space and the parietal cortex - massachusetts institute of technology
TRANSCRIPT

Space and the parietal cortexMasud Husain1,2 and Parashkev Nachev2
1 Institute of Cognitive Neuroscience, University College, London, 17 Queen Square, London WC1N 3AR, UK2 Imperial College London, Charing Cross Hospital Campus, London W6 8RF, UK
Review TRENDS in Cognitive Sciences Vol.11 No.1
Current views of the parietal cortex have difficultyaccommodating the human inferior parietal lobe (IPL)within a simple dorsal versus ventral stream dichotomy.In humans, lesions of the right IPL often lead to syn-dromes such as hemispatial neglect that are seeminglyin accord with the proposal that this region has a crucialrole in spatial processing. However, recent imaging andlesion studies have revealed that inferior parietalregions have non-spatial functions, such as in sustain-ing attention, detecting salient events embedded in asequence of events and controlling attention over time.Here, we review these findings and show that spatialprocesses and the visual guidance of action are onlypart of the repertoire of parietal functions. Althoughsub-regions in the human superior parietal lobe andintraparietal sulcus contribute to vision-for-actionand spatial functions, more inferior parietal regionshave distinctly non-spatial attributes that are neitherconventionally ‘dorsal’ nor conventionally ‘ventral’ innature.
IntroductionStudies in monkeys originally led to the hypothesis thatthe dorsal visual stream of cortical pathways, extendingfrom the primary visual cortex to the posterior parietalcortex (PPC), has a special role in spatial perception,whereas the ventral stream to the temporal cortex has akey role in object perception [1]. Later, that view waschallenged and revised to the proposal that the dorsalstream and PPC have a crucial role in directing visuallyguided actions [2]. However, it is becoming increasinglyapparent that both these models have difficulty in captur-ing some aspects of human parietal function. In particular,recent findings have begun to question a role for the humaninferior parietal lobe (IPL) solely in spatial processes or inthe visual guidance of action – ‘vision-for-action’.
Here, we review emerging trends in this field andsuggest that a reconsideration of human parietal functionis necessary. Close inspection of data from functionalimaging and lesion studies reveals that parts of the humanIPL are involved in functions that do not involve visuallyguided action, shifts of spatial attention or spatial memory.Instead, sub-regions within the IPL seem to be involved inthe detection of salient new items embedded in a sequenceof events and in maintaining or controlling attention overtime. We start by considering the current unease thatexists over human IPL function.
Corresponding author: Husain, M. ([email protected]).Available online 28 November 2006.
www.sciencedirect.com 1364-6613/$ – see front matter � 2006 Elsevier Ltd. All rights reserve
Current models of inferior parietal lobe functionIn Goodale and Milner’s view, the dorsal ‘vision-for-action’system operates in real time to compute the absolutemetrics of the target and its position in the egocentricco-ordinates of the effector (eye or limb) [3]. Thus, accord-ing to this scheme, the dorsal stream delivers informationdirect to the motor system for immediate use for reaching,grasping or eye movements. By contrast, the ventralstream is dedicated to ‘vision-for-perception’, but mighthave a role in movement planning based on memory of theobject and its relationship to other items.
Such a distinction between dorsal and ventral systemsis supported particularly clearly by the results of lesions ofthe human superior parietal lobe (SPL; Figure 1), whichoften lead to optic ataxia or misreaching to visual targets[2,4]. The regions of the PPC that have proven far moredifficult to fit into any dorsal–ventral dichotomy are theIPL and nearby temporoparietal junction (TPJ). Milnerand Goodale acknowledged this from the outset; theyspeculated that these regions might be a high-level repre-sentational system fed largely by the ventral stream andsubserving perceptual awareness [2,4]. Their proposalwould be consistent with some aspects of hemispatialneglect, a syndrome that often follows lesions of the IPLor TPJ, particularly in the right hemisphere [5,6]. Indivi-duals with neglect often fail to be aware of contralesionalobjects (to the left for individuals with right parietaldamage), even when given unlimited time to explore theworld around them.
A different perspective on the IPL has been offered bythe scheme proposed by Rizzolatti and Matelli, who sug-gest that it might be better to consider the superior andinferior parts of the PPC as belonging to two differentstreams [7]. According to this model, the SPL is part ofa distinct ‘dorso-dorsal’ system dedicated to the onlinecontrol of action, whereas the IPL is part of the ‘ventro-dorsal’ stream that is necessary for action understandingand spatial perception. This model originated from struc-tural and functional considerations of the macaque brainbut it has also been applied to the human PPC. In fact, it ispossible that in the human brain, the ventro-dorsal systemis different in the two hemispheres because deficits ofaction control (in the form of ideomotor apraxia) are moreprominent after left-hemisphere lesions, whereas deficitssuch as hemispatial neglect are more frequent and severeafter right parietal damage.
