spatial variability of the structure of the harpacticoida (copepoda) crustacean assemblages in...
TRANSCRIPT

51
ISSN 0001-4370, Oceanology, 2007, Vol. 47, No. 1, pp. 51–59. © Pleiades Publishing, Inc., 2007.Original Russian Text © E.S. Chertoprud, M.V. Chertoprud, L.A. Garlitskaya, A.I. Azovsky, D.V. Kondar’, 2007, published in Okeanologiya, 2007, Vol. 47, No. 1, pp. 59–68.
INTRODUCTION
Despite the low taxonomic ranking, representativesof the copepod crustaceans of the Harpacticoida ordersignificantly differ both in their morphology and intheir modes of life. The diversity of the life forms ofmarine harpacticoids allow them to dwell over a widespectrum of habitats from mud and phytal zones torocky pools and pieces of decaying timber. In so doing,many species are strictly confined to certain habitats,which results in the high biotopic variability of theirassemblages [11, 35]. As to the geographical variabilityof the harpacticoid fauna, it is suggested that the spe-cies composition of these highly mobile and rapidlyspreading copepods features only minor changes fromone region to another [35, 37]. This suggestion is justi-fied by the fact that a significant part of the knownmarine species are cosmopolitan and only a few of thespecies feature relatively restricted habitats [27, 34, 40,44]. Meanwhile, to date, this opinion has received noreliable confirmation. In this paper, we consider thespatial variability in the harpacticoid assemblages oversoft sediments in shallow-water and intertidal zones ofthe seas off Europe.
There are rather abundant data about the shelf har-pacticoids of different regions; meanwhile, their studiesfeatured a patchy character. In particular, in WesternEurope (Fig. 1), almost all the harpacticoid surveyswere carried out off the coasts of four countries: Ger-many, Italy, France, and Great Britain. Because of thefragmentation of the different surveys, it is not surpris-ing that only rare publications are devoted to comparing
lists of harpacticoids from the different regions and tothe general biogeography of the group. Meanwhile,when the assemblages are compared on an interregionalscale, it occurs that the similarity and differences
MARINEBIOLOGY
Spatial Variability of the Structure of the Harpacticoida (Copepoda) Crustacean Assemblages in Intertidal and Shallow-
Water Zones of European Seas
E. S. Chertoprud
a
, M. V. Chertoprud
a
, L. A. Garlitskaya
b
, A. I. Azovsky
a
, and D. V. Kondar’
c
a
Faculty of Biology, Moscow State University, Moscow, Russia
b
Odessa Branch, Kovalevskii Institute of Biology of the Southern Seas, National Academy of Sciences of Ukraine,Odessa, Ukraine
c
Shirshov Institute of Oceanology, Russian Academy of Sciences, Moscow, Russia
Received February 14, 2006; in final form, March 29, 2006
Abstract
—On the basis of original and published data for 26 sites off Europe (the Barents, White, Black, Cas-pian, North, Baltic, Mediterranean, and Adriatic seas and the Atlantic Ocean) we assessed the variability of thenear-shore benthic harpacticoid assemblages. A set of six life forms is recognized that retains its compositionregardless of the geographical location and species composition of the assemblage. It is shown that the geo-graphical variability of harpacticoid assemblages is low and intraregional and biotopical variations in the struc-ture of these assemblages are often greater than their interregional (geographical) variability. It seems that themain factors that control the composition of species and life forms of harpacticoids are the characteristics of thesediments and the hydrodynamical processes that determine the structure of the sediments and the salinity.
DOI:
10.1134/S0001437007010080
2 22
2
2
2
2
2 22
2
21
211
122
22
2
2
Fig. 1.
Location of the stations used:
1
—author’s surveys;
2
–published materials.

52
OCEANOLOGY
Vol. 47
No. 1
2007
CHERTOPRUD et al.
between the assemblages feature no dependence on thedistance between them. The harpacticoid faunas fromadjacent bights may differ from one another as stronglyas from the faunas from another sea [22, 24]. This kindof seemingly irregular mosaic pattern is caused by thefact that, when comparing the regions, one can hardlyseparate the geographical component of the variabilityfrom the intraregional and biotopic components. Thestrong differences between the harpacticoid assem-blages from different habitats are emphasized by theexistence of several groups of species in the same typeof biotope. These variations may be caused not only byenvironmental factors but also (to a greater extent) bybiotic interactions; alternatively, they may have a prob-abilistic character [11]. Thus, there emerges the prob-lem of the proportions of individual variability compo-nents in the total spatial heterogeneity of the fauna dis-tribution.
In this paper, for the first time, we performed a com-parison between the bottom harpacticoid communitiesfrom the seas washing Europe in the north, west, andsouth. The objective of our study was to compare theinterregional (geographical) and intraregional variabil-ities of the structure of the harpacticoid assemblages. Inaddition, we tried to recognize the principal factors thatdefine the diversity of the assemblages of this group onthe scale of the European continent.
MATERIALS AND METHODS
We analyzed the spatial variability of the near-shoreharpacticoid fauna using the original and publisheddata collected at 26 sites in the Barents, White, Black,Caspian, North, Baltic, Mediterranean, and Adriaticseas and in the Atlantic Ocean (Fig. 1). At 5 sites, weused the materials collected by the authors; they arelocated in the White (1 site), Black, and Caspian seas.For 21 sites, published data were used: the White(2 sites, one of which coincides with the site of ourstudies), Barents (1), North (2), Baltic (4), Black (1),Mediterranean (5), and Adriatic (2) seas and the Atlan-tic Ocean (5). The distances between individual sitesare no less than 60–70 km.
The selection of published data for our analysis wasperformed with respect to three criteria. First, the pub-lication should contain a detailed list of the harpacti-coid species encountered. A series of papers also con-tained data on the species abundances; however, thesekinds of publications were rare. Second, the studyshould consider a limited area of the intertidal or uppersubtidal zone (not greater than 50 km in size). Third, thestudies should be based on representative materials. Inthe publications selected for our purposes, the samplingeffort per unit length of the shore usually comprised20
−
30 cm
2
per 1 km. Unfortunately, most of the studiesused were missing detailed descriptions of the biotopesin which the materials were collected. Therefore, it wasfeasible to perform an analysis of the biotopic variabil-ity only for selected regions.
