single molecular observation of the interaction of groel with substrate proteins
TRANSCRIPT

Article No. jmbi.1999.3129 available online at http://www.idealibrary.com on J. Mol. Biol. (1999) 292, 965±972
COMMUNICATION
Single Molecular Observation of the Interaction ofGroEL with Substrate Proteins
Ryo Yamasaki1, Masaru Hoshino1, Tetsuichi Wazawa3, Yoshiharu Ishii3
Toshio Yanagida2,3, Yasushi Kawata4, Takashi Higurashi4
Kazuko Sakai1,4, Jun Nagai4 and Yuji Goto1*
1Institute for Protein ResearchOsaka University, 3-2Yamadaoka, Suita, Osaka565-0871, Japan2Department of PhysiologyOsaka University MedicalSchool, 2-2 Yamadaoka, SuitaOsaka, 565-0871, Japan3Single Molecule ProcessesProject, ICORP, JST, 2-4-14Senba-Higashi, Mino, Osaka562-0035, Japan4Department of BiotechnologyFaculty of EngineeringTottori University, Tottori680-0945, Japan
E-mail address of the [email protected]
Abbreviations used: CD, circularb-lactoglobulin with the free thiol gCy5-red-blg, reduced and carboxymb-lactoglobulin with about one amiCy5; DTT, dithiothreitol; Gdn-HCl,hydrochloride; TMR, tetramethylrhb-lactoglobulin with the free thiol gTMR.
0022-2836/99/400965±8 $30.00/0
To understand the mechanism of GroEL-assisted protein folding, weobserved the interaction of ¯uorescence-labeled GroEL with ¯uorescence-labeled substrate proteins at the single molecule level by total internalre¯ection ¯uorescence microscopy. GroEL with a A133C mutation in theequatorial domain was labeled with a ¯uorescent dye, tetramethylrhoda-mine. As substrate proteins, we used the largely denatured and partlydenatured forms of bovine b-lactoglobulin, both labeled with another ¯u-orescent dye, Cy5. The complexes formed by GroEL with these substrateswere characterized by size-exclusion gel chromatography. The recoveredcomplexes were then observed by ¯uorescence microscopy. For both sub-strates, agreement of the ¯uorescent spots for tetramethylrhodamine andCy5 indicated formation of the complex at the single molecule level.Similar observation of macroscopic binding by size-exclusion chromatog-raphy and microscopic binding by the ¯uorescence microscopy was donefor the folding intermediate of Cy5-labeled bovine rhodanese. The ¯uor-escence microscopy opens a new avenue for studying the interaction ofGroEL with substrate proteins.
# 1999 Academic Press
Keywords: chaperonin; ¯uorescence microscopy; GroEL; b-lactoglobulin;single molecular detection
*Corresponding authorThe chaperonin GroEL from Escherichia coli isone of the most extensively studied molecular cha-perones (Fenton & Horwich, 1997; Coyle et al.,1997, 1999; Bukau & Horwich, 1998). It is com-posed of two heptameric rings of 57 kDa subunitsstacked back to back, and each subunit consists ofapical, intermediate and equatorial domains (Braiget al., 1994, 1995; Figure 1(a)). It has been assumedthat chaperonin GroEL in cooperation with a co-chaperonin, the heptameric ring of the 10 kDaGroES, and ATP hydrolysis assists the correct andef®cient folding of substrate proteins. First, sub-
ing author:
dichroism; Cy5-blg,roup labeled by Cy5;ethylated
no group labeled byguanidine
odamine; TMR-blg,roup labeled by
strate proteins in unfolded or intermediate confor-mational states are bound to the hydrophobicapical domains of GroEL so as to prevent theiraggregation and misfolding. Upon binding of co-chaperonin GroES, the trapped intermediates arereleased into the hydrophilic cavity of the GroEL/GroES complex, resulting in ef®cient folding.When the GroES lid is opened, the refolded pro-teins come out while still denatured proteins arebound to GroEL again, thus repeating the bindingand release cycle.
In addition to its interaction with the foldingintermediate, several modes of interaction havebeen reported for GroEL. It interacts with misfoldedor trapped states, unless the aggregation is notextensive, providing a chance to restart the foldingto the native structure (Todd et al., 1994; Smith &Fisher, 1995; Ranson et al., 1995). GroEL is able tointeract with different conformations of the sameprotein (Gervasoni et al., 1998; Kawata et al., 1998).Furthermore, GroEL interacts with proteins in thenative state and consequently unfolds them (Zahn
# 1999 Academic Press
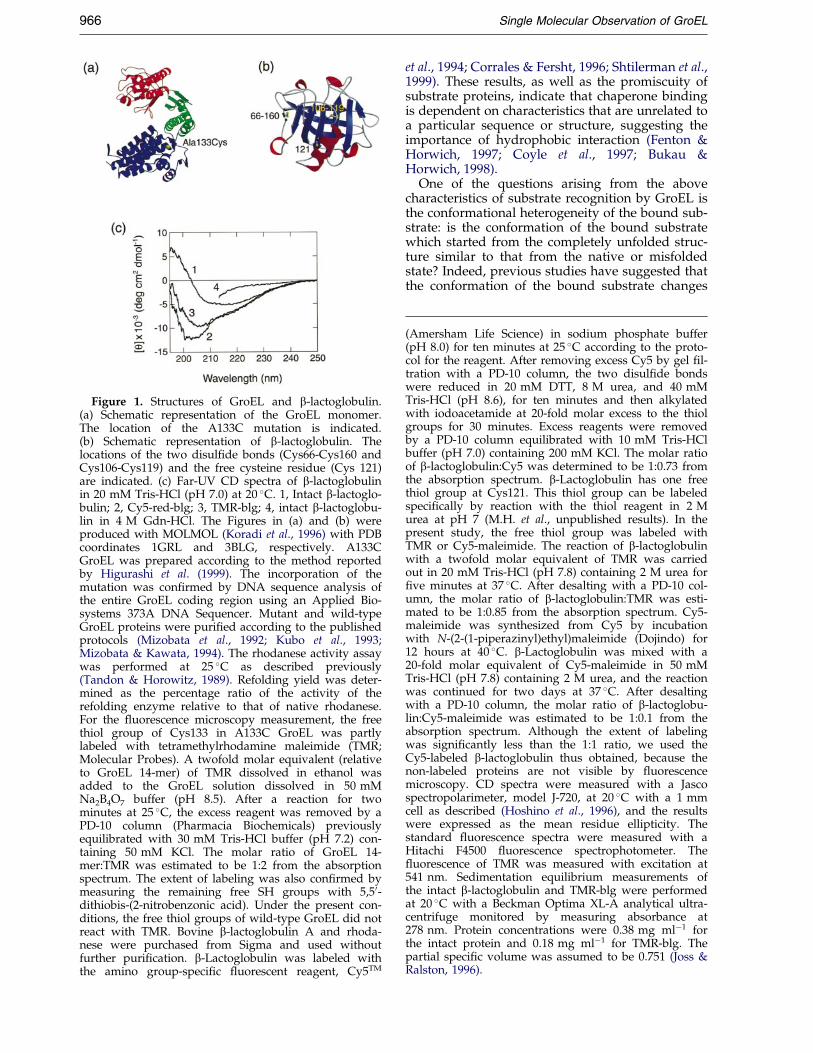
