single locus microsatellite development for the … papers/jtas vol. 28 (2) sep...single locus...
TRANSCRIPT

PertanikaJ. Trop. Agric. Sci. 28 (2): 87-94 (2005)ISSN: 1511-3701
©Universiti Putra Malaysia Press
Single Locus Microsatellite Development for the Malaysian Giant FreshwaterPrawns, Macrobrachium rosenbergii
!SUBHA BHASSU, ]ROSLYHASSAN, 2KHATIJAH YUSOFF, 3ZAINODDIN JAMARI,2SITI SHAPOR SIRAJ & *TAN SOON GUAN
1 Freshwater Fisheries Research Center, Batu Berendam, 75350 Melaka, Malaysia2Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
^National Prawn Fry Production and Research Centre (NAPFRE), MalaysiaE-mail: [email protected]
Keywords: Genetic diversity, M. rosenbergii microsatellites
ABSTRAKJJdang genus M. rosenbergii, Bate 1868 (Palaemonidae) adalah kumpulan pelbagai jenis krustasea decopod yangdidapaii di kawasan tropika, laut, estuari dan air tawar. Pada masa ini, stok liar telah digunakan untuk ternakanpengeluaran benih di kebanyakan tempat di Malaysia. Walau bagaimanapun, pengambilan stok liarsecara berterusanakan mengakibatkan kepupusan dan kehilangan kepelbagaian genetik. Oleh itu adalah penting bagi seseorangmempunyai kepekaan terhadap maklumat genetik untuk tujuan pemuliharaan dan pembiakan. Kajian inimenggunakan teknik RAMS untuk menghasilkan penanda lokus mikrosatelit tunggal. Sebanyak 13 kawasanmikrosatelit telah dikesan. Hasil kajian menujukkan kejayaan menggunakan teknik ini untuk memperoleh mikrosatelitdalam M. rosenbergii. Kajian ini akan diteruskan untuk mencari klonyang mengandungi mikrosatelit dan merekabentuk primer bagi setiap kawasan mikrosatelit. Jujukannya adalah sama dengan dua spesies lain iaitu ikan zebradan Perna viridis. Kewujudan persamaan pada kedua-dua spesies (ikan Zebra dan Perna viridis) akan digunakansebagai 'model* bagi M. rosenbergii untuk masa hadapan.
ABSTRACTPrawns of the genus Macrobrachium Bate, 1868 (Crustacea: Palaemonidae) are a highly diverse group of decopodcrustaceans found in circumtropical marine-, estuarine- and freshwaters. At present, wild stocks have been used tocultivate commercial cultures in many areas in Malaysia. Such repeated harvesting from wild stocks, however, willeventually lead to extinction and loss of genetic diversity. Therefore, it is important to know the genetic make up of thesewild stocks in order to manage the populations for conservation and breeding purposes. In the present study, a novelmethod known as Random Amplified Microsatellites (RAMS) was used to develop single locus microsatellite markers. Atotal of 13 microsatellite regions were detected. The findings of this study showed a successful detection of the microsatellitesequences in M. rosenbergii using RAMS. This study will be continued to identify more clones that contain microsatellitesequences and to design the forward and reverse primers for each microsatelite region. M. rosenbergii sequences werehomologous with Danio rerio (Zebra fish) and Perna viridis (green lipped mussel).
INTRODUCTIONMacrobrachium rosenbergii, the giant freshwaterprawn (Family: Palaemonoidea, subfamily:Palaemoninae) is a component of thebiodiversity in Malaysia's diverse waterecosystems. It inhabits tropical and subtropicalzones of the world, in lakes, rivers, swamps,
irrigation ditches, canals and ponds, as well asin estuarine areas. Most species requirebrackish water in the initial stages of their lifecycle, and a minority complete their life cyclein inland saline and freshwater lakes. Theirnumbers in these natural habitats has beendeclining due to overexploitation and habitat

SUBHA BHASSU ETAL.
