sodininkystĖ ir darŽininkystĖ
TRANSCRIPT

LIETUVOS AGRARINIŲ IR MIŠKŲ MOKSLŲ CENTRO FILIALOSODININKYSTĖS IR DARŽININKYSTĖS INSTITUTO IR
ALEKSANDRO STULGINSKIO UNIVERSITETO MOKSLO DARBAISCIENTIFIC WORKS OF THE INSTITUTE OF HORTICULTURE, LITHUANIAN RESEARCH CENTRE FOR AGRICULTURE AND FORESTRY AND ALEKSANDRAS STULGINSKIS UNIVERSITY
SODININKYSTĖ IR DARŽININKYSTĖ
34(3–4)
Eina nuo 1983 m.Published since 1983
Babtai 2015

UDK 634/635 (06)
Redaktorių kolegijaEditorial BoardDr. Audrius SASNAUSKAS (LAMMC SDI, žemės ūkio mokslai, agronomija)prof. habil. dr. Pavelas DUCHOVSKIS (LAMMC SDI, žemės ūkio mokslai, agronomija) dr. Edite KAUFMANE (Latvija, Latvijos valstybinis sodininkystės institutasbiomedicinos mokslai, biologija)dr. Aleksandras KMITAS (ASU, žemės ūkio mokslai, agronomija) dr. Pranas VIŠKELIS (LAMMC SDI, žemės ūkio mokslai, agronomija)dr. Giedrė SAMUOLIENĖ (LAMMC SDI, žemės ūkio mokslai, agronomija)prof. habil. dr. Vidmantas STANYS (LAMMC SDI, žemės ūkio mokslai, agronomija)prof. habil. dr. Algirdas SLIESARAVIČIUS (ASU, žemės ūkio mokslai, agronomija)
Redakcinė mokslinė tarybaEditorial Scientific CouncilDr. Audrius SASNAUSKAS – pirmininkas (Lietuva), prof. habil. dr. Pavelas DUCHOVSKIS (Lietuva), dr. Kalju KASK (Estija), dr. Edite KAUFMANE (Latvija), prof. habil. dr. Zdzisław KAWECKI (Lenkija),dr. Giedrė SAMUOLIENĖ (Lietuva), prof. habil.dr. Albinas LUGAUSKAS (Lietuva), habil. dr. Maria LEJA (Lenkija), prof. habil. dr. Lech MICHALCZUK (Lenkija), prof. dr. Ala SILAJEVA (Ukraina), prof. habil. dr. Algirdas SLIESARAVIČIUS (Lietuva), prof. habil. dr. Vidmantas STANYS (Lietuva), prof. dr. Viktor TRAJKOVSKI (Švedija)
Redakcijos adresas:Lietuvos agrarinių ir miškų mokslų centro filialas Sodininkystės ir daržininkystės institutasLT-54333 Babtai, Kauno r. Tel. (8 37) 55 52 10Faksas (8 37) 55 51 76 El. paštas [email protected]
Address of the Editorial Office: Institute of Horticulture, Lithuanian Research Centre for Agriculture and Forestry LT-54333 Babtai, Kaunas district, Lithuania Phone +370 37 55 52 10 Telefax +370 37 55 51 76 E-mail [email protected]
Leidinio adresas internete www.lsdi.ltLeidinys cituojamas CAB Abstracts, EBSCO Publishing, VINITI duomenų bazėse
ISSN 0236-4212 © Lietuvos agrarinių ir miškų mokslų centro filialas Sodininkystės ir daržininkystės institutas, 2015 © Aleksandro Stulginskio universitetas, 2015

3
LIETUVOS AGRARINIŲ IR MIŠKŲ MOKSLŲ CENTRO FILIALO SODININKYSTĖS IR DARŽININKYSTĖS INSTITUTO IR ALEKSANDRO STULGINSKIO UNIVERSITETO MOKSLO DARBAI. SODININKYSTĖ IR DARŽININKYSTĖ. 2015. 34(3–4).
Paprastosios žemuogės derėjimas bei antocianinų kiekis ir sudėtis uogose vandens deficito sąlygomis
Rytis Rugienius, Tadeušas Šikšnianas, Vidmantas Bendokas, Vidmantas StanysLietuvos agrarinių ir miškų mokslų centro filialas Sodininkystės ir daržininkystės institutas, Kauno g. 30, LT-54333 Babtai, Kauno r., el. paštas [email protected]
Vidutinės oro temperatūros kilimas didina laistymo poreikį auginant uoginius augalus lauko ir šiltnamio sąlygomis. Nors vandens deficitas dažnai sumažina uogų derlių ir dydį, tačiau gali praturtinti jų biocheminę sudėtį, nes dėl patirto streso kaupiasi fenoliniai junginiai. Mūsų tyrimo tikslas buvo ištirti žemuogių derėjimą, antocianinų kiekį ir sudėtį uogose atskirų skynimų metu, augalus auginant įprastinėmis bei vandens deficito sąlygomis. 2013 ir 2014 m., žemuoges auginant šiltnamyje, kas 4–7 dienas derėjimo metu įvertinta vidutinė uogos masė bei derlius, aukšto slėgio chromatografijos metodu nustatytas antocianinų kiekis ir sudėtis. Tyrimų metu paaiškėjo, kad vandens deficito sukelto streso įtaka uogų derlingumo rodikliams – uogų dydžiui ir derliui – priklauso nuo vandens deficito laipsnio ir oro temperatūros. 2013 m. vandens normos sumažinimas 50 proc. derėjimo rodiklių nepaveikė, o 2014 m. – suvėlino uogų nokimą. 2013 m. išlaisčius tik 25 proc. vandens normos, uogų masė ir derlius derėjimo pabaigoje sumažėjo, 2014 m. – uogos padidėjo, bet gerokai sumažėjo derlius. Nustatyta, kad vandens deficitas 2013 m. atskirų skynimų metu padidino antocianinų kiekį žemuogių uogose iki dviejų kartų o 2014 m. – iki 38 procentų. Bendras antocianinų kiekio padidėjimas 2013 m. sudarė 45, o 2014 m. – 18 procentų. Jis buvo maksimalus, kai oro temperatūra buvo ne aukštesnė kaip 20 °C. Pelargonidin-3-glikozido ir cianidin-3-glikozido kiekis uogose derėjimo pradžioje sudarė atitinkamai 48 ir 36 proc. visų antocianinų. Šių antocianinų santykis derėjimo metu kito priklausomai nuo oro temperatūros ir augalo fiziologinės būklės.
Reikšminiai žodžiai: derlingumas, Fragaria vesca, lietinimas, oro temperatūra, pi-gmentai, sausra, šiltnamis.
Įvadas. Klimato kaitos sąlygomis netgi vidutinėje klimato juostoje, kurioje yra Lietuva, vis dažniau susiduriama su drėgmės trūkumu ir pernelyg aukšta tempera-tūra. Tai ypač svarbu auginant uoginius augalus šiltnamiuose, kur auginimo sąnau-dos yra daug didesnės negu lauke (Klamkowski, Treder, 2008). Drėgmės trūkumas mažina sodininkystės konkurencingumą ir verčia jos atsisakyti tose vietose, kur anksčiau ji buvo gana sėkminga. Šiuo metu pasaulyje intensyviai kuriamos nau-

4
jos auginimo technologijos kiek įmanoma mažinant vandens naudojimą (Giné Bor-donaba, Terry, 2009). Tai nėra lengva, nes sausros sukeliamas stresas dažnai turi neigiamą poveikį uoginių augalų derliui ir kokybei. Tačiau vandens trūkumas gali padidinti sausą masę ir kai kurių žmogaus sveikatai naudingų medžiagų kiekį (Kim ir kt., 2009; Giné Bordonaba, Terry, 2010). Todėl būtini detalesni augalų tyrimai sie-kiant ne tik sumažinti nuostolius dėl sausros sukeliamo streso, bet ir padidinti vaisių ir uogų maistines, juslines ir funkcines savybes (Nora ir kt., 2012). Yra žinoma, kad antocianinai kaupiasi vaisiuose ir uogose dėl įvairių stresą sukeliančių veiksnių: sausros, šalčio, vėjo, druskų ir kt. Flavonoidams priklausantys antocianinai yra na-tūralūs augalų pigmentai, ne tik suteikiantys spalvą, bet ir apsaugantys augalus nuo ultravioletinių spindulių, oksidacinio streso, mažinantys laisvųjų radikalų kiekį (Al-bert ir kt., 2009). Antocianinai svarbūs daugelio ligų, taip pat ir vėžio, profilaktikai (Zafra-Stone ir kt., 2007).
Tirtas antocianinų kaupimasis braškėse, giminingoje žemuogėms rūšyje, veikiant įvairiems aplinkos veiksniams (Klamkowski, Treder, 2008; Ghaderi ir kt., 2015). Lietuvoje antocianinai tirti serbentuose, vyšniose ir šilauogėse (Anisimovienė ir kt., 2009, 2013). Žemuogė dėl žinomos genetinės struktūros diploidinio genomo laikoma viena iš modelinių rūšių erškėtinių (Rosacea) šeimoje. Be to, susidomėjimas žemuo-gių auginimu Lietuvoje auga, vykdoma šių augalų selekcija (Rugienius ir kt., 2013). Iki šiol nepakanka duomenų apie žemuogių derlių ir antocianinų kaupimąsi uogose derėjimo metu. Dėl biologinių rūšies savybių derėjimas trunka nuo 3 iki 6 savaičių ir yra dažnai veikiamas nepastovių orų.
Darbo tikslas – ištirti žemuogių uogų derlių ir dydį, antocianinų kiekį ir sudėtį jose atskirų skynimų metu, augalus auginant įprastinėmis bei vandens deficito (kai lietinimo normos – 50 ir 25 proc.) sąlygomis. Šie tyrimai svarbūs kuriant tausojančias ir biocheminę uogų sudėtį gerinančias žemuogių ir braškių auginimo technologijas.
Tyrimo objektas, metodai ir sąlygos. Ž e m u o g i ų d e r ė j i m o v a n d e n s d e f i c i t o s ą l y g o m i s t y r i m a i. Tyrimai atlikti 2013–2014 m. Lietuvos agrari-nių ir miškų mokslų centro (LAMMC) filialo Sodininkystės ir daržininkystės instituto Sodo augalų genetikos ir biotechnologijos skyriuje. Tyrimui naudoti ‘Rojan’ veislės žemuogių (F. vesca var. semperflorens) augalai. Iš prieš metus sudaigintų sėklų iš-augę augalai buvo laikomi puodeliuose durpių substrate lauke. 2013 m. sausio mė-nesį žemuogės įneštos į nešildomą šiltnamį, kur pradėjo žydėti balandžio 2 dieną. 2014 m. augalai žiemojo ir augo lauke, žydėti pradėjo gegužės 5 dieną. Pradėjusios derėti žemuogės buvo perneštos į nešildomą šiltnamį. Bandymas pradėtas uogoms pradėjus derėti: 2013 m. – balandžio 22 d., 2014 m. – gegužės 29 dieną. Visiškai sunokusios uogos rinktos kas 4–7 dienas: 2013 m. – iki gegužės 25 d., 2014 m. – iki liepos 7 d., iš viso 6–8 kartus. Iki derėjimo pradžios augalai laistyti taikant normalų režimą. Vidutinė oro temperatūra lauke tyrimo metu svyravo nuo 12 iki 24 °C, mak-simali – nuo 14 iki 28 °C (1 pav.). Tiriant žemuogių derėjimą ir antocianinų kaupimą-si uogose drėgmės trūkumo sąlygomis, žemuogių augalai, augantys šiltnamyje, nuo derėjimo pradžios buvo laistomi taikant tris skirtingus režimus: 1) augalai, kaip ir anksčiau, laistyti pagal normalų režimą, esant poreikiui taip, kad substrato paviršius nuolat būtų drėgnas, išliejant vidutiniškai 470 ml vandens augalui; 2) išlaistyta pusė (50 proc.) normos, 3) išlaistyta ketvirtis (25 proc.) normos. Uogos buvo sveriamos,

5
užšaldomos ir laikomos –80 °C temperatūroje tolesniems antocianinų kiekio ir su-dėties tyrimams atlikti. Kiekviename iš trijų variantų buvo po 9 kiekvienos veislės augalus (trys pakartojimai po tris augalus).
1 pav. Vidutinė ir maksimali oro temperatūra lauke 2013 ir 2014 m.Fig. 1. Average and maximal air temperature outside in 2013 and 2014.
A n t o c i a n i n ų a n a l i z ė. Antocianinams išskirti naudoti 92 proc. koncentraci-jos metanolio tirpalai, parūgštinti HCl iki 0,1 N koncentracijos. Tiriamosios medžiagos ir ekstrahento santykis – 1 : 20 (g/ml) (Arnnok ir kt., 2012). Tirpalai koncentruoti evaporatoriuje 37 °C temperatūroje. Liekana ištirpinta parūgštintame vandenyje (pH 2,4). Antocianinų kokybinė ir kiekybinė sudėtis nustatyta taikant aukšto slėgio skysčių chromatografijos metodą. Skirstyta „Agilent 1200“ chromatografu su diodų matricos detektoriumi (bangos ilgis – 520 nm) ir „Eclipse XDB“ C18 kolonėle (4,6 × 150 mm) (Agilent Technologies Inc., Vokietija). Analizei atlikti naudotas judrios fazės (A – 10 proc. acto rūgštis, 1 proc. ortofosforo rūgštis, B – acetonitrilas) gradientas: 0 min. 95 proc. A, 5 min. 95 proc. A, 20 min. 75 proc. A, 25 min. 55 proc. A, 27 min. 95 proc. A ir 30 min. 95 proc. A, srauto greitis – 1 ml min.-1 (Durst, Wrolstad, 2001; Anisimovienė ir kt., 2013). Antocianinai identifikuoti ir jų kiekis nustatytas naudojant cianidin-3-O-glikozido (C3G), cianidin-3-O-rutinozido (C3R), cianidin-3-O-sambu-biozido (C3Sam), delfinidin-3-O-rutinozido (D3R), delfinidin-3-O-glikozido (D3G), pelargonidin-3-O-glikozido (Pel3G), peonidin-3-O-rutinozido (Peo3R) ir malvidin-3-O-glikozido (M3G) standartus. Tyrimams pasirinkti dažniausiai Fragaria gentyje aptinkami antocianinai. Bendras antocianinų kiekis apskaičiuotas sudėjus atskirų antocianinų kiekius.
Rezultatai. D e r l i n g u m o r o d i k l i ų k i t i m a s 2 0 1 3 m. Tyrimų re-zultatai rodo, kad derėjimo metu žemuogių vidutinė uogos masė mažėjo (2 pav. A). Palyginti su derėjimo pradžia, šis sumažėjimas sudarė 33 procentus. Pastebėta, kad

6
‘Rojan’ veislės vidutinė uogos masė, kai augalai laistyti panaudojant 50 proc. vandens normos, iš esmės nesiskyrė nuo normaliai (pagal poreikį) lietinamų augalų vidutinės uogos masės. Esant didesniam vandens deficitui (lietinimo norma – 25 proc.) nuo ketvirto skynimo (gegužės 3 d.) vidutinis uogos dydis svyravo: pradžioje sumažėjo, vėliau (gegužės 8 d.) – padidėjo, o dar vėliau (gegužės 21 d.) – gerokai sumažėjo. Normaliai laistytų žemuogių derlius iki ketvirto skynimo (gegužės 3 d.) buvo gana stabilus (2 pav. C).
2 pav. Žemuogių ‘Rojan’ vidutinė uogos masė (A ir B), vidutinis derlius iš augalo (C ir D) atskirų skynimų metu 2013 ir 2014 m.,
augalus laistant pagal poreikį (100 proc. normos), išlaistant pusę normos (50 proc.) ir ketvirtį normos (25 proc.).
Fig. 2. Average berry weight (A and B), average yield per plant (C and D) at separate harvests in 2013 and 2014, when plants were irrigated according to
requirements (100 % norm), half norm (50 %), quarter of norm (25 %).

7
Vėliau derlius svyravo, o septinto skynimo metu (gegužės 21 d.) gerokai pa-didėjo. Pastebėta, kad drėgmės trūkumas, kai lietinimo norma buvo 50 proc., uogų derliaus, kaip ir vidutinės uogos masės, nesumažino – jis išliko panašus kaip norma-liai laistytų augalų. Kai augalų laistymo norma buvo 25 proc., uogų derlius, kitaip nei augalus normaliai laistant ir laistymo normai esant 50 proc., nuo šešto skynimo (gegužės 13 d.) nebedidėjo: pasireiškė didelio vandens deficito neigiamas poveikis uogų skaičiui ir masei. Derliaus ir uogų dydžio svyravimai antroje derėjimo pusėje yra susiję su oro temperatūros kitimu. Ypatingos sąlygos susidarė 2013 m. gegužės 5–10 d., kai oro temperatūra pakilo iki 24–27 °C, buvo saulėta, todėl šiltnamyje oro temperatūra pakilo virš 35 °C. Iki gegužės 3 d. oro temperatūra svyravo tik apie 10–13 °C (1 pav.). Temperatūros šuolis prieš penktąjį skynimą (gegužės 8 d.) labai paveikė sunokusių uogų derlių ir jų dydžio rodiklius. Įdomu, kad tuo metu, normaliai laistant, vidutinė uogos masė sumažėjo, o laistymo normai esant 50 ir 25 proc., – pa-didėjo.
D e r l i n g u m o r o d i k l i ų k i t i m a s 2 0 1 4 m. Tyrimai atlikti mėnesiu vėliau negu 2013 m., nes augalai į šiltnamį įnešti tik prieš pat tyrimą, gegužės mėn. pabaigoje. Tyrimo rezultatai rodo, kad normaliai laistytų augalų vidutinė uogos masė iki trečio skynimo (birželio 13 d.) sumažėjo perpus (2 pav. B), o vėliau didėjo, bet pra-dinės uogos masės reikšmės nepasiekė ir derėjimo pabaigoje liko 38,5 proc. mažesnė. Vandens deficito sąlygomis augintų augalų antro ir trečio skynimo uogos vidutinė masė buvo gerokai didesnė negu normaliai laistytų augalų: atitinkamai 42,3 ir 57,7 proc., išlaisčius 25 proc. normos, ir atitinkamai 20 ir 33 proc., išlaisčius 50 proc. normos.
2014 metais normaliai laistytų augalų derėjimo maksimumas, kitaip nei 2013 metais, pasiektas pirmoje derėjimo laikotarpio pusėje, antro skynimo metu, birželio 6 d. (2 pav. D). Trečio skynimo metu jis sumažėjo ir išliko beveik nepakitęs iki de-rėjimo pabaigos. Išlaisčius 50 proc. vandens normos, augalų derėjimo maksimumas pasiektas trečio skynimo metu – birželio 13 dieną. Nors šių augalų derlius vėliau taip pat mažėjo, iki pat derėjimo pabaigos jis buvo didesnis negu normaliai laistytų augalų. Kai lietinimo norma buvo 25 proc., augalų derlius antro skynimo metu buvo daug mažesnis negu normaliai laistytų augalų. Vėlesnių skynimų metu ir vienų, ir kitų augalų derlius buvo panašus.
A n t o c i a n i n ų k i e k i s i r s u d ė t i s 2 0 1 3 m. Tiriant bendrą antocianinų kiekį žemuogių, laistytų pagal skirtingą režimą, uogose, 2013 m. balandžio–gegužės mėnesiais nustatyta, kad tyrimo pradžioje dėl vandens deficito uogose kaupėsi daugiau antocianinų. Išlaisčius 50 proc. vandens normos, ‘Rojan’ veislės uogose antocianinų kaupėsi 1,5 karto daugiau, o išlaisčius 25 proc. – 2,3 karto daugiau negu normaliai (pagal poreikį) laistytų augalų uogose (3 pav. A). Vėliau augalų, kurie buvo laistyti panaudojant 25 proc. vandens normos, uogose antocianinų kiekis derėjimo metu ma-žėjo, bet išliko didesnis nei normaliai laistytų augalų uogose iki pat penkto skynimo (gegužės 5 d.). Vėliau jų kiekis beveik susilygino. Antocianinų kaupimosi dinamika augalų, kurie buvo laistyti panaudojant 50 proc. vandens normos, uogose buvo panaši kaip ir normaliai laistytų augalų uogose, tačiau iki šešto skynimo pirmuoju atveju antocianinų kiekis uogose buvo šiek tiek didesnis negu antruoju.

8
3 pav. Antocianinų kiekis (A ir B) ir sudėtis (C–F) žemuogių, augintų šiltnamyje 2013 ir 2014 m., uogose, augalus laistant pagal poreikį
(100 proc. normos), išlaistant pusę normos (50 proc.) ir ketvirtį normos (25 proc.).
Fig. 3. Anthocyanin amount (A – B) and composition (C – F) at separate harvests in 2013 and 2014, when plants were irrigated according to
requirements (100 % norm), half norm (50 %), quarter of norm (25 %).

9
Derėjimo pradžioje pelargonidin-3-glikozidas ir cianidin-3-glikozidas sudarė atitinkamai 48 ir 36 proc. visų antocianinų (3 pav. C, E). Kiti antocianinai sudarė tik nedidelę dalį (16–20 proc.) bendro antocianinų kiekio. Iki ketvirto skynimo (gegužės 3 d.) šių dviejų antocianinų kiekiai buvo stabilūs. Vėliau Pel3G kiekis procentais pra-dėjo didėti, o C3G kiekis – mažėti. Derėjimo pabaigoje jų santykis jau buvo atitinkamai 65 ir 18 procentų. Vandens deficitas šių dviejų pagrindinių antocianinų santykį veikė nedaug. Derėjimo pirmoje pusėje dėl vandens deficito C3G kiekis šiek tiek mažėjo, o antroje pusėje – šiek tiek didėjo. Pel3G kiekis kito atvirkščiai proporcingai C3G kiekiui.
A n t o c i a n i n ų k i e k i s i r s u d ė t i s 2 0 1 4 m. Analizuojant antocianinų kaupimąsi uogose derėjimo metu, matyti, kad jis buvo pasiekęs maksimumą derėjimo pradžioje ir antroje derėjimo pusėje (3 pav. B). Derėjimo pradžioje, esant vandens deficitui, antocianinai kaupėsi ‘Rojan‘ veislės uogose taip pat, kaip ir kontrolinių, normaliai laistytų augalų uogose. Vėliau, antroje derėjimo laikotarpio pusėje, esant vandens deficitui, antocianinų kaupėsi daugiau negu kontrolinių augalų uogose, tačiau skirtumas buvo nedidelis. Nagrinėjant atskirų antocianinų kiekio kitimą derėjimo metu, matyti, kad 2014 m., kitaip nei 2013 m., pastebima cianidin-3-glikozido dalies didėjimo ir pelargonidin-3-glikozido dalies mažėjimo bendrame antocianinų kiekyje tendencija (3 pav. D, F). Derėjimo pradžioje šių dviejų pagrindinių antocianinų san-tykis buvo panašus kaip ir 2013 metais. Pastebėta, kad derėjimo antroje pusėje, esant vandens deficitui, C3G dalis buvo mažesnė negu normaliai laistytų augalų uogose, o Pel3G – didesnė.
Aptarimas. 2013 ir 2014 m. dauguma uogų, išskyrus paskutinių dviejų skyni-mų uogas, užsimezgė prieš lietinimo bandymo pradžią, todėl sumažintas vandens kiekis augalams laistyti uogų skaičių ir derlių paveikė minimaliai. Derėjimo metu uogų skaičius didėjo, nes pradėjo derėti žiedynų šoninės šakos. Braškės ir žemuo-gės žiedynas yra dvišakis (dichazis), kuriame didžiausias yra pirmos eilės žiedas, o tolimesnių eilių žiedai ir atitinkamai uogos yra vis mažesni. Todėl derėjimo metu, daugėjant iš žemesnių eilių žiedų susiformavusių uogų, jų dydis mažėjo, bet skaičius didėjo. Uogų nokimo dinamika 2013 ir 2014 m. šiek tiek skyrėsi. Pirmaisiais tyrimo metais derėjimo maksimumas pasiektas septintojo skynimo metu, tai yra praėjus 30 dienų nuo derėjimo ir bandymo pradžios. Antraisiais tyrimo metais šis maksimumas pasiektais per antrąjį ir trečiąjį skynimus – praėjus 6–14 dienų nuo derėjimo pra-džios. Matyt, tai lėmė oro temperatūros kitimo ypatumai skirtingais metais. 2013 m. temperatūra gerokai ir gana stabiliai pakilo antroje derėjimo pusėje, nuo ketvirto skynimo, o 2014 m. temperatūra pakilo jau antrą derėjimo savaitę, tai yra derėjimo pradžioje (1 pav.). Po to oro temperatūra vėl truputį krito ir nusistovėjo. Temperatū-ros pakilimas teigiamai paveikė derėjimą, bet, esant vidutinio dydžio vandens defi-citui, šis poveikis buvo ryškesnis. 2014 m. tai ypač buvo būdinga augalams, kurie laistyti panaudojant 50 proc. vandens normos. Derėjimo maksimumas šiuo atveju pasiekiamas vėliau negu normaliai laistytų augalų. Galima daryti prielaidą, kad dėl vandens deficito sukelto streso pristabdomas uogų nokimas ir daugiau užuomazgų žiedyno ašies viršuje (kurios yra didesnės negu žiedyno apatinėje dalyje) išlieka vė-lesnių skynimų metu, be to, vėliau susiformavo tam tikra fiziologinė ir biocheminė adaptacija stresui.
Praėjus savaitei nuo tos dienos, kai temperatūra pakilo, derėjimas sumažėjo, nes

10
derlingumo potencialas buvo išnaudotas. Didelis vandens deficitas, kai augalai buvo laistyti panaudojant 25 proc. vandens normos, augalus išsekino, dėl to derlius labai sumažėjo. Vandens deficitas dažniausiai mažina braškių lapų plotą, ūglių sausą masę, bendrą sausą masę, vandens kiekį ūgliuose, vandens potencialą, uogų dydį ir derlių (Giné Bordonaba, Terry, 2010; Klamkowski, Treder, 2008). Tačiau Liu ir bendraau-toriai (2007) nustatė, kad, nepaisant uogų masės mažėjimo, kiti derlingumo rodikliai, tokie kaip uogų skaičius, esant vandens deficitui, išlieka stabilūs. Pastebėta, kad vandens deficitas padidina vandens naudojimo efektyvumą (Liu ir kt., 2007; Boyer ir kt., 2015). Aprašyti atvejai, kai vandens deficitas netgi stimuliavo vynmedžio uogų nokimą (Castellarin ir kt., 2007). Kaip minėta, mūsų tyrimų atveju vidutinio dydžio vandens deficitas nesumažino derlingumo, bet ištęsė derėjimo laikotarpį. Literatūros duomenimis, reakcija į vandens deficito sukeltą stresą labai priklauso nuo genotipo (Ghaderi ir kt., 2015). Genotipų skirtumai susiję su anglies fiksavimo ir vandens ne-tekimo santykiu (Klamkowski, Treder, 2008).
Literatūros duomenimis, nepaisant neigiamo poveikio uogų dydžiui, lietinimo normos sumažinimas ketvirčiu nuo žydėjimo pradžios iki derėjimo padidino kai kurių veislių braškių vandens panaudojimo efektyvumą ir pagerino uogų kokybę ir skonį (Giné Bordonaba, Terry, 2010). Pastebėta, kad vandens deficito sukeltas stresas skatina įvairių junginių, tarp jų ir antocianinų, kaupimąsi braškių uogose (Terry ir kt., 2007). Mūsų atliktais tyrimais nustatyta, kad vandens deficitas padidino antocianinų kiekį žemuogių uogose atskirų skynimų metu: 2013 m. – iki dviejų kartų (balandžio 26 d.), o 2014m. – 38 proc. (birželio 20–27 d.). Bendras antocianinų kiekio padidėjimas 2013 m. sudarė 45, o 2014 m. – 18 procentų. Antocianinų kiekio variacijas tyrimo metais galima susieti su aplinkos sąlygų bei derlingumo rodiklių pokyčiais. Literatūros duomenimis, antocianinų kiekis priklauso nuo daugelio veiksnių, tarp jų – nuo genoti-po ir klimato sąlygų skirtingais metais (Najda ir kt., 2014; Ghaderi ir kt., 2015). Mūsų tyrimo duomenimis, bendras antocianinų kiekis buvo didžiausias, kai oro temperatūra nebuvo aukščiausia: 2013 m. – derėjimo pradžioje, 2014 m. – derėjimo antroje pusėje. Temperatūros pakilimas iš karto arba po tam tikro laiko sukeldavo antocianinų kiekio mažėjimą. He ir bendraautorių duomenimis (2010), dažniausiai neaukštos tempera-tūros (iki 25 °C) skatina antocianinų biosintezę, o aukštos temperatūros (apie 35 °C) yra susijusios su antocianinų degradacija. Mūsų tyrimo duomenimis, apie 15 °C oro temperatūra lauke teigiamai veikė antocianinų kaupimasi, o apie 25 °C – jų kiekį uo-gose mažino. Šviečiant saulei, šiltnamyje oro temperatūra paprastai buvo apie 10 °C aukštesnė nei lauke. Mūsų tyrimų duomenys rodo, kad aukštesnė temperatūra anto-cianinų kaupimąsi augalų, augintų vandens deficito sukelto streso sąlygomis, uogose taip pat veikė neigiamai.
Pastebėta, kad antocianinų kiekis ‘Rojan’ veislės uogose, augalams laistyti panau-dojant 25 proc. vandens normos, neigiamai koreliuoja su uogų derliumi (koreliacijos koeficientas r = –0,74). Mažėjant uogų masei, šiek tiek didėjo antocianinų kiekis.
Kaip minėta, derėjimo pradžioje Pel3G ir C3G sudarė atitinkamai 48 ir 36 proc. visų antocianinų. Kitų autorių duomenimis, C3G sudarė 49, 2 proc., Pel3G – 40,7 proc. visų antocianinų kiekio žemuogėse (Kawanobu ir kt., 2011). Remiantis daugelio autorių duomenimis, braškėse dominuoja Pel3G ir sudaro nuo 50 iki daugiau kaip 90 proc. visų antocianinų, kiti antocianinai – Pel3rut, C3G ir Pel3MG – retai viršija 30 proc.