Figure 1. Structures of GroEL and b-lactoglobulin.(a) Schematic representation of the GroEL monomer.The location of the A133C mutation is indicated.(b) Schematic representation of b-lactoglobulin. Thelocations of the two disul®de bonds (Cys66-Cys160 andCys106-Cys119) and the free cysteine residue (Cys 121)are indicated. (c) Far-UV CD spectra of b-lactoglobulinin 20 mM Tris-HCl (pH 7.0) at 20 �C. 1, Intact b-lactoglo-bulin; 2, Cy5-red-blg; 3, TMR-blg; 4, intact b-lactoglobu-lin in 4 M Gdn-HCl. The Figures in (a) and (b) wereproduced with MOLMOL (Koradi et al., 1996) with PDBcoordinates 1GRL and 3BLG, respectively. A133CGroEL was prepared according to the method reportedby Higurashi et al. (1999). The incorporation of themutation was con®rmed by DNA sequence analysis ofthe entire GroEL coding region using an Applied Bio-systems 373A DNA Sequencer. Mutant and wild-typeGroEL proteins were puri®ed according to the publishedprotocols (Mizobata et al., 1992; Kubo et al., 1993;Mizobata & Kawata, 1994). The rhodanese activity assaywas performed at 25 �C as described previously(Tandon & Horowitz, 1989). Refolding yield was deter-mined as the percentage ratio of the activity of therefolding enzyme relative to that of native rhodanese.For the ¯uorescence microscopy measurement, the freethiol group of Cys133 in A133C GroEL was partlylabeled with tetramethylrhodamine maleimide (TMR;Molecular Probes). A twofold molar equivalent (relativeto GroEL 14-mer) of TMR dissolved in ethanol wasadded to the GroEL solution dissolved in 50 mMNa2B4O7 buffer (pH 8.5). After a reaction for twominutes at 25 �C, the excess reagent was removed by aPD-10 column (Pharmacia Biochemicals) previouslyequilibrated with 30 mM Tris-HCl buffer (pH 7.2) con-taining 50 mM KCl. The molar ratio of GroEL 14-mer:TMR was estimated to be 1:2 from the absorptionspectrum. The extent of labeling was also con®rmed bymeasuring the remaining free SH groups with 5,50-dithiobis-(2-nitrobenzonic acid). Under the present con-ditions, the free thiol groups of wild-type GroEL did notreact with TMR. Bovine b-lactoglobulin A and rhoda-nese were purchased from Sigma and used withoutfurther puri®cation. b-Lactoglobulin was labeled withthe amino group-speci®c ¯uorescent reagent, Cy5TM
966 Single Molecular Observation of GroEL
et al., 1994; Corrales & Fersht, 1996; Shtilerman et al.,1999). These results, as well as the promiscuity ofsubstrate proteins, indicate that chaperone bindingis dependent on characteristics that are unrelated toa particular sequence or structure, suggesting theimportance of hydrophobic interaction (Fenton &Horwich, 1997; Coyle et al., 1997; Bukau &Horwich, 1998).
One of the questions arising from the abovecharacteristics of substrate recognition by GroEL isthe conformational heterogeneity of the bound sub-strate: is the conformation of the bound substratewhich started from the completely unfolded struc-ture similar to that from the native or misfoldedstate? Indeed, previous studies have suggested thatthe conformation of the bound substrate changes
(Amersham Life Science) in sodium phosphate buffer(pH 8.0) for ten minutes at 25 �C according to the proto-col for the reagent. After removing excess Cy5 by gel ®l-tration with a PD-10 column, the two disul®de bondswere reduced in 20 mM DTT, 8 M urea, and 40 mMTris-HCl (pH 8.6), for ten minutes and then alkylatedwith iodoacetamide at 20-fold molar excess to the thiolgroups for 30 minutes. Excess reagents were removedby a PD-10 column equilibrated with 10 mM Tris-HClbuffer (pH 7.0) containing 200 mM KCl. The molar ratioof b-lactoglobulin:Cy5 was determined to be 1:0.73 fromthe absorption spectrum. b-Lactoglobulin has one freethiol group at Cys121. This thiol group can be labeledspeci®cally by reaction with the thiol reagent in 2 Murea at pH 7 (M.H. et al., unpublished results). In thepresent study, the free thiol group was labeled withTMR or Cy5-maleimide. The reaction of b-lactoglobulinwith a twofold molar equivalent of TMR was carriedout in 20 mM Tris-HCl (pH 7.8) containing 2 M urea for®ve minutes at 37 �C. After desalting with a PD-10 col-umn, the molar ratio of b-lactoglobulin:TMR was esti-mated to be 1:0.85 from the absorption spectrum. Cy5-maleimide was synthesized from Cy5 by incubationwith N-(2-(1-piperazinyl)ethyl)maleimide (Dojindo) for12 hours at 40 �C. b-Lactoglobulin was mixed with a20-fold molar equivalent of Cy5-maleimide in 50 mMTris-HCl (pH 7.8) containing 2 M urea, and the reactionwas continued for two days at 37 �C. After desaltingwith a PD-10 column, the molar ratio of b-lactoglobu-lin:Cy5-maleimide was estimated to be 1:0.1 from theabsorption spectrum. Although the extent of labelingwas signi®cantly less than the 1:1 ratio, we used theCy5-labeled b-lactoglobulin thus obtained, because thenon-labeled proteins are not visible by ¯uorescencemicroscopy. CD spectra were measured with a Jascospectropolarimeter, model J-720, at 20 �C with a 1 mmcell as described (Hoshino et al., 1996), and the resultswere expressed as the mean residue ellipticity. Thestandard ¯uorescence spectra were measured with aHitachi F4500 ¯uorescence spectrophotometer. The¯uorescence of TMR was measured with excitation at541 nm. Sedimentation equilibrium measurements ofthe intact b-lactoglobulin and TMR-blg were performedat 20 �C with a Beckman Optima XL-A analytical ultra-centrifuge monitored by measuring absorbance at278 nm. Protein concentrations were 0.38 mg mlÿ1 forthe intact protein and 0.18 mg mlÿ1 for TMR-blg. Thepartial speci®c volume was assumed to be 0.751 (Joss &Ralston, 1996).

