solving nonlinear systems of first order ordinary ...fhegedus/00 - numerics/2013 solving... ·...
TRANSCRIPT

Received April 5, 2013, accepted May 27, 2013, date of publication June 17, 2013, date of current version July 1, 2013.
Digital Object Identifier 10.1109/ACCESS.2013.2269192
Solving Nonlinear Systems of First OrderOrdinary Differential Equations Using a GalerkinFinite Element MethodAHMAD AL-OMARI1, HEINZ-BERND SCHÜTTLER2, JONATHAN ARNOLD3, AND THIAB TAHA41Institute of Bioinformatics, University of Georgia, Athens, GA 30602, USA2Department of Physics and Astronomy, University of Georgia, Athens, GA 30602, USA3Department of Genetics, University of Georgia, Athens, GA 30602, USA4Department of Computer Science, University of Georgia, Athens, GA 30602, USA
Corresponding author: A. Al-Omari ([email protected])
This work was supported in part by the NSF under Grants NSF QSB-0425762 and the NSF DBI-1062213 and the Department of SystemsEngineering and Medical Bioinformatics, Yarmouk University, Irbid, Jordan.
ABSTRACT A new numerical technique to solve nonlinear systems of initial value problems for nonlinearfirst-order differential equations (ODEs) that model genetic networks in systems biology is developed. Thistechnique is based on finding local Galerkin approximations on each sub-interval at a given time grid ofpoints using piecewise hat functions. Comparing the numerical solution of the new method for a singlenonlinear ODE with an exact solution shows that this method gives accurate solutions with relative error1.88 × 10−11 for a time step 1 × 10−6. This new method is compared with the adaptive Runge Kutta(ARK) method for solving systems of ODEs, and the results are comparable for a time step 2 × 10−4. Itis shown that the relative error of the Galerkin method decreases approximately linearly with the log of thenumber of hat functions used. Unlike the ARKmethod, this newmethod has the potential to be parallelizableand to be useful for solving biological problems involving large genetic networks. An NSF commis-sioned video illustrating how systems biology helps us understand that a fundamental process in cells isincluded.
INDEX TERMS Biological clock, Galerkin method, finite element method, hat function, Newton–Raphsonmethod, ordinary differential equation, toggle switch, systems biology.
I. INTRODUCTIONIn the new cross-disciplinary field of systems biology merg-ing genomics, bioinformatics and engineering the focus ison using networks of genes and their products to predictfundamental processes in the cell [1]. The field began inthe 1990s with the assembly of biochemical pathways todescribe the functioning of entire cells [2]–[4]. The field wastransformedwith the development of new genomics technolo-gies [1], [5], [6] to measure how many genes and proteinsbehave simultaneously in cells.We are now poised to describethe cellular dynamics of an entire cellular network [7], [8].The challenge is to be able to simulate such large networks.The dynamics of these cellular networks are often describedby very large systems of ordinary differential equations [9].One of the major problems in systems biology is solvinglarge systems of ordinary differential equations describinghow genetic networks behave [10], a challenge arising in
other areas of science and engineering as well [11]. TheGalerkin method has been employed for solving differentkinds of ordinary differential equations [12]–[19]. Here weshow how Galerkin’s method can be used in conjunctionwith Finite Element Method (FEM) piecewise hat functionsto solve systems of nonlinear first-order ordinary differentialequations (ODEs). Here our method is applied to systemsof ODEs describing several genetic networks [20], [21]. Theimportance of these networks to our daily lives is summarizedin anNSF commissioned video attached [22]. The idea behindthe method is to find local Galerkin approximations to thesolutions of the ODEs on each sub-interval of a given meshusing a collection of hat functions. In addition to the fact thatthis method is a newmethod for solving any nonlinear systemof ODEs with high accuracy and stability that is comparablewith the ARK method, it has the potential to be parallelizableand to be useful for solving biological problems that depend
408 2169-3536/$31.00 2013 IEEE VOLUME 1, 2013

A. Al-Omari et al.: Solving Nonlinear Systems of First Order Ordinary Differential Equations
on solving systems of nonlinear ODEs modeling geneticnetworks [10]. Since the data to identify such networks aresparse and noisy (error being 10% of values measured orlarger), such biological problems can be solved quickly andwith acceptable accuracy and high stability when a smallnumber of hat functions is used as shown in (Fig. 1). Thehigh accuracy of the ARK method, as an example, is notneeded for these biological problems [23]. Our new approachachieves the required biological accuracy and if so desired,gives results as accurate as theARKmethod. The basic idea ofthe new approach is to approximate each element in the solu-tion of a system of nonlinear first-order ODEs by a piecewisehat function on one subinterval at a time. In this paper, thismethod is illustrated by solving an initial value problem of:1) a single nonlinear first-order differential equation; 2)a system of nonlinear first-order differential equations fora genetic network describing the toggle switch [24]; and3) a system of nonlinear first-order differential equationsfor a genetic network for the biological clock of the modelfungal system,Neurospora crassa [25] described in the video.The latter two initial value problems are central to systemsbiology. It is to be noted that parallelizing the computa-tion of the Jacobian matrix and the integration functionsin the Galerkin method can speed up the numerical com-putations of a system of nonlinear first-order differentialequations.
