sources of the scalp-recorded amplitude-modulation ... · j am acad audiol 13: 188-204 (2002)...
TRANSCRIPT

J Am Acad Audiol 13 : 188-204 (2002)
Sources of the Scalp-Recorded Amplitude-Modulation Following Response Shigeyuki Kuwada* Julia S. Andersont Ranjan Batrat Douglas C. Fitzpatrick§ Natacha Teissier* William R. D'Angelo*
Abstract
The scalp-recorded amplitude-modulation following response (AMFR),, is gaining recognition as an objective audiometric tool, but little is known about the neural sources that underlie this poten-tial . We hypothesized, based on our human studies and single-unit recordings in animals, that the scalp-recorded AMFR reflects the interaction of multiple sources . We tested this hypothesis using an animal model, the unanesthetized rabbit. We compared AMFRs recorded from the surface of the brain at different locations and before and after the administration of agents likely to enhance or suppress neural generators . We also recorded AMFRs locally at several stations along the audi-tory neuraxis. We conclude that the surface-recorded AMFR is indeed a composite response from multiple brain generators . Although the response at any modulation frequency can reflect the activ-ity of more than one generator, the AMFRs to low and high modulation frequencies appear to reflect a strong contribution from cortical and subcortical sources, respectively .
Key Words: Amplitude-modulation following response, auditory steady-state potentials, electrical audiometry, evoked potentials, temporal coding
Abbreviations : AMFR = amplitude-modulation following response ; ASSR = auditory steady-state response ; EFR = envelope following response ; GABA = y-aminobutyric acid ; HCI = hydrochloride ; KCI = potassium chloride ; MTF = modulation transfer function ; SAM = sinusoidally amplitude-modulated ; SSEP = auditory steady-state evoked potential
Sumario
Las respuestas de seguimiento de amplitud modulada (AMFR)II registradas en el craneo estan ganando reconocimiento como una herramiento audiometrica objetiva, pero poco se sabe sobre los fuentes neurales que generan este potencial . Se plante6 la hip6tesis, basada en nuestros estu-dios en humanos y en registros de unidad unica en animales, que los AMFR registrados en el craneo reflejan la interacci6n de multiples fuentes . Evaluamos esta hip6tesis utilizando un modelo animal : un conejo no anestesiado. Comparamos los AMFR registrados en diferentes sitios sobre la superficie del cerebro, asi como antes y despues de la administraci6n de agentes que estimu-Ian o suprimen la acci6n de generadores neurales . Tambien registramos localmente los AMFR en diferentes estaciones a to largo del neureje auditivo . Concluimos que los AMFR registrados en el craneo son verdaderamente una respuesta compuesta de multiples generadores cerebrales . Aunque las respuestas a cualquier frecuencia de modulaci6n pueden reflejar la actividad de mas de un generador, los AMFR producidos ante frecuencias de baja o alta modulaci6n parecen refle-jar una fuerte contribuci6n de fuentes corticales o subcorticales, respectivamente .
Palabras Clave: Respuesta de seguimiento de amplitud modulada, potenciales auditivos de estado estable, audiometria electrica, potenciales evocados, codificaci6n temporal
*Department of Neuroscience, University of Connecticut Health Center, Farmington, Connecticut ; tDepartment of Anatomy, University of Connecticut Health Center, Farmington, Connecticut ; $Department of Anatomy, University of Mississippi Medical Center, Jackson, Mississippi ; §Department of Otolaryngology, University of North Carolina, Chapel Hill, North Carolina "The terms amplitude-modulation following response (AMFR), auditory steady-state response (ASSR), steady-state evoked potential (SSEP), and envelope following response (EFR) are used interchangeably in the literature . Whereas other authors in these special issues use the term ASSR, author Kuwada prefers AMFR . (Editor) Reprint requests : Shigeyuki Kuwada, Department of Neuroscience, The University of Connecticut Health Center, 263 Farmington Ave ., Farmington, CT 06030-3405
188

Sources of the Scalp-Recorded AMFR/Kuwada et al
Abreviaturas : AMFR = respuesta de seguimiento de amplitud modulada ; ASSR = respuestas audifvas de estado estable ; EFR = respuesta de seguimiento de envolvente ; GABA = Acido gama-aminobutirico ; HCI = hidrocloruro ; KCI = cloruro de potasio ; MTF = fundbn de transferencia de modulaci6n ; SAM = amplitud modulada sinusoidalmente ; SSEP = potential auditivo de estado estable
T
he amplitude-modulation following response (AMFR) is a neural potential that follows the envelope of complex
sounds . This potential is also known as the enve-lope following response (EFR ; Dolphin and Mountain, 1992), auditory steady-state response (ASSR, Picton et al, 1987), and auditory steady-state evoked potential (SSEP; Rickards et al, 1994). The AMFR can be recorded from the scalp in response to sinusoidally amplitude-modulated (SAM) tones and can be used to assess hearing on a frequency-by-frequency basis across the audiometric range (250-8000 Hz) in adults (Kuwada et al, 1986 ; Rees et al, 1986 ; Picton et al, 1987 ; Griffiths and Chambers, 1991) and in neonates and children (Levi et al, 1992 ; Rickards et al, 1994 ; Aoyagi et al, 1996).
Despite the AMFR's growing reputation as an objective method for hearing evaluation, we know little about its neural generators . Scalp-recorded AMFRs to SAM tones in humans (Kuwada et al, 1986) and in gerbils (Dolphin and Mountain, 1992) appear to reflect the activ-ity of multiple generators . Different generators appear to dominate over different ranges of mod-ulation frequency. Generators with long activa-tion delays dominate at low frequencies, whereas generators with short activation delays domi-nate at high frequencies . The lengths of the delays implicate cortical generators at low fre-quencies and subcortical generators at high fre-quencies . The idea that multiple sources underlie the AMFR was supported by local recordings made earlier in the auditory cortex and inferior colliculus (Tielen et al, 1969). These recordings found that local AMFRs in the auditory cortex had maximum amplitude at low modulation fre-quencies (15-30 Hz) and were activated with long delays, whereas AMFRs in the inferior col-liculus could be recorded at higher modulation frequencies and had shorter delays . However, later studies that recorded AMFRs in animals after ablation of the auditory cortex discredited the idea that the primary auditory cortex was a major source (Tsuzuku, 1993 ; Kiren et al, 1994).
To investigate the sources of the AMFR, we employed an animal model, the unanesthetized rabbit. The advantage of the rabbit is that record-
ings can be made without the confounds of anes-thesia, and we have considerable data on the responses of single neurons to SAM tones at several levels along the auditory pathway. Using behavioral, pharmacologic, and cortical inacti-vation techniques, we conclude that the surface-recorded AMFR reflects the activity of multiple generators .
METHOD
Preparatory Surgery
Six adult Dutch Belted rabbits (-2-2.5 kg) with clean external ears were used . Three were surgically prepared for surface-recorded AMFRs (epidural) and three for locally recorded AMFRs from the superior olivary complex, inferior col-liculus, and auditory cortex . All surgical and postoperative care followed the National Insti-tutes of Health guidelines and was approved by the Institutional Animal Care Committee at the University of Connecticut Health Center .
Both types of recordings involved attaching a square brass rod to the skull to immobilize head movement during recordings . Under anesthesia (ketamine hydrochloride [HCll 35 mg/kg and xylazine 5 mg/kg), the scalp was retracted to expose the skull, and the brass rod was anchored to the skull with screws and dental acrylic. The rod was positioned parallel and to one side of the midsagittal suture.
Surface Epidural Electrodes : During the initial surgery, stainless steel screws (0-80 x 1/a°) were threaded into the skull and contacted the dural surface. Silver wire leads soldered to the screws were led into a plastic strip connector, and this assembly was also attached to the exposed skull with dental acrylic. In almost all cases, one active electrode was placed near the sagittal suture on the side contralateral to the stimulated ear about halfway between the coronal and the lambdoid suture, and the reference electrode was placed in the frontal sinus . A third skull screw served as a ground . In one animal, two additional screw electrodes were placed : one over the auditory cortex and the other near the midline and lambdoid suture .
189