Original Data
Th e W h i t e S e a
. A portion of the Kareliancoast of Kandalaksha Bay in the region of the Biologi-cal Station of Moscow State University and the villageof Chernaya Rechka was studied. The principal area ofthe studies was Gryaznaya Bight located close to theestuary of the Chernaya River. Harpacticoid sampleswere collected with a tube corer from the sandy–siltyintertidal zone in six inlets different in their morphol-ogy. The area of an individual sample was 1.8 cm
2
. Inall, during the summer seasons of 1999–2004, about 760samples of meiobenthos were collected. We describedthe ecology of the intertidal harpacticoids of this part ofthe coast in a series of papers [6, 9, 10, 17, 18].
Th e B l a c k S e a
. The materials were collectedin the subtidal zone of the southwestern coast near thedelta of the Danube River (sea depth 17 m) and abeamOdessa (sea depth 24 m) as well as in the near-shorezone of Grigor’evskii Lagoon (sea depth 0.5 m). Inshallow-water areas, the samples were taken with thehelp of a handle box corer
10
×
10
cm in size; in the sub-tidal zone, a grab corer was used. The sample areasranged from 78 to 100 cm
2
. In all, in 2003–2005, about30 samples of meiobenthos were collected. The harpac-ticoid fauna of Grigor’evskii and adjacent lagoons wasreported in [31].
Th e C a s p i a n S e a
. In 2003, a portion of thecoast of Bol’shoi Gyzyl Agaj Bay (Southeast Azer-baijan) was surveyed [25]. The samples were takenwith a 1.8 cm
2
corer from shallow-water areas (seadepth 0.1 m) and the supralittoral zone in three minorinlets. In all, about 20 samples of meiobenthos werecollected. These not too abundant materials are interest-ing because they represent the harpacticoid fauna of aportion of the Caspian coast rather than summarize iso-lated findings made in different years over the entirearea of the sea.
All the samples collected were washed through amill sieve with a mesh size of 70
µ
m. In the White andCaspian seas, only adult individuals were examined,while, in the Black Sea, copepodites were alsoincluded. No nauplius stages were taken into account.The harpacticoid individuals were mostly identified tothe species level; in selected cases, when the individu-als were deformed, the identification was restricted tothe genus level.
Published Data
The information about the harpacticoids of the siteslisted below was taken from publications: the north-western part of the White Sea (Kislaya Bay [13] and offCape Kartesh [12] on the Karelian coast of Kandalak-sha Bay); the southern coast of the Barents Sea (south-eastern inlet in Dal’nezenetskaya Bay [1]); the south-western part of the North Sea (the region of the NorthFrisian Islands [16, 29], a portion of the western coastof Belgium [33], the region off Southampton on the
OCEANOLOGY
Vol. 47
No. 1
2007
SPATIAL VARIABILITY OF THE STRUCTURE 53
southern coast of Great Britain in the English Channel[19]); the eastern Baltic Sea (Kattegat Strait, two sitesoff the coast of Germany west and east of Rostock, thearea near Borne Island off the coast of Sweden [14,15]); the East Atlantic (Galway Bay in the west of Ire-land, the north of the Brittany coast of France [22],areas off Plymouth in the southwest of England [32],and Arcachon Bay in the southwest of France [23]); theBlack Sea (southern termination of the Crimean Penin-sula, Donuzlav Lagoon [7]); the southern coasts of theAdriatic and Mediterranean seas (a series of lagoonsnear Venice, the Lesina lagoons in the southeast of Italy,Porto Pino Bay in the south of Sardinia Island [24],Smarlacca Lagoon in the southeast of Italy [28], bays inthe region of Salambo Biological Station on the north-ern coast of Tunisia [38], a series of lagoons nearMarseille [21], and a portion of the coast of Catalonia[41]).
In the species list reported, some species had oftenbeen included that invaded the benthic samples fromplankton or macrophytes. For example, two planktonicgenera—
Microsetella
(Ectinosomatidae family) and
Euterpina
(Euterpinidae family)—were noted. Amongthe phytal forms, most often were representatives of theTisbidae, Thalestridae, Porcellidiidae, Peltidiidae, andTegastidae families. The occurrence frequency and thediversity of these species in different regions stronglyvaried; they undoubtedly depended on the position ofthe phytal zone, which was not reported in most of thepapers. This made us exclude all the phytal speciesfrom our analysis.
Statistical Analysis of the Data
We compared the data from different regions usingtwo parameters: the Sörensen index of faunistic simi-larity (
D
) and the Czekanowski similarity index (
Cz
)applied to the quantitative data represented by the pro-portions calculated with respect to the abundances orspecies numbers (in order to compare the diversitywithin individual life forms). The intraregional varia-tions in the taxocene structure were estimated as theaverage pairwise similarity value between the sitesfrom the same sea. The interregional variations werecalculated as the mean pairwise similarity between thesites for each of the seas. The confidence level of thedifferences between the sets indices of intra- and inter-regional variabilities were determined using the Mann–Whitney nonparametric test.
In addition, in order to separate the interregionalcomponent of the variability, we applied the ANOSIMprocedure for similarity analysis [26]. This procedureimplies ranging of all the values of paired similaritybetween the stations; then, the value
R
= (
R
in
−
R
ex)/
n
is calculated, where
R
in is the mean similarity rankbetween the stations of the same region,
R
ex is themean similarity rank between the stations of differentregions, and
n
is the total number of similarity indices.The confidence level of the
R
-statistic inequality to zero
was estimated as the probability of obtaining the sameor greater values of differences at random distributionof the stations over the regions.
In parallel, when analyzing the spatial heterogeneityof the distribution of the group, we plotted the depen-dences of the taxonomic similarity between the regionson the distance between them over the latitude and thedependences of the species richness on the number ofstations considered. In order to estimate the speciesrichness, we used the Margalef index
(
I
):
I
= (
W
–1)/
, where
W
is the number of species and
N
isthe number of organisms assessed.
