al., 2010; Ogawa et al., 1985; Vriesema et al., 2000). This ability was confirmed in our studies (Figure 1 in paper I). An intracellular phase of S. aureus infection may protect the bacteria from phagocytosis by professional phagocytes (Fraunholz and Sinha, 2012) and may also protect the bacteria from antibiotics that are unable to act intracellulary (Krut et al., 2004). From paper I it cannot be concluded whether the pro-inflammatory response detected required the internalization of S. aureus, or if the adherent bacteria were enough. However, Beekhuizen et al (Beekhuizen and van de Gevel, 2007) concluded in a study on

Results and discussion
54
the effect of interferon-γ on HUVEC-S. aureus interaction, that activation of ECs is a consequence of interaction of S. aureus with the EC surface, rather than result that occurs after bacterial internalization.
Figure 11. Scanning electron microscopy (performed by M. Rohde) shows adherence and initial uptake of S. aureus into HUVEC after 1 h of incubation (paper I). HUVEC were cultured on 12 mm cover glass in 24-well plates and incubated with approximately 1 x 107 CFU/mL of bacteria in cell culture medium without antibiotics. White bar represent 1 μm.
S. aureus induction of apoptosis in endothelial cells (papers I and II) After incubation of internalized S. aureus with HUVEC for different periods of time, the cells or cell culture medium were prepared for the following analysis. For the flow cytometry experiments on ECs, unattached cells in the medium were collected by centrifugation. The monolayer of HUVEC in the wells was treated with trypsin, and after they were detached, they were transferred to new tubes. The pelleted cells from the same well were transferred to the trypsonized cells from the same well. After washing, the ECs were marked with different kinds of fluorochromes. For detection of apoptotic cells, Annexin V antibodies conjugated with fluorescein isotiocyanate (FITC) were used and propidiumiodide was added to detect necrotic cells. This experiment was done on cells infected for 4, 8, 12 and 24 h, respectively. In paper I, no sign of apoptosis was observed after 42 h of interaction with the non-cytotoxic strain used. In paper II, six of ten strains (two colonizing and four invasive) induced apoptosis after 12 and 24 h of incubation, but not after 4 h. There was also no

Results and discussion
55
difference between isolates in the regulation of genes connected to apoptosis, not even when comparing the apoptotic isolates with the non-apoptotic. There were only a limited number of differentially regulated genes connected to apoptosis, which supported the data from the apoptosis experiment. This may be explained by the short incubation time (4 h). None of the ten strains induced necrosis in HUVEC. Krut et al. (Krut et al., 2004) suggested an exclusive relationship between S. aureus and fibroblasts or keratinocytes in that either S. aureus or the host cells is killed. For the isolates in paper II, our results indicated that S. aureus isolates survive intracellularly without killing the ECs for at least 72 h. This may represent a niche for invasive S. aureus, which might be of importance in persistence and recurrence of infection as well as in the choice of antibiotic strategy. Of the antibiotics commonly used to treat S. aureus infections, only rifampicin is able to eliminate intracellular S. aureus (Krut et al., 2004). In a mouse peritonitis model a poor correlation between the intracellular accumulation of antibiotics and the actual intracellular effect was found (Sandberg et al., 2009). For measurement of soluble cytokines and chemokines with enzyme-linked immunosorbent assay (ELISA), the cell culture medium was centrifuged before the start of the analysis. In paper I, ten proteins were investigated (Figure 3 in paper I), while six were investigated in paper II (Figure 2 in paper II). For the detection of vascular cell adhesion molecule (VCAM)-1 on the surface of ECs, monoclonal FITC-conjugated antibodies were used.
Gene expression in HUVEC infected with S. aureus (papers I and II) After infection of HUVEC for 18 h as described in paper I and for 4 h, reported in paper II, RLT buffer was added to each well of infected HUVEC, in order to preserve the RNA in the cells. Total RNA was isolated and the amount and quality were investigated. To get a sufficient amount of RNA (1.5 μg) and to compensate for variation in HUVEC from different donors, equal amounts of RNA from HUVEC infected with the same bacteria isolate from three independent experiments were mixed. RNA was converted to ds cDNA, and thereafter cDNA was transcribed to cRNA with a mixture of unlabeled dNPTs and biotin-labeled CTP and UTP. Fragmentation of cRNA (100-150 bases) was performed, and thereafter the samples represented by denatured cRNA were loaded onto the microarray chip (22,283 probe sets representing 14,239 human genes). Hybridized cRNA was then fluorescently labeled by adding

Results and discussion
56
streptavidin-phycoerythrin. After scanning, the results were evaluated and genes with at least a threefold or greater change in signal intensity in two independent experiments compared to uninfected cells, were considered regulated.
In the study described in paper I, 194 probe sets representing 156 different genes were differentially regulated, 149 were up-regulated and seven were down-regulated. As expected, genes representing the innate immune response were differentially regulated. The following groups were found: genes associated with adhesion, antigen presentation, apoptosis, cytokines and chemokines, and genes associated with complement (Table 2 in paper I). The highest increase in mRNA levels was found in the group of cytokines and chemokines, and many of these were also detected with ELISA (Figure 3 in paper I). In the study described in paper II, there was no difference in gene regulation between the two groups of invasive and colonizing S. aureus isolates. However, there was a great variability in the number of differentially regulated genes in HUVEC (51-156) between the different isolates (Table 9). This variation could not be explained by any difference in the presence of virulence genes in the different S. aureus isolates (Tables 7 and 8). In paper II, a “core set of genes” in HUVEC was differentially regulated after infection with all ten isolates (Table 5 in paper II). There were 45 genes, classified into the following groups; apoptosis, cell metabolism, cytokines and chemokines, signaling and transcription factors and interferon-associated genes. One may speculate that these genes always become differentially regulated at the onset of an infection with S. aureus, independent of the isolates’ characteristics. The majority of these 45 genes may also be connected to an innate immune response in ECs. Grundmeier et al showed that different clinical isolates varied greatly in their ability to induce an inflammatory response in ECs but that many of the differentially regulated genes were involved in innate immunity (Grundmeier et al., 2010).

Results and discussion
57
Table 9. Number of differentially regulated genes at different fold changes*.
Strain FC up BI BII BIII BIV BV CI CII CIII CIV CV
2 71 125 145 156 88 117 156 126 106 51
5 17 37 44 45 21 35 50 40 26 13
10 10 17 19 21 12 15 23 17 13 5
FC down
2 22 101 49 12 130 31 181 78 41 138
5 2 5 3 1 4 0 5 2 1 8 *Fold change (FC) is the factor of regulation of mRNA from S. aureus infected HUVEC versus non-infected HUVEC. Strains designated “B” belong to the group of invasive isolates and isolates designated “C” belong to the group of colonizing isolates.

Fi
gure
12
. Hie
rarc
hic
al c
lust
er a
nal
ysis
of
hu
man
gen
es (
I) m
easu
red
in d
up
licat
e b
y A
ffym
etri
x G
ene
Ch
ip (
Hu
.U1
33
plu
s 2
.0)
afte
r 4
h o
f in
cub
atio
n o
f H
UV
EC
wit
h d
iffe
ren
t S.
au
reu
s. C
olu
mn
s re
pre
sen
t st
rain
s an
d is
ola
tes
des
ign
ated
“B
” b
elo
ng
to in
vasi
ve is
ola
tes
and
“C
” b
elo
ng
to c
olo
niz
ing
iso
late
s, “
N”
rep
rese
nt
un
infe
cted
cel
ls. T
he
ho
rizo
nta
l lin
es r
epre
sen
t d
iffe
ren
t ge
nes
as
sho
wn
in II
, wh
ich
is a
n e
nla
rged
sec
tio
n o
f I.
Gre
en c
olo
r sh
ow
s d
ow
n-r
egu
late
d g
enes
an
d
red
co
lor
up
-reg
ula
ted
gen
es
.
58