A third recent view of the PPC also makes a distinctionbetween the SPL and more ventral regions in the PPC,specifically the TPJ, but this hypothesis concerns a dichot-omy that incorporates visual-attention functions more
d. doi:10.1016/j.tics.2006.10.011

Figure 1. Posterior parietal cortex of macaque monkey (left) and human (right). The human posterior parietal cortex (PPC) is divided by the intraparietal sulcus (IPS) into
two parts: the superior parietal lobe (SPL) and the inferior parietal lobe (IPL). The IPL consists of the angular gyrus (Ang) and supramarginal gyrus (Smg) and borders the
superior temporal gyrus (purple) at a region that is often referred to as the temporoparietal junction (TPJ). In macaques, the PPC also consists of an SPL (area 5) and an IPL
(areas 7a and 7b) but, according to Brodmann [71], the homologues of these macaque regions are all confined to the human SPL (yellow), so the IPL in humans consists of
novel cortical areas. Subsequent anatomists such as Bailey and von Bonin [72] disagreed with this scheme, considering the IPL to be similar across both species. It remains
to be established whether there are new functional sub-regions within the human IPL.
Figure 2. Functional imaging studies. (a) Meta-analysis of activations associated
with spatial shifts of attention in healthy individuals demonstrate activations in the
superior parietal lobe (SPL) and intraparietal sulcus (IPS), plus dorsolateral frontal
regions (different colours correspond to findings from different studies). Similar
regions are also activated when participants perform spatial working memory
tasks. Adapted, with permission, from Ref. [8], � (2002) MacMillan Publishers Ltd.
(b) By contrast, regions in the inferior parietal lobe (IPL) and IPS, together with
more ventral frontal regions, are activated by salient events (cyan), sustained
attention (red) and non-spatial selective attention protocols (yellow) such as the
attentional-blink paradigm. Adapted from Ref. [18].
Review TRENDS in Cognitive Sciences Vol.11 No.1 31
directly. Corbetta and Shulman [8] argue that the SPL andparts of the intraparietal sulcus (IPS) have a role indirecting visual attention ‘top-down’ to locations or objectsin the scene and in selecting responses of effectors (eye orlimb). By contrast, the right TPJ acts ‘as a circuit breaker’,for example, by reorienting spatial attention when it hasbeen deployed incorrectly [8]. All these models provideimportant syntheses of a large body of data that hasemerged from studies of monkey and human PPC, butthere are other findings, which we consider later, thatare still not easily incorporated into these frameworks.
Functional imaging reveals non-spatial functionsNeuroimaging investigations in humans have consistentlydemonstrated that parts of the SPL or IPS are activated bytasks such as shifting spatial attention, engaging spatialworking memory, making saccadic eye movements orreaching to a visual target [8–13] (Figure 2a). In addition,more recent reports have demonstrated evidence in SPLand IPS for topographical representations of contralateralspace for saccades to remembered locations [14,15] or inremapping spatial locations across saccades [16,17]. Notethat the SPL and IPS activations are often associated withsimultaneous activity in the dorsolateral frontal lobe(Figure 2a), in accord with the view that the SPL is partof a dorsal frontoparietal system for directing spatialattention or action. Thus these neuroimaging findingsregarding the SPL and parts of the IPSwould be consistentwith many aspects of current models of PPC function.
However, the results of several other functional imagingstudies relating to the IPL and TPJ are very different. Theydemonstrate that these areas – and more ventral frontalregions – are consistently activated when healthy indivi-duals perform non-spatial tasks [18] (Figure 2b). They havebeen identified to be active when subjects maintain vigilantattention [9,19–22] orwhen they are presented infrequentlywithunusual, salient stimuli ina repetitive stream,as in the‘oddball’ paradigm [23–26], even when no spatial shifts of
www.sciencedirect.com

32 Review TRENDS in Cognitive Sciences Vol.11 No.1
attention, eye or limb movements are required. Moreover,these effects occur in visual, auditory or somatosensorytasks and are therefore not modality specific. Some parietalareas in or near to the IPS, togetherwith frontal regions, arealso activated when healthy subjects perform non-spatial,selective attention tasks [27–30] such as the attentional-blink protocol, a paradigm that allows measurement of thedynamic capacity of visual attention when stimuli are pre-sented sequentially at one spatial location. Thus regions inthe IPL, TPJ and parts of the IPS form a ventral frontopar-ietal circuit that seems very different from the dorsal onethat isactive in spatial perceptual, attentional,mnemonic oraction tasks (Figure 2b).
These neuroimaging findings suggest both spatial andnon-spatial functions are represented in the human PPC,butwith a gradient extending fromSPL through IPS to IPLof spatial to non-spatial functions [18] and some intermedi-ate regions in the IPS showing activation on both spatialand non-spatial tasks [28]. Corbetta and Shulman’s modelproposes a slightly different dichotomy between superiorand inferior parietal structures, with the TPJ involved indirecting attention to salient events, for example, whenobservers have to redeploy attention to an unexpectedperipheral stimulus [8,31]. However, in our view, theIPL and TPJ also play a more general, non-spatial rolein sustaining attention over time [18] that is not capturedby the Corbetta and Shulman model. New data fromtranscranial magnetic stimulation also support the viewthat this region might have a role in non-spatial encodingof salient stimuli [32], as do a range of findings from lesionsof the human PPC, which we discuss next.
