springer 6wdeoh 85
TRANSCRIPT

Springerhttp://www.jstor.org/stable/4601071 .
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Springer is collaborating with JSTOR to digitize, preserve and extend access to Behavioral Ecology andSociobiology.
http://www.jstor.org
This content downloaded from 130.212.18.200 on Thu, 20 Jun 2013 12:24:00 PMAll use subject to JSTOR Terms and Conditions

Behav Ecol Sociobiol (1995) 36: 243-250 ? Springer-Verlag 1995
Jan A. Randall Susan M. Hatch Evon R. Hekkala
Interspecific variation in anti-predator behavior in sympatric species of kangaroo rat
Received: 10 March 1994/Accepted after revision: 26 November 1994
Abstract Kangaroo rats, Dipodomys, occupy desert habitats with little cover and thus are under high pre- dation risk from diverse predators. The behavior used to assess predation risk or to escape capture is unknown. We therefore compared anti-predator behav- ior of two sympatric species of kangaroo rat of different sizes, D. merriami and D. spectabilis. We first exam- ined whether kangaroo rats use olfaction as a first line of defense against snake predation and tested the rats for their responses to scent extracted from two species of snake that live sympatrically with the kangaroo rats, the Mojave rattlesnake (Crotalus scutulatus) and the gopher snake (Pitophis melanoleucus). We also tested for species differences in anti-predator behavior through 1 5-min interactions between the kangaroo rats and free-moving gopher snakes. We found that D. spectabilis actively approached the scent of both rattlesnakes and gopher snakes more than controls of vegetable oil and evaporated solvent (Fig. 1). In con- trast, D. merriami did not differentiate snake odors from controls in the experimental arena, but they sniffed the sand where a free-moving snake had passed more than D. spectabilis. Both species successfully avoided predation in encounters with live snakes. Although total numbers of approaches and with- drawals were similar (Fig. 2), D. spectabilis spent significantly more time within striking distance of the snake than D. merriami. D. spectabilis approached the head of the snake in 930/, of its approaches and often engaged in nose to snout contact with the snake. If the snake struck, D. spectabilis jumped directly backward to avoid a strike and footdrummed at a safe distance. In contrast, D. merriami oriented to the snake more than D. spectabilis, but approached the head in only 41 %, of the approaches and rarely engaged in nose-to-
J. A. Randall (5A) S. M. Hatch E. R. Hekkala Department of Biology, San Francisco State University, San Francisco, California 94132, USA
snout contact. The snakes struck, hissed and decreased predatory approaches with D. spectabilis but not with D. merriami (Fig. 3). These results show that kanga- roo rats can behaviorally influence the risk of being preyed on by snakes. The two species differ, however, in how they react to snakes. The larger D. spectabilis confronts snakes while the smaller D. merriami moni- tors snakes from a safe distance and avoids them.
Key words Snake Anti-predator Dipodomys
Introduction
Predation is a major selection pressure on the behav- ior of small mammals (Lima and Dill 1990). In small desert rodents, predation risk constitutes an important foraging cost (Brown 1988). Mating, territorial defense and dispersal also expose desert rodents to predators (Daly et al. 1990; Randall 1993). High predation risk favors the evolution of large groups, but many small mammals are solitary and cannot rely on group pro- tection. Instead, solitary species must employ individ- ual defenses which often consist of cryptic coloration and avoidance of predators (Pulliam and Caraco 1984).
Kangaroo rats (Dipodomys; Heteromyidae) are soli- tary rodents inhabiting desert habitats with little cover. These rodents are considered to be at high predation risk and under constant pressure to minimize preda- tion from a variety of diverse predators: birds (barn owls, Tyto alba; great horned owls, Bubo virginianus; shrikes, Lanius ludovicianus), large mammals (coyotes, Canis latrans; kit foxes, Vulpes macrotis; bobcats, Felis rufus; badgers, Taxidea taxus) and both venomous and nonvenomous snakes (rattlesnakes, Crotalus spp; gopher snakes, Pituophis melanoleucus; whip snakes Masticophis flagellum) (Kotler 1984; Randall and Stevens 1987; Kotler et al. 1988; Daly et al. 1990, 1992; Longland and Price 1991; Pierce et al. 1992).
This content downloaded from 130.212.18.200 on Thu, 20 Jun 2013 12:24:00 PMAll use subject to JSTOR Terms and Conditions

244
At least three morphological and behavioral char- acteristics of kangaroo rats have been proposed as adaptations for predator avoidance in open terrain. First, bipedal locomotion provides speed and an erratic, zig-zagging path of escape (Bartholomew and Caswell 1951; Djawdan and Garland 1988; Djawdan 1993). Bipedality also enables kangaroo rats to escape attacks by snakes and aerial predators at close prox- imity; they leap perpendicular to the flight path of owls (Longland and Price 1991) and backward in response to snake strikes (Webster 1962). The enlarged tympanic bullae and hypertrophied middle ear volumes give kan- garoo rats excellent hearing, which may be an adapta- tion to detect low-frequency sounds made by approaching predators (Webster 1962; Webster and Webster 1971). This sensitivity to low-frequency sounds seems to be an adaptation to desert conditions, because non-desert heteromyids (Heteromys and Liomys) have relatively poor low-frequency hearing when compared to desert species (Webster and Webster 1984). Finally, avoidance of bright illumination provided by moon- light or its equivalent in artificial light decreases pre- dation from nocturnal predators, primarily owls (Kotler 1984, 1992; Kotler et al. 1988, 1993; Longland and Price 1991; Daly et al. 1992).
