space matters when defining effective management for ... · space matters when defining effective...
TRANSCRIPT

BIODIVERSITYRESEARCH
Space matters when defining effectivemanagement for invasive plants
Eliane S. Meier1*, Stefan Dullinger2,3, Niklaus E. Zimmermann4,
Daniel Baumgartner5, Andreas Gattringer2 and Karl H€ulber2,3
1Agroscope, Institute for Sustainability
Sciences ISS, Reckenholzstrasse 191, CH-
8046 Z€urich, Switzerland, 2Vienna Institute
for Nature Conservation & Analyses,
Giessergasse 6/7, A-1030 Vienna, Austria,3Department of Conservation Biology,
Vegetation and Landscape Ecology, Faculty
Centre of Biodiversity, University of Vienna,
Rennweg 14, A-1030 Vienna, Austria,4Dynamic Macroecology Group, Landscape
Dynamics Unit, Swiss Federal Research
Institute WSL, Zuercherstrasse 111, CH-8903
Birmensdorf, Switzerland, 5Economics and
Social Sciences Group, Regional Economics
and Development Unit, Swiss Federal
Research Institute WSL, Zuercherstrasse 111,
CH-8903 Birmensdorf, Switzerland
*Correspondence: Eliane S. Meier, Agroscope,
Institute for Sustainability Sciences ISS,
Reckenholzstrasse 191, CH-8046 Z€urich,
Switzerland.
E-mail: [email protected]
ABSTRACT
Aim Invasive alien species are a threat to biodiversity and can harm resident
plants, animals, humans and infrastructure. To reduce deleterious effects, effec-
tive management planning for invasive plants is required. Currently, the effec-
tiveness of management is primarily optimized locally through eradication of
individual populations. By contrast, spatial prioritization of control activities at
the landscape level has received less attention, despite its potential to improve
management planning in complex landscapes, especially under budget
constraints.
Location North-eastern Switzerland, Europe.
Methods We used a dynamic simulation model to evaluate the effectiveness of
spatially designed management planning for controlling the expansion of three
invasive alien plants (IAPs; Heracleum mantegazzianum, Impatiens glandulifera
and Reynoutria japonica) across a heterogeneous landscape in North-eastern
Switzerland. The model predicted the spread of IAPs from their current distri-
bution under constraints of 361 control options differing in local intensity,
frequency, duration, area and spatial prioritization of eradication measures.
Results Our results demonstrate that IAP-control actions under a restricted
budget are more effective if control actions are spatially prioritized. Most effec-
tive spatial treatments generally prioritized small populations in the case of the
annual species and large populations in the case of the perennial species. Fur-
ther, applying intensive control at early stages generally increased effectiveness
of control.
Main conclusions For IAP-management planning, our findings suggest that
control should be applied early when IAPs start spreading, to maximize success
or minimize costs. Further, spatial prioritization schemes are particularly useful
under limited financial means for IAP-management. Finally, our modelling
approach may serve as a proof of concept to evaluate the effectiveness of
control actions of various IAPs in complex landscapes.
Keywords
Alien species, biological invasions, complex landscapes, Heracleum mante-
gazzianum, Impatiens glandulifera, population growth, Reynoutria japonica,
riparian habitats, spatial spread, species distribution model.
INTRODUCTION
Alien invasive species reduce ecosystem functions and ser-
vices by out-competing native species, harming the health of
humans and domestic animals, damaging infrastructure,
providing feeding niches for other pests, altering nutrient
cycling or changing fire and water regimes (Vil�a et al., 2011).
Evaluating the efficiency of control actions to constrain bio-
logical invasions is hence an important issue (e.g. Brabec &
Py�sek, 2000; Shea et al., 2010). So far, most studies have
DOI: 10.1111/ddi.12201ª 2014 John Wiley & Sons Ltd http://wileyonlinelibrary.com/journal/ddi 1029
Diversity and Distributions, (Diversity Distrib.) (2014) 20, 1029–1043A
Jou
rnal
of
Cons
erva
tion
Bio
geog
raph
yD
iver
sity
and
Dis
trib
utio
ns

focused on local eradication measures by comparing, for
example, the effects of chemicals, mechanical treatments or
biological agents on local population growth and spatial
spread (Shea et al., 2010). Such insights improved control
actions of invasive alien plants (IAPs) at local scales (Mack
et al., 2000; Hulme, 2006; Wilson et al., 2011). However,
today many IAPs have already become widespread, and
because of limited financial resources, public authorities have
to set priorities at a landscape scale. Yet, studies at the land-
scape scale are rare (Krug et al., 2010), and thus, the ques-
tion when, where and how much control should be applied
within a landscape is still largely unsolved (Epanchin-Niell &
Hastings, 2010).
To improve the impact of landscape-wide management
planning, control actions may follow different prioritization
criteria (Ib�a~nez et al., 2009; Andrew & Ustin, 2010): Hulme
(2003), for example, advised prioritization of large popula-
tions for control because these are principal sources of prop-
agules. In contrast, other studies advocate to prioritize small
outlying populations because they may contribute most to
range expansion due to their large edge-to-area ratio and
because outlying populations are often surrounded by uncol-
onized but suitable habitats (Moody & Mack, 1988; Higgins
et al., 2000; Taylor & Hastings, 2004). Further, it has been
suggested to prioritize populations in large connected habi-
tats because spread is reduced with increasing habitat frag-
mentation (With, 2002). Others finally proposed to focus
control on the main pathways of dispersal vectors (Wads-
worth et al., 2000), such as running waters, animals or
humans (Damschen et al., 2008). To control hydrochorous
IAP, for example, populations near streams and especially at
upper reaches may be targeted to reduce the sources for
downstream colonization (Wadsworth et al., 2000), or those
at lower reaches may be prioritized where arriving propa-
gules have not yet established a seedbank. However, none of
these studies addressed the complexity of landscapes despite
the great influence of habitat suitability and fragmentation
on patterns of spread (Smolik et al., 2010; Meier et al., 2012;
Richter et al., 2013a,b).
In practice, landscape-wide IAP-management planning has
to improve effectiveness of control under limited financial
resources. To measure the effectiveness of control, the dura-
tion until a population falls below an arbitrary size or is
locally eradicated is a straightforward measure (Regan et al.,
2006), although only applicable for isolated populations (So-
low et al., 2008). Where individuals may continue to invade
from neighbouring areas, effectiveness of control may be
evaluated in terms of population growth or spatial spread
(Shea et al., 2010). For modelling financial limitations of
IAP-management planning, a scenario-based approach for
available public spending for IAP-control appears useful,
because cost functions of landscape-wide IAP-control actions
are complex and hence may not be deduced from theory as
assumed in bioeconomic models for IAP-control (Keller
et al., 2009; Epanchin-Niell & Hastings, 2010). Scenario
techniques can deal with cost-proxies that are advisable to
use at the landscape scale because sound data on specific
costs per unit area are either lacking or highly volatile
(Hauser & McCarthy, 2009).
Currently, most models of invasion control are based on
reaction-diffusion processes (e.g. Flather & Bevers, 2002), in-
tegro-difference equations (e.g. Kot et al., 1996), matrix
models (e.g. Shea & Kelly, 2004; Ramula et al., 2008), gravity
models (e.g. Bossenbroek et al., 2001) or cellular automata
(e.g. Higgins et al., 2000). Such models reliably determine
the effectiveness of IAP-control within simple landscapes
(Moody & Mack, 1988; Wadsworth et al., 2000). Although
some progress has been made regarding the effects of control
actions for widespread IAPs in heterogeneous landscapes
(e.g. Wadsworth et al., 2000; Blackwood et al., 2010), synop-
tic insights for complex landscapes are lacking (but see Krug
et al., 2010). To account for the complexity of real land-
scapes, a promising approach is to combine species distribu-
tion models (SDMs; Guisan & Zimmermann, 2000) that are
useful to assess habitat suitability variation across landscapes
with population spread models (Dullinger et al., 2012) that
cover meta-population dynamics.
The goal of our study is to use such a model to evaluate
the effectiveness of IAP-control actions for four spending
scenarios and four management goals applied to three wide-
spread IAPs in a complex landscape of the Swiss plateau.