Hemispatial, spatial and non-spatial deficitsIt has often been suggested that some components ofneglect syndrome, the characteristic clinical disorder fol-lowing right IPL damage, might best be understood using aspatial conceptual framework; for example, a lack of spaceexploration to one side of the body midline, to one side ofthe head axis or to one side of the direction of gaze. In fact,there is evidence for all three egocentric spatial referenceframes modulating the extent of neglect [33]. But thedisorder need not be strictly hemispatial – that is,neglected and non-neglected space might not be demar-cated by an abrupt plane that transects space in anyegocentric coordinate frame.
Kinsbourne suggested instead that each hemisphereorients attention towards the opposite side of space, witha stronger ‘vector’ exerted by left-hemisphere systems [34].Hence a strong rightward directional bias would beexpected after unilateral right parietal damage (becauseof unopposed left-hemisphere activity). The model predictsa gradient, rather than a sharp cut-off, in the distributionof visual attention from right to left – a proposal supportedby findings of a gradient of response times to stimulipresented at different spatial locations in neglect patients[35]. This model also shows that the unilateral nature ofneglect might emerge from a directional bias in just onecomponent of the syndrome; other components thatcontribute to neglect need not be directionally lateralized.
The key feature of Kinsbourne’s directional theory is thatit predicts hyperattention (i.e. better performance) to the
www.sciencedirect.com
right. By contrast, the hypothesis advanced by Heilmanet al. is slightly different, suggesting that there might besome deficits on the right (‘good’) in addition to the left(neglected) side of space [36]. The authors proposed that,although the right PPC might normally direct attention toboth sides of space, the left PPC directs attention only to theright hemispace. One implication of this model is that rightPPC damage might be associated with not only a severedeficit for itemsto the left, butalsoamilderdeficit for stimulito the right [37]. Recent studies show that there are indeeddeficits on the right side of space, but they are not necessa-rily milder than on the left.
Firstly, Duncan et al. have demonstrated reducedvisual processing and short-termmemory capacity, whichcan be equivalent in severity in left and right hemifields, inIPL patients who have varying degrees of neglect [38].Importantly, a subsequent study of non-neglect patientsrevealed that IPL- and TPJ-lesioned individuals weremore severely affected than those with SPL involvement[39]. Secondly, processing of visual information from theright visual field – but not the left – might be unselective,with information that is irrelevant to the task beinginappropriately prioritized in neglect patients [40].Thirdly, attention to transient onsets and offsets of visualstimuli is disrupted bilaterally in right IPL patients [41].Fourthly, detection of briefly presented stimuli isimpaired in both visual hemifields, but worse to the left[42]. Finally, spatial functions such as keeping track ofobject locations over intervening saccades or awareness ofchanges in their locationmight all be severely disrupted inpatients with right-IPL damage, even on the right side ofspace [43–46]. All these studies show that damage to theright IPLmight lead to deficits that are not confined to onehemispace.
Other investigations have revealed that parietaldamage can lead to deficits on non-spatial tasks even whenstimuli are presented at only one spatial location. Theattentional-blink protocol, which we discussed previouslyin the context of functional imaging studies, has also beenemployed in patients to index the dynamic control of visualattention when stimuli are presented sequentially at fixa-tion [47,48]. These investigations have demonstrated thatindividuals with IPL lesions – including those with spatialneglect – are dramatically impaired in visual-processingability, even when attention does not have to be shiftedacross space. By contrast, lesions of the SPL do not lead toattentional-blink deficits [48]. A second series of studieshas focused on tests of the ability to maintain vigilantattention. Resection of the right IPL leads to impairment inthe ability to sustain visual attention over prolonged inter-vals [49]. In addition, right-hemisphere neglect patientswho have parietal involvement are also impaired at main-taining vigilant attention on non-spatial tasks, regardlessof whether the stimuli are auditory or visual [50,51].
This wide range of findingsmakes it difficult to sustain asimple ‘spatial’ or indeed ‘hemispatial’ account for neglect.Instead, these results reveal that deficits occur on bothsides of space in the syndrome; some of them might bespatial in nature, but evidently others are not. The datasuggest that a combination of spatial and non-spatialimpairments exists in neglect [18], and this might explain
Review TRENDS in Cognitive Sciences Vol.11 No.1 33
why it has been difficult to frame the syndrome in terms ofany dorsal–ventral dichotomy (discussed in Ref. [4]).
Human and monkey parietal cortex might not be thesameOur review of the lesion and imaging data in humanssuggests the SPLmight have a key role in spatial functionsand vision-for-action. We consider this part of the humanparietal cortex to have strong similarities to the monkeyPPC. By contrast, some regions within the human IPLhave non-spatial functions that do not map easily to the‘dorsal’ stream (see also Refs [2,4]). In fact,monkey parietalcortex might not be a complete model for the human PPC.In our opinion and that of Milner [52], there is no goodevidence for a long-lasting and severe neglect syndrome inmonkeys such as that seen in humans [53]. Althoughparietal-cortex or white-matter damage in macaque mon-keys leads to various impairments in contralateral space,there is no description of their everyday behaviour, in ourview, that equals the profound deficits observed in humansby clinicians. Crucially, there is also no evidence for hemi-spheric differences for neglect-like symptoms in any mon-key model, whereas in humans neglect is far more commonand prominent after right-sided damage.