alterations. At present, wild stocks have beenused to cultivate commercial cultures in manyareas in Malaysia. Repeated harvesting fromthe wild stocks will lead to extinction andeventually loss of genetic diversity. To save thisimportant bio-resource, effective conservationand propagation-assisted rehabilitationstrategies are necessary. However, this may notbe feasible unless data is available forM. rosenbergii on stock structure and geneticvariation throughout its distribution range.The identification of polymorphic markerswith consistent scorable alleles is a crucial stepto generate population genetics data. A studyconducted by Yuzine etal (1996 using isozymesas genetic markers) revealed thatM rosenbergiifrom Rompin River, Linggi River and theHatchery Unit at Universiti Putra Malaysiahave low genetic variability when compared toP merguiensis. The low genetic variabilityshowed that these populations constitutespecies that need to be further investigated byusing DNA markers. A recent study using themitochondrial 16S ribosomal RNA geneplaced Malaysian M. rosenbergii into thewestern 'form' along with those fromThailand, Java and Vietnam. (Mather and deBruyn 2003). This information hasimplications for both the aquaculture industryand the conservation of wild stocks.
Population genetic markers includingallozymes, mitochondrial DNA andmicrosatellites can be the markers of choicefor addressing issues such as genetic variability,inbreeding, parentage assignments, speciesand strain identification and for theconstruction of high resolution genetic linkagemaps for aquaculture species. Thus for thisstudy, microsatellites was the marker of choice.The great popularity of microsatellite markersis mainly due to their presence in allprokaryotic and eukaryotic genomes, andtheir codominant, highly polymorphic andeasy to score characteristics. Thesecharacteristics make them more informativethan dominant markers such as RAPD, AFLP,and others. Microsatellites were used toestimate relative levels of genetic divergencewithin and among populations. Microsatellites
can also identify genetically distinctpopulations, which contribute to conservationefforts. To date, only six microsatellite locihave been developed for the eastern form ofM. rosenbergii and they failed to show anypolymorphism in the western form of M.rosenbergii These six loci exhibited null allelesin the western form. (Mather, pers. comm.).Thus it will be a breakthrough in the moleculargenetics of M. rosenbergii if steps are taken todevelop microsatellite markers by isolatingmicrosatellite regions and developing singlelocus microsatellite markers for this species.At present, a well developed protocol for singlelocus microsatellite identification has beendeveloped for Perna viridis (Teh et al 2002).Single locus microsatellites are highly repeatedDNA sequences that have been markers ofchoice in resolving genetic variability in lowvariability populations, genome mapping,parentage, kinship assignments and stockstructures. In this study, we have attempted todevelop microsatellite loci for M. rosenbergii.
MATERIALS AND METHODSPrawns were collected from the wild stocksfrom Muda River (Kedah), Perak River (Perak)and Rompin River (Pahang). The sampleswere kept on ice for transportation and storedat -80°C for the study. Identification of theprawns collected was based on the taxonomicidentification keys provided by New (2002).DNA was extracted from the muscle tissues bythe protocol of Winnepenninckx et al (1993).The microsatellite isolation techniquefollowed the procedure of Teh et al (2003).This involved the creation of an enrichedgenomic library based on the 5' anchored PCRtechnique. Degenerate primers were used forPCR and the DNA bands generated from eachof the primers were ligated into the TOPO TAcloning vector (Invitrogen, USA). Vectors withinserts were then transformed intocompetent'^, coli cells. The transformed cellswere then plated onto LB agar, andsubsequently selection for white colonies weremade after incubation at 37°C. Plasmid DNAswere extracted from positive clones, using theprotocol of Sambrook et al (1989) before
88 PERTANIKAJ. TROR AGRIC. SCI. VOL. 28 NO. 2 (2005)

SINGLE LOCUS MICROSATELLITE DEVELOPMENT FOR M. ROSENBERGII
sequencing on an ABI PRISM 377 DNAsequencer
Analysis of nucleotide sequences wascarried out with BLAST-N program. Thisprogram was used to identify similarities ofnucleotide sequences obtained with those ofpublished sequences of other closely relatedspecies and to determine the significance ofthese sequences.