Recent studies suggest species differences in how desert rodents respond to predation by snakes (Pierce et al. 1992; Kotler et al. 1993). However, because no behavioral observations were made in these studies, strategies used by desert rodents to assess predation risk or to escape capture from snakes are unknown. Desert rodents are probably similar to ground-dwelling squir- rels that live in open grasslands.They live in snake- inhabited environments and have developed specific predator defenses through millennia of coevolution with snakes (Goldthwaite et al. 1990; Towers and Coss 1991).
We selected two distinctly different, but sympatric, species of kangaroo rats for a comparison of anti- predator behavior directed at snakes by desert rodents (Randall 1993). The 130-g banner-tailed kangaroo rat, Dipodomys spectabilis, is territorial. Central to the indi- vidual territory of male and female rats are one or more large dirt mounds (approximately 3 m wide, 0.5 m high, and spaced 10-50 m apart) and the area immediately surrounding them (Schroder 1979; Randall 1984). Mounds are a scarce resource and contain burrows essential for shelter, seed storage and reproduction (Best 1972; Jones 1984). D. spectabilis footdrums indi- vidually distinct footdrumming signatures and chases away conspecific intruders in territorial defense (Randall 1984, 1989a, 1993, 1994). D. spectabilis also footdrums in the presence of snakes (Randall and Stevens 1987). In contrast, the approximately 40-g D. merriami occupies overlapping home ranges in which males overlap females and each other (Randall 1993). The rodents may use at least three diferent bur- rows in their home areas. They do not footdrum in
either a social or defensive context. D. merriami appears more social than D. spectabilis and tolerates neighbors in areas of homerange overlap (Randall 1989b).
We had two goals in this study. We first examined whether kangaroo rats use olfaction to detect snakes for subsequent defense. Detection of the presence of snakes by odor could be especially effective in locating an ambush predator that is motionless and visually cryptic on a dark night or in a burrow (Hennessy and Owings 1978; Towers and Coss 1990). In a previous study, D. spectabilis either approached and sniffed a tethered snake or sniffed the ground after a live snake had been there (Randall and Stevens 1987). We also examined species differences in anti-predator behavior through staged rat-snake interactions under controlled laboratory conditions. We predicted that the size and social differences in the two species could lead to different anti-predator behavior.
Methods
Animals
D. merriami was captured in the wild in 1987 and 1988 and D. spectabilis in 1988 near Portal, Arizona. The rats were main- tained in the laboratory on a 14:10 L:D light cycle in 42x22 x 20 cm plastic cages on 4 cm of sand with a tin can "burrow". The cages were cleaned and the sand changed weekly, and all rats were fed on altemate days with wild bird seed and a leaf of iceberg lettuce.
Gopher snakes, Pitophis melanoleucus, were used to stimulate anti-predator behavior in the two species of kangaroo rats. We chose gopher snakes because they had been observed preying on kangaroo rats in the field (Randall and Stevens 1987 and unpub- lished observations) and to eliminate the difficulties of handling ven- omous snakes. A total of seven snakes large enough to prey on kangaroo rats were used in the study. Six P. m. affinis, captured by hand on the roadsides near Portal, Arizona, in May-August 1990, ranged in snout-vent lengths from 0.75 to 1.2 m and weighed 200-650 g. Each of these snakes was used in an encounter from two to four times with each species of rat. The seventh snake, P. m. catenifer, which was 1 m long and weighed 980 g, was cap- tured in the Bay area and used in only one encounter with each species. We based our selection of snakes for an encounter on size and length of time since consumption of the last meal. Larger snakes were paired with larger rats and all snakes were deprived of food from 2-4 weeks to increase predatory behavior. The snakes were individually housed in glass and wood terrariums averaging 0.70 x 0.55 x 0.45 m with a 5 cm layer of Bed-o-cob at an average room temperature of 26?C and a 12L: 12D cycle. Live mice were offered weekly to all snakes when not in the study and water was present ad lib.
Procedures for olfactory Tests
A total of 19 sexually mature D. spectabilis (10 males and 9 females) and 24 D. merriami ( 12 males and 12 females) were tested four times each with four different stimuli, two controls and two snake odors, in a counterbalanced order in January 1990. Nine D. spectabilis and 12 D. mnerriami were tested each day in alternating sex and stimulus patterns so that 2 days separated each stimulus presenta- tion to a test subject.
This content downloaded from 130.212.18.200 on Thu, 20 Jun 2013 12:24:00 PMAll use subject to JSTOR Terms and Conditions

245
Snake odor consisted of lipids extracted from the skin (Graves and Halpern 1988) of two different species of snake that live sym- patrically with the kangaroo rats, the Mojave rattlesnake (Crotalus scutulatus) and the gopher snake. Snake scent was derived from fresh-frozen, road-killed snakes that had been thawed, rinsed with warm water and soaked for 12 h in a solution of methylene chlo- ride under an evaporation hood. (Care was taken to ensure that the head and cloacal region and any damaged tissue did not come into contact with the soaking solution.) The extract in the methyl- ene chloride was poured into a clean, weighed beaker and the sol- vent was allowed to evaporate, leaving a oily residue in the bottom of the beaker. We standardized concentrations by weighing the residue and redissolving it in methylene chloride in a ratio of 1 mil- ligram residue in 0.5 ml solvent. The other two stimuli were con- trols of methylene chloride solvent and 1 mg vegetable oil in 0.5 ml methylene chloride.