Results and discussion
59
The array results in both papers I and II were confirmed with real time RT-PCR on a selection of genes involved in the inflammatory response. These experiments were performed with the same pooled RNA samples that were used for the microarray analysis. For relative quantification, the housekeeping gene RPS9 was used for normalization. All analyzed genes, nine in paper I (Figure 2 in paper I) and seven in paper II (Figure 1 in paper II), confirmed the array results, with only one exception in paper II. The protein expression in HUVEC infected with S. aureus was also investigated to confirm the gene expression results. The levels of 10 soluble proteins in paper I and six in paper II, were measured after different incubation times. In paper I, nine of the ten tested proteins were elevated (Figure 3 in paper I), and in paper II all cytokines/chemokines were elevated, with no difference between the two groups of bacteria (Figure 2 in paper II). The expression of all analyzed proteins increased over time.
Prevalence and molecular epidemiology of S. aureus (paper III)
Differences in S. aureus prevalence have been reported depending on the population studied (Kluytmans et al., 1997). The prevalence varies between 10-85 % and a majority of these studies were on populations within hospital settings and on people with different conditions (Kluytmans et al., 1997). In recent studies on general populations, a great difference has also been found between different countries (den Heijer et al., 2013), and indications of difference between gender have been observed (Mernelius et al., 2013a; Olsen et al., 2013). In paper III, we found an overall prevalence of 49 % among all participating individuals sampled in three locations (anterior nares, throat and groin), independent of which body site was tested or how many sites were colonized. When only the anterior nares were studied, 31 % were colonized, and 34 % were colonized in the throat. Most studies have focused on colonization of the anterior nares, and the colonization rate for this body site in our investigation is in agreement with recent data from a study on a general population in Sweden (den Heijer et al., 2013). The anterior nares has been considered as the most consistent site of S. aureus colonization (Kluytmans et al., 1997; Williams, 1963), but recent studies have suggested that colonization of the throat is more prevalent (Lee et al., 2011; Marshall and Spelman, 2007; Nilsson and Ripa, 2006), and in paper III the highest colonization rate was found in the throat, which supports these findings. The variations in prevalence rates may be due to individual differences such as genetic (Olsen et al., 2012; van Belkum et al., 2007; van den Akker et al., 2006), (antibiotic) treatment, diseases

Results and discussion
60
and environmental factors. The variations may also be a consequence of differences in sampling technics, logistic factors and culture conditions (VandenBergh et al., 1999). It is known that decolonization of the nose usually has a decolonizing effect on the skin and perineum (Parras et al., 1995; Varga and White, 1961), as well as a reducing effect on postoperative infections (Perl et al., 2002). Whether decolonization of the throat has the same effect is still not known.
From epidemiological studies in hospital settings or in general populations, a diverse distribution of spa types has been found (Grundmann et al., 2010; Mernelius et al., 2013a; Miko et al., 2013; Sangvik et al., 2011). In paper III, a diverse distribution was also found. Three different geographical regions in the south of Sweden were included, and we did not find any difference in spa type distribution between the different regions. However, in one of the regions, signs of the clonal spread of one spa type were found, as this specific spa type was found in seven of nine individuals at the same nursing home. BURP cluster analysis of all spa types (n=98) found in the longitudinal study in paper III revealed 10 clusters and 14 singletons. The four largest clonal clusters (CC) were spa-CC 015, spa-CC 012, spa-CC 084 and spa-CC 005 and these included 45 % of all detected spa types. Two of these most common clusters were also the most common in the Swedish HITS-study (Mernelius et al., 2013b) from the same region as our study reported in paper III. Of the 35 individuals that carried different spa types at any time during the study period, 21 carried the different strains on the same occasion and 18 changed spa type over time. Three individuals carried spa types that were closely related, and in two of these individuals both spa types were present on the same sampling occasion. Because of the difference in the number of samples, mainly between the nares and throat, we are not able to draw any conclusions about how often an individual changes strains. It was showen in a study by Sakwinska et al. (Sakwinska et al., 2010) that replacement of S. aureus strains occurred quite infrequently but that the mutation rate in the spa locus was high.

Results and discussion
61
Colonization and persistent carriage of S. aureus (paper III) Usually, three categories of S. aureus carriage are described: persistent, intermittent and non-carriers (Eriksen et al., 1995; Hu et al., 1995; Kluytmans et al., 1997; Kuehnert et al., 2006; Nouwen et al., 2004; Wertheim et al., 2005b). Recently, a reclassification of the carriage groups has been suggested, namely persistent carriers and others (van Belkum et al., 2009). The definition of a persistent carrier varies from study to study, depending on how many samples they include, at what interval the samples are obtained, and how many positive cultures are needed to meet the classification of a persistent carrier (VandenBergh et al., 1999). Also, most studies are based on samples from the anterior nares. In paper III, we found 20 % persistent carriers in the anterior nares if three out of three samples were required to contain S. aureus. We found that 17 % were carrying the same spa type on all three occasions. Carriage rates based on two sampling occasions from the nares showed that S. aureus was isolated on both occasions in 22 % of the individuals, and in 19 % of the individuals S. aureus of the same spa type was found. This indicates that two sampling occasions may be enough to confirm if an individual is a persistent S. aureus carrier or not. Our rates are in agreement with other studies (Kluytmans et al., 1997; van Belkum et al., 2009). It is important to identify the group of persistent carriers because they have an increased risk of infection (Nouwen et al., 2004; Wertheim et al., 2004; von Eiff et al., 2001). In about 80 % of cases, S. aureus infections are caused by the carrier strain present on the nasal mucosa of the patient (Wertheim et al., 2004; von Eiff et al., 2001). Another piece of evidence that the strain carried in the nares is causing disease is that eradication of S. aureus from the anterior nares has proved to be effective in reducing the incidence of infection by the bacteria (Bode et al., 2010; Kluytmans and Wertheim, 2005; Perl et al., 2002). The importance of carrying S. aureus in the throat is still not known. In paper III, we found a rate of 7 % persistent S. aureus throat carriage. This is a lower figure than shown by Nilsson et al. (Nilsson and Ripa, 2006). Explanations for the low rate in our study could be the low number of individuals that agreed to samples being taken on three sampling occasions from this body site, and that the obtained throat samples were of poor quality. Due to the discrepancy in the number of sampling occasions for the different sampling sites, the throat and nares were analyzed separately in order to evaluate the carriage state. In the nares, 20 % of the individuals were colonized with S. aureus on three occasions during one year, but for the throat the corresponding rate was only 7 %. Also the carrier state for an individual was evaluated. An individual was considered as a carrier of S. aureus if the

Results and discussion
62
bacterium was found in one or more of the three sampling sites. Forty-four individuals were sampled at all three sampling sites on three sampling occasions, and 25 % of these individuals were colonized with S. aureus on all three occasions. In 20 % the same spa type was isolated on all the occasions. In 30 % of the individuals, S. aureus was not detected at any sampling site on any of the three occasions.
Antibiotic resistance found in S. aureus (papers II and III) In paper II, the presence of some resistance genes as virulence factors was investigated. Only resistance against β-lactamase was detected in the majority of the isolates (7 of 10 isolates). The investigated genes were; blaZ (β-lactamase), mecA (modified penicillin binding protein), ermA, ermC, linA, msrA, vatA, vatB, vga, vgaA, vgB (macrolides/lincosamines/streptogramines), aacA-aphD, aadD, aphA3 (aminoglycosides), sat (streptothricine), dfrA (trimethoprim), far1 (fusidic acid), mupR (mupirocin), tetK, tetM (tetracycline), vanA, vanB, vanZ (vancomycin resistance genes in enterococci). Overall, in paper III low antibiotic resistance was found among the nursing home residents and there was no association between resistance and spa type. The most prevalent resistance was that against ciprofloxacin. None of the isolates were resistant to vancomycin, gentamicin, erythromycin, linezolid or rifampicin and no MRSA isolates were detected. It has been suggested that nursing home residents may be a reservoir for antimicrobial resistance (Bonomo, 2000), but this was not observed in our study (paper III). Also, a study by van der Donk et al showed no higher prevalence of resistance among nursing home residents compared with the remaining population in the same area (van der Donk et al., 2013). Further studies correlating resistance to antibiotic load at a unit or in relation to various conditions of residents, should be conducted. Even in countries with a high prevalence of MRSA in hospital settings, the prevalence found among the nursing home residents was lower (Pfingsten-Wurzburg et al., 2011).