Specifically, we addressed the following two questions: (1)
Does effectiveness and specifications of the most effective
control options vary between management goals, financial
spending levels or species? and (2) is the effectiveness of con-
trol increased, if control follows a set of criteria for spatial
prioritization instead of being applied to randomly selected
locations? To tackle these questions, we simulated IAP-spread
over 15 years constrained by 361 control options differing in
local intensity, area treated, treatment frequency, duration of
treatments and spatial prioritization. We selected three IAPs
that preferentially spread in riparian habitats but differ in life
forms: Heracleum mantegazzianum Sommier et Levier (giant
hogweed), Impatiens glandulifera Royle (Himalayan balsam)
and Reynoutria japonica Houtt. (Japanese knotweed).
METHODS
Study area
Our study area comprised the Canton Zurich (1729 km2;
47°090 N–47°410 N, 8°210 E–8°590 E) located on the North-
eastern Swiss plateau. The number of IAPs in Switzerland is
above the European average (EEA/SEBI, 2010), and particu-
larly the area around Zurich is a hub of human-mediated
invasions owing to its high population density and high
coverage of agricultural areas (Nobis et al., 2009). The cli-
mate is oceanic, elevation ranges from 330 to 1289 m above
sea level, and numerous river valleys provide suitable habitats
for hydrochorous IAPs.
1030 Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd
E. S. Meier et al.

Species and environmental data
We selected three study species, which are all widespread in
the study area, spread in the same habitats, but differ in
life cycles: Heracleaum mantegazzianum (H.man), Impatiens
glandulifera (I.gla) and Reynoutria japonica (R.jap). They
were introduced in the Canton Zurich at the beginning of
the 20th century and were used as garden plants, forage
plants and/or for erosion control (Beerling & Perrins, 1993;
Py�sek & Prach, 1993). All study species spread rapidly
mainly into ruderal riparian habitats and out-compete
native species. Further, H.man harms the health of humans
and domestic animals due to its phototoxicity, whereas I.gla
and R.jap facilitate erosion of riverbanks and damage infra-
structure. H.man is monocarpic perennial, I.gla is annual,
and R.jap is polycarpic perennial but disperses in Europe
exclusively vegetatively (see Table S1 in Supporting Infor-
mation).
Species occurrences were derived from point and polygon
data on species abundance and area occupied in the Canton
of Zurich (AWEL, 2010). For H.man, I.gla and R.jap, we
used n = 1216, n = 2251 and n = 2346 observations, respec-
tively, taken between 2006 and 2010 by municipality employ-
ees, nature conservation authorities and volunteers. The
sampling is quite uneven resulting in ‘well-sampled munici-
palities’ and ‘insufficiently sampled municipalities’.
To describe habitat suitability and propagule pressure, we
selected 17 variables among a comprehensive set of environ-
mental descriptors. To characterize habitat suitability, in
terms of climate, we selected growing season degree-days
(°C d) from downscaled monthly temperature maps of cur-
rent climate (1950–2000; Hijmans et al., 2005). In terms of
topography, we computed an aspect value (0–100 for south-
to north-facing slopes), potential yearly global radiation
(kj m�2 d�1), and a topographical position index (m) from
a digital elevation model (Swisstopo, 2003). In terms of soil,
we selected coarse-fragment content (% volume of mineral
fragment in the soil; BFS & GEOSTAT, 2000). To character-
ize propagule pressure, we used the Cantonal structure plan
to compute a land-use variable with the categories ‘settle-
ment’, ‘agriculture’, ‘forest’, ‘nature reserve’ and ‘water
bodies’ (ARE, 2001). In addition, we calculated Euclidean
distances to the following objects as they might either serve
as preferred habitats or as migration corridors for the study
species: (1) open gravel pits (AWEL, 2008), uncovered
creeks, uncovered rivers, lakes (all from AWEL, 2007); (2)
buildings, forest edges, small roads, large roads, railroads,
bridges (all from Swisstopo, 2008); and (3) house construc-
tion activities (ARE, 2001). All predictor grids were gener-
ated at a 25-m spatial resolution.
Modelling IAP-spread
For modelling IAP-spread, we used species distribution
models to map habitat suitability. Based on these habitat suit-
ability maps, a hybrid model was used for simulating IAP-dis-
persal and spread simulations under different control options
for evaluating the effectiveness of control options (Fig. 1).
Species distribution modelling
Distributiondata
Environmentalpredictors
Samplingintensity
Habitat suitability
Hydrochorous Anemochorous
Landscape-wide management- Frequency- Duration- Local Intensity- Area- Spatial allocation pattern
Simulating populationdynamics and spreadstarting from current
observations
Ranking managementoptions
Spending
Effe
ct
Demographic modelling
Dispersal modelling
DistanceDistance
Habitat suitability
See
ds
See
ds
See
dsG
erm
inat
ion
Est
ablis
hmen
t
Habitat suitability
Habitat suitability
Figure 1 Overview of model set-up. Species distribution models were used to estimate habitat suitability. Habitat suitability
determined the rates of local demographic processes, which in turn controlled the number of individuals at a site and the number of
seeds produced by these individuals. Juveniles and adults were removed just before seeding according to 361 management options. Local
seed yields were re-distributed using to wind and secondary water dispersal kernels. Effectiveness of landscape-wide management was
estimated within four spending levels.
Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd 1031
Spatial management for controlling invasive plants

Mapping suitable habitats with SDMs
We mapped habitat suitability for SDMs using generalized
linear models (GLMs), because they allow for spatial autocor-
relation correction of presence–absence data (Dormann et al.,
2007). Such a correction is required because ongoing spread
from nascent foci has generated highly autocorrelated regio-
nal distribution patterns in all three species (Dullinger et al.,
2009). To correct for spatial autocorrelation, we applied spa-
tial eigenvector filtering (SEVM-v3; Bini et al., 2009), which
is provided by the R-library ‘spacemakeR’ (Dray, 2008; R
Development Core Team, 2011). Further, because our data
set only included reported presences, we generated a set of
pseudo-absences. To reduce the probability of generating
pseudo-absences at locations where IAPs are present, we
randomly generated 80% of these pseudo-absences in well-
sampled municipalities outside a 25 m buffer of recorded
IAP-occurrences. The total number of pseudo-absences was
set equal to the number of presences of each species.
Binomial GLMs were calibrated with presences and
pseudo-absences as the response and the 17 environmental
variables [to reduce multicollinearity problems all |rs| < 0.6
and all Variance Inflation Factor (VIF; Neter et al., 1983)
<1.82] and the significant spatial eigenvectors as predictor
variables. The environmental variables were entered both as
linear and quadratic terms, while the spatial eigenvectors
were entered only as linear terms. Variable selection included
a combined backward and forward stepwise procedure based
on the Bayesian information criterion (BIC) to avoid overfit-
ting (Burnham & Anderson, 2002) applied to the full model.
Model fit was evaluated by the adjusted D2, and for model
accuracy, we used Cohen’s kappa and the area under the
ROC curve (AUC) using a 10-fold cross-validation. Non-
colonizable areas such as buildings and lakes were classified
as unsuitable per definition.
The hybrid model: simulating IAP-dispersal
The hybrid model combines elements of SDMs, demographic
and dispersal models to simulate the spread of plant species
(see Dullinger et al., 2012 for details). It operates on a
two-dimensional grid of cells and simulates changes in the
abundance and the distribution of populations on this grid
in discrete, annual time steps. Habitat suitability maps from
SDMs are used to determine species and grid cell-specific
vital rates such as germination, juvenile survival, age of
maturity, seed production and clonal growth, which are sub-
ject to density-dependent growth regulation to avoid growth
beyond a cell-specific carrying capacity. These vital rates
drive the year-to-year dynamics in grid cells, such as the
number of individuals in age structured cohorts (juveniles,
adults, seeds in the seedbank), and determine the annual seed
yield from each cell. Seeds are subsequently dispersed by
wind dispersal kernels covering both short- and long-distance
propagule transport (WALD model; Katul et al., 2005). For
this study, we added an additional dispersal pathway, namely
the re-distribution of seeds and plant fragments along creeks
and rivers. We assumed that this secondary dispersal follows
a negative exponential kernel along flow direction only
(Groves et al., 2009). Model parameters were derived from
the literature or selected so that the simulations provided a
good fit to measured data (Table S1).