FIGURE 1. Display of several hat functions.
II. NUMERICAL METHODSThe Galerkin method is a very popular method for findingnumerical solutions to partial differential equations. As anexample for the Finite Element Method (FEM), we use theGalerkin method to approximate the solution of ordinary dif-ferential equations with a piecewise linear function as a sumof basis functions (Hat Functions). By using FEM and a weakformulation of the approximation method, which transfers theproblem from a system of ODEs to a system of algebraicequations, we find the solution for the ODEs by solvingthese algebraic equations using the Newton-Raphsonmethod.Three initial value problems are considered to show the accu-racy of our method. The first problem involves solving only asingle nonlinear first-order differential equation, and the othertwo cases involve solving two systems of nonlinear first-orderdifferential equations. In the first example the new method isas accurate as the ARK method, and the other two examplessolutions by the new method are comparable with the ARKmethod.
A. GALERKIN ALGORITHM FOR SOLVING SYSTEMS OFODEsA system of initial-value problems for nonlinear first orderODEs over a solution’s interval [0 L] can be defined as
y′=V (y, t); where y =
y1y2·
·
·
yS−1yS
, V =
V1V2·
·
·
VS−1VS
, y(a) = b;
where S is the number of variables in a system of ODEs andin particular, the number of molecular species in a geneticnetwork.Note that a single nonlinear first order ODE problem con-
sidered above can be solved as a special case of the abovesystem.An approximate solution is expanded in terms of basis
functions φj (t) as
yn (t) =N∑j=0
pn,jφj(t) (1)
N is the number of hat functions; pnj is a vector of unknownexpansion amplitudes that we are solving for; and n labels thedifferent molecular species; and the φj (t) is a finite-elementbasis function (hat function) defined on a grid of time pointstj by
φj (t) =
t−tj−1tj−tj−1
, tj−1 ≤ t ≤ tjtj+1−ttj+1−tj
, tj ≤ t ≤ tj+10, otherwise
For example, the initial condition of the first species is givenby y1(0) = p1,0 and the solution for a specific species (n)at a specific time point (j) is given by yn(tj) = pn,j, sinceφj(tj)= 1 and φk
(tj)= 0 for k 6= j.
An alternative to hat functions is using compactly sup-ported wavelets [26] or other types of finite-element basisfunctions, such as Hermite finite elements [23].
Using the residual form
Rn(t) =N∑j
pn,jφ′
j (t)− Vn(Yn(t), t) (2)
we impose a weak Galerkin formulation of an approximatesolution to solve for pnj as weight variables
0 =∫ tk+1
tk−1φk (t)Rndt. (3)
B. The Algorithm
0 =∫ tk+1
tk−1φk (t)Rndt (4)
VOLUME 1, 2013 409

A. Al-Omari et al.: Solving Nonlinear Systems of First Order Ordinary Differential Equations
For arbitrary pnj
Fk,n(pn,j) =∫ tk+1
tk−1φk (t)Rn(t)dt (5)
j = k − 1, k, k + 1; k = 1, . . . ,L − 1; n = 1, . . . , S (S is thenumber of species).
Define for a given fixed k; pk , pk−1 and pk+1; wherep = pk+1
f (p) =
Fk,1(pk−1, pk , p)Fk,2(pk−1, pk , p)
.
.
.
Fk,S (pk−1, pk , p)
≡f1(p)f2(p).
.
.
fS (p)
where p1,k are the solution points for the first ODE (speciesf1), pS,k is the solution points for the last ODE (species fS );k = 1, . . . ,L − 1.
Solving for f(p)= 0 is done by using the Newton-Raphsonmethod [27]. The procedure for a Newton-Raphson schemefor solving this system of nonlinear algebraic equations canbe described by
1) Setting the initial iteration value to zero and assigninginitial values for each variable,
2) Calculating the Jacobian matrix J ,
J (p) =
∂f1∂p1
∂f1∂p2
...∂f1∂pN
∂f2∂p1
∂f2∂p2
...∂f2∂pN
.
.
.
.
.
.
.
.
.
.
.
.∂fS∂p1
∂fS∂p2
...∂fS∂pN
3) Using the Newton-Raphson scheme to solve the sys-
tem of algebraic equations, which is obtained fromf(p) = 0, the solution is defined by
pi+1 = pi − Gi (6)
Gi = J−1(pi)f (pi) (7)
J (pi)Gi = f (pi) (8)
The matrix Gi can be obtained by using the Gaussianelimination method with scale partial pivoting [27].A central finite-difference formula has been used tofind an approximation to the partial derivatives of theJacobian matrix.For example, calculating a given value in the Jacobianis done by
∂fr∂pu∼=1pfr1pu
=fT (pu + δ)− fT (pu − δ)
2δ;T = 0 . . . S, u = 0 . . .N (9)
The δ has been assigned a small value such as 1×10−4,so that the final ODEs solutions for the above systemdo not change much by further reducing the δ value.