We tested the rats on 1-2 cm of clean sand in two different gal- vanized steel arenas with removeable sides that were scaled to the size of the rats: 70 cm in diameter and 45 cm high for D. merriami and 90 cm in diameter and 70 cm high for D. spectabilis. The rats had been habituated to the arenas for 5 min 2 weeks prior to the tests. Immediately before each test, either 0.5 ml of methylene chlo- ride or I mg of vegetable oil or snake extract in 0.5 ml of methyl- ene chloride was pipetted and mixed with a clean toothpick into 5.0 ml of clean sand and allowed to evaporate for 10 min. We placed the 5 ml of sand containing the stimulus in the center of one quad- rant of the arena, released the test subject in the center of the arena and timed the location of the rat in either the quadrant containing the stimulus or in the opposite quadrant (far) for each minute of a 4-min test. The other two quadrants were considered neutral areas. After each test, we removed the soiled sand, wiped clean the bot- tom of the arena with 95% ethanol, and poured a thin layer of new sand to cover the bottom of the arena. The arena was rotated clock- wise 450 for each test to control for position effects in the room. Testing occurred under red lights in the first 5 h after the onset of the dark cycle. We also observed D. spectabilis in their home cages for an additional 4 min post-test for signs of footdrumming.
Procedures for rat-snake interactions
We video-taped nine males and seven females of each species dur- ing encounters with live gopher snakes in a 4 x 1.2 x 1 m test arena with a wood floor that was covered with about 0.3 m of sifted sand. We filmed from a raised platform (2 x 2 x 0.63 m) 0.5 m from the test arena by focusing the video-camera either directly into the arena or from a mirror suspended at a 200 angle at the end of the arena opposite the camera. We video-taped from 25 October 1990 to 10 February 1991 under red lights with two cameras. A Sony EVC-X7 Video 8 camera with a low light intensity zoom lens was used to film the rats and a Cannon E53 8 mm video camera and recorder with a AF zoom lens to 7 lux filmed the snakes.
Filming began when a snake was released gently from a cloth bag on the opposite side of the arena from the test subject. An encounter was initiated when either the snake or the rat approached within 1 m and began to interact. The encounter was terminated 15 min after its initiation. Prior to each encounter, a rat spent 30-48 h in the experimental arena with its tin can "burrow" and a scattering of seeds. Approximately 2-3 h before beginning the encounter, we removed the tin can to facilitate interactions between rats and snakes and turned on the red lights. This usually occurred within I h after the lights went off in the normal light cycle in the room.
We analyzed each 1 5-min encounter in three 5-min segments for the following rat behavior: (1) approaches to within I m of the snake's head and body (marks at 1 m intervals on the sides of the arena facilitated measuring distances between animals), (2) fre- quency of withdrawal from within 1 m of the snake, (3) time spent within l m of the snake's head, (4) time spent within striking dis- tance of the snake mealsured as two body lengths (17 cm) of the
rat, (5) number of times a rat stopped activity, turned its head and oriented toward the snake, (6) frequency of jumping back from the snake, (7) frequency of kicking sand with hind feet, (8) frequency of sniffing tracks left by snakes' body in the sand, and (9) the fre- quency and distance from the snake of audible footdrumming. We analyzed all video tapes in real time with a Sony EV-Sl 8-mm VCR with slow motion, freeze frame and frame by frame advance and a Sony Triniton, color monitor (PMV 2530).
We also tabulated snake behavior from the video-tapes: (1) predatory approaches in which the snake moved with snout directed toward the prey flicking its tongue (Gillingham and Clark 1981), (2) withdraw by moving away from the rat, (3) frequency of strikes, and (4) frequency of hissing. We tested for changes in predatory behavior during the encounters by comparing the number of preda- tory approaches in the first and second 5-min periods during the first encounter a snake had with each species.
Data analysis
All data were analyzed using SYSTAT (Wilkinson 1990). We used a repeated-measures analysis of variance (ANOVA) to analyze data from the olfactory tests. In this model, sex was the between sub- jects variable and location (stimulus quadrant or far quadrant) and stimulus (solvent, oil, gopher snake, and rattlesnake odor) were the repeated measures variables. We analyzed data for each species sep- arately, because the large variation in the data of D. merriami greatly affected the outcome of any statistical test that included both species and was not representative of the behavior of D. spectabilis. All other comparisons, however, included species. We analyzed the behavioral responses to the gopher snakes in a 2 x 3 design, species x time, in which time was the repeated measure. When data contained zero responses and failed to meet criteria for parametric tests, we combined data for the 15-min tests and compared species using a nonparametric Mann-Whitney U-test. We report multi- variate F-statistics when possible because assumptions of homoge- neous variances and compound symmetry are not required (Wilkinson 1990). Data were normalized with a log transformation when necessary and reported as x ? SE.