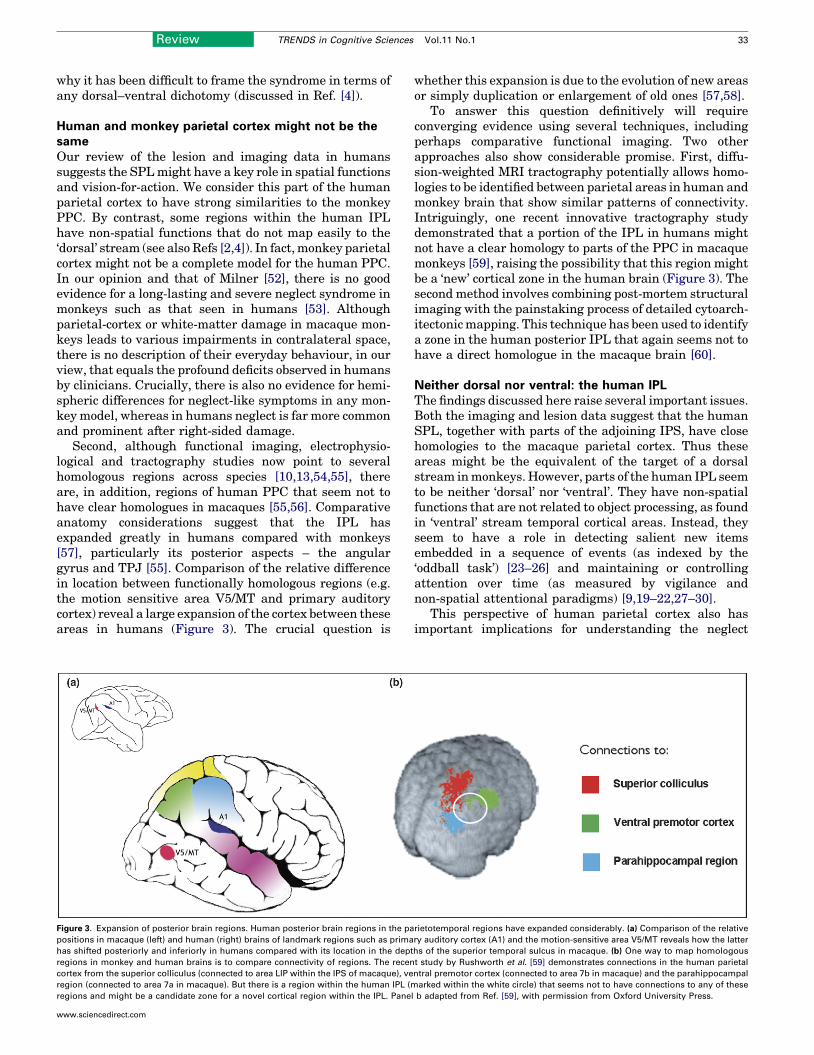
Second, although functional imaging, electrophysio-logical and tractography studies now point to severalhomologous regions across species [10,13,54,55], thereare, in addition, regions of human PPC that seem not tohave clear homologues in macaques [55,56]. Comparativeanatomy considerations suggest that the IPL hasexpanded greatly in humans compared with monkeys[57], particularly its posterior aspects – the angulargyrus and TPJ [55]. Comparison of the relative differencein location between functionally homologous regions (e.g.the motion sensitive area V5/MT and primary auditorycortex) reveal a large expansion of the cortex between theseareas in humans (Figure 3). The crucial question is
Figure 3. Expansion of posterior brain regions. Human posterior brain regions in the pa
positions in macaque (left) and human (right) brains of landmark regions such as prima
has shifted posteriorly and inferiorly in humans compared with its location in the dept
regions in monkey and human brains is to compare connectivity of regions. The recen
cortex from the superior colliculus (connected to area LIP within the IPS of macaque), ve
region (connected to area 7a in macaque). But there is a region within the human IPL (m
regions and might be a candidate zone for a novel cortical region within the IPL. Pane
www.sciencedirect.com
whether this expansion is due to the evolution of new areasor simply duplication or enlargement of old ones [57,58].
To answer this question definitively will requireconverging evidence using several techniques, includingperhaps comparative functional imaging. Two otherapproaches also show considerable promise. First, diffu-sion-weighted MRI tractography potentially allows homo-logies to be identified between parietal areas in human andmonkey brain that show similar patterns of connectivity.Intriguingly, one recent innovative tractography studydemonstrated that a portion of the IPL in humans mightnot have a clear homology to parts of the PPC in macaquemonkeys [59], raising the possibility that this region mightbe a ‘new’ cortical zone in the human brain (Figure 3). Thesecond method involves combining post-mortem structuralimaging with the painstaking process of detailed cytoarch-itectonicmapping. This technique has been used to identifya zone in the human posterior IPL that again seems not tohave a direct homologue in the macaque brain [60].
Neither dorsal nor ventral: the human IPLThe findings discussed here raise several important issues.Both the imaging and lesion data suggest that the humanSPL, together with parts of the adjoining IPS, have closehomologies to the macaque parietal cortex. Thus theseareas might be the equivalent of the target of a dorsalstream inmonkeys. However, parts of the human IPL seemto be neither ‘dorsal’ nor ‘ventral’. They have non-spatialfunctions that are not related to object processing, as foundin ‘ventral’ stream temporal cortical areas. Instead, theyseem to have a role in detecting salient new itemsembedded in a sequence of events (as indexed by the‘oddball task’) [23–26] and maintaining or controllingattention over time (as measured by vigilance andnon-spatial attentional paradigms) [9,19–22,27–30].