Spread simulations under different control options
To avoid negative effects from unbalanced sampling of spe-
cies occurrences (Marchetto et al., 2010), we added five sets
of randomly allocated new populations in insufficiently sam-
pled municipalities. The size of these hypothetical popula-
tions was randomly drawn from an interval defined by the
means and standard deviations of recorded populations
(118.9 � 57.5, 152.2 � 105.5, 108.8 � 89.3 for H.man, I.gla,
and R.jap, respectively). Populations were added until the
mean number of individuals per suitable habitat cell in insuf-
ficiently sampled municipalities equalled that in well-sampled
municipalities. In total, per set 2675, 5767 and 902 artificial
populations were generated for the three study species.
We used the hybrid model to simulate the spread of the
three IAPs from the five different sets of initial distributions
over 15 years. Different control options were simulated by
reducing specific populations (=numbers of juveniles and
adults) in specific years by predefined percentages as described
below. Control was assumed to take place prior to seed pro-
duction, that is, it effectively reduced the population’s seed
yield and hence its propagule pressure in the respective year.
Simulations were run under a ‘do nothing’ option (i.e. no
removal of plants) and under 360 different control options.
These options represent all possible combinations of five dif-
ferent treatments varying in frequency and duration (annual,
bi-annual and five-annual treatments, and treatments for the
first N years only, where N is the total number of bi- and
five-annual treatments), local intensities of treatments (25%,
55% and 90% of juveniles and adults removed per treated
cell, while seed bank remains unchanged) and proportional
eradication of all cells per species within predefined areas
(5%, 25% and 90%) following eight different spatial prioriti-
zation schemes (selecting cells according to the criteria:
randomly, large populations, small populations, isolated
populations, well-connected habitats, riverbanks, upper
reaches of rivers and lower reaches of rivers; Table 1). This
resulted in 5 9 3 9 3 9 8 = 360 treatment combinations.
The frequencies, durations and local intensities of treatments
were selected to mimic common practice, such as mechanical
treatments (removal of stems or roots), chemical treatments
(foliar- or soil-active chemicals), cultural treatments (grazing,
re-vegetation, competition plantations, fertilization or fire) or
biological treatments (augmentation or release of enemies
from native or invaded range), which usually vary in local
intensity between 25% and 90% (Andersen, 1994; Masters &
Sheley, 2001; De Micheli et al., 2006). The eight different spa-
tial treatment allocations were selected based on biological
reasoning. ‘Large populations’ may reflect core populations
1032 Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd
E. S. Meier et al.

that represent principal sources of propagules (Hulme, 2003).
‘Small populations’ have large edge-to-area ratios and hence
may contribute most to range expansion (Moody & Mack,
1988; Taylor & Hastings, 2004). To target large and small
populations, we defined population sizes by summing up
individuals of adjacent cells and removed populations in
decreasing or increasing order of population size until the
respective clearing area and local intensity were exhausted.
‘Outlier populations’ are surrounded by un-colonized habitats
open for colonization. To target outlier populations, we
removed populations in decreasing order based on the mini-
mal distance between centroids until the clearing area and
local intensity were exhausted. We targeted cells within ‘large
well-connected habitats’ following the percolation theory say-
ing that increasing fragmentation may effectively reduce
migration (Turner, 1989; With, 2002). Therefore, we defined
large connected habitats with FRAGSTATS 3.3 (McGarigal et al.,
2002) as focal cells within a moving window of 500 m radius
that had >40% of habitat per window and a largest patch
index of <80%. Within these large connected habitats, we ran-
domly removed colonized cells until the clearing area and
local intensity were exhausted. Colonized cells on ‘riverbanks’
were targeted, because rivers are the main dispersal pathways
of hydrochorous plants. We targeted ‘upper reaches of rivers’,
as this reduces the propagule sources for downstream coloni-
zation (Wadsworth et al., 2000). We targeted ‘lower reaches
of rivers’, as this reduces the colonizing propagule pressure
before establishing a seedbank. We defined upper and lower
reaches according to the distance to their sources, and for
treatments, we selected colonized cells randomly within a 50-
m buffer along rivers or river sections until the clearing area
and local intensity were exhausted. Finally, we ‘randomly’ tar-
geted colonized cells to test the null hypothesis that the spatial
allocation of treatments has no influence on its effectiveness.
Evaluating the effectiveness of control options
During each of the 5415 (three species 9 five initial distribu-
tions 9 361 control options; Table 1) model runs, a range
of variables was stored, and the effectiveness of control
options was evaluated by comparing the following indices of
management goals against the simulation under the ‘do
nothing’ option: (1) mean spread distance, (2) size of the
area occupied, (3) number of adult individuals and (4) the
number of juvenile individuals and seeds (representing
the regeneration potential). All results were averaged among
the five initial distributions of the species.
We performed a variation partitioning analysis to deter-
mine for each species the influence of initial condition, fre-
quency and duration, local intensity, area and spatial
prioritization on the four management goals (Borcard et al.,
1992). Therefore, we estimated for each predictor variable
(i.e. initial condition, frequency and duration, local intensity,
area and spatial prioritization) its individual contribution in
explaining the dependent variable (i.e. effects on a manage-
ment goal) by subtracting the adjusted deviance (adj.D2;
Guisan & Zimmermann, 2000) of the full linear model with-
out the specific predictor from the adj.D2 of the full linear
model.
We estimated costs for control actions yearly by a spending
proxy because financial data on IAP-management are not
documented in Switzerland. The spending proxy ranged from
0% to 100% derived from averaging treatment frequency
(e.g. yearly treatments correspond to 100% while five-annual
treatments correspond to 20%), treatment duration (treat-
ments over 15 years = 100%), number of treated cells (if the
same number of cells are treated as are occupied in ‘do noth-
ing’ = 100%) and number of treated juveniles and adults (if
the same number of juveniles and adults are treated as are
occurring in ‘do nothing’ = 100%). The spending proxy over
all 15 years of simulation was calculated by averaging these
yearly spending proxies. Further, as no continuous cost func-
tions for species control were available, four discrete spending
scenarios (very low, low, medium or high) have been devel-
oped to describe different levels of financial resources avail-
able for IAP-management (Table 2). Thresholds of these four
levels of spending scenarios were set qualitatively for incorpo-
rating ‘realistic’ management scenarios. The higher the
Table 1 Overview of treatment characteristics that were combined to set up 360 control options for each species and initial condition.
Additionally, we included a ‘do nothing’ option, which only addresses combinations of species and initial conditions.
Species Initial condition Spatial prioritization Area Local intensity Frequency and duration
Heracleum
mantegazzianum
1–5 Random 5% 25% Annual treatments
Large populations 50% 55% Bi-annual treatments
Small populations 90% 90% Five-annual treatments
Impatiens glandulifera Outlying populations No. of bi-annual treatments set yearly
at the beginningWell-connected habitats
Riverbanks
Reynoutria japonica Upper reaches of rivers No. of five-annual treatments set yearly
at the beginning
Lower reaches of rivers
‘Initial condition’ refers to the sets of artificial populations added in insufficiently sampled municipalities. ‘Spatial prioritization’ refers to the different
spatial prioritization schemes. ‘Area’ refers to proportional eradication of all cells per species within predefined areas. ‘Local intensity’ refers to the%
of juveniles and adults removed per treated cell. ‘Frequency and duration’ refers to the temporal frequency and duration control was applied.
Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd 1033
Spatial management for controlling invasive plants

spending levels are, the more financial resources for IAP-con-
trol are available to invest in labour (i.e. qualified workers),
in capital (i.e. equipment) and in their coordination (i.e.
leadership; Besanko et al., 2004). These four spending levels
allow for comparisons within and among similar control
options from a financial point of view.
RESULTS
Colonization of suitable habitats
SDM quality was high for all three species (Table 3, full
model) with AUC and Kappa values for H.man, I.glan and
R.jap models reaching 0.89, 0.91, 0.90 and 0.62, 0.66, 0.64,
respectively. The importance of single predictors for the
current distribution is given in Table 3.
Colonization rates of suitable habitats under the ‘do noth-
ing’ strategy differed strongly between the three species
(Table S2). The simulated increase in area over 15 years on
the Swiss plateau (Fig. 2) was highest for I.gla (from 0.7% to
9.7% within 689 km2 of suitable habitat area in the study
area), lower for H.man (from 0.2% to 5.3% within 925 km2)
and lowest for R.jap (from 0.2% to 1.1% within 735 km2).