11
ribą (Lopesda Silva ir kt., 2007; Kawanobuir kt., 2011). Mažesnis Pg ir Cy santykis yra vertinamas teigiamai, nes cyanidin-3-glikozido antioksidacinis aktyvumas dides-nis negu pelargonidin-3-glikozido (Satué-Gracia ir kt., 1997). Todėl žemuogės šiuo požiūriu galbūt yra šiek tiek vertingesnės negu braškės.
Atskirų antocianinų santykio kitimą derėjimo metu galima susieti su oro tempe-ratūros pokyčiais. Tyrimo metais Pel3G ir C3G santykis gerokai padidėjo pakilus oro temperatūrai arba praėjus keletui dienų po temperatūros pakilimo. 2013 m. tai įvyko derėjimo antroje pusėje, o 2014 m. – beveik sutapo su dviem temperatūros pakilimais derėjimo pradžioje ir pabaigoje. Atsižvelgiant į derėjimo laiką ir derėjimo tarpsnį (de-rėjimo pabaiga ar pradžia), augalo fiziologinė būklė buvo nevienoda. Tai galbūt turėjo įtakos uogų biocheminiams rodikliams. Mūsų tyrimo duomenimis, antocianinų sudėtį uogose veikė ir vandens deficitas, tačiau šis poveikis priklausė nuo derėjimo tarpsnio. Vandens deficito sąlygomis augintų augalų uogose antocianinų santykio pokyčiai šiek tiek atsiliko nuo šių pokyčių normaliai laistytų augalų uogose. Pastebėta, kad, esant žemesnei kaip 20 °C oro temperatūrai, vandens deficito sąlygomis Pel3G ir C3G san-tykis uogose buvo 25–43 proc. didesnis negu normaliai laistytų augalų uogose, o esant aukštesnei oro temperatūrai – beveik toks pat, kaip normaliai laistytų augalų uogose.
Antocianinų kaupimosi duomenys, surinkti derėjimo metu žemuoges auginant šiltnamyje, suteikia naujų žinių, galinčių padėti patobulinti žemuogių auginimo siste-mas, kuriomis siekiama sumažinti sąnaudas ir pagerinti uogų biocheminę sudėtį. Mūsų įsitikinimu, antocianinų sintezę lemiančių genų raiškos tyrimai, atliekami su mūsų tyrimų metu surinktomis žemuogėmis, tolesni šių modelinių erškėtinių šeimos augalų streso fiziologijos tyrimai padės atskleisti natūralių pigmentų kaupimosi dėsningumus, sukurti genetinius arba biocheminius žymeklius, o ateityje – efektyviai antocianinus kaupiančius augalus ir jų auginimo sistemą.
Išvados. 1. Vandens deficito sukelto streso įtaka uogų derlingumo rodikliams – uogų dydžiui ir derliui – priklauso nuo vandens deficito laipsnio ir klimato sąlygų. Laistymas panaudojant 50 proc. vandens normos 2013 m. derėjimo rodiklių nepa-veikė, o 2014 m. – suvėlino uogų nokimą. Laistymas panaudojant 25 proc. normos 2013 m. sumažino uogų masę ir derlių derėjimo pabaigoje, 2014 m. – padidino uogų dydį, bet gerokai sumažino derlių.
2. Mūsų atliktais tyrimais nustatyta, kad vandens deficitas padidino antocianinų kiekį žemuogių uogose atskirų skynimų metu: 2013 m. – iki dviejų kartų, 2014 m. – iki 38 procentų. Bendras antocianinų kiekio padidėjimas 2013 m. sudarė 45, o 2014 m. – 18 procentų. Šis padidėjimas buvo maksimalus, kai oro temperatūra buvo ne aukštesnė kaip 20 °C.
3. Pelargonidin-3-glikozido ir cianidin-3-glikozido kiekiai derėjimo pradžioje sudarė atitinkamai 48 ir 36 proc. visų antocianinų. Šių antocianinų santykis derėjimo metu kito priklausomai nuo oro temperatūros ir nuo augalo fiziologinės būklės.
Padėka. Mokslinis tyrimas finansuotas Lietuvos mokslo tarybos lėšomis, pro-jektas Nr. SVE-06/2012.
Gauta 2015 12 08 Parengta spausdinti 2015 12 17

12
Literatūra
1. Albert N. W., Lewis D. H., Zhang H., Irving L. J., Jameson P. E., Davies K. M. 2009. Light-induced vegetative anthocyanin pigmentation in Petunia. Journal of Experimental Botany, 60: 2 191–2 202.
2. Anisimovienė N., Jankauskienė J., Jodinskienė M., Bendokas V., Stanys V., Šikšnianas T. 2013. Phenolics, antioxidative activity and characterization of anthocyanins in berries of black currant interspecific hybrids. Acta Biochimica Polonica, 60(4): 767–772.
3. Anisimovienė N., Rubinskienė M., Viškelis P., Stackevičienė E., Stanys V., Šikšnianas T., Jankovska E., Sasnauskas A. 2009. Anthocyanins in currants, cherries, blueberries and antioxidative activity of berry extracts. Žemdirbystė / Agriculture, 96(3): 158–167.
4. Arnnok P., Ruangviriyachai C., Malachai R., Techawongstien S., Chanthai S. 2012. Determination of total phenolics and anthocyanin contents in the peri-carp of hot chilli pepper (Capsinum annuum L.). International Food Research Journal, 19: 235–243.
5. Boyer L. R., Brain P., Xu X. M., Jeffries P. 2015. Inoculation of drought-stressed strawberry with a mix edinoculum of two arbus cular mycorrhizal fungi: effects on population dynamics of fungal species in roots and consequential plant tole-rance to water deficiency. Mycorrhiza, 25(3): 215–227.
6. Castellarin S. D., Matthews M. A., Di Gaspero G., Gambetta G. A. 2007.Water deficits accelerate ripening and induce changes in gene expression regulating flavonoid biosynthesis in grape berries. Planta, 227(1): 101–112.
7. Durst R. W., Wrolstad R. 2001. Separation and characterization of anthocyanins by HPLC. Current protocols in food analytical chemistry, F1.3.1–F1.3.13.
8. Ghaderi N., Normohammadi, S., Javadi T. 2015. Morpho-physiological respon-ses of strawberry (Fragaria × ananassa) to exogenous salicylic acid applica-tion under drought stress. Journal of agricultural science and technology, 17: 167–178.
9. Giné Bordonaba J., Terry L. A. 2009. Differential effect of water deficit irriga-tion on fruit quality of five june-bearing strawberry varieties. Acta Horticulturae, 838: 187–191.
10. Giné Bordonaba J., Terry. L. A. 2010. Manipulating the taste-related composi-tion of strawberry fruits (Fragaria × ananassa) from different cultivars using deficit irrigation. Food Chemistry, 122(4): 1 020–1 026.
11. He F., Mu L., Yan G. L., Liang N, N., Pan Q. H., Wang J., Reeves M. J., Duan C. Q. 2010. Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules,15(12): 9 057 –9 091.
12. Kawanobu S., Yamaguchi M., Zushi K., Kondo K., Matsuzoe N. 2011. Identification and distribution of anthocyanins in strawberry varieties. Journal of Food, Agriculture and Environment, 9(1): 140–141.
13. Kim S. K., Na H. Y., Song J. H., Kim M. J., Son J. E., Bae R. N., Chun C., Kang H. J. 2009. Influence of water stress on fruit quality and yield of straw-berry cvs. ‘Maehyangand Seolhyang’. Acta Horticulturae, 842(1): 177–180.

13
14. Klamkowski K., Treder W. 2008. Response to drought stress of three strawberry varieties grown under greenhouse conditions. Journal of Fruit and Ornamental Plant Research, 16: 179–188.
15. Liu F., Savić S., Jensen C. R., Shahnazari A., Jacobsen S. E., Stikić R., Andersen M. N. 2007. Water relations and yield of lysimeter-grown strawberries under limited irrigation. Scientia Horticulturae, 111(2): 128–132.
16. Lopesda Silva F. L., Escribano-Bailon M. T., Perez-Alonso J. J., Rivas-Gonzalo J. C., Santos-Buelga C. 2007. Anthocyanin pigments in strawberry. LWT-Food Science and Technology, 40(2): 374–382.
17. Najda A., Dyduch-Siemińska M., Dyduch J., Gantner, M. 2014. Comparative analysis of secondary metabolites contents in Fragaria vesca L. fruits. Annals of Agricultural and Environmental Medicine, 21(2): 339–343.
18. Nora L., Dalmazo G. O., Nora F. R., Rombaldi C. V. 2012. Controlled water stress to improve fruit and vegetable postharvest quality. In: I. Md. Rahman, H. Hasegawa (ed.), Agricultural and Biological Sciences „Water Stress“. www.intechopen.com
19. Rugienius R., Šikšnianas T., Sasnauskas A. 2013. Žemuogių veislių, hibridų ir atrinktų linijų tyrimai. Sodininkystė ir daržininkystė, 32(1–2): 3–14.
20. Satué-Gracia M. T., Heinonen I. M., Frankel E. N. 1997. Anthocyanins as an-tioxidants on human low-density lipoprotein and lecithin-liposome systems. Journal of Agricultural and Food Chemistry, 45: 3 362 –3 367.
21. Terry L. A., Chope G. A., Bordonaba J. G. 2007. Effect of water deficit irrigation and inoculation with Botrytis cinerea on strawberry (Fragaria × ananassa) fruit quality. Journal of Agricultural and Food Chemistry, 55(26): 10 812–10 819.
22. Zafra-Stone S., Bagchi M., Bagchi D. 2007. Health benefits of edible berry anthocyanins: Novel antioxidant and anti-angiogenic properties. In: F. Shahidi, C. T. Ho (eds)., Antioxid ant Measurement and Application. American Chemical Society, Washington, 337–351.
SODININKYSTĖ IR DARŽININKYSTĖ. SCIENTIFIC ARTICLES. 2015. 34(3–4).
Cropping and anthocyanin content in berries of wild strawberry under water deficit stress
R. Rugienius, T. Šikšnianas, V. Bendokas, V. Stanys
Summary
The rise of average air temperature increases the demand of irrigation of small fruit plants in greenhouses and open fields. Despite berry weight and yield decrease under water deficit it could enrich berry biochemical composition due to the accumulation of stress induced phenolic compounds. The aim of our investigations was to evaluate cropping, anthocyanin amount and composition of wild strawberry fruits from plants grown under water deficit stress in a green-house. Every 4–7 days the average berry weight and yield, anthocyanin amount and content was evaluated using HPLC in 2013 and 2014. It was revealed in the study that the impact of

14
water deficit on berry weight and yield depends on the level of water deficit and air temperature. Cropping parameters were not affected by the decrease of an irrigation norm by 50 % in 2013. The cropping was delayed when 50 % norm water irrigation had been applied in 2014. The ir-rigation by the 25 % norm caused a decrease of the yield in both 2013 and 2014, but differently from the first year it increased the average berry weight in 2014. It was established in the study that the water deficit increased the anthocyanin amount in the berries at separate harvesting times to two times in 2013 and up to 38 % in 2014. The average increase of the anthocyanin amount was 45 % in 2013 and 18 % in 2014. This increase was maximal when air temperature did not exceed 20 °C. The amount of pelargonidin-3-glucoside and cyanidin-3-glucoside at in the beginning of cropping reached 48 and 36 % of all anthocyanins respectively. Pel3G/C3G ratio changed during the cropping season depending on air temperature and physiological state of the plants.
Key words: air temperature, drought, Fragaria vesca, greenhouse, irrigation, pigments, yield.

15
LIETUVOS AGRARINIŲ IR MIŠKŲ MOKSLŲ CENTRO FILIALO SODININKYSTĖS IR DARŽININKYSTĖS INSTITUTO IR ALEKSANDRO STULGINSKIO UNIVERSITETO MOKSLO DARBAI. SODININKYSTĖ IR DARŽININKYSTĖ. 2015. 34(3–4).
Dviejų vystymosi tarpsnių žirnių fotosintezės rodiklių atsakas į substrato rūgštumo ir UV-B spinduliuotės poveikį
Irena JanuškaitienėVytauto Didžiojo universitetas, Gamtos mokslų fakultetas, Vileikos g. 8-223, LT-44404 Kaunas, el. paštas [email protected]
Atliekant šį tyrimą buvo tirtas 1 ir 3 kJ m-2 d-1 UV-B spinduliuotės ir rūgštaus substrato (pH 4,8) poveikis skirtingų vystymosi tarpsnių sėjamojo žirnio (Pisum sativum L.) fotosin-tezės rodikliams. Žirniai buvo sėjami į vegetacinius indus su paruoštu neutralaus rūgštumo durpių substratu. Augalams pasiekus lapų vystymosi (BBCH 14–16) ir šoninių ūglių forma-vimosi tarpsnius (BBCH 21–23), stebėtas UV-B spinduliuotės ir substrato rūgštumo poveikis. Fotosintezės rodikliai matuoti fotosintezės intensyvumo matavimo įrenginiu LI-6400, o fotosintetiniai pigmentai nustatyti acetono ekstrakte spektrofotometriškai. UV-B spinduliuo-tės poveikis BBCH 14–16 tarpsnio sėjamųjų žirnių tirtiems rodikliams buvo stipresnis nei BBCH 21–23 tarpsnio: pirmuoju atveju vidutinis visų tirtų rodiklių sumažėjimas sudarė 34,2 proc., antruoju – tik 21,8 proc., palyginti su kontrolės augalais. Vystymosi tarpsnio, kaip veiksnio, įtaka buvo statistiškai reikšminga (p < 0,05). Rūgštus substratas BBCH 21–23 tarpsnio sėjamuosius žirnius pažeidė labiau (38 proc.) nei kontrolės augalus, tačiau vystymosi tarpsnio įtaka nebuvo statistiškai reikšminga (p > 0,05). Sėjamuosius žirnius veikiant komplek-siškai – UV-B spinduliuote ir rūgštinant substratą, BBCH 14–16 tarpsnio augalų bendras tirtų rodiklių sumažėjimas sudarė 46,9 proc., o BBCH 21–23 tarpsnio – 50,9 proc., tačiau vystymosi tarpsnio, kaip veiksnio, įtaka nebuvo statistiškai reikšminga (p > 0,05). Kompleksinis UV-B spinduliuotės ir rūgštaus substrato poveikis lėmė didesnius sėjamųjų žirnių pažeidimus negu pavienis, BBCH 14–16 tarpsnio žirnių tirti rodikliai sumažėjo labiau (p > 0,05).
Reikšminiai žodžiai: chlorofilai, fotosintezės rodikliai, sausa biomasė, substrato rūgš-tumas, UV-B, vystymosi tarpsnis.
Įvadas. Augalų atsakas į nepalankias aplinkos sąlygas labai priklauso ir nuo au-galo amžiaus (Larcher, 2003). Jaunų augalų lapuose, kurie augalams yra svarbiausi, streso įveikimo mechanizmai įsijungia pirmiausiai (Reifenrath, Muller, 2007). Tad šiems procesams skiriama labai daug energijos, kuri galėtų būti panaudota augalui augti ir vystytis. Be abejo, labiau subrendę augalai yra sukaupę didesnę masę, tuo pačiu ir didesnius energijos išteklius (Duchovskis, 1998; Januškaitienė, Dikšaitytė, 2014).

16
Aplinkos tarša rūgščiu lietumi tapo svarbiu veiksniu, darančiu įtaką augalų au-gimui, vystymuisi bei produktyvumui. Rūgštėjant dirvožemiams (pH < 5,5) padidėja nuodingų sunkiųjų metalų (ypač Al) aktyvumas ir tirpumas. Šie pokyčiai tiesiogiai veikia ląstelių sieneles, Ca homeostazę ir plazmines membranas (Ma, 2007). Šaknų sistemos pakenkimai sąlygoja vandens ir mitybinių medžiagų patekimo į augalus su-mažėjimą, o tai lemia ir viso augalo ląstelių biocheminius ir fiziologinius pakitimus (Haling ir kt., 2010). Rūgštūs dirvožemiai daro ne tik pavienius, bet ir kompleksinius pakenkimus (Shan ir kt., 2000).
Didesnė negu įprasta spinduliuotė sukelia įvairius augalo ląstelių pažeidimus (Hollosy, 2002). Pakitimai ir pažeidimai molekulių lygmeniu neabejotinai keičia augimo ir vystymosi procesus: genų aktyvumą, metabolizmą, fotosintezės aktyvumą (Wei ir kt., 2003; Jukunen-Tiitto ir kt., 2005). UV-B spinduliuotė gali sutrikdyti CO2 patekimą į augalą per žioteles bei CO2 fiksavimo reakcijas Kalvino cikle (Xu, Qiu, 2007). Naujausių tyrimų duomenys rodo, kad neigiamas UV-B spinduliuotės povei-kis fotosintezei labiausiai pasireiškia dėl antrosios fotosistemos (FS II) aktyvumo slopinimo (Kataria ir kt., 2014; Zhu, Yang, 2015). Tačiau gauta ir priešingų rezultatų: teigiama, kad UV-B spinduliuotė neveikia fotosintezės intensyvumo. Manoma, kad UV-B absorbuojantys pigmentai, tokie kaip flavonoidai, karotinoidai, gali patikimai apsaugoti fotosintezės sistemą nuo žalingo UV-B spinduliuotės poveikio (Agati, Tattini, 2010; Januskaitiene, 2011; Sakalauskiene ir kt., 2013).
Kompleksinio UV-B spinduliuotės ir rūgštaus substrato poveikio augalų augi-mui ir fotosintezei tyrimai yra labai negausūs. E. Paoletti (1998) nustatė, kad UV-B ir rūgštaus lietaus poveikis paprastajam bukui ir akmeniniam ąžuolui sukėlė aukščiausio lygio lapų audinių nekrozes. Šiems veiksniams veikiant atskirai, rūgštus lietus sukėlė didesnius augalų pažeidimus nei UV-B spinduliuotė (Paoletti, 1998; Agati ir kt., 2012). Panašus dėsningumas nustatytas paveikus UV-B spinduliuote valgomuosius ridikė-lius ir auginant juos rūgščiame substrate (Januskaitiene, 2011). Tačiau atliekant tyrimą su vasariniais rapsais gauti priešingi rezultatai – kompleksinis UV-B spinduliuotės ir rūgštaus substrato poveikis buvo stipresnis nei pavienis (Januškaitienė, 2013).
Darbo tikslas – įvertinti kompleksinį substrato rūgštumo ir UV-B spinduliuotės poveikį skirtingų vystymosi tarpsnių sėjamojo žirnio (Pisum sativum L.) fotosintezės rodikliams.
Tyrimo objektas, metodai ir sąlygos. Atliekant tyrimus, ‘Ilgių’ veislės sėja-mieji žirniai buvo sėjami po 20–25 sėklas į 5 l talpos vegetacinius indus su paruoštu neutralaus (pH 6,0–6,5) rūgštumo durpių substratu. Pasėti augalai iki sudygimo buvo auginami specialioje patalpoje (fotoperiodo trukmė – 14 val.). Praėjus aštuonioms dienoms po sudygimo – augalams pasiekus lapų vystymosi tarpsnį (BBCH 14–16), pusė augalų pradėti veikti UV-B spinduliuote ir padidintas substrato rūgštumas (Growth stages..., 2001). Kita dalis augalų pradėti veikti UV-B spinduliais ir padi-dintas substrato rūgštumas po savaitės, jiems pasiekus šoninių ūglių formavimosi tarpsnį (BBCH 21–23) (Growth stages..., 2001). Abiejų tarpsnių augalai buvo veikia-mi penkias dienas: kasdien žirniai buvo veikiami 1 ir 3 kJ m-2 d-1 UV-B spinduliuote, substrato rūgštumui padidinti iki pH 4,8 jis buvo laistomas 0,07 M koncentracijos H2SO4 tirpalu. Kontrolės augalai poveikio dienomis buvo laistomi distiliuotu vande-niu, kad augtų vienodomis sąlygomis su veikiamais rūgštimi augalais.

17
Paskutinę bandymo dieną, po UV-B poveikio praėjus 2 val., fotosintezės intensy-vumas (CO2 asimiliavimo greitis lape, µmol CO2 m
-2 s-1), transpiracijos intensyvumas (mmol H2O
-1 s-1), vandens naudojimo efektyvumas (µmol CO2 m-2 s-1/mmol H2O m-2
s-1) ir viduląstelinis CO2 kiekis (µmol CO2 mol air-1) matuoti fotosintezės intensyvumo matavimo įrenginiu LI-6400 (LI-COR, JAV). Kas 3 sekundes po 5 minutes registruoti atsitiktinai pasirinktų trijų kiekvieno tyrimų varianto augalų antrosios poros lapų fotosintezės parametrai. Iš šių duomenų skaičiuotas momentinis fotosintezės rodiklių vidurkis.
Tyrimų metu vidutinis lapo apšviestumas kvantais svyravo nuo 90 iki 150 μmol m-2 s-1, vidutinis santykinis drėgnis mėginio kameroje buvo 25–30 proc., anglies dvideginio koncentracija bandinio kameroje – 300–400 μmol CO2 mol-1, bloko ir lapo temperatūra – apie 25 °C, oro srauto greitis – 400 μmol s-1.
Chlorofilo a ir b kiekiai nustatyti paskutinę bandymo dieną pagal Wettstein metodą 100 % acetono ekstrakte su spektrofotometru DU800 VU/VIS (USA), kartojant tris kartus (Wettstein, 1957; Гавриленко ir kt., 1975). Sausai biomasei nustatyti augalų daigai buvo džiovinami +60 °C temperatūroje vieną parą. Vieno augalo sausos bio-masės vidurkis buvo skaičiuojamas iš penkių augalų.
Tirtų veiksnių poveikio reikšmingumas įvertintas dispersinės analizės metodu ANOVA, skaičiuojant F kriterijų ir jo p reikšmę, o lyginant pavienius atvejus skai-čiuota Stjudento t kriterijaus p reikšmė. Visuose paveiksluose pateikiamas vidurkis ± pasikliautinasis intervalas (P = 0,05).
Rezultatai. 1 paveiksle pateikti skirtingų vystymosi tarpsnių sėjamųjų žirnių fotosintezės intensyvumo pokyčiai esant skirtingam substrato rūgštumui (pH 6,5; pH 4,8) ir 1 bei 3 kJ m-2 d-1 UV-B spinduliuotės poveikiui.
Neutraliame (pH 6,5) substrate lapų vystymosi tarpsnio (BBCH 14–16) žirnius paveikus UV-B spinduliuote fotosintezės intensyvumas mažėjo. Didžiausi (net 4,7 karto (p < 0,05) nuostoliai gauti veikiant 3 kJ m-2 d-1 UV-B spinduliuote. BBCH 14–16 vystymosi tarpsnio žirnių fotosintezės intensyvumo nuostoliai rūgščiame substrate buvo kiek mažesni – 93 proc. (p < 0,05). Paveiktų šoninių ūglių formavimosi tarps-nio (BBCH 21–23) žirnių fotosintezės intensyvumas, didėjant UV-B spinduliuotei, dėsningai mažėjo abiejuose substratuose. Šio tarpsnio kontrolės augalų fotosintezė svyravo apie 2,9 µmol CO2 m
-2 s-1, o 1 ir 3 kJ m-2 d-1 UV-B spinduliuotės paveiktų – atitinkamai net 58,5 (p < 0,05) ir 73,5 proc. (p < 0,05) mažiau. Veikiant 1 kJ m-2 d-1 UV-B spinduliuote parūgštintą iki pH 4,8 substratą, žirnių fotosintezės intensyvumas sumažėjo šiek tiek mažiau nei veikiant neutralių substratą. Paskutinę bandymo dieną nustatytas 52,2 proc. (p < 0,05) fotosintezės intensyvumo sumažėjimas, palyginti su kontrole. Didesnė UV-B spinduliuotė parūgštintame substrate fotosintezės intensyvumą sumažino labai stipriai – net 2,3 karto (p < 0,05), palyginti su kontrole.
Lapų vystymosi (BBCH 14–16) tarpsnio žirnių transpiracijos intensyvumas dėl UV-B spinduliuotės poveikio mažėjo abiejuose substratuose (2 pav.). Didesni nuostoliai nustatyti substrate, kurio pH 4,8: dėl 1 ir 3 kJ m-2 d-1 UV-B spinduliuotės poveikio transpiracijos intensyvumas mažėjo atitinkamai 42 (p < 0,05) ir 64 proc. (p < 0,05). Dėl minėtos spinduliuotės poveikio transpiracijos intensyvumas pH 6,5 substrate mažėjo atitinkamai 12 (p < 0,05) ir 32 proc. (p < 0,05). Šoninių ūglių formavimosi tarpsnio (BBCH 21–23) žirnius tiek paveikus vien tik UV-B spinduliuote, tiek veikiant kompleksiškai – spinduliuote ir rūgštinant substratą, statistiškai reikšmingų skirtumų nenustatyta (p > 0,05) (2 pav.).

18
1 pav. Skirtingų vystymosi tarpsnių žirnių fotosintezės intensyvumo pokyčiai esant skirtingam substrato rūgštumui (pH 6,5 – kontrolė; pH 4,8) ir
1 bei 3 kJ m-2 d-1 UV-B spinduliuotės poveikiui. BBCH 14–16 – lapų vystymosi tarpsnis;
BBCH 21–23 – šoninių ūglių formavimosi tarpsnis.Fig. 1. Photosynthetic rate of pea at different growth stages under impact of
different substrate acidity (pH 6.5 – control; pH 4.8) and 1 and 3 kJ m-2 d-1 UV-B dose. BBCH 14–16 – leaf development stage;
BBCH 21–23 –formation of lateral shoots stage.
2 pav. Skirtingų vystymosi tarpsnių žirnių transpiracijos intensyvumo pokyčiai esant skirtingam substrato rūgštumui (pH 6,5 – kontrolė; pH 4,8) ir
1 bei 3 kJ m-2 d-1 UV-B spinduliuotės poveikiui. BBCH 14–16 – lapų vystymosi tarpsnis;
BBCH 21–23 – šoninių ūglių formavimosi tarpsnisFig. 2. Transpiration rate of pea at different growth stages under impact of
different substrate acidity (pH 6.5 – control; pH 4.8) and 1 and 3 kJ m-2 d-1 UV-B dose. BBCH 14–16 – leaf development stage;
BBCH 21–23 – formation of lateral shoots stage.

19
Skirtingų vystymosi tarpsnių žirnių viduląstelinio CO2 kiekio pokyčiai pateikti 3 paveiksle. Lapų vystymosi tarpsnio žirnius paveikus 1 ir 3 kJ m-2 d-1 UV-B spin-duliuote, statistiškai reikšmingas buvo tik 3 kJ poveikis: gautas 29 proc. (p < 0,05) padidėjimas, palyginti su kontrole. Substrato parūgštinimas viduląstelinį CO2 kiekį taip pat didino 23 proc. (p < 0,05), o kompleksinis substrato rūgštumo ir UV-B spin-duliuotės poveikis – 29 proc. (p < 0,05), palyginti su kontrolės augalais. Šoninių ūglių formavimosi tarpsnio žirnių viduląstelinio CO2 kiekio pokyčiai dėl minėtų veiksnių poveikio buvo gerokai silpnesni ir esminiai skirtumai gauti tik esant kompleksiniam substrato rūgštumo ir 3 kJ UV-B spinduliuotės poveikiui: viduląstelinis CO2 kiekis padidėjo 30 proc. (p < 0,05), palyginti su kontrole.
3 pav. Skirtingų vystymosi tarpsnių žirnių viduląstelinio CO2 kiekio pokyčiai esant skirtingam substrato rūgštumui
(pH 6,5 – kontrolė; pH 4,8) ir 1 bei 3 kJ m-2 d-1 UV-B spinduliuotės poveikiui. BBCH 14–16 – lapų vystymosi tarpsnis;
BBCH 21–23 – šoninių ūglių formavimosi tarpsnis.Fig. 3. Intercellular CO2 concentration of pea at
different growth stages under impact of different substrate acidity (pH 6.5 – control; pH 4.8) and 1 and 3 kJ m-2 d-1 UV-B dose.
BBCH 14–16 – leaf development stage; BBCH 21–23 – formation of lateral shoots stage.

20
4 paveiksle pateikta skirtingų vystymosi tarpsnių sėjamųjų žirnių vandens nau-dojimo efektyvumo pokyčiai esant skirtingam substrato rūgštumui (pH 6,5; pH 4,8) ir 1 bei 3 kJ m-2 d-1 UV-B spinduliuotės poveikiui. Šio rodiklio pokyčių dėsningu-mai gauti tokie pat, kaip ir fotosintezės intensyvumo: didžiausi nuostoliai gauti dėl 3 kJ UV-B spinduliuotės poveikio (2 pav.).
4 pav. Skirtingų vystymosi tarpsnių žirnių vandens naudojimo efektyvumo pokyčiai esant skirtingam substrato rūgštumui
(pH 6,5 – kontrolė; pH 4,8) ir 1 bei 3 kJ m-2 d-1 UV-B spinduliuotės poveikiui. BBCH 14–16 – lapų vystymosi tarpsnis;
BBCH 21–23 – šoninių ūglių formavimosi tarpsnis.Fig. 4. Water use effieciency of pea at different growth stages
under impact of different substrate acidity (pH 6.5 – control; pH 4.8) and 1 and 3 kJ m-2 d-1 UV-B dose.
BBCH 14–16 – leaf development stage; BBCH 21–23 – formation of lateral shoots stage.
Chlorifilų a ir b kiekį lapų vystymosi tarpsnio sėjamųjų žirnių lapuose labiausiai mažino rūgštus (pH 4,8) substratas ir 3 kJ UV-B spinduliuotė (5 pav.). Chlorifilų a ir b kiekį šoninių ūglių formavimosi tarpsnio žirnių lapuose labiausiai (atitinkamai 56 ir 57 proc.) mažino rūgštus (pH 4,8) substratas (p < 0,05 p). Silpna 1 kJ m-2 d-1 UV-B spinduliuotė tiek neutraliame, tiek rūgščiame substrate chlorofilų a ir b kiekius šio vystymosi tarpsnio žirnių lapuose padidino, palyginti su UV-B spinduliuote neveiktais augalais, bet statistiškai reikšmingai – tik rūgščiame substrate (pH 4,8).