This perspective of human parietal cortex also hasimportant implications for understanding the neglect
rietotemporal regions have expanded considerably. (a) Comparison of the relative
ry auditory cortex (A1) and the motion-sensitive area V5/MT reveals how the latter
hs of the superior temporal sulcus in macaque. (b) One way to map homologous
t study by Rushworth et al. [59] demonstrates connections in the human parietal
ntral premotor cortex (connected to area 7b in macaque) and the parahippocampal
arked within the white circle) that seems not to have connections to any of these
l b adapted from Ref. [59], with permission from Oxford University Press.

Box 1. Anatomy of neglect
The anatomical locus of neglect has recently become the subject of
controversy. The debate has centred on evidence derived from
patients with focal brain lesions, usually caused by stroke in the
territory of the middle cerebral artery (MCA). The region most
consistently implicated is the right IPL [5,6]. Recently, an alternative
crucial locus has been proposed in the superior temporal gyrus (STG)
[65,66]. The disparity between this new proposal and the results of
other studies has attracted considerable debate and has been
attributed to differences in clinical selection criteria, quality of
imaging and the methods of ensuring comparisons between homo-
logous structures across subjects. However, the size of the difference
between studies is too large to be accounted for by these factors
alone.
It is probable that a single locus for neglect is implausible. The
syndrome is functionally heterogeneous, incorporating several com-
ponent deficits [18,33,51] that are likely to result from disruption of
circuits distributed beyond the parietal lobe. Indeed, neglect might
occur following damage to lateral frontal regions, consistent with the
known frontoparietal circuits revealed by functional imaging in
healthy individuals (Figure 2). The syndrome also follows lesions to
subcortical structures and even in patients with strokes in the
distribution of the posterior cerebral artery (PCA) who do not have
direct involvement of lateral parietal or frontal regions [6].
A contributory factor in such cases might be the interruption in
functional connectivity between regions: such diaschetic effects are
suggested by the correlation between neglect and focal cortical
hypoperfusion following subcortical lesions [67]. More directly,
damage to frontoparietal fibres has also been shown to be associated
with neglect [62,68] whereas lesions affecting the splenium or
connections from the parahippocampal region to the parietal cortex
have been shown to be predictive of neglect following PCA stroke
[6,69,70]. Patients who have chronic neglect are more likely to have
involvement of multiple cortical areas, which reinforces this view.
The anatomy of neglect revealed by these studies suggests a
complexity that is hard to capture adequately by current lesion-
mapping techniques. Most methods rely on performing a statistical
test independently at each inter-subject aligned voxel, without any
attempt to capture correlations across different brain regions.
Because the structure of the vascular tree introduces a marked
inhomogeneity in the shape and distribution of stroke lesions, this
approach is inherently insecure. Indeed, the only global feature of the
lesion commonly modelled – overall lesion size – will inevitably
distort any spatial inference because of the strong inhomogeneity in
lesion volume across the brain (Figure I). Further progress in this field
will have to await the introduction of novel multivariate methods.
However, the investigation of functions that might have no
counterpart in non-human primates presents a special challenge. In
the absence of detailed neurophysiology and experimental lesion
studies, there is little to constrain speculation based on the abundant –
yet purely correlational – data generated by functional imaging. It is,
therefore, imperative that lesion-mapping methods are developed that
match the ease and flexibility of functional imaging and can be widely
used to test the predictions generated from it. Only by considering
the results of studies on healthy brains in combination with the effects
of lesions in patients is it possible to make strong inferences about
the functional specialization of regions within the human parietal
cortex.
Figure I. Spatial inhomogeneity of mean lesion volume. Voxel-wise map of the
rank correlation between total lesion volume and probability of damage, given a
clinical and radiological diagnosis of stroke, derived from a sample of 456
patients. Only voxels affected in eight or more patients are shown. Note the
strong centrifugal gradient of mean lesion size, with more peripheral (cortical)
voxels being involved in larger lesions. Scale gives value of Spearman’s rho.
34 Review TRENDS in Cognitive Sciences Vol.11 No.1
syndrome [18]. Recent studies of the anatomy of neglecthave been controversial (Box 1). Although the argumenthas often focussed on the contribution of single brainregions it is evident that most patients have large lesionsthat extend over several crucial areas. Moreover, the syn-drome is heterogeneous, with different patients havingdifferent combinations of cognitive deficit [51]. Many indi-viduals with neglect have brain damage that spans thecortex of the SPL, IPS and IPL [6,51,61] and the underlyingwhite matter [62]. Given what we know of the functions ofthese parietal regions, one would expect such damage tolead to a combination of spatial and non-spatial deficits inneglect patients [18], and this is exactly what has beenfound with careful testing using a battery of spatial andnon-spatial tasks [51].