Single Molecular Observation of GroEL 967
while it is bound to GroEL (Martin et al., 1991;Corrales & Fersht, 1995; Katsumata et al., 1996a).Even under the same solvent conditions, substrateproteins may bind to GroEL in various confor-mations (Gervasoni et al., 1998; Kawata et al., 1998).However, with conventional spectroscopicapproaches, it is dif®cult to clarify the confor-mational heterogeneity of the bound substrate.Among the approaches used so far, mass spec-troscopy combined with the H/2H exchange reac-tion of amide protons (Robinson et al., 1994; Coyleet al., 1999) might be the only method fordistinguishing the conformational heterogeneity ofthe bound substrate.
Single molecule analysis using, for example,atomic force microscopy or the optical tweezerstechnique is becoming popular as a criticalapproach for characterizing the structure and func-tion of protein molecules (Mehta et al., 1999). One ofthe most promising methods is ¯uorescencemicroscopy, in which the behavior of ¯uorescence-labeled protein molecules is observed at the singlemolecule level (Weiss, 1999; Ha et al., 1999).Funatsu et al. (1995) analyzed the ATP hydrolysiscycle of myosin subfragment-1. Using Cy3-labeledATP, they observed the turnover of ATP moleculeson myosin subfragment-1, the enzymatic character-istics being consistent with those obtained macro-scopically in bulk solution. On the other hand, Ishiiet al. (1999) studied the conformational dynamics of¯uorescence-labeled tropomyosin at the single mol-ecule level. They introduced Cy5 and Cy3 at theopposite ends of the molecule and measured the¯uorescence energy transfer between the two dyes,proving that the conformational change could beobserved at the single molecule level.
We expect that single molecule observation of¯uorescence-labeled GroEL and substrate proteinswill be useful for clarifying several unknownaspects of GroEL in the ATP-dependent reactioncycle. As the ®rst step for analyzing the GroELaction at the single molecule level, we prepared acouple of ¯uorescence-labeled substrates, as wellas ¯uorescence-labeled GroEL, and examined theirdirect interaction by total internal re¯ection ¯uor-escence microscopy.
bbb-Lactoglobulin, a substrate protein
As a model substrate protein, we chose bovineb-lactoglobulin, a major component of cow's milk(Hambling et al., 1992; Figure 1(b)). b-Lactoglobulinis an intriguing model for clarifying the mechanismof the a-helix to b-sheet (a! b) transition of pro-teins, a key issue for understanding the foldingand biological function of a number of protein mol-ecules (Prusiner, 1997; Booth et al., 1997; Dalal et al.,1997). Bovine b-lactoglobulin A, with a molecularmass of 18,400 (162 amino acid residues), and twodisul®de bonds and one free thiol group, is a pre-dominantly b-sheet protein consisting of a b-barrelof eight continuous antiparallel b-strands (bA ÿ bH)
shaped into a ¯attened cone or calyx, and onemajor a-helix (Brownlow et al., 1997; Qin et al.,1998; Kuwata et al., 1998, 1999; Figure 1(b)). Thefolding of bovine b-lactoglobulin is accompaniedby an intramolecular a! b transition due to theinconsistency of local and non-local interactions(Shiraki et al., 1995; Hamada et al., 1996). NMR isnow beginning to clarify the mechanism of thea! b transition of b-lactoglobulin (Kuwata et al.,1998, 1999).
Because native b-lactoglobulin did not form astable complex with GroEL measured by size-exclu-sion gel chromatography, we prepared two types ofmodi®ed b-lactoglobulin species, which, as will bedescribed, exhibited signi®cant interaction withGroEL. First, two disul®de bonds of b-lactoglobulinwere reduced by dithiothreitol (DTT) in 8 M urea,and the resulting ®ve thiol groups were alkylatedby iodoacetamide. Then, the reduced and carboxy-methylated b-lactoglobulin was reacted with a ¯u-orescent dye, Cy5, so that about one Cy5 moleculewas introduced to each b-lactoglobulin (Cy5-red-blg). Whereas b-lactoglobulin is a predominantly b-sheet protein, Cy5-red-blg was denatured substan-tially, as measured by far-UV CD, although someresidual structure, probably including some helicalstructures, remained in comparison with thatdenatured in 6 M Gdn-HCl (Figure 1(c)).
b-Lactoglobulin has a free thiol group at Cys121,which is buried underneath the major helix(Figure 1(b)). We prepared a b-lactoglobulinderivative in which the free thiol group wasspeci®cally labeled with Cy5 (Cy5-blg) or tetra-methylrhodamine (TMR-blg). The modi®cation ofthe free thiol group by the bulky ¯uorescentreagents resulted in partial denaturation of the pro-tein molecule, as measured by CD (Figure 1(c)).The extent of denaturation was less than that ofCy5-red-blg, and probably contained non-nativehelical structures.
The oligomeric state of Cy5-blg was examinedby sedimentation equilibrium experiments atpH 7.0. For the intact b-lactoglobulin, the molecu-lar mass was 33,000(�1000), consistent with thedimeric structure (Hambling et al., 1992). For TMR-blg, the molecular mass was 18,700(�500). No sig-ni®cant tendency of aggregation was observed forthe intact b-lactoglobulin or TMR-blg. The resultsclearly indicate that the alkylation of Cys121 bybulky groups like TMR results in the dissociationof dimer into monomer. We consider that the sameis true for Cy5-blg and Cy5-red-blg.