Results
Olfactory Test
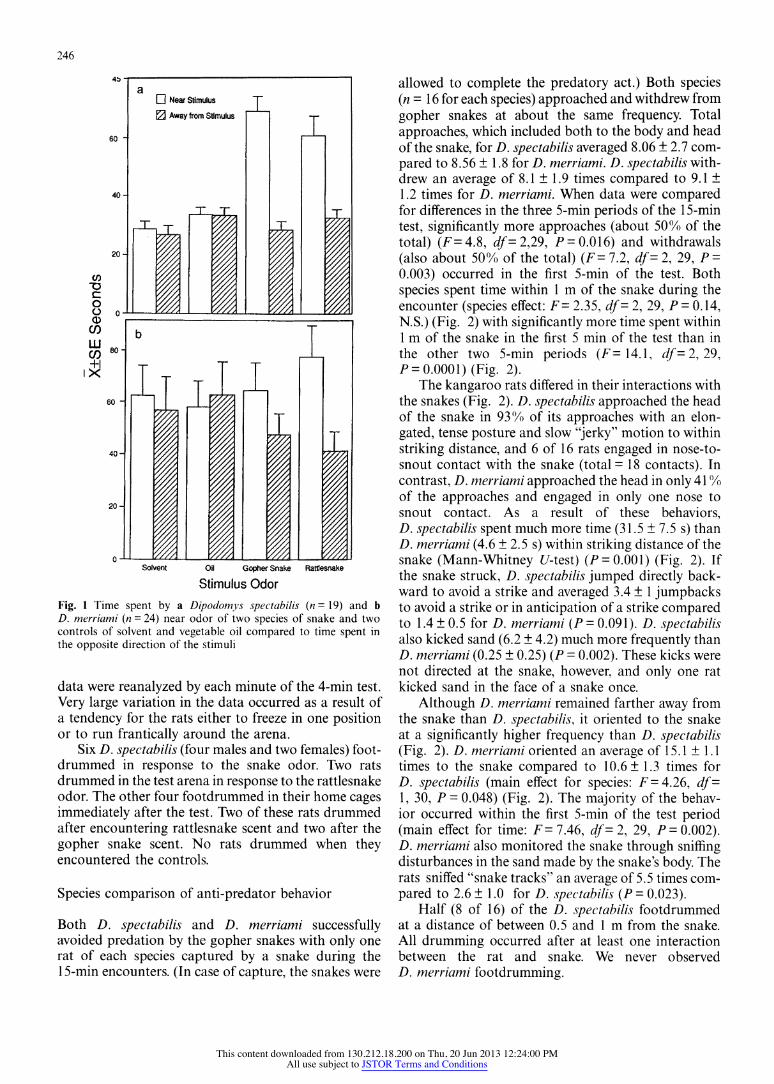
D. spectabilis clearly responded to snake odor. The rats actively approached the scent of both rattlesnakes and gopher snakes, sniffed the area and sometimes dug in the odor deposit. They spent significantly more time inspecting snake odor than the controls (main effect for stimulus: F= 15.5, df = 3, 15, P = 0.0001) as well as more total time in the stimulus quadrant than in the quad- rant opposite the stimulus (main effect for location: F= 90.6, df = 1, 17, P = 0.0001) (Fig. 1). This effect was a result of the time spent inspecting the snake odor. The rats spent almost twice as long in the quadrant with the snake odor as they spent away from the stimulus (inter- action of location and stimulus: F= 26.9, df = 3, 15, P = 0.0001). No difference was found in the rats' response to the odor of the two snake species and none of the data analyzed for sex differences was significant.
In contrast to D. spectabilis, the data for D. merri- ami yielded no significant differences in any of the analyses (Fig. 1). This result was also found when the
This content downloaded from 130.212.18.200 on Thu, 20 Jun 2013 12:24:00 PMAll use subject to JSTOR Terms and Conditions
246
4b -a a EL Near Stimulus T 3 Away from Stimulus
60-
40-
20-
a) co b
Cl)80 +1
60-
40-
20-
Solvent Oil Gopher Snake Ratfesnake
Stimulus Odor Fig. 1 Time spent by a Dipodomys spectabilis (n = 19) and b D. merriami (n = 24) near odor of two species of snake and two controls of solvent and vegetable oil compared to time spent in the opposite direction of the stimuli
data were reanalyzed by each minute of the 4-min test. Very large variation in the data occurred as a result of a tendency for the rats either to freeze in one position or to run frantically around the arena.
Six D. spectabilis (four males and two females) foot- drummed in response to the snake odor. Two rats drummed in the test arena in response to the rattlesnake odor. The other four footdrummed in their home cages immediately after the test. Two of these rats drummed after encountering rattlesnake scent and two after the gopher snake scent. No rats drummed when they encountered the controls.