New experimental approaches to the neglect syndrome,using a combination of high-resolution anatomy and beha-vioural tests designed to assess spatial or non-spatialfunctions, have begun to show how the heterogeneity inthe syndromemight be explained in this way. For example,
www.sciencedirect.com
those neglect patients who have posterior damage and adeficit in keeping track of spatial locations across saccadeshave lesions that include a small zone in the IPS [43]. Bycontrast, neglect patients in which the posterior cortex islesioned but this small area of the IPS is spared do notshow this deficit. Similarly, lesions of the right TPJ – butnot the SPL – lead to impairments in responding to rare,salient events such as when a target is presented at anunexpected location [63], whereas damage near the IPS isassociated with interference from right-sided irrelevantdistractors [61]. The interaction of spatial and non-spatialfactors, and the precise combination of these, might becrucial in determining the manifestations of neglect acrossdifferent patients [18].
Concluding remarksCurrent models of the parietal cortex have difficulties incapturing elements of human IPL function. Our reviewpoints to the conclusion that parts of the human IPL do notfit a role in spatial processing or vision-for-action, as might

Box 2. Questions for future research
� Just as novel parietal regions might have emerged in humans,
might there also be specializations in monkey parietal regions that
are not present in man (e.g. related to the special visuomotor
abilities that might be required for an arboreal existence)?
� What are the differences in underlying cognitive deficits between
neglect following left- and right-hemisphere lesions? For exam-
ple, the data reviewed here would suggest that major deficits in
sustained attention would not be expected following left-IPL
lesions. Better characterisation of neglect following left-hemi-
sphere lesions would be an important step forward.
� Although several dissociable behavioural components of neglect
have now been clearly identified, their corresponding anatomical
loci have proved challenging to establish. Component deficits
might be masked or confounded by interactions between different
components. Similarly, a faithful analysis of the underlying
anatomy might require lesion-mapping methods that capture
correlations between different anatomical areas. Can such richly
multivariate analyses be successfully performed with the neces-
sarily constrained datasets used in human lesion-mapping
research?
� The fine functional parcellation of the cortex achieved with fMRI is
yet to be matched by a correspondingly detailed map of the
connections between remote areas. Therefore, the behavioural
effects of white-matter lesions are hard to relate to the underlying
functional architecture. Would the development of a detailed
standardized map of white-matter tracts improve our under-
standing of the consequences of damage that involves parietal
grey- and white-matter regions?
Review TRENDS in Cognitive Sciences Vol.11 No.1 35
be assumed from considerations of the monkey PPC.Rather, some regions in the human IPL seem to participatein the detection of salient new items embedded in a rapidsequence of events and inmaintaining or controlling atten-tion over time. This view of parietal cortex can also accom-modate findings that demonstrate both homologous andnon-homologous sub-regions of parietal cortex in macaquemonkeys and humans (Figure 3). Monkeys with parietallesions might not demonstrate the full-blown neglect syn-drome observed in humans because they do not have thecomplement of spatial and non-spatial deficits that arecommon following human lesions [18].
In addition, hemispheric differences between left andright IPL are an important feature of the human parietalcortex. Whereas right IPL and TPJ lesions commonly leadto neglect, damage to the homologous region in the lefthemisphere in humans often leads to the syndrome ofapraxia [64]. However, some left IPL patients might alsoshow evidence of right-sided neglect. Clearly, the precisedistinctions in function between left and right IPL remainto be fully established (see Box 2 for Questions for futureresearch). But it is evident that such substantial differ-ences in function across the hemispheres has not beenobserved in monkeys, demonstrating the need to be cau-tious when making extrapolations from monkey studies tohuman parietal cortex.
AcknowledgementsOur research is funded by The Wellcome Trust.
References1 Mishkin, M. et al. (1983) Object vision and spatial vision: two cortical
pathways. Trends Neurosci. 6, 414–417
www.sciencedirect.com
2 Milner, A.D. and Goodale, M.A. (1995) The Visual Brain in Action,Oxford University Press
3 Goodale, M.A. et al. (2004) Two distinct modes of control for object-directed action. Prog. Brain Res. 144, 131–144
4 Milner, A.D. (1997) Neglect, extinction, and the cortical streams ofvisual processing. In Parietal Lobe Contributions to Orientation in 3DSpace (Thier, P. and Karnath, H-O., eds), pp. 3–22, Springer-Verlag