4) Calculating fT (pu + δ), fT (pu − δ) by using the Gaussquadrature rule [27] since fT is an integration functionin this approach.
5) Iterating until the convergence of all variables isachieved. Tolerance of 1 ×10−6 is sufficient for solv-ing the above system by using the Newton-Raphsonmethod.
6) Note that solving such a system of algebraic equationsis obtained sequentially. For example, the solution ofthe vector f (pk+1) at fixed time is obtained from thesolution of the vectors f (pk−1) and f (pk ).What makes this Galerkin approach so attractive is thestability properties of the algorithm and the ability tocontrol rigorously the error [14], [15], [18], [19].
In (Fig. 2) the solution of such systems is shown for thefirst initial value problem described below using our proposedmethod, and the solution oscillates around the exact solution.Therefore, to achieve a reasonably accurate solution withthe lowest possible number of hat functions, we proposethat the initial guess for the Newton-Raphson method on theoscillation time to be the average of pk−1 and pk insteadof just pk. We check for an oscillation on pk−2, pk−1 andpk and we assign the average of pk−1 and pk to pk if theoscillation happens on these points. The accuracy of the finalsolution is based on the number of hat functions used forthe solution (as the number of hat functions increases, theaccuracy increases).
FIGURE 2. The solutions for the single nonlinear ODE and the effect ofaveraging on the numerical solution.
(Fig. 2) below shows part of the solution for the nonlinearsingle ODE during the interval [0 10] using 1000 hat func-tions. The numerical solution without averaging is shown inblue, the exact solution in red, and the solution with averagingin green.
C. THE THREE INITIAL VALUE PROBLEMS CONSIDEREDARE1) An initial value problem of a single nonlinear first ODE
y′ = −y− y2; (10)
with intial condition y(0) = 1
410 VOLUME 1, 2013

A. Al-Omari et al.: Solving Nonlinear Systems of First Order Ordinary Differential Equations
that has the exact solution:
y(t) = 1/(−1+ 2et ) (11)
2) An initial value problem of a system of nonlinear firstorder ODEs for a genetic network of the toggle switch[24] as specified by
du/dt = α1/(1+ vβ )− u (12)
dv/dt = α2/(1+ uγ )− v (13)
where; u(0)= 0;v(0)= 0;α1 = 2, α2 = 4, β = 2, γ = 2
∝1= 2,∝2= 4, β = 2, γ = 2
3) An initial value problem of a system of nonlinear firstorder ODEs for a genetic network of the biologicalclock of Neurospora crassa [25] as specified by
f ′1 = A(fG-f1)wn− A′f1
f′r = S3(fG-f1)+ S4f1-D3frf′p = L3fr-D6fp
w′ = E2up-D8w-nA(fG − f1)wn+ nA′f1-Pwf
mp
u′p = L1ur1-D4up-E2up
u′r1 = C1ur0fp-D7ur1u′r0 = V1-D1ur0-C1ur0fp
All of the parameter values of this clock network problemare given in Tables 1 and 2 [25].
III. RESULTS AND DISCUSSIONSThe new method yields solutions for fixed and specific timesteps, and the accuracy is as high as the ARKmethod if a largenumber of hat functions are considered as shown in Table 3and (Fig. 3). On the other hand, the accuracy of the solution
FIGURE 3. Maximum relative error (on a log scale) decreases linearly withthe log of the number of hat functions for the single nonlinear ODEcompared with the exact solution. Different numbers of hat functions(1 ×102, 1 ×103, 1 ×104, 1 ×105, 1 ×106, and 1 ×107) over the solutioninterval [0 10] are used.
TABLE 1. Initial conditions at t = 0 for clock.
TABLE 2. Parameters values.
TABLE 3. Errors in the solution over the time period [0 10] are shown as afunction of the number of hat functions for the single ODE (Example 1).The maximum relative errors have been calculated using differentnumbers of hat functions and the solutions compared with the ARKmethod (with absolute and relative errors for ARK equal to 1 ×10−13).