For the calculations, we used the PRIMER
5(PRIMER-E Ltd., [26])
and ECOS (Moscow State Uni-versity software packages).
RESULTS AND DISCUSSION
In the course of our analysis of the variability anddiversity of the harpacticoid taxocens, we estimated thevariations of their principal characteristics on the intra-and interregional scales. The variability of the speciescomposition and the sets of life forms (the functionalstructure and the taxonomic composition of the group)were considered in detail.
Species Richness
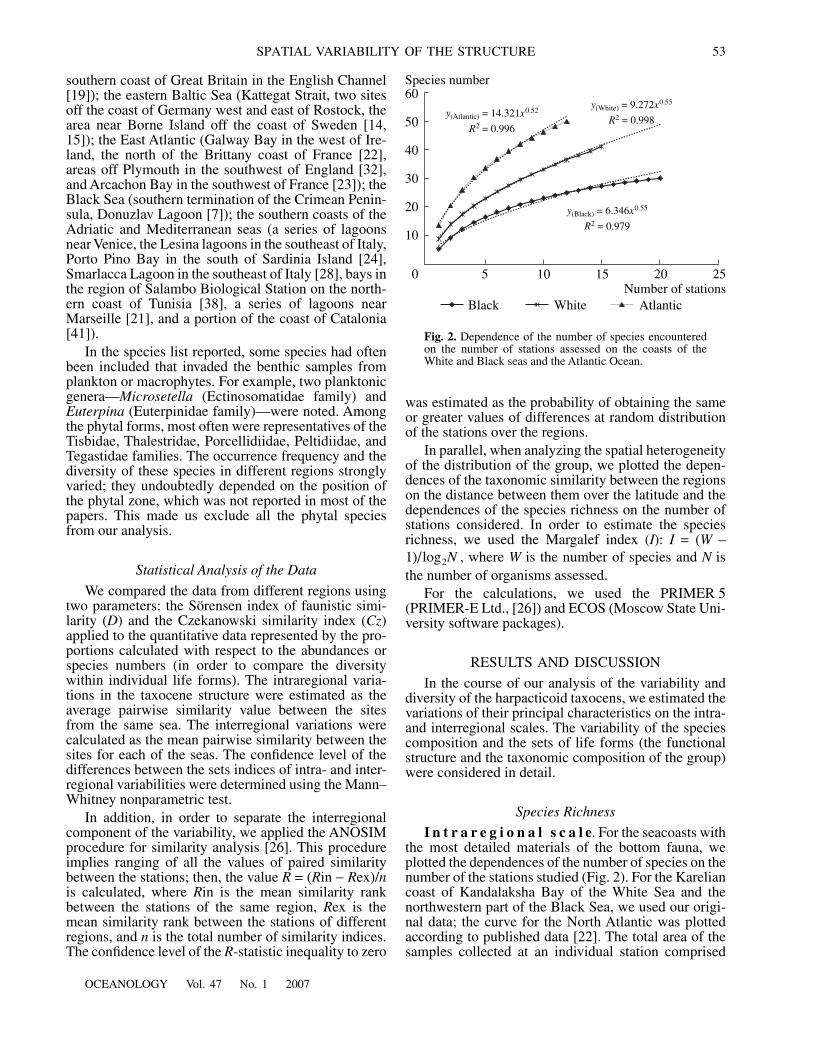
I n t r a r e g i o n a l s c a l e
. For the seacoasts withthe most detailed materials of the bottom fauna, weplotted the dependences of the number of species on thenumber of the stations studied (Fig. 2). For the Kareliancoast of Kandalaksha Bay of the White Sea and thenorthwestern part of the Black Sea, we used our origi-nal data; the curve for the North Atlantic was plottedaccording to published data [22]. The total area of thesamples collected at an individual station comprised
log N2
Species number
60
50
40
30
20
10
0 5 10 15 20 25
Black White AtlanticNumber of stations
y
(Atlantic)
= 14.321
x
0.52
R
2
= 0.996
y
(Black)
= 6.346
x
0.55
R
2
= 0.979
y
(White)
= 9.272
x
0.55
R
2
= 0.998
Fig. 2.
Dependence of the number of species encounteredon the number of stations assessed on the coasts of theWhite and Black seas and the Atlantic Ocean.

54
OCEANOLOGY
Vol. 47
No. 1
2007
CHERTOPRUD et al.
100, 30, and 10 cm
2
for the Black Sea, the AtlanticOcean, and the White Sea, respectively. In each of thesethree cases, the sampling network covered a part of thecoast from 20 km (Atlantic) to 50–70 km (Black andWhite seas).
In all the regions examined, the number of speciesregularly grows with the increase in the number of sta-tions considered. In so doing, in the Black Sea, despitethe greatest volume of the samples, the mean number ofspecies per station is significantly lower than in theWhite Sea and the Atlantic (6.3, 9.2, and 14.3, respec-tively). The low local species richness of harpacticoidsin the Black Sea is probably caused by the local biotopefeatures and, probably, by the features of sampling andsample processing [44]. The species richness estimatedby the Margalef index equals
4.51
±
2.45, 3.78
±
1.28
,and
2.07
±
1.54
in the Atlantic, the Black Sea, and theWhite Sea, respectively. It is characteristic that, in theBlack Sea, which represents a relatively young lowsaline marine basin, low species diversities were notedfor many other marine taxa [4].
The following fact is of even greater interest: thegrowth rates of the species’ richness depending on thenumber of stations are close in all three of the seasassessed above. This is suggested by the small range ofvariations (0.52–0.55) of the exponent values in thespecies–stations relations for all of the regions (Fig. 2).Thus, the inhomogeneity of the distributions of the tax-onomic composition and, probably, the harpacticoiddiversity in different areas assessed are virtually thesame, being rather high. For example, a twofoldincrease in the number of stations causes a growth inthe species number by a factor of 1.5. The species listseven from neighboring inlets significantly differ fromone another and, as was shown by qualitative data, maybe dominated by absolutely different groups of species.For example, at the stations of the Brittany coast of theAtlantic Ocean, over similar sediments, the position ofthe first dominant is subsequently occupied by the spe-cies of the
Canuella, Harpacticus, Amphiascoides
, and
Asellopsis
genera. A sharp change in the assemblagesof intertidal species over the same type of sand wasdescribed in detail in [11].