Journal of the American Academy of Audiology/Volume 13, Number 4, April 2002
Local Microelectrodes : One animal was used to record AMFRs from the superior olivary com-plex and inferior colliculus, and two animals were used to record AMFRs from the auditory cortex . A small hole (2 x 2 mm) in the skull just rostral to the lambdoid suture and ~ 2 mm lat-eral to the midline allowed access to the inferior colliculus and superior olivary complex. Access to the auditory cortex was via a rectangular hole in the skull (2 x 4 mm, ~ 3 mm posterior to the bregma and - 11 mm lateral to the mid-line) that overlaid the dorsal part of the auditory cortex . Recordings were made through glass-coated Pt/W microelectrodes (~1 MOhm). In one animal, we used a microwire array to make local AMFR recordings in the auditory cortex. This array consisted of two rows of 12 wires each. Sep-aration of the wires was 250 ~Lm both within and between rows . Each wire was 50 p,m in diameter (<200 kOhms impedance) and insu-lated except for the tip.
In two of the rabbits used for surface record-ings, a chronic cannula was inserted into the external jugular vein and fed into the subclavian vein during the initial surgery. The other end of the cannula was brought up subdermally and fixed onto the skull surface with dental acrylic for the convenient administration of drugs . Heparinized saline (50 U/mL) was injected (1 mL) daily to keep the catheter patent . In another rabbit, a small hole (3 mm in diameter) was drilled over each cortical hemisphere . These holes were used to apply a potassium chloride (KCI) (15% or 25% solution)-soaked cotton pled-gett to the dural surface to induce cortical spread-ing depression .
Acoustic Stimulation
A custom-fitted ear mold for each rabbit was constructed from ear impression compound (Audilin). A sound delivery tube was inserted through the mold, and its tip was within 2 cm of the tympanum . The mold was made while the animal was anesthetized .
SAM tones were generated digitally using a Uni-versity of Wisconsin-based system (Rhode, 1976) and delivered through a Beyer DT-48 earphone connected to the sound delivery tube in the custom-fitted ear mold. Except where noted, the sounds were presented to the ear contralateral to the recording site. Modulation depth was either 80 or 100 percent. Where appropri-ate, stimuli were presented in a block-randomized order.
Amplitude (dB SPL re : 20 ~tPA) and phase at frequencies from 60 to 40,000 kHz were mea-
sured just prior to sacrificing the animal. The earphone was connected to the sound delivery tube, and, in most animals, the tip of a cali-brated probe tube connected to a 1/2" microphone (Bruel and Kjaer) was placed near the tympa-num via a small sealed hole in the external bony meatus . In one animal, the tip of the probe was at the end of the sound delivery tube .
Recording Procedure
Each rabbit was tested, unanesthetized, over a period of several months . Daily recording ses-sions were conducted in a double-walled sound-proofed room . During data collection, the rabbit was placed in a custom plexiglass cradle and its body was restrained in a zippered Lycra sleeve . The rabbit's head was clamped in a fixed position via the head bar. The rabbit was monitored con-tinuously by video camera. Each session usually lasted 1 to 3 hours, but if the rabbit fidgeted, the session was terminated. All rabbits were first given preparatory sittings in which to become accustomed to the recording environment.
Surface potentials were recorded differen-tially between active and reference electrodes, with a third electrode used for ground . The interelectrode resistance was < 12 kOhms. Two amplifiers (EG&G Pare) in series were used to record (total gain 20,000-50,000) and filter (0.3-3000Hz, 12 dB/octave rolloff) the scalp potentials . The SAM tones were delivered con-tinuously, and a pulse at the zero crossing of each modulation cycle was used to trigger an A/D converter to begin digitizing the scalp poten-tials (> 25 points/ modulation cycle) . Potentials were averaged over 10 cycles of the modulation frequency, in blocks of 50 to 2500 trials, and several blocks at each modulation frequency were usually collected. To ensure that there was no electrical or magnetic contamination in the AMFR from the earphones, we tested in each animal the potentials to different carrier and modulation frequencies when the sound deliv-ery tube was plugged. We could not detect any artifacts.
The local recordings were conducted in a similar manner. Prior to these recordings, single-unit recordings were made to ensure that the local recordings were from the auditory nuclei of interest. This was later confirmed by histologic examination of the relevant brain sections . Recordings were made with an extracellular amplifier (Dagan 2400) coupled to a conventional amplifier (EG&G Pare) using the same filter
190

Sources of the Scalp-Recorded AMFR/Kuwada et al
settings (0.3-3000 Hz) . Microwire recordings were made with a different amplifier (TDT Bioamp) and filters (5-3000 Hz) .
To provide a point of comparison to the rab-bit, we included the AMFRs from one human subject with normal hearing . Recording tech-
niques were similar to those for surface record-ings in the rabbit . The scalp potentials were recorded differentially between the vertex and the contralateral earlobe (50,000-100,000 gain ; filters, 0.3-3000 Hz), while the other earlobe served as ground . The resistance between any two electrodes was less than 5 kOhms. During the recording, the subject read or sat quietly in a comfortable chair in a soundproofed booth . The SAM tones were delivered monaurally through mu-metal shielded earphones calibrated for both intensity and phase .
Pharmacologic Procedures
We tested the effects on the surface-recorded AMFR of systemically administered cocaine, sodium pentobarbital, and ketamine HCl/ xylazine and of KCI-induced cortical depres-sion . The effect of cocaine was tested on two rabbits, cocaine and sodium pentobarbital on separate occasions in one rabbit, and ketamine HCl/xylazine and KCI on separate occasions in another rabbit . Different drug treatments in a single animal were separated by at least 2 days . Cocaine (1-1 .5 mg/kg) and sodium pentobarbi-tal were injected through the indwelling catheter. Since rabbits metabolize sodium pentobarbital rather rapidly, we attempted to achieve a steady anesthetic state by giving an initial bolus of sodium pentobarbital (12-25 mg/kg) followed by supplements (12 mg/kg) at approximately 20-to 30-minute intervals. Ketamine HCI/xylazine (35 mg/kg, 5 mg/kg) was injected intramuscularly and lasted about 1 hour.
Spreading depression is confined to the hemisphere where the electrical, mechanical, or chemical (KCI) stimulation is applied and is marked by the absence of the electroencephalo-gram . It creates a deep, reversible inhibition of all cortical functions (Leao, 1944; Bures et al, 1974). It is long lasting (2-3 hours) and does not appear to cause irreversible neuronal injury outside the area of KCI application (Nedergard and Hansen, 1988). In one rabbit, we reversibly inactivated the cortex by applying KCI (15% or 25% solution) to the dural surface to induce cor-tical spreading depression . A cotton pellet soaked in KCI or control solution of saline (0.9%) was
applied to the dural surface. Recordings were ini-tiated about 7 minutes after application of the KCI . After each session, the dura was flushed with saline, filled with a dab of antibacterial oint-ment (Bacitracin), and capped with an elastopolymer (Rolyan) .
Analysis of the AMFR
Figure lA illustrates a surface-recorded AMFR from a rabbit to a SAM tone (modulation = 67 Hz). In this example, the AMFR represents an average of 1000 trials, each trial being 10 modulation cycles in length . The spectrum of the AMFR was analyzed with a discrete Fourier transform (Fig . 1B). This yielded an estimate of the amplitude (peak to peak) at the modulation frequency, which was taken to be the amplitude of the AMFR. The Fourier transform also yielded
A
h
to Ftv
0 Modulation Cycles 10
s 0 a a
AMFR = 5.3 pV, Phase = 0.58 cycles Noise = 0.42 ^ SD = 0 .14 )tV w Criterion = (AMFR-Noise)/SD = 34 .8
E a III III
e7 134 201
Modulation Frequency (Hz)
Figure 1 Waveform (A) and spectrum (B) of a surface-recorded amplitude-modulation following response (AMFR) from a rabbit to a sinusoidally amplitude-mod-ulated tone (modulation frequency = 67 Hz, carrier fre-quency = 2000 Hz, level = 80 dB SPL) . A, Waveform of the AMFR displayed over 10 cycles of the modulation fre-quency (1000 averages). B, Discrete Fourier transform of the waveform in A yields the amplitude (5 .3 p V) and phase (0 .58 cycles) at the modulation frequency and an estimate of the noise in the three frequency bins (arrows; mean = 0.42 pLV) surrounding the modulation frequency (exclud-ing the immediately adjacent bins). The criterion for the
presence of an AMFR is that the amplitude at the mod-ulation frequency (5 .3 p V) minus the mean amplitude of the noise (0 .42 wV) divided by the standard deviation of the noise (0.14 p .V) must exceed 3. In this example, this calculation (34.8) far exceeds our criterion (>3) . p-p = peak-to-peak amplitude.
191