Macroscopic observation
Cy5-red-blg
The interaction of b-lactoglobulin derivativeswith wild-type GroEL was monitored by HPLCsize-exclusion gel chromatography (Figure 2). Allthe modi®ed forms of b-lactoglobulin eluted at aposition much later than that of tetradecamericGroEL. The elution positions of the modi®ed forms
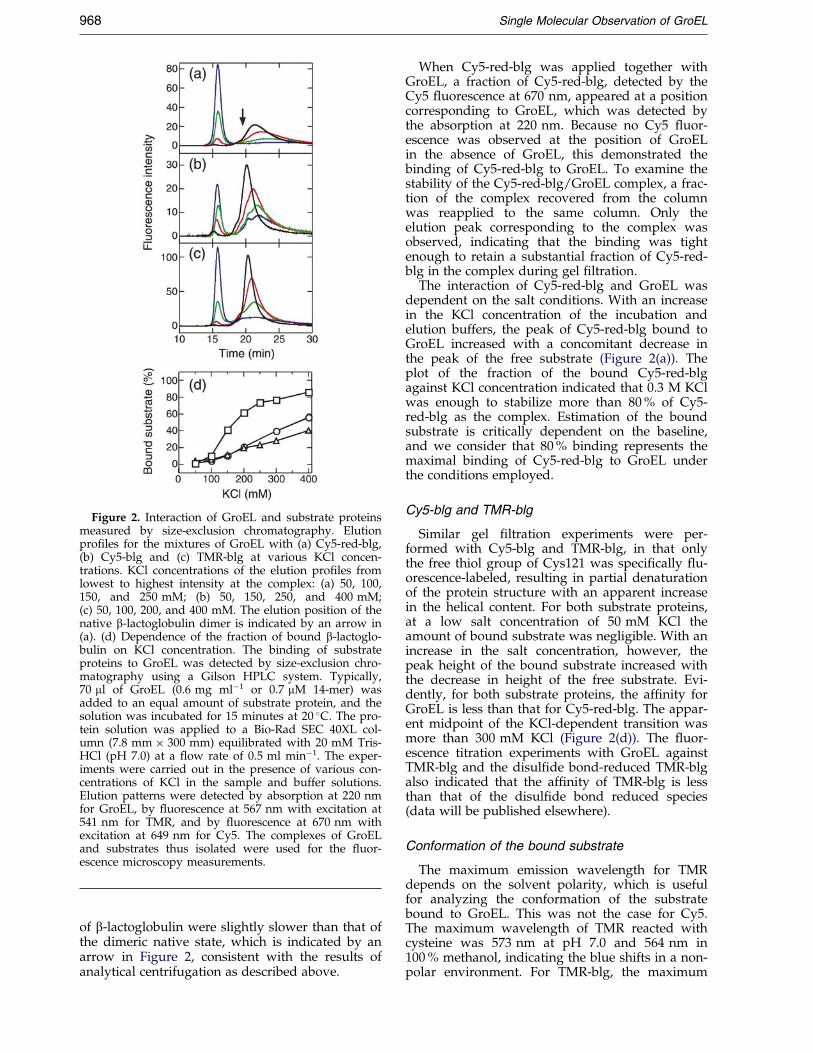
Figure 2. Interaction of GroEL and substrate proteinsmeasured by size-exclusion chromatography. Elutionpro®les for the mixtures of GroEL with (a) Cy5-red-blg,(b) Cy5-blg and (c) TMR-blg at various KCl concen-trations. KCl concentrations of the elution pro®les fromlowest to highest intensity at the complex: (a) 50, 100,150, and 250 mM; (b) 50, 150, 250, and 400 mM;(c) 50, 100, 200, and 400 mM. The elution position of thenative b-lactoglobulin dimer is indicated by an arrow in(a). (d) Dependence of the fraction of bound b-lactoglo-bulin on KCl concentration. The binding of substrateproteins to GroEL was detected by size-exclusion chro-matography using a Gilson HPLC system. Typically,70 ml of GroEL (0.6 mg mlÿ1 or 0.7 mM 14-mer) wasadded to an equal amount of substrate protein, and thesolution was incubated for 15 minutes at 20 �C. The pro-tein solution was applied to a Bio-Rad SEC 40XL col-umn (7.8 mm � 300 mm) equilibrated with 20 mM Tris-HCl (pH 7.0) at a ¯ow rate of 0.5 ml minÿ1. The exper-iments were carried out in the presence of various con-centrations of KCl in the sample and buffer solutions.Elution patterns were detected by absorption at 220 nmfor GroEL, by ¯uorescence at 567 nm with excitation at541 nm for TMR, and by ¯uorescence at 670 nm withexcitation at 649 nm for Cy5. The complexes of GroELand substrates thus isolated were used for the ¯uor-escence microscopy measurements.
968 Single Molecular Observation of GroEL
of b-lactoglobulin were slightly slower than that ofthe dimeric native state, which is indicated by anarrow in Figure 2, consistent with the results ofanalytical centrifugation as described above.
When Cy5-red-blg was applied together withGroEL, a fraction of Cy5-red-blg, detected by theCy5 ¯uorescence at 670 nm, appeared at a positioncorresponding to GroEL, which was detected bythe absorption at 220 nm. Because no Cy5 ¯uor-escence was observed at the position of GroELin the absence of GroEL, this demonstrated thebinding of Cy5-red-blg to GroEL. To examine thestability of the Cy5-red-blg/GroEL complex, a frac-tion of the complex recovered from the columnwas reapplied to the same column. Only theelution peak corresponding to the complex wasobserved, indicating that the binding was tightenough to retain a substantial fraction of Cy5-red-blg in the complex during gel ®ltration.
The interaction of Cy5-red-blg and GroEL wasdependent on the salt conditions. With an increasein the KCl concentration of the incubation andelution buffers, the peak of Cy5-red-blg bound toGroEL increased with a concomitant decrease inthe peak of the free substrate (Figure 2(a)). Theplot of the fraction of the bound Cy5-red-blgagainst KCl concentration indicated that 0.3 M KClwas enough to stabilize more than 80 % of Cy5-red-blg as the complex. Estimation of the boundsubstrate is critically dependent on the baseline,and we consider that 80 % binding represents themaximal binding of Cy5-red-blg to GroEL underthe conditions employed.