Results and discussion
63
Multiclonality of S. aureus (papers III and IV)
The bacterial isolates or pelleted bacteria from enrichment broth were lysed at 95 °C before DNA isolation. The PCR-reaction was performed with the same primers as for spa typing (Kahl et al., 2005) with the difference that a GC-clamp was attached to the forward primer. The PCR products were then analyzed on a polyacrylamide gradient gel, using a DCode universal mutation detection system. The gel contained a gradient of urea and formamid, ranging from 15 % to 50 %. After running the gel electrophoresis at 62 ⁰C for 14 h, the gel was stained with SYBERGold and the presence of separated bands was visualized in UV transillumination, using a charge-coupled device (CCD) camera. In paper III, we identified a number of individuals colonized with S. aureus at more than one body site on the same sampling occasion. In some of these individuals, S. aureus belonging to two different spa types was detected. The issue of multiclonality is important and is not addressed in clinical laboratory diagnostics, where often only one isolate is analyzed. This might lead to an inconclusive report as a more virulent and antibiotic resistant strain may be missed, and in epidemiological investigations misleading results may be reported. This was illustrated in paper III, where five individuals were colonized with one resistant and one non-resistant isolate of S. aureus, and these isolates belonged to different spa types. Whether the two strains isolated from an individual were also present at one body site could not be evaluated in this study due to the study protocol, but this question should be further investigated. In order to detect different clones of S. aures in one sample, a method that is independent of culture was developed in paper IV. This method is based on DGGE and amplification of the spa gene. First, the PCR conditions required were optimized, and then the attachment of the GC-clamp to the spa primer was evaluated. The discriminatory power and the sensitivity of the method were evaluated, and we found that it was possible, from the spa gene, to detect PCR-products with the same length but different sequences. Moreover, the sensitivity was found to be 1:1000 when a serial dilution of a mixture of two S. aureus isolates was amplified. This is a great advantage but another benefit is that no prior isolation from a culture is needed. A disadvantage is that the method is time consuming and, like other gel-based methods, the result is based on individual assessment. The spa DGGE method was also tested on samples previously obtained in an investigation of an MRSA outbreak (Figure 13). In this outbreak, two different but closely related S. aureus spa types (t015 and t069) were detected (Table 10).

Results and discussion
64
Table 10. Comparison of repeat succession of two closely related spa types. t069 08-16-02-16-34-13-17-34-16-16-34
t015 08-16-02-16-34-13-17-34-16-34
Broth samples from colonized individuals (n=13) were analyzed with the new spa DGGE method. The results showed that two individuals were simultaneously colonized with both t015 and t069 at one body site and one individual was colonized with the different strains at different body sites on the same sampling occasion.
Figure 13.Overview of colonized individuals within an MRSA outbreak with the detection of two different spa types. *In samples from two of the individuals both spa types were simultaneously detected with DGGE (Figure 2 in paper IV). Line A represents nursing home residents, line B staff at the nursing home, and line C relatives to staff. The ovals represent females and rectangles represent males. Samples were taken from the nares (N), throat (T) and groin (G).
For some of the individuals colonized with S. aureus in paper III, broth samples were collected and frozen. In the future, these samples will be analyzed with the spa DGGE method developed in paper IV. With the results from paper III, it will be interesting to investigate how commonly multiple colonization occures in this population of nursing home residents. The result may then be compared with our result in paper III where we found a rate of 23 % with simultaneous colonization of different clones of S. aureus at different body sites.

Conclusion
65
CONCLUSION The focus of this thesis is on the pathogenesis and molecular epidemiology of S. aureus. The interaction model, S. aureus and HUVEC, was suitable for evaluateing the virulence of S. aureus. To study the epidemiology of S. aureus, antibiotic resistance and transmission patterns including multiclonality, nursing home residents are important and suitable populations.
Paper I:
The microarray revealed the induction of broad activity of the innate immune response in HUVEC after infection with S. aureus.
An intracellular phase of S. aureus infection was observed and an intracellular survival period of at least 42 h was documented.
Paper II:
Genotyping by microarray revealed no differences between invasive and non-invasive isolates of S. aureus in their content of virulence genes.
The gene expression in HUVEC after infection with S. aureus revealed no differences, when the results of invasive and non-invasive isolates were compared.
Among the genes induced in HUVEC by all S. aureus isolates, the dominance of the innate immune response was confirmed.
Apoptosis of HUVEC was induced by isolates representing the invasive as well as the non-invasive groups. None of the ten isolates induced necrosis.
An intracellular phase of S. aureus infection was confirmed, since all ten isolates survived for at least 42 h, as documented by viable count.

Conclusion
66
Paper III:
High diversity among strains of S. aureus colonizing the population of nursing home residents.
Possible spread of one clone within one of the nursing homes.
Overall, the prevalence of antibiotic-resistant strains was low and no MRSA was found.
Antibiotic resistance was more prevalent in multiclonal carriers and simultaneous detection of resistant and non-resistant strains was registered.
There was relatively high prevalence of S. aureus among the individuals in this population of elderly.
Paper IV:
The newly developed DGGE method for detection of different S. aureus spa types in one sample was sensitive and had a high discriminatory power.
The spa-DGGE method may be useful for checking clinical samples to ensure the detection of possible multiclonal infections thereby guaranteeing correct treatment.
The method was useful in the investigation of an MRSA outbreak comprising two different spa types, and may be used as a tool for epidemiological investigations.

Conclusion
67
Benefits for individuals and future perspectives S. aureus is a highly pathogenic bacterium and at the same time a frequent commensal in humans. In this thesis no easily detected virulence factors characterizing an isolate as highly virulent or of low virulence were found. This means that all S. aureus isolates must be regarded as potentially pathogenic in a clinical situation. The documentation of the intracellular phase of the infection has implications for the choice of antibiotic treatment. Although surveillance of antibiotic resistance is low today, due to the fear of a future spread of resistant clones, increased surveillance is warranted. Nursing home residents comprising a group of individuals that have frequent contact with hospital settings and therefore represent an important and convenient group that may be the basis for an early warning system of antibiotic resistance. The newly-developed DGGE method for multiclonal detection of S. aureus, offers the possibility to ascertain the correct antibiotic treatment. It also increases the quality of epidemiological investigations. The low prevalence of S. aureus in the groin samples indicates that this body site may be omitted in screening programs. On the other hand, a high prevalence was found in throat samples, which indicates that this body site should be included in screening programs. The importance of throat colonization for autologous infections and for transmission should be further evaluated.

Acknowledgements
68
ACKNOWLEDGEMENTS I would like to express my sincere gratitude to all who, in one way or another, have helped and supported me during these years and who in different ways have contributed to the completion of this thesis. I thank you all, and especially:
Per-Eric Lindgren, my main supervisor, for support, advice and guidance over the years.
Sture Löfgren, Erik Kihlström, Jan Strindhall and Andreas Matussek, my co-supervisors, for support, advice and guidance over the years. A special thanks to Sture for the support during the last year.
All my great colleagues and friends, present and past, at the Department of Clinical Microbiology, Ryhov County Hospital for invaluable support, understanding and great encouragement over all these years. You are the best!
All my friends at the section of Molecular Biology at the Department of Clinical Microbiology, Ryhov County Hospital, it is you who made this to the world's best place to work.
The girls at the office (the kiss and cry corner) and to you who sometimes visit us, for help, great support and discussions about almost everything.
Anna J, Anna Ö, Diana, Gun and Vera for help with figures and other “technical” things.
All co-authors in Sweden and Germany for making my articles possible.
All my great friends for love, support and just being my friends and for that some of you never stop asking questions.
Jonas because you wanted to run with me.
My family for letting me “be me”.
This work was supported by grants from Futurum – the Academy of Healthcare, Jönköping County Council and the Health Research Council in the South-East of Sweden (FORSS).