Journal of the American Academy of Audiology/Volume 13, Number 4, April 2002
estimates of the amplitude and standard devi-ation of the surrounding noise. The noise esti-mates were derived from the three flanking bins, excluding the immediately adjacent bin, around the modulation frequency (arrows) . We considered the AMFR to be present if the ampli-tude at the modulation frequency minus the amplitude of the noise divided by the standard deviation of the noise was > 3. The present example far exceeds this criterion (34.8) . The dis-crete Fourier transform also provides an estimate of the phase of the AMFR. We use the well-known relationship between phase and fre-quency in a linear system to estimate the neural delay. If the relationship between the phase of the AMFR and modulation frequency is linear (our criterion is that r > .97), then the slope of this function provides an estimate of the neural delay. The neural delay estimates were corrected for the influence of the recording system (i .e ., delays due to filtering) .
m v
N t a
0
noise
RESULTS
AMFR as a Function of Modulation Frequency
The amplitude and timing of AMFRs in the rabbit were similar to those in humans . Figure 2, A and B, shows the modulation transfer func-tions (MTF) (i.e ., amplitude and phase of the AMFR as a function of modulation frequency) from a rabbit (dashed lines indicate the noise level; see Fig. 1) . The amplitude displays a series of peaks and valleys . The larger two peaks are at low modulation frequencies (15-70 Hz), whereas the smaller two peaks are at high mod-ulation frequencies. The presence of peaks and valleys suggests the interaction of different gen-erators, with the peaks being the regions where the responses from different sources are in phase and the valleys the regions where the responses are out of phase. In Figure 213, the phase of the AMFRs in the peak regions, excluding those in
1 .5, C Human
0
Rabbit
"I
-9 -criterion >3 0 criterion <3
2 .7 msec
0 200 400 600 800
,/26 msec
100 300
8 msec
0 0 50 100 150 200 250 300
Modulation Frequency (Hz)
Figure 2 The modulation transfer function (MTF) of a rabbit and a human both display peaks and valleys. Rabbit surface recording was from a near midline screw placed midway between the bregma and the lambdoid suture (i .e ., vertex) referenced to a screw in the frontal sinus. Human scalp recordings were from the vertex referred to the earlobe contralateral to the stimulated ear. A, MTF for a rabbit to sinusoidally amplitude-modulated (SAM) tones (carrier = 5000 Hz, level = 90 dB SPL) across a range of modulation frequencies (14-799 Hz). B, Phase versus modulation plot displays four linear segments . C, MTF for a human adult with normal hearing to SAM tones (carrier = 1 kHz, level = 70 dB SPL) across a range of modulation frequencies (26-261 Hz). In both A and C, solid circles indicate AMFRs with criterion >3 and open circles criterion <3 ; the dashed line indicates the mean noise at each modulation frequency. D, The slope of the phase versus modulation frequency plot shows three linear segments (three different neural delays), each associated with a particular range of modulation frequencies . The neural delays decrease with modulation fre-quency. To illustrate linearity of the phase plots, modulation frequency is plotted on a linear scale, whereas the peak-to-peak (p-p) amplitude is plotted on the conventional log scale.
d e12 msec
192
Sources of the Scalp-Recorded AMFR/Kuwada et al
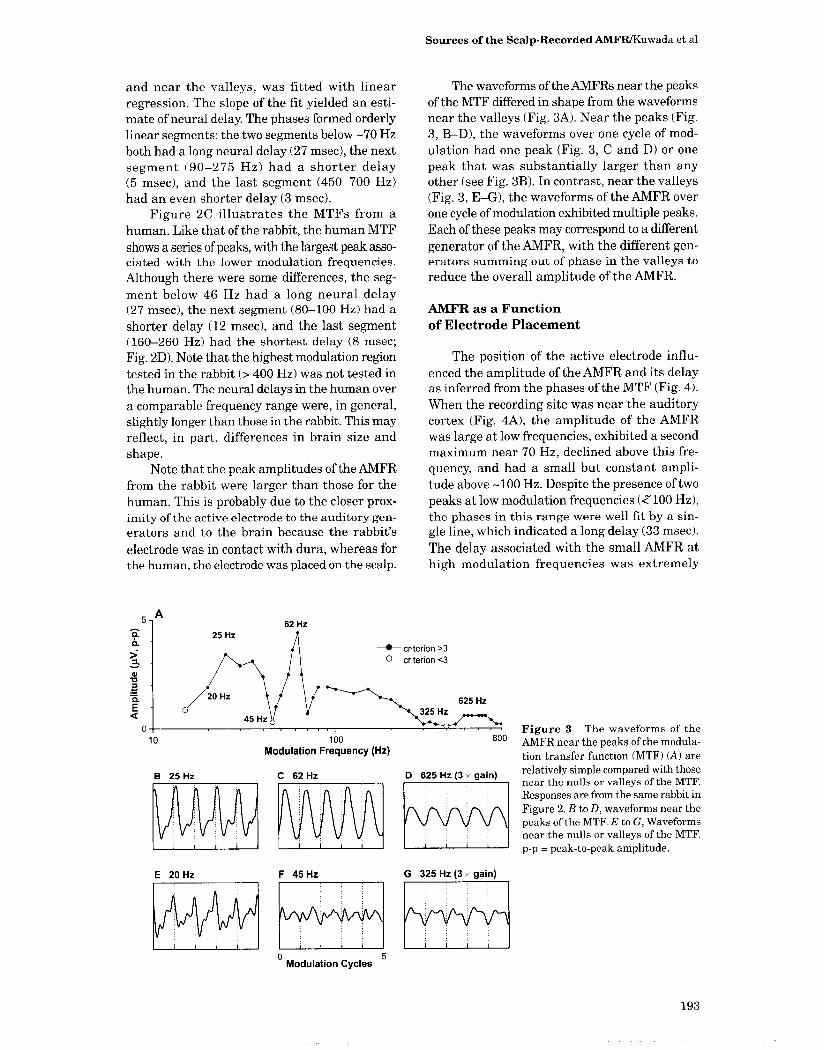
and near the valleys, was fitted with linear regression . The slope of the fit yielded an esti-mate of neural delay. The phases formed orderly linear segments : the two segments below -70 Hz both had a long neural delay (27 msec), the next segment (90-275 Hz) had a shorter delay (5 msec), and the last segment (450-700 Hz) had an even shorter delay (3 msec).
Figure 2C illustrates the MTFs from a human. Like that of the rabbit, the human MTF shows a series of peaks, with the largest peak asso-ciated with the lower modulation frequencies . Although there were some differences, the seg-ment below 46 Hz had a long neural delay (27 msec), the next segment (80-100 Hz) had a shorter delay (12 msec), and the last segment (160-260 Hz) had the shortest delay (8 msec ; Fig. 2D). Note that the highest modulation region tested in the rabbit (> 400 Hz) was not tested in the human. The neural delays in the human over a comparable frequency range were, in general, slightly longer than those in the rabbit . This may reflect, in part, differences in brain size and shape.
Note that the peak amplitudes of the AMFR from the rabbit were larger than those for the human. This is probably due to the closer prox-imity of the active electrode to the auditory gen-erators and to the brain because the rabbit's
electrode was in contact with dura, whereas for the human, the electrode was placed on the scalp .
The waveforms of the AMFRs near the peaks of the MTF differed in shape from the waveforms near the valleys (Fig. 3A) . Near the peaks (Fig . 3, B-D), the waveforms over one cycle of mod-ulation had one peak (Fig . 3, C and D) or one peak that was substantially larger than any other (see Fig. 3B). In contrast, near the valleys (Fig . 3, E-G), the waveforms of the AMFR over one cycle of modulation exhibited multiple peaks. Each of these peaks may correspond to a different generator of the AMFR, with the different gen-erators summing out of phase in the valleys to reduce the overall amplitude of the AMFR.
AMFR as a Function of Electrode Placement
The position of the active electrode influ-enced the amplitude of the AMFR and its delay as inferred from the phases of the MTF (Fig . 4) . When the recording site was near the auditory cortex (Fig . 4A), the amplitude of the AMFR was large at low frequencies, exhibited a second maximum near 70 Hz, declined above this fre-quency, and had a small but constant ampli-tude above -100 Hz . Despite the presence of two peaks at low modulation frequencies (< 100 Hz), the phases in this range were well fit by a sin-gle line, which indicated a long delay (33 msec). The delay associated with the small AMFR at high modulation frequencies was extremely
t criterion >3 O criterion <3
625 Hz
B 25 Hz C 62 Hz i1 . h
Ij U U y E 20 Hz F 45 Hz
W
0 Modulation Cycles 5
800
D 625 Hz (3 x gain)
G 325 Hz (3 x gain)
Figure 3 The waveforms of the AMFR near the peaks of the modula-tion transfer function (MTF) (A) are relatively simple compared with those near the nulls or valleys of the MTF. Responses are from the same rabbit in Figure 2. B to D, waveforms near the peaks of the MTF. E to G, Waveforms near the nulls or valleys of the MTF. p-p = peak-to-peak amplitude.
193