RESULTS AND DISCUSSIONA total of 16 degenerate RAMS primers werescreened to check for amplification. Allshowed amplification. However, PCRoptimization was done for some of theseprimers. A single degenerate primercontaining (G\G)5 microsatellite repeat motifswas used to detect the presence ofmicrosatellite sequences. The primersequence of Bp 11 (5' to 3') is(K)3(YH)2Y(CAG)5, K= G/T, V= G/C/A, Y=T,H=A,C or T. The size of the amplified bandsranged from 200bp to lOOObp (Figure 1).
M 1Figure 1: Banding pattern observed in one of the
individuals in M. rosenbergii usingRAMS primer, Bpll. Lane M: 100 bp marker,
Lane 1: Bpll primer bands
The whole PCR product of primer Bp-11was cloned and almost 200 clones with anaverage insert size of 200 bp were isolated. A
total of 12 positive clones were sequenced.Eight of the clones contained perfect CAGrepeats and cryptic simple (GG)n(GGG)nrepeats while the rest of the clones had noinserts (Table 1). These results suggest that the(CAG)n repeats are abundant in the genomeof M. rosenbergii
Nineteen cryptic simple regions weredetected in the eight clones. Cryptic simpleregions were found preferentially in the non-coding regions of the genomes but they arealso found in the coding regions (Goldstein1999). According to Tautz et al (1986),microsatellites probably arose from crypticsimple regions. On the other hand,microsatellites may have undergone mutationsto become regions of cryptic simplicity.
Random Amplified Microsatellites(RAMS) is proven to be an efficient startingpoint for developing single locus microsatellitemarkers in M. rosenbergii. This technique hasbeen used to characterize species such asGremmeniella abietina ( Han tula et al. 1996);Pinus radiata (Fisher et al. 1998); Vigna radiata( Kumar et al 2002) and blue mussel, Mytilusedulis (Presa et al 2002). RAMS markers haveprovided useful information about populationstructures of these species. The findings of ourstudy showed the success of using this methodas a starting point for detecting microsatellitesequences in M. rosenbergii The microsatellitesequences that we have obtained will be usedto design primers based on the uniquesequences flanking each microsatellite region.These primer pairs will then be tested for theirabilities to amplify single locus microsatellitemarkers in M. rosenbergii The development ofpolymorphic microsatellite loci will be usefulin rebuilding wild stock populations,contributing significantly to environmentalconservation and it will serve to providevaluable information for broodstockmanagement of M. rosenbergii
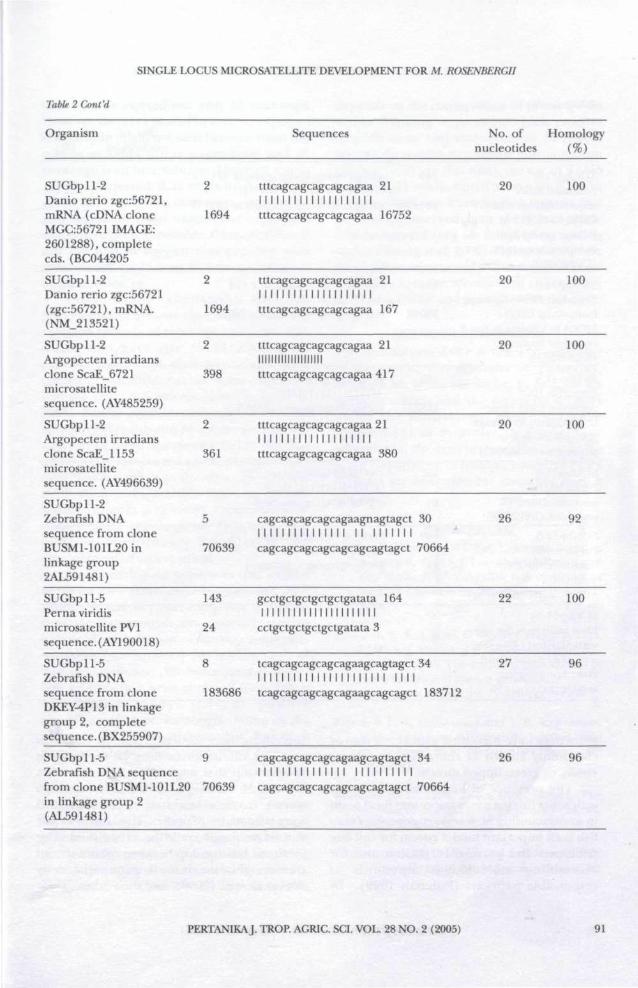
Blast analysis was performed to identifyhomology sequences between M. rosenbergiiwith its closely related species. The list ofsequences between M. rosenbergii and otherspecies are shown in Table 2. The analysisshowed that the majority of the homologous
PERTANIKAJ. TROP. AGRIC. SCI. VOL. 28 NO. 2 (2005) 89

SUBHABHASSU ETAL.