Species comparison of anti-predator behavior
Both D. spectabilis and D. merriami successfully avoided predation by the gopher snakes with only one rat of each species captured by a snake during the 1 5-min encounters. (In case of capture, the snakes were
allowed to complete the predatory act.) Both species (n = 16 for each species) approached and withdrew from gopher snakes at about the same frequency. Total approaches, which included both to the body and head of the snake, for D. spectabilis averaged 8.06 ? 2.7 com- pared to 8.56 ? 1.8 for D. merriami. D. spectabilis with- drew an average of 8.1 ? 1.9 times compared to 9.1 + 1.2 times for D. merriami. When data were compared for differences in the three 5-min periods of the 1 5-min test, significantly more approaches (about 50% of the total) (F= 4.8, df =2,29, P =0.016) and withdrawals (also about 50% of the total) (F= 7.2, df = 2, 29, P = 0.003) occurred in the first 5-min of the test. Both species spent time within I m of the snake during the encounter (species effect: F= 2.35, df = 2, 29, P= 0.14, N.S.) (Fig. 2) with significantly more time spent within 1 m of the snake in the first 5 min of the test than in the other two 5-min periods (F= 14.1, df 2, 29, P = 0.0001) (Fig. 2).
The kangaroo rats differed in their interactions with the snakes (Fig. 2). D. spectabilis approached the head of the snake in 93%'/o of its approaches with an elon- gated, tense posture and slow "jerky" motion to within striking distance, and 6 of 16 rats engaged in nose-to- snout contact with the snake (total = 18 contacts). In contrast, D. merriami approached the head in only 41 % of the approaches and engaged in only one nose to snout contact. As a result of these behaviors, D. spectabilis spent much more time (31.5 ? 7.5 s) than D. merriami (4.6 ? 2.5 s) within striking distance of the snake (Mann-Whitney U-test) (P = 0.00 1) (Fig. 2). If the snake struck, D. spectabilis jumped directly back- ward to avoid a strike and averaged 3.4 ? 1 jumpbacks to avoid a strike or in anticipation of a strike compared to 1.4 ? 0.5 for D. merriami (P = 0.091). D. spectabilis also kicked sand (6.2 ? 4.2) much more frequently than D. merriami (0.25 ? 0.25) (P = 0.002). These kicks were not directed at the snake, however, and only one rat kicked sand in the face of a snake once.
Although D. merriami remained farther away from the snake than D. spectabilis, it oriented to the snake at a significantly higher frequency than D. spectabilis (Fig. 2). D. merriami oriented an average of 15.1 ? 1.1 times to the snake compared to 10.6 ? 1.3 times for D. spectabilis (main effect for species: F 4.26, df= 1, 30, P = 0.048) (Fig. 2). The majority of the behav- ior occurred within the first 5-min of the test period (main effect for time: F= 7.46, df = 2, 29, P = 0.002). D. merriami also monitored the snake through sniffing disturbances in the sand made by the snake's body. The rats sniffed "snake tracks" an average of 5.5 times com- pared to 2.6 ? 1.0 for D. spectabilis (P= 0.023).
Half (8 of 16) of the D. spectabilis footdrummed at a distance of between 0.5 and 1 m from the snake. All drumming occurred after at least one interaction between the rat and snake. We never observed D. merriamni footdrumming.
This content downloaded from 130.212.18.200 on Thu, 20 Jun 2013 12:24:00 PMAll use subject to JSTOR Terms and Conditions

247
a
0
6 6
05s
4-
0-
Cl 3
0r2 (D
0~ 4-
w
(1)3
(D Ca
C 1O 2
E 0
E
E
45
40 40
w O 535 -
-H Ix -...030- CD 0) c25 (Ui
~20-
C 10 (U E s 1=
0- 0 5 10 15
Minutes Fig. 2 Time spent by D. spectabilis * (n = 16) and D. merriami o (n = 16) a orienting to b, within 1 m, and c within striking distance of gopher snakes for 5-min intervals of a 15-min encounter
Snake behavior
The gopher snakes exhibited predatory behavior dur- ing the tests. They approached quickly and directly while tracking the rats' movement with their heads and
tongue. Once predator and prey were less than 0.5 m apart, the snake began to coil slowly while still moni- toring movement of the rat and, if either the rat or snake moved closer, the snake would strike, often from a partial coil. The snakes exhibited predatory approaches and withdrawals at the same frequency when paired with both species (Table 1).
Differences in the behavior of the rats was reflected in the behavior of the snakes. The snakes struck more often in encounters with D. spectabilis than with D. merriami (Table 1). At least one strike occurred in 10 of 16 encounters with D. spectabilis compared to one strike in only 2 of 16 encounters with D. merriami. Frequency of hissing also differed with more hisses directed at D. spectabilis than D. merriami (Table 1).
The gopher snakes significantly decreased preda- tory approaches directed toward D. spectabilis after the first 5 min of the encounter. The seven snakes approached an average of 3.75 ? 0.92 times in the first 5 min compared to only 0.90 ? 0.40 times in the sec- ond 5 min (Wilcoxon sign test: P = 0.031) (Fig. 3). This behavior was not found for D. merriami. The snakes continued to approach D. merriami at about the same frequency during the second 5-min period of the test (1.90 ? 0.27 approaches) as in the first 5-min period (2.41 ? 0.24 approaches) (P = 1.0, N.S).