Journal of the American Academy of Audiology/Volume 13, Number 4, April 2002
A Near Auditory Cortex
0
B Near Midline
a a
6_1C Near Midbrain 4
>=L d v w
33 msec
2 .6 msec
Figure 4 Different surface record-ing sites yield different modulation transfer functions (MTFs) and neural delays . The amplitude (left column) and phase (right column) MTFs at different surface recording sites in one rabbit (A-C). In all cases, the car-rier frequency (2000 Hz, 80 dB SPL) and the position of the reference elec-trode (frontal sinus) were the same . Solid circles indicate amplitude-modulation following responses with criterion >3 and open circles with cri-terion <3 . A, Recordings from a surface
4.4 msec electrode placed over the dorsal part
m
4) N L a
-~ 0 i ~ 10 100 0 100
Modulation Frequency (Hz)
short (2.6 msec). Near the midline, medial to the auditory cortex (Fig. 4B), the MTF was bimodal, with two peaks of roughly equal amplitude sep-arated by a valley. The phases associated with the low-frequency peak were well fit by a straight line, and the delay was shorter than over the auditory cortex but was still relatively long (22 msec). The phases associated with the peak at higher modulation frequencies were also well fit by a straight line, but the delay was longer (4.4 msec) than at similar frequencies over the auditory cortex . Note that the amplitudes at high frequencies near midline were greater than over the auditory cortex and that the peak at these frequencies extended below 100 Hz, into the low-frequency region over the auditory cor-tex. Finally, at the third, more caudal recording site (Fig . 4C, near midbrain), the MTF was sim-ilar to that medial to the auditory cortex, except that the peak at low frequencies was now smaller than the peak at high modulation frequencies.
The shifts in amplitude and delay at low and high frequencies support the idea that multiple generators are present. At high frequencies, there appear to be two generators . One gener-ator is associated with an extremely short delay (2.6 msec) and is picked up by an electrode over the auditory cortex. At more medial locations, the
200 00
of the auditory cortex (see Methods). B, Recordings from a surface electrode placed near the midline, midway between the bregma and the lamb-ddoid suture . C, Recordings from a surface electrode placed near the lam-boid suture, - 2 mm from midline. p-p = peak-to-peak amplitude.
signal from this generator is either absent or is overshadowed by a stronger signal from a gen-erator with a slightly longer delay (-4.5 msec). At low frequencies, amplitudes decline as the electrode is moved away from the auditory cor-tex, suggesting that this is the chief source at these frequencies. The shifts in delay suggest an additional source, but these could also be a result of inadequate sampling of the relatively large volume of the auditory cortex .
Effects of Behavioral and Pharmacologic Stimulation on the AMFR
If the AMFR reflects contributions from dif-ferent sources, then sources higher in the path-way might be more affected by behavioral state than sources lower in the pathway. Since low and high modulation frequencies are associated with long and short delays, respectively, they are also likely to be associated with centers higher and lower in the auditory pathway. We tested this idea by measuring the AMFR at low and high modu-lation frequencies, with and without behavioral stimulation that consisted of gently touching the rabbit at irregular intervals as an attempt to keep it alert. The AMFRs to 52 and 201 Hz, before and during behavioral stimulation, are shown
194

Sources of the Scalp-Recorded AMFR/Kuwada et al
A Control
0
Figure 5 Behavioral stimulation enhanced the amplitude-modulation following response (AMFR) at a low modulation frequency but not at a high modulation frequency. Carrier frequency was 2000 Hz at 80 dB SPL. The control response was taken with the rabbit alone in the soundproofed room, whereas behavioral stimula-tion was taken while the experimenter gently tapped the rabbit's head and nose . A and B, AMFR to a low modu-lation frequency (52 Hz). The ampli-tude of the AMFR increased by a factor of 2 during behavioral stimulation. C and D, AMFR to a high modulation frequency (201 Hz). At this modulation frequency, behavioral stimulation had little or no effect on the AMFR .
J
o s Modulation Cycles
in Figure 5 . Behavioral stimulation increased the amplitude of the AMFR at low modulation rates by about a factor of two (Fig. 5, A and B : 3 .9 vs 7.6 V,V), but at high modulation rates, a change was barely noticeable (Fig . 5, C and D : 0.98 vs 1.03 wV) . Thus, AMFRs at low modula-tion frequencies are influenced by behavioral stimulation, whereas those at high modulation fre-quencies are not . This supports the idea that the AMFRs over different ranges of frequency are dominated by different sources, with the source at low modulation frequencies lying in a higher center such as the cortex, which is more likely influenced by behavioral stimulation.
Using the same rationale, we tested the effects of a stimulant (cocaine) at a low (62 Hz)
A Control
B Behavioral Stimulation
Cocaine
and a high modulation frequency (200 Hz). Fig-ure 6 shows that as with behavioral stimulation, cocaine caused a marked increase in the AMFR at a low modulation frequency (Fig . 6, A-B) but no change at the high modulation frequency (Fig. 6, D-E). Approximately 80 minutes after the injection of cocaine, the AMFRs at low mod-ulation frequencies returned to their control levels (Fig . 6C).
Since stimulation (behavioral or cocaine) elevated the AMFRs to low modulation fre-quencies, we reasoned that a depressant (sodium pentobarbital) should attenuate the AMFRs at these frequencies . Pentobarbital eliminated the two lowest-frequency peaks of the MTF, replac-ing them with a broad plateau of roughly con-
D E F
0 5 Modulation (cycles)
Figure 6 Cocaine enhanced the amplitude-modulation following response (AMFR) at a low modula-tion frequency but not at a high mod-ulation frequency. Organization similar to Figure 5. Carrier frequency was 5000 Hz at 90 dB SPL. The responses under cocaine were taken within a 10-minute interval immedi-ately after an intravenous injection of cocaine (1 .5 mg/kg) and the post-cocaine response (C and F) -80 min-utes after the injection of cocaine. A to C, AMFR to a low modulation fre-quency (62 Hz) . The amplitude increased by a factor of ~3 immedi-ately after injection of cocaine (B) and returned to near control levels by -80 minutes (C). D to F, AMFR to a high modulation frequency (200 Hz). At this modulation frequency, cocaine had little or no effect on the AMFR.