I
n t e r r e g i o n a l s c a l e
. When comparing thespecies richness in different seas, all the data availablewere subdivided into two groups. The first groupincluded the materials for portions of the coast less than5 km long (individual lagoons and bights); the secondgroup contained the materials on the harpacticoid faunafor the coastal sections from 20 to 50 km long. Whenanalyzing selected published data, we faced the prob-lem related to the strong variations in the data volumesat different stations. Meanwhile, usually, the smallnumber of samples at a point was compensated for bytheir large size, while the small area of the samples wasaccompanied by their great number. The characteristictotal area of all the samples assessed for an individualbight comprised 90–120 cm
2
and reached 1000 cm
2
for
a lengthy portion of the coast. In so doing, the nest-typesample distribution used in selected studies [21, 22, 24,41] allowed one to cover significant spatial areas usinga small volume of the materials.
The highest harpacticoid diversity was noted in theintertidal zones of the Subarctic White and Barentsseas: the average values were 38.5 species per individ-ual bight and 45.5 species over a lengthy portion of thecoast. The Atlantic and the North Sea are second withrespect to the species richness (16 and 43.8 species,respectively). The third richest was the MediterraneanSea with the corresponding values of 11 and 32.5 spe-cies. The lowest species diversity values were observedin the Caspian, Black, and Baltic sea (about 10.3 spe-cies in a bight and only 18.4 species over the coast).
The decrease in the species richness of harpacti-coids (as well as many other groups of marine organ-isms) in desalinated (
10–18
‰) basins of the Caspian,Baltic, and Black seas was mentioned in a series ofreviews [2, 35, 37]. The decrease in the abundance andspecies richness of benthic copepods in brackish waterswas also shown for selected estuarine systems [8, 39,43]. The lower (as compared to the northern regions)diversity of harpacticoids in shallow-water areas ofsubtropical coasts is probably related to the tempera-ture factor. For example, in lagoons of the Black andMediterranean seas, the summer water temperaturereaches
25–28°ë
, which reduces the oxygen solubilityin the water down to a critical level [35]. In shallow-water areas, even the period of the harpacticoid repro-duction is shifted to colder seasons of the year [3, 24].Meanwhile, selected species dwelling in the subtidalzone of the Mediterranean Sea under a more constantand lower temperature reproduce all year round [37].
The relative poorness of the local coastal faunas ofthe temperate belt as compared to the Subarctic regionsmay be related to the following reasons. On the onehand, the considered shallow-water areas of the NorthSea and the Atlantic are more polluted than those of theWhite or Barents seas because of the higher economicactivity on the coasts. A significant part of the studies inthe Atlantic Ocean and the North Sea used here [22, 32,33] were performed immediately after the major cata-strophic accident of the
Amoco Cadiz
tanker in theregion of the English Channel (March 1978). On theother hand, in the temperate zone, the intertidal mac-robenthos is far more abundant and rich in species thanin the freezing and damaged by floating ice Subarcticintertidal zone [5]. It is possible that the decrease in theabundance and richness of the macrobenthos in the lat-ter regions has a positive effect on the meiobenthosdiversity due to the decrease in the tension over trophiclinks.
On the whole, the results on the interregional vari-ability of the species richness of harpacticoids are pre-liminary and require further tests using additionalmaterials.

OCEANOLOGY
Vol. 47
No. 1
2007
SPATIAL VARIABILITY OF THE STRUCTURE 55
Life Forms and Functional Structure
S p e c t r u m o f l i f e f o r m s
. The general listof species for the regions considered by us contains 254harpacticoid species belonging to 25 families. We sub-divided these species into six life forms with respect totheir morphology and dwelling conditions. Theirdetailed description is presented in our paper devoted tothe diversity of harpacticoid taxocens in the intertidalzone of the White Sea [11]. The descriptions of the lifeforms presented below are complemented and includean extended list of taxa. We consider only the taxa onthe genus level and assume that the species of the samegenus of harpacticoids, as a rule, refer to the same lifeform, though representatives of selected genera (forexample,
Schisopera
and
Laophonte
) are rather diversein their morphology.
E p i b e n t h i c s p e c i e s
. (1) Specialized inter-stitial species. These are relatively large copepods withwell-developed limbs supplied with long bristles. As arule, their bodies feature cylindrical or peachlikeshapes. In addition, the bodies of the organisms dwell-ing over mud surfaces have flat outgrowths and pricklesthat prevent them from sinking into the sediment [36].In our materials, this form is widely represented andincludes 50 genera from 14 families. A brief list of themost common taxa contains the following genera:
Can-uella
(Canuellidae family);
Microarthridion
and
Tachidius
(Tachidiidae family);
Harpacticus
(Harpacti-cidae family);
Schizopera, Stenhelia
, and
Robertsonia
(Diosaccidae family);
Enhydrosoma
and
Cletodes
(Cletodidae family); and
Asellopsis, Heterolaophonte,Laophonte, Platychelipus, Pseudonychocamptus
, and
Paronychocamptus
(Laophontidae family).(2) Sand-burrowing species. This form is repre-
sented in three families: the
Itunella
(Canthocamptidaefamily), the
Huntemannia
and
Nannopus genera (Hunt-emanniidae family), and the Rhizothrix and Tryphoemagenera (Rhizothricidae family). The organisms of thesespecies usually have cylindrical bodies; when extractedfrom the sediment, many crustaceans bend in an arc-shaped mode (Huntemannia, Tryphoema). The limbs ofthese copepods are shortened; a part of their bristles arereduced or transformed into thick prickles. This helpsthem to draw apart sediment particles and dig theirchannels.