Table 1. Number of strikes, hisses, predatory approaches and with- drawal of gopher snakes in response to encounters (n = 16 for each species) with two species of kangaroo rat (Dipodom_ys)
D. merriami D. spectabilis
Behavior Strike frequency 3 37 Hiss frequency 1 22 Predatory approach 94 85 Withdraw 19 28
5
4 o- I U)
CZ
0~
0
0 5 10 15
Minutes
Fig. 3 Comparison of predatory approaches of gopher snakes (n = 7) to D. spectabilis@* and D. merriami o for each 5-mmn inter- val of a 1 5-mmn encounter
This content downloaded from 130.212.18.200 on Thu, 20 Jun 2013 12:24:00 PMAll use subject to JSTOR Terms and Conditions

248
Discussion
Kangaroo rats can behaviorally influence the risk of being preyed upon by gopher snakes. The two species of kangaroo rat in this study, however, differed in how they responded to predatory behavior of snakes: D. merriami monitored the location of gopher snakes from a distance and avoided them while D. spectabilis actively approached to within striking distance, jumped back to avoid a strike and footdrummed at a safe dis- tance. These desert rodents, therefore, employ two different, but equally successful, strategies to assess and influence the risk of being preyed on by gopher snakes: confrontation and avoidance.
Before interacting with a snake, both D. spectabilis and D. mnerriami may use olfactory information to detect its presence. Rattlesnakes and gopher snakes depend on crypsis to ambush their prey (Sweet 1985; Radcliffe et al. 1986). Because snakes lay motionless in burrow tunnels or behind rocks and bushes on dark nights, scent may be the only effective means that desert rodents have to detect the presence of a snake and thus thwart any opportunity for an ambush. We found responses of D. spectabilis to snake odor consistent with their anti-predator behavior in the laboratory, and the active inspection of snake scent followed by foot- drumming also has been observed in the field (J.A. Randall, unpublished observations). The strong response to snake odor in the laboratory tests suggests that D. spectabilis responded specifically to the snake scents. However, because our study did not include a control for vertebrate odor, additional tests are neces- sary to show that D. spectabilis discriminates snake scent from other vertebrate scents.
The response of D. nierriami to snake odor in the test arena was less definitive than for D. spectabilis. The lack of a clear response to the snake extracts prob- ably resulted from the tendency of D. merriami either to freeze and remain inconspicuous or to attempt to escape from the small arena, rather than a disinterest in snake odors. This seems especially true because D. merriami inspected snake odors in encounters with live snakes in the larger arena. D. merriami may be more likely to inspect predator odors in a large, famil- iar space than in the small, somewhat unfamiliar arena used for olfactory tests in this study.
Attraction to snake odors seems a common rodent behavior. California ground squirrels, Spermophilus beechyii (Coss and Owings 1978; Hennessy and Owings 1978), Formosan squirrels, Callosiuris erythraeus thawvanensis (Tamura 1989) and Siberian chipmunks, Eutainiais sibiricus, (Kobayashi 1987) actively inspect snake scent. In contrast, the scent of mammalian preda- tors often repels small mammals and is avoided (Weldon 1990; Epple et al. 1993 and references therein). Therefore, the scent from predators can function either
to attract or to repel mammalian prey with responses specifically adapted to the mode of search and capture by the predator.
The anti-predator behavior observed in this study was directed toward nonvenomous gopher snakes, and the kangaroo rats did not discriminate between gopher snake and rattlesnake scent. We do not know whether kangaroo rats differentiate between live gopher snakes and rattlesnakes and thus alter their anti-predator behavior in response to the threat of a venomous snake. However, if the behavior of D. spectabilis is consistent with other rodents, the rats probably can discriminate the risk of venomous from nonvenomous snakes and adjust their anti-predator behavior accordingly. For instance, S. beecheyi and S. columbianus discriminate the difference between gopher and rattlesnake odors (Hennessy and Owings 1978; Towers and Coss 1990, 1991) and direct similar anti-predator behavior that differs in frequency and intensity toward both rattle- snakes and gopher snakes (Coss and Owings 1978; Towers and Coss 1990, 1991). The squirrels approach within striking distance of both venomous and non- venomous snakes, but snake-experienced squirrels tend to harass the more dangerous rattlesnakes more than gopher snakes (Towers and Coss 1990, 1991).
What benefits might D. spectabilis gain from engag- ing in such risky anti-predator behavior? First, con- frontation probably affords less risk to the larger D. spectabilis than to the smaller D. merriarni. Second, if confrontation and footdrumming by D. spectabilis cause a decline in the predatory behavior of snakes, as seen in the decrease in predatory approaches of the gopher snakes after the first few minutes of an encounter, the less risky option is to confront the snake and make it leave the area of the mound rather than to risk continued predation attempts. In natural obser- vations in the field, snakes left the vicinity of the mound in response to approaches and footdrumming by the mound owner (Randall and Stevens 1987). Because sur- vival of D. spectabilis depends on occupation of a mound and the rats are unable to relocate without a large cost, it is probably to their advantage to deter a dangerous predator from the territory as quickly as possible.
Although D. spectabilis interacts with snakes alone instead of in a group, its anti-predator behavior may function similar to mobbing behavior in social species of squirrel (Curio 1978). Ground squirrels, prairie dogs, Cynomys ludovicianus, and Siberian squirrels mob and aggressively harass snakes until the snakes move away (Owings and Coss 1977; Halpin 1983; Owings and Loughry 1985; Loughry 1987; Towers and Coss 1990). Prairie dogs and California ground squirrels increase the intensity of snake harassment if the snake does not move, and prairie dogs engage in jump-yipping and footdrumming (Hennessy and Owings 1978; Owings and Owings 1979; Owings and Loughry 1985; Loughry
This content downloaded from 130.212.18.200 on Thu, 20 Jun 2013 12:24:00 PMAll use subject to JSTOR Terms and Conditions

249
1987). Ground squirrels also kick dirt at both gopher snakes and rattlesnakes (Towers and Coss 1990, 1991). D. spectabilis, however, neither bites nor kicks sand at snakes, and snakes showed no signs of distress or con- fusion in encounters either in the laboratory or in the field (Randall and Stevens 1987). On the other hand, the snake may find the approach of a rat directly up to its nose aversive, and the hissing of some of the smaller snakes was defensive.