is still acceptable for biological problems if a fewer numberof hat functions is considered as shown in (Fig. 4–Fig. 5).The maximum global relative error has been computed for
the single nonlinear first ODE, for the genetic network of thetoggle switch, and for the genetic network of the biologicalclock of Neurospora crassa using the following formula:
Max global relative error (14)
= Maxk2Maxn|Y
Aprx.k (tn)− Y Exactk (tn)|
Maxn|YAprx.k (tn)| +Maxn|Y Exactk (tn)|
(15)
VOLUME 1, 2013 411
A. Al-Omari et al.: Solving Nonlinear Systems of First Order Ordinary Differential Equations
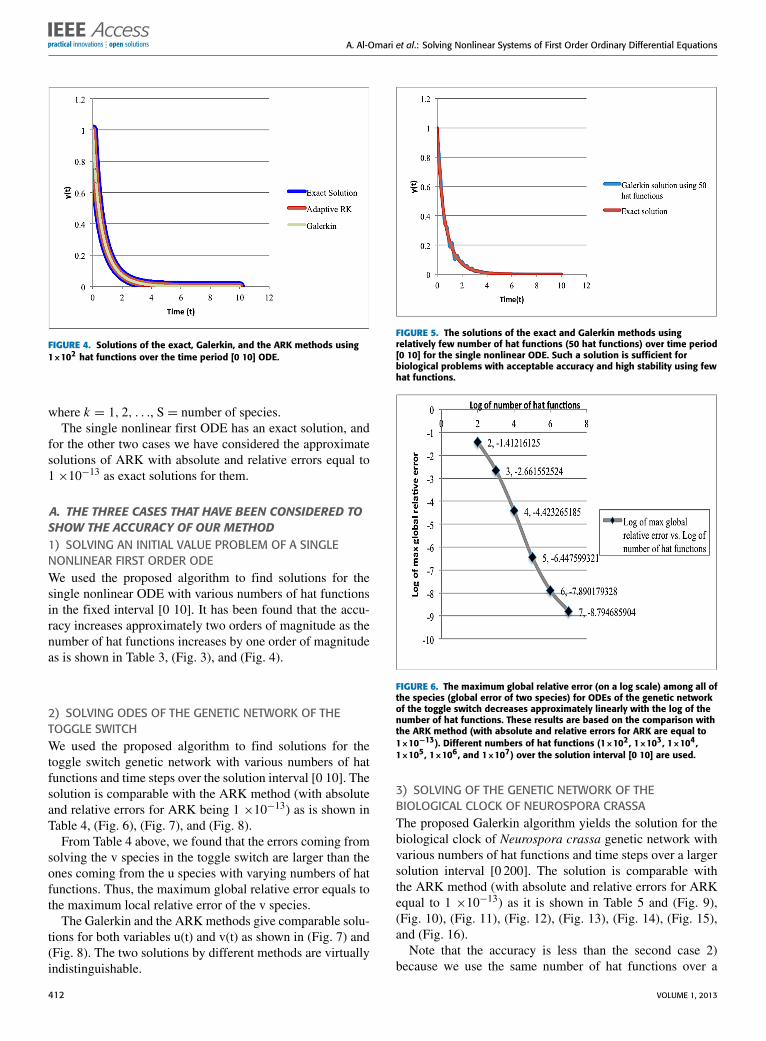
FIGURE 4. Solutions of the exact, Galerkin, and the ARK methods using1×102 hat functions over the time period [0 10] ODE.
where k = 1, 2, . . ., S = number of species.The single nonlinear first ODE has an exact solution, and
for the other two cases we have considered the approximatesolutions of ARK with absolute and relative errors equal to1 ×10−13 as exact solutions for them.
A. THE THREE CASES THAT HAVE BEEN CONSIDERED TOSHOW THE ACCURACY OF OUR METHOD1) SOLVING AN INITIAL VALUE PROBLEM OF A SINGLENONLINEAR FIRST ORDER ODEWe used the proposed algorithm to find solutions for thesingle nonlinear ODE with various numbers of hat functionsin the fixed interval [0 10]. It has been found that the accu-racy increases approximately two orders of magnitude as thenumber of hat functions increases by one order of magnitudeas is shown in Table 3, (Fig. 3), and (Fig. 4).
2) SOLVING ODES OF THE GENETIC NETWORK OF THETOGGLE SWITCHWe used the proposed algorithm to find solutions for thetoggle switch genetic network with various numbers of hatfunctions and time steps over the solution interval [0 10]. Thesolution is comparable with the ARK method (with absoluteand relative errors for ARK being 1 ×10−13) as is shown inTable 4, (Fig. 6), (Fig. 7), and (Fig. 8).
From Table 4 above, we found that the errors coming fromsolving the v species in the toggle switch are larger than theones coming from the u species with varying numbers of hatfunctions. Thus, the maximum global relative error equals tothe maximum local relative error of the v species.
The Galerkin and the ARKmethods give comparable solu-tions for both variables u(t) and v(t) as shown in (Fig. 7) and(Fig. 8). The two solutions by different methods are virtuallyindistinguishable.
FIGURE 5. The solutions of the exact and Galerkin methods usingrelatively few number of hat functions (50 hat functions) over time period[0 10] for the single nonlinear ODE. Such a solution is sufficient forbiological problems with acceptable accuracy and high stability using fewhat functions.
FIGURE 6. The maximum global relative error (on a log scale) among all ofthe species (global error of two species) for ODEs of the genetic networkof the toggle switch decreases approximately linearly with the log of thenumber of hat functions. These results are based on the comparison withthe ARK method (with absolute and relative errors for ARK are equal to1×10−13). Different numbers of hat functions (1×102, 1×103, 1×104,1×105, 1×106, and 1×107) over the solution interval [0 10] are used.
3) SOLVING OF THE GENETIC NETWORK OF THEBIOLOGICAL CLOCK OF NEUROSPORA CRASSAThe proposed Galerkin algorithm yields the solution for thebiological clock of Neurospora crassa genetic network withvarious numbers of hat functions and time steps over a largersolution interval [0 200]. The solution is comparable withthe ARK method (with absolute and relative errors for ARKequal to 1 ×10−13) as it is shown in Table 5 and (Fig. 9),(Fig. 10), (Fig. 11), (Fig. 12), (Fig. 13), (Fig. 14), (Fig. 15),and (Fig. 16).