Effectiveness of control
The size of the treated area had the largest influence on the
effectiveness of control options, followed by the spatial prior-
itization pattern (Table 4). In contrast, initial conditions had
the least predictive power for any management goal of any
species.
Across all management options, the reduction in mean
spread distance was significantly less successful than the
reduction of area, reproduction and regeneration potential
(Kruskal–Wallis test, P < 0.001), because some management
options even increased mean spread distances compared with
the ‘do nothing’ option (Fig. 3).
Specifications of most effective control options
More than two-thirds of the most effective control options
per spending level, management goal and species apply the
highest possible local intensity over the entire duration of
the simulation (Table 5). Moreover, all most effective control
options have a high initial spending and thereby achieved
higher effects at lower overall costs than control options with
low initial spending (Fig. S1).
Specifications of most effective control options differed
between spending levels and species, but did not differ
between management goals. Among spending levels, the most
effective control options differed significantly in treatment
area, frequency, local intensity and duration (all Kruskal–
Wallis tests, P < 0.001), but not in spatial prioritization
schemes (Kruskal–Wallis test, P = 0.437). Among manage-
ment goals, the most effective options of IAP-control did not
Table 2 Four scenarios of spending proxies that were used to define thresholds of spending levels
Spending
level Implications for management agencies
Specifications
Spatial
prioritization
Area
(%)
Local intensity
(%)
Frequency and
duration
Very low Few workers, poor equipment, poor leadership Random* 5 25 Five-annual
Low Few workers, poor equipment, fair leadership Random* 5 25 Bi-annual
Medium Many workers, fair equipment, fair leadership Random* 50 25 Bi-annual
High Many workers, good equipment, strong
leadership
Random* 90 25 Annual
*Because no optimal spatial prioritization pattern is known, random allocation patterns are assumed for all spending levels.
Table 3 Deviance explained (adj.D2) for individual predictors
and the full model after spatial eigenvector filtering to explain
the spatial distribution and habitat suitability
Predictor
Heracleum
mantegazzianum
Impatiens
glandulifera
Reynoutria
japonica
Degree-days 0.05 0.01 0.05
Aspect value 0.00 0.01 0.00
Potential yearly global
radiation
0.02 0.02 0.02
Topographical
position
0.04 0.02 0.08
Coarse-fragment
content
0.01 0.01 0.05
Land use 0.04 0.09 0.06
Gravel pits 0.02 0.04 0.01
House building
activities
0.04 0.03 0.05
Buildings 0.03 0.03 0.04
Forest edges 0.01 0.06 0.00
Small roads 0.03 0.05 0.01
Large roads 0.06 0.01 0.04
Rail roads 0.01 0.04 0.04
Bridges 0.05 0.01 0.07
Creeks 0.02 0.03 0.00
Non-covered rivers 0.02 0.03 0.04
Lakes 0.02 0.01 0.02
Full model 0.24 0.33 0.24
Bold face numbers indicate values ≥ 0.05.
1034 Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd
E. S. Meier et al.

differ significantly (Kruskal–Wallis test, all P > 0.901).
Among species, the most effective control options differed
only significantly in their spatial prioritization schemes
(Kruskal–Wallis test, P = 0.004). For perennial species (i.e.
H.man and R.jap), treating large populations was generally
most effective, while for the annual I.gla treating small popu-
lations was a more successful strategy.
Effects of spatial prioritization of most effective
control options
Across spending levels, management goals and species, the
most effective randomly applied control actions was 2.7%
less effective than the most effective management option
under spatial prioritization schemes and required 0.5% more
(a) (b) (c)
Figure 2 Current and future distribution of (a) Heracleum mantegazzianum, (b) Impatiens glandulifera and (c) Reynoutria japonica in
Canton Zurich, Switzerland. Black squares represent current observations, grey areas represent areas with a high probability of current
occurrence (threshold at max. Kappa), and red areas are predicted to be colonized after 15 years of simulation without control.
Table 4 Deviance explained (adj.D2) of the effects on management goals by the full model and individual predictors derived by
variation partitioning (most important factors are indicated in bold face)
Species Management goal (response)
R2 of predictor
Full
model
Initial
condition
Spatial
prioritization Area
Local
intensity
Frequency and
duration
Heracleum mantegazzianum Effect on area 0.972 0.000 0.277 0.501 0.086 0.117
Effect on adults 0.966 0.000 0.359 0.476 0.053 0.086
Effect on spread 0.780 0.000 0.237 0.361 0.081 0.108
Effect on regeneration 0.973 0.000 0.379 0.447 0.057 0.098
Impatiens glandulifera Effect on area 0.948 0.000 0.171 0.555 0.062 0.167
Effect on adults 0.965 0.000 0.158 0.535 0.000 0.215
Effect on spread 0.627 0.027 0.126 0.236 0.042 0.201
Effect on regeneration 0.953 0.000 0.250 0.481 0.064 0.166
Reynoutria japonica Effect on area 0.957 0.001 0.08 0.609 0.173 0.102
Effect on adults 0.961 0.000 0.113 0.641 0.116 0.099
Effect on spread 0.481 0.001 0.030 0.175 0.219 0.059
Effect on regeneration 0.914 0.000 0.117 0.610 0.096 0.099
Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd 1035
Spatial management for controlling invasive plants

(a) (b) (c)
1036 Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd
E. S. Meier et al.

spending (Table S3). Regarding spending levels, spatial prior-
itization increased the success of most effective control
actions at very low (Mann–Whitney U-test, all P < 0.010)
and under low or medium spending (all between P = 0.009
and P = 0.180), but not at high spending levels (all
P > 0.258). Regarding the different management goals, spa-
tial prioritization tended to reduce the colonized area, adults
and regeneration (Mann–Whitney U-test, all P < 0.104), but
did not reduce mean spread distances (P = 0.307). Regarding
species, control options with specific spatial prioritization
schemes increased effectiveness for I.gla (Mann–Whitney
U-test, all P < 0.138) and R.jap (all P < 0.078), but not for
H.man (all P > 0.178) compared with random spatial control
actions.
DISCUSSION
Effectiveness of IAP-control actions
The lack of a landscape-scale perspective can limit the effec-
tiveness of IAP-control measures. Here, we explicitly took
this perspective by simulating spatio-temporal spread in a
real landscape. We showed that (i) specific spatial prioritiza-
tion schemes increase the effectiveness of IAP-control, (ii)
the effect of spatial prioritization increases with decreasing
budgets available for control, (iii) most effective spatial pri-
oritization schemes differ between species, (iv) IAP-control
actions are most effective when extensive control is applied
early, and (v) ineffective IAP-control actions increase mean
spread distances.
The spatial prioritization scheme is (apart from the area
treated by control measures) the most important factor to
enhance effectiveness of IAP-control actions. Thus, in line
with Andrew and Ustin (2010), prioritizing control on popu-
lations that most strongly contribute to spatial spread and
overall population growth is also effective in complex land-
scapes. However, in line with the controversial findings on
most effective spatial prioritization schemes by previous
studies (Moody & Mack, 1988; Higgins et al., 2000; Wads-
worth et al., 2000; Hulme, 2003; Taylor & Hastings, 2004),
we found no one-fits-all best prioritization scheme. The
effectiveness of a spatial prioritization scheme depends on
the available budget, the target species and its invasion stage,
and the habitat configuration of the landscape. Moreover,
even though some spatial prioritization patterns may be
highly effective, they may not be straightforward to apply in
a real landscape. For instance, spatial prioritization
patterns that can be delivered as map products to practitioners
(e.g. treatment of upper reaches of rivers) may be easier to
implement than theoretically motivated choices of spatial
prioritization patterns (e.g. treatment of small populations).
Spatial prioritization of IAP-control is particularly impor-
tant under budget restrictions. The higher the spending level,
the less important is the spatial prioritization of control:
effectiveness is most influenced by the area that can be trea-
ted. However, the larger the area, the less different are the
spatial prioritization patterns, and thus, the lower is the
influence from the spatial prioritization patterns. Further,
the most effective spatial prioritization schemes did rarely
differ between spending levels. This is not in line with Taylor
and Hastings (2004) who propose to select spatial prioritiza-
tion patterns according to the annual budget available (i.e.
removal of low-density areas for low and medium budgets
and removal of high-density areas for high budgets).