Confronting a predator may also function to com- municate the futility of further hunting (Curio et al. 1978). D. spectabilis confronts a snake first and then footdrums from a safe distance. Randall and Stevens ( 1987) and Randall (1993) proposed a "predator deter- rence" hypothesis in which the rats could footdrum as an individual alarm signal to inform the snake it has been detected and to leave (Woodland et al. 1980). Footdrumming could also convey health and ability to avoid predation (Dawkins and Krebs 1978). These and other hypotheses for the function of footdrumming at snakes are under investigation.
Avoidance of snakes by D. merriami also seems to be an effective anti-predator strategy. By staying a safe distance away, D. merriami avoids the risk of being within striking distance of the snake. Hungry snakes, however, may engage in predatory behavior longer in the absence of direct confrontation, which requires more vigilance by the rat. In encounters with D. mer- riamni, the snakes continued to approach for a longer time than they did in the encounters with D. spectabilis. In contrast to D. spectabilis, D. merriami can move away from a snake into a different burrow within its home range, and a snake is probably little threat if the rat can monitor its location. D. merriami has been observed foraging near large, coiled rattlesnakes with- out adverse consequences (J.A. Randall, unpublished observations), and the rats successfully avoided preda- tion by rattlesnakes in experimental enclosures if they had been cued to a snake's presence (Pierce etal. 1992). The better detection of snakes and ability to jump back- ward to escape a strike in open habitats may be at least one reason why kangaroo rats often avoid foraging in brushy areas (Randall 1993).
Acknowledgements We are grateful to Allie Rich for her assistance with the olfactory tests. This research was supported by a grant from the National Science Foundation (BNS 89-08827) to J.A. Randall. The experimental procedures in this study were approved by the University Animal Care and Use Committee.
References
Bartholomew GA, Caswell HH (1951) Locomotion in kangaroo rats and its adaptive significance. J Mammal 32:155-169
Best TL (1972) Mound development by a pioneer population of the banner-tailed kangaroo rat, Dipoclomys spectabilis Goldman, in eastern New Mexico. Am Midl Nat 87:201-206
Brown JS (1988) Patch use as an indicator of habitat preference, predation risk, and competition. Behav Ecol Sociobiol 22: 37-47
Coss RG, Owings DH (1978) Snake-directed behavior by snake naive and experienced California ground squirrels in a simulated burrow. Z Tierpsychol 48:421-435
Curio E (1978) The adaptive significance of mobbing. I. Teleonomic hypotheses and prediction. Z Tierpsychol 48:175-183
Curio E, Ernst U, Vieth W (1978) Cultural transmission of enemy recognition: one function of mobbing. Science 202:899-901
Daly M, Wilson M, Behrends PR, Jacobs LF (1990) Characteristics of kangaroo rats, Dipodon1vs merriami, associated with differen- tial predation risk. Anim Behav 40:380-389
Daly M, Behrends PR, Wilson MI, Jacobs LF (1992) Behavioural modulation of predation risk: moonlight avoidance and cre- puscular compensation in a nocturnal desert rodent, Dipodorns merricani. Anim Behav 44:1-9
Dawkins R, Krebs JR (1978) Animal signals: information or manipulation. In: Krebs JR, Davies NB (eds) Behavioural ecol- ogy. Blackwell, Oxford pp 282-308
Djawdan M (1993) Locomotor performance of bipedal and quadrupedal heteromyid rodents. Funct Ecol 7:195-202
Djawdan M, Garland T Jr (1988) Maximal running speeds of bipedal and quadrupedal rodents. J Mammal 69:765-772
Epple G, Mason JR, Nolte DL, Campbell DL (1993) Effects of predator odors on feeding in the mountain beaver (Aplodoontia rufa). J Mammal 74:715-722
Gillingham JC, Clark DG (1981) An analysis of prey searching behavior in the western diamondback rattlesnake, Crotalus aitrox. Behav Neur Biol 32:235-240
Goldthwaite RO, Coss RG, Owings DH (1990) Evolutionary dis- sipation of an antisnake system: differential behavior by California and arctic ground squirrels in above- and below- ground contexts. Behaviour 112:246-269
Graves BM, Halpern M (1988) Neonate plains garter snakes (Thaminnophis radlix) are attracted to conspecific skin extracts. J Comp Psychol 102:251-253
Halpin ZT (1983) Naturally occurring encounters between black- tailed prairie dogs (Cvnomvs ludovicianus) and snakes. Am Mid] Nat 109:50-54
Hennessy DF, Owings DH (1978) Snake species discrimination and the role of olfactory cues in the snake-directed behavior of the California ground squirrel. Behaviour 67:115-124