Journal of the American Academy of Audiology/Volume 13, Number 4, April 2002
1 0 1 00 600
300 400 500 600 Modulation Frequency (Hz)
Figure 7 Sodium pentobarbital attenuated the ampli-tude of the AMFR and shortened the neural delay at low modulation frequencies, whereas the responses to high modulation frequencies were relatively unaffected . Con-trol responses are the same as in Figure 2. Sodium pen-tobarbital was administered via an indwelling intravenous catheter. Recordings were made during and after an ini-tial bolus of sodium pentobarbital (25 mg/kg) followed by supplements (12 mg/kg) at approximately 20- to 30-minute intervals. A, Control and sodium pentobarbital modulation transfer functions (MTFs) . B, Control and sodium pentobarbital phase versus frequency plots. Cor-responding frequency regions in the MTF and phase plots are displayed as dotted vertical lines. p-p = peak-to-peak amplitude.
stant amplitude (Fig. 7A). Thus, the amplitude decreased at the peaks but increased at fre-quencies where valleys were present. At inter-mediate frequencies (region 3), the amplitude decrease was small but consistent, and at the highest frequencies (region 4), they hardly changed at all. The phase portion of the MTF also changed under pentobarbital (Fig. 7B). At low frequencies, the slope of the fitted regres-sion line decreased (Fig. 7B), whereas at higher frequencies, there was barely any change . Fur-thermore, the slope at low frequencies (region 2) under pentobarbital was 5 msec, which matched the slope associated with the third peak observed in the control (region 3) . All of the changes are consistent with the suppression of a generator at low frequencies that had been activated with a long delay. The remaining sig-nal at these frequencies under pentobarbital appears to be generated by a source with a shorter delay.
Further evidence for source interactions was the changes in the AMFR waveform under
sodium pentobarbital, cocaine, and ketamine . Figure 8A compares the AMFR waveform under sodium pentobarbital and cocaine at a modula-tion frequency (45 Hz) that corresponded to a val-ley in the MTF (see Fig. 3) . Where two peaks per modulation cycle were present prior to admin-istration of pentobarbital (control), one narrow and one broad, under pentobarbital, the broad peak was attenuated, leaving only the narrow peak . In contrast, under cocaine, a single, large ,peak was present at the phase where the broad peak had been previously. The amplitude of the single peak under cocaine was over 5 times the amplitude of the broad peak prior to cocaine administration. The changes under pentobarbital and cocaine are consistent with the presence of two generators, one of which was selectively depressed by pentobarbital and enhanced by cocaine . Along with Figure 6, the effects of cocaine are consistent with the presence of two generators, which are active over different but overlapping ranges of frequency.
6 Modulation Cycles 5
Figure S Sodium pentobarbital, cocaine, and keta-mine selectively altered the waveform of the amplitude-modulation following response at or near the nulls in their modulation transfer functons . Control, sodium pento-barbital, cocaine, and ketamine HCI/xylazine were taken in separate sessions in the same rabbit using the same recording configuration (vertex referenced to frontal sinus) . A, Control, pentobarbital, and cocaine responses to a 45-Hz sinusoidally amplitude-modulated (SAM) tone (carrier = 5000 Hz, 90 dB SPL). B, Control and ketamine responses to a 47-Hz SAM tone (carrier = 2000 Hz, 80 dB SPL) . In both A and B, as a visual aid, vertical dashed lines are positioned at the initial peak of the control response.
196

Sources of the Scalp-Recorded AMFR/Kuwada et al
Figure 8B compares the AMFR, with and without ketamine at a slightly different modu-lation frequency (47 Hz) than that used in Fig-ure 8A (45 Hz) but still near a valley in the MTF (see Fig. 3) . Here the waveform over one cycle of modulation displayed three peaks in the unanesthetized state (control). Under ket-amine, the waveform was simplified . It appeared that ketamine did not affect the source associ-ated with the initial peak in the cycle but depressed the activity of the sources associated with the subsequent peaks.
Identification of Sources of the AMFR
The AMFRs at low frequencies appeared to be activated at long delays and also appeared to
be selectively modulated by a depressant and a stimulant . To investigate the possibility that AMFRs at these frequencies were cortical in origin, we reversibly inactivated the cortex using
KCI-induced spreading depression while record-ing the AMFR from a surface electrode . Figure 9A shows the amplitude (left panel) and phase
(right panel) MTFs to a range of low (23-37 Hz) and high (101-201 Hz) modulation frequencies in the unanesthetized animal . As usual, the
responses at low modulation frequencies were large and their delays long relative to the higher modulation frequencies . Applying KCI to the
A Control
B Ipsilateral KCI
n
22 msec
27 msec
a 41 C Contralateral KCI 2-1
'9 -criterion >3 -o--criterion <3
A
100 200 0 so Modulation (Hz)
cortex ipsilateral to the stimulated ear had min-
imal, if any, effect on the AMFR (Fig. 9B) . This
suggests that the sources of the AMFR do not
reside in the cortex on the same side as the stimulated ear. However, KCI applied to the
cortex contralateral to the stimulated ear (Fig . 9C) selectively attenuated the AMFRs at low modulation frequencies, whereas those at high
modulation frequencies remained relatively unchanged . Furthermore, after KCI application, the phase slopes at the low modulation fre-quencies appeared to align with those at the high modulation frequencies . These changes in
the surface-recordedAMFR suggest that a major
source at low modulation frequencies is located in the cortex opposite to the stimulated ear . In contrast, the AMFRs at high modulation fre-
quencies were relatively unaffected by cortical depression, suggesting that their origin lies in subcortical regions .
To determine if the different delays observed in the surface-recorded AMFR over different ranges of modulation frequency corresponded to delays in structures along the auditory pathway,
we made local recordings of the AMFR in the auditory cortex (Fig. 10A), inferior colliculus (Fig . 10B), and superior olivary complex (Fig .
10C) . The MTFs show that the peak modulation frequency decreases from the superior olivary complex to the auditory cortex (Fig . 10, left
4 msec
3 msec
4 msec
Figure 9 Cortical inactivation of the cerebral cortex contralateral to the stimulated ear attenuated the ampli-tude of the amplitude-modulation fol-
lowing response and shortened the neural delay at low modulation fre-quencies, whereas inactivation to the ipsilateral cortex had little if any effect
at all on modulation frequencies . Car-
rier frequency was 2000 Hz at 80 dB
SPL, and recording configuration was vertex referenced to frontal sinus. Right
and left columns display the ampli-tude and phase modulation transfer functions, respectively. Owing to time constraints imposed by cortical inacti-
vation, we tested only a limited range of low modulation (23-37 Hz) and high modulation frequencies (101-201 Hz). A, Control responses (unanesthetized).
B, Responses under potassium chlo-ride (KCI)-induced inactivation of the cortex ipsilateral to the stimulated ear. C, Responses under KCI-induced inac- _
10 15o 0 zoo tivation of the cortex contralateral to the stimulated ear. p-p = peak-to-peak amplitude.
197

Journal of the American Academy of Audiology/Volume 13, Number 4, April 2002
A Superior Olivary Complex
B Inferior Colliculus
A
C Auditory Cort ex 1, d 3
a .a N
d
E V
v
a N
d N A
a E `o z 0
0 10 100 1000 100 200 Modulation Frequency (Hz)
panel) . It is approximately 250 Hz for the supe-rior olivary complex, 90 Hz for the inferior col-liculus, and 20 Hz for the auditory cortex . The
corresponding phase plots also show an increase in neural delay from the superior olivary com-plex to the auditory cortex . In addition, each
C p
y 8- d
6-0 N U) t 4 a
2- m 11 msec
3.23.2 msec _~
Figure 10 The peaks of locally recorded amplitude-modulation fol-lowing responses shift to lower mod-
2 .9 msec ulation frequencies and the phase
tex to SAM tones (carrier = 2000 Hz, 70 dB SPL) . Recordings are single ended.
slopes increase at higher stations along the auditory pathway. Right and left columns display the ampli-tude and phase modulation transfer functions, respectively. A, Local recordings in the superior olivary complex to sinusoidally amplitude modulated (SAM) tones (carrier = 6000 Hz, 70 dB SPL). B, Local record-ings in inferior colliculus to SAM tones (carrier = 8000 Hz, 55 dB SPL) . C, Local recordings in auditory cor-
27 3 msec 1 T
0 0 i --i--r--r 0 20 40 60 80 100 120 140 160 180 200 0 20 40 60
Modulation Frequency (Hz)
msec
19 msec ~- 20 - 11 msec
~ 16 lomsec 2
24 11 msec
15 11 msec
19 - 11 msec 1
80 100 120 140 160 180 200
Figure 11 Local recordings in the auditory cortex can display similar nulls and similar neural delays . Recordings are from a subset of adja-cent microwires (eight) in an array chronically implanted in the auditory cortex (see Methods) . Carrier fre-quency was 4000 Hz at 50 dB SPL. Each wire was referenced to the same microwire in the array that did not show amplitude-modulation follow-ing response activity. A, Normalized amplitude modulation transfer func-tions (MTFs) from the eight microwires . Although peak amplitudes occurred at different modulation fre-quencies, all MTFs showed a null at 71 Hz . B, Mean MTF and standard errors of the responses in A. C, Phase plots from the eight microwires . For illustrative purposes, the phase plots from each wire have been incremented by one cycle. D, Mean phase plots and standard error of the nonincremented phase plots in C.
198

