spread of a distinct stx2- encoding phage prototype among...
TRANSCRIPT

0
JVI00986-12 revised 1
2
Spread of a distinct Stx2-encoding phage prototype among E. coli O104:H4 3
strains from outbreaks in Germany, Norway and Georgia 4
5
Lothar Beutin 1*, Jens Andre Hammerl 1*, Eckhard Strauch 1, Jochen Reetz 1, Ralf 6
Dieckmann 1, Ylanna Kelner-Burgos 1, Annett Martin 2, Angelika Miko 1, Nancy A. 7
Strockbine 3, Björn Arne Lindstedt 4, Detlef Horn 5, Hella Monse 5, Bruno Huettel 6, Ines 8
Müller 6, Kurt Stüber 6, and Richard Reinhardt 6 9
10
1 Federal Institute for Risk Assessment, Department of Biological Safety, Diedersdorfer 11
Weg 1, D-12277 Berlin, Germany 12
2 Federal Institute for Risk Assessment, Epidemiology, Biostatistics and Mathematical 13
Modelling, Scientific Services, Diedersdorfer Weg 1, D-12277 Berlin, Germany 14
3 Centers for Disease Control and Prevention, Escherichia and Shigella Reference Unit, 15
Atlanta, United States 16
4 Norwegian Institute of Public Health, Dept. of Foodborne Infections, Oslo, Norway 17
5 Chemisches und Veterinäruntersuchungsamt Rhein-Ruhr-Wupper, CVUA-RRW, Krefeld, 18
Germany 19
6 Max-Planck-Genomzentrum, Carl-von-Linné-Weg 10, D-50829 Köln, Germany 20
21
Running title: Stx2 phage of enteroaggregative E. coli O104:H4 22
23
Keywords: Shiga toxin, bacteriophage, STEC, EHEC, O104:H4, outbreak24
Copyright © 2012, American Society for Microbiology. All Rights Reserved.J. Virol. doi:10.1128/JVI.00986-12 JVI Accepts, published online ahead of print on 18 July 2012
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

1
* Corresponding Authors: 25
Dr. Lothar Beutin, National Reference Laboratory for Escherichia coli, Federal Institute 26
for Risk Assessment (BfR), Diedersdorfer Weg 1, D-12277 Berlin, Germany 27
E-mail: [email protected] 28
Phone: +49 30 18412 2259 29
Fax: +49 30 18412 2983 30
31
Dr. Jens Andre Hammerl, Department of Biological Safety, Federal Institute for Risk 32
Assessment (BfR), Diedersdorfer Weg 1, D-12277 Berlin, Germany 33
E-mail: [email protected] 34
Phone: +49 30 18412 2057 35
Fax: +49 30 18412 2000 36
37
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

2
ABSTRACT 38
Shiga toxin 2 (Stx2)-producing Escherichia coli (STEC) O104:H4 caused one of the 39
worlds largest outbreaks of hemorrhagic colitis and hemolytic uremic syndrome in 40
Germany in 2011. These strains have evolved from enteroaggregative E. coli (EAEC) by 41
the acquisition of the Stx2 genes and have been designated as enteroaggregative 42
hemorrhagic E. coli. Nucleotide sequencing has shown that the Stx2 gene is carried by 43
prophages integrated into the chromosome of STEC O104:H4. We studied the properties 44
of Stx2-encoding bacteriophages which are responsible for the emergence of this new type 45
of E. coli pathogen. For this, we analyzed Stx bacteriophages from STEC O104:H4 strains 46
from Germany (in 2001 and 2011), Norway (2006) and the Republic of Georgia (2009). 47
Viable Stx2-encoding bacteriophages could be isolated from all STEC strains except for 48
the Norwegian strain. The Stx2 phages formed lysogens on E. coli K-12 by integration 49
into the wrbA locus resulting in Stx2 production. The nucleotide sequence of the Stx2 50
phage P13374 of a German STEC O104:H4 outbreak was determined. From the 51
bioinformatic analyses of the prophage sequence of 60,894 base pairs 79 open reading 52
frames were inferred. Interestingly, the Stx2 phages from the German 2001 and 2011 53
outbreak strains were found to be identical and closely related to the Stx2 phages from the 54
Georgian 2009 isolates. Major proteins of the virion particles were analysed by mass 55
spectrometry. Stx2 production in STEC O104:H4 strains was inducible by mitomycin C 56
and was compared to Stx2 production of E. coli K-12 lysogens. 57
58
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

3
INTRODUCTION 59
60
In Germany between May and July 2011, Shiga toxin-producing E. coli (STEC) 61
O104:H4 were identified as the cause of one of the world's largest outbreaks of STEC (19). 62
The outbreak spread to other European and non-European countries and finally accounted for 63
3,842 cases of human infections with a high percentage of patients developing the life-64
threatening hemolytic uremic syndrome (HUS) (17). Until 2011, O104:H4 was considered a 65
rare STEC serotype (8,47), with only a few sporadic cases of human STEC O104:H4 66
infection reported in some European and Asian countries (51). The first STEC O104:H4 67
strains were isolated in 2001 from the feces of siblings with hemorrhagic colitis (HC) and 68
HUS in Cologne, Germany (7,17). 69
The causative STEC O104:H4 strain recovered in the outbreak 2011 differs from 70
E. coli O157:H7 and other enterohemorrhagic E. coli (EHEC) strains because it is negative 71
for the EHEC virulence plasmid and for the genes encoding the attaching/effacing (A/E) 72
mechanism (7,51). On the genome level, STEC O104:H4 strains were shown to most similar 73
to the previously characterized enteroaggregative E. coli (EAEC) strain 55989 and were found 74
to carry typical virulence genes of EAEC strains. (7,46). Accordingly, the term 75
enteroaggregative hemorrhagic E. coli (EAHEC) was proposed for this new group of human 76
pathogenic E. coli (10). Interestingly, EAEC specific virulence genes were not found in all 77
STEC O104:H4 isolates collected before 2011 and significant differences in the EAEC 78
specific virulence genes and EAEC plasmids were found by comparing the German STEC 79
O104:H4 strains from 2001 and from 2011 (7,26). 80
Similar to many classical EHEC strains, the STEC O104:H4 outbreak isolate produces 81
the Shiga toxin 2a (Stx2a). This toxin type is associated with classical EHEC (16,38,46), in 82
particular with EHEC O157:H7 strains. The stx2a genes are located on the genome of 83
lambdoid bacteriophages integrated in the chromosome of their bacterial hosts (31,38,39). 84
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

4
Augmented colonization of the humans by STEC O104:H4 through aggregative adherence 85
fimbriae might result in high quantities of Stx2a delivered into the host intestine. This was 86
suggested as the causative mechanism leading to the high numbers of patients with severe 87
clinical illness in STEC O104:H4 infected humans (7). 88
Stx2a was previously shown to be associated with severe clinical illness in humans 89
(20,21) and the amount of Stx2a produced by EHEC O157:H7 strains was found to correlate 90
with the development of severe clinical illness in a gnotobiotic piglet model (3). The 91
production of Stx by STEC can be significantly enhanced by certain antimicrobial and by 92
DNA-damaging agents such as ciprofloxacin and mitomycin C, respectively (6,13,48). An 93
increase in Stx production caused by exposure to agents such as ciprofloxacin likely results 94
from induction of the prophage. Stx production was also found to be stimulated by 95
neutrophils and norepinephrine in the gastrointestinal tract indicating that the inducibility of 96
Stx might play a role for in vivo pathogenesis (14,54). The genetic composition of Stx-97
encoding bacteriophages could have an influence on bacterial host specificity and on Stx 98
production (13,48,55). A number of Stx2-encoding bacteriophages have been analysed for the 99
nucleotide sequence and significant differences in global nucleotide composition and in the 100
regulatory genes affecting Stx production were found (31,39,45,49). Genomic sequencing of 101
STEC O104:H4 strains revealed that these carry Stx2a-encoding prophages (40,43). 102
In this work we characterized the Stx2a-encoding short-tailed DNA bacteriophage 103
(podovirus) P13374 from the STEC O104:H4 strain CB13374 which was isolated in the 104
course of outbreak investigations from an opened package of retailed sprouted seeds from a 105
household of an infected patient (53). We analyzed P13374 for its genetic composition, 106
biological properties and bacterial host specificity. Genomic sequencing and analysis of 107
restriction endonuclease digestion patterns indicate that the bacteriophage P13374 is highly 108
similar to the Stx2a phages carried by STEC O104:H4 strains isolated from human patients in 109
Germany in 2001 and in 2011. Additionally, a comparison to phages which are present in 110
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

5
Stx2a producing STEC O104:H4 strains from Norway and Georgia revealed close 111
similarities. 112
113
MATERIALS AND METHODS 114
115
Bacteria and culture conditions. Bacterial strains used in this work were from the 116
collection of the National Reference Laboratory for Escherichia coli (NRL-E. coli, BfR, 117
Berlin, Germany). STEC O104:H4 strains were serotyped for the O and H antigens and 118
confirmed by Real Time PCR for the presence of stx2, aggR, terB, wzxO104 genes as previously 119
described (53). In order to discriminate between different virulotypes of STEC O104:H4, the 120
strains were additionally investigated for the EAEC heat stable enterotoxin (astA) and for 121
sequence encoding aggregative adherence fimbriae I (aggA) and III (agg3A) as described (7). 122
The strains used for induction and isolation of Stx2-encoding bacteriophages are listed in 123
Table 1. The STEC O104:H4 strain RKI 11-022027 (CB13344) was isolated from a female 124
patient with HUS at the onset of the STEC O104:H4 outbreak in May 2011 in northern 125
Germany (19). Strain CB13374 was isolated from an opened package of sprouted seeds from 126
the household of a patient infected with STEC O104:H4. The sprouts originated from the 127
farm incriminated as the food source of the STEC O104:H4 2011 outbreak strain (4,53). 128
STEC strain CB8983 was isolated in 2001 from a five years old girl with hemorrhagic colitis 129
in Cologne, Germany. Subsequently, the girl developed HUS and was hospitalized. The 130
clinical case report reveals that the strain HUSEC 41 (STEC O104:H4) was isolated from the 131
same patient after she was hospitalized with HUS (7) (L. Beutin, unpublished data). STEC 132
O104:H4 strain 1106-3060-1 (CB13437) originated from a patient in Norway who became 133
infected in 2006. Two STEC O104:H4 strains, 2009EL-2050 (CB13771) and 2009EL-2071 134
(CB13772) were isolated in 2009 from patients with bloody diarrhea in the Republic of 135
Georgia (12). A collection of 31 enteroaggregative E. coli (EAEC) strains was used for 136
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