References
69
REFERENCES Armstrong-Esther, C.A., 1976. Carriage patterns of Staphylococcus aureus in a healthy non-hospital
population of adults and children. Ann Hum Biol 3, 221-227. Bannerman, T.L., Hancock, G.A., Tenover, F.C., Miller, J.M., 1995. Pulsed-field gel electrophoresis as a
replacement for bacteriophage typing of Staphylococcus aureus. J Clin Microbiol 33, 551-555. Baum, C., Haslinger-Loffler, B., Westh, H., Boye, K., Peters, G., Neumann, C., Kahl, B.C., 2009. Non-
spa-typeable clinical Staphylococcus aureus strains are naturally occurring protein A mutants. J Clin Microbiol 47, 3624-3629.
Becker, K., Friedrich, A.W., Lubritz, G., Weilert, M., Peters, G., Von Eiff, C., 2003. Prevalence of genes encoding pyrogenic toxin superantigens and exfoliative toxins among strains of Staphylococcus aureus isolated from blood and nasal specimens. J Clin Microbiol 41, 1434-1439.
Beekhuizen, H., van de Gevel, J.S., 2007. Gamma interferon confers resistance to infection with Staphylococcus aureus in human vascular endothelial cells by cooperative proinflammatory and enhanced intrinsic antibacterial activities. Infect Immun 75, 5615-5626.
Berends, E.T., Horswill, A.R., Haste, N.M., Monestier, M., Nizet, V., von Kockritz-Blickwede, M., 2010. Nuclease expression by Staphylococcus aureus facilitates escape from neutrophil extracellular traps. J Innate Immun 2, 576-586.
Birtles, A., McCarthy, N., Sheppard, C.L., Rutter, H., Guiver, M., Haworth, E., George, R.C., 2005. Multilocus sequence typing directly on DNA from clinical samples and a cultured isolate to investigate linked fatal pneumococcal disease in residents of a shelter for homeless men. J Clin Microbiol 43, 2004-2008.
Blair, J.E., Williams, R.E., 1961. Phage typing of staphylococci. Bull World Health Organ 24, 771-784. Bode, L.G., Kluytmans, J.A., Wertheim, H.F., Bogaers, D., Vandenbroucke-Grauls, C.M., Roosendaal,
R., Troelstra, A., Box, A.T., Voss, A., van der Tweel, I., van Belkum, A., Verbrugh, H.A., Vos, M.C., 2010. Preventing surgical-site infections in nasal carriers of Staphylococcus aureus. N Engl J Med 362, 9-17.
Bonomo, R.A., 2000. Multiple antibiotic-resistant bacteria in long-term-care facilities: An emerging problem in the practice of infectious diseases. Clin Infect Dis 31, 1414-1422.
Brown, E.M., Thomas, P., 2002. Fusidic acid resistance in Staphylococcus aureus isolates. Lancet 359, 803.
Buckingham Lela , F.M.L., 2007. Molecular Diagnostics fundamentals, methods, & clinical applications. F. A. Davis Company, Philadelphia.
Bustin, S.A., 2000. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J Mol Endocrinol 25, 169-193.
Chambers, H.F., Deleo, F.R., 2009. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat Rev Microbiol 7, 629-641.
Chamchod, F., Ruan, S., 2012. Modeling the spread of methicillin-resistant Staphylococcus aureus in nursing homes for elderly. PLoS One 7, e29757.
Chavakis, T., Preissner, K.T., Herrmann, M., 2007. The anti-inflammatory activities of Staphylococcus aureus. Trends Immunol 28, 408-418.
Chavakis, T., Wiechmann, K., Preissner, K.T., Herrmann, M., 2005. Staphylococcus aureus interactions with the endothelium: the role of bacterial "secretable expanded repertoire adhesive molecules" (SERAM) in disturbing host defense systems. Thromb Haemost 94, 278-285.
Chen, F.J., Siu, L.K., Lin, J.C., Wang, C.H., Lu, P.L., 2012. Molecular typing and characterization of nasal carriage and community-onset infection methicillin-susceptible Staphylococcus aureus isolates in two Taiwan medical centers. BMC Infect Dis 12, 343.

References
70
Cheng, A.G., DeDent, A.C., Schneewind, O., Missiakas, D., 2011. A play in four acts: Staphylococcus aureus abscess formation. Trends Microbiol 19, 225-232.
Cheung, A.L., Bayer, A.S., Zhang, G., Gresham, H., Xiong, Y.Q., 2004. Regulation of virulence determinants in vitro and in vivo in Staphylococcus aureus. FEMS Immunol Med Microbiol 40, 1-9.
Cines, D.B., Pollak, E.S., Buck, C.A., Loscalzo, J., Zimmerman, G.A., McEver, R.P., Pober, J.S., Wick, T.M., Konkle, B.A., Schwartz, B.S., Barnathan, E.S., McCrae, K.R., Hug, B.A., Schmidt, A.M., Stern, D.M., 1998. Endothelial cells in physiology and in the pathophysiology of vascular disorders. Blood 91, 3527-3561.
Coico Richard , S.G., 2009. Immunology a short course. John Wiley & Sons, New Jersey. Cookson, B.D., Aparicio, P., Deplano, A., Struelens, M., Goering, R., Marples, R., 1996. Inter-centre
comparison of pulsed-field gel electrophoresis for the typing of methicillin-resistant Staphylococcus aureus. J Med Microbiol 44, 179-184.
Cosgrove, K., Coutts, G., Jonsson, I.M., Tarkowski, A., Kokai-Kun, J.F., Mond, J.J., Foster, S.J., 2007. Catalase (KatA) and alkyl hydroperoxide reductase (AhpC) have compensatory roles in peroxide stress resistance and are required for survival, persistence, and nasal colonization in Staphylococcus aureus. J Bacteriol 189, 1025-1035.
Cunliffe, A.C., 1949. Incidence of Staphylococcus aureus in the anterior nares of healthy children. Lancet 2, 411-414.
Cuny, C., Friedrich, A., Kozytska, S., Layer, F., Nubel, U., Ohlsen, K., Strommenger, B., Walther, B., Wieler, L., Witte, W., 2010. Emergence of methicillin-resistant Staphylococcus aureus (MRSA) in different animal species. Int J Med Microbiol 300, 109-117.
David, M.Z., Taylor, A., Lynfield, R., Boxrud, D.J., Short, G., Zychowski, D., Boyle-Vavra, S., Daum, R.S., 2013. Comparing pulsed-field gel electrophoresis with multilocus sequence typing, spa typing, staphylococcal cassette chromosome mec (SCCmec) typing, and PCR for panton-valentine leukocidin, arcA, and opp3 in methicillin-resistant Staphylococcus aureus isolates at a U.S. Medical Center. J Clin Microbiol 51, 814-819.
de Kraker, M.E., Jarlier, V., Monen, J.C., Heuer, O.E., van de Sande, N., Grundmann, H., 2012. The changing epidemiology of bacteraemias in Europe: trends from the European Antimicrobial Resistance Surveillance System. Clin Microbiol Infect.
DeLeo, F.R., Chambers, H.F., 2009. Reemergence of antibiotic-resistant Staphylococcus aureus in the genomics era. J Clin Invest 119, 2464-2474.
den Heijer, C.D., van Bijnen, E.M., Paget, W.J., Pringle, M., Goossens, H., Bruggeman, C.A., Schellevis, F.G., Stobberingh, E.E., Team, A.S., 2013. Prevalence and resistance of commensal Staphylococcus aureus, including meticillin-resistant S aureus, in nine European countries: a cross-sectional study. Lancet Infect Dis 13, 409-415.
Dijkshoorn, L., Ursing, B.M., Ursing, J.B., 2000. Strain, clone and species: comments on three basic concepts of bacteriology. J Med Microbiol 49, 397-401.
Dinges, M.M., Orwin, P.M., Schlievert, P.M., 2000. Exotoxins of Staphylococcus aureus. Clin Microbiol Rev 13, 16-34, table of contents.
Doan Thao , M.R., Viselli Susan , Waltenbaugh Carl 2013. Immunology. Lippincott Williams & Wilkins, Baltimore and Philadelphia.
Edwards, A.M., Bowden, M.G., Brown, E.L., Laabei, M., Massey, R.C., 2012. Staphylococcus aureus extracellular adherence protein triggers TNFalpha release, promoting attachment to endothelial cells via protein A. PLoS One 7, e43046.
Enright, M.C., Day, N.P., Davies, C.E., Peacock, S.J., Spratt, B.G., 2000. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol 38, 1008-1015.
Enright, M.C., Robinson, D.A., Randle, G., Feil, E.J., Grundmann, H., Spratt, B.G., 2002. The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA). Proc Natl Acad Sci U S A 99, 7687-7692.