(3) Mud-burrowing species. The organisms of thesespecies usually have spin-shaped bodies, which helpstheir sliding inside the mud. The swimming limbs arelong and well equipped. In our materials, they aremostly represented by the species of the Ectinosoma-tidae family: the Bradya, Pseudobradya, Ectinosoma,and Halectinosoma genera and by the Brianola genus(Canuellidae family).
I n t e r s t i t i a l s p e c i e s. (1) Specializedepibenthic species. These harpacticoids are mainlysmall organisms with an average body length of 400µm. Their limbs, furcal branches, and the bristleslocated on them are often reduced. Their body shapes
are of two types: worm-shaped bodies (Cyllindropsil-lidae, Leptastacidae, Leptopontiidae, Ectinosomatidae,and Darcythompsonidae families), and flattened dors-oventral lanceolate bodies (Paramesochridae family).This life form is represented by 19 genera: Darcyth-ompsonia and Leptocaris (Darcythompsonidae fam-ily); Arenosetella and Hastigerella (Ectinosomatidaefamily); Evansula, Cylindropsyllus, and Stenocaris(Cyllindropsillidae family); Arenocaris, Leptastacus,Paraleptastacus, and Psammastacus (Leptastacidaefamily); Arenopontia and Leptopontia (Leptopontiidaefamily); and, finally, Apodopsyllus, Kliopsyllus, Lep-topsyllus, Paramesochra, Remanea, and Scottopsyllus(Paramesochridae family).
(2) Nonspecific interstitial species. The organismsof these species have elongated cylindrical bodiessimilar to those of the epibenthic representatives.Their principal adaptation mechanism to interstitialdwelling is related to their small sizes (on average,about 400 µm). This form includes 23 genera belong-ing to 5 families: Parastenhelia (Parastenheliidaefamily); Amphiascus, Amphiascoides, Amphiascopsis,Bulbamphiascus, Paramphiascopsis, Pseudamphias-copsis, Rhyncholagena, Teissierella, and Typhlamph-iascus (Diosaccidae family); Mesochra, Nannomeso-chra, and Psammocamptus (Canthocamptidae fam-ily); Ameira, Ameiropsis, Interleptomesochra,Leptomesochra, Nitocra, Parapseudoleptomesochra,Pseudoleptomesochrella, Sarsameira, Proameira, andPsyllocamptus (Ameiridae family); and Orthopsyllus(Orthopsyllidae family).
This subdivision with respect to life forms is notabsolute, and the boundaries between individual formsmay be rather fuzzy. Nevertheless, we tried to compilethe most detailed classification scheme insensible tominor variations in the taxocen structures.
F u n c t i o n a l s t r u c t u r e. The geographicalvariability of the functional structures of harpacticoidassemblages is low. For example, the set of life formsrecognized in the intertidal zone of the White Sea [11]is completely applicable to the taxocens off the coastsof Europe. The life-mode spectra in different seas aresimilar to a great extent (the mean value of the Sörensenindex of similarity was 0.80 ± 0.12) (see table). On theintraregional scale (in different areas of the same sea),these spectra vary slightly greater (with a mean similar-ity of 0.72 ± 0.11). The life-mode spectra of harpacti-coids established for an individual region may beextended over the coasts of entire continents.
It is characteristic that the species diversity withinindividual forms is rather stable both over local shal-low-water areas and in different seas. The ANOSIMprocedure showed that the influence of the geographi-cal variability on the proportions of the species of dif-ferent life forms in the assemblages is statistically reli-able, though very weak. The R-statistic value is as smallas 0.224, while the probability of the occasional gainingof this value is extremely low (a confidence level o 1.5%).

56
OCEANOLOGY Vol. 47 No. 1 2007
CHERTOPRUD et al.
The similarity between the seas with respect to the lifeform spectra estimated using the Czekanowski indexequals 0.82 ± 0.08, while that between the areas of aselected sea is somewhat lower being 0.73 ± 0.15. Usu-ally, specialized epibenthic and nonspecialized intersti-tial species are the most diverse involving, on the aver-age, 49 and 23% of the total species richness, respec-tively. The third diverse are mud-burrowing andspecialized interstitial forms (10 and 11% of the totalspecies richness, respectively). Meanwhile, the varia-tions of the diversity within the life forms between indi-vidual biotopes are significantly greater than those onthe regional scale and the similarity is as low as 0.46 ±0.16 (the data on the White and Mediterranean seas andthe Atlantic Ocean). The proportions between the abun-dances of species with different specializations regu-larly change depending on the type of the sediment. Forexample, in the Mediterranean Sea, the abundances ofinterstitial species on coarse-grained washed sands andon silty grounds comprise 42 and not greater than 20%of the total abundance, respectively. In the White Sea[11] and in the Atlantic Ocean, interstitial forms domi-nate over washed medium-grained and all types ofcoarse-grained sands (63–73% of the total abundance),while, on silted fine- and medium-grained sediments,their proportion is negligibly small (0.03–4%).
Thus, on the intraregional scale (between individualparts of the coast and local biotopes), the differences inthe spectra of life forms are greater than on the interre-gional scale (between the entire seas). This implies thatthe functional structure of the harpacticoid taxocens isdetermined, first, by the environmental factors that
operate within an individual bight rather than by thosecharacteristic of the entire sea or climatic zone. The sig-nificant differences in the water temperatures, salinityvalues, and pollution between the seacoasts comparedsuggest that, under the influence of these factors, the setof the life forms of harpacticoids almost do not change.According to our data [6] and to the opinion of someother scientists [20, 30, 35, 42], the spatial distributionof the life forms of harpacticoids is mostly controlledby the characteristics of the sediments and by thehydrodynamical processes, which determine the struc-ture of the sediments. For example, interstitial speciesdwell over coarse-grained washed sediments, whileepibenthic species prefer more fine-grained silted sub-strates rich in organic matter and mud-burrowing spe-cies are most abundant on the sediments rich in silty–clayey matter [6, 12, 35].