21
Chlorofilų a ir b santykio skirtingų vystymosi tarpsnių žirnių lapuose pokyčiai pateikti 6 paveikle. Tiek dėl UV-B spinduliuotės, tiek dėl rūgštaus substrato poveikio chlorofilų a ir b santykis mažėjo, išskyrus pavienį substrato rūgštumo (pH 4,8) poveikį šoninių ūglių formavimosi tarpsnio žirniams: chlorofilų a ir b santykis padidėjo, bet statistiškai nereikšmingai (p > 0,05). Chlorofilų a ir b santykis neutraliame substrate (pH 6,5) augusių lapų vystymosi tarpsnio žirnių lapuose dėl UV-B spinduliuotės povei-kio sumažėjo daugiau nei šoninių ūglių formavimosi tarpsnio žirnių lapuose: veikiant 3 kJ UV-B spinduliuote, chlorofilų a ir b santykis buvo atitinkamai 28 ir 22 proc. (p < 0,05) mažesnis nei kontrolės augalų lapuose.
5 pav. Chlorofilo a ir b kiekio skirtingų vystymosi tarpsnių žirnių lapuose pokyčiai esant skirtingam substrato rūgštumui (pH 6,5 – kontrolė; pH 4,8) ir
1 bei 3 kJ m-2 d-1 UV-B spinduliuotės poveikiui. BBCH 14–16 – lapų vystymosi tarpsnis;
BBCH 21–23 – šoninių ūglių formavimosi tarpsnis; ŽM – žalia masė.Fig. 5. Chlorophyll a and b content of pea at different growth stages under impact of different substrate acidity (pH 6.5 – control; pH 4.8) and 1 and
3 kJ m-2 d-1 UV-B dose. BBCH 14–16 – leaf development stage; BBCH 21–23 – formation of lateral shoots stage; FW – fresh weight.

22
6 pav. Chlorofilų a ir b santykio skirtingų vystymosi tarpsnių žirnių lapuose pokyčiai esant skirtingam substrato rūgštumui (pH 6,5 – kontrolė; pH 4,8) ir
1 bei 3 kJ m-2 d-1 UV-B spinduliuotės poveikiui. BBCH 14–16 – lapų vystymosi tarpsnis;
BBCH 21–23 – šoninių ūglių formavimosi tarpsnis; ŽM – žalia masė.Fig. 6. Chlorophyll a and b ratio of pea at leaf development (BBCH 14–16) and
formation of lateral shoots (BBCH 21–23stages) under impact of different substrate acidity (pH 6.5 – control; pH 4.8) and 1 and 3 kJ m-2 d-1 UV-B dose. BBCH 14–16 – leaf development stage;
BBCH 21–23 – formation of lateral shoots stage; FW – fresh weight.
Lapų vystymosi ir šoninių ūglių formavimosi tarpsnių sėjamųjų žirnių, paveik-tų 1 ir 3 kJ m-2 d-1 UV-B spinduliuote ir augusių skirtingo rūgštumo substratuose (pH 6,5; pH 4,8), vieno augalo antžeminės dalies sausos biomasės kiekiai tyrimo pabaigoje pateikti 7 paveiksle. Dėl UV-B spinduliuotės poveikio abiejų vystymosi tarpsnių žirnių biomasė tiek neutraliame, tiek ir rūgščiame substratuose dėsningai mažėjo ir nuo kontrolės augalų (pH 6,5) skyrėsi statistiškai reikšmingai (p < 0,05). Tiek dėl pavienio, tiek dėl kompleksinio tirtų veiksnių poveikio lapų vystymosi tarps-nio žirnių biomasės nuostoliai buvo didesni nei tų pačių veiksnių veiktų šoninių ūglių formavimosi tarpsnio žirnių. Pavyzdžiui, esant 3 kJ UV-B spinduliuotės poveikiui lapų vystymosi tarpsnio žirnių biomasės nuostoliai buvo 35 proc. (p < 0,05), o šoninių ūglių formavimosi tarpsnio – 31 proc. (p < 0,05), palyginti su kontrolės augalais. Veikiant kompleksiškai, neigiamas poveikis dar sustiprėjo: veikiant 3 kJ UV-B spinduliuote ir substrato rūgštumui esant pH 4,8, lapų vystymosi tarpsnio ir šoninių ūglių formavimosi tarpsnių žirnių sausos biomasės nuostoliai buvo atitinkamai 44 ir 42 proc. (p < 0,05), palyginti su kontrole.

23
7 pav. Skirtingų vystymosi tarpsnių žirnių antžeminės dalies vidutinės augalo sausos biomasės pokyčiai,
esant skirtingam substrato rūgštumui (pH 6,5 – kontrolė; pH 4,8) ir 1 bei 3 kJ m-2 d-1 UV-B spinduliuotės poveikiui.
BBCH 14–16 – lapų vystymosi tarpsnis; BBCH 21–23 – šoninių ūglių formavimosi tarpsnis.
Fig. 7. One plant dry weight of over ground biomass of pea at different growth stages under impact of different substrate acidity
(pH 6.5 – control; pH 4.8) and 1 and 3 kJ m-2 d-1 UV-B dose. BBCH 14–16 – leaf development stage;
BBCH 21–23 – formation of lateral shoots stage.
Skirtingų vystymosi tarpsnių sėjamųjų žirnių visų tirtų rodiklių vidutinis su-mažėjimas (%), esant pavieniam bei kompleksiniam UV-B spinduliuotės ir rūgštaus substrato poveikiui, pateikiamas lentelėje. Iš lentelėje pateiktų duomenų matyti, jog pavienis UV-B spinduliuotės poveikis didesnę neigiamą įtaką darė tirtiems lapų vys-tymosi tarpsnio augalų rodikliams: vidutinis sumažėjimas sudarė 34,2 proc., palyginti su kontrole, o šoninių ūglių formavimosi tarpsnio augalų – 21,8 procento.

24
Lentelė. Visų tirtų rodiklių vidutinis sumažėjimas (%), palyginti su kontrole, esant skirtingam vystymosi tarpsniui ir skirtingam pasirinktų stresorių poveikiuiTable. The average decrease (%) of all investigated indicators, compared to the control, at different growth stages and different effects of investigated stressors
PoveikisEffect
Sumažėjimas, palyginti su kontrole Decrease compared with control, %
lapų vystymosi tarpsnis (BBCH 14–16)
Leaf development stage (BBCH 14–16)
šoninių ūglių formavimosi tarpsnis (BBCH 21–23)
Lateral shoots formation stage (BBCH 21–23)
Pavienis UV-B poveikis Single UV-B effect
34,2 21,8
Pavienis substrato pH poveikis Single substrate pH effect
36,4 38,0
UV-B ir substrato pH poveikis UV-B and substrate pH effect
46,9 50,9
Visų poveikių vidurkis Average of all effects
39,2 36,9
Atlikus dispersinę analizę nustatyta, kad dėl pavienio UV-B spinduliuotės poveikio vystymosi tarpsnio, kaip veiksnio, įtaka buvo statistiškai reikšminga (p < 0,05). Rūgštus substratas sukėlė didesnius šoninių ūglių formavimosi tarpsnio augalų pažeidimus (38, proc.). Sėjamuosius žirnius veikiant kompleksiškai – UV-B spinduliuote ir rūgštinant substratą (pH 4,8), vidutinis BBCH 14–16 tarpsnio augalų tirtų rodiklių sumažėjimas sudarė 46,9 proc., o BBCH 21–23 tarpsnio – 50,9 procento. Tačiau at-likus dispersinę analizę nustatyta, kad tiek dėl pavienio substrato rūgštumo, tiek dėl kompleksinio pasirinktų stresorių poveikio vystymosi tarpsnio, kaip veiksnio, įtaka nebuvo statistiškai reikšminga (p > 0,05).
Aptarimas. Augalai turi įvairių apsaugos mechanizmų, padedančių kovoti su sukeltu stresu. Suaktyvinę savo apsaugos sistemas, jie gali įveikti stresą, bet kai jis trunka ilgai arba yra stiprus, gynybinės sistemos nualinamos ir tai labai pakenkia augalui arba sukelia jo žūtį (Alexieva ir kt., 2003).
UV-B spinduliuotės ir kitų veiksnių poveikis augalams gali kisti ir priklausomai nuo augalo rūšies (Qaderi ir kt., 2007). Tačiau daugeliu atvejų dėl UV-B poveikio sumažėja augalų imlumas šviesai ir keičiasi fotosintezės proceso intensyvumas. Šio tyrimo metu taip pat labiausiai tiek fotosintezės, tiek kiti rodikliai sumažėjo tuomet, kai žirniai buvo veikti didesne (3 kJ m-2 d-1) UV-B spinduliuote (1–6 pav.). Neigiamas UV-B poveikis fotosintezei siejamas su lapų anatominiais, morfologiniais, citologiniais, fotosintetinių fermentų ir pigmentų sudėties pokyčiais (Zhu, Yang, 2015; Bernal ir kt., 2015). Fotosintezės intensyvumo sumažėjimą lemia antrosios fotosistemos aktyvumo slopinimas bei CO2 fiksavimo reakcijų Kalvino cikle sutrikimas (Albert ir kt., 2011; Zhu, Yang, 2015; Xu, Qiu, 2007).

25
Nedidelės (1 kJ m-2 d-1) UV-B spinduliuotės poveikis tirtiems abiejų tarpsnių (lapų vystymosi ir šoninių ūglių formavimosi) žirniams tiek neutraliame (pH 6,5), tiek ir rūgščiame (pH 4,8) substrate buvo silpnesnis. Taip pat kompleksinis rūgštaus substrato poveikis ir 1 kJ UV-B spinduliuotės poveikis buvo net silpnesnis nei vien rūgštaus substrato poveikis abiejų vystymosi tarpsnių žirniams (1, 4 ir 5 pav.). Panašus dėsnin-gumas buvo nustatytas ir atliekant kitą tyrimą su ridikėliais, kai ridikėlių fotosintezės intensyvumo nuostoliai, veikiant UV-B spinduliuote ir auginant rūgščiame substrate, buvo mažesni, palyginti su fotosintezės nuostoliais padidinus vien substrato rūgštumą. UV-B spinduliuotė stimuliavo rūgščiuose substratuose augusių ridikėlių fermentines fotosintezės reakcijas ir vandens naudojimo efektyvumą (Januskaitiene, 2011).
Literatūroje pateikiama skirtingų duomenų apie UV-B spinduliuotės poveikį fo-tosintezės pigmentams. Vieni autoriai teigia, kad UV-B spinduliuotė labai sumažina chlorofilų ir karotinoidų kiekį ir net vienos rūšies augalų skirtingų veislių reakcija gali labai skirtis (Carletti ir kt., 2003). Kiti mano, kad chlorofilo a ir karotinoidų kiekis lieka stabilus, o tik šiek tiek sumažėja chlorofilo b kiekis (Barsing ir kt., 2000), treti teigia, kad, atvirkščiai, UV-B poveikis skatina sintetinti daugiau chlorofilų ir karotinoidų (Singh ir kt., 2014). UV-B poveikis gali skatinti spinduliuotę absorbuojančių pigmentų sintezę ir tuo pačiu didinti augalų atsparumą (Bernal ir kt., 2015). Šio tyrimo metu nustatytas nežymus chlorofilų a ir b kiekių padidėjimas, esant 1 kJ m-2 d-1 UV-B spin-duliuotės poveikiui tiek neutraliame, tiek ir rūgščiame substrate abiejuose vystymosi etapuose, palyginti su šiuo spinduliuote neveiktų augalų rodikliu (5, 6 pav.). Tačiau 3 kJ m-2 d-1 UV-B spinduliuotė pigmentų kiekį mažino stipriau, ypač rūgščiame substra-te. Tirti veiksniai sėjamųjų žirnių biomasę veikė neigiamai (7 pav.). Yra duomenų, kad sausos biomasės sumažėjimą lemia dėl neigiamo UV-B spindulių poveikio sumažėjęs bendras fotosintezės intensyvumas (Singh ir kt., 2014).
Apibendrinant skirtingų vystymosi tarpsnių sėjamųjų žirnių visų tirtų rodiklių vidutinį sumažėjimą, esant pavieniam bei kompleksiniam UV-B spinduliuotės ir rūgštaus substrato poveikiui (Lentelė), ir atsižvelgiant į dispersinės analizės rezultatus, galima teigti, jog vystymosi tarpsnis tirtų rodiklių kitimui statistiškai reikšmingos įta-kos turėjo tik veikiant UV-B spinduliuote. Didinant tik substrato rūgštumą ir veikiant kompleksiškai (veikiant UV-B spinduliuote rūgščiame substrate augančius žirnius), taikyti veiksniai reikšmingos įtakos neturėjo.
Išvados. Pavienis UV-B spinduliuotės poveikis lapų vystymosi tarpsnio sėja-mojo žirnio tirtiems rodikliams buvo stipresnis nei šoninių ūglių formavimosi tarps-nio augalams. Atlikus dispersinę analizę nustatyta, kad tiek pavienio UV-B spin-duliuotės poveikio, tiek vystymosi tarpsnio, kaip veiksnio, įtaka buvo statistiškai reikšminga (p < 0,05).
Kompleksinis UV-B spinduliuotės ir rūgštaus substrato poveikis lėmė didesnius sėjamųjų žirnių pakenkimus negu pavienis šių veiksnių poveikis.

26
Tiek dėl pavienio substrato rūgštumo, tiek dėl kompleksinio UV-B spinduliuotės ir substrato rūgštumo poveikio tirtų abiejų vystymosi tarpsnių žirnių rodikliai skyrėsi nežymiai: vystymosi tarpsnio, kaip veiksnio, įtaka nebuvo statistiškai reikšminga (p > 0,05).
Padėka. Nuoširdus ačiū Jurgitai Mikelionytei už pagalbą atliekant tyrimus.
Gauta 2015 11 18 Parengta spausdinti 2015 12 15
Literatūra
1. Agati G., Azzarello E., Pollastri S., Tattini M. 2012. Flavonoids as antioxidants in plants: location and functional significance. Plant Sci., 196: 67–76.
2. Albert K. R., Mikkelsen T. N., Ro-Poulsen H., Arndal M. F., Michelsen A. 2011. Ambient UV-B decreases PS II performance and net photosynthesis in high arc-tic Salix arctica. Environ. Exp. Bot., 72: 439–447.
3. Alexieva V., Ivanov S., Sergiev I., Karanov E. 2003. Interaction between stres-ses. Bulgarian Journal of Plant Physiology., Spec. Issue: 1–17.
4. Bernal M., Verdaguer D., Badosa J., Abadia A., Llusia J., Penuelas J., Nunez-Olivera E., Llorens L. 2015. Effects of enhanced UV radiation and water availa-bility on performance, biomass production and photoprotective mechanisms of Laurus nobilis seedlings. Environmental and Experimental Botany, 109: 264–275.
5. Carletti P., Masi A., Grill D., Tausz M., Ferretti M. 2003. Changes in antioxidant and pigment pool dimensions in UV-B irradiated maize seedlings. Environmental and Experimental Botany, 50: 149–157.
6. Duchovskis P. 1998. Problems of resistance to abiotic factors of horticultural plants in Lithuania and their solution. Sodininkystė ir daržininkystė, 17(3): 3–11
7. Growth stages of mono- and dicotyledonous plants. 2001. In: U. Meier (ed.), BBCH Monograph. German Federal Biological Research Centre for Agriculture and Forestry, 10–11.
8. Haling R. E., Simpson R. J., Delhaize E., Hocking P. J., Richardson A. E. 2010. Effect of lime on root growth, morphology and the rhizosheath of cereal see-dlings growing in an acid soil. Plant Soil, 327: 199–212
9. Hollosy F. 2002. Effects of ultraviolet radiation on plant cell. Micron, 33: 179–197.
10. Januskaitienė I. 2011. Effects of substrate acidity and UV-B radiation on photo-synthesis of radishes. Central European Journal of Biology, 6(4): 624–631.
11. Januškaitienė I, Dikšaitytė A. 2014. Surplus fertilization effect on photosynt-hetic response of pea to Cd stress at different growth stages. Polish journal of environmental studies, 23(5): 1 599–1 607.
12. Januškaitienė I. 2013. Low UV-B radiation and substrate acidity impact on pho-tosynthetic parameters of Brassica napus. Biologija, 59(2): 219–230.

27
13. Jukunen-Tiitto R., Haggman H., Aphalo P. J., Lavola A., Tegelberg R., Veteli T. 2005. Growth and defense in deciduous trees and shrubs under UV-B. Environmental Pollution, 137: 404–414.
14. Kataria S., Jajoo A., Guruprasad K. N. 2014. Impact of increasing Ultraviolet-B (UV-B) radiation on photosynthetic processes. Journal of Photochemistry and Photobiology B: Biology, 137: 55–66.
15. Ma J. F. 2007. Syndrome of aluminum toxicity and diversity of aluminum resi-stance in higher plants. Int. Rev. Cyt., 264: 225–252.
16. Paoletti E. 1998. UV-B and acid rain effects on beech (Fagus sylvatica L.) and holm oak (Quercus ilex L.) leaves. Chemosphere, 36(4–5): 835–840.
17. Qaderi M. M., Reid D. M., Yeung E. C. 2007. Morphological and physiological responses of canola (Brassica napus) siliquas and seeds to UVB and CO2 under controlled environment conditions. Environmental and Experimental Botany, 60: 428–437.
18. Reifenrath K., Muller C. 2007. Species-specific and leaf-age dependent effects of ultraviolet radiation on two Brassicaceae. Phytochemistry, 68: 875–885.
19. Sakalauskienė S., Januškaitienė I., Juknys, R., Miliauskienė J. 2013. The effect of UV-B radiation on phytochemical properties of Spinacia oleracea. Proceedings of the 6th international scientific conference „Rural development 2013“, 6(2): 227–231.
20. Shan Y., Izuta T., Totsuka T. 2000. Phenological disorder, cold death of api-cal shoots of red pine subjected to combined exposures of simulated acid rain and soil acidification, and implications for forest decline. Water, Air, and Soil Pollution, 117: 191–203.
21. Singh S., Agrawal S. B., Agrawal M. 2014. UVR8 mediated plant protective res-ponses under low UV-B radiation leading to photosynthetic acclimation. Journal of Photochemistry and Photobiology B: Biology, 137: 67–76.
22. Wei G., Zheng Y., Slusser J. R., Heisler G. M. 2003. Impact of enhanced ultravi-olet B radiation on growth and leaf photosynthetic reaction of soybean (Glicine max). Physiologia Plantarum, 52: 353–362.
23. Wettstein D. 1957. Chlorophyll Letale und der submikroskopische Formweschsel der Plastiden. Experimental Cell Research, 12: 427–432.
24. Xu K., Qiu B. 2007. Responses of superhigh-yield hybrid rice Liangyoupeijiu to enhancement of ultraviolet – B radiation. Plant Science, 3: 139–149.
25. Zhua P., Yang L. 2015. Ambient UV-B radiation inhibits the growth and physio-logy of Brassica napus L. on the Qinghai-Tibetan plateau. Field Crops Research, 171: 79–85.
26. Гавриленко В. Ф., Ладыгина М. Е., Хандобина Л. М. 1975. Большой практикум по физиологии растений. Москва.

28
SODININKYSTĖ IR DARŽININKYSTĖ. SCIENTIFIC ARTICLES. 2015. 34(3–4).
The response of photosynthetic parameters of peas in two growth stages to substrate acidity and UV-B radiation effect
I. Januškaitienė
Summary
The aim of this work was to investigate the combined impact of substrate acidity (pH 4.8) and 1 and 3 kJ m-2 d-1 UV–B radiation on two growth stage pea (Pisum sativum L.) photosynthetic parameters. The investigated plants were sown in vegetative pots with prepared peat substrate. When the plants were at a leaf development stage (BBCH 14–16) and the stage of formation of lateral shoots (BBCH 21–23) the impact of substrate acidity and UV-B radiation was started. Photosynthetic parameters were measured with portable photosynthesis system LI-6400 and photosynthetic pigments were analyzed in acetone extract using a spectrophotometer on the last (5th) day of the experiments. UV-B radiation effect on the investigated indicators of BBCH 14–16 stage peas were stronger than on BBCH 21–23 stage, when the average decrease of all investigated indicators was 34.2 % compared with the control plants. Meanwhile, the decrease of BBCH 21–23 stage was 21.8 %, and the effect of the growth stage, as a factor, was statistically significant (p < 0.05). Substrate acidity had higher (38 %) negative effect on BBCH stage 21–23 peas, but the effect of growth stage was statistically insignificant (p > 0.05). The combined UV-B radiation and substrate acidity affect the parameters of BBCH stage 14–16 peas decreased by 46.9 %, and those of BBCH 21–23 peas slightly more – by 50.9 %, but the growth stage, as a factor, was statistically insignificant (p > 0.05). The combined UV-B radia-tion and substrate acidity effect resulted in higher losses on peas than single ones, and higher losses (p > 0.05) were detected for BBCH 14–16 stage peas.
Key words: Chlorophyll, dry biomass, growth stage, photosynthetic parameters, subs-trate acidity, UV-B.

29
LIETUVOS AGRARINIŲ IR MIŠKŲ MOKSLŲ CENTRO FILIALO SODININKYSTĖS IR DARŽININKYSTĖS INSTITUTO IR ALEKSANDRO STULGINSKIO UNIVERSITETO MOKSLO DARBAI. SODININKYSTĖ IR DARŽININKYSTĖ. 2015. 34(3–4).
Ekologinėmis sąlygomis augintų brokolinių kopūstų sodinimo laiko įtaka produktyvumui
Roma Starkutė, Ona Bundinienė, Vytautas ZalatoriusLietuvos agrarinių ir miškų mokslų centro filialas Sodininkystės ir daržininkystės institutas, Kauno g. 30, LT-54333 Babtai, Kauno r.
2012–2014 metais Lietuvos agrarinių ir miškų mokslų centro (LAMMC) filiale Sodininkystės ir daržininkystės institute buvo tirta ir įvertinta ekologinėmis sąlygomis augintų brokolinio kopūsto (Brassica oleracea L. var. italica Plenck.) hibridų – ‘Fiesta’ ir ‘Belstar’ – sodinimo laiko įtaka rudens sezono derliaus produktyvumui Lietuvos agroklimato sąlygomis.
Atlikus tyrimus nustatyta, kad abu tirti brokolinio kopūsto hibridai tinkami ekologiškai auginti rudens sezono derliui Lietuvos agroklimato sąlygomis.
Ekologinėmis sąlygomis rudeniniam derliui išaugintų brokolinių kopūstų, sodintų birželio mėn. antrojo dešimtadienio pabaigoje, vidutinis bendras derlius buvo 4,01 t ha-1, so-dintų birželio mėn. trečiojo dešimtadienio pabaigoje – 3,19 t ha-1, sodintų liepos mėn. antrojo dešimtadienio viduryje – 3,62 t ha-1, o prekinio derliaus išeiga buvo atitinkamai 52,4, 53,3 ir 60,8 procento. Sodinant daigus birželio mėn. antrojo dešimtadienio pabaigoje ir liepos mėn. antrojo dešimtadienio viduryje, didžiausia buvo hibrido ‘Belstar’ prekinio derliaus išeiga, o sodinant birželio mėn. trečiojo dešimtadienio pabaigoje – hibrido ‘Fiesta’.
Didžiausias bendras ir prekinis derlius gautas sodinant daigus birželio mėn. antrojo de-šimtadienio pabaigoje ir liepos mėn. antrojo dešimtadienio viduryje. Sodinant birželio mėn. trečiojo dešimtadienio pabaigoje, dėl didelių karščių sąlygos daigams prigyti ir augti buvo nepalankios, todėl gautas mažesnis žiedynų derlius.
Nuėmus brokolinių kopūstų pagrindinį derlių, esant palankioms meteorologinėms sąly-goms šoninių ūglių derlių dar galima imti maždaug tris savaites. Vėliau šoninių ūglių žiedynai smulkėja, todėl derlius būna nekokybiškas ir mažas. Tyrimų duomenimis, žiedynai greičiausiai (per 55–70 dienų) susiformavo sodinant daigus birželio mėn. antrojo dešimtadienio pabaigoje, o vėliausiai (per 80–96 dienas) – pasodinus liepos mėn. antrojo dešimtadienio viduryje. Esant sausai ir karštai vasarai derliaus nuėmimo pradžia gali vėluoti maždaug 10 dienų.
Lietuvos agroklimato sąlygomis paskutinis terminas, kai galima sėti (sodinti) brokoli-nius kopūstus rudens derliui, yra birželio mėn. antrojo dešimtadienio pradžia (liepos mėn. antrojo dešimtadienio vidurys). Vėlai rudenį (spalio mėn. antrasis dešimtadienis) iškritus gausesniam kritulių kiekiui, nuo ant brokolinių kopūstų žiedynų užsilaikiusių lietaus lašų ir dėl didelio drėgmės kiekio į galvutę gali patekti bakterinio šlapiojo puvinio sukėlėjas (Erwinia carotovora subsp. carotovora) (Santipracha, 2007). Be to, tuo laikotarpiu jau būna šalnų, dėl to pablogėja žiedynų kokybė.
Reikšminiai žodžiai: brokoliniai kopūstai, ekologinis auginimas, hibridai, rudeninis derlius, sodinimo laikas.

30
Įvadas. Lietuvoje (Baltramaitytė, Rutkovienė, 2000), kaip ir visoje Europoje, sparčiai didėja susidomėjimas ekologiškais produktais. Mokslininkų atlikti tyrimai rodo, kad vartotojai ekologiškus produktus vertina dėl maisto saugos, maistinės vertės, geresnio skonio, palyginti su tradiciškai užauginta produkcija (Skulskis, Girgždienė, 2009; Skulskis ir kt. 2011; Fotini Arvaniti, Zampelas, 2006; Shafie, Rennie, 2012). Todėl daug dėmesio skiriama ekologiniame ūkyje auginamų daržovių asortimentui, parenkamoms veislėms, sodinimo laikui ir užaugintai produkcijai (Lammerts van Bueren ir kt., 2011).
Brokolinis kopūstas (Brassica oleracea L. convar. botrytis L. Alef. var. italica Plenck.) kilęs iš Viduržemio jūros rytinio regiono ir Mažosios Azijos (Decoteau, 2000). Lietuvoje šios daržovės plačiai auginti ir naudoti maistui pradėtos palyginti neseniai. Brokoliniuose kopūstuose randama daug įvairių vitaminų: vitamino C, B grupės vitaminų, vitamino U, beta karoteno ir kitų. Naujausi tyrimai parodė, kad reguliarus brokolinių daržovių vartojimas gali sumažinti besivystančių chroniškų ligų, tokių kaip vėžys, širdies ir kraujagyslių sistemos ligos, II tipo cukrinis diabetas, riziką (Moreno ir kt., 2010; Parente ir kt., 2013).
Brokolinius kopūstus auginti ekologinėmis sąlygomis yra gana sudėtinga, nes jie itin reiklūs maisto medžiagoms, juos puola kenkėjai. Optimali augimo ir vystymosi tem-peratūra – 16–20 °C (Decoteau, 2000; Iglesias-Acosta ir kt., 2010). Vidurio Europoje vyraujančios karštos vasaros mažina žiedynų dydį ir blogina jų kokybę (Grabowska ir kt., 2014). Todėl auginant brokolinius kopūstus ekologinėmis sąlygomis labai svarbu tinkamai parinkti veislę, sodinimo laiką, kad ši vertinga daržovė galėtų būti vartojama šviežia kuo ilgesnį laiką.
Darbo tikslas – ištirti ir įvertinti ekologinėmis sąlygomis auginamų brokolinio kopūsto hibridų tinkamumą rudens derliui, nustatyti tinkamiausią sodinimo laiką Lietuvos agroklimato sąlygomis.
Tyrimo objektas, metodai ir sąlygos. Tyrimai atlikti 2012–2014 me-tais Lietuvos agrarinių ir miškų mokslų centro filialo Sodininkystės ir dar-žininkystės instituto ekologiniame lauke. Dirvožemis – karbonatingasis sekliai glėjiškas priesmėlio ant lengvo priemolio išplautžemis (Idg8-k/Calc(ar)i – Epihypogleyic Luvisols – LVg-p-w-cc) (Buivydaitė ir kt., 2001). Dirvožemio armuo – 22–25 cm. Ariamasis sluoksnis neutraliai šarminis (pH 7,5), mažo azotingumo (Nmin. – 37,3–38,5 kg ha-1), vidutinio humusingumo (2,76 %), didelio fosforingumo (181–214 mg kg-1), vidutinio kalingumo (121–122 mg kg-1), kalcingas, magningas.
Auginti brokolinio kopūsto (Brassica oleracea L. var. italica Plenck.) hibridai: ‘Fiesta’ ir ‘Belstar’.
S o d i n i m o l a i k a s :– 06-19 (birželio mėn. antrojo dešimtadienio pabaiga);– 06-29 (birželio mėn. trečiojo dešimtadienio pabaiga);– 07-13 (liepos mėn. antrojo dešimtadienio vidurys).Daigai buvo išauginti polimerinėse daigyklose. Sudygę jie augo 35–40 dienų. Apskaitomojo laukelio dydis – 8,4 m2 (3 m × 2,8 m). Kiekvienas variantas kartotas
po 4 kartus. Laukeliai buvo išdėstyti sisteminiu būdu. Priešsėlis – vikių ir avižų mišinys žaliajai trąšai. Augalai buvo auginami pagal
ekologiniuose ūkiuose taikomas technologijas. Pavasarį, dirvai pasiekus fizinę brandą,