5 Vallar, G. and Perani, D. (1986) The anatomy of unilateral neglect afterright-hemisphere stroke lesions. A clinical/CT-scan correlation study inman. Neuropsychologia 24, 609–622
6 Mort, D.J. et al. (2003) The anatomy of visual neglect. Brain 126, 1986–1997
7 Rizzolatti, G. and Matelli, M. (2003) Two different streams form thedorsal visual system: anatomy and functions.Exp. Brain Res. 153, 146–157
8 Corbetta, M. and Shulman, G.L. (2002) Control of goal-directed andstimulus-driven attention in the brain. Nat. Rev. Neurosci. 3, 215–229
9 Vandenberghe, R. et al. (2001) Functional specificity of superiorparietal mediation of spatial shifting. Neuroimage 14, 661–673
10 Culham, J.C. and Valyear, K.F. (2006) Human parietal cortex in action.Curr. Opin. Neurobiol. 16, 205–212
11 Connolly, J.D. et al. (2003) FMRI evidence for a ‘parietal reach region’in the human brain. Exp. Brain Res. 153, 140–145
12 LaBar, K.S. et al. (1999) Neuroanatomic overlap of working memoryand spatial attention networks: a functional MRI comparison withinsubjects. Neuroimage 10, 695–704
13 Culham, J.C. et al. (2006) The role of parietal cortex in visuomotorcontrol: what have we learned from neuroimaging? Neuropsychologia44, 2668–2684
14 Sereno, M.I. et al. (2001) Mapping of contralateral space in retinotopiccoordinates by a parietal cortical area in humans. Science 294, 1350–1354
15 Schluppeck, D. et al. (2006) Sustained activity in topographic areas ofhuman posterior parietal cortex during memory-guided saccades.J. Neurosci. 26, 5098–5108
16 Medendorp, W.P. et al. (2003) Gaze-centered updating of visual spacein human parietal cortex. J. Neurosci. 23, 6209–6214
17 Merriam, E.P. et al. (2003) Spatial updating in human parietal cortex.Neuron 39, 361–373
18 Husain, M. and Rorden, C. (2003) Non-spatially lateralizedmechanisms in hemispatial neglect. Nat. Rev. Neurosci. 4, 26–36
19 Pardo, J.V. et al. (1991) Localization of a human system for sustainedattention by positron emission tomography. Nature 349, 61–64
20 Sturm, W. et al. (1999) Functional anatomy of intrinsic alertness:evidence for a fronto-parietal-thalamic-brainstem network in theright hemisphere. Neuropsychologia 37, 797–805
21 Manly, T. et al. (2003) Enhancing the sensitivity of a sustainedattention task to frontal damage: convergent clinical and functionalimaging evidence. Neurocase 9, 340–349
22 Adler, C.M. et al. (2001) Changes in neuronal activation withincreasing attention demand in healthy volunteers: an fMRI study.Synapse 42, 266–272
23 Downar, J. et al. (2002) A cortical network sensitive to stimulussalience in a neutral behavioral context across multiple sensorymodalities. J. Neurophysiol. 87, 615–620
24 Clark, V.P. et al. (2000) Responses to rare visual target and distractorstimuli using event-related fMRI. J. Neurophysiol. 83, 3133–3139
25 Marois, R. et al. (2000) A stimulus-driven approach to object identityand location processing in the human brain. Neuron 25, 717–728
26 Linden, D.E. et al. (1999) The functional neuroanatomy of targetdetection: an fMRI study of visual and auditory oddball tasks.Cereb. Cortex 9, 815–823
27 Coull, J.T. and Frith, C.D. (1998) Differential activation of rightsuperior parietal cortex and intraparietal sulcus by spatial andnonspatial attention. Neuroimage 8, 176–187
28 Wojciulik, E. and Kanwisher, N. (1999) The generality of parietalinvolvement in visual attention. Neuron 23, 747–764
29 Marois, R. et al. (2000) Neural correlates of the attentional blink.Neuron 28, 299–308
30 Coull, J.T. and Nobre, A.C. (1998) Where and when to pay attention:the neural systems for directing attention to spatial locations and totime intervals as revealed by both PET and fMRI. J. Neurosci. 18,7426–7435

36 Review TRENDS in Cognitive Sciences Vol.11 No.1
31 Chambers, C.D. et al. (2004) Fast and slow parietal pathways mediatespatial attention. Nat. Neurosci. 7, 217–218
32 Mevorach, C. et al. (2006) Opposite biases in salience-based selectionfor the left and right posterior parietal cortex. Nat. Neurosci. 9, 740–742
33 Behrmann, M. and Geng, J.J. (2002) What is ‘left’ when all is said anddone? Spatial coding and hemispatial neglect. In The Cognitive andNeural Bases of Spatial Neglect (Karnath, H-O. et al., eds), pp. 85–100,Oxford University Press
34 Kinsbourne, M. (1993) Orientational bias model of unilateral neglect:evidence from attentional gradients within hemispace. In UnilateralNeglect: Clinical and Experimental Studies (Robertson, I.H. andMarshall, J.C., eds), pp. 63–86, Lawrence Erlbaum
35 Smania, N. et al. (1998) The spatial distribution of visual attention inhemineglect and extinction patients. Brain 121, 1759–1770
36 Heilman, K.M. and Van Den Abell, T. (1980) Right hemispheredominance for attention: the mechanism underlying hemisphericasymmetries of inattention (neglect). Neurology 30, 327–330
37 Bartolomeo, P. and Chokron, S. (1999) Left unilateral neglect or righthyperattention? Neurology 53, 2023–2027
38 Duncan, J. et al. (1999) Systematic analysis of deficits in visualattention. J. Exp. Psychol. Gen. 128, 450–478