Note that the accuracy is less than the second case 2)because we use the same number of hat functions over a
412 VOLUME 1, 2013

A. Al-Omari et al.: Solving Nonlinear Systems of First Order Ordinary Differential Equations
TABLE 4. Errors in the solution over the time period [0 10] are shown as a function of number of hat functions for the toggle switch (Example 2). The maxlocal and global relative errors have been calculated using different number of hat functions and the solutions compared with the ARK method (withabsolute and relative errors for ARK equal to 1 ×10−13).
TABLE 5. Maximum local relative errors of the Galerkin solution over the time period [0 200] for varied numbers of hat functions for the clock model(Example 3). The maximum local and global relative errors have been calculated using different numbers of hat functions and the solutions, comparedwith the ARK method (with absolute and relative errors for ARK equal to 1 ×10−13).
FIGURE 7. The solutions of u(t) using the Galerkin method with 1× 102
hat functions and the ARK method over the time period [0 10] for thegenetic network of the toggle switch.
larger solution interval [0 200] for the genetic network of thebiological clock ofNeurospora crassa instead of [0 10] for theformer two cases. On the other hand, we still can see in Table 5that a total of 10,000 hat functions is sufficient to obtain arelative error that is 0.07 or 7% or less. Again in (Fig. 9) thereis a linear relation between the maximum global relative errorand the number of hat functions on a log-log plot.
The solutions for this dynamical system in (Fig. 10),(Fig. 11), (Fig. 12), (Fig. 13), (Fig. 14), (Fig. 15), and (Fig. 16)using the Galerkin and ARK methods are indistinguishableusing 1000 hat functions. Although the max global relativeerror using 1000 hat functions over the interval [0 200] is
FIGURE 8. The solutions of v(t) using the Galerkin method with 1 ×102
hat functions and the ARK method over the time period [0 10] for thegenetic network of the toggle switch.
of order of 30% as it is shown in Table 5, in the figureswe show that the solution is sufficiently good for biologicalproblems.
B. A POTENTIAL PARALLELIZATION SCHEME FOR THEGALERKIN METHODUnlike the ARK method, which is inherently sequential forsolving systems of ODEs, the Galerkin method as statedbefore can be parallelized by parallelizing the Jacobianmatrix’s calculation and the integration functions. This par-allelization will speed up the numerical method of solving asystem of nonlinear first-order differential equations. More-
VOLUME 1, 2013 413

A. Al-Omari et al.: Solving Nonlinear Systems of First Order Ordinary Differential Equations
FIGURE 9. The maximum global relative error (on a log scale) among thewhole species (seven species) for ODEs of the genetic network of thebiological clock of Neurospora crassa decreases approximately linearlywith the log of the number of hat functions compared with the ARKmethod (with absolute and relative errors for ARK equal to 1×10−13).Different numbers of hat functions (2×102, 1×103, 1×104, 1×105, 1×106,and 1×107) over solution interval [0 200] are used.
FIGURE 10. The solution of f1(t) using the Galerkin and the ARK methodusing 1 ×103 hat functions over the time period [0 200] for the geneticnetwork of the biological clock of Neurospora crassa.
over, this new method allows us to parallelize the ensemblemethod [25] for identifying genetic networks from real dataon each variable (or species). Briefly, the ensemble methodsuggests that instead of identifying one unique parameteriza-tion of the model, we aim to identify an ensemble of modelsconsistent with available experimental data and use MonteCarlo simulation techniques to generate random samples ofmodel parameterizations (an ensemble) that represent thedata well. This sampling process is captured in an animationwithin the associated NSF commissioned video [22]. In otherwords, in the ODE solving scenario a unique solution willbe found by specifying the initial conditions. In contrast, inthe ensemble method since we don’t know the initial con-
FIGURE 11. The solution of fr(t) using the Galerkin and the ARK methodsusing 1 ×103 hat functions over the time period [0 200] for the geneticnetwork of the biological clock of Neurospora crassa.
FIGURE 12. The solution of fp(t) using the Galerkin and the ARK methodsusing 1 ×103 hat functions over the time period [0 200] for the geneticnetwork of the biological clock of Neurospora crassa.
ditions and other parameters values, which are required forsolving systems of ODE, a Monte Carlo procedure is usedto generate several initial conditions and parameters values,and while the Monte Carlo runs, it finds parameters thatmake the predicted solution closer to the experimental data.Finding the parameters could be done by using theMetropolisprocedure [25], which minimizes the Chi-squared statisticcomparing the experimental data and the predicted solution.Mainly, there are two stages in the ensemble method: theequilibration stage that is used to find parameters values thatmake the ODE solution converge to the experimental dataand the accumulation stage, which is used to accumulatemany sets of these parameters (i.e., the ensemble) that fitthe experimental data well. Averaging over the ensembleallows an assessment of fit to the experimental data. Thus,averaging several solutions of the ODEs with different initialconditions that fit the experimental data will be found from arandom sample of parameters that reproduce the experimentaldata.