Most effective spatial prioritization schemes differ between
species. For the perennial IAPs, most pronounced effects
were achieved by primarily treating large populations. Large
populations are generally older, and thus, for monocarpic
perennials (i.e. H.man) and clonally reproducing perennials
(i.e. R.jap), they contain a higher proportion of adults that
produce seeds or fragments, respectively, compared with
small populations. Contrarily, for the annual species (i.e.
I.gla), best effects were achieved by primarily treating small
populations before they spread radially into vacant habitats
(Moody & Mack, 1988; Taylor & Hastings, 2004). Thus, life
history traits are important when defining effective control
actions for invasive species (Ramula et al., 2008).
In line with Hulme (2006), we demonstrate that IAP-con-
trol actions are most effective when extensive control is
applied during early invasion stages. Given the same total
resource input early, control was more effective than apply-
ing treatment at regular intervals. Control options with high
initial spending achieved higher effects at lower overall costs
than those with low initial spending (Fig. S1). Moreover,
when extensive control is applied early, yearly effects
increased while yearly spending decreased over the 15 simu-
lation years (Fig. S1). Thus, control options with high initial
investment resulted in a clear reduction in incurring
expenses after a few years.
Ineffective IAP-control actions may even increase mean
spread distances. Regarding the management goal to reduce
spread distances, our simulations suggest that ineffective
management may induce overcompensatory effects by
increasing germination rates, juvenile survival and clonal
propagation for species that reproduce and spread within
1 year (i.e. I.gla and R.jap). This may lead to spread dis-
tances being larger than without any control actions. This
agrees with results from local-scale studies showing that the
Figure 3 Effectiveness and spending of management options over 15 years for the management goals ‘reducing spread’, ‘reducing area’,
‘reducing adults’ and ‘reducing regeneration potential’ for (a) Heracleum mantegazzianum, (b) Impatiens glandulifera and (c) Reynoutria
japonica. Black and grey crosses represent management options with and without spatial prioritization. Dotted black lines separate the
four spending levels (vertical). The most effective management option for each spending level is given as coloured dot (blue-very low
spending, green-low, yellow-medium, red-high). The solid horizontal line indicates the effect of ‘do nothing’.
Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd 1037
Spatial management for controlling invasive plants

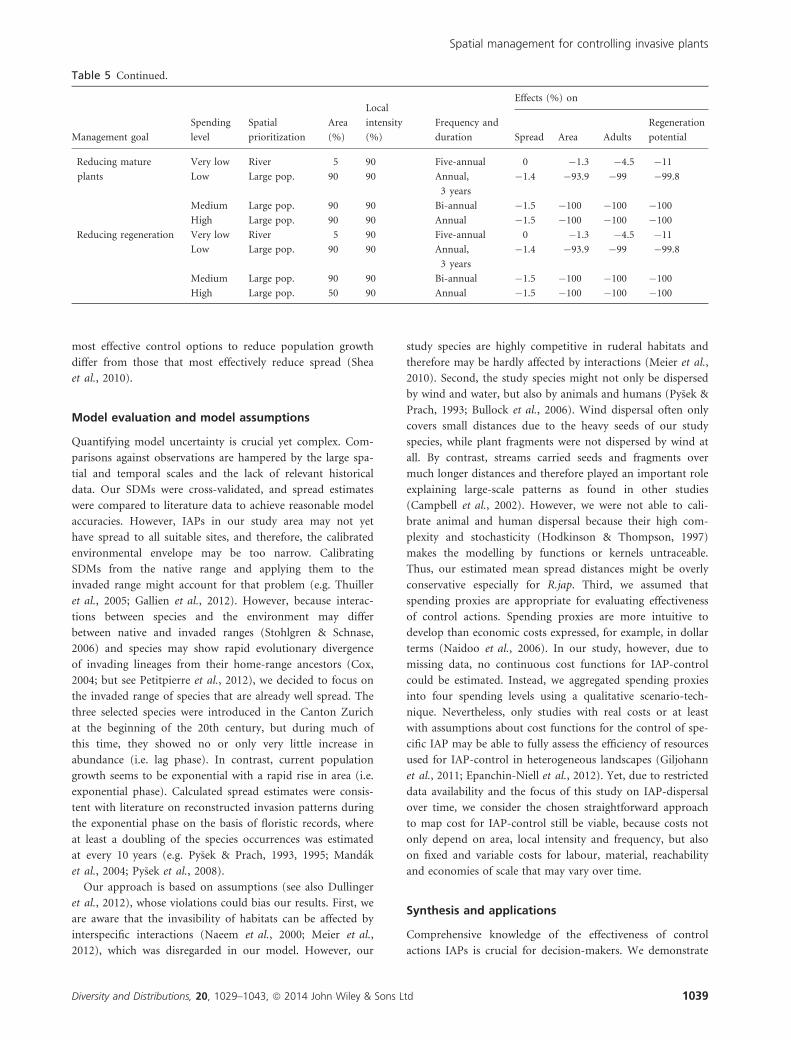
Table 5 Overview of most effective control options for each species, management goal and spending level. Effects were evaluated by
comparing the indices of the four management goals against the simulation under the ‘do nothing’ option
Management goal
Spending
level
Spatial
prioritization
Area
(%)
Local
intensity
(%)
Frequency and
duration
Effects (%) on
Spread Area Adults
Regeneration
potential
Heracleum mantegazzianum
Reducing spread Very low Connected
hab.
5 90 Five-annual �0.2 �2.1 �5.7 �4.2
Low Large pop. 90 90 Five-annual �6.5 �72.3 �94.7 �86.2
Medium Large pop. 90 90 Bi-annual �8.4 �100 �100 �100
High Large pop. 90 90 Annual �8.4 �100 �100 �100
Reducing area Very low Connected
hab.
5 90 Five-annual �0.2 �2.1 �5.7 �4.2
Low Small pop. 90 90 Five-annual �6.5 �73.5 �93.7 �85.1
Medium Large pop 90 90 Bi-annual �8.4 �100 �100 �100
High Large pop. 90 90 Annual �8.4 �100 �100 �100
Reducing mature
plants
Very low Connected
hab.
5 90 Five-annual �0.2 �2.1 �5.7 �4.2
Low Large pop. 90 90 Five-annual �6.5 �72.3 �94.7 �86.2
Medium Small pop. 90 90 Bi-annual �8.3 �100 �100 �100
High Large pop. 90 90 Annual �8.4 �100 �100 �100
Reducing regeneration Very low Connected
hab.
5 90 Five-annual �0.2 �2.1 �5.7 �4.2
Low Large pop. 90 90 Five-annual �6.5 �72.3 �94.7 �86.2
Medium Large pop. 90 90 Bi-annual �8.4 �100 �100 �100
High Small pop. 90 90 Annual �8.4 �100 �100 �100
Impatiens glandulifera
Reducing spread Very low Small pop. 5 55 Annual,
3 years
�0.5 �3.8 �3.8 �3.9
Low Small pop. 90 90 Five-annual �2.2 �28 �34.2 �34.1
Medium River 90 90 Bi-annual �3.6 �31.4 �45.1 �24.9
High Large pop. 90 90 Annual �9.2 �92.8 �99.3 �99.5
Reducing area Very low Small pop. 5 25 Five-annual �0.4 �4 �3.9 �3.9
Low Small pop. 90 90 Five-annual �2.2 �28 �34.2 �34.1
Medium River 90 90 Bi-annual �3.6 �31.4 �45.1 �24.9
High Small pop. 90 90 Annual �8.8 �93.8 �99.6 �99.6
Reducing mature
plants
Very low Small pop. 5 25 Five-annual �0.4 �4 �3.9 �3.9
Low Small pop. 90 90 Five-annual �2.2 �28 �34.2 �34.1
Medium Connected
hab.
90 90 Bi-annual �2.8 �27.4 �47 �48.3
High Small pop. 90 90 Annual �8.8 �93.8 �99.6 �99.6
Reducing regeneration Very low Small pop. 5 90 Annual,
3 years
�0.4 �3.9 �3.8 �3.9
Low Small pop. 90 90 Five-annual �2.2 �28 �34.2 �34.1
Medium Connected
hab.