Cy5-blg and TMR-blg
Similar gel ®ltration experiments were per-formed with Cy5-blg and TMR-blg, in that onlythe free thiol group of Cys121 was speci®cally ¯u-orescence-labeled, resulting in partial denaturationof the protein structure with an apparent increasein the helical content. For both substrate proteins,at a low salt concentration of 50 mM KCl theamount of bound substrate was negligible. With anincrease in the salt concentration, however, thepeak height of the bound substrate increased withthe decrease in height of the free substrate. Evi-dently, for both substrate proteins, the af®nity forGroEL is less than that for Cy5-red-blg. The appar-ent midpoint of the KCl-dependent transition wasmore than 300 mM KCl (Figure 2(d)). The ¯uor-escence titration experiments with GroEL againstTMR-blg and the disul®de bond-reduced TMR-blgalso indicated that the af®nity of TMR-blg is lessthan that of the disul®de bond reduced species(data will be published elsewhere).
Conformation of the bound substrate
The maximum emission wavelength for TMRdepends on the solvent polarity, which is usefulfor analyzing the conformation of the substratebound to GroEL. This was not the case for Cy5.The maximum wavelength of TMR reacted withcysteine was 573 nm at pH 7.0 and 564 nm in100 % methanol, indicating the blue shifts in a non-polar environment. For TMR-blg, the maximum

Single Molecular Observation of GroEL 969
wavelength at pH 7.0 was 577 nm and was shiftedto 583 nm upon complete unfolding in the pre-sence of 4 M Gdn-HCl. The red shift indicates thatTMR is exposed to a polar environment uponunfolding. For comparison, in the presence of 95 %(v/v) methanol at pH 7.0, the maximum wave-length of TMR-blg was 567 nm. When a ®vefoldmolar excess of GroEL was added to TMR-blg, themaximum wavelength shifted slightly to a shorterwavelength (575 nm). A similar shift in the maxi-mum wavelength was also observed for the com-plex of TMR-blg and GroEL recovered from theHPLC column. These results indicate that theenvironment of TMR in TMR-blg is relatively non-polar and does not change signi®cantly upon bind-ing to GroEL. No signi®cant change in maximalwavelength of TMR upon binding to GroEL wasobserved for the disul®de bond-reduced TMR-blg,which has a maximum at 576 nm. These resultssuggest that the substrate proteins bound to GroELretain a partially folded conformation as observedin solution.
Mechanism of the salt effects
Several examples of modulation of the GroEL-substrate interactions by salt have been reported(Itzhaki et al., 1995; Hoshino et al., 1996; Katsumataet al., 1996a,b; Pack et al., 1999). In our previousstudy (Hoshino et al., 1996), the effects of salt onthe interaction of positively charged cytochrome c(pI � 10) and GroEL were examined. The inter-action was strong under low salt conditions, butan increase in salt concentration resulted in adecrease of af®nity. This salt effect was explainedby a decrease in electrostatic attraction between thepositively charged substrate and the negativelycharged GroEL, in particular around the apicaldomain responsible for substrate recognition. Simi-lar but opposing effects have been reported for a-lactalbumin, where salt stabilizes the interaction(Katsumata et al., 1996a,b). Because a-lactalbuminis an acidic protein with pI � 4, this salt effect canbe explained by a decrease in the electrostaticrepulsion upon addition of salt. b-Lactoglobulin isan acidic protein with pI � 5. Thus, stabilization ofthe interaction by salt is similar to that of a-lactal-bumin, i.e. the shielding of unfavorable electro-static repulsions at a high salt concentration.
Fluorescence microscopy observation
A133C GroEL
In order to detect GroEL at the single moleculelevel, it is necessary to speci®cally label it with anappropriate ¯uorescent dye. We prepared a mutantGroEL protein in which the alanine residue at pos-ition 133 in the equatorial domain of the GroEL sub-unit was mutated to cysteine (A133C GroEL;Figure 1(a)). This mutation site was selected on thebasis of the criterion that the residue would be fully
exposed to solvent, and thus the chaperonin func-tion would not be perturbed by the mutation. Themutant A133C GroEL was easily and speci®callymodi®ed with TMR taking advantage of the intro-duced cysteine residue, and the resulting TMR-labeled GroEL (TMR-GroEL) showed the sameATPase activity as the wild-type GroEL (data notshown). When we examined the refolding activityof TMR-GroEL with rhodanese as a substrate, up to75 % of the rhodanese activity was recovered in thepresence of GroES and ATP, which was the samerefolding activity as that mediated by the wild-typeGroEL protein (data not shown). These results indi-cate that TMR-GroEL has the same chaperoninactivity as the wild-type protein.
Single molecule observation
The chromatography patterns of the complexesformed between Cy5-labeled substrate proteinsand TMR-GroEL were similar to those of wild-typeGroEL shown in Figure 2. Stable complexes elutedat the position of GroEL were recovered from thecolumn, and observed by total internal re¯ection¯uorescence microscopy. The absorption spectrumof the recovered complex indicated that the stoichi-ometry of Cy5-red-blg to TMR-GroEL was 0.8 to 1,close to the stoichiometric binding of substrate andGroEL tetradecamer. The ratio of Cy5-blg to TMR-GroEL could not be determined because the label-ing ef®ciency of Cy5-blg was low. Even though thelabeling ef®ciency or stoichiometry of binding wasnot high, single molecule observation was possiblebecause we did not observe the species without¯uorescence.
The complexes examined by ¯uorescencemicroscopy were those between the substantiallydenatured Cy5-red-blg and TMR-GroEL (Figure 3(a)and (b)) and between the partly denatured Cy5-blgand TMR-GroEL (Figure 3(c) and (d)). The locationof GroEL was identi®ed by the ¯uorescence of TMR(upper panels), and that of substrate proteins wasdetermined by the ¯uorescence of Cy5 (lowerpanels). For both cases, we observed many brightspots with both Cy5 ¯uorescence and TMR ¯uor-escence, indicating the complex of the substrate andGroEL.