31
laukas lygintas ir du kartus kultivuotas. Prieš sodinimą patręšta ekologiškomis orga-ninėmis trąšomis Italpollina 4-4-4 (1,2 t ha-1). Daigai sodinti pagal sodinimo tankumo schemą – 0,70 × 0,35 m. Vegetacijos metu papildomai netręšta, esant reikalui, nuo kenkėjų pukšta Nimazaliu (0,5 % tirpalas). Laukas buvo nelaistomas. Pasėlis ravėtas rankomis.
Derlius buvo imtas atrankiniu būdu, išpjaunant jau susiformavusius, bet dar neperaugusius žiedynus. Nustatytas bendras ir prekinis derlius, šoninių ūglių derlius, pagrindinio ir šoninio ūglio žiedyno masė.
Tyrimų duomenys įvertinti dispersinės analizės metodu naudojant kompiuterinę programą ANOVA iš paketo SELEKCIJA (Tarakanovas, Raudonius, 2003).
Bandymų duomenų statistinis patikimumas įvertintas nustatant mažiausią esminio skirtumo ribą (R05).
M e t e o r o l o g i n ė s s ą l y g o s. Bandymų vykdymo metais meteorologinės sąlygos buvo skirtingos. Brokoliniai kopūstai, kaip ir visos kopūstinės daržovės, nereiklūs šilumai ir mėgsta vėsesnius orus, todėl karšti orai, ypač kai trūko drėgmės, stabdė jų vegetaciją.
2012 metai brokoliniams kopūstams augti buvo palankūs: užteko šilumos ir drėgmės (1 lentelė).
1 lentelė. Meteorologinės sąlygosTable 1. Meteorological conditions during investigation years
Babtai, „iMetos“ meteorologinės stoties duomenysData of iMetos meteorological station, Babtai
Mėnuo Month
Vidutinė oro temperatūra Average air temperature, °C
Krituliai Precipitation, mm
2012 m. 2013 m. 2014 m.
daugiametis vidurkis long-term average
2012 m. 2013 m. 2014 m.
daugiametis vidurkis long-term average
Birželis June
14,6 18,0 14,8 15,9 99,6 35,8 84,8 71,2
Liepa July
18,7 18,3 20,4 17,3 135,4 63,6 51,2 75,3
Rugpjūtis August
16,5 17,8 23,2 16,7 81,0 76,2 22,0 78,4
Rugsėjis September
13,1 11,9 12,1 12,1 59,8 86,4 0 58,0
Spalis October
6,9 8,4 7,4 7,1 69,8 45,5 47,0 50,5
Vid. / Avg. 06–10
14,0 14,9 15,6 13,8 89,1 61,5 23,5 66,82

32
Šilumos ir kritulių kiekis buvo artimas daugiamečiam vidurkiui, išskyrus liepos mėnesį, kai iškritęs kritulių kiekis buvo dvigubai didesnis už normą, lyginant su dau-giamečiu vidurkiu, tačiau brokolinių kopūstų augimui ir derliui tai neturėjo neigiamos įtakos.
2013 metai buvo vidutiniškai palankūs augalams augti. Birželis buvo karštas ir trūko drėgmės, todėl daigai prigijo sunkiai. Kritulių iškrito perpus mažiau nei daugia-metis vidurkis. Liepos mėnuo taip pat buvo nepalankus augalams augti: nors orai šiek tiek atvėso, bet iškritęs kritulių kiekis buvo 12 mm mažesnis už daugiametį vidurkį. Todėl gautas mažesnis brokolinių kopūstų derlius, ypač tų, kurių daigai buvo sodinti birželio mėn. trečiojo dešimtadienio pabaigoje. Rugpjūčio ir rugsėjo mėnesiais atvėsę orai ir iškritęs didesnis kritulių kiekis paskatino brokolinių kopūstų augimą. Tai turėjo didelės teigiamos įtakos brokoliniams kopūstams, pasodintiems liepos mėn. antrojo dešimtadienio viduryje.
2014 m. birželio mėnesio meteorologinės sąlygos buvo palankios pasodintiems augalams prigyti. Vidutinė oro temperatūra buvo 14,8 °C, t. y. 1,1 °C žemesnė už vidu-tinę daugiametę. Iškritęs kritulių kiekis buvo 13,6 mm didesnis už daugiametį vidurkį. Liepos ir rugpjūčio mėnesiais vyravę karšti ir sausi orai stabdė brokolinių kopūstų augimą, dėl to derlius pradėtas imti dviem savaitėmis vėliau. Sausas buvo ir rugsėjis.
Rezultatai. Ekologinėmis sąlygomis augintų brokolinių kopūstų derlius skirtin-gais metais buvo nevienodas (2 lentelė).
2 lentelė. Sodinimo terminų įtaka brokolinių kopūstų bendram ir prekiniam der-liuiTable 2. Influence of planting dates on total and marketable yield of broccoli, t ha-1
Tyrimų metai Investigation year
Suminis derlius Total yield
Prekinis derlius Commercial yield
sodinimo laikas planting time R05
sodinimo laikas planting time R05
I II III I II III‘Fiesta’ H
2012 3,00 4,30 3,80 0,28 2,1 2,1 3,1 0,232013 5,37 2,10 3,75 0,25 0,9 0,6 0,9 0,152014 4,00 3,95 3,80 0,23 3,0 2,9 2,6 0,202012–2014 4,16 3,45 3,78 0,25 2,0 1,9 2,2 0,19
‘Belstar’ H2012 3,70 3,40 3,80 0,33 1,8 1,6 3,1 0,232013 4,32 1,80 3,35 0,25 2,3 0,8 1,9 0,212014 3,52 3,55 3,20 0,21 2,1 2,0 1,9 0,212012–2014 3,85 2,92 3,45 0,26 2,1 1,5 2,3 0,18Hibridų vidurkis Hybrid average
4,01 3,19 3,62 0,25 2,1 1,7 2,2 0,18
2012 metais brokolinių kopūstų ‘Fiesta’ H didžiausias bendras derlius (4,3 t ha-1) gautas daigus pasodinus birželio mėn. trečiojo dešimtadienio pabaigoje, o ‘Belstar’ H
33
(3,8 t ha-1) – daigus pasodinus liepos mėn. antrojo dešimtadienio viduryje. Šiek tiek mažesnis (0,1 t ha-1) ‘Belstar’ H derlius gautas pasodinus daigus birželio mėn. antrojo dešimtadienio viduryje. Didžiausias abiejų hibridų prekinis derlius (3,1 t ha-1) gautas daigus pasodinus liepos mėn. antrojo dešimtadienio viduryje. 2012 metais brokolinių kopūstų ‘Fiesta’ H prekinis derlius buvo vidutiniškai 9,1 proc. didesnis negu ‘Belstar’ H. 2013 metais, esant karštai vasarai ir trūkstant drėgmės, abiejų hibridų – ‘Fiesta’ ir ‘Belstar’ – didžiausias bendras derlius (atitinkamai 5,37 ir 4,32 t ha-1) gautas pasodinus brokolinius kopūstus birželio mėn. antrojo dešimtadienio pabaigoje. Pasodinus daigus birželio mėn. trečiojo dešimtadienio pabaigoje, gautas mažas tiek bendras, tiek prekinis abiejų hibridų derlius. Labai mažą prekinį derlių (vidutiniškai 0,8 t ha-1) 2013 metais išaugino brokoliniai kopūstai ‘Fiesta’ H, nepriklausomai nuo sodinimo laiko.
2014 metais didesnių skirtumų tarp brokolinių kopūstų ‘Fiesta’ H, sodintų skir-tingu laiku, bendro ir prekinio derliaus nenustatyta: bendras derlius įvairavo nuo 3,8 iki 4,0 t ha-1, prekinis – nuo 2,6 iki 3,0 t ha-1. Brokolinių kopūstų ‘Belstar’ H bendras ir prekinis derlius (atitinkamai 2,92 ir 1,5 t ha-1) buvo mažiausias daigus pasodinus birželio mėn. trečiojo dešimtadienio pabaigoje.
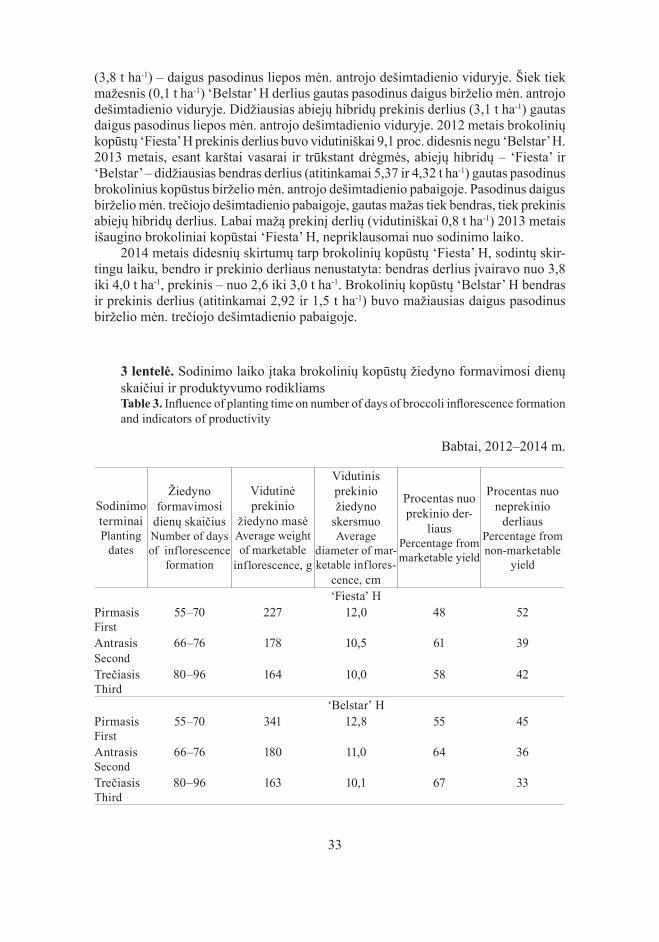
3 lentelė. Sodinimo laiko įtaka brokolinių kopūstų žiedyno formavimosi dienų skaičiui ir produktyvumo rodikliamsTable 3. Influence of planting time on number of days of broccoli inflorescence formation and indicators of productivity
Babtai, 2012–2014 m.
Sodinimo terminai Planting
dates
Žiedyno formavimosi
dienų skaičius Number of days of inflorescence
formation
Vidutinė prekinio
žiedyno masėAverage weight of marketable
inflorescence, g
Vidutinis prekinio žiedyno
skersmuoAverage
diameter of mar-ketable inflores-
cence, cm
Procentas nuo prekinio der-
liaus Percentage from marketable yield
Procentas nuo neprekinio
derliaus Percentage from non-marketable
yield
‘Fiesta’ HPirmasis First
55–70 227 12,0 48 52
Antrasis Second
66–76 178 10,5 61 39
Trečiasis Third
80–96 164 10,0 58 42
‘Belstar’ HPirmasis First
55–70 341 12,8 55 45
Antrasis Second
66–76 180 11,0 64 36
Trečiasis Third
80–96 163 10,1 67 33

34
Brokolinių kopūstų žiedyno susiformavimo laikui didelę įtaką turėjo sodinimo laikas. Vidutiniais duomenimis, žiedynai greičiausiai (per 55–70 dienų) susiformavo sodinant birželio mėn. antrojo dešimtadienio pabaigoje, o vėliausiai (per 80–96 die-nas) – pasodinus liepos mėn. antrojo dešimtadienio viduryje.
Brokolinių kopūstų, pasodintų birželio mėn. antrojo dešimtadienio pabaigoje, žiedyno masė buvo 38 proc. didesnė negu pasodintų liepos mėn. antrojo dešimtadienio viduryje. Prekinių žiedynų masė svyravo nuo 163 iki 341 g. Ta pati tendencija nusta-tyta išmatavus žiedyno skersmenį: didesnis buvo birželio mėn. antrojo dešimtadienio pabaigoje pasodintų kopūstų žiedyno skersmuo. Jis įvairavo nuo 10,0 iki 12,8 cm. Prekinio ir neprekinio derliaus santykis labai priklausė nuo pagrindinio derliaus nu-ėmimo laiko. Pirmojo sodinimo laiko brokolinių kopūstų pagrindinį derlių nuėmus anksčiau, iš lapų pažastyse esančių užuomazgų išaugo daugiau šoninių ūglių žiedynų.
Aptarimas. Auginant augalus, taip pat ir brokolinius kopūstus, ekologinėmis sąlygomis, kyla dvi pagrindinės problemos: apsauga nuo ligų ir aprūpinimas mais-to medžiagomis, nes sintetinių mineralinių trąšų, organinių trąšų, kurioms gaminti naudotos sintetinės mineralinės medžiagos, ir augalų apsaugos preparatų naudojimą griežtai riboja ES reglamentai (Tarybos reglamentas (EB) Nr. 834/2007; Komisijos reglamentas (EB) Nr. 889/2008; Pekarskas, 2008). Todėl norint gauti kokybišką eko-loginėmis sąlygomis auginamų brokolinių kopūstų derlių, reikia pasirinkti tokias veisles, kurios būtų atsparios ligoms ir stresą keliančioms aplinkos sąlygoms (Lam-merts van Bueren ir kt., 2011; Farnham, Björkman, 2011), ir tinkamą sodinimo laiką (Nooprom ir kt., 2013).
Užsienyje ir Lietuvoje atliktais tyrimais nustatyta, kad auginant brokolinius kopūstus įprastinėmis (Jamil Ahmed, Wajid Siddique, 2004) ir ypač ekologinėmis sąlygomis, didelės įtakos jų vystymuisi ir derliui turi šviesos, temperatūros ir kritulių kiekis. Kadangi brokoliniai kopūstai – vėsesnio klimato augalai, todėl temperatūrai pakilus virš +25 °C, jie blogiau auga ir vystosi. Karitonas, Viškelis (1999) teigė, kad vasaros derliui pasodinti brokoliniai kopūstai augo ir formavo žiedynus greičiau negu rudeninio derliaus augalai. Farnham, Björkman (2011) bei mūsų atliktais tyrimais nustatyta, kad geresnės kokybės žiedynų derlius gaunamas auginant brokolinius ko-pūstus rudens derliui.
Lietuvoje atlikti tyrimai patvirtino užsienio mokslininkų teiginį, kad veislė (hibri-das) yra vienas svarbiausių veiksnių, darančių įtaką brokolinių kopūstų derlingumui ir kokybei taikant tiek intensyvią, tiek ekologinę žemdirbystės sistemas (Lammerts van Bueren ir kt., 2011).
Sari ir kt. (2000) teigia, kad pagrindiniai žiedynai sudaro 20–25 proc. viso der-liaus, 75–80 proc. derliaus gaunama iš šoninių žiedynų. Ta pati tendencija nustatyta ir institute atlikus tyrimus. Priklausomai nuo brokolinių kopūstų derliaus nuėmimo laiko prekinis derlius sudarė 55 proc., neprekinis žiedynų derlius – 45 procentus.
LAMMC filiale Sodininkystės ir daržininkystės institute atliktais tyrimais nusta-tyta, kad ‘Fiesta’ ir ‘Belstar’ hibridai tinkami auginti ekologiškai Lietuvos agroklimato sąlygomis.
Nustatyta, kad vėliausiai galima sodinti brokolius rudens derliui liepos mėn. an-trojo dešimtadienio viduryje. Pasodinus šias daržoves vėliau, žiedynams kenkia rudens šalnos ir gausesnis kritulių kiekis. Ant brokolinių kopūstų žiedynų užsilaikę lietaus

35
lašai ir didelis drėgmės kiekis gali sukelti infekciją – augalai gali susirgti bakteriniu šlapiuoju puviniu (Erwinia carotovora subsp. carotovora) (Santipracha, 2007).
Išvados. 1. Ekologinėmis sąlygomis išaugintų brokolinių kopūstų vidutinis bendras derlius buvo didžiausias pasodinus daigus birželio mėn. antrojo dešimtadie-nio pabaigoje – 4,01 t ha-1, birželio mėn. trečiojo dešimtadienio pabaigoje – 3,19 t ha-1 ir liepos mėn. antrojo dešimtadienio viduryje – 3,62 t ha-1.
2. Ekologinėmis sąlygomis rudeniniam derliui išaugintų brokolinių kopūstų vidu-tinė prekinio derliaus išeiga sudarė 52,4 proc. pasodinus daigus birželio mėn. antrojo dešimtadienio pabaigoje, 53,3 proc. – pasodinus birželio mėn. trečiojo dešimtadienio pabaigoje ir 60,8 proc. – liepos mėn. antrojo dešimtadienio viduryje.
3. Didžiausias žiedyno skersmuo ir masė buvo brokolinius kopūstus sodinant birželio mėn. antrojo dešimtadienio pabaigoje.
4. Žiedynai greičiausiai (per 55–70 dienų) susiformavo sodinant birželio mėn. antrojo dešimtadienio pabaigoje, vėliausiai (per 80–96 dienas) – pasodinus liepos mėn. antrojo dešimtadienio viduryje. Esant sausai ir karštai vasarai derliaus nuėmimo pradžia gali vėluoti maždaug 10 dienų.
Gauta 2015 08 09 Parengta spausdinti 2015 09 30
Literatūra
1. Baltramaitytė D., Rutkovienė V. 2000. Kviečių ir miežių grūdų derliaus ir koky-bės tyrimai skirtingose žemdirbystes sistemose. Vagos, 48(l): 12.
2. Buivydaitė V. V., Vaičys M., Juodis J., Motuzas A. 2001. Lietuvos dirvožemių klasifikacija. Vilnius.
3. Decoteau D. R. 2000. Vegetable Crops. Prentice Hall. Upper Saddle River, NJ, USA, 464.
4. Farnham M. W., Björkman T. 2011. Evaluation of experimental broccoli hybrids developed for summer production in the Eastern United States. Hort. Science, 46(6): 858–863.
5. Fotini Arvaniti A., Zampelas A. 2006. Organic Food: Buying More Safety or Just Peace of Mind? A Critical Review of the Literature. Critical Reviews in Food Science and Nutrition, 46: 23–56.
6. Grabowska A., Kunicki E., Kalisz A., Wojciechowska R., Leja M., Sękara A. 2014. Chilling stress applied to broccoli transplants of different age affects yield of the plants cultivated in summer. Hort. Science (Prague), 41: 71–79.
7. Iglesias-Acosta M., Martínez-Ballesta C. M., Teruel J. A., Carvajal M. 2010. The response of broccoli plants to high temperature and possible role of root aquaporins. Environmental and Experimental Botany, 68: 83–90.
8. Jamil Ahmed M., Wajid Siddique. 2004. Effect of Sowing Dates on Growth and Yield of Broccoli (Brassica oleracea L.) under Rawalakot Conditions. Asian Journal of Plant Sciences, 3: 167–169.
9. Karitonas R., Viškelis P. 1999. Brokolinių veislių ir hibridų tyrimas ir biohemi-nės sudėties įvertinimas. Sodininkystė ir daržininkystė, 18(1): 86–97.

36
10. Komisijos reglamentas (EB) Nr. 889/2008, kuriuo nustatomos išsamios Tarybos reglamento (EB) Nr. 834/2007 dėl ekologinės gamybos ir ekologiškų produktų ženklinimo įgyvendinimo taisyklės dėl ekologinės gamybos, ženklinimo ir kon-trolės (OL L 250, 2008 9 18, p. 1). http://eur-lex.europa [žiūrėta 2012 09 21].
11. Lammerts van Bueren E. T., Jones S. S., Tammd L., Murphy K. M., Myers J. L., Leifertf C., Messmer M. M. 2011. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. NJAS – Wageningen Journal of Life Sciences, 58(3–4): 193–205.
12. Moreno A. D., Pérez-Balibrea S., Ferreres F., Gil-Izquierdo A., García-Viguera C. 2010. Acylated anthocyanins in broccoli sprouts. Food Chemistry, 123: 358–363.
13. Nooprom K., Santipracha Q., Te-chato S. 2013. Effect of planting date and va-riety on growth and yield of broccoli during the dry season in southern Thailand. International Journal of Plant, Animal and Environmental Sciences, 3: 121–124.
14. Parente P. C., Reis Lima J. M., Texeira-Lemos E., Moreira M. M., Aquiles A. Barros, Luís F. Guido. 2013. Phenolic Content and Antioxidant Activity Determination in Broccoli and Lamb’s Lettuce. International Journal of Biological, Food, Veterinary and Agricultural Engineering, 7: 7.
15. Pekarskas J. 2008. Tręšimas ekologinės gamybos ūkiuose. Kaunas.16. Santipracha Q. 2007. Vegetable Crop Variety and Growing Season of Southern
Thailand. Text and Journal Publication, Bangkok. 17. Sari N., Yildiz Dasgan H., Abak K. 2000. Effect of sowing times on yield and
head size of broccoli grown in the GAP area, Turkey. Acta Horticulturae (ISHS), 533: 299–306.
18. Shafie F. A., Rennie D. 2012. Consumer perceptions towards organic food. Procedia-Social and Behavioral Sciences, 49: 360–367.
19. Skulskis V., Girgždienė V. 2009. Ekologiškų maisto produktų vartojimas ir jį skatinantys informaciniai šaltiniai Lietuvoje. Žemės ūkio mokslai, 16(3–4): 154–161.
20. Skulskis V., Girgždienė V., Melnikienė R. 2011. Ekologiškų maisto produktų tiesioginis pardavimas. Vadybos mokslas ir studijos – kaimo verslų ir jų infras-truktūros plėtra, 1(25): 208–215.
21. Tarakanovas P., Raudonius S. 2003. Agronominių tyrimų statistinė analizė, tai-kant kompiuterinę programą ANOVA, STAT, SPLIT–PLOT iš paketo IRRISTAT. Akademija, Kėdainių r.
22. Tarybos reglamentas (EB) Nr. 834/2007 dėl ekologinės gamybos ir ekologiš-kų produktų ženklinimo, kuriuo panaikinamas Reglamentas (EEB) Nr. 2092/91 (OL L 189, 2007 7 20, p. 1). http://eur-lex.europa [žiūrėta 2012 08 01].

37
SODININKYSTĖ IR DARŽININKYSTĖ. SCIENTIFIC ARTICLES. 2015. 34(3–4).
Influence of planting time of broccoli grown under organic conditions on productivity
R. Starkutė, O. Bundinienė, V. Zalatorius
Summary
In 2012–2014 the influence of planting time on productivity of autumn seasonal yield of broccoli (Brassica oleracea L. var. italica Plenck.) hybrids ‘Fiesta’ and ‘Belstar’ grown under organic conditions in Lithuanian agroclimate was investigated and evaluated at the Institute of Agriculture, LRCAF.
Having carried out the experiments it was established that both broccoli hybrids investi-gated are suitable to grow as autumn seasonal yield under Lithuanian agroclimatic conditions.
The total average yield of broccoli planted at the end of the second 10-day period of June under organic conditions and harvested in autumn was 4.01 t ha-1, the yield of those planted at the end of the third 10-day period of June was 3.19 t ha-1, and the yield of those planted in the middle of the second 10-day period in July was 3.62 t ha-1; marketable yield output was 52.4, 53.3 and 60.8 percent respectively. The highest marketable yield output was demonstrated by hybrid ‘Belstar’ when it had been planted at the end of the second 10-day period of June and the middle of the second 10-day period in July, however, hybrid ‘Fiesta’ demonstrated the highest marketable yield output when it had been planted at the end of the third 10-day period of June.
The highest total and marketable yield was obtained, when seedlings had been planted at the end of the second 10-day period of June and the middle of the second 10-day period in July. Planting at the end of the third 10-day period of June resulted in lower yield of florets due to high summer temperatures, which caused unfavourable conditions for seedlings to establish and grow.
After harvesting the main crop of broccoli and under favourable meteorological condi-tions the yield of side shoots can still be harvested for three weeks approximately. Later on side shoot florets become smaller, therefore, harvest tends to be of low quality and quantity. According to the investigations florets formed earliest (in 55–70 days) when seedlings planted at the end of the second 10-day period of June, and latest (in 80–96 days) – when planted in the middle of the second 10-day period in July. In dry and hot summer, harvest start can be delayed by 10 days approximately.
The deadline of broccoli sowing (planting) for autumn harvest under Lithuanian agro-climatic conditions is beginning of the second 10-day period of June (middle of the second 10-day period of July). Late in autumn (second 10-day period of October) with more abundant precipitation the causal agent of bacterial soft rot (Erwinia carotovora subsp. carotovora) can be transferred to the head due to raindrops retained by broccoli florets and excessive moisture (Santipracha, 2007). In addition, during this period frosts happen which deteriorate floret quality.
Key words: autumn yield, broccoli, hybrids, organic growing, planting time.

38
LIETUVOS AGRARINIŲ IR MIŠKŲ MOKSLŲ CENTRO FILIALO SODININKYSTĖS IR DARŽININKYSTĖS INSTITUTO IR ALEKSANDRO STULGINSKIO UNIVERSITETO MOKSLO DARBAI. SODININKYSTĖ IR DARŽININKYSTĖ. 2015. 34(3–4).
Lapinių kopūstų, auginamų rudens derliui, produktyvumo tyrimai
Roma Starkutė, Ona Bundinienė, Vytautas ZalatoriusLietuvos agrarinių ir miškų mokslų centro filialas Sodininkystės ir daržininkystės institutas, Kauno g. 30, LT-54333 Babtai, Kauno r.
Auginti lapinių garbanotųjų kopūstų hibridai: ‘Winetou’ ir ‘Redbor’. Tyrimai atlikti Lietuvos agrarinių ir miškų mokslų centro (LAMMC) filiale Sodininkystės ir daržininkys-tės institute 2011–2013 metais. Lietuvos klimato sąlygomis lapinių kopūstų derlius įvairavo nuo 5,24 iki 8,65 kg m-2, prekinio derliaus išeiga buvo 60–68 proc., perdirbimo išeiga – 25–32 procentai.
Hibridas ‘Winetou’ išaugino vidutiniškai 5,5 proc. didesnį bendrą ir 4,7 proc. didesnį prekinį lapų derlių nei hibridas ‘Redbor’. Abu hibridai, sodinti birželio mėn. antrojo dešim-tadienio pabaigoje, palyginti su kitu sodinimo laiku, užaugino didžiausią bendrą derlių, o sodinant birželio mėn. trečiojo dešimtadienio pabaigoje buvo gautas didžiausias prekinis lapų derlius. Sausųjų medžiagų kiekis, pašalinus lapų gyslas, įvairavo nuo 13,5 iki 16 pro-centų. Didžiausias sausųjų medžiagų derlius gautas sodinant kopūstus birželio mėn. antrojo dešimtadienio pabaigoje.
Reikšminiai žodžiai: derlius, lapinis kopūstas, sodinimo laikas.
Įvadas. Lapiniai garbanotieji kopūstai (angl. kale arba borecole; vok. Grünkohl; lot. Brassica oleracea convar. acephala var. sabellica), priklausomai nuo veislės, gali užaugti nuo 45 cm iki dviejų metrų, o lajos skersmuo gali siekti iki 70 cm. Šaknys yra gerai išsivysčiusios, bet išsidėsčiusios dirvos paviršiuje (Parvu, 2005). Šie kopūstai Europoje buvo žinomi dar nuo antikos laikų ir plačiai auginami iki viduramžių. Pagal klasifikaciją šios daržovės artimesnės laukinėms formoms negu kultūrinėms (Balcău ir kt., 2012). Visos dabartinės lapinių kopūstų formos turi vie-ną bendrą protėvį – varietetą Sabellian kale, kuris buvo žinomas senovės romėnų ir graikų jau IV mūsų eros amžiuje (Snogerup ir kt., 1990). Istoriškai lapiniai garba-notieji kopūstai yra pagrindinė daržovė šalto klimato regionuose, nes iš kopūstinių daržovių jie atspariausi žemoms temperatūroms (Altinok, Karakya, 2003). Optimali lapinių kopūstų, kaip ir kitų kopūstinių daržovių, augimo temperatūra – +16–20 °C (Decoteau, 2000).
Lapiniai kopūstai – Lietuvoje retai auginama daržovė, tačiau dėl savo maistinių ir gydomųjų savybių turėtų būti labiau įtraukiama į žmonių mitybos racioną. Ši daržovė išsiskiria vertinga biochemine sudėtimi, kuri lemia ne tik itin gerą maistinę vertę,

39
bet ir gydomąją šių daržovių galią. Lapiniuose kopūstuose gausu angliavandenių (5,3–8,6 proc.), azotinių medžiagų (1,3–3,8 proc.), sieros, fosforo, kalio, folio rūgš-ties, ląstelienos, vitaminų A, B1, B6, C, P, U, riebalų (0,2–0,8 proc.), įvairių fermentų, mineralinių druskų, fitoncidų, geležies, antioksidantų (Acikgoz, 2011; Emebu, Anyika, 2011; Ahmed, Beigh, 2009; Korus ir kt., 2010). Juose randama daug liuteino ir karo-tinoidų, gerinančių regėjimą (Lefsrud ir kt., 2007; Sikora ir kt., 2008), didelis kiekis sausųjų medžiagų (Rosa, Heaney, 1996). Lapiniuose kopūstuose gausu įvairių fenolio junginių, glikozidų. Jie ypač vertingi gydant širdies ligas (Grønbæk, 2014). Tačiau dėl šių medžiagų lapai yra karstelėjusio, aštroko specifinio skonio, kuris ypač stipriai pasireiškia, jei dirvoje trūksta drėgmės ir vyrauja karšti orai (Carlson ir kt., 1987). Lapinių kopūstų augimui ir vystymuisi didelės įtakos turi klimato ir auginimo sąlygos (Babik, 2003), optimalus derliaus nuėmimo laikas (D‘Antuono, Neri, 2003).
Darbo tikslas – įvertinti garbanotųjų lapinių kopūstų hibridų morfologinius požymius, optimalų sodinimo laiką Lietuvos agroklimato sąlygoms ir įtaką derliui ir kokybei.
Tyrimo objektas, metodai ir sąlygos. Lauko bandymai įrengti Lietuvos agra-rinių ir miškų mokslų centro filialo Sodininkystės ir daržininkystės instituto eks-perimentinėje bazėje. Tyrimai atlikti 2011–2013 metais. Auginti garbanotųjų lapinių kopūstų (Brassica oleracea L. convar. acephala var. sabellica) hibridai: ‘Winetou’ ir ‘Redbor’. Lapiniai kopūstai auginti iš daigų, išaugintų polimerinėse daigyklose, pri-pildytose paruošto durpių ir smėlio mišinio santykiu 3 : 1. Čia jie augo 40–55 dienas.
Sodinimo laikas:– birželio mėnesio antrojo dešimtadienio pabaiga; – birželio mėnesio trečiojo dešimtadienio pabaiga; – liepos mėnesio antrojo dešimtadienio vidurys.Visų trijų sodinimo terminų lapinių kopūstų derlius buvo nuimtas vienu metu –
rudenį, po šalnų: 2011-11-02; 2012-10-05; 2013-10-15.Lapinių kopūstų daigai pasodinti 4–5 tikrųjų lapelių tarpsnio lygiame dirvos
paviršiuje pagal sodinimo schemą – 70 × 60 cm. Tyrimų variantai kartoti po keturis kartus. Bandymo apskaitomojo laukelio plotas – 3,36 m2. Laukelyje augo po 8 augalus.
Dirvožemis – karbonatingasis sekliai glėjiškas priesmėlio ant lengvo priemolio iš-plautžemis (IDg8-k/Calc(ar)i – Epihypogleyic Luvisols – LV-p-w-cc) (Buivydaitė ir kt., 2001). Dirvožemio armuo – 20–25 cm. Dirva lapiniams kopūstams sodinti rudenį giliai suarta, pavasarį lyginta ir iki sodinimo kelis kartus kultivuota. Prieš sodinimą išbertos ir įterptos kompleksinės trąšos YaraMila CROPCARE 11-11-21, norma – 800 kg ha-1.
Daigams prigijus (praėjus 15–20 dienų po sodinimo) lapiniai kopūstai patręšti amonio salietra, po 20 dienų – patręšti per lapus 0,2 % kompleksinių trąšų tirpalu. Vegetacijos metu pasėlis ravėtas, purenti tarpueiliai, esant poreikiui augalai laistyti. Biometriniai matavimai atlikti prieš daigų sodinimą į lauką, vegetacijos viduryje ir derliaus nuėmimo metu. Nustatyta augalo masė (g), išmatuotas aukštis (cm), suskai-čiuotas lapų skaičius (vnt.) ant augalo. Atlikta derliaus apskaita. Apskaičiuotas bendras ir prekinis (lapai sveiki, ilgesni kaip 10 cm) lapų derlius su gyslomis ir lapų derlius be gyslų (apdorotų lapų). Sausųjų medžiagų kiekis nustatytas džiovinant mėginius 105 °C temperatūroje iki nekintamos masės (LST ISO 751:2000). Tyrimų duomenys statistiškai įvertinti ANOVA programa (Tarakanovas, Raudonius, 2003).
Vegetacijos metu meteorologinės sąlygos fiksuotos automatine meteorologinių duomenų registravimo stotimi ,,iMetos D“. Tyrimo metais meteorologinės sąlygos buvo skirtingos (Pav.).