6
susceptibility tests to the STEC O104:H4 Stx2a phage (see Table S1 in the supplemental 137
material) and characterized for serotypes and for the presence of the aggR gene as indicator 138
for the presence of the aggregative adherence mechanism (24). Escherichia coli K-12 strain 139
C600 was used as recipient for propagation and lysogenization of Stx bacteriophages (50). 140
C600 derivative strains lysogenizated with different Stx phages and phage lambda were 141
described previously and are listed in Table 3. Media for cultivation of bacteria, preparation 142
of phage lysates and phage susceptibility were used as previously described (50). 143
Isolation, propagation, and purification of Stx2 phages from STEC O104:H4 144
strains. Single colonies of the respective E. coli strains were grown in Luria-Broth (LB) to 145
exponential growth. Induction of lysogenic bacteriophages was performed by adding 146
0.5 µg/ml mitomycin C to the growing bacterial cultures which were further incubated 147
overnight. Bacteriophages were isolated from single plaques grown on the E. coli K-12 strain 148
C600 as described previously (27). High titer phage lysates (109-1011 pfu/ml) were obtained 149
from E. coli C600 as described (50) and were used for isolation of phage DNA, 150
characterization of phage envelope proteins and for phage infection studies. 151
Lysogenization of the E. coli K-12 strain C600 with Stx2a-encoding bacteriophages 152
from STEC O104:H4 strains. Serial dilutions of phages released in mitomycin C induced 153
cultures of O104:H4 strains were spotted on a lawn of the stx-negative E. coli K-12 strain 154
C600. Isolation of lysogenic C600 (Stx phage) strains was performed as previously described 155
(50). The presence of the Stx phage in C600 was confirmed by stx2-specific PCR as described 156
(53) and by mitomycin C induction of viable Stx2 phages from C600 lysogens. 157
Susceptibility of enteroaggregative E. coli (EAEC) to the Stx2 phage from 158
CB13374. Cultures of 31 EAEC strains belonging to 22 different serotypes from the 159
collection of the NRL-E. coli (see Table S1 in the supplemental material) were grown 160
overnight at 37°C in LB-broth. 100 µl of the culture was added to 3 ml of molten LC-Top 161
agar and immediately poured onto LB-agar as described (50). 20 µl portions of P13374 162
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

7
lysates (1011 pfu/ml) and of freshly mitomycin C induced cultures of the STEC O104:H4 163
strain CB13374 were spotted on EAEC strains and the plates were incubated for 20-22 h at 164
37°C. Lysis of bacteria and plaque formation was monitored after overnight incubation. 165
Strains C600 and CB13374 were used as positive and negative control for plaque formation, 166
respectively. In order to investigate the possible transfer of the stx2 gene to EAEC strains by 167
transduction bacteria grown in zones spotted with P13374 were subcultured on LB-agar and 168
grown overnight at 37°C. DNA extracted from subcultured bacteria was investigated for the 169
presence of the stx2 gene by Real Time PCR as described (53). 170
Determination of Stx2 production. Stx production by bacteria was measured 171
semiquantitavely by the Ridascreen Verotoxin enzyme immunoassay (Ridascreen-EIA, R-172
biopharm Darmstadt, Germany). Bacterial samples for the Stx-EIA were grown at 37°C in 173
Tryptic Soy Broth (TSB) supplemented with or without 0.5 µg/ml mitomycin C under 174
aeration as described previously (5). Undiluted and serial dilutions of bacterial culture fluid 175
were tested from overnight cultures of bacteria showing titers of about 109 cfu/ml. The 176
Ridascreen EIA test results were recorded photometrically with a Dynatech MRX Microplate 177
reader at 450 nm with 630 nm reference wavelength. 178
Statistical analysis. Independent t-tests were used to test for a difference (mean value 179
OD450) between two independent groups (STEC wildtype strains grown in the presence or 180
absence of mitomycin C as well as C600 Stx phage lysogens grown in the presence or 181
absence of mitomycin C, respectively). All analyses were performed with PASW Statistics 182
(v18.0.2). The statistical analyses were 2-tailed, and p values <0.05 were considered 183
significant. 184
Morphology of isolated bacteriophages. To determine the morphological 185
characteristics of isolated phages by transmission electron microscopy (TEM), the negative-186
staining procedure was used. Briefly, Pioloform-carbon-coated, 400-mesh copper grids (Plano 187
GmbH, Wetzlar, Germany) were placed in drops from supernatants of samples collected and 188
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

8
primed for TEM for 10 min, fixed with a 2.5% aqueous glutaraldehyde (Taap Laboratories, 189
Aldermaston, U.K.) solution for 1 min and stained with 2% aqueous uranyl acetate (Merck, 190
Darmstadt, Germany) solution for 1 min. 191
In addition, the ultrathin sectioning method was performed for electron microscopic 192
investigations of the E. coli K-12 strain TPE2364 (C600) infected with phages lysates of the 193
sprouts strain CB13374. For these investigations infected and mitomycin C-induced bacterial 194
cell suspensions were concentrated by centrifugation. The culture supernatant was removed 195
and the bacterial pellet was suspended in a 2.5% aqueous glutaraldehyde (Taap Laboratories) 196
solution for 24 h and centrifuged at 8,000 rpm for 5 min. The supernatant was removed and 197
the pellet was mixed with a 3% aqueous low melting point agarose (MP Biomedicals, 198
Eschwege, Germany) solution, centrifuged shortly at 14,000 rpm for 5 s and immediately 199
refrigerated for 10 min. This agarose/bacteria cell pellet was cut into little cubes and soaked in 200
1% aqueous osmium tetroxide (Electron Microscopic Science, Hatfield, U.K.) solution before 201
embedding in Epon Resin (Agar Scientific, Stansted, U.K.) according to standard procedures. 202
Thereafter, ultrathin sections were cut, post-contrasted with 2% aqueous uranyl acetate 203
(Merck) solution for 20 min and 2.7% alkaline lead citrate (Serva, Heidelberg, Germany) 204
work solution for 15 min. The negative stained specimens and the ultrathin sections were 205
examined with a JEM-1010 electron microscope (Jeol, Tokyo, Japan) at 80 kV acceleration 206
voltages. Photographs and measurements were performed using a digital camera MegaView II 207
and a computer assisted analysis program (SIS Analysis, Olympus, Münster, Germany). 208
SDS-PAGE and in-gel digestion of P13374 proteins. Phage P13374 proteins were 209
extracted from concentrated phage particles, reduced with 10 mM DTT for 5 min at 95°C and 210
SH groups were subsequently alkylated with 50 mM iodoacetamide for 45 min in the dark at 211
room temperature. Samples were heated for 5 min in sample buffer at 95°C and subjected to 212
sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) according to the 213
procedure of Laemmli (29). Protein samples were electrophoresed on precast gradient gels 214
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

9
(Criterion Tris-HCl 4-20%, Biorad, Munich, Germany) in a Bio-Rad Protean II 215
electrophoresis system. Gels were washed with water and stained using BioSafe Coomassie 216
Stain (BioRad). Bands of interest were excised and cut into smaller pieces using a clean 217
scalpel. Gel pieces were washed one after another with water, 50% acetonitrile (ACN) and 218
20 mM NH4HCO3. Gel pieces were washed twice with 20 mM NH4HCO3/ ACN (50:50 v/v) 219
to destain the proteins, dehydrated with 100% ACN and dried. Tryptic in-gel digestion was 220
carried out overnight at 37°C using a 12.5 ng/ml solution of proteomics-grade recombinant 221
trypsin (Roche, Mannheim, Germany) in 25 mM NH4HCO3/ 10% ACN/ 5 mM CaCl2. Tryptic 222
peptides were extracted with 50% ACN in 5% trifluoroacetic acid (TFA) for 30 min. The 223
extracted peptides were dried, reconstituted with 0.3% TFA in 60% ACN and directly 224
subjected to MALDI TOF-TOF MS/MS analysis. Alternatively, dried peptides were 225
reconstituted in 0.1% TFA and purified using C18 Zip-Tip according to the manufacturers 226
instructions (Millipore, Schwalbach, Germany) and subjected to MALDI TOF-TOF MS/MS 227
analysis. 228
MALDI sample preparation and MALDI-TOF TOF MS-MS analysis. 1 µl of 229
extracted P13374 peptides was spotted on a MALDI stainless steel target plate and mixed 230
with 1 µl of matrix solution (5 mg/ml α-cyano-4-hydroxycinnamic acid in 0.3% TFA/ 60% 231
ACN). Zip-Tip purified samples were eluted with 5 µl 50% ACN in 0.1% TFA and 1 µl was 232
spotted onto a 800 µm AnchorChip Target plate and dried. Subsequently, 1 µl of matrix 233
solution was added (0.7 mg/ml α-cyano-4-hydroxycinnamic acid in 0.1% TFA/ 85% ACN/ 234
1 mM NH4H2PO4). For external calibration Peptide Calibration Standard II was used (Bruker 235
Daltonics, Bruker). MALDI-TOF TOF MS-MS analysis was performed on a Ultraflex II 236
TOF/TOF instrument (Bruker Daltonics) operated in the reflector mode for MALDI-TOF 237
peptide mass fingerprint (PMF) or LIFT mode for MALDI TOF-TOF analysis. PMF and 238
LIFT spectra were interpreted with the Mascot software (Matrix Science Ltd, London, UK). 239
Database searches (NCBInr), through Mascot were performed via BioTools 3.0 software 240
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