90 90 Bi-annual �2.8 �27.4 �47 �48.3
High Small pop. 90 90 Annual �8.8 �93.8 �99.6 �99.6
Reynoutria japonica
Reducing spread Very low Small pop. 5 90 Five-annual �0.1 �4.5 �3.7 �0.9
Low Large pop. 90 90 Annual,
3 years
�1.4 �93.9 �99 �99.8
Medium Large pop. 90 90 Bi-annual �1.5 �100 �100 �100
High Large pop. 50 90 Annual �1.5 �100 �100 �100
Reducing area Very low Small pop. 5 90 Five-annual �0.1 �4.5 �3.7 �0.9
Low Large pop. 90 90 Annual,
3 years
�1.4 �93.9 �99 �99.8
Medium Large pop. 90 90 Bi-annual �1.5 �100 �100 �100
High Large pop. 90 90 Annual �1.5 �100 �100 �100
1038 Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd
E. S. Meier et al.
most effective control options to reduce population growth
differ from those that most effectively reduce spread (Shea
et al., 2010).
Model evaluation and model assumptions
Quantifying model uncertainty is crucial yet complex. Com-
parisons against observations are hampered by the large spa-
tial and temporal scales and the lack of relevant historical
data. Our SDMs were cross-validated, and spread estimates
were compared to literature data to achieve reasonable model
accuracies. However, IAPs in our study area may not yet
have spread to all suitable sites, and therefore, the calibrated
environmental envelope may be too narrow. Calibrating
SDMs from the native range and applying them to the
invaded range might account for that problem (e.g. Thuiller
et al., 2005; Gallien et al., 2012). However, because interac-
tions between species and the environment may differ
between native and invaded ranges (Stohlgren & Schnase,
2006) and species may show rapid evolutionary divergence
of invading lineages from their home-range ancestors (Cox,
2004; but see Petitpierre et al., 2012), we decided to focus on
the invaded range of species that are already well spread. The
three selected species were introduced in the Canton Zurich
at the beginning of the 20th century, but during much of
this time, they showed no or only very little increase in
abundance (i.e. lag phase). In contrast, current population
growth seems to be exponential with a rapid rise in area (i.e.
exponential phase). Calculated spread estimates were consis-
tent with literature on reconstructed invasion patterns during
the exponential phase on the basis of floristic records, where
at least a doubling of the species occurrences was estimated
at every 10 years (e.g. Py�sek & Prach, 1993, 1995; Mand�ak
et al., 2004; Py�sek et al., 2008).
Our approach is based on assumptions (see also Dullinger
et al., 2012), whose violations could bias our results. First, we
are aware that the invasibility of habitats can be affected by
interspecific interactions (Naeem et al., 2000; Meier et al.,
2012), which was disregarded in our model. However, our
study species are highly competitive in ruderal habitats and
therefore may be hardly affected by interactions (Meier et al.,
2010). Second, the study species might not only be dispersed
by wind and water, but also by animals and humans (Py�sek &
Prach, 1993; Bullock et al., 2006). Wind dispersal often only
covers small distances due to the heavy seeds of our study
species, while plant fragments were not dispersed by wind at
all. By contrast, streams carried seeds and fragments over
much longer distances and therefore played an important role
explaining large-scale patterns as found in other studies
(Campbell et al., 2002). However, we were not able to cali-
brate animal and human dispersal because their high com-
plexity and stochasticity (Hodkinson & Thompson, 1997)
makes the modelling by functions or kernels untraceable.
Thus, our estimated mean spread distances might be overly
conservative especially for R.jap. Third, we assumed that
spending proxies are appropriate for evaluating effectiveness
of control actions. Spending proxies are more intuitive to
develop than economic costs expressed, for example, in dollar
terms (Naidoo et al., 2006). In our study, however, due to
missing data, no continuous cost functions for IAP-control
could be estimated. Instead, we aggregated spending proxies
into four spending levels using a qualitative scenario-tech-
nique. Nevertheless, only studies with real costs or at least
with assumptions about cost functions for the control of spe-
cific IAP may be able to fully assess the efficiency of resources
used for IAP-control in heterogeneous landscapes (Giljohann
et al., 2011; Epanchin-Niell et al., 2012). Yet, due to restricted
data availability and the focus of this study on IAP-dispersal
over time, we consider the chosen straightforward approach
to map cost for IAP-control still be viable, because costs not
only depend on area, local intensity and frequency, but also
on fixed and variable costs for labour, material, reachability
and economies of scale that may vary over time.
Synthesis and applications
Comprehensive knowledge of the effectiveness of control
actions IAPs is crucial for decision-makers. We demonstrate
Table 5 Continued.
Management goal
Spending
level
Spatial
prioritization
Area
(%)
Local
intensity
(%)
Frequency and
duration
Effects (%) on
Spread Area Adults
Regeneration
potential
Reducing mature
plants
Very low River 5 90 Five-annual 0 �1.3 �4.5 �11
Low Large pop. 90 90 Annual,
3 years
�1.4 �93.9 �99 �99.8
Medium Large pop. 90 90 Bi-annual �1.5 �100 �100 �100
High Large pop. 90 90 Annual �1.5 �100 �100 �100
Reducing regeneration Very low River 5 90 Five-annual 0 �1.3 �4.5 �11
Low Large pop. 90 90 Annual,
3 years
�1.4 �93.9 �99 �99.8
Medium Large pop. 90 90 Bi-annual �1.5 �100 �100 �100
High Large pop. 50 90 Annual �1.5 �100 �100 �100
Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd 1039
Spatial management for controlling invasive plants

that taking a landscape-scale perspective on IAP-control sig-
nificantly enhances the effectiveness of control. Specifically,
our simulations peak in four recommendations: (i) IAP-
management planning should set spatial priorities of control
at a landscape level. Yet, spatial priorities must account for
specific landscape characteristics (e.g. the configuration of
suitable habitats), the stage of invasion and species traits,
because there is no one-fits-all control option that ensures
most effective control for all species and landscapes. (ii) Spa-
tial prioritization of IAP-control is particularly important
under budget restrictions. (iii) A careful selection of IAP-
control options is crucial, because spread of invading species
can be increased by ineffective control compared with doing
nothing. (iv) Given the same overall input of resources,
cumulative early control is more promising than intervening
bit by bit.
From a practical view point, optimal control options may
not be equally easy to implement. For instance, one may
provide practitioners with maps for controlling species in
highly connected habitats or along reaches of rivers. Advising
them to focus on small or large populations is much more
complex, because population sizes are highly volatile and
hence may not easily be mapped. This calls, however, for
both sound monitoring to assist management planning and
early treatment ascertaining rapid success.
Finally, the hybrid modelling approach used in this study
may substantially improve IAP-management planning at a
strategic level. Our modelling approach allows combining
IAP monitoring with an integrated tool for evaluating past,
present and planned future actions for IAP-control. In com-
bination with more detailed data on costs for IAP-control,
the modelling approach presented in this paper may help to
spend sparse financial resources in more efficient ways than
based exclusively on currently used bioeconomic optimiza-
tion models.
ACKNOWLEDGEMENTS
We thank T W€ust for technical assistance, Canton Zurich
(AWEL, Biosecurity) for access to cantonal databases, the EU
FP6 Grant GOCE-CT-2007-036866 (ECOCHANGE) for
funding and the Swiss Multi-Science Computing Grid
(SMSCG) for providing parts of the computing resources.
REFERENCES
Andersen, U.V. (1994) Sheep grazing as a method of control-
ling Heracleum mantegazzianum. Ecology and management
of invasive riverside plants (ed. by L.C. Waal, L.E. Child,
P.M. Wade and J.H. Brock), 217 p. Wiley, Chichester.
Andrew, M.E. & Ustin, S.L. (2010) The effects of temporally
variable dispersal and landscape structure on invasive spe-
cies spread. Ecological Applications, 20, 593–608.
ARE (2001) Kantonaler Richtplan (ed. Amt f€ur Raumord-
nung und Vermessung Kanton Z€urich).
AWEL (2007) €Okomorphologische Erhebung der Fliess-
gew€asser im Kanton Z€urich (ed. Amt f€ur Abfall Wasser
Energie und Luft AWEL).
AWEL (2008) Kieskataster des Kantons Z€urich (ed. Amt f€ur
Abfall Wasser Energie und Luft AWEL).
AWEL (2010) Neophytenkataster des Kantons Z€urich (ed.