414 VOLUME 1, 2013

A. Al-Omari et al.: Solving Nonlinear Systems of First Order Ordinary Differential Equations
FIGURE 13. The solution of w(t) using the Galerkin and the ARK methodsusing 1×103 hat functions over the time period [0 200] for the geneticnetwork of the biological clock of Neurospora crassa.
FIGURE 14. The solution of up(t) using the Galerkin and the ARK methodsusing 1×103 hat functions over the time period [0 200] for the geneticnetwork of the biological clock of Neurospora crassa.
UsingARK in the ensemblemethod implies that the systemof ODEs should be re-solved for each proposed ensembleMonte Carlo updating step, and solving for the time step t+hrequires the solution at the prior t. To apply a parallelizedGalerkin method version instead of the ARKmethod version,suppose there are (n) hat functions subintervals with (k) testgrid points. The purpose of these test grid points is to samplethe quality of the solution, for each of the subintervals asshown in (Fig. 17). On the one hand, Monte Carlo simulationwill propose a set of parameters values and hat functionamplitudes, which are required for solving the system ofODEs using FEM explained in this paper, for each subinter-val. On the other hand, one or more subinterval(s) could beassigned to one slave processor that will solve for the systemof ODEs given these parameters on its subinterval(s). Thenfor each of these test grid points within this subinterval(s),the method evaluates the left hand side of the differentialequation and the right hand side of the differential equation
FIGURE 15. The solution of ur1(t) using the Galerkin and the ARKmethods using 1×103 hat functions over the time period [0 200] for thegenetic network of the biological clock of Neurospora crassa.
FIGURE 16. The solution of ur0(t) using the Galerkin and the ARKmethods using 1×103 hat functions over the time period [0 200] for thegenetic network of the biological clock of Neurospora crassa.
FIGURE 17. n subinterval with a hat function for each of thesesubintervals divided into k test grid points used to estimate the quality ofthe solution. One or more hat function(s) could be assigned to one slaveprocessor.
and from the difference between the left hand side and theright hand side will find the residual, which is given inEquation 13. After that each processor will calculate its chi-
VOLUME 1, 2013 415

A. Al-Omari et al.: Solving Nonlinear Systems of First Order Ordinary Differential Equations
square statistic and send the result back to a master processor.The master processor adds the resulting chi-squared statis-tics up and either accepts or rejects the proposed parame-ters and amplitudes based on the Metropolis procedure. Thepotential parallelizing procedures for the Galerkin methodare either through Message Passing Interface (MPI) or MPIwith Graphics Processing Units (GPUs). The result is a newparallel ensemble method, which we call the super-ensemblemethod [23] because it combines the Monte Carlo search forparameters with an approximation to the ODE solution (bythe Galerkin Method).
IV. CONCLUSIONA new method for solving systems of initial-value prob-lems for nonlinear First-order Ordinary Differential Equa-tions using the Galerkin finite elements method piecewise hatfunctions has been developed that gives as accurate a solutionas the Adaptive Runge Kutta method when a large numberof hat functions are used, and acceptable accuracy for thebiological problems when a fewer number of hat functionsis used. On the other hand, unlike the adaptive Runge Kuttamethod, this method has the potential to be parallelizable andto be useful for solving biological problems that depend onsolving large systems of nonlinear ODEs describing geneticnetworks and other systems in engineering. Moreover, thismethod yields solutions not for arbitrary time steps but fordesirable fixed time steps.
As shown above we produce trajectories from the GalerkinMethod comparable with the adaptive Runge Kutta method(with relative and absolute errors for the ARK are equal to1 ×10−13) by using a low number of hat functions (100hat functions for the first two cases over the interval [0 10]and 1000 hat functions for the last case over the interval[0 200]). Developing this method to be faster than the tradi-tional ODE solvers is a potential study in the future especiallywhen biological problems with large networks are consid-ered. Identifying large networks is complicated by havingmany parameters and limited data [28]. One solution to thisproblem is the use of ensemble methods [25]. A parallelizedODE solver enables faster sampling of the parameter spacein ensemble methods to identify what we know (i.e., is sup-ported across the ensemble) and what we do not know (i.e.,is not supported across the ensemble) about a large systemof ODEs.
REFERENCES[1] T. Ideker, V. Thorsson, J. A. Ranish, R. Christmas, J. Buhler, J. K.
Eng, R. Bumgarner, D. R. Goodlett, R. Aebersold, and L. Hood,‘‘Integrated genomic and proteomic analyses of a systematically per-turbed metabolic network,’’ Science, vol. 292, no. 5518, pp. 929–934,May 2001.
[2] I. M. Keseler, J. Collado-Vides, A. Santos-Zavaleta, M. Peralta-Gil, S.Gama-Castro, L. Muniz-Rascado, C. Bonavides-Martinez, S. Paley, M.Krummenacker, T. Altman, P. Kaipa, A. Spaulding, J. Pacheco, M. Laten-dresse, C. Fulcher, M. Sarker, A. G. Shearer, A. Mackie, I. Paulsen, R.P. Gunsalus, and P. D. Karp, ‘‘EcoCyc: A comprehensive database ofEscherichia coli biology,’’ Nucleic Acids Res., vol. 39, pp. D583–D590,Jan. 2011.