Taxonomic Composition and Domination Structure
In order to estimate the similarity between the har-pacticoid assemblages from the above-described sea-coasts, we compared their compositions on differenttaxonomic levels—species, genus, and family—usingthe Sörensen similarity index. It was noted that, whilecomparing different seas or different regions of thesame sea, the similarity of the species composition islow; that of the genus composition is noticeably higher.The highest similarity of the species is in the familycomposition (see table). It is characteristic that theintraregional variability of the genus and family com-positions is confidently higher than their interregionalvariability. The ANOSIM method yields similar results(for genera, the R-statistic equals 0.42 at a confidencelevel of 0.1%, while that for families is 0.22 at a confi-dence level of 1%). In so doing, the variations of thespecies composition on both the interregional andintraregional scales are close and almost equal. TheR-statistic equals 0.56 (at a confidence level of 0.1%),which also confirms the significant geographical vari-ability of the fauna.
In parallel, we estimated the variability of the har-pacticoid fauna with respect to the differences in thegeographical latitudes of the regions compared (Fig. 3).The similarity between the faunas regularly thoughweakly decrease with the increase in the distancebetween the stations. Again, the variability of the spe-cies composition was the highest; that of the genuscomposition was somewhat lower, while the familycomposition was the most stable. Many mass and seem-ingly flourishing species and genera were noted atalmost all the latitudes considered (approximately from38 to 70°N) For example, the Halectinosoma cur-ticorne, Stenhelia palustris, and Mesochra pigmaeaspecies, as well as the representatives of the Ectino-soma, Canuella, Microarthridion, Laophonte, andother genera, were encountered in shallow-water areasof the White, North, Baltic, Mediterranean, and Blackseas and the Atlantic Ocean. The low geographical vari-
Comparison between the taxonomic and functional composi-tions of harpacticoid taxocens on the inter- and itraregionalscales (Sörensen similarity index)
Level of comparison
Mean similarity (± mean deviation) Reliability of
the differences (Mean annual–Whitney test)within
a region between
the regions*
Species 0.23 ± 0.13 0.24 ± 0.09 0.480
Genera 0.40 ± 0.16 0.48 ± 0.10 0.072
Families 0.62 ± 0.14 0.71 ± 0.10 0.012
Life forms 0.72 ± 0.11 0.80 ± 0.12 0.025
* The Caspian Sea is excluded from the analysis because of thesmall volume of the data in this region.

OCEANOLOGY Vol. 47 No. 1 2007
SPATIAL VARIABILITY OF THE STRUCTURE 57
ability of the harpacticoid fauna has been repeatedlynoted before [35, 37]. For example, in the coastal zoneof Europe, K. Lang distinguished only two areas signif-icantly differing in the species composition, namely,the Mediterranean and boreal areas, which are sepa-rated from one another by a transition zone about athousand kilometers long.
The results of the application of the method usedshow that, on the levels higher than the species level,the interregional variability of the taxonomic composi-tion of the harpacticoid assemblages is noticeablylower than their intraregional variability. Even withinthe same region, one can observe a wide spectrum oftypes of taxonomic structure, and the extension of thescale of the analysis provides no fundamental changesin the variability level. On the scale of tens and hun-dreds of kilometers, the regional variability of the dis-tribution and species richness of harpacticoids is usu-ally related to the topography of the coastline, whichcontrols the deposition setting and grain-size composi-tion of the sediments, and to the salinity ranges [24, 45].Therefore, in our case, the lowest intraregional variabil-ity with a mean value of the Sörensen similarity indexof 0.32 ± 0.14 is confined to the Black and Mediterra-nean seas, where most of the stations are located in sim-ilar macrobiotopes (in the former case, represented bymuddy sediments of the upper subtidal zone and, in thelatter case, located in brackish-water bays). Mean-while, in the North Sea, where the stations are locatedboth in shallow-water bights and off open coasts, theintraregional variability is significantly higher (with amean similarity of 0.09 ± 0.03).
In the harpacticoid assemblages examined, the dom-ination structure changes significantly stronger thantheir taxonomic composition. In geographically neigh-boring regions, not only different species but also dif-ferent families may dominate in terms of their occur-
rence frequency and abundance. For example, when thedata on the relative abundances are used on the specieslevel, the similarity estimated by the Czekanowskiindex is 0.09 ± 0.13; on the genus level, it increases upto 0.18 ± 0.16, and, on the family level, it makes up upto 0.31 ± 0.16. The low similarity between the regionsin the domination structure may be explained by eco-logical rather than by biogeographical reasons. Thevariability of the harpacticoid abundance is caused bytheir microdistribution; it depends on the season and isstrongly related to even minor variations in the charac-teristics of the sediments and salinity [6, 35, and oth-ers]. The volume of the data used by us is insufficient tosmooth these effects.
Most of the harpacticoid assemblages assessed byus may be regarded as parallel in terms of Kusakin [5],since they are mostly composed of different but conge-neric species. Earlier, we showed that, in the intertidalbiotopes of the White Sea, the parallel character of thestructures of harpacticoid taxocens might be encoun-tered on a scale as small as a few kilometers [11]. Prob-ably, this phenomenon is rather common for the assem-blages of these organisms.
Perspectives of the Classification of Harpacticoida Taxocens
The estimation of the heterogeneity of the speciesrichness distribution of harpacticoids over the coasts ofselected European seas allows one to suggest a ratherhigh diversity of their taxocens. For example, on a por-tion of the coast of the White Sea only a few tens ofkilometers long, we distinguished six different types ofassemblages [6, 11]. Due to this reason, when analyz-ing the spatial and biogeographical variabilities of thegroup, it seems reasonable to make a preliminary clas-sification of its taxocens and to represent their diversity
0.2
0 500
Sörensen similarity index
Distance, km
Species Genera Families
1000 1500 2000 2500 3000 3500 4000
0.1
0.3
0.4
0.5
0.6
0.7
0.8
1.0
0.9
Fig. 3. Similarity between the regions in terms of species, genus, and family compositions as functions of the latitudinal distancebetween them.

58
OCEANOLOGY Vol. 47 No. 1 2007
CHERTOPRUD et al.
as a mosaic composed of elements with different simi-larity degrees.