10
(Bruker Daltonics) using combined PMF and MS/MS (peptide fragment fingerprinting) 241
datasets. The peptide sequences obtained were localised in the annotated P13374 genome 242
sequence using WU-blast 2.0 (Gish, W., http://www.proweb.org/). 243
Isolation, sequencing, and bioinformatic analysis of the CB13374 Stx prophage. 244
The Stx prophage sequence of P13374 was deduced from one contig of the whole genome 245
sequencing (WGS) project (unpublished data) of E. coli O104:H4 strain CB13374. Genomic 246
DNA of the bacteria was extracted and purified by the use of the RTP Bacteria DNA Mini Kit 247
(Stratec, Berlin, Germany). Determination of the CB13374 genomic sequence was performed 248
with the Roche 454 genome sequencer FLX system. Libraries for 454 sequencing were 249
prepared and sequenced according to Roche’s 454 protocols and as previously described (44). 250
Assembling of the reads resulted in several contigs with an average sequence coverage of 251
more than 30-fold per consensus base. The genome sequence of the prophage was confirmed 252
by partial nucleotide sequencing of DNA from isolated phage particles. From these data the 253
final P13374 genome sequence of 60,882 bp was identified and used for further analyses. 254
Protein-encoding open reading frames (ORFs) were predicted using the algorithms of 255
Accelrys Gene version 2.5 (Accelrys Inc.) and ORF-Finder (http://www.ncbi.nlm.nih.gov/) 256
with manual curation. For the ORF analysis, the presence of ATG, GTG, or TTG as a 257
potential start codon and a length of at least 25 encoded amino acids were stated. To compare 258
the gene products of the P13374 ORFs to the annotated proteins of the non redundant 259
GenBank database BLASTp-searches (http://www.ncbi.nlm.nih.gov/blast/) were performed at 260
the NCBI homepage. For gene products with low e-values (>0.001) or no sequence similarity, 261
the PSI-BLAST algorithm was used (1). Identification of putative tRNA genes was performed 262
using tRNAscan-SE (30). Putative Rho-independent transcription terminators were identified 263
using TRANSTERM (9,15). 264
Analysis of integration sites of Stx2 phages. STEC O104:H4 strains and C600 Stx2 265
phage lysogens were investigated for possible prophage insertion site with primers developed 266
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

11
previously (11) (Table 2). PCR amplification products obtained with STEC O104:H4 strains 267
and E. coli K-12 C600 phage lysogens were investigated by nucleotide sequencing. 268
Database submission: The complete nucleotide sequence of phage P13374 was submitted to 269
EMBL under accession number HE664024. The sequences of the left and right integration 270
sites of the investigated Stx2a phages in wildtype EAEHEC and in E. coli K-12 strains were 271
submitted (left site: accessions HE775132-41; right site: accessions HE775266-75). 272
Additionally, the phage attachment sites in the viral genomes were deposited with the 273
accession numbers HE775127-31 (see Table S2 in the supplemental material). 274
275
RESULTS 276
277
Isolation of Stx2-encoding bacteriophages from STEC O104:H4 strains. Six STEC 278
O104:H4 strains from Germany (isolated in 2001 and 2011), Norway (2006) and the Republic 279
of Georgia (2009) were used as source for isolation of temperate Stx2-encoding 280
bacteriophages (Table 1). All strains were found to be positive by PCR for stx2, aggR and 281
wzxO104 genes but were different in regard to the presence of astA, aggA and agg3A genes 282
(Table 1). 283
Induction of temperate bacteriophages using mitomycin C was successful for all 284
O104:H4 strains. Stx2a-encoding bacteriophages were isolated from the German (CB8983, 285
CB13344 and CB13374) and the Georgian (CB13771 and CB13772) STEC O104:H4 strains. 286
The Norwegian strain CB13437 released only bacteriophages which did not encode stx2 287
genes (see below). All phages formed pinhead-sized plaques on the lawn of strain C600. The 288
presence of the stx2a gene in the purified bacteriophage DNA was confirmed by PCR as 289
described (53). C600 lysogenic for Stx2 phages were isolated as described in Material and 290
Methods section. The bacteriophage released from CB13437 did not form lysogens on C600. 291
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

12
To study the relationship of the phages purified phage DNAs were subjected to 292
restriction enzyme digestion with PvuI, EcoNI and BsrBI. The Stx2a phages isolated from 293
strains CB8983 (Germany 2001) as well as CB13344 and CB13374 (both Germany 2011) 294
were identical for their restriction profiles with all enzymes tested (Fig. 1). Similar findings 295
were obtained with Stx2a phages from other STEC O104:H4 collected from human patients 296
in the 2011 outbreak in Germany (data not shown). The restriction profiles of the Stx2a 297
phages isolated from the Georgian strains CB13771 and CB13772 were highly similar to each 298
other but showed differences in their restriction patterns with all three enzymes tested when 299
compared to the Stx2a phages isolated from the German 2001 and 2011 isolates (Fig. 1). The 300
Norwegian O104:H4 strain CB13437 did not release Stx phages although the strain was 301
positive for Stx2a phage associated sequences at the attachment site in the wrbA locus. 302
Lysogenization of E. coli with Stx2 phages isolated from STEC O104:H4. Stx2a-303
encoding bacteriophages derived from the German and Georgian O104:H4 strains were found 304
to lysogenize C600 resulting in derivative strains TPE2364 (derived from phage P13374 305
released by strain CB13374), TPE2369 (phage P8983), TPE2381 (phage P13771), TPE2383 306
(phage P13772) and TPE2385 (phage P13344) (Table 3). The C600 lysogens were positive 307
for the stx2a gene and converted to Stx2 production as tested by Stx-EIA (Fig. 2). Induction of 308
lysogenic strains with mitomycin C resulted in release of Stx2a phages which were 309
morphologically similar to those of their wildtype host strains. The integration site of all 310
Stx2a phages in C600 was identical to those found in the STEC wildtype strains (see below). 311
Quantification of Shiga toxin production in cultures of STEC O104:H4 and C600 312
lysogens with and without mitomycin C treatment. Stx production was measured with 313
bacteria grown in TSB supplemented with and without mitomycin C. All six STEC O104:H4 314
strains listed in Table 1 produced significantly more (p=0.033) Stx in the presence of 315
mitomycin C compared to untreated cultures. For Stx production there were no significant 316
differences found (p=0.074) between the tested STEC O104:H4 strains and the results 317
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

13
obtained for the six strains are summarized in Fig. 2 A. In the presence of mitomycin C, Stx 318
was still detectable in culture fluid diluted between 1:400 and 1:600. In cultures without 319
mitomycin C treatment, Stx was not detectable in dilutions of 1:100 and higher. 320
C600 lysogens harboring Stx2 phages from STEC O104:H4 wildtype strains (TPE2364, 321
TPE2369, TPE2381, TPE2383 and TPE2385 from Table 3) produced significantly higher 322
amounts of Stx than the wildtype STEC O104:H4 strains when grown in the presence 323
(p=0.023) and without (p<0.001) mitomycin C (summarized in Fig. 2B). 324
In contrast to the STEC O104:H4 wildtype strains, there were no significant differences 325
for Stx production between the C600 lysogens when grown with or without mitomycin C 326
(p=0.397). In order to determine if this was due to an increasing spontaneous induction of Stx 327
phages in the P13374 K-12 lysogen we explored the differences in the spontaneous phage 328
release between TPE2364 (Table 3) and CB13374 over a 24 h bacterial growth period. This 329
was investigated in three independent experiments and different time intervals (6 h, 12 h and 330
24 h) by titration of cell-free supernatants of both strains on E. coli C600. In comparison to 331
the STEC strain CB13374 a higher level of one to two orders of magnitude of released phages 332
in cell-free supernatants of TPE2364 was observed (data not shown). These findings could 333
explain the relatively higher Stx2 levels found in the C600 lysogens. 334
The amounts of Stx2a produced by STEC wildtype strains were found similar to that 335
produced by an Stx2a EHEC O157:[H7] strain (CB6161, stx2a) in cultures with (p=0.605) and 336
without (p=0.81) mitomycin C (data not shown). 337
Morphology of phage particles. Negative staining shows that all investigated strains 338
except CB13437 (Table 1) release phages with a short tail and a hexagonal head (virus 339
family: Podoviridae). Phage tails were 12.7 nm (±1.4 nm; n = 20) in the width and their 340
length diversified up to 40 nm (on average approximately 26 nm). The diameters of the phage 341
heads were 68.5 nm (±2.3 nm) measured between the sides parallels, respectively 70.7 nm 342
(±1.4 nm) measured diagonal to the tail. The phages from all strains (CB8983, CB13344, 343
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

14
CB13374, CB13771 and CB13772) as well as from their C600 lysogens (Table 3) were 344
morphologically very similar to each other (Fig. 3) resembling the Stx2a phage 933W of 345
EHEC O157:H7 strain EDL933 which is also a podovirus (34,39,41,56). In ultrathin sections 346
of C600 infected with P13374, several stages of the phage development were observed (phage 347
attachment, propagation and liberation by bacteriolysis) (Fig. 4A and B). Upon mitomycin C 348
induction the Norwegian strain CB13437 yielded a morphologically different Stx-negative 349
phage of the virus family Myoviridae (Fig. 5). 350
Complete nucleotide sequencing and annotation of Stx2 prophage P13374. The 351
complete nucleotide sequence of the Stx2 prophage P13374 was deduced from the whole 352
genome sequence of the German E. coli O104:H4 outbreak strain CB13374 and confirmed by 353
partial sequencing of relevant regions of the phage DNA of virion particles. Based on this 354
analysis, a genomic sequence of 60,894 bp with a mean G+C content of 50.3% for the 355
temperate phage P13374 was inferred. The annotated prophage genome also includes the left 356
and right core sequences of the attachment sites (13 nt). From detailed bioinformatic analyses 357
(for details see Materials and Methods section) 79 open reading strands (ORFs) with good 358
coding potential were predicted. Furthermore, the investigated prophage genome comprises 359
fourteen putative Rho-independent transcription terminators and three transfer RNAs (tRNAs) 360
(see Table S3 in the supplemental material). The modular organization of the prophage and 361
the specific features of the bioinformatic analyses are summarized in Fig. 6. Further detailed 362
information is given in the supplemental material Table S3. Out of 79 gene products that have 363
been predicted for phage P13374, 62 gene products revealed values of identity ranging from 364
38-100% to annotated proteins of previously characterized Stx-encoding phages (Table S3). 365
According to the predicted function of the suggested gene products on P13374 the genome 366
can be divided into two parts (Fig. 6), left and right half. Moreover, the whole genome 367
organization of P13374 is strongly reminiscent to the Enterobacteria phage sequences TL-368
2011c (JQ011318.1) and VT2phi_272 (HQ424691.1) which originate from E. coli pathotypes 369
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