[3] S. Okuda, T. Yamada, M. Hamajima, M. Itoh, T. Katayama, P. Bork,S. Goto, and M. Kanehisa, ‘‘KEGG Atlas mapping for global analysisof metabolic pathways,’’ Nucleic Acids Res., vol. 36, pp. W423–W426,Jul. 2008.
[4] R. Overbeek, N. Larsen, G. D. Pusch, M. D’Souza, E. Selkov, Jr., N. Kyrpi-des, M. Fonstein, N. Maltsev, and E. Selkov, ‘‘WIT: Integrated system forhigh-throughput genome sequence analysis and metabolic reconstruction,’’Nucleic Acids Res., vol. 28, pp. 123–125, Oct. 2000.
[5] J. L. DeRisi, V. R. Iyer, and P. O. Brown, ‘‘Exploring the metabolic andgenetic control of gene expression on a genomic scale,’’ Science, vol. 278,pp. 26–37, Oct. 1997.
[6] S. J. Maerkl and S. R. Quake, ‘‘A systems approach to measuring thebinding energy landscapes of transcription factors,’’ Science, vol. 315,no. 5809, pp. 233–237, Jan. 2007.
[7] J. R. Karr, J. C. Sanghvi, D. N. Macklin, M. V. Gutschow, J. M. Jacobs, B.Bolival, Jr., N. Assad-Garcia, J. I. Glass, and M. W. Covert, ‘‘A whole-cellcomputational model predicts phenotype from genotype,’’ Cell, vol. 150,no. 2, pp. 389–401, Jul. 2012.
[8] M. W. Covert, E. M. Knight, J. L. Reed, M. J. Herrgard, and B. O. Palsson,‘‘Integrating high-throughput and computational data elucidates bacterialnetworks,’’ Nature, vol. 429, no. 6987, pp. 82–96, May 2004.
[9] A. K. Chakraborty and J. Das, ‘‘Pairing computation with experimentation:A powerful coupling for understanding T cell signalling,’’ Nature Rev.Immunol., vol. 10, no. 1, pp. 59–71, Jan. 2010.
[10] X. Tang, W. Dong, J. Griffith, R. Nilsen, A. Matthes, K. B. Cheng,J. Reeves, H. B. Schüttler, M. E. Case, and J. Arnold, ‘‘Systems biologyof the qa gene cluster in Neurospora crassa,’’ PloS ONE, vol. 6, no. 6, p.e20671, 2011.
[11] S. H. Strogatz, ‘‘Exploring complex networks,’’ Nature, vol. 410, pp. 268–276, Mar. 2001.
[12] M. Delfour, F. Trochu, and W. Hagar, ‘‘Discontinuous Galerkin methodsfor ordinary differential equations,’’ Math. Comput., vol. 36, no. 154, pp.455–473, 1981.
[13] M. Delfour and F. Dubeau, ‘‘Discontinuous polynomial approximations inthe theory of one-step, hybrid andmultistepmethods for nonlinear ordinarydifferential equations,’’ Math. Comput., vol. 47, pp. 169–189, Jul. 1986.
[14] C. Johnson, ‘‘Error estimates and adaptive time-step control for a class ofone-step methods for stiff ordinary differential equations,’’ SIAM J. Numer.Anal., vol. 25, no. 4, pp. 908–926, 1988.
[15] F. Dubeau, A. Ouansafi, and A. Sakat, ‘‘Galerkin methods for nonlinearordinary differential equation with impulses,’’ Numer. Algorithms, vol. 33,nos. 1–4, pp. 215–225, 2003.
[16] H. Temimi, S. Adjerid, and M. Ayari, ‘‘Implementation of the discontinu-ous Galerkin method on a multi-story seismically excited building model,’’Eng. Lett., vol. 18, no. 1, pp. 18–27, 2010.
[17] B. L. Hulme, ‘‘One-step piecewise polynomial Galerkin methods for initialvalue problems,’’Math. Comput., vol. 26, no. 118, pp. 415–426, 1972.
[18] D. Estep, ‘‘A posteriori error bounds and global error control for approxi-mation of ordinary differential equations,’’ SIAM J. Numer. Anal., vol. 32,no. 1, pp. 1–48, 1995.
[19] K. Bottcher and R. Rannacher, ‘‘Adaptive error control in solving ordi-nary differential equations by the discontinuous Galerkin method,’’ Inst.AppliedMathematics, Univ. Heidelberg, Heidelberg, Germany, Tech. Rep.,Feb. 1997.
[20] J. Arnold, R. T. Thiab, and L. Deligiannidis, ‘‘GKIN: A tool for drawinggenetic networks,’’ Netw. Biol., vol. 2, no. 1, pp. 26–37, 2012.
[21] B. Aleman-Meza, Y. Yu, H. B. Schüttler, J. Arnold, and T. R. Taha, ‘‘KIN-SOLVER: A simulator for computing large ensembles of biochemical andgene regulatory networks,’’ Comput. Math. Appl., vol. 57, no. 3, pp. 420–435, 2009.