References
71
Eriksen, N.H., Espersen, F., Rosdahl, V.T., Jensen, K., 1995. Carriage of Staphylococcus aureus among 104 healthy persons during a 19-month period. Epidemiol Infect 115, 51-60.
Ernsth Bravell, M., Westerlind, B., Midlov, P., Ostgren, C.J., Borgquist, L., Lannering, C., Molstad, S., 2011. How to assess frailty and the need for care? Report from the Study of Health and Drugs in the Elderly (SHADES) in community dwellings in Sweden. Arch Gerontol Geriatr 53, 40-45.
European Center for Disease Prevention and Control (ECDC). Annual Surveillance Report. Reporting on 2009 surveillance data and 2010 epidemic intelligence data. Stockholm: ECDC, 2011. http://www.ecdc.europa.eu/en/publications/Publications/1111_SUR_Annual_Epidemiological_Report_on_Communicable_Diseases_in_Europe.pdf (accessed april 25, 2013)
Fischer, S.G., Lerman, L.S., 1983. DNA fragments differing by single base-pair substitutions are separated in denaturing gradient gels: correspondence with melting theory. Proc Natl Acad Sci U S A 80, 1579-1583.
Flock, J.I., Froman, G., Jonsson, K., Guss, B., Signas, C., Nilsson, B., Raucci, G., Hook, M., Wadstrom, T., Lindberg, M., 1987. Cloning and expression of the gene for a fibronectin-binding protein from Staphylococcus aureus. EMBO J 6, 2351-2357.
Foster, T.J., 2005. Immune evasion by staphylococci. Nat Rev Microbiol 3, 948-958. Foster, T.J., Hook, M., 1998. Surface protein adhesins of Staphylococcus aureus. Trends Microbiol 6,
484-488. Fraunholz, M., Sinha, B., 2012. Intracellular Staphylococcus aureus: live-in and let die. Front Cell
Infect Microbiol 2, 43. Friedrich, A.W., Daniels-Haardt, I., Kock, R., Verhoeven, F., Mellmann, A., Harmsen, D., van Gemert-
Pijnen, J.E., Becker, K., Hendrix, M.G., 2008. EUREGIO MRSA-net Twente/Munsterland--a Dutch-German cross-border network for the prevention and control of infections caused by methicillin-resistant Staphylococcus aureus. Euro Surveill 13.
Fuda, C., Hesek, D., Lee, M., Morio, K., Nowak, T., Mobashery, S., 2005a. Activation for catalysis of penicillin-binding protein 2a from methicillin-resistant Staphylococcus aureus by bacterial cell wall. J Am Chem Soc 127, 2056-2057.
Fuda, C.C., Fisher, J.F., Mobashery, S., 2005b. Beta-lactam resistance in Staphylococcus aureus: the adaptive resistance of a plastic genome. Cell Mol Life Sci 62, 2617-2633.
Gold, B., Hanson, M., Dean, M., 2001. Two rare confounding polymorphisms proximal to the factor V Leiden mutation. Mol Diagn 6, 137-140.
Goodyear, C.S., Silverman, G.J., 2003. Death by a B cell superantigen: In vivo VH-targeted apoptotic supraclonal B cell deletion by a Staphylococcal Toxin. J Exp Med 197, 1125-1139.
Grumann, D., Nubel, U., Broker, B.M., 2013. Staphylococcus aureus toxins - Their functions and genetics. Infect Genet Evol.
Grundmann, H., Aanensen, D.M., van den Wijngaard, C.C., Spratt, B.G., Harmsen, D., Friedrich, A.W., European Staphylococcal Reference Laboratory Working, G., 2010. Geographic distribution of Staphylococcus aureus causing invasive infections in Europe: a molecular-epidemiological analysis. PLoS Med 7, e1000215.
Grundmeier, M., Tuchscherr, L., Bruck, M., Viemann, D., Roth, J., Willscher, E., Becker, K., Peters, G., Loffler, B., 2010. Staphylococcal strains vary greatly in their ability to induce an inflammatory response in endothelial cells. J Infect Dis 201, 871-880.
Haggar, A., Ehrnfelt, C., Holgersson, J., Flock, J.I., 2004. The extracellular adherence protein from Staphylococcus aureus inhibits neutrophil binding to endothelial cells. Infect Immun 72, 6164-6167.
Hallin, M., Deplano, A., Denis, O., De Mendonca, R., De Ryck, R., Struelens, M.J., 2007. Validation of pulsed-field gel electrophoresis and spa typing for long-term, nationwide epidemiological surveillance studies of Staphylococcus aureus infections. J Clin Microbiol 45, 127-133.
Harmsen, D., Claus, H., Witte, W., Rothganger, J., Claus, H., Turnwald, D., Vogel, U., 2003. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J Clin Microbiol 41, 5442-5448.

References
72
Hartman, B., Tomasz, A., 1981. Altered penicillin-binding proteins in methicillin-resistant strains of Staphylococcus aureus. Antimicrob Agents Chemother 19, 726-735.
Hienz, S.A., Schennings, T., Heimdahl, A., Flock, J.I., 1996. Collagen binding of Staphylococcus aureus is a virulence factor in experimental endocarditis. J Infect Dis 174, 83-88.
Higuchi, R., Fockler, C., Dollinger, G., Watson, R., 1993. Kinetic PCR analysis: real-time monitoring of DNA amplification reactions. Biotechnology (N Y) 11, 1026-1030.
Hoeger, P.H., Lenz, W., Boutonnier, A., Fournier, J.M., 1992. Staphylococcal skin colonization in children with atopic dermatitis: prevalence, persistence, and transmission of toxigenic and nontoxigenic strains. J Infect Dis 165, 1064-1068.
Hu, L., Umeda, A., Kondo, S., Amako, K., 1995. Typing of Staphylococcus aureus colonising human nasal carriers by pulsed-field gel electrophoresis. J Med Microbiol 42, 127-132.
Hussain, M., von Eiff, C., Sinha, B., Joost, I., Herrmann, M., Peters, G., Becker, K., 2008. eap Gene as novel target for specific identification of Staphylococcus aureus. J Clin Microbiol 46, 470-476.
Imamura, T., Tanase, S., Szmyd, G., Kozik, A., Travis, J., Potempa, J., 2005. Induction of vascular leakage through release of bradykinin and a novel kinin by cysteine proteinases from Staphylococcus aureus. J Exp Med 201, 1669-1676.
Ito, T., Katayama, Y., Hiramatsu, K., 1999. Cloning and nucleotide sequence determination of the entire mec DNA of pre-methicillin-resistant Staphylococcus aureus N315. Antimicrob Agents Chemother 43, 1449-1458.
Jaffe, E.A., Nachman, R.L., Becker, C.G., Minick, C.R., 1973. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J Clin Invest 52, 2745-2756.
Jones, N., Bohnsack, J.F., Takahashi, S., Oliver, K.A., Chan, M.S., Kunst, F., Glaser, P., Rusniok, C., Crook, D.W., Harding, R.M., Bisharat, N., Spratt, B.G., 2003. Multilocus sequence typing system for group B streptococcus. J Clin Microbiol 41, 2530-2536.
Josefsson, E., Higgins, J., Foster, T.J., Tarkowski, A., 2008. Fibrinogen binding sites P336 and Y338 of clumping factor A are crucial for Staphylococcus aureus virulence. PLoS One 3, e2206.
Kahl, B.C., Mellmann, A., Deiwick, S., Peters, G., Harmsen, D., 2005. Variation of the polymorphic region X of the protein A gene during persistent airway infection of cystic fibrosis patients reflects two independent mechanisms of genetic change in Staphylococcus aureus. J Clin Microbiol 43, 502-505.
Kaneko, J., Kamio, Y., 2004. Bacterial two-component and hetero-heptameric pore-forming cytolytic toxins: structures, pore-forming mechanism, and organization of the genes. Biosci Biotechnol Biochem 68, 981-1003.
Kato, F., Kadomoto, N., Iwamoto, Y., Bunai, K., Komatsuzawa, H., Sugai, M., 2011. Regulatory mechanism for exfoliative toxin production in Staphylococcus aureus. Infect Immun 79, 1660-1670.
Kawabata, S., Morita, T., Miyata, T., Kaida, S., Igarashi, H., Iwanaga, S., 1986. Difference in enzymatic properties between "staphylothrombin" and free alpha-thrombin. Ann N Y Acad Sci 485, 27-40.
Kim, H.K., Thammavongsa, V., Schneewind, O., Missiakas, D., 2012. Recurrent infections and immune evasion strategies of Staphylococcus aureus. Curr Opin Microbiol 15, 92-99.
Kluytmans, J., van Belkum, A., Verbrugh, H., 1997. Nasal carriage of Staphylococcus aureus: epidemiology, underlying mechanisms, and associated risks. Clin Microbiol Rev 10, 505-520.
Kluytmans, J.A., Wertheim, H.F., 2005. Nasal carriage of Staphylococcus aureus and prevention of nosocomial infections. Infection 33, 3-8.
Koch, T.K., Reuter, M., Barthel, D., Bohm, S., van den Elsen, J., Kraiczy, P., Zipfel, P.F., Skerka, C., 2012. Staphylococcus aureus proteins Sbi and Efb recruit human plasmin to degrade complement C3 and C3b. PLoS One 7, e47638.
Krut, O., Sommer, H., Kronke, M., 2004. Antibiotic-induced persistence of cytotoxic Staphylococcus aureus in non-phagocytic cells. J Antimicrob Chemother 53, 167-173.