15
O103:H25 and O157:H7, respectively (28). The annotation of the P13374 genome was 370
arranged according to the organization of the prophage sequence starting with the putative 371
genes int (ORF01) and exc (ORF02) coding for a phage integrase and excisionase, 372
respectively. Both genes are closely related to their functional counterparts on a cryptic 373
prophage sequence of the E. coli O26:H11 strain 11368 (NC_013361.1) and previously 374
described Stx-encoding phages (933W, VT2-Sakai, Stx1-converting phage and Stx2-375
converting phage I & II). Further genes on the left half of the genome code for proteins 376
involved in regulatory circuits, anti-termination, replication, nucleotide metabolism, Shiga 377
toxin production, and host cell lysis. The core sequences of the genetic switch of P13374 378
show a similar organisation to Lambda-like phages but consist only of the two antagonistic 379
repressor genes cI and cro which code for the prophage repressor CI and the lytic repressor 380
Cro. Whereas the two repressor proteins could be identified due to their close relationships to 381
Lambda-like phage repressors, it was not possible to find operator-sites in the intergenic 382
region between these repressor genes. Furthermore, genes encoding proteins like CII and CIII 383
which are involved in the regulation of the genetic switch of Lambda-like phages could not be 384
identified on the genome of P13374. This finding suggests that P13374 carries a regulation 385
mechanism for the lysogenic / lytic cycle which is only distantly related to Lambda-like 386
phages. Further analyzes have to be performed to determine the specificity of the repressors 387
(Cro and CI) to its operator sequences. Downstream of the repressor genes cI and cro 388
functional counterparts of genes for a bacteriophage-related helicase and primase were 389
identified. Both gene products might be of key importance for the replication of the phage 390
genome. While helicases act on DNA to separate the double-stranded DNA into single-391
strands, primases catalyze the synthesis of a short RNA or ssDNA fragments to initiate 392
complementary strand synthesis. Downstream of the Shiga toxin subunits A and B encoding 393
genes the functional module for cell wall degradation and the lysis of the host cell was 394
identified. This module consists of the two essential genes which are typically transcribed 395
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

16
from adjacent ORFs and serve as functional counterparts to holins (Gp50) and endolysins 396
(Gp51). 397
The right half of the P13374 genome begins with the small and the large subunit of the 398
terminase genes (terS and terL, Fig. 6). Both putative proteins are closely related to their 399
functional counterparts on phage genomes 933W, VT2-Sakai and other Shiga toxin-400
converting phages (86, Stx1, Stx2-I and Stx2-II). Further genes of the right half of the P13374 401
genome might represent a cluster of structural proteins involved in virion morphology and 402
synthesis (Fig. 6). Almost all annotated P13374 gene products encoding proteins for the 403
phage morphology have functional counterparts to proteins of the virion structure module of 404
Stx-converting phages (Fig. 6). An unusual feature of the tail gene cluster (gp65, gp66 & 405
gp69) is the 2,043 bp open reading frame ORF65. The putative gene product of this ORF has 406
functional counterparts in characterized phages which are of key importance for the 407
adsorption specificity of a phage to its host cell. From the bioinformatic analyses it can be 408
suggested that the specificity of phage P13374 follows the presence of short (gp65) and distal 409
(gp66, gp69) tail fibers. Self-comparison analyses of the phage tail fiber protein (gp65) shows 410
a large insertion sequence which consists of several imperfect direct repeats (data not shown). 411
Some putative gene products of P13374 were confirmed by SDS-PAGE and mass 412
spectrometric analyses as the main structural proteins of the virion particle (see below). 413
Genetic similarity of phage P13374 with other E. coli phages. Overall the gene 414
content and the structural organization of several functional modules are similar to most of the 415
previously described Stx-converting phages. As summarized in Fig. 7 gene products of TL-416
2011c and VT2phi272 show the highest degree of similarity to the predicted P13374 proteins. 417
58 and 57 putative gene products of phage P13374 (79 gene products total) are related (51 to 418
100%) to TL-2011c and VT2phi272, respectively. The highest degree of similarity to 419
previously characterized Stx-converting phages was determined to 933W (AF125520), Min27 420
(NC_010237), VT2-Sakai (AP000422) and Stx1 & Stx2-converting bacteriophages 421
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

17
(AP005153 and AP0044202). The Enterobacteria phages HK97 (NC_002167) and Lambda 422
(J02459) and the Stx2c phage 2851 (FM180578) share only lower numbers of related proteins 423
with P13374 (see Table S3 in the supplemental material). 424
Except for three singular stretches (genome region: 5,250 to 6,750; 29,100 to 31,300 425
and 40,900 to 43,800) of the P13374 genome that exhibit no significant homology to known 426
Stx-encoding phages, the remaining P13374 DNA is 99-100% identical to E. coli phage TL-427
2011c (Fig. 7). On average, P13374 and TL-2011c DNAs are 90% identical. Interestingly, the 428
one major difference between the two phage genomes concerns the putative outer membrane 429
lipoprotein (Bor protein precursor) homolog whose coding sequence is present on TL-2011c 430
genome and which is widespread among a number of other lambdoid coliphages. A functional 431
counterpart of the TL-2011c Bor-like protein which might promote bacterial resistance to 432
serum complement killing could not be identified on P13374. Further differences were 433
identified in the tail gene clusters of P13374 and TL-2011c. In contrast to P13374 which 434
comprise the information for a long tail fiber adhesin (gp66) downstream of tail fiber protein 435
encoding ORF65, a related gene product on TL-2011c could not be identified. Therefore it 436
can be speculated that TL-2011c and P13374 form different types of tail fibers. Further 437
differences between the two phages are restricted to sequences for which no function can be 438
predicted. 439
Analysis of P13374 proteins. To verify the bioinformatic analysis of the phage 440
genome, major protein bands from phage preparations were excised from one-dimensional 441
SDS PAGE gels and subjected to tryptic in-gel digestion followed by tandem MALDI-TOF 442
mass spectrometry (MS/MS) identification. In this way four abundant proteins could be 443
assigned to putative coding regions (ORF58, 60, 61, 78) of the genome sequence of phage 444
P13374 (Fig. 6, Table 4). These ORFs most likely represent structural phage proteins. 445
The E. coli wrbA locus is the integration site of Stx2a phages from STEC O104:H4 446
strains. STEC O104:H4 strains and C600 Stx2 phage lysogens were investigated by PCR for 447
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

18
different known Stx phage insertion site as previously described (11), see Fig. 8. A 500 bp 448
product was obtained for all O104:H4 strains and C600 Stx2 phage lysogens (Table 3) with 449
primers ML1 and ML2 (Table 2) spanning the region between the E. coli wrbA gene and the 450
phage integrase gene, thus defining the left integration site of the prophage. Nucleotide 451
sequence analysis of the PCR fragments obtained with ML1+ML2 primers indicated that the 452
Stx2 prophages are integrated in the 5’-end of the coding region of the E. coli wrbA gene 453
(AP009048 in E. coli K-12 W3110). The core region for attachment in the bacterial genome 454
attB is a 13 bp long sequence GTTTCAATATGTC. Due to the insertion of the Stx2 phage the 455
coding region of the wrbA gene is altered. 456
Primer orf79 binding to the 3’-region of the P13374 prophage sequence and primer 457
wrbA-R8 (Table 2) were used to identify the right integration site of the prophage into the 458
interrupted wrbA gene. A 256 bp PCR-fragment was investigated for its nucleotide sequence. 459
PCR amplification of the phage DNA from P13374 virion particles using primers orf79 and 460
ML2 (int gene) yielded a PCR product of 473 bp and revealed the phage attachment site attP 461
GTTTCAACATGTC. The attachment sites attB and attP were found to be different in one 462
nucleotide position (underlined nucleotides, see also Fig. 8). 463
From lysates of phages P8983, P13344, P13374, P13771 and P13772 the same PCR 464
product of 473 bp was amplified, whereas no PCR product was obtained with the stx-negative 465
phage from strain CB13437 as template. Nucleotide sequencing showed that all phage 466
genomes in virion particles are closed between ORF79 (hypothetical protein) and the 467
integrase (ORF01) gene. Thus, the sequence analyses of phage DNAs revealed that the 468
prophage genomes between ORF79 and the integrase gene are joined which leads to circular 469
permuted genomes of the phage. 470
Immunity of C600 Stx phage lysogens to superinfection with Stx2a phages from 471
STEC O104:H4 strains. To investigate the immunity between the Stx2 phages from STEC 472
O104:H4 strains and other Stx-encoding bacteriophages plaque assays were performed. For 473
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

19
these examinations phage lysates originating from the German and Georgian STEC O104:H4 474
strains were used on different C600 Stx1- and Stx2-encoding phages lysogens (Table 3). 475
C600 was used as a phage negative control. The Stx2 phages from the German STEC 476
O104:H4 strains did not form plaques on C600 lysogens carrying the phages P8983, P13344 477
and P13374. On all other C600 derivates including strains TPE2381 and TPE2383 which 478
carry the Stx2a phages from the Georgian STEC O104:H4 strains a lytic phage activity was 479
determined. In contrast, the Stx2 phages from the Georgian O104:H4 strains (P13771 and 480
P13772) did not lyse C600 lysogen carrying the Stx2 phages from the German outbreak 481
strains (TPE2364, TPE2369 and TPE2385) nor from the Georgian O104:H4 strains (TPE2381 482
and TPE2383). 483
Investigation of enteroaggregative E. coli (EAEC) for susceptibility to the Stx2a-484
encoding bacteriophage P13374. To obtain information about the host range of the Stx2a 485
phage P13374, phage preparations with approx. 2 x 109 pfu were plated on enteroaggregative 486
E. coli strains. However, none of 31 EAEC strains belonging to 22 different O:H serotypes 487
(see Table S1 in the supplemental material) showed plaque formation or lysis. C600 was used 488
as phage sensitive control. Subcultures of bacteria grown in zones spotted with P13374 489
preparations were found negative for the stx2 gene by PCR indicating that lysogenization with 490
P13374 had not occurred. The 31 STEC strains were further investigated by PCR for the 491
presence of an intact wrbA gene which is the E. coli host integration site for P13374. All but 492
one of the EAEC strains (CB7373, O111:H10) yielded a 596 bp PCR product with primers 493
wrbA-F / wrbA-R which amplify the complete wrbA gene encompassing the bacterial 494
attachment site for P13374 integration. These results indicate that despite the presence of the 495
potential phage integration site, phage P13374 is restricted for its host range among E. coli 496
belonging to the EAEC group. 497
498
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