Amt f€ur Abfall Wasser Energie und Luft AWEL) http://
www.gis.zh.ch/dokus/awel/abfallwirtschaft/neophyten.html
Beerling, D.J. & Perrins, J.M. (1993) Impatiens glandulifera
Royle (Impatiens roylei Walp.). Journal of Ecology, 81, 367–
382.
Besanko, D., Dranove, D., Shanley, M. & Schaefer, S. (2004)
Economics of strategy, 3rd edn. Wiley, New York, Chiches-
ter.
BFS & GEOSTAT (2000) BEK200 - Digitale Bodeneignungsk-
arte der Schweiz.
Bini, L.M., Diniz, J.A.F., Rangel, T. et al. (2009) Coefficient
shifts in geographical ecology: an empirical evaluation of
spatial and non-spatial regression. Ecography, 32, 193–204.
Blackwood, J., Hastings, A. & Costello, C. (2010) Cost-effec-
tive management of invasive species using linear-quadratic
control. Ecological Economics, 69, 519–527.
Borcard, D., Legendre, P. & Drapeau, P. (1992) Partialling
out the spatial component of ecological variation. Ecology,
73, 1045–1055.
Bossenbroek, J.M., Kraft, C.E. & Nekola, J.C. (2001) Predic-
tion of long-distance dispersal using gravity models: Zebra
mussel invasion of inland lakes. Ecological Applications, 11,
1778–1788.
Brabec, J. & Py�sek, P. (2000) Establishment and survival of
three invasive taxa of the genus Reynoutria (Polygonaceae)
in mesic mown meadows: a field experimental study. Folia
Geobotanica, 35, 27–42.
Bullock, J., Shea, K. & Skarpaas, O. (2006) Measuring plant
dispersal: an introduction to field methods and experimen-
tal design. Plant Ecology, 186, 217–234.
Burnham, K.P. & Anderson, D.R. (2002) Model selection and
multimodel inference: a practical information-theoretic
approach, 2nd edn. Springer, New York.
Campbell, G.S., Blackwell, P.G. & Woodward, F.I. (2002)
Can landscape-scale characteristics be used to predict plant
invasions along rivers? Journal of Biogeography, 29, 535–
543.
Cox, G.W. (2004) Alien species and evolution: the evolutionary
ecology of exotic plants, animals, microbes, and interacting
native species. Island Press, Washington, DC.
Damschen, E.I., Brudvig, L.A., Haddad, N.M., Levey, D.J.,
Orrock, J.L. & Tewksbury, J.J. (2008) The movement ecol-
ogy and dynamics of plant communities in fragmented
landscapes. Proceedings of the National Academy of Sciences
USA, 105, 19078–19083.
De Micheli, A., Bollens, U., Gelpke, G., Streit, B. & Fischer,
D. (2006) Bericht und Empfehlung zur Bek€ampfung des Jap-
ankn€oterichs. Kantone Aargau, Bern, Glarus, Luzern, Wallis
und Z€urich.
1040 Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd
E. S. Meier et al.

Dormann, C.F., McPherson, J.M., Ara�ujo, M.B., Bivand, R.,
Bolliger, J., Carl, G., Davies, R.G., Hirzel, A., Jetz, W., Kis-
sling, W.D., K€uhn, I., Ohlem€uller, R., Peres-Neto, P.R.,
Reineking, B., Schr€oder, B., Schurr, F.M. & Wilson, R.
(2007) Methods to account for spatial autocorrelation in
the analysis of species distributional data: a review. Ecogra-
phy, 30, 609–628.
Dray, S. (2008) SpacemakeR: spatial modelling. R package
version 0.0-3. Available at: http://cran.r-project.org
Dullinger, S., Kleinbauer, I., Peterseil, J., Smolik, M. & Essl,
F. (2009) Niche based distribution modelling of an invasive
alien plant: effects of population status, propagule pressure
and invasion history. Biological Invasions, 11, 2401–2414.
Dullinger, S., Gattringer, A., Thuiller, W. et al. (2012)
Extinction debt of high-mountain plants under twenty-
first-century climate change. Nature Climate Change, 2,
619–622.
EEA/SEBI (2010) Number of the listed ‘worst’ terrestrial and
freshwater invasive alien species threatening biodiversity in
Europe. European Environment Agency, Expert Group on
trends in invasive alien species.
Epanchin-Niell, R.S. & Hastings, A. (2010) Controlling estab-
lished invaders: integrating economics and spread dynam-
ics to determine optimal management. Ecology Letters, 13,
528–541.
Epanchin-Niell, R.S., Haight, R.G., Berec, L., Kean, J.M. &
Liebhold, A.M. (2012) Optimal surveillance and eradica-
tion of invasive species in heterogeneous landscapes. Ecol-
ogy Letters, 15, 803–812.
Flather, C.H. & Bevers, M. (2002) Patchy reaction-diffusion
and population abundance: the relative importance of hab-
itat amount and arrangement. The American Naturalist,
159, 40–56.
Gallien, L., Douzet, R., Pratte, S., Zimmermann, N.E. & Thu-
iller, W. (2012) Invasive species distribution models – how
violating the equilibrium assumption can create new
insights. Global Ecology and Biogeography, 21, 1126–1136.
Giljohann, K.M., Hauser, C.E., Williams, N.S.G. & Moore,
J.L. (2011) Optimizing invasive species control across
space: willow invasion management in the Australian Alps.
Journal of Applied Ecology, 48, 1286–1294.
Groves, J.H., Williams, D.G., Caley, P., Norris, R.H. & Cait-
cheon, G. (2009) Modelling of floating seed dispersal in a
fluvial environment. River Research and Applications, 25,
582–592.
Guisan, A. & Zimmermann, N.E. (2000) Predictive habitat
distribution models in ecology. Ecological Modelling, 135,
147–186.
Hauser, C.E. & McCarthy, M.A. (2009) Streamlining ‘search
and destroy’: cost-effective surveillance for invasive species
management. Ecology Letters, 12, 683–692.
Higgins, S.I., Richardson, D.M. & Cowling, R.M. (2000)
Using a dynamic landscape model for planning the man-
agement of alien plant invasions. Ecological Applications,
10, 1833–1848.
Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G. & Jarvis,
A. (2005) Very high resolution interpolated climate sur-
faces for global land areas. International Journal of Clima-
tology, 25, 1965–1978.
Hodkinson, D.J. & Thompson, K. (1997) Plant dispersal: the
role of man. Journal of Applied Ecology, 34, 1484–1496.
Hulme, P.E. (2003) Biological invasions: winning the science
battles but losing the conservation war? Oryx, 37, 178–193.
Hulme, P.E. (2006) Beyond control: wider implications for
the management of biological invasions. Journal of Applied
Ecology, 43, 835–847.
Ib�a~nez, I., Silander, J.A., Allen, J.M., Treanor, S.A. & Wilson,
A. (2009) Identifying hotspots for plant invasions and fore-
casting focal points of further spread. Journal of Applied
Ecology, 46, 1219–1228.
Katul, G.G., Porporato, A., Nathan, R., Siqueira, M., Soons,
M.B., Poggi, D., Horn, H.S. & Levin, S.A. (2005) Mecha-
nistic analytical models for long-distance seed dispersal by
wind. The American Naturalist, 166, 368–381.
Keller, R.P., Lodge, D.M., Lewis, M.A. & Shogren, J.F. (2009)
Bioeconomics of invasive species: integrating ecology, econom-
ics, policy, and management. Oxford University Press,
Oxford, UK.
Kot, M., Lewis, M.A. & van den Driessche, P. (1996) Dis-
persal data and the spread of invading organisms. Ecology,
77, 2027–2042.
Krug, R.M., Roura-Pascual, N. & Richardson, D.M. (2010)
Clearing of invasive alien plants under different budget sce-
narios: using a simulation model to test efficiency. Biologi-
cal Invasions, 12, 4099–4112.
Mack, R.N., Simberloff, D., Lonsdale, W.M., Evans, H.,
Clout, M. & Bazzaz, F.A. (2000) Biotic invasions: causes,
epidemiology, global consequences, and control. Ecological
Applications, 10, 689–710.
Mand�ak, B., Py�sek, P. & B�ımov�a, K. (2004) History of the
invasion and distribution of Reynoutria taxa in the Czech
Republic: a hybrid spreading faster than its parents. Preslia,
76, 15–64.