A spot with only TMR ¯uorescence, correspond-ing to the free GroEL, was occasionally observed.On the other hand, a spot with only Cy5 ¯uor-escence was rarely observed. These ®ndings areconsistent with the preparation of the complex byrecovering the GroEL fraction by size-exclusionchromatography.
Complex between rhodanese and GroEL
The substrate proteins described above cannotfold into the native state. Although we tried to iso-late the kinetic complex of GroEL and the foldingintermediate of intact b-lactoglobulin, we wereunable to do so. Instead, as a model of the kineticcomplex between GroEL and the foldable inter-
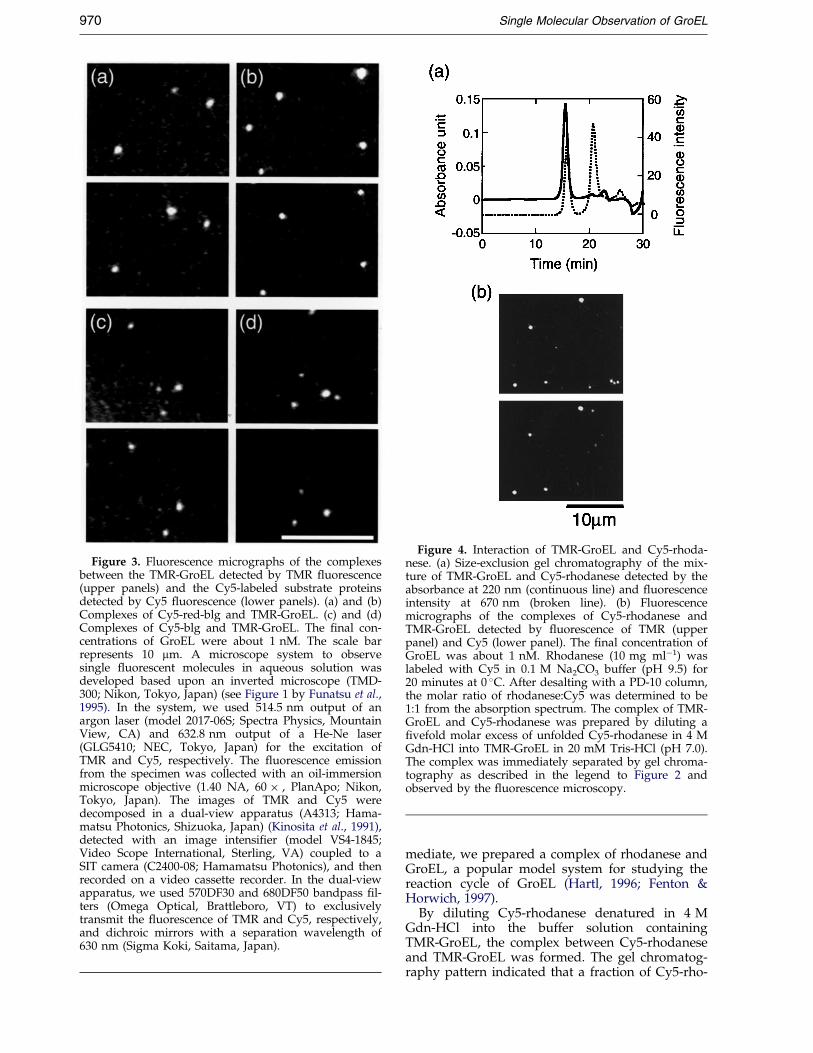
Figure 3. Fluorescence micrographs of the complexesbetween the TMR-GroEL detected by TMR ¯uorescence(upper panels) and the Cy5-labeled substrate proteinsdetected by Cy5 ¯uorescence (lower panels). (a) and (b)Complexes of Cy5-red-blg and TMR-GroEL. (c) and (d)Complexes of Cy5-blg and TMR-GroEL. The ®nal con-centrations of GroEL were about 1 nM. The scale barrepresents 10 mm. A microscope system to observesingle ¯uorescent molecules in aqueous solution wasdeveloped based upon an inverted microscope (TMD-300; Nikon, Tokyo, Japan) (see Figure 1 by Funatsu et al.,1995). In the system, we used 514.5 nm output of anargon laser (model 2017-06S; Spectra Physics, MountainView, CA) and 632.8 nm output of a He-Ne laser(GLG5410; NEC, Tokyo, Japan) for the excitation ofTMR and Cy5, respectively. The ¯uorescence emissionfrom the specimen was collected with an oil-immersionmicroscope objective (1.40 NA, 60 � , PlanApo; Nikon,Tokyo, Japan). The images of TMR and Cy5 weredecomposed in a dual-view apparatus (A4313; Hama-matsu Photonics, Shizuoka, Japan) (Kinosita et al., 1991),detected with an image intensi®er (model VS4-1845;Video Scope International, Sterling, VA) coupled to aSIT camera (C2400-08; Hamamatsu Photonics), and thenrecorded on a video cassette recorder. In the dual-viewapparatus, we used 570DF30 and 680DF50 bandpass ®l-ters (Omega Optical, Brattleboro, VT) to exclusivelytransmit the ¯uorescence of TMR and Cy5, respectively,and dichroic mirrors with a separation wavelength of630 nm (Sigma Koki, Saitama, Japan).
Figure 4. Interaction of TMR-GroEL and Cy5-rhoda-nese. (a) Size-exclusion gel chromatography of the mix-ture of TMR-GroEL and Cy5-rhodanese detected by theabsorbance at 220 nm (continuous line) and ¯uorescenceintensity at 670 nm (broken line). (b) Fluorescencemicrographs of the complexes of Cy5-rhodanese andTMR-GroEL detected by ¯uorescence of TMR (upperpanel) and Cy5 (lower panel). The ®nal concentration ofGroEL was about 1 nM. Rhodanese (10 mg mlÿ1) waslabeled with Cy5 in 0.1 M Na2CO3 buffer (pH 9.5) for20 minutes at 0 �C. After desalting with a PD-10 column,the molar ratio of rhodanese:Cy5 was determined to be1:1 from the absorption spectrum. The complex of TMR-GroEL and Cy5-rhodanese was prepared by diluting a®vefold molar excess of unfolded Cy5-rhodanese in 4 MGdn-HCl into TMR-GroEL in 20 mM Tris-HCl (pH 7.0).The complex was immediately separated by gel chroma-tography as described in the legend to Figure 2 andobserved by the ¯uorescence microscopy.