40

41
Pav. Meteorologinės sąlygos tyrimo metais. „iMetos“ meteorologinės stoties duomenys.
Fig. Meteorological conditions during investigation years, data of iMetos meteorological station.
Babtai, 2011–2013 m.
M e t e o r o l o g i n ė s s ą l y g o s lapiniams kopūstams augti buvo vidutiniškai palankios. Lapiniai kopūstai, kaip ir visos kopūstinės daržovės, mėgsta vėsų ir drėgną orą, o visais tyrimo metais liepos ir rugpjūčio mėnesiais jų augimą lėtino aukšta tem-peratūra: 2011 metais ji siekė +28,9 °C, o 2012 ir 2013 metais – atitinkamai +30,3 ir +33,3 °C, tačiau liepos mėnesį iškritęs pakankamas kritulių kiekis tiek 2011, tiek 2012 metais atgaivino karščių nualintus augalus (Pav.). 2013 m. liepos ir rugpjūčio mėnesiais taip pat buvo karšta ir trūko drėgmės. Tačiau rugsėjo mėnesį iškritęs pa-kankamas kritulių kiekis, atvėsusios ir pailgėjusios naktys paskatino lapinių kopūstų lapų masės augimą.
Rezultatai. Skirtingais metais lapinių kopūstų derlius buvo nevienodas. Tai la-bai priklausė nuo meteorologinių sąlygų. Vidutiniais duomenimis, lapiniai kopūstai ‘Winetou’ H užaugino vidutiniškai 13,6 proc. didesnį bendrą ir 12,2 proc. didesnį prekinį lapų derlių negu ‘Redbor’ H (1 lentelė).

42
1 lentelė. Lapinių kopūstų derliusTable 1. Kale yield
Babtai, 2011–2013 m.
Metai Year
Derlius (kg m-2) ir sodinimo terminaiYield and planting date
bendrastotal R05
LSD05
prekinismarketable R05
LSD05I II III I II III‘Winetou’ H
2011 8,65 8,28 7,73 0,57 5,75 5,88 5,24 0,122012 7,85 7,23 6,89 0,51 4,35 4,20 4,00 0,412013 6,79 6,50 5,31 0,42 3,75 3,87 3,62 0,452011–2013 7,76 7,34 6,64 0,50 4,62 4,65 4,29 0,33
‘Redbor’ H2011 7,33 7,36 6,88 0,62 4,90 5,10 4,77 0,362012 6,22 6,26 6,35 0,47 3,90 4,15 3,26 0,352013 5,64 6,16 5,24 0,22 3,69 3,26 3,14 0,342011–2013 6,39 6,59 6,15 0,44 4,16 4,17 3,72 0,35
Didžiausias ‘Winetou’ H ir ‘Redbor’ H prekinis lapų derlius (atitinkamai 4,65 ir 4,17 t ha-1) gautas augalus pasodinus birželio mėn. trečiojo dešimtadienio pabaigoje.
Pasodinus lapinius kopūstus birželio mėn. antrojo dešimtadienio pabaigoje, gau-tas didesnis bendras, bet mažesnis prekinis derlius, nes dalis apatinių lapų nudžiūvo. Pasodinus lapinių kopūstų daigus liepos mėn. antrojo dešimtadienio viduryje, abiejų hibridų tiek bendras, tiek prekinis derlius buvo mažiausias.
2 lentelė. Sodinimo laiko įtaka lapinių kopūstų apdorotų lapų derliui ir sausųjų medžiagų kiekiui Tabale 2. Influence of kale planting time for processing leaves and dry matter yield
Babtai, 2011–2013 m.
Metai Year
Derlius ir sodinimo terminaiYield and planting date
apdorotų lapų derliusprocessing yield, kg m-2 R05
LSD05
sausųjų medžiagų kiekis nuo apdorotų lapų derliaus
dry matter of processing yield, kg m-2
R05LSD05
I II III I II III1 2 3 4 5 6 7 8 9
‘Winetou’ H2011 2,98 2,87 2,87 0,21 0,58 0,44 0,30 0,022012 2,15 2,29 2,24 0,19 0,55 0,43 0,34 0,022013 1,90 2,39 1,48 0,20 0,38 0,39 0,31 0,022011–2013 2,34 2,52 1,86 0,20 0,50 0,42 0,32 0,02

43
1 2 3 4 5 6 7 8 9‘Redbor’ H
2011 2,62 2,55 2,54 0,37 0,41 0,39 0,26 0,022012 2,02 1,86 1,62 0,34 0,43 0,40 0,27 0,022013 1,56 1,93 1,71 0,33 0,37 0,37 0,23 0,022011–2013 2,07 2,11 1,96 0,35 0,38 0,40 0,25 0,02
Vidutiniais duomenimis, apdorotų (be gyslų) lapinių kopūstų ‘Winetou’ H lapų der-lius buvo vidutiniškai 9,27 proc. didesnis už ‘Redbor’ H derlių (2 lentelė). Didžiausias lapinių kopūstų ‘Winetou’ H ir ‘Redbor’ H apdorotų lapų derlius (atitinkamai 2,52 ir 2,11 kg m-2) gautas augalus pasodinus birželio mėn. trečiojo dešimtadienio pabaigoje. Didžiausias sausųjų medžiagų derlius gautas lapinius kopūstus pasodinus birželio mėn. antrojo dešimtadienio pabaigoje.
3 lentelė. Lapinių kopūstų ‘Winetou’ H biometrinių rodiklių kitimas per vege-tacijąTable 3. Kale plant weight change during vegetation
Babtai, 2011–2013 m.
Matavimo objektas Measuring object, cm
Dienų skaičius po pasodinimo Number of days after planting
0 30 prieaugis growth 60 prieaugis
growth 90 prieaugis growth
Augalo aukštis Plant height, cm
10 33 +23 60 +27 74 +14
Augalo masė Plant weight, g
1,52 176 +174,48 970 +794 2 180 +1 210
Augalo lapų skaičius, vnt. Plant leaf number (units)
4 17 +13 21 +4 50 +29
Atlikus augalo biometrinius matavimus nustatyta, kad abu lapinių kopūstų hibridai stipriausiai augo vegetacijos pradžioje ir viduryje (3 lentelė). Po daigų pasodinimo į lauką praėjus 30 ir 60 dienų, augalai paaugo atitinkamai 23 ir 27 cm, o vegetacijos pabaigoje per 30 dienų (t. y. praėjus 90 dienų po pasodinimo) – tik 14 cm. Lapinio kopūsto lapų masė didėjo atvirkščiai proporcingai augalo aukščiui. Praėjus 30 dienų po pasodinimo (vegetacijos pradžioje) augalo masė buvo 174,48 g, o didžiausias masės prieaugis – 1 240 ar 1 210 g – nustatytas antroje vegetacijos pusėje (praėjus 90 dienų po pasodinimo), kai sutrumpėjo diena ir atvėso naktys. Daugiausia lapų kopūstai užaugino praėjus 90 dienų po pasodinimo.
Aptarimas. Lapiniai kopūstai gali būti auginami sėjant sėklas tiesiai į lau-ką arba sodinant daigus (Balcău ir kt., 2012; 2013). Korus (2010) teigia, kad
2 lentelės tęsinysTable 2 continued

44
lapiniai kopūstai, išauginti iš daigų, užaugina stabiliai didelį ir didelės mais-tinės vertės lapų derlių. Bendras derlius gali sudaryti apie 10 kg m-2, o prekinis – apie 7,0 kg m-2. Mūsų tyrimo duomenimis, Lietuvos agroklimato sąlygomis lapinių kopūstų bendras derlius sudarė 6,8, o prekinis – 6,4 kg ha-1. Jų augimas labai priklau-sė nuo meteorologinių sąlygų. Pagrindinė lapų masė pradėjo augti esant žemesnei oro temperatūrai ir sutrumpėjus dienai – rugsėjo mėnesį. Tai patvirtina ir kiti auto-riai (Harper, Compton, 1980). Esant sausai vasarai lapinius kopūstus būtina laistyti, pageidautina – lašeliniu būdu.
Rao ir Horn (1986) tyrimų duomenimis, rudenį, praėjus 85–95 dienoms po paso-dinimo, lapiniai kopūstai sukaupia daugiausia sausųjų medžiagų. Harper ir Compton (1980) teigia, kad lapiniuose kopūstuose sausųjų medžiagų daugėja nuo rugsėjo iki lapkričio mėnesio.
Mokslininkų Hagen ir kt. (2009), Steindal ir kt. (2015) tyrimų rezultatai rodo, kad šaltis lapinių kopūstų lapuose padidina tirpaus cukraus kiekį ir sumažina nesočiųjų riebalų, gliukozinolatų ir eruko rūgšties kiekį, todėl lapai pasidaro ne tokie aitrūs, pagerėja skonis. Remiantis šiais šaltiniais, derlius bandymų lauke imtas rudenį, po pirmųjų šalnų. Priklausomai nuo metų meteorologinių sąlygų tai buvo spalio mėnuo ar lapkričio mėn. pirmasis dešimtadienis.
Išvados. 1. Lietuvos agroklimato sąlygomis lapinius kopūstus geriausia sodinti birželio mėn. trečiojo dešimtadienio pabaigoje.
2. Lapiniai kopūstai ‘Winetou’ H užaugino didesnį lapų derlių negu ‘Redbor’ H. 3. Didžiausias lapų masės priaugis gautas rugsėjo mėnesį, kai atvėso oras ir su-
trumpėjo diena, t. y. praėjus 90 dienų po pasodinimo.4. Didžiausias sausųjų medžiagų derlius gautas lapinių kopūstų daigus pasodinus
birželio mėn. antrojo dešimtadienio pabaigoje.
Gauta 2015 09 09 Parengta spausdinti 2015 10 06
Literatūra
1. Acikgoz E. F. 2011. Mineral, vitamin C and crude protein contents in kale (Brassica oleraceae var. acephala) at different harvesting stages. African Journal of Biotechnology, 10(75): 17 170–17 174.
2. Ahmed S., Beigh S. H. 2009. Ascorbic acid carotenoids, total phenolic content and antioxidant activity of various genotypes of Brassica olerecea acephala. Journal of Medical and Biological Sciences, 3(1): 1–7.
3. Altinok S., Karakaya A. 2003. Effect of growth season on forage yields of diffe-rent Brassica cultivars under Ankara conditions. Turkish Journal of Agriculture and Forestry, 27: 85–90.
4. Babik I. 2003. Wpływ niektórych czynników klimatycznych i uprawowych na plon i jakość warzyw przeznaczonych do przetwórstwa. Ogólnopolska Konf. „Uprawa warzyw dla przetwórstwa“. Instytut Warzywnictwa, Skierniewice.

45
5. Balcău S. L., Apahidean M., Zaharia A., Gocan T., Boca D. 2013. The Effects of Some Technological Sequences and Environmental Factors on Plant Development and Yield of Kale (Brassica oleracea var. acephala L.). Bulletin of University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca. Horticulture,70(1): 34–43.
6. Balcău S. L., Apahidean M., Zaharia A., Pop D. 2012. Results regarding the ways to produce seedlings of Brassica oleracea var. acephala. Bulletin USAMV. Horticulture, 69(1): 64–70.
7. Buivydaitė V. V., Vaičys M., Juodis J., Motuzas A. 2001. Lietuvos dirvožemių klasifikacija. Vilnius.
8. Carlson D. G., Daxenbichler M. E., VanEtten C. H., Kwolek W. F.,Williams P. H. 1987. Glucosinolates in crucifer vegetables: broccoli, Brussels sprouts, caulif-lower, collards, kale, mustard greens, and kohlrabi. Journal of the American Society for Horticultural Science, 112(1): 173–178.
9. D’Antuono L. F., Neri R. 2003. Traditional crop revised: yield and quality of palm-tree kale grown as a mechanised industrial crop, as a function of cutting height. Acta Horticulturae, 598: 123–127.
10. Decoteau D. R. 2000. Vegetable Crops. Prentice Hall Inc., Upper Saddle River, New Jersey, 232–237.
11. Emebu P. K., Anyika J. U. 2011. Proximate and Mineral Composition of Kale (Brassica oleracea) Grown in Delta State, Nigeria. Pakistan Journal of Nutrition, 10: 190–194.
12. Grønbæk M. 2014. Effects of cultivation strategies on phytochemicals and sen-sory properties of cabbage (Brassica oleracea L. var. capitata L.) and curly kale (Brassica oleracea L. var. sabellica L.): doctoral dissertation. Aarhus University, Science and Technology, Institute for Fødevarer Department of Food Science, Institute for Fødevarer-Planter, Fødevarer og Bæredygtighed Department of Food Science-Plant, Food & Sustainability, 136.
13. Hagen S. F., Borge G. I., Solhau K. A., Bengtsson G. B. 2009. Effect of cold storage and harvest date on bioactive compounds in curly kale (Brassica olera-cea L. var. acephala). Postharvest Biology and Technology, 51: 36–42.
14. Harper F., Compton I. J. 1980. Sowing date, harvest date and the yield of forage brassica crops. Grass and Forage Science, 35(2): 147–157.
15. Kopsell D. A., Lefsrud M. G., Kopsell D. E., Curran-Celentano J. 2005. Air Temperature Affects Biomass and Carotenoid Pigment Accumulation in Kale and Spinach Grown in a Controlled Environment. HortScience, 40: 2 026–2 030.
16. Korus A. 2010. Level of vitamin C, polyphenols and antioxidant and enzymatic activity in three varieties of kale (Brassica oleracea L. var. acephala) at different stages of maturity. International Journal of Food Properties, 14: 1 069–1 080.
17. Lefsrud M. G., Kopsell D. A., Wenzel A., Sheehan J. 2007. Changes in kale (Brassica oleracea L. var. acephala) carotenoid and chlorophyll pigment con-centrations during leaf ontogeny. Scientia Horticulturae, 112(2): 136–141.
18. LST ISO 751: 2000. Vaisių ir daržovių gaminiai. Vandenyje netirpių sausųjų medžiagų nustatymas, 3.

46
19. Parvu C. 2005. Enciclopedia plantelor, plante din flora României, vol. IV, ed. Tehnică, București.
20. Rao S. C., Horn F. P. 1986. Planting season and harvest date effects on dry matter production and nutrition value of Brassica spp. in Southern Great Plains. Agronomy Journal, 78: 327–333.
21. Rosa E., Heaney R. 1996. Seasonal variation in protein, mineral and glucosi-nolate composition of Portuguese cabbages and kale. Animal Feed Science and Technology, 57(1–2): 111–127.
22. Sikora E., Cieślik E., Leszczyńska T., Filipiak-Florkiewicz A., Pisulewski P. 2008. The antioxidant activity of selected cruciferous vegetable subjected to aquathermal processing. Food Chemistry, 107: 55–59.
23. Snogerup S., Gustafsson M., von Bothmer R. 1990. Brassica sect. Brassica (Brassicaceae) I. Taxonomy and Variation. Willdenowia, 19(2): 271–365.
24. Steindal A. L. H., Rødven R., Hansen E., Mølmann J. 2015. Effects of photope-riod, growth temperature and cold acclimatisation on glucosinolates, sugars and fatty acids in kale. Food chemistry, 174: 44–51.
25. Tarakanovas P., Raudonius S. 2003. Agronominių tyrimų statistinė anali-zė, taikant kompiuterinę programą ANOVA, STAT, SPLIT-PLOT iš paketo SELEKCIJA ir IRRISTAT. Akademija, Kėdainių r.
SODININKYSTĖ IR DARŽININKYSTĖ. SCIENTIFIC ARTICLES. 2015. 34(3–4).
Investigations of productivity of kale grown for autumn harvest
R. Starkutė, O. Bundinienė, V. Zalatorius
Summary
Curly kale hybrids ‘Winetou’ and ‘Redbor’ were grown. Experiments were carried out at the Institute of Horticulture, Lithuanian Research Centre for Agriculture and Forestry (IH LRCAF) in 2011–2013. Under Lithuanian climatic conditions kale yield ranged from 5.24 to 8.65 kg m-2, marketable yield output was 60–68 % and processing output – 25–32 %.
Hybrid ‘Winetou’ produced 5.5 % higher total and 4.7 % higher marketable leaf yield compared with hybrid ‘Redbor’. Both hybrids planted at the end of the second 10-day period of June produced the highest total average yield compared with other times of planting, and those planted at the end of the third 10-day period of June produced the highest marketable leaf yield. Having removed leaf veins dry matter content ranged from 13.5 to 16 percent. The highest dry matter yield was obtained, when kale had been planted at the end of the second 10-day period of June.
Key words: yield, kale, planting time.

47
LIETUVOS AGRARINIŲ IR MIŠKŲ MOKSLŲ CENTRO FILIALO SODININKYSTĖS IR DARŽININKYSTĖS INSTITUTO IR ALEKSANDRO STULGINSKIO UNIVERSITETO MOKSLO DARBAI. SODININKYSTĖ IR DARŽININKYSTĖ. 2015. 34(3–4).
Daržinių braškių ligų sukėlėjai ir jų paplitimo Lietuvoje įvertinimas
Neringa Rasiukevičiūtė, Rytis Rugienius, Jūratė Bronė ŠikšnianienėLietuvos agrarinių ir miškų mokslų centro filialas Sodininkystės ir daržininkystės institutas, Kauno g. 30, LT-54333 Babtai, Kauno r., el. paštas [email protected]
Vieni plačiausiai Lietuvoje auginamų uoginių augalų yra daržinės braškės (Fragaria × ananassa Duch.). Viena žalingiausių braškių ligų yra kekerinis uogų puvinys, dėl jo neten-kama nuo 15 iki 50 proc. derliaus. Nuo 2013 m. Lietuvos agrarinių ir miškų mokslų centro (LAMMC) filialo Sodininkystės ir daržininkystės instituto tyrėjai dalyvauja ERA-NET pro-jekte EUPHRESCO II „Braškių patogenų tyrimas ir paplitimo įvertinimas (Assessment and testing of strawberry pathogens (SPAT)“. Šio projekto dalyviai yra mokslo institucijos iš sep-tynių šalių: Estijos, Austrijos, Airijos, Olandijos, Ispanijos, Rusijos ir Lietuvos. Įgyvendinant šį projektą siekiama įvertinti esamą augalų apsaugos situaciją Europos braškynuose, nustatyti patogenų sudėtį, paplitimą, diagnostinių metodų efektyvumą ir jų tobulinimo galimybes. Iki šiol Europoje trūksta visuotinai adaptuotų, universalių patogenų identifikavimo metodų. Molekuliniai Botrytis cinerea tyrimai, atlikti įvairiose pasaulio šalyse, parodė didelę atskirų šio patogeno kamienų genetinę įvairovę. Pietų ir Vidurio Europos šalyse pradėti tyrimai pes-ticidų poveikio patogenų populiacijų struktūrai nustatyti. Skirtingų veiksnių įtaka patogeno virulentiškumui, atskirų kamienų paplitimui, esant nevienodoms agroklimato sąlygoms, dar nėra pakankamai ištirta. Būtina atlikti kompleksinius daržinių braškių ligas sukeliančių mikroorganizmų vietinių kamienų tyrimus, taikant naujausius molekulinius tyrimo metodus ir suvienodinant juos su kitomis ES šalimis.
Siekiant įvertinti braškių fitosanitarinę būklę Lietuvoje, tyrimo metu buvo parengtas klausimynas braškių augintojams. Apklausa parodė, kad visuose ūkiuose vyravo kekerinio puvinio (Botrytis cinerea) (42 proc.) ir braškių miltligės (Podospaera aphanis) (38 proc.) sukėlėjai. Nustatyta Lietuvoje surinktų B. cinerea izoliatų genetinė variacija.
Reikšminiai žodžiai: Botrytis cinerea, Fragaria × ananassa, SPAT.
Įvadas. Braškės – vienos labiausiai vartojamų desertinių uogų Lietuvoje. 2015 m. Lietuvoje deklaruota 519 ha braškynų. Kekerinis puvinys (Botrytis cinerea) yra viena žalingiausių braškių ligų. Dėl Botrytis spp. sukeliamų puvinių Lietuvoje ir pasaulyje netenkama nuo 15 iki 50 proc. uogų derliaus (Giraud ir kt., 1999; Albertini ir kt., 2002; Fournier ir kt., 2003; Valiuškaitė ir kt., 2008; Asadolahi ir kt., 2013).

48
Ligos sulėtina augalų augimą, sumažina augalų produktyvumą ir pablogina produk-cijos prekinę kokybę (Elad ir kt., 2007; Valiuškaitė ir kt., 2008, 2010, 2013). Botrytis genties patogeninių rūšių biologinė įvairovė didelė ir iki šiol pakankamai neištirta. Nėra iki galo įvertinti Botrytis spp. paplitimas, virulentiškumas, atsparumas pes-ticidams, priklausomai nuo grybo populiacijos genetinės struktūros bei aplinkos. Trūksta visuotinai priimtų, universalių metodų patogenams identifikuoti. Moleku-liniai B. cinerea tyrimai, atlikti įvairiose pasaulio šalyse, parodė didelę atskirų šio patogeno kamienų genetinę įvairovę (Giraud ir kt., 1999; Albertini ir kt., 2002; Four-nier ir kt., 2003; Fekete ir kt., 2012; Asadolahi ir kt., 2013). Pietų ir Vidurio Europos šalyse pradėti pesticidų įtakos patogenų populiacijų struktūrai tyrimai (Veloukas ir kt., 2014; Grabke ir kt., 2014). Tačiau skirtingų veiksnių įtaka patogeno virulentiš-kumui, atskirų kamienų paplitimui, esant nevienodoms agroklimato sąlygoms, dar nėra pakankamai ištirta. Todėl Lietuvoje aktualu atlikti platesnius kompleksinius ligas sukeliančių mikroorganizmų vietinių kamienų tyrimus, taikant naujausius mo-lekulinius tyrimo metodus.
Tradicinis Botrytis spp. rūšių indentifikavimas yra netikslus, todėl pastaruoju metu, norint tiksliau apibūdinti grybų rūšis, taikomi DNR sekvenavimo, polimerazės grandininės reakcijos (PGR), tikrojo laiko kiekybinės PGR (RL-kPGR), mikrosatelitų (SSR) analizės ir kiti molekulinių tyrimų metodai (De Vienne ir kt., 2003; Agrios, 2005; Elad ir kt., 2007).
Atliekant patogenų identifikavimo tyrimus pradėti taikyti molekuliniai metodai atskleidė, kad B. cinerea populiacija skirstoma į dvi virulentiškas grupes, jos pava-dintos filogenetinėmis rūšimis. Nustačius, kuriai grupei priklauso patogenas, galima spręsti apie jo atsparumą fungicidui fenheksamidui, nes I grupės izoliatai atsparūs, o II – jautrūs šiam fungicidui (Fournier ir kt., 2005; Elad ir kt., 2007; Asadollahi ir kt., 2013). B. cinerea giminingos rūšys skiriasi morfologiniais požymiais ir dažnai aptin-kamos viename areale, be to, skirtingi jų augalai šeimininkai (Fournier ir kt., 2003; Fekete ir kt., 2012).
Nuo 2013 m. LAMMC Sodininkystės ir daržininkystės instituto tyrėjai dalyvauja ERA NET (EUPHRESCO II) projekte „Braškių patogenų tyrimas ir paplitimo įvertini-mas (Assessment and testing of strawberry pathogens (SPAT)“, jo dalyviai yra mokslo organizacijos iš septynių šalių: Estijos, Austrijos, Airijos, Olandijos, Ispanijos, Rusijos ir Lietuvos. Projektu siekiama įvertinti esamą augalų apsaugos situaciją Europos braš-kynuose, nustatyti patogenų sudėtį, paplitimą, taikomų diagnostinių metodų efektyvu-mą ir galimybes juos patobulinti bei atlikti bendrus tyrimus (Rugienius ir kt., 2015).
Darbo tikslas – ištirti braškių patogenų paplitimą pagrindiniuose Lietuvos braš-kynuose ir išanalizuoti Botrytis cinerea polimorfizmą.
Tyrimo objektas, metodai ir sąlygos. 2013–2015 m., vykdant projektą, LAMMC Sodininkystės ir daržininkystės institute atlikti braškių patogenų identifi-kavimo ir paplitimo tyrimai. Siekiant įvertinti braškių fitosanitarinę būklę skirtin-gose Europos šalyse, 2013 m., remiantis SPAT projekto dalyvių (tarp jų – ir Lietuvos) pasiūlymais, bendru sutarimu buvo parengtas klausimynas šalių – SPAT dalyvių – ūkininkams. Jis buvo išverstas į lietuvių kalbą, pagal jį parengta anketa ir pateikta 26-iems stambiausiems Lietuvos braškių augintojams.
Klausimynu buvo siekiama išsiaiškinkit ūkio tipą (daigų ar uogų augintojas),

49
ūkininko kompetenciją, nustatyti naujus kvalifikacijos tobulinimo šaltinius: ar ūki-ninkai lanko specialius kursus (1), ar skaito leidinius apie braškių auginimą (2), ar konsultuojasi su agrofirmų konsultantais (3), ar mokosi iš kitų braškių augintojų (4), ar domisi internete pateikta informacija (5). Ūkininkų buvo prašoma nurodyti braškių auginimo sistemą savo ūkyje: nemulčiuotose lysvėse (1), šiaudais mulčiuotose lysvėse (2), plėvele mulčiuotose lysvėse (3), šiltnamiuose (4). Klausimyne buvo pateiktos braškių ligų iliustracijos ir teiraujamasi, kokiomis pagrindinėmis ligomis serga braškės jų ūkyje. Braškių pažeidimo intensyvumas buvo vertinamas balais (balai, atitinkantys pažeistų audinių plotą procentais nuo viso paviršiaus ploto: 1 = 0–5 %, 2 = 5–25 %, 3 = 25–50 %, 4 = > 50 %.
Braškių uogų fragmentai su tipiškais Botrytis spp. sukeltų puvinių požymiais buvo surinkti iš 6 skirtingų rajonų 2012–2014 metais. Iš viso paimti 273 mėginiai, iš jų išskirti izoliatai (1 pav.) ir DNR.
Braškių pažeistos dalys sterilintos 1 proc. natrio hipochlorito tirpale, išdėliotos į sterilias Petri lėkštutes su PDA terpe ir inkubuotos termostate 22 ± 2 °C temperatūroje 7 dienas. Kolonijoms užaugus, iš jų išskirtos monokultūros. Per inkubacijos laikotarpį susiformavęs micelis mentele surinktas nuo Petri lėkštelės (~200 mg), sušaldytas skystame azote ir grūstuvėje sutrintas piestele iki smulkių miltelių. Izoliatų kolekcijos DNR buvo išskirta naudojant DNR išskyrimo rinkinį (Thermo Fisher Scientific Baltics, Lietuva) pagal gamintojo rekomendacijas. DNR ištirpinta vandenyje, laikoma –20 °C temperatūroje.
1 pav. Botrytis spp. izoliatas, auginamas ant bulvių dekstrozės agaro terpės.Fig. 1. Botrytis spp. isolate on potato dextrose agar medium.