The recognition of individual types of harpacticoidassemblages should be performed on the basis of theirrelatively stable functional structure, which is virtuallycommon for all the European coasts considered andwithin individual regions. The greatest variability of thefunctional structure is observed when comparing dif-ferent biotopes; the classification should start with theirdescription. The taxonomic composition, being a morevariable parameter, plays a subordinate role. It is char-acteristic that the higher the taxonomic rank the lowerthe geographical variations in its composition. Thisallows one to recognize groups of genera and familiescommon for all the regions and, if necessary, to distin-guish sets of species specific to selected areas. As arule, the quantitative characteristics of the harpacticoidpopulations are not important for taxocen identificationbecause of their high spatial and temporal variability.
The multilevel comparison of taxocens from differ-ent European seacoasts showed that a significant part ofthe spatial variations of their structure fall on the intrar-egional and, in selected cases, biotopic scales. This isespecially important, because it allows one to suggestthat the classification elaborated for an individualregion may be readily extended over any other region aswell.
ACKNOWLEDGMENTS
This study was supported by the Russian Founda-tion for Basic Research, projects no. 04–05–64734 andno. 06-04-48106.
REFERENCES
1. I. Ya. Agarova, M. N. Voronova, V. V. Gal’tsova, et al.,“Distribution and Ecology of the Bottom Fauna on theDal’nii Plyazh Intertidal Shoal,” in Ecological Studies ofSandy Intertidal Zones (KF AN SSSR, Apatity, 1976),pp. 95–186 [in Russian].
2. E. V. Borutskii, Fauna of the USSR. Vol. 3, Issue 4. Crus-taceans. Freshwater Harpacticoids (AN SSSR, Mos-cow, 1952) [in Russian].
3. L. V. Vorob’eva, Yu. P. Zaitsev, and I. I. Kulakova, Inter-stitial Meiofauna of Sandy Beaches of the Black Sea(Naukova Dumka, Kiev, 1992) [in Russian].
4. V. N. Eremeev and V. E. Zaika, “Introduction,” inPresent-Day Condition of the Biological Diversity of theCoastal Waters off the Crimea (Black Sea Sector)(INBYuM, Sevastopol, 2003), pp. 13–15 [in Russian].
5. O. G. Kusakin, “Intertidal Communities,” in Biologiyaokeana (Nauka, Moscow, 1977), Vol. 2, pp. 111–133 [inRussian].
6. P. V. Rybnikov, D. V. Kondar’, and A. I. Azovskii, “Prop-erties of the White Sea Littoral Sediments and TheirInfluence oon the Fauna and Distribution of Harpacti-coida,” Okeanologiya 43 (1), 97–108 (2003) [Oceanol-ogy 43 (1), 91–102 (2003)].
7. N. G. Sergeeva, “Meiobentos of the Donuzlav Lagoon,”in Present-Day Condition of the Biological Diversity ofthe Coastal Waters off the Crimea (Black Sea Sector)(INBYuM, Sevastopol, 2003), pp. 251–258 [in Russian].
8. E. S. Chertoprud, Extended Abstract of Candidate’s Dis-sertation in Biology (Moscow, 2005) [in Russian].
9. E. S. Chertoprud and A. I. Azovsky, “Seasonal Dynamicsof the Populations of Intertidal Harpacticoids (Harpacti-coida: Copepoda) in the White Sea,” Okeanologiya45 (6), 1–9 (2005) [Oceanology 46, (1), 71–80 (2006)].
10. E. S. Chertoprud, A. I. Azovsky, and F. V. Sapozhnikov,“Colonization of Azoic Sediments of Different Grain-Size Composition by Littoral Harpacticoida (Copep-oda),” Okeanologiya 45 (4), 637–646 (2005) [Oceanol-ogy 45 (5), 698–706 (2005)].
11. E. S. Chertoprud, M. V. Chertoprud, D. V. Kondar’, et al.,“Taxocen Diversity of Harpacticoids (Copepoda) in theSandy–Silty Intertidal Zone of Kandalaksha Bay of theWhite Sea,” Okeanologiya 46 (4), 1–10 (2006) [Ocean-ology 46, (4)].
12. L. L. Chislenko, “Harpacticoids of the Karelian Coast ofthe White Sea,” in Hydrobiological Studies off the Kare-lian Coast of the White Sea (Nauka, Leningrad, 1967),pp. 48–196 [in Russian].
13. E. A. Chusova, “Microdistribution of Different Stages ofMesochra lilljeborgi Boeck and Stenchlia palustrisBradi (Harpacticoida) in the Upper Layer of the SandyIntertidal Zone in Kislaya Bight of Kandalaksha Bay, theWhite Sea,” in Biology of the White Sea (Mosk. Gos.Univ., Moscow, 1980), Vol. 5, pp. 93–102 [in Russian].
14. G. Arlt, “Taxonomy and Ecology of Some Harpacticoids(Crustacea, Copepoda) in the Baltic Sea and Kattegat,”Zool. Ans. Syst. 140 (3), 45–85 (1983).
15. G. Arlt, “Temporal and Spatial Meiofauna Fluctuation inan Inlet of the South-West Baltic (Dars-Zingst BoadenChain) with Special Reference to the Harpacticoida(Copepoda, Crustacea),” Hydrobiol. 73 (3), 297–308(1988).
16. W. Armonies, “Meiofaunal Emergence from IntertidalSediment Measured in the Field: Significant Contribu-tion to Nocturnal Planktonic Biomass in ShallowWaters,” Helgolander Meeresunters 43, 29–43 (1989).
17. A. I. Azovsky, E. S. Chertoprud, and M. A. Saburova,“Variability of the Spatial and Temporal Structure of theMicrobenthic and Meiobenthic Communities of theIntertidal Zone of the White Sea,” Est. Coast. and Shelf.Sci. 60 (4), 663–671 (2004).
18. A. I. Azovsky, M. A. Saburova, E. S. Chertoprud, andI. G. Polikarpov, “Selective Feeding of Littoral Harpac-ticoids on Diatom Algae: Hungry Gourmands?,” Mar.Biol. 148, 327–337 (2005).