Sources of the Scalp-Recorded AMFR/Kuwada et al
phase plot displays two linear segments corre-
sponding to different neural delays . In each
structure, the shorter delays are associated with higher modulation frequencies, where the
amplitude is smaller. The longer delays may reflect the activity of the neurons intrinsic to the structure, and the shorter delays may reflect the
activity of the inputs to that structure . If this is so, then the long delay at low modulation frequencies observed in surface-recorded AMFRs
(-27 msec) may correspond to the activity of neu-rons in the auditory cortex. However, the shorter
delays observed in surface recordings (-5 and
-3 msec) do not appear to correspond to the delays of neurons in the inferior colliculus or the
superior olivary complex; instead, they seem to
represent the delays of neurons feeding into
these structures . We noticed considerable variability in the
locally recorded AMFRs within a structure . This
is probably because the electrode tips were small
and sampled a small area surrounding the elec-trode tip . To sample the AMFRs in the largest
structure more effectively, we made local record-
ings in the primary auditory cortex using a
microwire array. As expected, different electrodes
displayed different MTF shapes . However, a sub-
set of the array (eight of the microwires) in close
proximity to each other (four in one row and four
in the other row) displayed remarkable similar-
ities in their MTFs (Fig . 11, A and B) and phase
plots (Fig . 11, C and D) . The carrier frequency
(4 kHz) and level (50 dB SPL) were the same for
all of the recordings shown . Note that although
the position of the peaks varies among the wires,
all had a common null near 70 Hz (see Fig. 11A) .
The mean MTF shows a plateau between 20 and
40 Hz and a peak near 120 Hz, separated by a null
near 70 Hz (Fig . 12B) . The remaining microwires
(16/24) did not display this null . The phase plots
indicate that the lower modulation frequencies
(21-61 Hz) are associated with longer delays
compared with those at higher modulation fre-
quencies (91-181 Hz) . Do the locally recorded AMFRs in the audi-
tory cortex match the activity of neurons in this
structure? Figure 12 displays the distribution of
the best modulation frequency (A) and the mean synchrony (B) of single cortical neurons recorded
extracellularly with a microelectrode . The best
modulation frequency and the highest synchrony
are both about 10 to 15 Hz . This corresponds, in
general, to the modulation frequency associated
with the peak amplitude of locally recorded AMFRs from the auditory cortex (see Figs . 10C
and 11) .
A Best Modulation Frequency 2s a r--,
z
0 r U
fI
n1 ~I II II II II II II II Ir-,n~ 1-2 4-6 6-12 16-24 32-48 64-96 128-192
Modulation Frequency (Hz)
Figure 12 Neurons in the auditory cortex prefer low
modulation frequencies. The cortical neurons (n = 114)
were usually studied with the same range of modulation
frequencies (1-400 Hz in 0.5-octave steps) using carriers
at their best pure-tone frequency (-2-20 kHz) and usu-
ally at a stimulus level of 70 dB SPL. Responses are all
to sinusoidally amplitude-modulated tones presented to
the contralateral ear (re: recording site). A, Distribution
of best modulation frequency (i .e ., the modulation fre-
quency that evoked the most spikes and had significant
synchrony (p < .001, Rayleigh coefficient) . B, Mean syn-
chrony and standard error across modulation frequency.
For each neuron, only responses that showed significant
synchrony (p < .001, Rayleigh coefficient) were included .
The synchrony across neurons was averaged at each
modulation frequency. See Kuwada and Batra (1999) for
details of synchrony and best modulation analysis and
Fitzpatrick and colleagues (2000) for details of cortical
recordings .
DISCUSSION
B y analyzing waveforms, varying electrode placement, performing pharmacologic manipulations, and recording both local AMFRs
and single units, we have provided evidence that the surface-recorded AMFR reflects con-
tributions from multiple sources. We will, in turn, discuss the neural substrates of the AMFR, compare our results with other studies investi-
gating the sources of the AMFR, and discuss the use of the AMFR as an audiometric tool .
Neural Substrates of the AMFR
At every level of the auditory system, neu-
rons can temporally follow the envelopes of mod-
199

Journal of the American Academy of Audiology/Volume 13, Number 4, April 2002
ulated signals. However, the upper limit of mod-ulation frequencies that neurons can follow decreases as the information ascends along the auditory pathway (see Frisina, 2001, for a review). In the auditory nerve, the MTF is low-pass in shape, with a corner cutoff frequency of about 800 Hz for neurons tuned above - 2kHz (Joris and Yin, 1992). For most neurons in the inferior colliculus, the MTFs are bandpass in shape, with the bulk of best modulation fre-quencies below - 100 Hz (Rees and Moller, 1983 ; Batra et al, 1989 ; Krishna and Semple, 2000). Cortical neurons also display bandpass MTFs, and the bulk of their best modulation frequen-cies are below - 20 Hz (Schreiner and Urbas, 1986, 1988 ; see also Fig. 12). Our locally recorded AMFRs also showed a similar decrease in the ability to follow high-frequency envelopes at progressively higher stations along the audi-tory pathway. Since neurons in all structures can follow low modulation frequencies, the surface-recorded AMFR to these frequencies can reflect contributions from all levels of the auditory pathway. In contrast, responses to higher and higher modulation frequencies can only reflect contributions from lower and lower auditory structures .
Neurons in the same structure can process envelopes differently. For example, in the coch-lear nucleus, neurons with an onset discharge pattern show higher synchrony to envelopes than those with a sustained discharge pattern (Rhode and Greenberg, 1994). In the superior oli-vary complex, the phase of the response of sus-tained neurons to envelopes is often 180 degrees out of phase to that of neurons with an off-discharge pattern (Kuwada and Batra, 1999). Even the same neurons can show different responses under different circumstances. In the cochlear nucleus and at higher stations, the MTFs of some neurons can change from lowpass to bandpass depending on the sound level (Moller, 1974; Rees and Moller, 1983 ; Frisina, 2001 ; Krishna and Semple, 2000). Moreover, their MTFs often show two peaks separated by a suppressive region (Krishna and Semple, 2000). Thus, the valleys and associated com-plex waveforms (doubling or tripling ; e.g ., Fig. 3) in the surface-recorded AMFRs could repre-sent the heterogeneous response of neurons within a structure, as well as the different responses between structures .
Neurons in the cortex of the rabbit can follow envelopes up to - 200 Hz . However, most cortical neurons have their best modulation fre-quencies (amplitude and synchrony) below
-15 Hz (see Fig. 12). A similar distribution has been reported in the auditory cortex of the cat (Schreiner and Urbas, 1988). Furthermore, this best modulation frequency corresponds to the peak frequency for locally recorded AMFRs in the auditory cortex of the rabbit (e.g ., see Figs . 10C and 11). In the midbrain, local recordings of the AMFR show a peak around -100 Hz (see Fig. 11B). However, the most prominent peak in the surface-recorded AMFR of the rabbit usually occurs neither at the peak activity of the cortex nor at the peak activity in the inferior collicu-lus. Thus, it is unlikely that the surface-recorded AMFR is a simple reflection of a single genera-tor but instead is likely to be the result of mul-tiple generators sensed by the surface electrode. Similar events may underlie the circa 40-Hz potentials in humans . That is, the large circa 40-Hz potential associated with human AMFRs or with click trains at that frequency (Galambos et al, 1981) is likely to be attributable to source interactions that reinforce over that frequency range rather than to entrainment of underlying 40-Hz neural oscillators .
Identifying the Generators in the Surface-Recorded AMFR
Our results indicate that the surface-recorded AMFR has multiple generators . Fur-thermore, different generators appear to dominate over different ranges of modulation fre-quency and in different recording configura-tions. At low modulation frequencies (<80 Hz), the cortex appears to be a major source . At higher modulation frequencies, there appears to be at least two generators that are likely sub-cortical. One generator has a delay of - 5 msec and corresponds to perhaps a midbrain or pon-tine source . The other has a delay of - 3 msec and corresponds perhaps to the superior olivary complex or the cochlear nucleus.
The scalp or surface-recorded AMFR reflects both near- and far-field sources; therefore, the contributions of these sources depend on brain size, in general, and the distance from the record-ing electrode, in particular. For example, the surface-recorded MTF in the rabbit has more peaks than that in the human, suggesting more source summation, especially at low modula-tion frequencies. This suggests that the gener-ators in the rabbit are in closer proximity to the surface electrode than in humans. As another example, the scalp-recorded AMFRs in gerbils (Dolphin and Mountain, 1992) display not only synchrony to envelopes but also robust syn-
200