20
DISCUSSION 499
500
Enteroaggregative hemorrhagic E. coli (STEC) O104:H4 is a new type of emerging 501
pathogen which has been identified as the causative agent in one of the world’s largest 502
outbreaks of foodborne illness in humans resulting in 855 HUS and in 53 fatal cases (7,19). 503
There are several models for the evolution of the German STEC O104:H4 strains. STEC 504
O104:H4 could have evolved from an enteroaggregative E. coli (EAEC) ancestor by the 505
uptake of Stx2 phages into the chromosome of the bacterium (10,40). It is also possible that 506
an Stx2 O104:H4 strain was the progenitor of both, HUSEC41 (Germany 2001) and the 2011 507
outbreak strains (LB226692) as described by Mellmann et al. 2011 (32). The latter model 508
could explain our findings, that the the Stx2 phages found in CB8983 (Germany 2001) and 509
CB13344, CB13374 (Germany 2011) were highly similar to each other (this work). The 510
increased human virulence potential of the STEC O104:H4 strain compared to any EAEC 511
O104:H4 progenitor probably results from production of phage encoded Stx2a combined with 512
the efficient enteroaggregative colonization of the human intestine. These properties in 513
combination are believed to be the cause of maximum delivery of Stx into the human host 514
resulting in augmented numbers of cases with severe clinical illness (7). 515
The presence of Stx phages in EAEC strains is unusual and apart from EAEC 516
O104:H4 only a few other EAEC serotype strains were reported to carry stx2 genes (25,33). 517
In this work we were interested to explore the characteristics of the Stx phages present in 518
STEC O104:H4 strains isolated in the 2011 German outbreak and from earlier, 519
epidemiologically unrelated episodes of human infections with STEC O104:H4. 520
Contaminated sprouted seeds produced at a horticultural farm in northern Germany 521
were identified as the food source of the STEC O104:H4 2011 outbreak strain (4,18). We 522
determined the nucleotide sequence of the Stx2 phage present in the STEC O104:H4 strain 523
CB13374 isolated from sprouted seeds produced at this farm. As this strain is believed to be 524
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

21
the origin of the pandemic spread of STEC O104:H4 in May/June 2011 we compared its Stx2 525
phage to those present in STEC O104:H4 strains from human patients in Germany (2001 and 526
2011), Norway (2006) and Georgia (2009). Although all of the STEC O104:H4 strains carried 527
stx2 genes, Stx2 phages could only be isolated from the German and the Georgian O104:H4 528
strains. All of these phages were identified as podoviruses and were morphologically similar 529
to the Stx2 phage 933W carried by EHEC O157:H7 strain EDL933 (34). Similar to phage 530
933W, the Stx2a phages harbored by the German and the Georgian O104:H4 strains were 531
found to be integrated in the wrbA locus in the wildtype STEC O104:H4 and in E. coli K-12 532
phage lysogenic strains (37). Although we found evidence for the presence of a similar Stx2a 533
prophage in the Norwegian STEC O104:H4 strain CB13437, viable Stx phages could not be 534
isolated, instead a morphologically different bacteriophage of the Myoviridae family which 535
does not encode Stx was released from CB13437. Interestingly, myoviruses were not found in 536
mitomycin C induced cultures of the German and the Georgian O104:H4 strains. 537
Surprisingly, the Stx2 phages isolated from the 2011 German outbreak STEC 538
O104:H4 strains (sprouts and patients) were indistinguishable for their restriction 539
endonuclease profiles from the Stx2 phage harbored by the STEC O104:H4 strain CB8983 540
which was isolated in 2001 from the feces of a child with HC in Germany. Phage genome 541
sequencing of P8983 revealed >99.9% identity with the genome of P13374 (R. Reinhardt, 542
unpublished results). In contrast, Stx2 phages from STEC O104:H4 strains from two patients 543
in Georgia (2009) showed clearly altered endonuclease restriction profiles compared to the 544
German STEC O104:H4 strains. Differences between the German and the Georgian O104:H4 545
Stx2 phages were also found in their bacterial host range and superinfection susceptibility. 546
The phage encoded prophage repressor protein binds efficiently to specific sites in the 547
immunity region on the phage genome. This mechanism is highly specific as most of the 548
temperate phages only bind to its own DNA and not that from other closely related phages. 549
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

22
All Stx2 phages from STEC O104:H4 strains showed lysis and plaque formation on 550
E. coli K-12 strains harboring phage Lambda or different lambdoid Stx1 and Stx2 phages. 551
These observations suggest that the Stx phages from the STEC O104:H4 strains and 552
previously characterized Stx1- and Stx2-encoding phages from other STEC serotypes exhibit 553
different prophage repressor specificities. This is in strong concordance with the findings 554
obtained from the DNA sequence analysis of the primary immunity region of phage P13374. 555
No related Lambda-like or other Stx phage-like prophage repressor or Cro repressor binding 556
sites (operators) could be identified in the predicted immunity region (cI to cro) of P13374. 557
Differences between the German and the Georgian E. coli O104:H4 derived Stx2 558
phages were also found in their bacterial host range and superinfection susceptibility. The 559
Stx2 phages from the German O104:H4 strains possessed a more extended lysis spectrum 560
compared to the Stx2 phages from the Georgian O104:H4 strains pointing to possible 561
differences in their immunity regions. This needs to be confirmed by nucleotide analysis of 562
the Stx2 phages harbored by the Georgian STEC O104:H4 strains. 563
We therefore investigated EAEC strains of different serotypes for their susceptibility 564
to P13374 as a prototype of the highly virulent Stx2 phage from the German STEC O104:H4 565
strains and despite the presence of an undisrupted wrbA gene in most of the EAEC recipient 566
strains, these strains were found to be resistant to a high infective dose of P13374. We do not 567
know the mechanisms which confer resistance to the Stx2 phage but this observation might 568
indicate that the generation of STEC by uptake of Stx phages in nature is a rare event. 569
Nevertheless, under so far unknown conditions Stx phages are able to infect EAEC strains 570
thus pointing to an extended host range. Previous findings about the presence of Stx phages 571
and stx genes in Enterobacteriaceae other than E. coli also show this possibility (35,36,52). 572
It is remarkable that despite the genomic differences found between the German STEC 573
O104:H4 strains isolated in 2001 and in 2011 both harbor the same Stx2 phage. Strain 574
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

23
CB8983 from 2001 differs from the outbreak 2011 strains in the composition of its EAEC 575
plasmid, the presence of a plasmid encoding heat stable enterotoxins and the presence of 576
genes conferring antibiotic resistance (7). 577
The original source of human infections and outbreaks with STEC O104:H4 reported 578
from different countries has not been identified in any case. As EAEC and STEC strains were 579
not found associated with animals but only with humans, a human source is supposed as most 580
likely (4). Based on similarities found between STEC O104:H4 strains that caused the 2011 581
outbreak in Germany and a satellite outbreak in France in June of the same year, a batch of 582
fenugreek seeds imported from Egypt which served for sprout production in Germany and 583
France was suspected as the origin of the outbreaks in both countries (18). Analysis of SNPs 584
(single-nucleotide polymorphisms) of STEC O104:H4 strains isolated in Germany and in 585
France in 2011 revealed a higher diversity in the French strains than in the German isolates. 586
This is surprising as the French outbreak was confined to only 15 persons. This finding raises 587
questions about the analysis of the source of the STEC O104:H4 outbreak in Europe in 2011 588
(22). 589
The high similarity observed between the Stx2 phages present in STEC O104:H4 590
strains isolated in Germany in 2001 and 2011 is surprising in light of the Egyptian origin of 591
the suspect seeds, the amount of time between their isolation, the considerable genetic 592
heterogeneity generally seen among Stx phages in nature (23) and the finding that the phages 593
harbored by the Georgian isolates showed distinct differences to the German STEC O104:H4 594
phages. The sequence of the Norwegian Stx2 prophage in strain CB13437 has yet to be 595
determined. 596
It has previously been shown, that STEC O104:H4 can survive in a viable but non-597
culturable state (VBNC) over long periods of time (2). During and after the outbreak it has 598
also been shown that asymptomatic human carriers may serve as a natural reservoir for STEC 599
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

24
O104:H4 strains (4,42). Further research is needed on Stx2 phages present in other STEC 600
O104:H4 strains to determine their potential for long-term survival outside the bacterial host. 601
As with many STEC and EHEC strains, Stx production in STEC O104:H4 strains 602
could be increased with mitomycin C resulting in a significant enhancement of Stx2 603
production and delivery (5,48). Similar results were reported with ciprofloxacin as inducer for 604
stx2 transcription, Stx2 production as well as release of Stx phages from STEC O104:H4 605
strains (6). However, a basal level of of Stx2 production was still observed in the absence of 606
mitomycin C in STEC O104:H4 wildtype strains confirming previous findings made on other 607
STEC serotype strains harboring Stx2 encoding bacteriophages (13). In our study, the amount 608
of Stx2 produced by the STEC O104:H4 strains was not significantly different from that of an 609
Stx2a-producing EHEC O157:H7 strain. Similar results were reported when ciprofloxacin 610
was used as inducer to compare Stx2 production of STEC O104:H4 and O157:H7 strains (6). 611
These findings suggest that the augmented virulence of STEC O104:H4 is not related to 612
enhanced production of Stx2, but rather to the efficient colonization of the STEC strains 613
allowing sustained delivery of Stx2a into the host intestine. Genetic studies of additional Stx 614
prophages from STEC O104:H4 strains will allow us to determine if P13374 is a prototype 615
phage for the conversion of EAEC strains to Shiga toxin production. 616
617
ACKNOWLEDGMENTS 618
The authors are grateful to Sabine Haby, Antje Konietzny, Karin Pries, Katja Steege and 619
Maria-Margarida Vargas for technical assistance. J. A. Hammerl was supported by a grant of 620
the BMBF-funded project “SiLeBAT.” 621
622
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