50
Botrytis cinerea identifikuoti buvo sudarytas toks PGR reakcijos mišinys: 1 µl mėginio DNR (20 ng), 12,5 µl PGR master reakcijos mišinio (2x), 9,5 µl vandens, po 1 µl (12,5 mol/µl) kiekvieno pradmens – Bc108+ ir Bc563. Visas reakcijos tūris – 25 µl. Temperatūros režimas buvo parinktas pagal Rigotti ir kt. (2006). PGR reakcijos atliktos „Mastercycler“ (Eppendorf, Vokietija) termocikleryje. PGR produktai B. cine-rea identifikuoti buvo frakcionuoti elektroforeze 1,5 proc. agarozės gelyje, dažytame etidžio bromido dažu. DNR fragmentų dydžiams nustatyti naudotas „GeneRulerTM DNA Ladder mix“ (Thermo Fisher Scientific Baltics, Lietuva). B. cinerea izoliatai, atlikus DNR restrikciją, suskirstyti į dvi filogenetines rūšis. Elektroforezės gelyje ma-tomas tiriamas fragmentas: I grupė (atspari fenheksamidui) – 480 bp, II grupė (jautri fenheksamidui) – 360 bp.
Duomenys statistiškai įvertinti ANOVA programa (Tarakanovas, Raudonius, 2003).
Rezultatai ir aptarimas. Lietuvos agrarinių ir miškų mokslų centro filiale So-dininkystės ir daržininkystės institute 2013–2015 m. buvo įvertinta braškių fitosa-nitarinė būklė Lietuvos braškynuose. Tiriamuoju laikotarpiu iš 6 skirtingų rajonų surinkti 273 mėginai, iš jų išskirti izoliatai (1 lentelė).
1 lentelė. Botrytis genties grybų mėginių kolekcija, 2012–2014 m. Table 1. Botrytis spp. isolate collection, 2012–2014
Rajonas District
Vietovė Locality
Metai Year
Veislė Cultivar
Mėginių skaičius
Number of samples
Prienų
nežinoma unknown
2014 ‘Marmolada’, ‘Sonata’, ‘Felicita’ 17
Klebiškis 2014 nežinoma unknown
6
ŠiauliųAdomiškiai 2014 ‘Malvina’ 20Kantminiai 2014 ‘Darselect’, ‘Elene’, ‘Syria’ 22Maniušiai 2014 ‘Sonata’, ‘Syria’ 30
Kauno
nežinoma unknown
2012 nežinoma unknown
1
Kauno m. 2012 ‘Elkat’ 42013 ‘Elkat’ 5
Babtų mstl. 2012 ‘Darselect’, ‘Elkat’ 72013 ‘Darselect’, ‘Venta’, ‘Elkat’ 822014 ‘Darselect’, ‘Elkat’ 10
Radviliškio Velžiai 2014 ‘Pandora’, ‘Sonata’ 19Panevėžio Sodeliškių sen. 2014 ‘Selvik’, ‘Rumba’, ‘Elkat’, ‘Felicita’, ‘Filut’ 19
Kėdainių Akademija 2014 ‘Senga Sengana’ 7Labūnava 2014 ‘Syria’, ‘Vikat’, ‘Pegasus’, ‘Pandora’ 24
Iš viso Total
273

51
Daugiausia izoliatų išskirta iš ‘Elkat’, ‘Syria’, ‘Sonata’, ‘Pandora’ ir ‘Darselect’ veislių braškių, kurios buvo auginamos dažniausiai.
Siekiant įvertinti braškių fitosanitarinę būklę įvairiose Europos šalyse, 2013 m., remiantis SPAT projekto dalyvių pasiūlymais, šalių – SPAT dalyvių – ūkininkams buvo parengtas klausimynas. Iš 26 ūkininkams išsiųstų anketų buvo gautos tik 5 užpildytos anketos. Į anketas atsakė 19 proc. respondentų. Anketos gautos iš šių braš-kių augintojų: 1 ūkis – Kantaminių k., Šiaulių r., 2 ūkis – Bajoriškių k., Lazdijų r., 3 ūkis – Sodeliškių sen., Panevėžio r., 4 ūkis – Smalininkų k., Jurbarko r. ir 5 ūkis – Milgaudžių k., Tauragės r.
Išanalizavus anketų duomenis nustatyta, kad 29 proc. apklaustųjų yra braškių daigų, 71 proc. – braškių uogų augintojai (2 pav.). Norėdami patobulinti žinias braškių auginimo bei priežiūros klausimais, 27 proc. ūkininkų lanko specializuotus kursus, 27 proc. konsultuojasi su specializuotais konsultantais.
2 pav. Braškių augintojų grupės ir jų kvalifikacijos kėlimo šaltiniai.Fig. 2. Strawberry grower groups and sources of education on strawberry production.
Anketų duomenys parodė, kad 37 proc. braškių augintojų braškes augina šiaudais mulčiuotose lysvėse, 25 proc. – nemulčiuotose, 25 proc. – plėvele mulčiuotose ir tik 13 proc. – šiltnamiuose (3 pav.). Iš anketų duomenų nustatyta, kad daugelyje ūkių auginamos ‘Darselect’, ‘Elkat’, ‘Asia’ ir ‘Sonata’ veislių braškės. Uselis ir kt. (2008) tyrimais nustatė, kad geriausiai peržiemoja balta plastikine plėvele mulčiuotos braš-kės, o nemulčiuotos braškės peržiemojo prasčiau. Taip pat Uselis ir kt. (2009) tyrimais nustatė, kad, siekiant išlaikyti augalus sveikus, gauti didžiausią derlių ir pelną, labai svarbu pasirinkti tinkamą braškių auginimo sistemą ir tinkamas auginti veisles.

52
3 pav. Tirtuose braškių augintojų ūkiuose naudotos braškių auginimo sistemos.Fig. 3. Information about strawberry growing systems.
Anketų duomenimis, tirtuose ūkiuose braškių ligų paplitimas ir intensyvumas nevienodas (4 pav.). Ligų paplitimą įvertinus 0–5 balų sistema daugiausia ligų nusta-tyta Bajoriškių k., Lazdijų r., esančiame braškių ūkyje (2 ūkis), čia taip pat labiausiai buvo pažeisti augalai.
4 pav. Grybinių ligų patogenų paplitimo intensyvumas braškyne skirtinguose ūkiuose, 2014 m.
Fig. 4. Strawberry infected by fungal pathogens on different farms, 2014.

53
Mažiausiai ligų pakenktų braškių buvo Smalininkų k., Jurbarko r., esančiame ūkyje (4 ūkis). Ligų plitimo įvertinimas parodė, kad įvairiose vietovėse susidarė nevienodos sąlygos ligoms plisti.
Visuose ūkiuose vyravo kekerinio puvinio (Botrytis cinerea) (42 proc.) ir braškių miltligės (Podospaera aphanis) (38 proc.) sukėlėjai. Nustatyta, kad Bajoriškių k., Lazdijų r., esančiame ūkyje (2 ūkis) buvo daugiau patogenų pažeistų augalų nei kituose ūkiuose. Iš anketų duomenų nustatyta, kad Lietuvoje braškės dažniausiai pažeidžiamos kekerinio puvinio ir lapų ligų. Tą rodo Valiuškaitės ir kt. (2008, 2010) bei Uselio ir kt. (2006) atlikti tyrimai. Todėl galima teigti, kad ūkininkai, atsakę į anketos klausimus, yra kvalifikuoti ir gana objektyviai vertina braškių ligų paplitimą savo ūkiuose.
Detaliau išanalizavus anketų duomenis nustatyta, kad braškių lapus dažniausiai pažeidžia šie patogenai: Mycosphaerella fragariae – 12 proc., Marssonina potentil-lae – 16 proc. ir Podospaera aphanis – 16 proc., uogas – Botrytis cinerea – 18 proc. (5 pav.). Uselis ir kt. (2006) tyrimais nustatė, kad Mycosphaerella fragariae padarytų pažeidimų intensyvumas sudaro 14–15,7 procento.
5 pav. Braškių augalų ir uogų ligų sukėlėjai, 2014 m.Fig. 5. Strawberry disease infections, 2014.
Iš surinktų izoliatų išskirta DNR buvo naudota patogeno Botrytis spp. genetinės įvairovės analizei atlikti. Atlikus sudėtinę PGR reakciją, iš 79 tirtų mėginių daugiausia nustatyta II grupės, t. y. fungicidui fenheksamidui jautrių, o I grupės mėginių, atsparių šiam fungicidui, rasta daug mažiau (6 pav., 2 lentelė).

54
6 pav. B. cinerea izoliatų identifikavimas naudojant PGR. Elektroforegramoje matyti tiriami DNR fragmentai – 360 bp ir 480 bp.
Fig 6. B. cinerea isolate identification after PGR. Identification fragment is 360 bp and 480 bp.
Tyrimų laikotarpiu atlikus PGR analizę nustatyti 73 mėginiai (II grupė), amplifi-kuojantys 480 bp dydžio DNR fragmentą, ir 6 (I grupės) – 360 bp fragmentą (7 pav.) Iš tirtų izoliatų atsparių fungicidui fenheksamidui (I grupės) buvo 6 (7,59 proc.), o jautrių (II grupės) – 73 (92,41 proc.) (2 lentelė).
2 lentelė. B. cinerea filogenetinių grupių paplitimas Table 2. Botrytis cinerea phylogenetic group strains
VietaLocation
VeislėCultivar
GrupėGroup
I (atspari)(resistant)
II (jautri)(sensitive)
1 2 3 4Babtai (Kauno r.) ‘Venta’ 2 5
‘Darselect’ 2 15‘Elkat’ 1 7‘Darselect’ 0 1
Sodeliškių sen. (Panėvėžio r.)‘Felicita’ 0 1‘Filut’ 0 2‘Felicita’ 0 1‘Elkat’ 0 2
Klebiškis (Prienų r.) ‘Marmalade’ 0 1‘Sonata’ 0 1‘Felicita’ 0 1
Maniušių k. (Šiaulių r.) ‘Sonata’ 0 1‘Syria’ 1 2

55
1 2 3 4Labūnava (Kėdainių r.) ‘Pegasus’ 0 3
‘Vilkat’ 0 1‘Darsect’ 0 1
Akademija (Kėdainių r.) ‘Senga sengana’ 0 4Velžiai (Radviliškio r.) ‘Pandora’ 0 6
‘Sonata’ 0 3Adomiškių k. (Šiaulių r.) Nežinoma
unknown0 7
Nežinoma (Prienų r.) Nežinomaunknown
0 3
Gruzdžiai (Šiaulių r.) ‘Syria’ 0 4‘Elene’ 0 2
Iš visoTotal
6 73
Gauti rezultatai rodo, kad tiriamuoju laikotarpiu Lietuvoje vyravo B. cinerea II grupė – jautri veikliajai medžiagai fenheksamidui. Remiantis literatūros duomenimis nustatyta, kad II grupė vyrauja ir pasaulyje (Giraud ir kt., 1999; Fournier ir kt., 2005; Fournier ir Giraud, 2008). Giraud ir kt. (1999) tyrimais nustatyta, kad I grupės izoliatų buvo 2,3 proc., o II grupės – 97,7 procento. DNR polimorfizmo tyrimai parodė, kad I grupės variacija didesnė nei II grupės (Giraud ir kt., 1999; Fournier ir kt., 2003).
Išvados. 1. Lietuvos braškių augintojų apklausa parodė, kad visuose ūkiuose vyravo kekerinio puvinio (Botrytis cinerea) (42 proc.) ir braškių miltligės (Podos-paera aphanis) (38 proc.) sukėlėjai. Braškių ligų paplitimas ir jų pažeistų augalų skaičius Lietuvos braškynuose skiriasi.
2. Dvylikoje skirtingų Lietuvos braškynų surinkti 273 Botrytis spp. mėginiai, su-kurta izoliatų kolekcija, išskirtos šių izoliatų grynos kultūros bei DNR molekuliniams tyrimams atlikti.
3. Nustatyta Lietuvoje surinktų B. cinerea izoliatų genetinė variacija. Botrytis cinerea populiacijoje vyrauja (92,41 proc.) atsparūs fungicidui fenheksamidui (II grupės) izoliatai.
Padėka. Tyrimai atlikti pagal ERA-NET projektą EUPRESCO II, finansuoja-mą Lietuvos Respublikos žemės ūkio ministerijos (sutarties Nr. TM-13-3).
Gauta 2015 11 18 Parengta spausdinti 2015 12 17
2 lentelės tęsinysTable 2 continued

56
Literatūra
1. Agrios G. E. 2005. Plant pathology. Amsterdam.2. Albertini C. G., Fournier E., Leroux P. 2002. Erubicol 14α-demethylase
gene (cyp 51) polymorphism and speciation in Botrytis cinerea. Mycological Research, 106: 1 176–1 178.
3. Asadollahi M., Fekete E., Karaffa L., Flipphi M., Árnyasi M., Esmaeili M., Váczy K. Z., Sándor E. 2013. Comparison of Botrytis cinerea populations iso-lated from two open-field cultivated host plants. Microbiological Research, 168(6): 379–388. http://doi: 10.1016/j.micres.2012.12.008
4. De Vienne D., Santoni S., Falque M. 2003. Principal sources of molecu-lar markers. In: D. de Vienne (ed.), Molecular Markers in Plant Genetics and Biotechnology. Science Publishers, Inc., Enfield, NH, 3–46.
5. Elad Y., Williamson B., Ttudzynski P., Delen N. 2007. Botrytis: Biology, Pathology and Control. Dordrecht, The Netherlands.
6. Fekete E., Fekete E., Irinyi L., Karaffa L., Arnyasi M., Asadollagi M., Sandor E. 2012. Genetic diversity of a Botrytis cinerea cryptic species complex in Hungary. Microbiological Research, 167: 283–291.
7. Fournier E., Giraud T., Albertini A., Brygoo Y. 2005. Partition of the Botrytis cinerea complex in France using multiple gene genealogies. Mycologia, 97: 1 251–1 267.
8. Fournier E., Giraud T. 2008. Sympatric genetic differentiation of a generalist pathogenic fungus, Botrytis cinerea, on two different host plants, grapevine and bramble. Journal of Evolutionary Biology, 21(1): 122–132. Doi: 10.1111/j.1420-9101.2007.01462.x
9. Fournier E., Levis C., Fortini D., Leroux P., Giraud T., Brygoo Y. 2003. Characterization of Bc-hch, the Botrytis cinerea homolog of the Neurospora crassa het-c vegetative incompatibility locus, and its use as a population marker. Mycologia, 95(2): 251–261.
10. Giraud T., Fortini D., Levis C., Lamarques C., Leroux P., LoBuglio K., Brygoo Y. 1999. Two sibling species of the Botrytis cinerea complex, transposa and Vacuma, are found in sympatry on numerous host plants. Phytopathologia, 89: 967–973. http://doi: 10.1094/PHYTO.1999.89.10.967
11. Grabke A., Fernández-Ortuño D., Amiri A., Li X., Peres N. A., Smith A., Schnabel G. 2014. Characterization of iprodione resitance in Botrytis cinerea from strawberry and blackberry. Phytopathology, 104: 396–402.
12. Rigotti S., Viret O., Gindro K. 2006. Two new primers highly specific for the de-tection of Botrytis cinerea Pers. Fr. Phytopathologia Mediterranea, 45: 253–260.
13. Rugienius R., Šikšnianienė J. B., Rasiukevičiūtė N. 2015. Braškių patogenų ty-rimas ir paplitimo įvertinimas. Mokslinės konferencijos pranešimai „Agrariniai ir miškininkystės mokslai: naujausi tyrimų rezultatai ir inovatyvūs sprendimai“, 5: 104–105.
14. Tarakanovas P., Raudonius S. 2003. Agronominių tyrimų duomenų statistinė analizė taikant kompiuterines programas ANOVA, STAT, SPLIT-PLOT iš pake-to SELEKCIJA ir IRRISTAT. Akademija, Kėdainių r.

57
15. Uselis N., Lanauskas J., Valiuškaitė A., Viškelis P. 2009. Braškių veislių tyrimas, auginant jas profiliuotoje dirvoje po priedanga. Sodininkystė ir daržininkystė, 28(4): 51–60.
16. Uselis N., Lanauskas J., Zalatorius V., Duchovskis P., Brazaityte A., Urbonaviciute A. 2008. Evaluation of the methods of soil cultivation growing dessert strawberries in beds. Sodininkystė ir daržininkystė, 27(2): 295–305.
17. Uselis N., Valiuškaitė A., Raudonis L. 2006. Incidence of fungal leaf diseases and phytophagous mites in different strawberry cultivars. Agronomy research, 4: 421–425.
18. Valiuškaitė A., Raudonis L., Survilienė E. 2008. Control of grey mould and white leaf spot in strawberry. Zemdirbyste-Agriculture, 95(3): 221–226.
19. Valiuškaitė A., Survilienė E., Duchovskienė L, Rasiukevičiūtė N., Tamošiūnas R. 2013. Sodo ir daržo augalų apsaugos nuo ligų ir kenkėjų naujausi tyrimai. Sodininkystė ir daržininkystė, 32(3–4): 129–137.
20. Valiuškaitė A., Uselis N., Survilienė E. 2010. iMETOS®sm Botrytis sp. progno-zavimo modelio efektyvumo tyrimai braškyne. Sodininkystė ir daržininkystė, 29(3): 13–21.
21. Veloukas T., Kalogeropoulou P., Markoglou A. N., Karaglanidis G. S. 2014. Fitness and competitive ability of Botrytis cinerea field isolates with dual re-sistance to SDHI and QoI fungicides, associated with several sdhB and cytb G143A mutations. Phytopathology, 104: 347–356.
SODININKYSTĖ IR DARŽININKYSTĖ. SCIENTIFIC ARTICLES. 2015. 34(3–4).
Estimation of Strawberry Pathogen Prevalence in Lithuania
N. Rasiukevičiūtė, R. Rugienius, J. B. Šikšnianienė
Summary
Strawberry (Fragaria × ananassa) is one of the most popular fruits in Lithuania. According to the data by the Statistical Department of 2015 519 ha of strawberries were grown in Lithuania. One of the most important strawberry diseases is grey mould, it causes yield losses from 15 % up to 50 %. The molecular investigations of B. cinerea in different countries revealed significant genetic diversity of this pathogen. The researchers of the Institute of Horticulture (LRCAF) have been involved in the ERA-NET project EUPHRESCO II Assessment and test-ing of strawberry pathogens (SPAT) since 2013. There are 7 partners from Estonia, Austria, Ireland, the Netherlands, Spain, Russia and also Lithuania participating in this project. The aim of this project is to evaluate the plant protection situation in strawberry plantations in Europe, identify the composition of pathogens, their prevalence, efficiency of diagnostic methods and improvement opportunities. The lack of fast and accurate methodology in Europe makes it necessary to characterize the distribution of the pathogens. B. cinerea research revealed high genetic diversity among different strains of the pathogen. Pesticides that affect the pathogen population structure are being investigating in Europe. Different factors affecting the influence of the pathogen virulence and incidence in different agroclimatic conditions are not explored. It is necessary to carry out the investigation of local pathogenic microorganism strains using

58
the latest molecular methods.In order to assess the phytosanitary condition of Lithuanian strawberry farms a question-
naire was prepared. The survey showed that on all farms 42 % of grey mould (B. cinerea) and 38 % of powdery mildew (P. aphanis) dominated on all farms. During the research a genetic variation among isolate collection was found.
Key words: Botrytis cinerea, Fragaria × ananassa, SPAT.

59
SCIENTIFIC WORKS OF THE INSTITUTE OF HORTICULTURE, LITHUANIAN RESEARCH CENTRE FOR AGRICULTURE AND FORESTRY AND ALEKSANDRAS STULGINSKIS UNIVERSITY. SODININKYSTĖ IR DARŽININKYSTĖ. 2015. 34(3–4).
Possibility to control diseases of white cabbage with Cabrio Duo
Elena Survilienė, Neringa Rasiukevičiūtė, Laisvūnė DuchovskienėInstitute of Horticulture, Lithuanian Research Centre for Agriculture and Forestry, Kauno 30, LT-54333 Babtai, Kaunas distr., Lithuania, e-mail [email protected]
The objective of studies was to establish the influence of new fungicide Cabrio Duo (a. i. pyraclostrobin 40 g l-1 + dimethomorph 72 g l-1) and compare its effects with the standard fungicide Signum (a. i. boskalid 267 g ha-1 + pyraclostrobin 67 g ha-1) to control such dise-ases as Alternaria leaf spot, grey mould, ring spot, sclerotinia and bacterial rots in cabbage during vegetation period and storage. The investigations were carried out in the Institute of Horticulture, LRCAF in 2010–2011. Three fungicide applications with Cabrio Duo at the 1.25 l ha-1 and 2.5 l ha-1 rates and Signum at the rate of 1.0 kg ha-1 were made on ‘Mentor’ H and ‘Jaguar’ H cultivars during the second part of plant vegetation (growth stage BBCH 41–46). After harvesting the samples of cabbage heads were kept in cold storage for four months. The obtained data showed that fungicide treatments allowed for reducing the incidence of diseases and reduced losses during the four–month storage period. The efficiency of Cabrio Duo at the 1.25 l ha-1 and 2.5 l ha-1 rates to control disease did not differ compared with the standard fungicide Signum at the rate of 1.0 kg ha-1, so it can be used in plant protection systems.
Key words: Alternaria leaf spot, bacterial rot, cabbage, efficiency, grey mould, ring spot, sclerotinia rot, fungicides.
Introduction. Cabbage is one of the main vegetables grown in Lithuania, so much attention is paid to its quality. Very often various infectious diseases, which cause the disorders of vegetable growth and development, damage cabbage more than other agricultural plants. The main economically important diseases and their pathogens of cabbages are as follows: dark leaf spot, Alternaria leaf spot, grey mould, sclerotinia and bacterial rots (Bedlan, 1998; Datta, Gopal, 1999; Geeson, 1983). These diseases do especially big damages both to grown and stored vegetables (Azevedo et al., 2000; Bedlan, 2001; Agblor, Waterer, 2001; Surviliene et al., 2010).
By reducing the spread of infection along with all the cultural practices of technological requirements, an important focus is on plant protection products. To prevent fungal spoilage it is a common practice in many countries to spray cabbages

60
with systemic and/or protective fungicides in the field prior to storage (Bedlan, 1998; Adamicki, Robak, 2000; Bartlett et al., 2002; Callens et al., 2005; Mazzini, 2009).
The aim of the investigation was to establish the influence of new fungicide Cabrio Duo and compare its effects with the standard fungicide Signum to control diseases in cabbage during vegetation period and storage.
Materials and Methods. The investigations were carried out at the Institute of Horticulture, Lithuanian Research Centre for Agriculture and Forestry during 2010–2011 to evaluate the effect of fungicide sprays on the management of diseases on white cabbage.
The trial field was a marginally rolling plain with microrelief. The soil of the experimental site is Epicalcari-Epihypogleyic Cambisol, CMg-p-w-cap; with a tex-ture of light clay loam and medium clay loam. The arable layer was weakly alkaline (pH 7.3–7.6), medium humus-rich (2.06–2.07 %).
The seedlings of white cabbage (Brassica oleracea L. convar. Capitata (L) Alef. var. alba) cultivars ‘Mentor’ H and ‘Jaguar’ H (Bejo Zaden, Holland) were planted in the beginning of May, rate – 31 700 seedlings ha-1 (70.0 cm × 45.0 cm). Both cabbage cultivars are popular among growers, characterized by resistance to Fusarium disease, a good storage ability and were grown under intensive cultivation technologies ac-cepted at the Institute of Horticulture and involved: fertilization before planting with (N) 200 kg ha-1, (P2O5) 120 kg ha-1, (K2O) 120 kg ha-1 and additionally with Ca (NO3)2 400 kg ha-1; weed control with Lontrel 300 SL 0.5 l ha-1, Lentagran WP 2.0 kg ha-1, hoed inter-rows at the depth of 5–8 cm and weeded; pest control with Decis Mega 50 EW 0.15 l ha-1, Fastac 50 EC 0.2 l ha-1 and Proteus 110 OD 0.75 l ha-1, if necessary. During both experimental years summer wheat was a preceding crop for cabbage.
The trials were arranged in the plots replicated four times, plot size – 22.4 m-2 (8 m × 2.8 m), 4 replications and random plot distribution (40 plants/12 m2). The trial design involved the following fungicides with active ingredients at the following rates: pyraclostrobin 40 g l-1 + dimethomorph 72 g l-1 (Cabrio Duo) and boskalid 267 g ha-1 + pyraclostrobin 67 g ha-1 (Signum, as standard). Three fungicide applications every 10–14 days were made on tested cabbage at the stage of growth BBCH 41–46, when it is usually recommended for the production of cabbage for longer storage. Surfactant Kinetic at the rate of 0.15 l ha-1 was added to the spray solution (volume of water 500 l ha-1). The last spray was conducted not later than 21 days before harvesting.
The cabbage was harvested in the middle of October. Cabbage heads (20 units from every replication) were kept in cold storage (at the temperature of 0–2 °C, relative air humidity – 90 %) for four (November–February) months after harvest. Lesions of diseases were assessed every month. Disease incidence and severity were assessed on 20 cabbage plants per plot according to the generally accepted experimental methods, calculating the percentage of infected leaves and the percentage of infected leaf area (Žemės ūkio augalų kenkėjai…, 2002). Statistical processing of the data was done using Duncan’s multiple range test at P = 0.05 (Tarakanovas, Raudonius, 2003).
The weather conditions during 2010 and 2011 vegetation periods was favourable for cabbage growth. The mean temperature was higher during all investigation months as compared with the multiyear average, but without low or high extremes. There was a lot of rain during the growing season, especially in May, July and August (amount

61
of precipitation was 1.4–1.7 times higher than the multiyear average in 2010). In 2011 it was very rainy in June and July, with the precipitation of 4 and 2 times higher com-pared with the multi-annual average. A higher temperature combined with abundant rain was favourable for infection processes.
Results and Discussion. During 2010–2011 trials Alternaria leaf spot (Alter-naria brassicae (Berk.) Sacc., Alternaria brassicicola (Schwein.) Wiltshire), grey mould ((Botryotinia fuckeliana (de Bary) Whetzel, anamorfa – Botrytis cinerea Pers.), ring spot (Mycosphaerella brassicicola (Duby) Lindau), sclerotinia rot (Scle-rotinia sclerotiorum (Lib.) de Bary) and bacterial rot (Erwinia carotovora subsp. carotovora (syn. Pectobacterium carotovorum subsp. carotovorum Jones) were es-tablished on white cabbage. The prevalence of diseases was not uniform. During both years, Alternaria leaf spot was the main and most widespread disease on the cabbage (Fig. 1.). The prevalence of Alternaria leaf spot was up to 72.97–89.6 % in an untreated plot, the prevalence of other diseases was sporadic. Grey mould was also significant; prevalence was up to 9.1–18.0 %. The difference between the tested ‘Mentor’ H and ‘Jaguar’ H cultivars of cabbage was not found.
Fig. 1. Disease prevalence in white cabbage ‘Mentor’ H and ‘Jaguar’ H in 2010–2011.
1 pav. Ligų paplitimas ‘Mentor’ H ir ‘Jaguar’ H veislių baltagūžių kopūstų lauke, 2010–2011 m.
The first single symptoms of Alternaria leaf spot were observed in early August. The first applications with fungicides were carried out on 18 August (2011) and on 25 August (2010). According to the test data the severity of Alternaria leaf spot in

62
the untreated (control) treatment increased by 11.5–16.25 % (from 8.75–11.75 % to 20.25–28.0 %) on average during the period from the middle of August until the beginning of October (Fig. 2.).
Fig. 2. Severity of Alternaria leaf spot in white cabbage in untreated plot during 2010–2011 growing season.
2 pav. Alternariozės intensyvumas kontrolės variante 2010–2011 m. vegetacijos periodu.
Meanwhile, the severity of Alternaria leaf spot decreased after 3 applications with fungicides as well as with standard fungicides Signum and Cabrio Duo. The intensity of disease was 6.9 times lower in the treatment with Cabrio Duo at the rate of 1.25 l ha-1 and respectively, 7.65 – with Cabrio Duo at the rate of 1.25 l ha-1 and 8.54 – with Signum at the rate of 1.0 kg ha-1 (Table 1).
The efficiency of Cabrio Duo at the rate of 1.25 l ha-1 following 21 days after the last (third) application was 85.24 % on average and at the rate of 2.5 l ha-1 – 86.75 % respectively. It is noted, that the different rates of Cabrio Duo had no significant effect on Alternaria leaf spot severity, herewith on efficiency. The efficiency of Cabrio Duo did not differ from the standard fungicide Signum, efficiency following 21 days after the last (third) application was 88.07 % on average. Significant differences between trial data compared with standard fungicide Signum are absent (Table 1).

63
Table 1. Efficiency of fungicide to control Alternaria leaf spot in ‘Mentor’ H and ‘Jaguar’ H cabbages1 lentelė. Fungicido efektyvumas kontroliuojant alternariozės plitimą ir intensyvumą ‘Mentor’ H ir ‘Jaguar’ H veislių kopūstų lauke
Babtai, 2010–2011
Treatments Variantas
Alternaria leaf spotAlternariozė (%) Efficiency
Efektyvumas (%)incidencepaplitimas
severityintensyvumas
‘Mentor’ HControlKontrolė
86.25 b 28.0 b –
Cabrio Duo 1.25 l ha-1 23.13 a 4.63 a 83.46 aCabrio Duo 2.5 l ha-1 20.63 a 4.13 a 85.25 aSignum 1.0 kg ha-1 11.88 a 2.88 a 89.71 a
‘Jaguar’ HControlKontrolė
79.38 b 20.25 b –
Cabrio Duo 1.25 l ha-1 13.13 a 2.63 a 87.01 aCabrio Duo 2.5 l ha-1 11.88 a 2.38 a 88.25 aSignum 1.0 kg ha-1 13.75 a 2.75 a 86.42 a* Means in columns followed by the same letter are not different significantly (P = 0.05) Duncan’s multiple range test / * Stulpeliuose pateiktos reikšmės, pažymėtos tomis pačiomis raidėmis, pagal Dunkano kriterijų (P = 0,05) iš esmės nesiskiria.
The studies showed that ‘Mentor’ H variety of cabbage was more damaged by Botrytis infection than ‘Jaguar’ H. Both rates of Cabrio Duo had no significant im-pact on the spreading of grey mould disease. The biological efficiency of Cabrio Duo was low – still 21.37 % at the rate of 1.25 l ha-1 and 42.86 % at the rate of 2.5 l ha-1 in ‘Mentor’ H variety, and 75.0 % and 37.4 % in ‘Jaguar’ H respectively (Table 2). The same can be said about the impact of Signum. Better effects against grey mould were observed in Signum: in ‘Mentor’ H variety it was 71.43 %, but only 25.0 % in ‘Jaguar’ H (Table 2).
The difficult situation of Botrytis infection control could be explained by Botrytis fungus resistance. The control of Botrytis diseases is difficult to obtain. Sprays with a number of broad-spectrum or systemic fungicides control Botrytis sp. fungi on a wide variety of crops. However, the control of Botrytis diseases in the field through chemical sprays was only partially successful. The application of fungicides just before or after harvest is desirable but this practice is restricted by the toxicological risks of their residues. Botrytis strains resistant to several systemic and even to some broad-spectrum fungicides were found in various crops (Agrios, 2005; Leroux, 2007).