19. P. R. O. Barnett, “Distribution and Ecology of Harpacti-coid Copepods, of an Intertidal Mudflat,” Intern. Rev. derGesamten Hydrobiologia 53 (2), 177–209 (1968).
20. G. F. Blanchard, “1990. Overlapping Microscale Disper-sion Patterns of Meiofauna and Microphytobenthos,”Mar. Ecol.: Proc. Ser. 68 (1-2), 101–111.
21. P. Bodin, “Recherches sur la Systématique et la Distribu-tion des Copépodes Harpacticoïdes des Substratas Meu-bles des Environs de Marseille,” Rec. Trav. St. Mar. End33, 107–183 (1964).

OCEANOLOGY Vol. 47 No. 1 2007
SPATIAL VARIABILITY OF THE STRUCTURE 59
22. P. Bodin and D. F. Jakson, “A Comparison of the Inter-tidal Harpacticoid Copepod Assemblages of SandyBeaches in Galway Bay (Ireland) and Northern Brittany(France),” Mar. Biol. Ass. 69, 573–588 (1989).
23. J. Castel and P. Lassere, “Opportunistic Copepods inTemperate Lagoons of Arcachon Bay: Differential Dis-tribution and Temporal Heterogeneity,” Estuar. Coast.Mar. Sci. 9, 357–368 (1979).
24. V. U. Ceccherelli and M. Mistri, “Ecological and Zoo-geographical Study of Some Association of Harpacti-coids,” Boll. Zool. 57, 73–81 (1990).
25. E. S. Chertoprood and P. N. Kornev, “On the Harpacti-coid Fauna of the Caspian Sea, Including the Descriptionof Schizopera rybnikov sp.n. (Copepoda: Harpacticoida:Diosaccidae),” Arthropoda Selecta 14 (3) 13–23 (2005).
26. K. R. Clarke and R. M. Warwick, Change in MarineCommunities: An Approach to Statistical Analysis andInterpretation (2nd edition: PRIMER-E PlymouthMarine, 2001).
27. B. C. Coull and S. S. Herman, “Zoogeographic and Par-allel Levelbottom Communities of the Meiobenthic Har-pacticoida (Crustacea, Copepoda) of Bermuda,” Oecolo-gia 5, 392–399 (1970).
28. C. Cristoni, M. A. Colangelo, and V. U. Ceccherelli,“Spatial Scale and Meiobenthic Copepod Recolonisa-tion: Testing the Effect of Disturbance Size in the Sea-grass Habitat,” Mar. Biol. 298, 49–70 (2004).
29. Die Biodiversität in der Deutschen Nord- und Ostsee,Ed. by S.A. Gerlach (Bundesanstalt für Gewässerkunde,Koblenz, Germany, 2004. (V. 2), pp. 133–141.
30. J. W. Fleeger and A. W. Decho, “Spatial Variability ofInterstitial Meiofauna: A Review,” Stygol 3 (1), 35–54(1987).
31. L. Garlitska, “Species Diversity and Type of Harpacti-coid Copepod Distribution in Water Areas with HighAnthropogenic Influence,” in The International Work-shop on Black Sea Benthos, Ed. by B. Ozturk et al.(Istanbul, Turkey, 2004), pp. 181–189.
32. J. M. Gee, “Impact of Epibenthic Predation on EstuarineIntertidal Harpacticoid Copepod Populations,” Mar.Biol. 96, 479–510 (1987).
33. J. C. R. Govaere, D. Van Damme, C. Heip, andL. A. P. De Coninck, “Benthic Communities in theSouthern Bight of the North Sea and Their Use in Eco-logical Monitoring,” Helgoländer Meeresunters 33,507–521 (1980).
34. G. R. F. Hicks, “Species Composition and Zoogeogra-phy of Marine Phytal Harpacticoid Copepods from CookStrait, and Their Contribution to Total Phytal Meio-fauna,” N. Z .J. Mar. Freshwat. Res. 11, 441–469 (1977).
35. G. R. F. Hicks and B. C. Coull, “The Ecology of MarineMeiobenthic Copepod,” Oceanogr. Mar. Biol. Ann Rev.21, 67–175 (1983).
36. R. Huys and G. A. Boxshall, Copepod Evolution (RaySociety, London, 1991).
37. K. Lang, Monographi der Harpacticiden (A-B NordiskaBokhandeln, Stockholm, 1948).
38. A. Monard, “Les Harpacticoïdes Marins de la Region deSalambò,” Bull. Stat. Oceanogr. Salambò 34, 1–94(1935).
39. C. G. Moore, “Meiofauna of the Industrialised Estuaryand Firth of Forth, Scotland,” Proc. Royal Soc. Edin-burgh 93, 415–430 (1987).
40. F. D. Por, “A Study of the Levantine and Pontic Harpac-ticoida (Crustacea, Copepoda),” Zool. Verh. 64, 1–128(1964).
41. J. Soyer, “Bionomie Benthique du Plateau Continentalde la Côte Catalane Française. III. Les Peuplements deCopépodes Harpacticoïdes (Crustacea),” Vie Milieu Ser.B 21, 337–511 (1970).
42. B. Sun, J. W. Fleeger, and R. S. Carney, “SedimentMicrotopography and the Small-Scale Spatial Distribu-tion of Meiofauna,” Exp. Mar. Biol. Ecol. 167 (1), 73–90(1993).
43. D. Van Damme, C. Heip, and K. A. Willems, “Influenceof Pollution on the Harpacticoid Copepods of Two NorthSea Estuaries,” Hydrobiologia 112, 143–160 (1984).
44. J. B. J. Wells, “Biogeography of Benthic HarpacticoidCopepods of the Marine Littoral and Continental Shelf,”Syllogeus 58, 126–135 (1986).
45. D. R. Woods and J. H. Tijtjen, “Horizontal and VerticalDistribution of Meiofauna in the Venezuela Basin,” Mar.Geol. 68, 233–241 (1985).