[22] M. O’Brien and M. Walton. (2010). ‘‘Biological clocks,’’ Science Nation[Online]. Available: http://www.nsf.gov/news/special_reports/science_nation/biologicalclocks.jsp
[23] Y. Huang, ‘‘Parameter estimation of chemical reaction networks,’’ M.S.thesis, Dept. Phys. Astron., Univ. Georgia, Athens, GA, USA, 2007.
[24] T. S. Gardner, C. R. Cantor, and J. J. Collins, ‘‘Construction ofa genetic toggle switch in Escherichia coli,’’ Nature, vol. 403, no. 6767,pp. 339–342, 2000.
[25] Y. Yu, W. Dong, C. Altimus, X. Tang, J. Griffith, M. Morello,L. Dudek, J. Arnold, and H. B. Schüttler, ‘‘A genetic network for theclock of Neurospora crassa,’’ Proc. Nat. Acad. Sci., vol. 104, no. 8,pp. 2809–2814, 2007.
416 VOLUME 1, 2013

A. Al-Omari et al.: Solving Nonlinear Systems of First Order Ordinary Differential Equations
[26] I. Daubechies, ‘‘Orthonormal bases of compactly supported waveletsII. Variations on a theme,’’ SIAM J. Math. Anal., vol. 24, no. 2,pp. 499–519, 1993.
[27] E. W. Cheney and D. R. Kincaid, Numerical Mathematics and Computing.Pacific Grove, CA, USA; Brooks/Cole, 2012.
[28] H. de Jong and D. Ropers, ‘‘Strategies for dealing with incomplete infor-mation in the modeling of molecular interaction networks,’’ BriefingsBioinformat., vol. 7, no. 4,
AHMAD AL-OMARI received the B.Sc. degreein electrical and computer engineering fromYarmouk University, Irbid, Jordan, in 2004. Hewas a Teaching Assistant and a Lab Engineer withthe Department of Electrical and Computer Engi-neering, Yarmouk University, from 2004 to 2010.He worked on more than 30 different projects oncomputers, communication, and electronic engi-neering. He received a grant of $100 000 fromYarmouk University to pursue the Ph.D. degree in
bioinformatics. He is currently a Research Assistant and the Ph.D. degree inbioinformatics under the supervision of Prof. J. Arnold with the Universityof Georgia, Atlanta, GA, USA, on systems biology and genetic networks.His current research interests include parallel computation, systems biology,finite elements method, machine learning and pattern recognition, biologicalcircuits and gene networks, and numerical analysis.
HEINZ-BERND SCHÜTTLER received theDiploma degree in physics from Technische Uni-versität München, Garching, Germany, in 1981,and the Ph.D. degree in physics from the Uni-versity of California, Los Angeles, CA, USA, in1984. After post-doctoral appointments with theUniversity of California, Santa Barbara, CA, USA,and the Argonne National Laboratory, Lemont,IL, USA, he joined the Faculty of the Depart-ment of Physics and Astronomy, University of
Georgia, Atlanta, GA, USA, in 1987, where he currently serves as a Pro-fessor of physics. His current research interests include the applicationsof computational statistical mechanics in biology and condensed matterphysics. His recent notable contributions in computational biology includethe development of the ensemble network simulation (ENS) method for thereconstruction of biological circuits and ENS-based methods for maximallyinformative next experiment design.
JONATHAN ARNOLD received the B.S. degreein mathematics from Yale University, New Haven,CT, in 1975, and the M.Phil. and Ph.D. degrees instatistics from Yale University, in 1978 and 1982,respectively. In 1982, he joined the Departmentsof Statistics and Genetics, College of Arts and Sci-ences, University of Georgia, Atlanta, GA, USA,where he was a Professor of genetics, statistics,and physics and astronomy. He has authored or co-authored over 130 papers in journals, book chap-
ters, and conference proceedings in the subjects of statistical genetics, popu-lation genetics, computational biology, and fungal genomics. He has servedon the NSF Computational Biology Panel, DOE Genome Panel, and as aCharter Member on the NIH Genetic Variation and Evolution Study Sectionfrom 2005 to 2008. His current research interests include the developmentand identification of genetic networks of fundamental processes in the modelsystem Neurospora crassa, i.e., Computing Life. He was elected an AAASFellow in 2011.
THIAB TAHA received the Ph.D. degree fromClarkson University, Potsdam, NY, USA, in 1982.In 1982, he joined the Computer Science Depart-ment, University of Georgia, Atlanta, GA, USA.His current research interests include scientific anddistributed computing and software developmentfor solving problems in nonlinear waves, opti-cal fiber communication systems, and biochemicalreaction networks. He has authored more than 60research papers. He is a Senior Editor of Math-
ematics and Computers in Simulation Journal and the Vice President ofIMACS. He received the M. G. Michael Award for Research in the Sciencesat UGA in 1985 and was a Fulbright Scholar from 1995 to 1996. Currently,he is the Director of the CUDA Teaching Center at UGA.
VOLUME 1, 2013 417