970 Single Molecular Observation of GroEL
mediate, we prepared a complex of rhodanese andGroEL, a popular model system for studying thereaction cycle of GroEL (Hartl, 1996; Fenton &Horwich, 1997).
By diluting Cy5-rhodanese denatured in 4 MGdn-HCl into the buffer solution containingTMR-GroEL, the complex between Cy5-rhodaneseand TMR-GroEL was formed. The gel chromatog-raphy pattern indicated that a fraction of Cy5-rho-

Single Molecular Observation of GroEL 971
danese was bound to TMR-GroEL (Figure 4(a)).The complex was recovered from the gel and thenobserved by ¯uorescence microscopy (Figure 4(b)).As was the case for b-lactoglobulin derivatives, wedetected several bright spots with both Cy5 ¯uor-escence and TMR ¯uorescence, demonstrating for-mation of the complex.
We examined the effects of addition of ATP tothe GroEL/rhodanese complex recovered by size-exclusion chromatography. When observed by ¯u-orescence microscopy, spots corresponding to freeGroEL and aggregated rhodanese were observedseparately (data not shown), indicating that Cy5-rhodanese was dissociated from the complex, pro-ducing the aggregated forms.
Conclusion
We have shown that the interaction of substrateproteins with GroEL can be observed directly atthe single molecule level by using speci®callylabeled substrate proteins and GroEL with totalinternal re¯ection ¯uorescence microscopy. Thisapproach opens a new avenue for studying theinteraction of GroEL and substrate proteins. In thepresent study, we ®xed neither GroEL nor sub-strate proteins to the plate for ¯uorescence obser-vation. The ®xing of GroEL or substrate proteinswill enable us to follow the ATP and GroES-depen-dent reaction cycle of GroEL, which should shedfurther light on the mechanism of chaperonin-assisted protein folding.
Acknowledgments
We thank Miyo Sakai for the measurements of sedi-mentation equilibrium. This work was supported byGrants-in-Aid for Scienti®c Research from the Ministryof Education, Science, Culture and Sports of Japan.
References
Booth, D. R., Sunde, M., Bellotti, V., Robinson, C. V.,Hutchinson, W. L., Fraser, P. E., Hawkins, P. N.,Dobson, C. M., Radford, S. E., Blake, C. C. F. &Pepys, M. B. (1997). Instability, unfolding andaggregation of human lysozyme variants under-lying amyloid ®brillogenesis. Nature, 385, 787-793.
Braig, K., Otwinowski, Z., Hegde, R., Boisvert, D. C.,Joachimiak, A., Horwich, A. L. & Sigler, P. B.(1994). The crystal structure of the bacterial chaper-onin GroEL at 2.8 AÊ . Nature, 371, 578-586.
Braig, K., Adams, P. D. & BruÈ nger, A. T. (1995). Confor-mational variability in the re®ned structure of thechaperonin GroEL at 2.8 AÊ resolution. Nature Struct.Biol. 2, 1083-1094.
Brownlow, S., Cabral, J. H. M., Cooper, R., Flower, D. R.,Yewdall, S. J., Polikarpov, I., North, A. C. T. &Sawyer, L. (1997). Bovine b-lactoglobulin at 1.8 AÊ
resolution - still an enigmatic lipocalin. Structure, 5,481-495.
Bukau, B. & Horwich, A. L. (1998). The Hsp70 andHsp60 chaperone machines. Cell, 92, 351-366.
Corrales, F. J. & Fersht, A. R. (1995). The folding ofGroEL-bound barnase as a model for chaperonin-mediated protein folding. Proc. Natl Acad. Sci. USA,92, 5326-5330.
Corrales, F. J. & Fersht, A. R. (1996). Toward a mechan-ism for GroEL-GroES chaperone activity: anATPase-gated and -pulsed folding and annealingcage. Proc. Natl Acad. Sci. USA, 93, 4509-4512.
Coyle, J. E., Jaeger, J., Groû, M., Robinson, C. V. &Radford, S. E. (1997). Structural and mechanisticconsequences of polypeptide binding by GroEL.Fold. Des. 2, R93-R104.
Coyle, J. E., Texter, F. L., Ashcroft, A. E., Masselos, D.,Robinson, C. V. & Radford, S. E. (1999). GroELaccelerates the refolding of hen lysozyme withoutchanging its folding mechanism. Nature Struct. Biol.6, 683-690.
Dalal, S., Balasubramanian, S. & Regan, L. (1997). Trans-muting a-helices and b-sheets. Fold. Des. 2,R71-R79.
Fenton, W. & Horwich, A. (1997). GroEL-mediatedprotein folding. Protein Sci. 6, 743-760.
Funatsu, T., Harada, Y., Tokuknaga, M., Saito, K. &Yanagida, T. (1995). Imaging of single ¯uorescentmolecules and individual ATP turnovers by singlemyosin molecules in aqueous solution. Nature, 374,555-559.
Gervasoni, P., Gehrig, P. & PluÈ ckthun, A. (1998). Twoconformational states of b-lactamase bound toGroEL: a biophysical characterization. J. Mol. Biol.275, 663-675.
Ha, T., Ting, A. Y., Liang, J., Caldwell, W. B., Deniz,A. A., Chemla, D. S., Schultz, P. G. & Weizz, S.(1999). Single-molecule ¯uorescence spectroscopy ofenzyme conformational dynamics and cleavagemechanism. Proc. Natl Acad. Sci. USA, 96, 893-898.
Hamada, D., Segawa, S. & Goto, Y. (1996). Non-nativea-helical intermediate in the refolding of b-lacto-globulin, a predominantly b-sheet protein. NatureStruct. Biol. 3, 868-874.
Hambling, S. G., MacAlpine, A. S. & Sawyer, L. (1992).b-Lactoglobulin. In Advanced Dairy Chemistry (Fox,P. F., ed.), pp. 141-190, Elsevier, Amsterdam.
Hartl, F. U. (1996). Molecular chaperones in cellular pro-tein folding. Nature, 381, 571-580.
Higurashi, T., Nosaka, K., Mizobata, T., Nagai, J. &Kawata, Y. (1999). Unfolding and refolding ofEscherichia coli chaperonin GroES is expressed by athree-state model. J. Mol. Biol. 291, 703-713.