References
73
Kuehnert, M.J., Kruszon-Moran, D., Hill, H.A., McQuillan, G., McAllister, S.K., Fosheim, G., McDougal, L.K., Chaitram, J., Jensen, B., Fridkin, S.K., Killgore, G., Tenover, F.C., 2006. Prevalence of Staphylococcus aureus nasal colonization in the United States, 2001-2002. J Infect Dis 193, 172-179.
Lahteenmaki, K., Kuusela, P., Korhonen, T.K., 2000. Plasminogen activation in degradation and penetration of extracellular matrices and basement membranes by invasive bacteria. Methods 21, 125-132.
Larionov, A., Krause, A., Miller, W., 2005. A standard curve based method for relative real time PCR data processing. BMC Bioinformatics 6, 62.
Laupland, K.B., Church, D.L., Mucenski, M., Sutherland, L.R., Davies, H.D., 2003. Population-based study of the epidemiology of and the risk factors for invasive Staphylococcus aureus infections. J Infect Dis 187, 1452-1459.
Lee, C.J., Sankaran, S., Mukherjee, D.V., Apa, Z.L., Hafer, C.A., Wright, L., Larson, E.L., Lowy, F.D., 2011. Staphylococcus aureus oropharyngeal carriage in a prison population. Clin Infect Dis 52, 775-778.
Liu, W.T., Marsh, T.L., Cheng, H., Forney, L.J., 1997. Characterization of microbial diversity by determining terminal restriction fragment length polymorphisms of genes encoding 16S rRNA. Appl Environ Microbiol 63, 4516-4522.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402-408.
Lotze, M.T., Tracey, K.J., 2005. High-mobility group box 1 protein (HMGB1): nuclear weapon in the immune arsenal. Nat Rev Immunol 5, 331-342.
Lowy, F.D., 1998. Staphylococcus aureus infections. N Engl J Med 339, 520-532. Maiden, M.C., Bygraves, J.A., Feil, E., Morelli, G., Russell, J.E., Urwin, R., Zhang, Q., Zhou, J., Zurth, K.,
Caugant, D.A., Feavers, I.M., Achtman, M., Spratt, B.G., 1998. Multilocus sequence typing: a portable approach to the identification of clones within populations of pathogenic microorganisms. Proc Natl Acad Sci U S A 95, 3140-3145.
Marshall, C., Spelman, D., 2007. Re: is throat screening necessary to detect methicillin-resistant Staphylococcus aureus colonization in patients upon admission to an intensive care unit? J Clin Microbiol 45, 3855.
Massimi, I., Park, E., Rice, K., Muller-Esterl, W., Sauder, D., McGavin, M.J., 2002. Identification of a novel maturation mechanism and restricted substrate specificity for the SspB cysteine protease of Staphylococcus aureus. J Biol Chem 277, 41770-41777.
Matussek, A., Stark, L., Dienus, O., Aronsson, J., Mernelius, S., Lofgren, S., Lindgren, P.E., 2011. Analyzing multiclonality of Staphylococcus aureus in clinical diagnostics using spa-based denaturing gradient gel electrophoresis. J Clin Microbiol 49, 3647-3648.
Matussek, A., Taipalensuu, J., Einemo, I.M., Tiefenthal, M., Lofgren, S., 2007. Transmission of Staphylococcus aureus from maternity unit staff members to newborns disclosed through spa typing. Am J Infect Control 35, 122-125.
Mazmanian, S.K., Liu, G., Jensen, E.R., Lenoy, E., Schneewind, O., 2000. Staphylococcus aureus sortase mutants defective in the display of surface proteins and in the pathogenesis of animal infections. Proc Natl Acad Sci U S A 97, 5510-5515.
McAdow, M., Missiakas, D.M., Schneewind, O., 2012. Staphylococcus aureus secretes coagulase and von Willebrand factor binding protein to modify the coagulation cascade and establish host infections. J Innate Immun 4, 141-148.
McCarthy, A.J., Breathnach, A.S., Lindsay, J.A., 2012. Detection of mobile-genetic-element variation between colonizing and infecting hospital-associated methicillin-resistant Staphylococcus aureus isolates. J Clin Microbiol 50, 1073-1075.
McCarthy, A.J., Lindsay, J.A., 2012. The distribution of plasmids that carry virulence and resistance genes in Staphylococcus aureus is lineage associated. BMC Microbiol 12, 104.
Medzhitov, R., Janeway, C.A., Jr., 1998. Self-defense: the fruit fly style. Proc Natl Acad Sci U S A 95, 429-430.

References
74
Mehndiratta, P.L., Bhalla, P., 2012. Typing of Methicillin resistant Staphylococcus aureus: a technical review. Indian J Med Microbiol 30, 16-23.
Mehndiratta, P.L., Bhalla, P., Ahmed, A., Sharma, Y.D., 2009. Molecular typing of methicillin-resistant Staphylococcus aureus strains by PCR-RFLP of SPA gene: a reference laboratory perspective. Indian J Med Microbiol 27, 116-122.
Melin, S., Melin, S., Haeggman, S., Melin, S., Haeggman, S., Olsson-Liljequist, B., Sjolund, M., Nilsson, P.A., Isaksson, B., Lofgren, S., Matussek, A., 2009. Epidemiological typing of methicillin-resistant Staphylococcus aureus (MRSA): spa typing versus pulsed-field gel electrophoresis. Scand J Infect Dis 41, 433-439.
Melles, D.C., Gorkink, R.F., Boelens, H.A., Snijders, S.V., Peeters, J.K., Moorhouse, M.J., van der Spek, P.J., van Leeuwen, W.B., Simons, G., Verbrugh, H.A., van Belkum, A., 2004. Natural population dynamics and expansion of pathogenic clones of Staphylococcus aureus. J Clin Invest 114, 1732-1740.
Melles, D.C., van Leeuwen, W.B., Snijders, S.V., Horst-Kreft, D., Peeters, J.K., Verbrugh, H.A., van Belkum, A., 2007. Comparison of multilocus sequence typing (MLST), pulsed-field gel electrophoresis (PFGE), and amplified fragment length polymorphism (AFLP) for genetic typing of Staphylococcus aureus. J Microbiol Methods 69, 371-375.
Mernelius, S., Lofgren, S., Lindgren, P.E., Blomberg, M., Olhager, E., Gunnervik, C., Lenrick, R., Thrane, M.T., Isaksson, B., Matussek, A., 2013a. The effect of improved compliance with hygiene guidelines on transmission of Staphylococcus aureus to newborn infants: The Swedish Hygiene Intervention and Transmission of S aureus study. Am J Infect Control.
Mernelius, S., Lofgren, S., Lindgren, P.E., Matussek, A., 2013b. The role of broth enrichment in Staphylococcus aureus cultivation and transmission from the throat to newborn infants: results from the Swedish hygiene intervention and transmission of S. aureus study. Eur J Clin Microbiol Infect Dis.
Miko, B.A., Hafer, C.A., Lee, C.J., Sullivan, S.B., Hackel, M.A., Johnson, B.M., Whittier, S., Della-Latta, P., Uhlemann, A.C., Lowy, F.D., 2013. Molecular characterization of methicillin-susceptible Staphylococcus aureus clinical isolates in the United States, 2004 to 2010. J Clin Microbiol 51, 874-879.
Monecke, S., Coombs, G., Shore, A.C., Coleman, D.C., Akpaka, P., Borg, M., Chow, H., Ip, M., Jatzwauk, L., Jonas, D., Kadlec, K., Kearns, A., Laurent, F., O'Brien, F.G., Pearson, J., Ruppelt, A., Schwarz, S., Scicluna, E., Slickers, P., Tan, H.L., Weber, S., Ehricht, R., 2011. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS One 6, e17936.
Monti, G., Tonetto, P., Mostert, M., Oggero, R., 1996. Staphylococcus aureus skin colonization in infants with atopic dermatitis. Dermatology 193, 83-87.
Moreillon, P., Entenza, J.M., Francioli, P., McDevitt, D., Foster, T.J., Francois, P., Vaudaux, P., 1995. Role of Staphylococcus aureus coagulase and clumping factor in pathogenesis of experimental endocarditis. Infect Immun 63, 4738-4743.
Mulligan, M.E., Arbeit, R.D., 1991. Epidemiologic and clinical utility of typing systems for differentiating among strains of methicillin-resistant Staphylococcus aureus. Infect Control Hosp Epidemiol 12, 20-28.
Mullis, K., Faloona, F., Scharf, S., Saiki, R., Horn, G., Erlich, H., 1986. Specific enzymatic amplification of DNA in vitro: the polymerase chain reaction. Cold Spring Harb Symp Quant Biol 51 Pt 1, 263-273.
Mulvey, M.R., Chui, L., Ismail, J., Louie, L., Murphy, C., Chang, N., Alfa, M., Canadian Committee for the Standardization of Molecular, M., 2001. Development of a Canadian standardized protocol for subtyping methicillin-resistant Staphylococcus aureus using pulsed-field gel electrophoresis. J Clin Microbiol 39, 3481-3485.
Muyzer, G., de Waal, E.C., Uitterlinden, A.G., 1993. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59, 695-700.