64
Table 2. Efficiency of fungicide to control diseases in ‘Mentor’ H and ‘Jaguar’ H cabbages2 lentelė. Fungicido efektyvumas kontroliuojant ligų plitimą ‘Mentor’ H ir ‘Jaguar’ H veislių kopūstų lauke
Babtai, 2010–2011
Treatments Variantas
Grey mould Kekerinis puvinys
(%)
Efficiency Efektyvu-
mas (%)
Sclerotinia rot
Sklerotinis puvinys (%)
Efficiency Efektyvu-
mas (%)
Ring spot Žiedinė
dėmėtligė (%)
Efficiency Efektyvu-
mas (%)
‘Mentor’ HControl Kontrolė
8.75 b –
not established nenustatyta
0.25 b –
Cabrio Duo 1.25 l ha-1 6.88 ab 21.37 0.13 ab 48.0Cabrio Duo 2.5 l ha-1 5.0 ab 42.86 0 ab 100Signum 1.0 kg ha-1 2.5 a 71.43 0 ab 100
‘Jaguar’ HControl Kontrolė 5.0 b – 2.5 ab –
not established nenustatyta
Cabrio Duo 1.25 l ha-1 1.25 a 75.0 3.13 ab 0Cabrio Duo 2.5 l ha-1 3.13 ab 37.4 4.38 ab 0Signum 1.0 kg ha-1 3.75 ab 25.0 1.25 ab 50.0* Means in columns followed by the same letter are not different significantly (P = 0.05) Duncan’s multiple range test / * Stulpeliuose pateiktos reikšmės, pažymėtos tomis pačiomis raidėmis, pagal Dunkano kriterijų (P = 0,05) iš esmės nesiskiria.
The impact of fungicide Cabrio Duo and Signum on Sclerotinia rot was con-troversial, as well as in Botrytis case. In one case efficiency was up to 50 % in the other – zero, as in ‘Jaguar’ H cultivar. Therefore, the conclusions about the efficacy of fungicides are complicated; we did not receive any significant difference between the results. It should be noted that it was difficult to estimate the prevalence of diseases before the appearance of characteristic features. It was influenced by the occurrence of bacterial disease.
Ring spot (M. brassicicola) was not a widely spread disease (Fig. 1.). The infection established only in 2010 on ‘Mentor’ H varieties of cabbage (Table 2). Disease severity ranged from 0.25 % in untreated plots to 0.13 % in treatment plots with Cabrio Duo at the 1.25 l ha-1 application rate. However, significant differences between trial data compared with standard fungicide Signum were absent.
Our experimental findings suggest that all fungicides applied at the stage of development of harvestable vegetative cabbage parts (growth stage BBCH 41–46) significantly suppressed Alternaria black rot in stored cabbage. The mean biological efficacy of the fungicides ranged from 66.67 % to 100 % and it corresponded to good and excellent effect (Table 3).

65
Table 3. Efficiency of fungicides on disease incidence on ‘Mentor’ H and ‘Jaguar’ H cabbages after 4 months storage3 lentelė. Fungicidų efektyvumas ‘Mentor’ H ir ‘Jaguar’ H veislių kopūstus palaikius sandėlyje 4 mėnesius
Babtai, 2010–2011
Treatment Variantas ALT (%)
Efficiency Efektyvumas
(%)BTR (%)
Efficiency Efektyvumas
(%)BAC (%)
Efficiency Efektyvumas
(%)‘Mentor’ H
Control Kontrolė
37.5 b – 62.5 b –
not establishednenustatyta
Cabrio Duo 1.25 l ha-1 0 ab 100 a 50.0 ab 20.0 aCabrio Duo 2.5 l ha-1 0 ab 100 a 12.5 ab 80.0 aSignum 1.0 kg ha-1 12.5 ab 66.67 a 37.5 ab 40.0 a
‘Jaguar’ HControl Kontrolė
12.5 b – 25 ab – 37.5 b –
Cabrio Duo 1.25 l ha-1 0 ab 100 a 25 b 0 a 12.5 ab 66.7 aCabrio Duo 2.5 l ha-1 0 ab 100 a 0 ab 100 b 25 ab 33.3 aSignum 1.0 kg ha-1 0 ab 100 a 12.5 ab 50 b 25 ab 33.3 a* Means in columns followed by the same letter are not different significantly (P = 0.05) Duncan’s multiple range test / * Stulpeliuose pateiktos reikšmės, pažymėtos tomis pačiomis raidėmis, pagal Dunkano kriterijų (P = 0,05) iš esmės nesiskiria.ALT – alternaria leaf spot / alternariozė; BTR – grey mould / kekerinis puvinys; BAC – bacterial rot / bakterinis puvinys.
According to the test data ‘Mentor’ H variety cabbage was mostly affected by grey mould after four months storage compared with ‘Jaguar’ H; disease incidence was 62.5 % in untreated cabbage, it was 50.0 % in cabbage treated with Cabrio Duo at the rate of 1.25 l ha-1 – and 12.5 % at the rate of 2.5 l ha-1; it was 37.5 % – treated with Signum at the rate of 1.0 kg ha-1 respectively (Table 3). Significant differences between trial data were absent.
However, ‘Jaguar’ H cabbage was more susceptible to bacterial infection (Table 3). Soft-rot bacterium, Erwinia carotovora subsp. carotovora (syn. Pectobacterium caro-tovorum subsp. carotovorum) enters through growth cracks or wounds caused by cold temperatures, insects, other disease organisms, or by mechanical means. Under warm and humid conditions uninjured tissue may become infected through natural openings. Prolonged moisture from rain or irrigation and mild temperatures encourage disease development (Agrios, 2005).
According to the test data application with fungicides Cabrio Duo and Signum had a small but positive effect on cabbage yield, the increase of cabbage head marketable yield increased up to 1.6 %–6.38 %. Cabrio Duo as well as Signum had no phytotoxic

66
effect on cabbage plants. Significant differences between trial data compared with standard fungicide Signum are absent. Yield increase was significantly higher compared with the untreated cabbage only in ‘Mentor’ H (Table 4).
Table 4. Impact of fungicides on yield of ‘Mentor’ H and ‘Jaguar’ H cabbages 4 lentelė. Fungicidų poveikis ‘Mentor’ H ir ‘Jaguar’ H veislių kopūstų derlingumui
Babtai, 2010–2011
Treatments Variantas
Marketable yield (tones ha-1) Prekinis derlius, t ha-1
Relative number Santykiniais skaičiais (%)
‘Mentor’ HControlKontrolė
136.0 a 100
Cabrio Duo 1.25 l ha-1 142.25 bc 104.6Cabrio Duo 2.5 l ha-1 143.82 bc 105.75Signum 1.0 kg ha-1 144.67 c 106.38
‘Jaguar’ HControlKontrolė
137.2 ab 100
Cabrio Duo 1.25 l ha-1 139.9 ab 101.97Cabrio Duo 2.5 l ha-1 139.5 ab 101.68Signum 1.0 kg ha-1 139.4 ab 101.6* Means in columns followed by the same letter are not different significantly (P = 0.05) Duncan’s multiple range test / * Stulpeliuose pateiktos reikšmės, pažymėtos tomis pačiomis raidėmis, pagal Dunkano kriterijų (P = 0,05) iš esmės nesiskiria.
A good effect of strobilurin-containing fungicides, such as Signum and Cabrio Duo, was obtained. Strobilurin fungicides have become a very valuable tool for managing diseases. Strobilurin groups of fungicides are broad-spectrum fungicides active against a wide range of diseases in many crops. They are excellent inhibitors of spore germination and known for their protectant activity (Ganeshan, Chethana, 2009). They are effective against several different plant pathogenic fungi (Shokes, Snajder, 1999; Šidlauskienė, 2001; Bartlett et al., 2002). For all parameters evaluated, strobilurin-containing fungicides gave similar or even better results in comparison with the fungicides containing mancozeb, mancozeb + metalaxyl, benalaxyl, triazole, iprodione to control diseases such as Phytophthora porri in leek, Erysiphe heraclei and Alternaria dauci in carrot, Mycosphaerella spp., Albugo candida and Alternaria spp. in Brussels sprouts, Botrytis cinerea and Sclerotinia sclerotiorum in outdoor lettuce (Callens et al., 2005; Robak, Adamicki, 2007).
Although there is insufficient research data about the effects of fungicide Cabrio Duo, but some reports of its use in horticulture are positive. New formulation dimet-homorph + pyraclostrobin (Cabrio Duo) showed an efficacy similar to that provided by the best chemical references, metalaxyl-m + copper oxychloride and dimetomorph +

67
copper respectively (Gengotti et al., 2010). Cabrio Duo is used in preventive and curative way to control downy mildew (Peronospora destructor) in garlic and onion; downy mildew (Pseudoperonospora cubensis) and powdery mildew (Erysiphe cicho-racearum) in Cucurbits; mildew (Bremia lactucae) in lettuce; late blight (Phytophthora infestans) and early blight (Alternaria sp.) in potatoes and tomatoes (Tarlazzi et al., 2008; Mazzini, 2009; Ganeshan, Chethana, 2009; Gengotti et al., 2009; Ihsan et al., 2013; Cabrio Duo EC, 2014). Experimental trials were carried out in order to evaluate the efficacy of dimethomorph + pyraclostrobin, as well as heat air when applied to basil seeds naturally infected by Peronospora belbahrii, the causal agent of downy mildew of basil, a pathogen which is seed transmitted (Gilardi et al., 2015). Capriotti et al. re-ported the use of Cabriotop, a combination product of pyraclostrobin + metiram, which was effective for both early blight and late blight on grapevine and tomato (Capriotti et al., 2005). The most effective fungicides against blueberry ripe rot caused by fungus Colletotrichum sp. in blueberry (Vaccinium corymbosum L.) were Signum and Cabrio Duo, both containing an active ingredient pyraclostrobin (Volkova et al., 2011).
Registered investigation of Cabrio Duo is now continued at the Institute of Horticulture, LRCAF; especially since their toxicological properties and environ-mental impact make them suitable to be introduced in the list of fungicides allowed in the Integrated Production Guidelines for the disease control (Gengotti et al., 2010).
Conclusions. The cabbage varieties tested on the resistance to diseases were relatively unequal. Cabbage cultivar ‘Mentor’ H was damaged by diseases more than ‘Jaguar’ H and its protection against diseases must be given more attention. ‘Jaguar’ H cabbage variety is susceptible to bacterial rot, so its harvesting and storage must meet all technological requirements.
Alternaria leaf spot has been the main and most widespread disease on cabbage. Ring spot is not a widespread disease in Lithuania at this moment, but under changing climate conditions it may become relevant. Grey and sclerotinia rots in many cases are a consequence of pest spreading and control of these diseases is inseparable from pest control.
The efficiency of Cabrio Duo at the rates of 1.25 l ha-1 and 2.5 l ha-1 to control disease did not differ compared with the standard fungicide Signum at the rate of 1.0 kg ha-1. Different rates of Cabrio Duo had no significant effect on most studied diseases during vegetation and four-month storage period. Cabrio Duo has consistent characteristics to control diseases, as well Signum; it can be successfully used in plant protection systems.
Acknowledgement. This work was carried out within the framework of the long-term research program “Horticulture: agro-biological basics and technologies” implemented by the Lithuanian Research Centre for Agriculture and Forestry.
Gauta 2015 11 23 Parengta spausdinti 2015 12 09

68
References
1. Adamicki F., Robak J. 2000. Effect of fungicide treatments before harvest on the incidence of grey mould caused by Botrytis cinerea Per., quality and storage ability of white cabbage. Vegetable Crops Research Bulletin, 53: 45–53.
2. Agblor Sh., Waterer D. 2001. Cabbage – post-harvest handling and storage. http://www.4.agr.ge.ca/
3. Agrios G. N. 2005. Plant Pathology. Elsevier-Academic Press, San Diego, CA.4. Azevedo S. S., Mariano R de L. R., Michereff S. J. 2000. Survey of the intensity
of black rot and Alternaria leaf spot of cabbage in Agreste of Pernambuco and de-termination of sample size for disease quantification. Summa Phytopathologica, 26(3): 299–306.
5. Bartlett D. W., Clough J. M., Godwin J. R., Hall A. A., Hamer M., Parr-Dobrzanski B. 2002. The strobilurin fungicides. Pest management science, 58(7): 649–662.
6. Bedlan G. 1998. Apllications-Management zur Bekampfung der Botrytis-Lagerfau Chinakhl. Gesunde Pflanzen, 50(6): 162–164.
7. Bedlan G. 2001. Krankheiten und Nichtparasitäre Schädligungen des Chinakohls während der Lagerung und deren Vermeidung. Pflanzenschutz, 17(4): 1–17.
8. Cabrio Duo EC. 2014. http://www.terralia.com/vademecum_de_productos_fitosanitarios_y_nutricionales/index.php?proceso=registro&numero=9933&id_marca=22077&base=2014/
9. Callens D., Sarrazyn R., Evens W. 2005. Signum, a new fungicide for control of leaf diseases in outdoor vegetables. Proceedings of the 57th International sympo-sium on crop protection. Gent, Belgium, 70(3): 199–207.
10. Capriotti M., Marchi A., Coatti M., Manaresi M. 2005. Cabriotop: the broad spectrum fungicide for the control of the main grapevine and tomato diseases. Informatore Fitopatologico, 55: 38–45.
11. Datta A., Gopal M. 1999. Safety evaluation of the fungicide iprodione on cau-liflower. Bulletin of Environmental Contamination and Toxicology, 62(4): 496–501.
12. Ganeshan G., Chethana B. S. 2009. Bioefficacy of Pyraclostrobin 25% EC against early blight of tomato. World Appl. Sci. J., 7(2): 227–229.
13. Geeson J. D. 1983. Brassica. In: C. Dennis (eds.), Postharvest Pathology of Fruits and Vegetables. Acad. Press, 125–156.
14. Gengotti S., Sbrighi C., Antoniacci L., Bugiani R. 2010. Efficacy evaluation of different fungicides for the control of potato late blight in Italy. PPO-Special Report No. 14 of 12th EuroBlight workshop. 3–6 May, Arras, France, 287–288.
15. Gengotti S., Sbrighi C., Antoniacci L. 2009. New fungicides to control downy mildew on melon. Informatore-Agrario, 65(21): 50–52.
16. Gilardi G., Pintore I., Demarchi S., Gullino M. L., Garibaldi A. 2015. Seed dres-sing to control downy mildew of basil. Phytoparasitica, 43: 531–539.
17. Ihsan J., Mohsan M., Ali Q., Mohyudin G. 2013. Evaluation of different fungi-cides for the management of downy mildew (Pseudoperonospora cubensis) of muskmelon. Pakistan Journal of Phytopathology, 25(02): 123–126.

69
18. Leroux P. 2007. Chemical control of Botrytis and its resistance to chemical fungicides. In: Y. Elad et al. (eds.), Botrytis: Biology, Pathology and Control. Springer, Netherlands, 195–222.
19. Mazzini F. 2009. Cabrio Duo: a new fungicide mixture for horticulture against Peronospora and Alternaria. Informatore Agrario Supplemento, 65(26): 14–15.
20. Robak J., Adamicki F. 2007. The effect of pre-harvest treatment with fungicide on the storage potential of root vegetables. Vegetable crops research bulletin, 67: 187–196.
21. Shokes F. M., Snajder K. 1999. Disease control on peanut with a strobilurin fungicide. Fungicide Nematocide Tests, 54: 414–415.
22. Survilienė E., Valiuškaitė A., Duchovskienė L., Kavaliauskaitė D. 2010. Influence of fungicide treatment on grey mould of cabbage. Vegetable crops research bulletin, 73: 133–142.
23. Šidlauskienė A. 2001. Effect of pesticides on prevalence of Alternaria leaf spot (Alternaria spp.) in white cabbage seed plants. Sodininkystė ir daržininkystė, 20(4): 79–85.
24. Tarakanovas P., Raudonius S. 2003. Agronominių tyrimų duomenų statistinė analizė, taikant kompiuterines programas ANOVA, STAT, SPLIT-PLOT iš pa-keto SELEKCIJA ir IRRISTAT. Akademija, Kėdainių r.
25. Tarlazzi, S., Marchi A., Manaresi M. 2008. Cabrio Duo: a new formulation of dimethomorph and pyraclostrobin effective against the late blight of tomato and downy mildew of vegetable crops. Giornate Fitopatologiche Cervia, 2: 149–153.
26. Volkova J., Baženova A., Rancāne R., Vilka L. 2011. Managment of Ripe Rot by fungicides in Commercial Blueberry Plantations. Ražas svētki „Vecauce – 2011“: LLU mācību un pētījumu saimniecībai Vecauce – 90, 70–73.
27. Žemės ūkio augalų kenkėjai, ligos ir jų apskaita. 2002. J. Šurkus, I. Gaurilčikienė (sudaryt.). Lietuvos žemdirbystės institutas, Akademija, Kėdainių r.
SODININKYSTĖ IR DARŽININKYSTĖ. MOKSLO DARBAI. 2015. 34(3–4).
Galimybė kontroliuoti baltagūžių kopūstų ligas su Cabrio Duo
E. Survilienė, N. Rasiukevičiūtė, L. Duchovskienė
Santrauka
Tyrimų tikslas – nustatyti fungicido Cabrio Duo (v. m. piraklostrobinas 40 g l-1 + dime-tomorfas 72 g l-1) efektyvumą stabdant alternariozės, kekerinio ir sklerotinio puvinių, žiedinės dėmėtligės ir bakterinio puvinio plitimą baltagūžių kopūstų lauke vegetacijos metu ir laikant kopūstus sandėlyje ir palyginti su standartinio fungicido Signum (v. m. boskalidas 267 g ha-1 + piraklostrobinas 67 g ha-1) efektyvumu. Tyrimai atlikti Lietuvos agrarinių ir miškų mokslų centro filiale Sodininkystės ir daržininkystės institute 2010–2011 m. ‘Mentor’ H ir ‘Jaguar’ H veislių kopūstai buvo nupurkšti 3 kartus fungicidais Cabrio Duo (normos – 1,25 ir 2,5 l ha-1) ir Signum (norma – 1,0 kg ha-1) antroje vegetacijos pusėje (augimo tarpsnis BBCH 41–46). Nuėmus derlių, kopūstų ėminiai buvo laikyti sandėlyje 4 mėnesius. Gauti rezultatai rodo,

70
kad purškimas fungicidais leido kontroliuoti ligų plitimą tiek vegetacijos metu, tiek laikant kopūstus sandėlyje. Fungicido Cabrio Duo efektyvumas, kai jo normos buvo 1,25 ir 2,5 l ha-1, nesiskyrė nuo standartinio fungicido Signum (norma – 1,0 kg ha-1) efektyvumo, todėl jis gali būti naudojamas kopūstų apsaugos nuo ligų sistemoje.
Reikšminiai žodžiai: alternariozė, bakterinis puvinys, efektyvumas, fungicidai, kekerinis puvinys, kopūstas, sklerotinis puvinys, žiedinė dėmėtligė.

71
LIETUVOS AGRARINIŲ IR MIŠKŲ MOKSLŲ CENTRO FILIALO SODININKYSTĖS IR DARŽININKYSTĖS INSTITUTO IR ALEKSANDRO STULGINSKIO UNIVERSITETO MOKSLO DARBAI. SODININKYSTĖ IR DARŽININKYSTĖ. 2015. 34(3–4).
Alternatyvių augalų apsaugos priemonių įtaka braškių augumui ir produktyvumui bei kekerinio puvinio plitimui
Neringa Rasiukevičiūtė1, Alma Valiuškaitė1, Nobertas Uselis1, Živilė Lukšienė2
1 Lietuvos agrarinių ir miškų mokslų centro filialas Sodininkystės ir daržininkystės institutas, Kauno g. 30, LT-54333 Babtai, Kauno r., el. paštas [email protected]
2 Vilniaus universiteto Taikomųjų mokslų institutas, Saulėtekio g. 10, LT-10223 Vilnius
Tyrimai atlikti Lietuvos agrarinių ir miškų mokslų centro (LAMMC) filiale Sodininkystės ir daržininkystės institute 2012–2015 m., siekiant ištirti skirtingų alternatyvių augalų apsaugos priemonių – chlorofilino darinių – poveikį kekerinio puvinio plitimui braškyne. Palyginimui buvo pasirinkti įprastiniai fungicidai: Signum ir Switch 62,5 WG, naudojami braškių apsaugai nuo kekerinio puvinio. Atliekant tyrimą naudoti chlorofilino dariniai turėjo teigiamą poveikį uogų derliui ir kokybei, nors ir neprilygo tradiciniams fungicidams, davusiems stabilų vidutinį 5,51 t ha-1 derliaus priedą. Naudotos alternatyvios augalų apsaugos priemonės, nors ir ne iš esmės, gerino braškių kerelių augumą ir išsivystymą. Apdorojus braškes chlorofilino dariniu Na-Chl-a, kekerinio puvinio pažeistų uogų visais tyrimų metais buvo mažiau, palyginti su neapdorotomis.
Reikšminiai žodžiai: Botrytis cinerea, braškės, chlorofilino dariniai, kerelių augumas, kerelių išsivystymas, Signum, Switch 62.5 WG.
Įvadas. Europos Sąjungos (ES) direktyvose yra nustatyti reikalavimai sugriež-tinti ir sumažinti pesticidų naudojimą, todėl imta intensyviai ieškoti naujų, efekty-vesnių neterminių antibakterinių maisto technologijų, padedančių veiksmingai pa-šalinti ligas sukeliančius mikroorganizmus nesumažinant maisto mitybinės vertės. Fizinio poveikio maisto saugos priemonės yra naujos, perspektyvios ir intensyviai plėtojamos technologijos, turinčios stiprų antibakterinį poveikį (Lukšienė, 2005; Lukšienė, Paškevičiūtė, 2011; Lukšienė, 2014).
Žinoma, kad daugelis patogeninių mikroorganizmų gali tapti atsparūs cheminiams pesticidams ir dezinfektantams (Yuk ir kt., 2006). Pasaulyje ieškoma alternatyvių augalų apsaugos produktų ne vien siekiant įgyvendinti ES direktyvą, bet ir dėl to,

72
kad perteklinis pesticidų naudojimas gali sukelti toksinį poveikį aplinkai, augalams, gyvūnams ir ypač žmonėms. Alternatyviai augalų apsaugai naudojami įvairūs eteriniai aliejai, turintys įvairių antimikrobinių savybių, bet nenuodingi augalams (Yuk ir kt., 2006; Lukšienė, 2014; Hesami ir kt., 2014). Hesami ir kt. (2014) in vitro tyrimais nustatė, kad pistacijų eterinis aliejus, kaip alternatyvi braškių apsaugos priemonė, 10 proc. sumažino Botrytis cinerea plitimą. Kaip alternatyvi augalų apsaugos priemonė naudojamos chitozano plėvelės: jomis padengus uogas, šios apsaugomos nuo puvinių. Chitozano plėvelių pagrindą sudaro gamtinis aminopolisacharidinis polimeras, turintis ir antimikrobinį poveikį (Moussa ir kt., 2013).
Viena iš alternatyvių augalų apsaugos priemonių yra fotosensibilizacija. Ji paremta dviejų netoksiškų veiksnių – matomos šviesos ir fotojautrios medžiagos – sąveika de-guonies aplinkoje. Metodas patrauklus tuo, kad šviesos spinduliai veikia tik paviršinį maisto medžiagos sluoksnį ir nekeičia jo juslinių savybių, bet gana veiksmingai nai-kina patogeninius mikroorganizmus, yra nenuodingi aplinkai ir nesukelia atsparumo. Fotosensibilizacinės medžiagos greitai išblunka, veikiamos tiek saulės, tiek dirbtinio šviesos šaltinio šviesos. Viena iš fotosensibilizacijai atlikti naudojamų medžiagų – chlo-rofilinas – yra vandenyje tirpus maisto priedas (E141), naudojamas maisto pramonėje kaip dažas, taip pat gaminant papildus bei kosmetikos priemones (Lukšienė ir kt., 2005; Lukšienė, Žukauskas, 2009; Lukšienė ir kt., 2010; Lukšienė, 2014).
Esminius desertinių uogų nuostolius sukelia uogų užterštumas patogeniniais mikroorganizmais. Viena pagrindinių braškių uogų nuostolius sukeliančių ligų yra kekerinis puvinys (Botrytis cinerea). Dėl augalų ligų patiriami ne tik derliaus nuostoliai, bet ir suprastėja augalų būklė, augalai prasčiau dera ir žiemoja (Uselis ir kt., 2006; Valiuškaitė ir kt., 2008; Valiuškaitė ir kt., 2010; Rasiukevičiūtė ir kt., 2013). Labai svarbu surasti priemones, padedančias sumažinti uogų užterštumą Botrytis cinerea, išauginti sveiką ir saugų maisto produktą ir pailginti uogų vartojimo trukmę.
Darbo tikslas – nustatyti chlorofilino darinių poveikį desertinių braškių augumo ir produktyvumo rodikliams bei kekerinio puvinio plitimui.
Tyrimo objektas, metodai ir sąlygos. Tyrimai atlikti Lietuvos agrarinių ir miš-kų mokslų centro filiale Sodininkystės ir daržininkystės institute 2012–2015 metais. Braškynas įveistas 2012 m. rugpjūčio mėnesį sodinant ‘Darselect’ veislės daigus. Braškės augintos žemose, juoda plėvele mulčiuotose, lašeliniu būdu lietinamose trieilėse lysvėse. Laukelio plotas – 3,2 m2. Laukeliai išdėstyti atsitiktine tvarka pa-kartojimų blokais. Variantai kartoti keturis kartus.
Siekiant ištirti skirtingų chlorofilino darinių – Na-Chl a, b, c (Na-Chl 1.5 × 10-3 M) (Lukšienė, Buchovec, 2009), kaip alternatyvios augalų apsaugos priemonės (AAAP), skirtingų koncentracijų poveikį, purkšta keturis kartus braškėms žydint (61–65 augimo tarpsnis pagal BBCH skalę) (BBCH Monograph, 2001). Palyginimui buvo pasirinkti įprastiniai fungicidai: Signum ir Switch 62,5 WG, naudojami braškių apsaugai nuo kekerinio puvinio. Šiais fungicidais buvo purškiama po du kartus braškėms žydint. 2013–2015 m. bandymo schema pateikiama 1 lentelėje.
Uogų užsikrėtimas puviniais nustatomas masinio braškių derėjimo metu, vizualiai vertinant uogas, atskirai sveriant sveikas bei supuvusias uogas, apskaičiuojant užsikrė-tusių uogų procentą (Žemės ūkio augalų kenkėjai, ligos ir jų apskaita, 2002). Nustatytas sveikų ir supuvusių uogų derlius (t ha-1). Braškių kerelių augumo, išsivystymo ir

73
produktyvumo rodikliai įvertinti po braškių derėjimo: 2013-07-03, 2014-07-22 ir 2015-06-09. Braškių kerelių augumas ir išsivystymas nustatytas suskaičiuojant vidutinį augalo ragelių, lapų, žiedynų ir ūsų skaičių.
2013–2014 m. žiema buvo labai šalta: vidutiniškai per 10 dienų oro temperatūra nukrisdavo iki –30 °C. Per tokį speigą tiriamąsias braškes dengė tik plona sniego danga. Meteorologiniai duomenys fiksuoti iMETOS®sm (Pessl Instruments, Austrija) meteorologine stotele.
1 lentelė. Tyrimams naudotos augalų apsaugos priemonėsTable 1. Experimental design of plant protection products, LRCAF IH, 2013–2015
ProduktasProduct
Veiklioji medžiaga Active ingredient
NormaRate
Tyrimo metaiYear of investigation
FungicidaiFungicides
Signum Boskalidas 267 g kg-1 + piraklostrobinas 37 g kg-1
3,17 g / 3 l
2013–2015Switch 62,5 WG Ciprodinilas 375 g kg-1 + fludioksonilas 250 g kg-1
1,76 g / 3 l
Alternatyvi augalų apsaugos priemonėAlternative plant protection measures
Na-Chl-a Chlorofilino darinys 1,5 × 10-3 MChlorophyllin derivatives
3 g / 3 l 2013
Na-Chl-b Chlorofilino darinys 1,5 × 10-3 MChlorophyllin derivatives
0,3 g / 3 l 2014–2015
Na-Chl-c Chlorofilino darinys 1,5 × 10-3 MChlorophyllin derivatives
3 g / 3 l 2015
Tyrimo duomenys apdoroti dispersinės analizės metodu pagal Dunkano kriterijų (P = 0,05), naudojant kompiuterio programą ANOVA, nustatytas mažiausias esminis skirtumas (R05) (Tarakanovas, Raudonius, 2003).
Rezultatai ir aptarimas. Pasaulyje yra kuriamos ir tobulinamos mikrobiolo-ginės braškių užkrėstumo mažinimo technologijos, galinčios padėti ne tik pailginti uogų vartojimo trukmę, bet ir sunaikinti patogeninius mikroorganizmus (Lukšienė, 2014).
Nustatyta, kad apdorojus braškes chlorofilino dariniu Na-Chl-a kekeriniu puviniu sergančių uogų visais tyrimų metais buvo mažiau negu kontrolės variante. 2013 m. apdorojus braškes Na-Chl-a gautas vidutiniškai 0,58 t ha-1 didesnis derlius, palyginti su kontrolės variantu (1 pav.). Atitinkamai kekerinio puvinio pažeistų uogų buvo 0,61 proc. mažiau nei kontrolės variante. 2015 m. purškimas Na-Chl-c 4,11 proc. sumažino kekerinio puvinio pažeistų uogų skaičių, palyginti su kontrolės variantu. 2014 m. derlius buvo mažas dėl šaltos žiemos ir pavasario šalnų: dalis žiedų nušalo, kai kur kereliai iššalo. AAAP turėjo teigiamą poveikį uogų derliui ir kokybei, nors ir neprilygo tradiciniams fungicidams: purškiant pastaraisiais gautas stabilus vidutinis

74
5,51 t ha-1 derliaus priedas (1 pav.).Literatūroje nurodoma, kad chlorofilinas sunaikina tokius patogenų, kaip Bacillus
cereus ir Listeria monocytogenes (Lukšiene, Paškevičiūtė, 2011; Lukšiene ir kt., 2013). Nustatyta, kad Listeria monocytogenes patogenų in vitro dėl fotosensibilizacijos po-veikio sumažėjo 98 proc. (Lukšiene, Paškevičiūtė, 2011). Moussa ir kt. (2013) nustatė, kad chitozano ir sidabro plėvelės efektyviai slopina Botrytis cinerea augimą in vitro.
1 pav. Alternatyvių augalų apsaugos priemonių įtaka uogų kekerinio puvinio plitimui
Fig. 1. Effect of alternative plant protection measures on grey mould, LRCAF IH, 2013–2015.
Labai svarbu palaikyti braškių kerelių būklę joms baigiant žydėti, prieš pat derliaus nuėmimą. Laiku netaikant efektyvių augalų apsaugos priemonių, braškių derėjimo pabaigoje augalus dažnai apninka ligos, kenkėjai, kerelių būklė suprastėja (Uselis ir kt., 2009).
Pirmą kartą atliekant lauko bandymus nustatyta, kad panaudojus alternatyvias augalų apsaugos priemones 2013 m. visuose bandymo variantuose braškių kerelių išsivystymas buvo panašus. Kai kuriuose laukeliuose, kur buvo panaudoti tradiciniai augalų apsaugos produktai ir AAAP, augančių kerelių ragelių, lapų, žiedynų ir ūsų

75
skaičius, tenkantis vienam augalui, varijavo neperžengdamas paklaidos ribų ir iš esmės nesiskyrė (2 pav.). Panaudotos priemonės turi įtakos braškių kerelių išsivystymui tik ateinančiais metais.
2 pav. Chlorofilino darinių ir fungicidų poveikis ‘Darselect’ veislės braškių kerelių augumui ir išsivystymui.
Fig. 2. Chlorophyllin derivative and fungicide measures on strawberry ‘Darselect’ plants growth and productivity, LRCAF IH, 2013–2015.
Antraisiais ir trečiaisiais tyrimų metais panaudotos AAAP pagerino (nors ir ne-esmingai) braškių kerelių augumą bei išsivystymą ir padidino braškių produktyvumą (2 pav.). Tik fungicidų Switch ir Signum derinys braškių išsivystymą ir produktyvumą pagerino iš esmės. Braškių kereliai suformavo iš esmės daugiausia žiedynų ir buvo gautas iš esmės didesnis derlius, palyginti su kontrolės variantu.
Trečiaisiais tyrimų metais AAAP pagerino (nors ir neesmingai) braškių kerelių augumą ir išsivystymą. Po nepalankių žiemojimo sąlygų tiek kontrolės, tiek ir kituose variantuose apie penktadalis braškių kerelių dėl stipraus pašalimo išnyko ir brašky-nas išretėjo. Tik tuose laukeliuose, kur braškės buvo apdorotos fungicidais Switch ir Signum, kereliai ne tik buvo daug geresnės būklės, bet ir jų išretėjo apie 3 kartus mažiau negu panaudojus kitas priemones ar be jų (2 lentelė).