Jones WT (1984) Natal philopatry in bannertailed kangaroo rats. Behav Ecol Sociobiol 15:151-155
Kobayashi T (1987) Does the Siberian chipmunk respond to the snake by identifying it? J Ethol 5:137-144
Kotler BP (1984) Risk of predation and the structure of desert rodent communities. Ecology 65:689-701
Kotler BP (1992) Behavioral resource depression and decaying per- ceived risk of predation in two species of coexisting gerbils. Behav Ecol Sociobiol 30:239-244
Kotler BP, Brown JS, Smith RJ, Wirtz WO 11( 1988) The effects of morphology and body size on rates of owl predation on desert rodents. Oikos 53:145-152
Kotler BP, Brown JS, Slotow RH, Goodfriend WL, Strauss M (1993) The influence of snakes on the foraging behavior of ger- bils. Oikos 67:309-316
Lima SL, Dill LM (1990) Behavioral decisions made under the risk of predation: a review and prospectus. Can J Zool 68:619-640
Longland WS, Price MV (1991) Direct observations of owls and heteromyid rodents: can predation risk explain microhabitat use? Ecology 72:2261-2273
Loughry WJ (1987) The dynamics of snake harassment by black- tailed prairie dogs. Behaviour 103:27-48
Owings DH, Coss RG (1977) Snake mobbing by California ground squirrels: adaptive variation and ontogeny. Behaviour 62:50-69
Owings DH, Owings SC (1979) Snake-directed behavior by black- tailed prairie dogs (Cvxnoniov s ludoviciansus). Z Tierpsychol 49: 35-54
This content downloaded from 130.212.18.200 on Thu, 20 Jun 2013 12:24:00 PMAll use subject to JSTOR Terms and Conditions

250
Owings DH, Loughry WJ (1985) Variation in snake-elicited jump- yipping by black-tailed prairie dogs: ontogeny and snake specificity. Z Tierpsychol 70:177-200
Pierce BM, Longland WS, Jenkins SH (1992) Rattlesnake preda- tion on desert rodents: microhabitat and species-specific effects on risk. J Mammal 73:859-865
Pulliam HR, Caraco T (1984) Living in groups: Is there an optimal group size? In: Krebs JR, Davies NB (eds) Behavioural ecology: an evolutionary approach, Sinauer, Sunderland, pp 122-147
Radcliffe GW, Esteop K, Boyer T, Chizar D (1986) Stimulus con- trol of predatory behavior in red spitting cobras (Naja mossam- bica pallicla) and prairie rattlesnakes (Crotalus v. viridis). Anim Behav 34:804-814
Randall JA (1984) Territorial defense and advertisement by foot- drumming in bannertail kangaroo rats (Dipodomys spectabilis) at high and low population densities. Behav Ecol Sociobiol 16: 11-20
Randall JA (1989a) Individual footdrumming signatures in banner- tailed kangaroo rats, Dipodomys spectabilis. Anim Behav 38: 620-630
Randall JA (1989b) Neighbor recognition in a solitary mammal (Dipodomys merriami). Ethology 81:123-133.
Randall JA (1993) Behavioural adaptations of desert rodents (Heteromyidae). Anim Behav 45:263-287
Randall JA (1994) Discrimination of footdrumming signatures by kangaroo rats (Dipodomys spectabilis). Anim Behav 47:45-54
Randall JA, Stevens CM (1987) Footdrumming and other anti- predator responses in the bannertail kangaroo rat (Dipodomys spectabilis). Behav Ecol Sociobiol 20:187-194
Schroder GD (1979) Foraging behavior and home range utilization of the bannertail kangaroo rat (Dipodomys spectabilis). Ecology 60:657-665
Sweet SS (1985) Geographic variation, convergent crypsis and mimicry in gopher snakes (Pituphis melanoleucus) and Wester rattlesnakes (Crotalus viridis). J Herpetol 19:55-67
Tamura N (1989) Snake-directed mobbing by Formosan squirrel Callosciurus ervthraeus thaiwanensis. Behav Ecol Sociobiol 24: 175-180
Towers, SR, Coss RG (1990) Confronting snakes in the burrow: snake-species discrimination and antisnake tactics of two California ground squirrel populations. Ethology 84:177-192
Towers SR, Coss RG (1991) Antisnake behavior of Columbian ground squirrels (Spermophilus columbianus). J Mammal 72: 776-783
Webster DB (1962) A function of the enlarged middle-cavity of the kangaroo rat Dipodomys. Physiol Zool 35:248-255
Webster DB, Webster M (1971) Adaptive value of hearing and vision in kangaroo rat predator avoidance. Brain Behav Evol 4:310-322
Webster DB, Webster M (1984) The specialized auditory system of kangaroo rats. Sensory Physiol 8:161-196
Weldon PJ (1990) Responses by vertebrates to chemicals from predators. In: Macdonald DW, Muller-Schwarze D, Natynczuk SE (eds) Chemical signals in vertebrates 5. Oxford University Press, Oxford, pp 500-521
Woodland DJ, Jaafar Z, Knight ML (1980) The "pursuit deter- rent" function of alarm signals. Am Nat 115:748-753
Wilkinson L (1990) SYSTAT: the system for statistics. Systat, Evanston
Communicated by G.M. Klump
This content downloaded from 130.212.18.200 on Thu, 20 Jun 2013 12:24:00 PMAll use subject to JSTOR Terms and Conditions