References
75
Nilsson, P., Ripa, T., 2006. Staphylococcus aureus throat colonization is more frequent than colonization in the anterior nares. J Clin Microbiol 44, 3334-3339.
Noble, W.C., Valkenburg, H.A., Wolters, C.H., 1967. Carriage of Staphylococcus aureus in random samples of a normal population. J Hyg (Lond) 65, 567-573.
Noskin, G.A., Rubin, R.J., Schentag, J.J., Kluytmans, J., Hedblom, E.C., Smulders, M., Lapetina, E., Gemmen, E., 2005. The burden of Staphylococcus aureus infections on hospitals in the United States: an analysis of the 2000 and 2001 Nationwide Inpatient Sample Database. Arch Intern Med 165, 1756-1761.
Nouwen, J.L., Ott, A., Kluytmans-Vandenbergh, M.F., Boelens, H.A., Hofman, A., van Belkum, A., Verbrugh, H.A., 2004. Predicting the Staphylococcus aureus nasal carrier state: derivation and validation of a "culture rule". Clin Infect Dis 39, 806-811.
O'Neill, A.J., Chopra, I., 2006. Molecular basis of fusB-mediated resistance to fusidic acid in Staphylococcus aureus. Mol Microbiol 59, 664-676.
O'Neill, A.J., Larsen, A.R., Henriksen, A.S., Chopra, I., 2004. A fusidic acid-resistant epidemic strain of Staphylococcus aureus carries the fusB determinant, whereas fusA mutations are prevalent in other resistant isolates. Antimicrob Agents Chemother 48, 3594-3597.
O'Riordan, K., Lee, J.C., 2004. Staphylococcus aureus capsular polysaccharides. Clin Microbiol Rev 17, 218-234.
Ogawa, S.K., Yurberg, E.R., Hatcher, V.B., Levitt, M.A., Lowy, F.D., 1985. Bacterial adherence to human endothelial cells in vitro. Infect Immun 50, 218-224.
Olive, D.M., Bean, P., 1999. Principles and applications of methods for DNA-based typing of microbial organisms. J Clin Microbiol 37, 1661-1669.
Oliveira, D.C., de Lencastre, H., 2011. Methicillin-resistance in Staphylococcus aureus is not affected by the overexpression in trans of the mecA gene repressor: a surprising observation. PLoS One 6, e23287.
Oliveira, D.C., Tomasz, A., de Lencastre, H., 2002. Secrets of success of a human pathogen: molecular evolution of pandemic clones of meticillin-resistant Staphylococcus aureus. Lancet Infect Dis 2, 180-189.
Olofsson, M., Lindgren, P.E., Ostgren, C.J., Midlov, P., Molstad, S., 2012. Colonization with Staphylococcus aureus in Swedish nursing homes: a cross-sectional study. Scand J Infect Dis 44, 3-8.
Olofsson, M., Toepfer, M., Ostgren, C.J., Midlov, P., Matussek, A., Lindgren, P.E., Molstad, S., 2013. Low level of antimicrobial resistance in Escherichia coli among Swedish nursing home residents. Scand J Infect Dis 45, 117-123.
Olsen, K., Danielsen, K., Wilsgaard, T., Sangvik, M., Sollid, J.U., Thune, I., Eggen, A.E., Simonsen, G.S., Furberg, A.S., 2013. Obesity and Staphylococcus aureus Nasal Colonization among Women and Men in a General Population. PLoS One 8, e63716.
Olsen, K., Falch, B.M., Danielsen, K., Johannessen, M., Ericson Sollid, J.U., Thune, I., Grimnes, G., Jorde, R., Simonsen, G.S., Furberg, A.S., 2012. Staphylococcus aureus nasal carriage is associated with serum 25-hydroxyvitamin D levels, gender and smoking status. The Tromso Staph and Skin Study. Eur J Clin Microbiol Infect Dis 31, 465-473.
Osterlund, A., Eden, T., Olsson-Liljequist, B., Haeggman, S., Kahlmeter, G., Swedish Study Group on Fusidic Acid-resistant Staphylococcus, a., 2002. Clonal spread among Swedish children of a Staphylococcus aureus strain resistant to fusidic acid. Scand J Infect Dis 34, 729-734.
Pallen, M.J., Loman, N.J., Penn, C.W., 2010. High-throughput sequencing and clinical microbiology: progress, opportunities and challenges. Curr Opin Microbiol 13, 625-631.
Parras, F., Guerrero, M.C., Bouza, E., Blazquez, M.J., Moreno, S., Menarguez, M.C., Cercenado, E., 1995. Comparative study of mupirocin and oral co-trimoxazole plus topical fusidic acid in eradication of nasal carriage of methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother 39, 175-179.

References
76
Peacock, S.J., Moore, C.E., Justice, A., Kantzanou, M., Story, L., Mackie, K., O'Neill, G., Day, N.P., 2002. Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infect Immun 70, 4987-4996.
Perl, T.M., Cullen, J.J., Wenzel, R.P., Zimmerman, M.B., Pfaller, M.A., Sheppard, D., Twombley, J., French, P.P., Herwaldt, L.A., Mupirocin, The Risk Of Staphylococcus Aureus Study, T., 2002. Intranasal mupirocin to prevent postoperative Staphylococcus aureus infections. N Engl J Med 346, 1871-1877.
Peter, W., 2006. Understanding immunology. Pearson Education Limited, Essex, Great Britain. Pfaffl, M.W., 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic
Acids Res 29, e45. Pfingsten-Wurzburg, S., Pieper, D.H., Bautsch, W., Probst-Kepper, M., 2011. Prevalence and
molecular epidemiology of meticillin-resistant Staphylococcus aureus in nursing home residents in northern Germany. J Hosp Infect 78, 108-112.
Pinchuk, I.V., Beswick, E.J., Reyes, V.E., 2010. Staphylococcal enterotoxins. Toxins (Basel) 2, 2177-2197.
Potempa, J., Watorek, W., Travis, J., 1986. The inactivation of human plasma alpha 1-proteinase inhibitor by proteinases from Staphylococcus aureus. J Biol Chem 261, 14330-14334.
Prokesova, L., Potuznikova, B., Potempa, J., Zikan, J., Radl, J., Hachova, L., Baran, K., Porwit-Bobr, Z., John, C., 1992. Cleavage of human immunoglobulins by serine proteinase from Staphylococcus aureus. Immunol Lett 31, 259-265.
Rakeman, J.L., Bui, U., Lafe, K., Chen, Y.C., Honeycutt, R.J., Cookson, B.T., 2005. Multilocus DNA sequence comparisons rapidly identify pathogenic molds. J Clin Microbiol 43, 3324-3333.
Reimer, L.G., Wilson, M.L., Weinstein, M.P., 1997. Update on detection of bacteremia and fungemia. Clin Microbiol Rev 10, 444-465.
Roben, P.W., Salem, A.N., Silverman, G.J., 1995. VH3 family antibodies bind domain D of staphylococcal protein A. J Immunol 154, 6437-6445.
Rooijakkers, S.H., van Kessel, K.P., van Strijp, J.A., 2005. Staphylococcal innate immune evasion. Trends Microbiol 13, 596-601.
Sabat, A.J., Budimir, A., Nashev, D., Sa-Leao, R., van Dijl, J., Laurent, F., Grundmann, H., Friedrich, A.W., Markers, E.S.G.o.E., 2013. Overview of molecular typing methods for outbreak detection and epidemiological surveillance. Euro Surveill 18, 20380.
Sakwinska, O., Blanc, D.S., Lazor-Blanchet, C., Moreillon, M., Giddey, M., Moreillon, P., 2010. Ecological temporal stability of Staphylococcus aureus nasal carriage. J Clin Microbiol 48, 2724-2728.
Sandberg, A., Hessler, J.H., Skov, R.L., Blom, J., Frimodt-Moller, N., 2009. Intracellular activity of antibiotics against Staphylococcus aureus in a mouse peritonitis model. Antimicrob Agents Chemother 53, 1874-1883.
Sangvik, M., Olsen, R.S., Olsen, K., Simonsen, G.S., Furberg, A.S., Sollid, J.U., 2011. Age- and gender-associated Staphylococcus aureus spa types found among nasal carriers in a general population: the Tromso Staph and Skin Study. J Clin Microbiol 49, 4213-4218.
Scriba, T.J., Sierro, S., Brown, E.L., Phillips, R.E., Sewell, A.K., Massey, R.C., 2008. The Staphyloccous aureus Eap protein activates expression of proinflammatory cytokines. Infect Immun 76, 2164-2168.
Selander, R.K., Caugant, D.A., Ochman, H., Musser, J.M., Gilmour, M.N., Whittam, T.S., 1986. Methods of multilocus enzyme electrophoresis for bacterial population genetics and systematics. Appl Environ Microbiol 51, 873-884.
Shopsin, B., Gomez, M., Montgomery, S.O., Smith, D.H., Waddington, M., Dodge, D.E., Bost, D.A., Riehman, M., Naidich, S., Kreiswirth, B.N., 1999. Evaluation of protein A gene polymorphic region DNA sequencing for typing of Staphylococcus aureus strains. J Clin Microbiol 37, 3556-3563.
Sila, J., Sauer, P., Kolar, M., 2009. Comparison of the prevalence of genes coding for enterotoxins, exfoliatins, panton-valentine leukocidin and tsst-1 between methicillin-resistant and