Hoshino, M., Kawata, Y. & Goto, Y. (1996). Interactionof GroEL with conformational states of horse cyto-chrome c. J. Mol. Biol. 262, 575-587.
Ishii, Y., Funatsu, T., Yoshida, T., Wazawa, T. &Yanagida, T. (1999). Fluorescence resonance energytransfer between single ¯uorophores attached to acoiled-coil protein in aqueous solution. Chem. Phys.247, 163-173.
Itzhaki, L. S., Otzen, D. E. & Fersht, A. R. (1995). Natureand consequence of GroEL-protein interactions. Bio-chemistry, 34, 14581-14587.
Joss, L. A. & Ralston, G. B. (1996). b-Lactoglobulin B:a proposed standard for the study of reversibleself-association reactions in the analytical ultra-centrifuge? Anal. Biochem. 236, 20-26.
Katsumata, K., Okazaki, A. & Kuwajima, K. (1996a).Effect of GroEL on the refolding kinetics of a-lactal-bumin. J. Mol. Biol. 258, 828-838.

972 Single Molecular Observation of GroEL
Katsumata, K., Okazaki, A., Tsurupa, G. P. &Kuwajima, K. (1996b). Dominant forces in therecognition of a transient folding intermediate ofa-lactalbumin by GroEL. J. Mol. Biol. 258, 828-838.
Kawata, Y., Hongo, K., Mizobata, T. & Nagai, J. (1998).The chaperonin GroE-facilitated folding of the disul-®de-bonded and non-bonded Taka-amylase A fromAspergillus oryzae. Protein Eng. 11, 1293-1298.
Kinosita, K., Itoh, H., Ishiwata, S., Hirano, K.,Nishizaka, T. & Hayakawa, T. (1991). Dual-viewmicroscopy with a single camera: real-time imagingof molecular orientations and calcium. J. Cell Biol.115, 67-73.
Koradi, R., Billeter, M. & WuÈ thrich, K. (1996). MOLMOL- a program for display and analysis of macromol-ecular structures. J. Mol. Graph. 14, 51-55.
Kubo, T., Mizobata, T. & Kawata, Y. (1993). Refoldingof yeast enolase in the presence of the chaperoninGroE. J. Biol. Chem. 268, 19346-19351.
Kuwata, K., Hoshino, M., Era, S., Batt, C. A. & Goto, Y.(1998). a! b transition of b-lactoglobulin as evi-dence by heteronuclear NMR. J. Mol. Biol. 283, 731-739.
Kuwata, K., Hoshino, M., Forge, V., Era, S., Batt, C. A.& Goto, Y. (1999). Solution structure and dynamicsof bovine b-lactoglobulin. Protein Sci. In the press.
Martin, J., Langer, T., Boteva, R., Schramel, A., Horwich,A. L. & Hartl, F.-U. (1991). Chaperonin-mediatedprotein folding at the surface of GroEL througha `molten globule'-like intermediate. Nature, 352, 36-42.
Mehta, A. D., Rief, M., Spudich, J. A., Smith, D. A. &Simmons, R. M. (1999). Single-molecule biomecha-nics with optical methods. Science, 283, 1689-1695.
Mizobata, T. & Kawata, Y. (1994). The guanidine-induced conformational changes of the chaperoninGroEL from Escherichia coli: evidence for the exist-ence of an unfolding intermediate state. Biochim.Biophys. Acta, 1209, 83-88.
Mizobata, T., Akiyama, Y., Ito, K., Yumoto, N. &Kawata, Y. (1992). Effects of the chaperonin GroEon the refolding of tryptophanase from Escherichiacoli. J. Biol. Chem. 267, 17773-17779.
Pack, C.-G., Nishimura, G., Tamura, M., Aoki, K.,Taguchi, H., Yoshida, M. & Kinjyo, M. (1999).Analysis of interaction between chaperonin GroELand its substrate using ¯uorescence correlationspectroscopy. Cytometry, 36, 247-253.
Prusiner, S. B. (1997). Prion diseases and the BSE crisis.Science, 278, 245-251.
Qin, B. Y., Bewley, M. C., Creamer, L. K., Baker, H. M.,Baker, E. N. & Jameson, G. B. (1998). Structuralbasis of the Tanford transition of bovine b-lacto-globulin. Biochemistry, 37, 14014-14023.
Ranson, N. A., Dunster, N. J., Burston, S. G. & Clarke,A. R. (1995). Chaperonins can catalyse the reversalof early aggregation steps when a protein misfolds.J. Mol. Biol. 250, 581-586.
Robinson, C. V., Groû, M., Eyles, S. J., Ewbank, J. J.,Mayhew, M., Hartl, F. U., Dobson, C. M. &Radford, S. E. (1994). Conformation of GroEL-bound a-lactalbumin probed by mass spectrometry.Nature, 372, 646-651.
Shiraki, K., Nishikawa, K. & Goto, Y. (1995). Tri¯uoro-ethanol-induced stabilization of the a-helical struc-ture of b-lactoglobulin: implication for non-hierarchical protein folding. J. Mol. Biol. 245, 180-194.
Shtilerman, M., Lorimer, G. H. & Englander, S. W.(1999). Chaperonin function: folding by forcedunfolding. Science, 284, 822-825.
Smith, K. E. & Fisher, M. T. (1995). Interaction betweenthe GroE chaperonins and rhodanese. J. Biol. Chem.270, 21517-21523.
Tandon, S. & Horowitz, P. M. (1989). Reversible foldingof rhodanese. J. Biol. Chem. 264, 9859-9866.
Todd, M. J., Viitanen, P. V. & Lorimer, G. H. (1994).Dynamics of the chaperonin ATPase cycle: impli-cations for facilitated protein folding. Science, 265,659-665.
Weiss, S. (1999). Fluorescence spectroscopy of single bio-molecules. Science, 283, 1676-1683.
Zahn, R., Spitzfaden, C., Ottiger, M., WuÈ thrich, K. &PluÈ ckthun, A. (1994). Destabilization of the com-plete protein secondary structure on binding to thechaperone GroEL. Nature, 368, 261-265.
Edited by A. R. Fersht
(Received 3 June 1999; received in revised form 16 August 1999; accepted 20 August 1999)