25
Reference List 623
624
1. Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller, and 625
D. J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein 626
database search programs. Nucleic Acids Res. 25:3389-3402. 627
2. Aurass, P., R. Prager, and A. Flieger. 2011. EHEC/EAEC O104:H4 strain linked with 628
the 2011 German outbreak of haemolytic uremic syndrome enters into the viable but 629
non-culturable state in response to various stresses and resuscitates upon stress relief. 630
Environ. Microbiol. 13:3139-3148. 631
3. Baker, D. R., R. A. Moxley, and D. H. Francis. 1997. Variation in virulence in the 632
gnotobiotic pig model of O157:H7 Escherichia coli strains of bovine and human 633
origin. Adv. Exp. Med. Biol. 412:53-58. 634
4. Beutin, L. and A. Martin. 2012. Outbreak of Shiga toxin-producing Escherichia coli 635
(STEC) O104:H4 infection in Germany causes a paradigm shift with regard to human 636
pathogenicity of STEC strains. J. Food Prot. 75:408-418. 637
5. Beutin, L., H. Steinruck, G. Krause, K. Steege, S. Haby, G. Hultsch, and B. Appel. 638
2007. Comparative evaluation of the Ridascreen(R) Verotoxin enzyme immunoassay 639
for detection of Shiga-toxin producing strains of Escherichia coli (STEC) from food 640
and other sources. J. Appl. Microbiol. 102:630-639. 641
6. Bielaszewska, M., E. A. Idelevich, W. Zhang, A. Bauwens, F. Schaumburg, A. 642
Mellmann, G. Peters, and H. Karch. 2012. Effects of Antibiotics on Shiga Toxin 2 643
Production and Bacteriophage Induction by Epidemic Escherichia coli O104:H4 644
Strain. Antimicrob. Agents Chemother. 56:3277-3282. 645
7. Bielaszewska, M., A. Mellmann, W. Zhang, R. Kock, A. Fruth, A. Bauwens, G. 646
Peters, and H. Karch. 2011. Characterisation of the Escherichia coli strain associated 647
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

26
with an outbreak of haemolytic uraemic syndrome in Germany, 2011: a 648
microbiological study. Lancet Infect Dis. 11:671-676. 649
8. Blanco, J., M. Blanco, J. Blanco, A. Mora, M. P. Alonso, E. A. Gonzalez, and M. I. 650
Bernardez. 2001. Epidemiology of Verocytotoxigenic Escherichia coli (VTEC) in 651
ruminants., p. 113-148. In: G. Duffy, P. Garvey, and D. A. McDowell (eds.), 652
Verocytotoxigenic E. coli. Food & Nutrition Press, Inc., Trumbull. 653
9. Brown, C. M., M. E. Dalphin, P. A. Stockwell, and W. P. Tate. 1993. The 654
translational termination signal database. Nucleic Acids Res. 21:3119-3123. 655
10. Brzuszkiewicz, E., A. Thurmer, J. Schuldes, A. Leimbach, H. Liesegang, F. D. 656
Meyer, J. Boelter, H. Petersen, G. Gottschalk, and R. Daniel. 2011. Genome 657
sequence analyses of two isolates from the recent Escherichia coli outbreak in 658
Germany reveal the emergence of a new pathotype: Entero-Aggregative-659
Haemorrhagic Escherichia coli (EAHEC). Arch. Microbiol. 193(12):883-91. 660
11. Bugarel, M., L. Beutin, F. Scheutz, E. Loukiadis, and P. Fach. 2011. Identification 661
of genetic markers for differentiation of Shiga toxin-producing, enteropathogenic and 662
avirulent strains of Escherichia coli O26. Appl. Environ. Microbiol. 77:2275-2281. 663
12. CDC. 2011. Investigation update: Outbreak of Shiga toxin-producing E. coli O104 664
(STEC O104:H4) infections associated with travel to Germany. Centers for Disease 665
Control and Prevention, Atlanta, USA http://www.cdc.gov/ecoli/2011/ecolio104/. 666
13. de Sablet, T., Y. Bertin, M. Vareille, J. P. Girardeau, A. Garrivier, A. P. Gobert, 667
and C. Martin. 2008. Differential expression of stx2 variants in Shiga toxin-668
producing Escherichia coli belonging to seropathotypes A and C. Microbiology 669
154:176-186. 670
14. Dowd, S. E. 2007. Escherichia coli O157:H7 gene expression in the presence of 671
catecholamine norepinephrine. FEMS Microbiol. Lett. 273:214-223. 672
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

27
15. Ermolaeva, M. D., H. G. Khalak, O. White, H. O. Smith, and S. L. Salzberg. 673
2000. Prediction of transcription terminators in bacterial genomes. J. Mol. Biol. 674
301:27-33. 675
16. Ethelberg, S., K. E. Olsen, F. Scheutz, C. Jensen, P. Schiellerup, J. Enberg, A. M. 676
Petersen, B. Olesen, P. Gerner-Smidt, and K. Molbak. 2004. Virulence factors for 677
hemolytic uremic syndrome, Denmark. Emerg. Infect. Dis. 10:842-847. 678
17. European Food Safety Authority. 2011. Shiga toxin-producing E. coli (STEC) 679
O104:H4 2011 outbreaks in Europe: Taking stock. EFSA Journal 9:2390. 680
18. European Food Safety Authority, EFSA. Tracing seeds, in particular fenugreek 681
(Trigonella foenum-graecum) seeds, in relation to the Shiga-toxin-producing E. coli 682
(STEC) O104:H4 2011 outbreaks in Germany and France. European Food Safety 683
Authority. 1-23. 5-7-2011. 684
19. Frank, C., M. Faber, M. Askar, H. Bernard, A. Fruth, A. Gilsdorf, M. Hohle, H. 685
Karch, G. Krause, R. Prager, A. Spode, K. Stark, and D. Werber. 2011. Large and 686
ongoing outbreak of haemolytic uraemic syndrome, Germany, May 2011. Euro. 687
Surveill. 16. 688
20. Friedrich, A. W., M. Bielaszewska, W. L. Zhang, M. Pulz, T. Kuczius, A. 689
Ammon, and H. Karch. 2002. Escherichia coli harboring Shiga toxin 2 gene 690
variants: frequency and association with clinical symptoms. J. Infect. Dis. 185:74-84. 691
21. Friedrich, A. W., J. Borell, M. Bielaszewska, A. Fruth, H. Tschape, and H. 692
Karch. 2003. Shiga toxin 1c-producing Escherichia coli strains: phenotypic and 693
genetic characterization and association with human disease. J. Clin. Microbiol. 694
41:2448-2453. 695
22. Grad, Y. H., M. Lipsitch, M. Feldgarden, H. M. Arachchi, G. C. Cerqueira, M. 696
Fitzgerald, P. Godfrey, B. J. Haas, C. I. Murphy, C. Russ, S. Sykes, B. J. Walker, 697
J. R. Wortman, S. Young, Q. Zeng, A. Abouelleil, J. Bochicchio, S. Chauvin, T. 698
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

28
Desmet, S. Gujja, C. McCowan, A. Montmayeur, S. Steelman, J. Frimodt-Moller, 699
A. M. Petersen, C. Struve, K. A. Krogfelt, E. Bingen, F. X. Weill, E. S. Lander, C. 700
Nusbaum, B. W. Birren, D. T. Hung, and W. P. Hanage. 2012. Genomic 701
epidemiology of the Escherichia coli O104:H4 outbreaks in Europe, 2011. Proc. Natl. 702
Acad. Sci. U.S.A. 109:3065-3070. 703
23. Herold, S., H. Karch, and H. Schmidt. 2004. Shiga toxin-encoding bacteriophages-704
genomes in motion. Int. J. Med. Microbiol. 294:115-121. 705
24. Huang, D. B., A. Mohanty, H. L. Dupont, P. C. Okhuysen, and T. Chiang. 2006. 706
A review of an emerging enteric pathogen: enteroaggregative Escherichia coli. J. Med. 707
Microbiol. 55:1303-1311. 708
25. Iyoda, S., K. Tamura, K. Itoh, H. Izumiya, N. Ueno, K. Nagata, M. Togo, J. 709
Terajima, and H. Watanabe. 2000. Inducible stx2 phages are lysogenized in the 710
enteroaggregative and other phenotypic Escherichia coli O86:HNM isolated from 711
patients. FEMS Microbiol. Lett. 191:7-10. 712
26. Kim, J., K. Oh, S. Jeon, S. Cho, D. Lee, S. Hong, S. Cho, M. Park, D. Jeon, and S. 713
Kim. 2011. Escherichia coli O104:H4 from 2011 European outbreak and strain from 714
South Korea. Emerg. Infect. Dis. 17:1755-1756. 715
27. Koch, C., S. Hertwig, R. Lurz, B. Appel, and L. Beutin. 2001. Isolation of a 716
lysogenic bacteriophage carrying the stx(1(OX3)) gene, which is closely associated with 717
Shiga toxin-producing Escherichia coli strains from sheep and humans. J. Clin. 718
Microbiol. 39:3992-3998. 719
28. L'abee-Lund, T. M., H. J. Jorgensen, K. O'Sullivan, J. Bohlin, G. Ligard, P. E. 720
Granum, and T. Lindback. 2012. The highly virulent 2006 Norwegian EHEC 721
O103:H25 outbreak strain is related to the 2011 German O104:H4 outbreak strain. 722
PLoS One 7:e31413. 723
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

29
29. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head 724
of bacteriophage T4. Nature 227:680-685. 725
30. Lowe, T. M. and S. R. Eddy. 1997. tRNAscan-SE: a program for improved detection 726
of transfer RNA genes in genomic sequence. Nucleic Acids Res. 25:955-964. 727
31. Makino, K., K. Yokoyama, Y. Kubota, C. H. Yutsudo, S. Kimura, K. Kurokawa, 728
K. Ishii, M. Hattori, I. Tatsuno, H. Abe, T. Iida, K. Yamamoto, M. Onishi, T. 729
Hayashi, T. Yasunaga, T. Honda, C. Sasakawa, and H. Shinagawa. 1999. 730
Complete nucleotide sequence of the prophage VT2-Sakai carrying the verotoxin 2 731
genes of the enterohemorrhagic Escherichia coli O157:H7 derived from the Sakai 732
outbreak. Genes Genet. Syst. 74:227-239. 733
32. Mellmann, A., D. Harmsen, C. A. Cummings, E. B. Zentz, S. R. Leopold, A. Rico, 734
K. Prior, R. Szczepanowski, Y. Ji, W. Zhang, S. F. McLaughlin, J. K. Henkhaus, 735
B. Leopold, M. Bielaszewska, R. Prager, P. M. Brzoska, R. L. Moore, S. 736
Guenther, J. M. Rothberg, and H. Karch. 2011. Prospective genomic 737
characterization of the German enterohemorrhagic Escherichia coli O104:H4 outbreak 738
by rapid next generation sequencing technology. PLoS One 6:e22751. 739
33. Morabito, S., H. Karch, P. Mariani-Kurkdjian, H. Schmidt, F. Minelli, E. Bingen, 740
and A. Caprioli. 1998. Enteroaggregative, Shiga toxin-producing Escherichia coli 741
O111:H2 associated with an outbreak of hemolytic-uremic syndrome. J. Clin. 742
Microbiol. 36:840-842. 743
34. O'Brien, A. D., L. R. Marques, C. F. Kerry, J. W. Newland, and R. K. Holmes. 744
1989. Shiga-like toxin converting phage of enterohemorrhagic Escherichia coli strain 745
933. Microb. Pathog. 6:381-390. 746
35. Ooka, T., K. Seto, K. Kawano, H. Kobayashi, Y. Etoh, S. Ichihara, A. Kaneko, J. 747
Isobe, K. Yamaguchi, K. Horikawa, T. A. Gomes, A. Linden, M. Bardiau, J. G. 748
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