39 Peers, P.V. et al. (2005) Attentional functions of parietal and frontalcortex. Cereb. Cortex 15, 1469–1484
40 Snow, J.C. and Mattingley, J.B. (2006) Goal-driven selective attentionin patients with right hemisphere lesions: how intact is the ipsilesionalfield? Brain 129, 168–181
41 Battelli, L. et al. (2003) Bilateral deficits of transient visual attention inright parietal patients. Brain 126, 2164–2174
42 Russell, C. et al. (2004) Attention modulates the visual field in healthyobservers and parietal patients. Neuroreport 15, 2189–2193
43 Mannan, S. et al. (2005) Revisiting previously searched locations invisual neglect: role of right parietal and frontal lesions in misjudgingold locations as new. J. Cogn. Neurosci. 17, 340–354
44 Pisella, L. et al. (2004) Impaired working memory for location but notfor colour or shape in visual neglect: a comparison of parietal and non-parietal lesions. Cortex 40, 379–390
45 Parton, A. et al. (2006) Space re-exploration in hemispatial neglect.Neuroreport 17, 833–836
46 Heide, W. et al. (1995) Cortical control of double-step saccades:implications for spatial orientation. Ann. Neurol. 38, 739–748
47 Husain, M. et al. (1997) Abnormal temporal dynamics of visualattention in spatial neglect patients. Nature 385, 154–156
48 Shapiro, K. et al. (2002) Control of visuotemporal attention byinferior parietal and superior temporal cortex. Curr. Biol. 12, 1320–1325
49 Rueckert, L. and Grafman, J. (1998) Sustained attention deficits inpatients with lesions of posterior cortex.Neuropsychologia 36, 653–660
50 Robertson, I.H. et al. (1997) Auditory sustained attention is amarker ofunilateral spatial neglect. Neuropsychologia 35, 1527–1532
51 Buxbaum, L.J. et al. (2004) Hemispatial neglect: subtypes,neuroanatomy, and disability. Neurology 62, 749–756
Endeav
Coming soon in the quarterly magazine for
Earthquake theories in the early mScience in fiction - attempts to make a scie
The birth of botanical Dro
Endeavour is available on Science
www.sciencedirect.com
52 Milner, A.D. (1987) Animal models for the syndrome of spatial neglectin humans. In Neurophysiological and Neuropsychological Aspects ofSpatial Neglect (Jeannerod, M., ed.), pp. 259–288, North-Holland
53 Lynch, J.C. and McLaren, J.W. (1989) Deficits of visual attention andsaccadic eye movements after lesions of parietooccipital cortex inmonkeys. J. Neurophysiol. 61, 74–90
54 Grefkes, C. and Fink, G.R. (2005) The functional organization of theintraparietal sulcus in humans and monkeys. J. Anat. 207, 3–17
55 Orban, G.A. et al. (2004) Comparative mapping of higher visual areasin monkeys and humans. Trends Cogn. Sci. 8, 315–324
56 Orban, G. et al. (2006) Mapping the parietal cortex of human and non-human primates. Neuropsychologia 44, 2647–2667
57 Zilles, K. and Palomero-Gallagher, N. (2001) Cyto-, myelo-, andreceptor architectonics of the human parietal cortex. Neuroimage14, S8–S20
58 Sereno, M.I. and Tootell, R.B. (2005) From monkeys to humans: whatdo we now know about brain homologies? Curr. Opin. Neurobiol. 15,135–144
59 Rushworth, M.F. et al. (2006) Connection patterns distinguish 3regions of human parietal cortex. Cereb. Cortex 16, 1418–1430
60 Caspers, S. et al. (2006) The human inferior parietal cortex:cytoarchitectonic parcellation and interindividual variability.Neuroimage 33, 430–448
61 Vandenberghe, R. et al. (2005) Attentional responses to unattendedstimuli in human parietal cortex. Brain 128, 2843–2857
62 Doricchi, F. and Tomaiuolo, F. (2003) The anatomy of neglect withouthemianopia: a key role for parietal–frontal disconnection?Neuroreport14, 2239–2243
63 Friedrich, F.J. et al. (1998) Spatial attention deficits in humans: acomparison of superior parietal and temporal-parietal junction lesions.Neuropsychology 12, 193–207
64 Goldenberg, G. and Karnath, H-O. (2006) The neural basis of imitationis body part specific. J. Neurosci. 26, 6282–6287
65 Karnath, H-O. et al. (2001) Spatial awareness is a function of thetemporal not the posterior parietal lobe. Nature 411, 950–953
66 Karnath, H-O. et al. (2004) The anatomy of spatial neglect based onvoxelwise statistical analysis: a study of 140 patients.Cereb. Cortex 14,1164–1172
67 Hillis, A.E. et al. (2005) Anatomy of spatial attention: insights fromperfusion imaging and hemispatial neglect in acute stroke. J. Neurosci.25, 3161–3167
68 Thiebaut de Schotten, M. et al. (2005) Direct evidence for a parietal-frontal pathway subserving spatial awareness in humans. Science 309,2226–2228
69 Park, K.C. et al. (2006) Deafferentation–disconnection neglect inducedby posterior cerebral artery infarction. Neurology 66, 56–61
70 Bird, C.M. et al. (2006) Visual neglect after right posterior cerebralartery infarction. J. Neurol. Neurosurg. Psychiatry 77, 1008–1012
71 Brodmann, K. (1909) Vergleichende Lokalisationslehre derGroßhirnrinde, Barth
72 Bailey, P. and von Bonin, G. (1951) The Isocortex of Man, University ofIllinois Press
our
the history and philosophy of science:
odern period by F. Willmoth
nce out of literary criticism by J. Adams
sophila by S. Leonelli
Direct, www.sciencedirect.com