chrony to the carrier frequency. Robust syn-chrony to carrier frequencies is not present in neurons at or above the level of the inferior col-liculus in rabbits (Stanford et al, 1992) and is not
detectable to carrier frequencies much above --500 Hz in the surface-recorded AMFRs of rab-bits and humans (Batra et al, 1986) . This sug-gests that surface recordings in the gerbil detect almost equally contributions from structures below and above the midbrain . Moreover, they reported a consistent valley between 200 and 300 Hz, above which estimates of neural delays became highly unreliable . The likely explanation is the smaller brain size and, consequently, the smaller distance of the recording electrode from the generators in the gerbil compared with that of the rabbit and human. Thus, the enumeration
of sources of the AMFR with different delays is simplified in larger brains with widely spaced generators .
The cortex appears to be optimally acti-vated by low modulation frequencies and
subcortical structures by high modulation fre-
quencies . Compared with low modulation frequencies, at high modulation frequencies (>150 Hz), the contributions of the cortex are minimal based on the preferred modulation fre-
quencies of local cortical AMFRs and of cortical neurons . Moreover, the AMFRs to low modula-tion frequencies have long neural delays,
whereas those to higher modulations have shorter neural delays . Finally, the AMFRs at low
modulation frequencies are markedly affected by behavioral and pharmacologic interventions, whereas those at high modulation frequencies are not .
In general, since inhibition cumulatively increases along the auditory neuraxis, agents that potentiate inhibition should have an increas-ing effect at higher and higher structures . A major inhibitory transmitter is y-aminobutyric acid (GABA), and sodium pentobarbital is known to potentiate GABA-mediated inhibition (Barker and Ransom, 1978). Sodium pentobarbital depresses the activity of neurons in the inferior colliculus (Kuwada et al, 1989) and auditory cortex (Fitzpatrick et al, 2000). Consistent with our reasoning, sodium pentobarbital had little, if any, effect on the responses to the highest modulation frequencies (see Fig. 7, region 4), which had the shortest delays and are likely to be generated by lower brainstem sources (e .g., cochlear nucleus or superior olivary complex) . However, it reduced the AMFR at intermediate modulation frequencies (see Fig. 7, region 3), which had a longer delay and are likely to be gen-
Sources of the Scalp-Recorded AMFR/Kuwada et al
erated by sources in the midbrain or pons, and caused a profound reduction in amplitude at the lowest frequencies (see Fig. 7, regions 1 and 2), which had the longest delays and are likely generated by cortical sources.
The variability in the delay of the AMFR at low frequencies, which we believe to be cortical
in origin, may have been due to the variation in the processing of amplitude-modulated sounds in different cortical areas . The cortical contri-butions to the AMFR are complex because there are several cortical areas that code sound envelopes (e.g ., Schreiner and Urbas, 1986, 1988) . Although neurons in all areas respond to low modulation frequencies, some areas in the cat's cortex are tuned to very low frequencies
(<7 Hz, secondary auditory cortex, posterior
auditory field, ventral posterior auditory field), others to intermediate frequencies (--14 Hz, pri-mary auditory cortex), and still others to higher frequencies (- 30 Hz, anterior auditory field) . Furthermore, the neural latencies in these areas can differ substantially. For example, neurons in the primary auditory cortex show latencies between - 10 and 20 msec, whereas those in the posterior auditory field show latencies between 20 and 100 msec (e.g ., Schreiner and Urbas,
1988). In general, our surface- and locally recorded AMFRs displayed features consistent with the heterogeneity of such cortical sources. For example, the surface-recorded neural delays to low modulation frequencies ( :580 Hz) ranged between 19 and 33 msec, and those from local cortical recordings ranged from 15 to 27 msec . The peaks and valleys in this range (e.g., Fig. 7) suggest that they are the result of contributions from multiple sources within the cortex .
Comparison with Other Studies
There are studies that have investigated the sources of event-related potentials (middle latency response and the possibly related 40-Hz potential), and most have used an anesthetized animal . Since we have shown that anesthesia has a major effect at low modulation frequencies, we consider here only studies that have used an unanesthetized preparation. Our results are in disagreement with studies that did use an unanesthetized animal model to investigate the sources of the surface-recorded AMFR. Kiren and colleagues (1994) measured the phase coher-ence of the surface-recorded AMFR to SAM tones in cats and reported that bilateral ablation of the auditory cortex had little effect on this mea-sure, whereas lesions of the inferior colliculus
201

Journal of the American Academy of Audiology/Volume 13, Number 4, April 2002
contralateral to the sound stimulation made the AMFR undetectable at all modulation fre-quencies (20-200 Hz) . They concluded that the inferior colliculus was the prominent generator and that the auditory cortex was not a genera-tor for the surface-recorded AMFR. At first glance, their results are not in accord with our findings . However, they based their conclusion solely on changes in the phase coherence, which is a measure of detectability, and provided no information about amplitude . If that had been our only measure, we also would have concluded that the cortex was not a generator of the AMFR as the AMFR was still detectable at most low fre-quencies . However, at these frequencies, the amplitude of the AMFR was markedly attenu-ated after inactivation of the contralateral cor-tex, and its neural delay was considerably shorter and similar to that at higher modulation frequencies (see Fig. 9) . A study similar to that of Kiren and colleagues (1994) investigated the effects of cortical and collicular lesions on the 40-Hz click-evoked steady-state response (Tsuzuku, 1993) . Although focal ablation of the primary auditory cortices caused only a small reduction (-10%), bilateral lesions of the inferior colliculi produced a marked reduction (-40%) in the 40-Hz response . The small reduction after ablation of the primary auditory cortex is in contrast to the marked reduction we observed under cortical inactivation . One resolution may be that areas of the cortex other than the primary auditory cortex may be the major contributors to the surface-recorded AMFR and 40-Hz response . Another possibility is that the 40-Hz response evoked by clicks (a broad-band stimu-lus) activates different generators than the AMFRs to SAM tones . Support for this latter pos-sibility is the finding that contralateral ablation of the inferior colliculus abolished the AMFR to SAM tones (Kiren et al, 1994), whereas the 40-Hz click response was still detectable, albeit reduced (Tsuzuku, 1993) .
Use of the AMFR as an Audiometric Tool
Although the AMFR is most robust at low modulation frequencies in humans, its use as an audiometric tool at these frequencies is prob-lematic because the amplitude changes with state. For example, Cohen and colleagues (1991) showed that the AMFRs to modulation frequen-cies around 40 Hz were attenuated in sleeping adults . If behavioral state is controlled by tasks such as reading, then the AMFR at low modula-tion frequencies can be used as an audiometric tool
(Kuwada et al, 1986). However, the use of elec-trical audiometry is most important in assessing hearing in infants, and it is difficult to control their behavioral state. For this reason, the AMFR at low modulation frequencies has been viewed as unsuitable for electrical audiometry.
Another reason for the disfavor of the circa 40-Hz AMFR is that it is difficult to detect in sleeping and sedated infants. This is consistent with our findings that, in rabbits, anesthesia markedly attenuates the AMFR to these modu-lation frequencies. Furthermore, cortical inacti-vation through KCl-induced spreading depression had similar attenuating effects to sodium pen-tobarbital . Since sleep or sedation is known to dramatically reduce cortical activity, it is not surprising that the circa 40-Hz AMFR was dif-ficult to measure in sleeping or sedated infants. So, the skepticism about the utility of the AMFR was partly due to its variability with behavioral state and also partly because it was tested in sedated or sleeping children .
Later, it was discovered that AMFRs to higher modulation frequencies (80-110 Hz) were relatively unaffected by sleep in adults (Cohen et al, 1991), and soon thereafter, a similar obser-vation was made in sedated or sleeping infants (Aoyagi et al, 1992 ; Levi et al, 1992). Although the amplitudes at these frequencies are smaller than those at low modulation frequencies, the noise level is also lower so that the detection of the AMFR is not compromised. We have also found that the AMFRs in the rabbit were less affected by behavioral state, anesthesia, stim-ulants, and cortical inactivation for higher mod-ulation frequencies.
The reasons for the superiority of the circa 80-Hz AMFRs over the circa 40-Hz AMFRs as an audiometric tool may be attributable to several factors . First, this higher modulation frequency may optimally drive subcortical sources that are relatively unaffected by behavioral state and anesthesia . Second, this frequency eliminates the cortical contribu-tions because cortical neurons are optimally driven by lower modulation frequencies. Third, sedation or sleep also eliminates cortical sources and may reduce extraneous noise. Fourth, the smaller size of the infant's brain may increase the detectability of subcortical activity compared with the larger adult brain. Fifth, since the detectability of the AMFR depends on the number of averages, twice the time is needed for the same number of aver-ages for a 40-Hz response compared with an 80-Hz response . In this way, less time is
202