30
Mainil, L. Beutin, Y. Ogura, and T. Hayashi. 2012. Clinical Significance of 749
Escherichia albertii. Emerg. Infect. Dis. 18:488-492. 750
36. Paton, A. W. and J. C. Paton. 1996. Enterobacter cloacae producing a Shiga-like 751
toxin II-related cytotoxin associated with a case of hemolytic-uremic syndrome. J. 752
Clin. Microbiol. 34:463-465. 753
37. Perna, N. T., G. Plunkett, III, V. Burland, B. Mau, J. D. Glasner, D. J. Rose, G. F. 754
Mayhew, P. S. Evans, J. Gregor, H. A. Kirkpatrick, G. Posfai, J. Hackett, S. 755
Klink, A. Boutin, Y. Shao, L. Miller, E. J. Grotbeck, N. W. Davis, A. Lim, E. T. 756
Dimalanta, K. D. Potamousis, J. Apodaca, T. S. Anantharaman, J. Lin, G. Yen, 757
D. C. Schwartz, R. A. Welch, and F. R. Blattner. 2001. Genome sequence of 758
enterohaemorrhagic Escherichia coli O157:H7. Nature 409:529-533. 759
38. Persson, S., K. E. Olsen, S. Ethelberg, and F. Scheutz. 2007. Subtyping method for 760
Escherichia coli Shiga toxin (verocytotoxin) 2 variants and correlations to clinical 761
manifestations. J. Clin. Microbiol. 45:2020-2024. 762
39. Plunkett III, G., D. J. Rose, T. J. Durfee, and F. R. Blattner. 1999. Sequence of 763
Shiga toxin 2 phage 933W from Escherichia coli O157:H7: Shiga toxin as a phage 764
late-gene product. J. Bacteriol. 181:1767-1778. 765
40. Rasko, D. A., D. R. Webster, J. W. Sahl, A. Bashir, N. Boisen, F. Scheutz, E. E. 766
Paxinos, R. Sebra, C. S. Chin, D. Iliopoulos, A. Klammer, P. Peluso, L. Lee, A. O. 767
Kislyuk, J. Bullard, A. Kasarskis, S. Wang, J. Eid, D. Rank, J. C. Redman, S. R. 768
Steyert, J. Frimodt-Moller, C. Struve, A. M. Petersen, K. A. Krogfelt, J. P. 769
Nataro, E. E. Schadt, and M. K. Waldor. 2011. Origins of the E. coli strain causing 770
an outbreak of hemolytic-uremic syndrome in Germany. N. Engl. J. Med. 365:709-771
717. 772
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

31
41. Rietra, P. J., G. A. Willshaw, H. R. Smith, A. M. Field, S. M. Scotland, and B. 773
Rowe. 1989. Comparison of Vero-cytotoxin-encoding phages from Escherichia coli of 774
human and bovine origin. J. Gen. Microbiol. 135:2307-2318. 775
42. Robert Koch Institut. Abschließende Darstellung und Bewertung der 776
epidemiologischen Erkenntnisse im EHEC O104:H4 Ausbruch. Deutschland 2011. 777
pp. 1-43. 1-9-2011. Berlin, Robert Koch Institut. 778
43. Rohde, H., J. Qin, Y. Cui, D. Li, N. J. Loman, M. Hentschke, W. Chen, F. Pu, Y. 779
Peng, J. Li, F. Xi, S. Li, Y. Li, Z. Zhang, X. Yang, M. Zhao, P. Wang, Y. Guan, Z. 780
Cen, X. Zhao, M. Christner, R. Kobbe, S. Loos, J. Oh, L. Yang, A. Danchin, G. F. 781
Gao, Y. Song, Y. Li, H. Yang, J. Wang, J. Xu, M. J. Pallen, J. Wang, M. 782
Aepfelbacher, and R. Yang. 2011. Open-source genomic analysis of Shiga toxin-783
producing E. coli O104:H4. N. Engl. J. Med. 365:718-724. 784
44. Roschanski, N., S. Klages, R. Reinhardt, M. Linscheid, and E. Strauch. 2011. 785
Identification of genes essential for prey-independent growth of Bdellovibrio 786
bacteriovorus HD100. J. Bacteriol. 193:1745-1756. 787
45. Sato, T., T. Shimizu, M. Watarai, M. Kobayashi, S. Kano, T. Hamabata, Y. 788
Takeda, and S. Yamasaki. 2003. Distinctiveness of the genomic sequence of Shiga 789
toxin 2-converting phage isolated from Escherichia coli O157:H7 Okayama strain as 790
compared to other Shiga toxin 2-converting phages. Gene 309:35-48. 791
46. Scheutz, F., N. E. Moller, J. Frimodt-Moller, N. Boisen, S. Morabito, R. Tozzoli, 792
J. Nataro, and A. Caprioli. 2011. Characteristics of the enteroaggregative Shiga 793
toxin/verotoxin-producing Escherichia coli O104:H4 strain causing the outbreak of 794
haemolytic uraemic syndrome in Germany, May to June 2011. Euro. Surveill. 16. 795
47. Scheutz, F. and N. A. Strockbine. 2005. Genus I. Escherichia, p. 607-624. In: G. M. 796
Garrity, D. J. Brenner, N. R. Krieg, and J. T. Staley (eds.), Bergey's Manual of 797
Systematic Bacteriology. 2nd ed. Springer. 798
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

32
48. Shimizu, T., Y. Ohta, and M. Noda. 2009. Shiga toxin 2 is specifically released from 799
bacterial cells by two different mechanisms. Infect. Immun. 77:2813-2823. 800
49. Strauch, E., J. A. Hammerl, A. Konietzny, S. Schneiker-Bekel, W. Arnold, A. 801
Goesmann, A. Puhler, and L. Beutin. 2008. Bacteriophage 2851 is a prototype 802
phage for dissemination of the Shiga toxin variant gene 2c in Escherichia coli 803
O157:H7. Infect. Immun. 76:5466-5477. 804
50. Strauch, E., R. Lurz, and L. Beutin. 2001. Characterization of a Shiga toxin-805
encoding temperate bacteriophage of Shigella sonnei. Infect. Immun. 69:7588-7595. 806
51. Takkinen, J., Struelens, M., Niskanen, T., Makela, P., Rizzi, V., Caprioli, A., and 807
Scheutz, F. 2011. Shiga toxin/verotoxin-producing Escherichia coli in humans, food 808
and animals in the EU/EEA, with special reference to the German outbreak strain 809
STEC O104. pp. 1-18. 1-6-2011. Stockholm, ECDC & EFSA. 810
52. Tschape, H., R. Prager, W. Streckel, A. Fruth, E. Tietze, and G. Bohme. 1995. 811
Verotoxinogenic Citrobacter freundii associated with severe gastroenteritis and cases 812
of haemolytic uraemic syndrome in a nursery school: green butter as the infection 813
source. Epidemiol. Infect. 114:441-450. 814
53. Tzschoppe, M., A. Martin, and L. Beutin. 2012. A rapid procedure for the detection 815
and isolation of enterohaemorrhagic Escherichia coli (EHEC) serogroup O26, O103, 816
O111, O118, O121, O145 and O157 strains and the aggregative EHEC O104:H4 strain 817
from ready-to-eat vegetables. Int. J. Food Microbiol. 152:19-30. 818
54. Wagner, P. L., D. W. Acheson, and M. K. Waldor. 2001. Human neutrophils and 819
their products induce Shiga toxin production by enterohemorrhagic Escherichia coli. 820
Infect. Immun. 69:1934-1937. 821
55. Wagner, P. L., J. Livny, M. N. Neely, D. W. Acheson, D. I. Friedman, and M. K. 822
Waldor. 2002. Bacteriophage control of Shiga toxin 1 production and release by 823
Escherichia coli. Mol. Microbiol. 44:957-970. 824
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

33
56. Willshaw, G. A., H. R. Smith, S. M. Scotland, A. M. Field, and B. Rowe. 1987. 825
Heterogeneity of Escherichia coli phages encoding Vero cytotoxins: comparison of 826
cloned sequences determining VT1 and VT2 and development of specific gene probes. 827
J. Gen. Microbiol. 133:1309-1317. 828
829
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