References
77
methicillin-susceptible isolates of Staphylococcus aureus at the university hospital in olomouc. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub 153, 215-218.
Smith, E.J., Visai, L., Kerrigan, S.W., Speziale, P., Foster, T.J., 2011. The Sbi protein is a multifunctional immune evasion factor of Staphylococcus aureus. Infect Immun 79, 3801-3809.
Song, M.D., Wachi, M., Doi, M., Ishino, F., Matsuhashi, M., 1987. Evolution of an inducible penicillin-target protein in methicillin-resistant Staphylococcus aureus by gene fusion. FEBS Lett 221, 167-171.
Stefani, S., Chung, D.R., Lindsay, J.A., Friedrich, A.W., Kearns, A.M., Westh, H., Mackenzie, F.M., 2012. Meticillin-resistant Staphylococcus aureus (MRSA): global epidemiology and harmonisation of typing methods. Int J Antimicrob Agents 39, 273-282.
Strindhall, J., Lindgren, P.E., Lofgren, S., Kihlstrom, E., 2002. Variations among clinical isolates of Staphylococcus aureus to induce expression of E-selectin and ICAM-1 in human endothelial cells. FEMS Immunol Med Microbiol 32, 227-235.
Strommenger, B., Kettlitz, C., Weniger, T., Harmsen, D., Friedrich, A.W., Witte, W., 2006. Assignment of Staphylococcus isolates to groups by spa typing, SmaI macrorestriction analysis, and multilocus sequence typing. J Clin Microbiol 44, 2533-2540.
Struelens, M.J., Hawkey, P.M., French, G.L., Witte, W., Tacconelli, E., 2009. Laboratory tools and strategies for methicillin-resistant Staphylococcus aureus screening, surveillance and typing: state of the art and unmet needs. Clin Microbiol Infect 15, 112-119.
Su, C., Lei, L., Duan, Y., Zhang, K.Q., Yang, J., 2012. Culture-independent methods for studying environmental microorganisms: methods, application, and perspective. Appl Microbiol Biotechnol 93, 993-1003.
Tenover, F.C., Arbeit, R.D., Goering, R.V., 1997. How to select and interpret molecular strain typing methods for epidemiological studies of bacterial infections: a review for healthcare epidemiologists. Molecular Typing Working Group of the Society for Healthcare Epidemiology of America. Infect Control Hosp Epidemiol 18, 426-439.
Tenover, F.C., Arbeit, R.D., Goering, R.V., Mickelsen, P.A., Murray, B.E., Persing, D.H., Swaminathan, B., 1995. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol 33, 2233-2239.
Tveten, Y., Jenkins, A., Kristiansen, B.E., 2002. A fusidic acid-resistant clone of Staphylococcus aureus associated with impetigo bullosa is spreading in Norway. J Antimicrob Chemother 50, 873-876.
van Belkum, A., Emonts, M., Wertheim, H., de Jongh, C., Nouwen, J., Bartels, H., Cole, A., Cole, A., Hermans, P., Boelens, H., Toom, N.L., Snijders, S., Verbrugh, H., van Leeuwen, W., 2007. The role of human innate immune factors in nasal colonization by Staphylococcus aureus. Microbes Infect 9, 1471-1477.
van Belkum, A., van Leeuwen, W., Kaufmann, M.E., Cookson, B., Forey, F., Etienne, J., Goering, R., Tenover, F., Steward, C., O'Brien, F., Grubb, W., Tassios, P., Legakis, N., Morvan, A., El Solh, N., de Ryck, R., Struelens, M., Salmenlinna, S., Vuopio-Varkila, J., Kooistra, M., Talens, A., Witte, W., Verbrugh, H., 1998. Assessment of resolution and intercenter reproducibility of results of genotyping Staphylococcus aureus by pulsed-field gel electrophoresis of SmaI macrorestriction fragments: a multicenter study. J Clin Microbiol 36, 1653-1659.
van Belkum, A., Verkaik, N.J., de Vogel, C.P., Boelens, H.A., Verveer, J., Nouwen, J.L., Verbrugh, H.A., Wertheim, H.F., 2009. Reclassification of Staphylococcus aureus nasal carriage types. J Infect Dis 199, 1820-1826.
van den Akker, E.L., Nouwen, J.L., Melles, D.C., van Rossum, E.F., Koper, J.W., Uitterlinden, A.G., Hofman, A., Verbrugh, H.A., Pols, H.A., Lamberts, S.W., van Belkum, A., 2006. Staphylococcus aureus nasal carriage is associated with glucocorticoid receptor gene polymorphisms. J Infect Dis 194, 814-818.
van der Donk, C.F., Rijnders, M.I., Donker, G.A., de Neeling, A.J., Nys, S., Stobberingh, E.E., 2013. Is living in a border region a risk for a high prevalence of resistance? Eur J Clin Microbiol Infect Dis 32, 989-995.

References
78
VandenBergh, M.F., Yzerman, E.P., van Belkum, A., Boelens, H.A., Sijmons, M., Verbrugh, H.A., 1999. Follow-up of Staphylococcus aureus nasal carriage after 8 years: redefining the persistent carrier state. J Clin Microbiol 37, 3133-3140.
Varga, D.T., White, A., 1961. Suppression of nasal, skin, and aerial staphylococci by nasal application of methicillin. J Clin Invest 40, 2209-2214.
Wertheim, H.F., Melles, D.C., Vos, M.C., van Leeuwen, W., van Belkum, A., Verbrugh, H.A., Nouwen, J.L., 2005a. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect Dis 5, 751-762.
Wertheim, H.F., van Leeuwen, W.B., Snijders, S., Vos, M.C., Voss, A., Vandenbroucke-Grauls, C.M., Kluytmans, J.A., Verbrugh, H.A., van Belkum, A., 2005b. Associations between Staphylococcus aureus Genotype, Infection, and In-Hospital Mortality: A Nested Case-Control Study. J Infect Dis 192, 1196-1200.
Wertheim, H.F., Vos, M.C., Ott, A., van Belkum, A., Voss, A., Kluytmans, J.A., van Keulen, P.H., Vandenbroucke-Grauls, C.M., Meester, M.H., Verbrugh, H.A., 2004. Risk and outcome of nosocomial Staphylococcus aureus bacteraemia in nasal carriers versus non-carriers. Lancet 364, 703-705.
Williams, R.E., 1963. Healthy carriage of Staphylococcus aureus: its prevalence and importance. Bacteriol Rev 27, 56-71.
Winn Washington , A.S., Janda William , Koneman Elmer , Procop Gary , Schreckenberger Paul , Woods Gail 2006. Koneman's Color Atlas and Textbook of Diagnostic Microbiology Lippincott Williams & Wilkins, Philadelphia.
von Eiff, C., Becker, K., Machka, K., Stammer, H., Peters, G., 2001. Nasal carriage as a source of Staphylococcus aureus bacteremia. Study Group. N Engl J Med 344, 11-16.
Vriesema, A.J., Beekhuizen, H., Hamdi, M., Soufan, A., Lammers, A., Willekens, B., Bakker, O., Welten, A.G., Veltrop, M.H., van De Gevel, J.S., Dankert, J., Zaat, S.A., 2000. Altered gene expression in Staphylococcus aureus upon interaction with human endothelial cells. Infect Immun 68, 1765-1772.
Yagupsky, P., Nolte, F.S., 1990. Quantitative aspects of septicemia. Clin Microbiol Rev 3, 269-279. Zecconi, A., Scali, F., 2013. Staphylococcus aureus virulence factors in evasion from innate immune
defenses in human and animal diseases. Immunol Lett 150, 12-22.