76
2 lentelė. Alternatyvių augalų apsaugos priemonių įtaka kerelių būklei ir išretėjimui 2014 metaisTable 2. Alternative plant protection system influence on strawberry growth conditions, LRCAF IH, 2014.
AugalaiPlants
Būklė, išvaizda balaisState, appearance score (0–10)
IšretėjimasLoss, %
KontrolėControl
6,5 a 23,3 b
Na-Chl-b 6,8 a 22,6 bSwitch / Signum 9,2 b 7,7 a* tomis pačiomis raidėmis pažymėtos reikšmės pagal Dunkano kriterijų (P = 0,05) iš esmės nesiskiria / *Means followed by the same letters do not differ significantly (P = 0.05) according to Duncan’s multiple range test
Išvados. 1. Alternatyvios augalų apsaugos priemonės – chlorofilino dariniai – stabdo braškių uogų kekerinio puvinio plitimą, nors ir neprilygsta tradiciniams augalų apsaugos produktams. Apdorojus braškes Na-Chl-a gautas vidutiniškai 0,58 t ha-1 didesnis derlius.
2. Nustatyta, kad braškių uogoms augant (61–65 tarpsnis pagal BBCH) ir nokstant (85 tarpsnis pagal BBCH) lauko sąlygomis panaudotos alternatyvios priemonės – chlorofilino dariniai – gerino braškių kerelių augumo ir išsivystymo rodiklius, bet nepadidino braškių kerelių ištvermingumo ekstremaliomis žiemojimo sąlygomis.
Padėka. Tyrimai finansuojami Lietuvos mokslo tarybos, vykdant nacionalinę mokslo programa „Sveikas ir saugus maistas“ Nr. SVE-02/2012.
Gauta 2015 11 16 Parengta spausdinti 2015 12 09
Literatūra
1. Growth stages of mono- and dicotyledonous plants. 2001. In: U. Meier (ed.), BBCH Monograph. German Federal Biological Research Centre for Agriculture and Forestry.
2. Hesami G., Bahramian S., Fatemi A., Hesami S. 2014. Effect of Pistacia atlan-tica subsp. Kurdica Essential oil and Acetic acid on Botrytis cinerea Growth in Culture Media and Strawberry Fruits. Bulletin of Environment, Pharmacology and Life Sciences, 3(2): 100–106.
3. Yuk H., Bartz J., Schneider K. 2006. The effectiveness of sanitizer treatment in inactivation of Salmonella spp. from bell pepper, cucumber, and strawberry. Journal of Food Science, 71: 95–99.

77
4. Lukšienė Ž., Buchovec I. 2009. Decontamination of food and food-related sur-faces. Patent No 5567.
5. Lukšienė Ž., Buchovec I., Viškelis P. 2013. Impact of high-power pulsed light on microbial contamination, health promoting components and shelf life of straw-berries. Food Technology and Biotechnology, 51(2): 284–292.
6. Lukšienė Ž. 2005. New approach to inactivate harmful and pathogenic microor-ganisms: photosensitization. Food Technology and Biotechnology, 43: 411–418.
7. Lukšienė Ž. 2014. Novel Approach to Control Pathogenic and Harmful Microorganisms in Nonthermal Way: Photosensitization. In: J. Boziaris (ed.), Novel Food preservation and microbial assessment techniques, 184–217.
8. Lukšienė Ž., Paškevičiūtė E. 2011. Novel approach to the microbial deconta-mination of strawberries: chlorophyllin-based photosensitization. Journal of Applied Microbiology, 110: 1 274–1 283.
9. Lukšienė Ž., Pečulytė D., Jurkoniene S., Puras R. 2005. Inactivation of pos-sible fungal food contaminants by photosensitization. Food Technology and Biotechnology, 43: 335–341.
10. Lukšienė Ž., Žukauskas A. 2009. Prospects of photosensitization in control of pathogenic and harmful microorganisms. Journal of Applied Microbiology, 107: 1 415–1 424.
11. Moussa S. H., Tayel A. A., Alsohim A. S., Abdallah R. R. 2013. Botryticidal Activity of Nanosized Silver-Chitosan Composite and Its Application for the Control of Gray Mold in Strawberry. Journal of Food Science, 78(10): M1589–1594.
12. Rasiukevičiūtė N., Valiuškaitė A., Survilienė-Radzevičė E., Supronienė S. 2013. Investigation of Botrytis cinerea risk forecasting model in strawberries. Proceedings of Latvian Academy of Science, Section B. Natural, Exact and Applied Sciences, 67(2): 195–198.
13. Tarakanovas P., Raudonius S. 2003. Agronominių tyrimų duomenų statistinė ana-lizė taikant kompiuterines programas ANOVA, STAT, SPLIT-PLOT iš paketo SELEKCIJA ir IRRISTAT. Akademija, Kėdainių r.
14. Uselis N., Lanauskas J., Valiuškaitė A., Viškelis P. 2009. Braškių veislių tyrimas, auginant jas profiliuotoje dirvoje po priedanga. Sodininkystė ir daržininkystė, 28(4): 51–60.
15. Uselis N., Valiuškaitė A., Raudonis L. 2006. Incidence of fungal leaf diseases and phytophagous mites in different strawberry cultivars. Agronomy research, 4: 421–425.
16. Valiuškaitė A., Raudonis L., Survilienė E. 2008. Control of grey mould and white leaf spot in strawberry. Zemdirbyste-Agriculture, 95(3): 221–226.
17. Valiuškaitė A., Uselis N., Survilienė E. 2010. iMETOS®sm Botrytis sp. progno-zavimo modelio efektyvumo tyrimai braškyne. Sodininkystė ir daržininkystė, 29(3): 13–21.
18. Žemės ūkio augalų kenkėjai, ligos ir jų apskaita. 2002. J. Šurkus, I. Gaurilčikienė (sudaryt.). Lietuvos žemdirbystės institutas, Akademija, Kėdainių r.

78
SODININKYSTĖ IR DARŽININKYSTĖ. SCIENTIFIC ARTICLES. 2015. 34(3–4).
The effect of alternative plant protection measures on grey mould spread, strawberry growth and productivity
N. Rasiukevičiūtė, A. Valiuškaitė, N. Uselis, Ž. Lukšienė
Summary
The research was carried out at the Institute of Horticulture, LRCAF in 2012–2015 in order to investigate the effect of different chlorophyllin derivative concentrations on strawberry grey mould. For comparison conventional fungicides (Signum and Switch 62.5 WG) were chosen against grey mould. Alternative plant protection measures resulted in an average 5.51 t ha-1 yield increase and had a positive effect on fruit yield and quality, although they were not equal to traditional fungicides. The alternative plant protection measures had a tendency to improve the growth and productivity of strawberry. The number of grey mould infected strawberries were reduced after the treatment with chlorophyllin derivative Na-Chl-a during the experimental year.
Key words: Botrytis cinerea, chlorophyllin derivatives, development, growth, Signum, strawberry, Switch 62.5 WG.

79
ATMINTINĖ AUTORIAMS, RAŠANTIEMS
Į MOKSLO DARBUS „SODININKYSTĖ IR DARŽININKYSTĖ“
Straipsnio rankraščio pateikimo ir priėmimo procedūra
Straipsnius redakcijai gali pateikti bet kurie Lietuvos ar užsienio šalies mokslo darbuotojai bei asmenys, dirbantys mokslinį darbą. Ne mokslo darbuotojo straipsnis turi būti parašytas kartu su mokslo darbuotoju.
Rankraštis redakcijai siunčiamas paštu dviem egzemplioriais, atspausdintas kompiuteriu popieriuje, laikantis toliau tekste nurodytų reikalavimų. Pateiktas straipsnio rankraštis užregistruojamas ir perduodamas redaktorių kolegijos na-riui, kuruojančiam šią sritį. Jis įvertina, ar rankraščio turinys ir forma atitinka svarbiausius periodiniams straipsniams keliamus reikalavimus. Rankraščiai, kurie buvo atmesti pirmojo vertinimo metu, su aiškinamuoju raštu grąžinami autoriui. Jeigu straipsnio tinkamumas nekelia abejonių, redaktorių kolegijos narys skiria du recenzentus.
Pataisytą rankraštį autorius turi atsiųsti el. paštu arba paštu redakcijai kartu su elektronine laikmena (diskeliu arba kompaktiniu disku) per dešimt dienų.
Rankraščio reikalavimai
Struktūra ir apimtis
Rankraščio forma turi atitikti periodiniams moksliniams straipsniams keliamus reikalavimus. Teksto ir jo sudedamųjų dalių seka tokia:
– Straipsnio pavadinimas (ne daugiau kaip 10 žodžių)Pavadinimas rašomas mažosiomis raidėmis paryškintu šriftu (Augalų adap-
tacijos prie šalčio molekulinio mechanizmo aspektai).– Autoriaus vardas, pavardėVardas, pavardė – mažosiomis raidėmis paryškintu šriftu (Viktor Sharma).
Jeigu yra keli autoriai, rašoma mažėjančia jų autorystės indėlio tvarka.– Institucija, adresas, elektroninis paštasRašomi mažosiomis raidėmis kursyvu (Italic) (Lietuvos agrarinių ir miškų
mokslų centro filialas Sodininkystės ir daržininkystės institutas).
Pagrindinis tekstas
– Santrauka (iki 1 400 rašybos ženklų, arba 250 žodžių)Labai glaustai pateikiami tikslai, sąlygos, svarbiausi rezultatai.– Reikšminiai žodžiai (ne daugiau kaip 10)– ĮvadasTrumpai išdėstoma nagrinėjama problema, ankstesni kitų panašių tyrimų
rezultatai, darbo reikalingumas, originalumas. Nurodomas darbo tikslas.– Tyrimo objektas, metodai ir sąlygos

80
– Rezultatai ir aptarimasGali būti rašomi kartu arba atskirai. Išdėstomi ir aptariami tyrimų metu surinkti
duomenys, palyginami su kitų autorių duomenimis, aiškinamos tirtų reiškinių prie-žastys, keliamos naujos idėjos, hipotezės. Jei rezultatai aptariami atskiroje dalyje, tuomet „Aptarimo“ skyrelyje aptariami, bet ne kartojami „Rezultatų“ skyrelyje pateikti duomenys.
– Išvados– Padėka (neprivaloma)– LiteratūraRekomenduojama į sąrašą įtraukti ne mažiau kaip 10 naujausių literatūros
šaltinių rašoma tema.– Santrauka lietuvių ir anglų kalbomis (600–1 400 sp. ženklų)Straipsnių, kurie parašyti remiantis netradiciniais bandymų duomenimis ir jų
rezultatais, struktūrinės teksto dalys gali būti ir kitokios.Straipsnis turi būti ne daugiau kaip 10 puslapių apimties kompiuteriu rinkto
teksto, įskaitant lenteles ir paveikslus (didesnės apimties straipsniai derinami su redaktorių kolegijos pirmininku).
Teksto parengimas
Straipsnis rašomas lietuvių ir anglų kalbomis ir spausdinamas Microsoft WORD teksto redaktoriumi Windows 2000, XP ar VISTA operacinėse siste-mose. Tekstas rašomas TIMES NEW ROMAN 12 dydžio šriftu, A4 formato (210 × 297 mm) lape, atstumas tarp rankraščio eilučių – 1 (single), išlyginamas iš abiejų pusių. Paraščių plotis: viršuje – 2 cm, apačioje – 2 cm, dešinėje –1,5 cm, kairėje – 3 cm. Straipsnis pateikiamas elektronine laikmena.
Paryškintai (Bold) rašoma: straipsnio pavadinimas visomis kalbomis, antraštės bei svarbiausi struktūros elementai (įvadas, tyrimo objektas, metodai ir sąlygos, rezultatai, aptarimas, išvados, padėka, literatūra, santrauka). Kursyvu (Italic) rašomi lotyniškieji augalų rūšių, genčių, ligų, kenkėjų, mikroorganizmų ir kitų biologinių objektų pavadinimai. Augalų veislių pavadinimai rašomi viengubose kabutėse (pvz., ‘Auksis’).
Cituojamas šaltinis tekste nurodomas lenktiniuose skliaustuose (autoriaus pavardė, metai).
Lentelės
Lentelėse neturi būti kartojama paveiksluose ar kitose iliustracijose pateikta informacija.
Lentelių tekstas rašomas lietuvių ir anglų kalbomis TIMES NEW ROMAN 12 dydžio šriftu, jeigu parašyti tekstai talpinami vienoje eilutėje, tarp jų dedamas ženklas /. Lentelės teksto dalys vertikaliomis ir horizontaliomis linijomis neatski-riamos. Horizontaliomis linijomis atskiriamos tik lentelės metrikos dalys ir lentelės

81
pabaiga. Lentelės padėtis puslapyje tik vertikali (Portrait).Bandymų veiksnių gradacijos lentelės neturi būti žymimos skaičiais, sudėtin-
gomis santrumpomis, o pateikiamos visos arba suprantamai sutrumpinto aprašo formos. Pateikiama santrumpa turi būti paaiškinama pirmosios lentelės (paveikslo) apraše.
Statistiniai duomenys, skaičiai ir skaitmenys
Pageidautina detaliai aprašyti taikytus tyrimų metodus ir nurodyti jų origi-nalius šaltinius. Labai svarbi informacija apie lauko, vegetacinių ir kt. bandymų išdėstymo schemą ir jos pasirinkimo motyvus. Lentelėse ir paveiksluose pateikiami duomenys privalo būti statistiškai įvertinti. Rodiklių žymėjimo santrumpos turi būti paaiškintos, jeigu jos neatitinka tarptautinių ISO standartų.
Paveikslai
Visa iliustracinė medžiaga – brėžiniai, grafikai, diagramos, fotografijos, pie-šiniai ir kt. – vadinami bendru paveikslų vardu. Tekstas juose rašomas lietuvių ir anglų kalbomis.
Paveikslai turi būti nespalvoti, padaryti Microsoft Office 2000, 2003, XP, VISTA paketų elektroninėje lentelėje EXCEL su suderintomis programomis ir pradine informacija. Redakcija pasilieka teisę keisti jų formatą pagal straipsnio ar viso leidinio dizainą.
Įrašai ir simboliai paveiksluose turi būti parašyti ne mažesniu kaip 10 dy-džio TIMES NEW ROMAN šriftu. Paveikslų blokų dalys turi būti sužymėtos raidėmis a, b, c ir t. t.
Literatūra
Į literatūros sąrašą turi būti įtraukiamos tik mokslinės publikacijos.Citavimo pavyzdžiai:
Vieno autoriaus knyga:1. Brazauskienė M. D. 2004. Agroekologija ir chemija. Naujasis lankas, Kaunas.2. Blažek J. 2001. Pìstujeme jablonì. Nakladatelství Brázda, s. r. o., Praha.
Kelių autorių knyga:1. Kawecki Z., Łojko R., Pilarek B. 2007. Mało znane rośliny sadownicze.
Wydawnictwo UWM, Olsztyn.2. Žemės ir miškų ūkio augalų pesticidų katalogas. 2005. G. Rimavičienė (su-
daryt.). Lietuvos žemės ūkio konsultavimo tarnyba, Akademija, Kėdainių r.

82
Straipsnis knygoje:1. Streif J. 1996. Optimum harvest date for different apple cultivar in the
‘Bodensee’ area. In: A. de Jager, D. Johnson, E. Hohn (eds.), Determination and Prediction of Optimum Harvest Date of Apples and Pears. COST 94. European Commission. Luxembourg, 15–20.
2. Byme D. H., Sherman W. B., Bacon T. A. 2000. Stone fruit genetic pool and its exploitation for growing under warm winter conditions. In: A. Erez (ed.), Temperature Fruit Crops in Warm Climates. Kluwer Academic Publishers, Netherlands, 157–230.
3. Keller R. R. J. 2002. Cryopreservation of Allium sativum L. (Garlic). In: L. E. Towill, Y. P. S. Bajas (eds.), Biotechnology in Agriculture and Forestry. Springer-Verlag, Berlin, Heidelberg, 50: 38–47.
Straipsnis konferencijos medžiagoje:
1. Kampuss K., Strautina S. 2004. Evaluation of blackcurrant genetic resour-ces for sustainable production. Proceedings of the International Workshop on Protection of Genetic Resources of Pomological Plants and Selection of Genitors with Traits Valuable for Sustainable Fruit Production. 22–25 August, Skierniewice, Poland, 147–158.
Vieno autoriaus žurnalo straipsnis:
1. Korban S. S. 1986. Interspecific hybridization in Malus. Hort. Science, 21(1): 41–48.
Žurnalo straipsnis (du ir daugiau autorių):
1. Sasnauskas A., Gelvonauskienė D., Gelvonauskis B. 2002. Evaluation of new scab resistance apple in the first-fifth years in orchard. Sodininkystė ir daržininkystė, 21(3): 29–37.
2. Balan V., Oprea M., Drosu S., Chireceanu C., Tudor V., Petrisor C. 2006. Maintenance of biodiversity of apricot tree phenotypes in Romania. Acta Horticulturae, 701: 207–214.
Straipsnis e. žurnale:
1. Stanys V., Mažeikienė I., Stanienė G., Šikšnianas T. 2007. Effect of phy-tohormones and stratification on morphogenesis of Paeonia lactiflora Pall. isolated embryos. Biologija, 18(1). http://images.katalogas.lt/maleidykla/Bio71/Bio_027_030.pdf
Duomenų bazė:FAO Stat Database. 2005. http://faostat.fao.org/faostat/
Disertacijos santrauka:
1. Čižauskas A. 2003. Valgomųjų svogūnų (Allium cepa L.) auginimo iš sėklų technologijos elementų tyrimai: daktaro disert. santr. Babtai.

83
GUIDELINES FOR THE PREPARATION AND SUBMISSION OF ARTICLES TO THE VOLUMES OF SCIENTIFIC WORKS
“SODININKYSTĖ IR DARŽININKYSTĖ“
Rules for Submission – Acceptance of Papers
Papers can be contributed by any Lithuanian and foreign scientific workers and persons carrying out scientific research. The latter papers will be accepted only when the co-author is a scientific worker.
Manuscripts should be sent by mail typed on the computer printed in two co-pies on paper taking into account the following instructions. The manuscript will be registered and submitted to the member of the Editorial Board in charge. He (she) will evaluate, if the contents and the form corresponds to the main require-ments for periodical articles. Manuscripts rejected during the first evaluation will be returned to the author with explanatory remarks. If the article is approved the member of the Editorial Board appoints two reviewers.
The author must return the corrected manuscript to the Editorial Board in ten days by e-mail or by mail in a diskette or CD.
Requirements to the Manuscript
Structure and LengthThe form of a manuscript has to correspond to the requirements for periodical
scientific articles. The paper should be organized in the following order:– Title (should not exceed 10 words)Title should be written in small letters in bold (Aspects of plant color accli-
mation molecular mechanism).– Author(s)’ name, surnameThe name and surname should be written in small letters in bold (Viktor
Sharma). If there are more than one author they are listed according to their input to the paper.
– Institution(s), address, email addressShould be written in small letters in Italics (Institute of Horticulture Lithuanian
Research Centre for Agriculture and Forestry).
The Main Text
– Abstract (should not exceed 1 400 characters or 250 words)Should contain the statement of the aims, methods and main results in
short.– Key words (should not exceed 10 words)– IntroductionShould present the investigated subject, results of earlier related research,
reasons of the study, innovation. Should indicate the aim of the investigation.

84
– Object, methods and conditions– Results and DiscussionMay be written in separate sections or conjugated. Collected data should be
presented and discussed, interpret them with references to the results obtained by other authors, explain the reasons of the investigated phenomena and raise new ideas, hypotheses. If results are written in separate section, „Discussion“ should not repeat results presented in “Results”.
– Discussion Should not repeat results presented in “Results” but should interpret them
with reference to the results obtained by other authors, explain the reasons of the investigated phenomena and raise new ideas, hypotheses.
– Conclusions– Acknowledgements (optional) – ReferencesShould be kept to a minimum of 10 latest references on this theme.– Summary in Lithuanian and English (600–1 400 characters)Articles written based on non-traditional trial data and the obtained results
may have other than traditional structural parts of a paper.The article should not exceed 10 pages of computer text, tables and figures
included (longer articles are agreed with the Chairman of the Editorial Board).
Text Preparation
Manuscripts should be submitted in Lithuanian and English, typed on a PC, using Microsoft WORD for Windows 2000 XP or VISTA word-processor format. The font to be typed – TIMES NEW ROMAN size 12, on A4 paper (210 × 297 mm), single spaced, justified. Margins: top – 2 cm, bottom – 2 cm, right – 1.5 cm, left – 3 cm. An article should be submitted an electronic carrier.
In bold the title of the a paper in all languages, headings and all main structural elements (introduction, object, methods and conditions, results, discussion, conclusions, acknowledgements, references, abstract) are written in bold. In Italics. Latin names of species, genera, diseases, pests micro-organisms and other biological objects are written. Names of plant cultivars should be placed within single quotation marks (for example, ‘Auksis’).
The quoted reference in the text is indicated in round brackets (author’s sur-name, year).
Tables
Information given in figures or other illustrations, should not be repeated in tables.
The text in tables is written in the Lithuanian and English languages. If Lithuanian and English texts are in one line they are separated by /. Do not use vertical and horizontal lines to separate parts of the text. A horizontal line separa-tes only headings of columns and the end of the table. Orientation in a page only

85
vertical (Portrait).Trial variants in tables should not be numbered or submitted in complicated
abbreviations. Tables should be self explanatory, and if there are abbreviations, they should be understandable. The used abbreviation should be explained in the description of the first table (figure).
Statistical Data, Figures, Numerals
It is desirable to describe in detail the applied research methods and indi-cate their original references. The information on the scheme (design) of field, vegetative and other trials and motivation of their choice is very important. Data presented in tables and figures must be statistically processed. Abbreviations of parameters should be explained, if they do not correspond to the international standard abbreviations (ISO).
Figures
All illustrations – drawings, graphs, diagrams, photographs, pictures, etc. are considered as figures. The text in them is written in Lithuanian and English.
Figures must be drafted in black colour in Microsoft Office 2000, 2003, XP, VISTA package, EXCEL electronic table with conformed programs and initial information. The Editorial Board has the right to change their format according to the design of the article or the whole publication.
Letters and symbols in figures should not be smaller than size 10 TIMES NEW ROMAN. Block parts of figures should be numbered consecutively by letters a, b, c, etc.
References
Scientific publications may be included into references.Titles of foreign journals, volumes of conference articles, etc., are not abbre-
viated. The reference list should be arranged in alphabetical order.
Examples of Quotation:
Book of One Author:
1. Brazauskienė M. D. 2004. Agroekologija ir chemija. Naujasis lankas, Kaunas.
2. Blažek J. 2001. Pìstujeme jablonì. Nakladatelství Brázda, s. r. o., Praha.
Books of several authors:
1. Kawecki Z., Łojko R., Pilarek B. 2007. Mało znane rośliny sadownicze. Wydawnictwo UWM, Olsztyn.
2. Žemės ir miškų ūkio augalų pesticidų katalogas. 2005. G. Rimavičienė (su-daryt.). Lietuvos žemės ūkio konsultavimo tarnyba, Akademija, Kėdainių r.

86
Article in Book:
1. Streif J. 1996. Optimum harvest date for different apple cultivar in the ‘Bodensee’ area. In: A.de Jager, D. Johnson, E. Hohn (eds.), Determination and Prediction of Optimum Harvest Date of Apples and Pears. COST 94. European Commission. Luxembourg, 15–20.
2. Byme D. H., Sherman W. B., Bacon T. A. 2000. Stone fruit genetic pool and its exploitation for growing under warm winter conditions. In: A. Erez (ed.), Temperature Fruit Crops in Warm Climates. Kluwer Academic Publishers, Netherlands, 157–230.
3. Keller R. R. J. 2002. Cryopreservation of Allium sativum L. (Garlic). In: L. E. Towill, Y. P. S. Bajas (eds.), Biotechnology in Agriculture and Forestry. Springer-Verlag, Berlin, Heidelberg, 50: 38–47.
Article in Conference Material:
1. Kampuss K., Strautina S. 2004. Evaluation of blackcurrant genetic resour-ces for sustainable production. Proceedings of the International Workshop on Protection of Genetic Resources of Pomological Plants and Selection of Genitors with Traits Valuable for Sustainable Fruit Production. 22–25 August, Skierniewice, Poland, 147–158.
Article in Scientific Journal:
1. Korban S. S. 1986. Interspecific hybridization in Malus. HortScience, 21(1): 41–48.
2. Sasnauskas A., Gelvonauskienė D., Gelvonauskis B. 2002. Evaluation of new scab resistance apple in the first-fifth years in orchard. Sodininkystė ir daržininkystė, 21(3): 29–37.
3. Balan V., Oprea M., Drosu S., Chireceanu C., Tudor V., Petrisor C. 2006. Maintenance of biodiversity of apricot tree phenotypes in Romania. Acta Horticulturae, 701: 207–214.
Article in E- Journal:
1. Stanys V., Mažeikienė I., Stanienė G., Šikšnianas T. 2007. Effect of phy-tohormones and stratification on morphogenesis of Paeonia lactiflora Pall. isolated embryos. Biologija, 18(1). http://images.katalogas.lt/maleidykla/Bio71/Bio_027_030.pdf
Database:
FAO Stat Database. 2005. http://faostat.fao.org/faostat/
Thesis Abstract:
1. Čižauskas A. 2003. Valgomųjų svogūnų (Allium cepa L.) auginimo iš sėklų technologijos elementų tyrimai: daktaro disert. santr. Babtai.

Turinys – Contents
R. Rugienius, T. Šikšnianas, V. Bendokas, V. Stanys
Paprastosios žemuogės derėjimas bei antocianinų kiekis ir sudėtis uogosevandens deficito sąlygomis ........................................................................................3Cropping and anthocyanin content in berries of wild strawberry under water deficit stress ...............................................................................................................13
I. Januškaitienė
Dviejų vystymosi tarpsnių žirnių fotosintezės rodiklių atsakas į substrato rūgštumo irUV-B spinduliuotės poveikį ......................................................................................15The response of photosynthetic parameters The response of photosyntheticparameters of peas in two growth stages to substrate acidity and UV-B radiationeffect to substrate acidity and UV-B radiation effect ................................................28
R. Starkutė, O. Bundinienė, V. Zalatorius
Ekologinėmis sąlygomis augintų brokolinių kopūstų sodinimo laiko įtaka produktyvumui ..........................................................................................................29Influence of planting time of broccoli grown under organic conditions on productivity ...............................................................................................................37
R. Starkutė, O. Bundinienė, V. Zalatorius
Lapinių kopūstų, auginamų rudens derliui, produktyvumo tyrimai ........................38Investigations of productivity of kale grown for autumn harvest .............................45
N. Rasiukevičiūtė, R. Rugienius, J. B. Šikšnianienė
Daržinių braškių ligų sukėlėjai ir jų paplitimo Lietuvoje įvertinimas ......................47Estimation of Strawberry Pathogen Prevalence in Lithuania ..................................57
E. Survilienė, N. Rasiukevičiūtė, L. Duchovskienė
Possibility to control diseases of white cabbage with Cabrio Duo ...........................59Galimybė kontroliuoti baltagūžių kopūstų ligas su Cabrio Duo ...............................69
N. Rasiukevičiūtė, A. Valiuškaitė, N. Uselis, Ž. Lukšienė
Alternatyvių augalų apsaugos priemonių įtaka braškių augumui ir produktyvumui bei kekerinio puvinio plitimui ...................................................................................71The effect of alternative plant protection measures on grey mould spread, strawberrygrowth and productivity ............................................................................................78
Atmintinė autoriams, rašantiems į mokslo darbus „Sodininkystė ir daržininkystė“ ... 79Guidelines for the preparation and submission of articles to the volumes of scientificworks “Sodininkystė ir daržininkystė” ....................................................................83

ISSN 0236-4212
Mokslinis leidinysLietuvos agrarinių ir miškų mokslų centro filialoSodininkystės ir daržininkystės instituto ir Aleksandro Stulginskio universiteto mokslo darbai „Sodininkystė ir daržininkystė“. T. 34(3–4). 1–88.
Redagavo ir skaitė korektūrą Danguolė Vanagaitė, Saulius ZagorskisKompiuteriu maketavo Larisa Kazlauskienė
SL 1070. 2015 12 16 5,5 sp. l. Tiražas 50 egz.Išleido Lietuvos agrarinių ir miškų mokslų centro filialasSodininkystės ir daržininkystės institutas, LT-54333 Babtai, Kauno r.
Spausdino UAB „Mudu2“, Studentų g. 12-64,LT-50243 KaunasUžsakymo Nr. 9943