34
Figure legends 830
831
Fig. 1. Restriction endonuclease digestion of Stx bacteriophage DNA. Restriction nuclease 832
digested bacteriophage DNA separated on 0.7% agarose digested with EcoNI (lanes 1-5), 833
BsrBI (lanes 6-10) and PvuI (lanes 11-15). Lanes: 1, 6, 11 phage P8983; lanes 2, 7, 12 phage 834
P13344; lanes 3, 8, 13 phage P13374; lanes 4, 9, 14 phage P13771 and lanes 5, 10, 15 phage 835
P13772. M = molecular weight standard (range 10-0.2 kb) 836
837
Fig. 2. Stx production of STEC O104:H4 wildtype strains and C600 phage lysogens. 838
Extinction values (OD450) obtained by measuring serial dilutions of the six STEC O104:H4 839
wildtype strains (Table 1) and five E. coli K-12 lysogens (Table 3) by the Stx-EIA. (A) , 840
STEC wildtype strains from Table 1 grown in the presence of mitomycin C; , STEC 841
wildtype strains grown without mitomycin C induction. (B) , C600 Stx phage lysogens 842
grown in the presence of mitomycin C, , C600 Stx phage lysogens grown without 843
mitomycin C. 844
845
Fig. 3. TEM-micrographs of Stx phages from STEC O104:H4 strains. Negatively stained 846
phages with a short tail (arrowheads) and hexagonal head isolated from E. coli K-12 (C600) 847
after infection with phage lysates of (A) strain CB8983, (B) strain CB13344, (C) strain 848
CB13771, and (D) strain CB13374 (bars = 50 nm). 849
850
Fig. 4. TEM micrographs of P13374 induction from lysogenic strain TPE2364. Ultrathin 851
sections of two bacterial cells (TPE2364) with maturating virion particles within the 852
cytoplasm indicated by arrows (bars = 500 nm). The cells showing signs of bacteriolysis (loss 853
of cytoplasm (A) and damaged cell wall as well as the formation of vesicles or opaque clumps 854
(B). (C) TEM micrograph of CsCl purified, negatively stained P13374 particles released by 855
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

35
strain TPE2364 (bar = 100 nm). Four phages with a short tail (arrows) and a hexagonal head. 856
In the upper part two phages with tail to tail interactions are shown. 857
858
Fig. 5. TEM-micrographs of myovirus phages isolated from the Norwegian STEC 859
O104:H4 strain CB13437. Negatively stained phages comprise a hexagonal head, contractile 860
tail and kinked tail fibers (Myoviridae). The phages were propagated on E. coli K-12 strain 861
C600 using phage lysates of the Norwegian strain CB13437. (A) Myovirus with non-862
contracted tail, (B) Myovirus with contracted tail (bars = 100 nm). 863
864
Fig. 6. Genome organization of prophage P13374. Putative genes are colored according to 865
the predicted functions of their products. The positions of putative Rho-independent 866
transcription terminators and transfer RNAs (tRNAs) are indicated. Further detailed 867
informations to the structural components of P13374 are given in the Supplemental material 868
Table S3. P13374 virion proteins which were identified by SDS-PAGE and mass 869
spectrometry are marked by asterisks (*). 870
871
Fig. 7. Relationship of P13374 to Stx phages and phage Lambda. The circular synthetic 872
plot shows the P13374 genome organization and the similarity of predicted gene products to 873
proteins of related Stx phages. The two outermost circles represent the + and – strand of 874
P13374 genome, respectively (colouring according to Fig. 6). Inward circles: phage TL-2011c 875
(accession JQ011318), phage VT2phi272 (HQ424691), phage 933W (AF125520), Phage 876
VT1-Sakai (AP000400), phage VT2-Sakai (AP000422), Stx1-converting phage (AP005153), 877
Stx2-converting phage (AP0044202), phage 2851 (FM180578) and Lambda (J02459). 878
879
Fig. 8. Integration of Stx phage P13374 into E. coli K-12 C600. (A) Integration of phage 880
P13374 into the wrbA gene of E. coli C600. Selected genes and the attachment site of the 881
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

36
bacteria (attB) and the phage (attP) are indicated. (B) Sequences flanking the attachment sites 882
in E. coli strain C600, phage P13374, and the left (attL) and right (attR) core sequences in 883
lysogenic C600 strains. wrbA* indicates a putative modified wrbA gene. 884
885
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

37
Table 1. STEC O104:H4 strains used for isolation of lysogenic Stx-encoding bacteriophages
. presence of virulence genes
Strain
no.
original
designation
source, origin,
year of isolation, disease
stx2 aggR wzxO104 aggA agg3A astA
CB13344 RKI 11-02027 a human, Germany, 2011, HUS + + + + - -
CB13374 - sprouted seeds, Germany. 2011 + + + + - -
CB8983 - human, Germany, 2001, HC + + + - + +
CB13437 1106-3060-1 b human, Norway, 2006, diarrhea + + + - + +
CB13771 2009EL-2050 c human, Georgia, 2009, HC + + + + - -
CB13372 2009EL-2071 c human, Georgia, 2009, HC + + + + - -
a) provided by the Robert Koch-Institute, Berlin, Germany,
b) from the collection of the Norwegian Institute of Public Health,
c) from the collection of the Centers for Disease Control and Prevention (CDC), USA
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
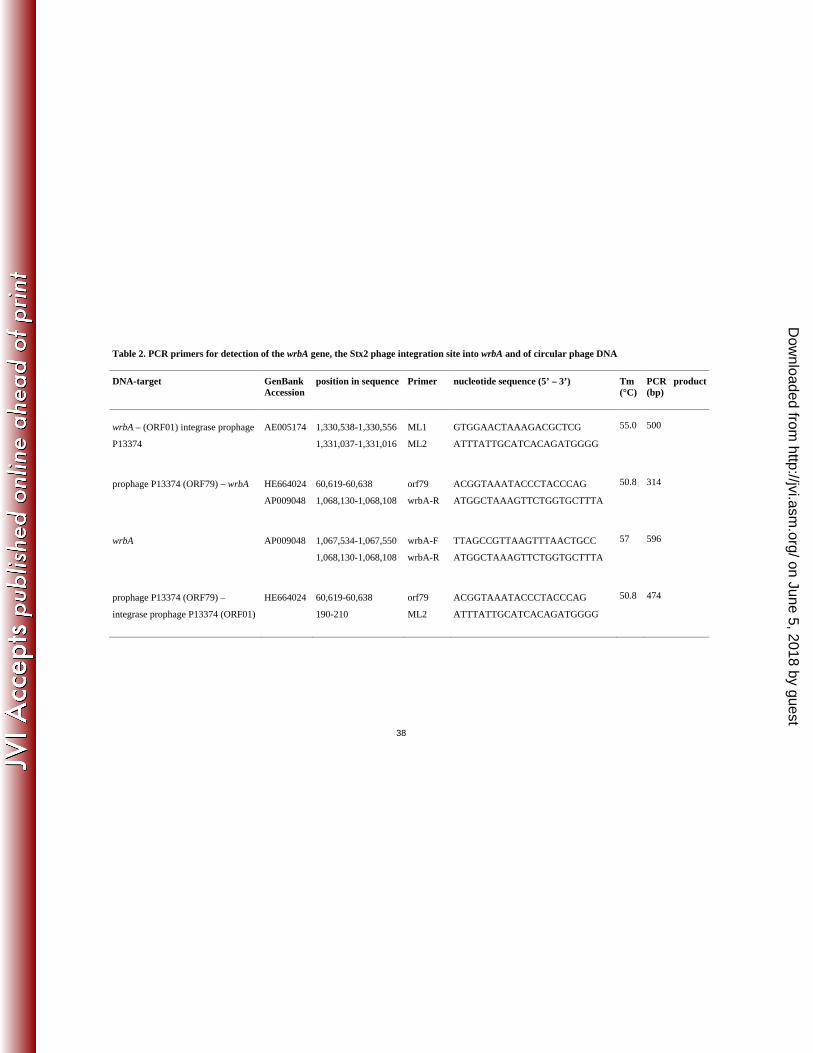
38
Table 2. PCR primers for detection of the wrbA gene, the Stx2 phage integration site into wrbA and of circular phage DNA
DNA-target GenBank Accession
position in sequence Primer nucleotide sequence (5’ – 3’) Tm (°C)
PCR product (bp)
wrbA – (ORF01) integrase prophage
P13374
AE005174 1,330,538-1,330,556
1,331,037-1,331,016
ML1
ML2
GTGGAACTAAAGACGCTCG
ATTTATTGCATCACAGATGGGG
55.0 500
prophage P13374 (ORF79) – wrbA HE664024
AP009048
60,619-60,638
1,068,130-1,068,108
orf79
wrbA-R
ACGGTAAATACCCTACCCAG
ATGGCTAAAGTTCTGGTGCTTTA
50.8 314
wrbA AP009048 1,067,534-1,067,550
1,068,130-1,068,108
wrbA-F
wrbA-R
TTAGCCGTTAAGTTTAACTGCC
ATGGCTAAAGTTCTGGTGCTTTA
57 596
prophage P13374 (ORF79) –
integrase prophage P13374 (ORF01)
HE664024 60,619-60,638
190-210
orf79
ML2
ACGGTAAATACCCTACCCAG
ATTTATTGCATCACAGATGGGG
50.8 474
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

39
Table 3. E. coli K-12 (C600) Stx phage lysogenic derivative strains
sensitivity to
strain lysogenic
for phage
phage source strain
(reference)
Shiga toxin P8983,
P13344,
P13374
P13371,
P13372
C600 - (49) - + +
TPE2385 P13344 CB13344 (53) Stx2a - -
TPE2364 P13374 CB13374 (53) Stx2a - -
TPE2369 P8983 CB8983 (this work) Stx2a - -
TPE2381 P13771 CB13771 (this work) Stx2a + -
TPE2383 P13772 CB13772 (this work) Stx2a + -
TPE1923 P933w EDL933 (49) Stx2a + +
TPE1914 PH19-B H19 (50) Stx1 + +
TPE1916 Lambda - - + +
TPE1841 P6220 CB6220 (27) Stx1c + +
TPE1874 CB2851 CB2851 (49) Stx2c + +
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

40
Table 4. Peptide identification by peptide mass fingerprinting (PMF) and peptide fragment fingerprinting (PFF) using tandem MALDI-
TOF mass spectrometry (MS/MS) and MASCOT database searching on tryptic digests of P13374 proteins
ORF
P13374
nucleotide
position
calculated
MW (kDa)
estimated MW
(kDa)a
peptides
found
Genbank
accession (GI)
Function, name
58 34,002 -
36,146
81.030 70-90 11 (PMF) 340730675 putative portal protein [Escherichia
coli O104:H4 str. 01-09591]
60 37,335 –
38,549
44.877 46-50 28 (PMF)
5 (PFF)
9632520
hypothetical protein 933Wp54
[Enterobacteria phage 933W]
61 38,605 –
38,994
13.496 <14 7 (PMF)
2 (PFF)
9632521 hypothetical protein 933Wp55
[Enterobacteria phage 933W]
78 51,952 –
60,372
309.315 >250 3 (PFF) 9632538
hypothetical protein 933Wp72
[Enterobacteria phage 933W]
a) from SDS-PAGE
on June 5, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from