Sources of the Scalp-Recorded AMFR/Kuwada et al
required to detect high than low modulation frequencies . Finally, the developing cortex may be poor at encoding any modulation fre-quencies, whereas subcortical structures may be closer to their adult abilities .
Joris PX, Yin TCT. (1992) . Response to amplitude-modulated tones in the auditory nerve of the cat. JAcoust Soc Am 91:215-232 .
Kiren T, Aoyagi M, Furuse H, Koike Y. (1994). An ex-perimental study on the generators of the amplitude-modulation following response . Acta Otolaryngol Suppl (Stockh) 511 :28-33 .
Acknowledgment . We thank Dr. Ahmed Khan for par-ticipation in the behavioral stimulation experiments and Dr. Sean J. O'Connor and Ms . Nancy Kluck for partici-pation in the cocaine experiments. This work was supported by National Institutes of Health grant R01 DC 01366 to S. Kuwada . The data in this manuscript were presented orally at the International Evoked Response Audiometry Study Group conference held in Vancouver, British Columbia, July 22-27, 2001.
REFERENCES
Aoyagi M, Kiren T, Kim Y, et al . (1992). Optimal modu-lation for amplitude-modulation following response in young children during sleep. Hear Res 65:253-261 .
Aoyagi M, Yamazaki Y, Yokota M, et al . (1996) . Frequency specificity of 80-Hz amplitude-modulation following response . Acta Otolaryngol Suppl (Stockh) 522:6-10.
Barker JL, Ransom BR. (1978) . Pentobarbitone phar-macology of mammalian central neurones grown in tissue culture. J Physiol (Lond) 280:355-372 .
Batra R, Kuwada S, Maher V. (1986) . The frequency-following response to continuous tones in humans . Hear Res 21:167-177 .
Batra R, Kuwada S, Stanford TR. (1989) . Temporal coding of envelopes and their interaural delays in the inferior colliculus of the unanesthetized rabbit. J Neurophysiol 61:257-268 .
Bures J, Buresova O, Krivanek J. (1974) . The Mechanism and Application of Leao's Spreading Depression of Electroencephalographic Actiuity . Prague : Academia .
Cohen LT, Rickards FW, Clark GM. (1991)Acomparison of steady-state evoked potentials to modulated tones in awake and sleeping humans . J Acoust Soc Am 90:2467-2479 .
Dolphin WF, Mountain DC . (1992) . The envelope follow-ing response : scalp potentials elicited in the Mongolian gerbil using sinusoidally AM acoustic signal . Hear Res 58:70-78 .
Fitzpatrick DC, Kuwada S, Batra R. (2000), Neural sen-sitivity to interaural time differences: beyond the Jeffress model. JNeurosci 20:1605-1615 .
Frisina RD . (2001) . Subcortical neural coding mecha-nisms for auditory temporal processing . Hear Res 158:1-27 .
Galambos R, Makeig S, Talmachoff P. (1981) . A 40 Hz auditory potential recorded from the human scalp. Proc Natl Acad Sci U S A 78:2643-2647 .
Griffiths SK, Chambers RD . (1991) . The amplitude modulation-following response as an audiometric tool . Ear Hear 12:235-241 .
Krishna BS, Semple MN . (2000) . Auditory temporal pro-cessing: responses to sinusoidally amplitude-modulated tones in the inferior colliculus . JNeurophysiol 84:255-273 .
Kuwada S, Batra R, Maher V (1986) . Scalp potentials of normal and hearing-impaired subjects in response to sinusoidally amplitude-modulated tones. Hear Res 21 :179-192 .
Kuwada S, Batra R, Stanford TR . (1989) . Monaural and binaural response properties of neurons in the inferior colliculus of the rabbit : effects of sodium pentobarbital . JNeurophysiol 61 :269-282 .
Kuwada S, Batra R. (1999) . Coding of sound envelopes by inhibitory rebound of the superior olivary complex in the unanesthetized rabbit. J Neurosci 19:2273-2287 .
Leao AA . (1944) . Spreading depression of activity in the cerebral cortex. JNeurophysiol 7:359-390 .
Levi EC, Folsom RC, Dobie RA . (1992) . Amplitude-modulation following response (AMFR) : effects of modulation rates, carrier frequency, age and state. Hear Res 68:42-52 .
Moller AR . (1974). Coding of units in the cochlear nucleus to sinusoidally amplitude-modulated tones. Exp Neural 45:104-117 .
Nedergard M, Hansen AJ . (1988) . Spreading depression is not associated with neuronal injury in the normal brain. Brain Res 449:395-398 .
Picton TW, Skinner CR, Champagne SC, et al . (1987) . Potentials evoked by sinusoidal modulation of the ampli-tude or frequency of a tone . JAcoust Soc Am 82:165-178 .
Rees A, Moller AR . (1983). Responses of neurons in the inferior colliculus of the rat to AM and FM tones. Hear Res 10 :301-330 .
Rees A, Green GGR, Kay RH . (1986). Steady-state evoked responses to sinusoidally amplitude-modulated sounds recorded in man. Hear Res 23 :123-133 .
Rhode WS . (1976) . A digital system for the auditory neu-rophysiological research . In : Brown P, ed . Current Computer Technology in Neurobiology . Washington, DC : Hemisphere, 543-567.
Rhode WS, Greenberg S. (1994) . Encoding of amplitude modulation in the cochlear nucleus of the cat . J Neurophysiol 56 :261-286 .
Rickards FW, Clark GM. (1984) . Steady-state evoked potentials to amplitude-modulated tones. In : Nodar RH, Barber C, eds. Euoked Potentials H. Boston : Butterworths, 163-168.
Rickards FW Tan LE, Cohen LT, et al . (1994) . Auditory steady-state evoked potentials in the newborns . Br J Audiol 28:327-337 .

Journal of the American Academy of Audiology/Volume 13, Number 4, April 2002
Schreiner CE, Urbas JV (1986) . Representation of ampli-tude modulation in the auditory cortex of the cat. I . The anterior auditory field (AAF) . Hear Res 21:227-241 .
Schreiner CE, Urbas JV (1988) . Representation of ampli-tude modulation in the auditory cortex of the cat. II . Comparison between cortical fields . Hear Res 32 :49-64 .
liculus and thalamus of the unanesthetized rabbit . J Neurosci 12 :3200-3216 .
Tielen AM, Kamp A, Lopes da Silva FH, et al . (1969) . Evoked responses to sinusoidally modulated sounds in unanesthetized dogs . Electroencephalogr Clin Neurophysiol 26:381-394 .
Stanford TR, Kuwada S, Batra R . (1992) . Comparison of interaural time sensitivity of neurons in the inferior col-
Tsuzuku T. (1993). 40-Hz steady state response in awake cats after bilateral chronic lesions in auditory cortices or inferior colliculi . Auris Nasus Larynx (Tokyo) 20:263-274 .