Marchetto, K.M., Jongejans, E., Shea, K. & Isard, S.A. (2010)
Plant spatial arrangement affects projected invasion speeds
of two invasive thistles. Oikos, 119, 1462–1468.
Masters, R.A. & Sheley, R.L. (2001) Principles and practices
for managing rangeland invasive plants. Journal of Range
Management, 54, 502–517.
McGarigal, K., Cushman, S.A., Neel, M.C. & Ene, E. (2002)
FRAGSTATS v3: spatial pattern analysis program for cate-
gorical maps. Computer software program produced by the
authors at the University of Massachusetts, Amherst.
Meier, E.S., Kienast, F., Pearman, P.B., Svenning, J.-C., Thu-
iller, W., Ara�ujo, M.B., Guisan, A. & Zimmermann, N.E.
(2010) Biotic and abiotic variables show little redundancy
in explaining tree species distributions. Ecography, 33,
1038–1048.
Meier, E.S., Lischke, H., Schmatz, D.R. & Zimmermann,
N.E. (2012) Climate, competition and connectivity affect
Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd 1041
Spatial management for controlling invasive plants

future migration and ranges of European trees. Global Ecol-
ogy and Biogeography, 21, 164–178.
Moody, M.E. & Mack, R.N. (1988) Controlling the spread of
plant invasions: the importance of nascent foci. Journal of
Applied Ecology, 25, 1009–1021.
Naeem, S., Knops, J.M.H., Tilman, D., Howe, K.M., Ken-
nedy, T. & Gale, S. (2000) Plant diversity increases resis-
tance to invasion in the absence of covarying extrinsic
factors. Oikos, 91, 97–108.
Naidoo, R., Balmford, A., Ferraro, P.J., Polasky, S., Ricketts,
T.H. & Rouget, M. (2006) Integrating economic costs into
conservation planning. Trends in Ecology & Evolution, 21,
681–687.
Neter, J., Wasserman, W. & Kutner, M.H. (1983) Applied lin-
ear regression models p. 547. Richard D. Irwin Inc., Home-
wood.
Nobis, M.P., Jaeger, J.A.G. & Zimmermann, N.E. (2009)
Neophyte species richness at the landscape scale under
urban sprawl and climate warming. Diversity and Distribu-
tions, 15, 928–939.
Petitpierre, B., Kueffer, C., Broennimann, O., Randin, C.,
Daehler, C. & Guisan, A. (2012) Climatic Niche shifts are
rare among terrestrial plant invaders. Science, 335, 1344–
1348.
Py�sek, P. & Prach, K. (1993) Plant invasions and the role of
riparian habiats: a comparison of four species alien to
central Europe. Journal of Biogeography, 20, 413–420.
Py�sek, P. & Prach, K. (1995) Invasion dynamics of Impatiens
glandulifera – A century of spreading reconstructed. Biolog-
ical Conservation, 74, 41–48.
Py�sek, P., Jaro�sik, V., M€ullerov�a, J., Pergl, J. & Wild,
J. (2008) Comparing the rate of invasion by Heracleum
mantegazzianum at continental, regional, and local scales.
Diversity and Distributions, 14, 355–363.
R Development Core Team (2011) R: a language and envi-
ronment for statistical computing. R Foundation for Statisti-
cal Computing, Vienna, Austria.
Ramula, S., Knight, T.M., Burns, J.H. & Buckley, Y.M. (2008)
General guidelines for invasive plant management based on
comparative demography of invasive and native plant popu-
lations. Journal of Applied Ecology, 45, 1124–1133.
Regan, T.J., McCarthy, M.A., Baxter, P.W.J., Panetta, F.D. &
Possingham, H.P. (2006) Optimal eradication: when to stop
looking for an invasive plant. Ecology Letters, 9, 759–766.
Richter, R., Dullinger, S., Essl, F., Leitner, M. & Vogl, G.
(2013a) How to account for habitat suitability in weed
management programmes? Biological invasions, 15, 657–
669.
Richter, R., Berger, U.E., Dullinger, S., Essl, F., Leitner, M.,
Smith, M. & Vogl, G. (2013b) Spread of invasive ragweed:
climate change, management and how to reduce allergy
costs. Journal of Applied Ecology, 50, 1422–1430.
Shea, K. & Kelly, D. (2004) Modeling for management of
invasive species: Musk thistle (Carduus nutans) in New
Zealand. Weed Technology, 18, 1338–1341.
Shea, K., Jongejans, E., Skarpaas, O., Kelly, D. & Sheppard,
A.W. (2010) Optimal management strategies to control
local population growth or population spread may not be
the same. Ecological Applications, 20, 1148–1161.
Smolik, M.G., Dullinger, S., Essl, F., Kleinbauer, I., Leitner,
M., Peterseil, J., Stadler, L.M. & Vogl, G. (2010) Integrating
species distribution models and interacting particle systems
to predict the spread of an invasive alien plant. Journal of
Biogeography, 37, 411–422.
Solow, A., Seymour, A., Beet, A. & Harris, S. (2008) The
untamed shrew: on the termination of an eradication pro-
gramme for an introduced species. Journal of Applied Ecol-
ogy, 45, 424–427.
Stohlgren, T.J. & Schnase, J.L. (2006) Risk analysis for
biological hazards: what we need to know about invasive
species. Risk Analysis, 26, 163–173.
Swisstopo (2003) DHM25 – Das digitale H€ohenmodell der
Schweiz.
Swisstopo (2008) VECTOR25 – Digitales Landschaftsmodell
der Schweiz.
Taylor, C.M. & Hastings, A. (2004) Finding optimal control
strategies for invasive species: a density-structured model for
Spartina alterniflora. Journal of Applied Ecology, 41, 1049–1057.
Thuiller, W., Richardson, D.M., Py�sek, P., Midgley, G.F.,
Hughes, G.O. & Rouget, M. (2005) Niche-based modelling
as a tool for predicting the risk of alien plant invasions at
a global scale. Global Change Biology, 11, 2234–2250.
Turner, M.G. (1989) Landscape ecology: the effect of pattern
on process. Annual Review of Ecology and Systematics, 20,
171–197.
Vil�a, M., Espinar, J.L., Hejda, M., Hulme, P.E., Jaro�s�ık, V.,
Maron, J.L., Pergl, J., Schaffner, U., Sun, Y. & Py�sek, P.
(2011) Ecological impacts of invasive alien plants: a
meta-analysis of their effects on species, communities and
ecosystems. Ecology Letters, 14, 702–708.
Wadsworth, R.A., Collingham, Y.C., Willis, S.G., Huntley,
B. & Hulme, P.E. (2000) Simulating the spread and man-
agement of alien riparian weeds: are they out of control?
Journal of Applied Ecology, 37, 28–38.
Wilson, J.R.U., Gairifo, C., Gibson, M.R. et al. (2011) Risk
assessment, eradication, and biological control: global
efforts to limit Australian acacia invasions. Diversity and
Distributions, 17, 1030–1046.
With, K.A. (2002) The landscape ecology of invasive spread.
Conservation Biology, 16, 1192–1203.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the
online version of this article:
Table S1 Traits of Heracleum mantegazzianum, Impatiens
glandulifera and Reynoutria japonica.
Table S2 Overview of colonization estimates of suitable
habitats.
1042 Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd
E. S. Meier et al.

Table S3 Overview of most effective management options
per species, spending level and management goal, compared
to most effective random management options.
Figure S1 Achieved effects for reducing spread, area, adults
and regeneration potential and required spending over
15 years of management.
BIOSKETCH
Eliane S. Meier is an ecologist at Agroscope Institute for
Sustainability Sciences ISS. Her research focuses on the
impact of local processes, such as species interactions, dis-
persal and landscape management, on the distribution of
species and biodiversity patterns in agricultural landscapes,
alpine and riparian ecosystems and forests.
Author contributions: E.S.M., D.B. and N.E.Z. designed the
research; E.S.M., K.H. and A.G. prepared and analysed the
data; E.S.M., S.D., K.H., N.E.Z. and D.B. wrote the paper.
Editor: Ingolf K€uhn
Diversity and Distributions, 20, 1029–1043, ª 2014 John Wiley & Sons Ltd 1043
Spatial management for controlling invasive plants