TABLE 1Microsatellite loci isolated from Macrobrachium rosenbergii using Bp-11 RAMS clone
and their GenBank accession numbers
Clone Type of repeat units
Perfect Imperfect Compound GenBank Accession
SUGbpll-1 (CAG)5,(TC)4, (GCT)5, (TAG)3
SUGbpll-2 (CAG)5
SUGbpll-3 (CAG)6 ,(TGC)4
SUGbpll-4 (CAG)5
SUGbpll-5 (CAG)5, (TGC)4
SUGbpll-6 (CAG)7, (TGC)4
SUGbpll-7 (CAG)5
SUGbp 11-34 (CAG)6
(GG)4(GGG)2(AG),,(GG)5(GGG)2(GG)5(GGG)2(GG)2(CA)2(CA)3
(GG)5(GGG)2(GG)5 (GGG)2(GG)2(GG)5(GGG)2(GG),(GGG)2
AY841999AY842000AY842001AY842002
AY842003AY842004AY842005AY842006
TABLE 2Nucelotide sequence comparisons of the five clones of M. rosenbergii with other species
Organism Sequences No. of Homologynucleotides (%)
Clone SUGbpll-1 8Cepaea nemoralismicrosatellite 343Cnelsequence(AF139019)
cagcagcagcagcaggaagagcaggagcagtagtagtag 46i l l i i i i i i i i n n i i n n n 111111 i i 1111cagcagcagcagcagcagcagcagcagcagtagtagtag 30
39 89
Clone SUBbpl 1-1 9Fonscochleaaccepta clone 130Sn49 microsatellitesequence.(AY805231)
agcagcagcagcaggaagagcaggagcagtagtagtag 46III IIIIIII MM I Mill MIMMMIMMagcagcagcagcagcagcagcagcagcagtagtagtag 167
39 87
SUGbpll-2Zebrafish DNAsequence fromclone CH211-HP18in linkagegroup 22,completesequence.(AL845526)
2 tttcagcagcagcagcagaa 21I l l l l l l l f MIMIMIM
16074 tttcagcagcagcagcagaa 160934
20 100
SUGbpll-2Zebrafish DNA 4sequence fromclone DKEY-4P13 183686in linkage group 2,(BX255907)
tcagcagcagcagcagaagnagtagct 30II II III I I II I III III I II M M
tcagcagcagcagcagaagcagcagct 183712
27 97
90 PERTANIKAJ. TROP. AGRIC. SCI. VOL. 28 NO. 2 (2005)
SINGLE LOCUS MICROSATELLITE DEVELOPMENT FOR M. ROSENBERG!!
TabU2ConCd
Organism Sequences No. of Homologynucleotides (%)
SUGbpll-2Danio rerio zgc:56721,mRNA (cDNA cloneMGC:56721 IMAGE:2601288), completecds. (BC044205
2 tttcagcagcagcagcagaa 21I I I III I II I I I I M I III I
1694 tttcagcagcagcagcagaa 16752
20 100
SUGbpll-2 2Danio rerio zgc:56721(zgc:56721), mRNA. 1694(NM_213521)
tttcagcagcagcagcagaa 21I l l l l l I I I I I I I I I I I 11 Itttcagcagcagcagcagaa 167
20 100
SUGbpll-2 2Argopecten irradiansclone ScaE_6721 398microsatellitesequence. (AY485259)
tttcagcagcagcagcagaa 21Illllllllllllllllllltttcagcagcagcagcagaa 417
20 100
SUGbpll-2 2Argopecten irradiansclone ScaE_l 153 361microsatellitesequence. (AY496639)
tttcagcagcagcagcagaa 21I l l l l l l l l l l l l l l l l l l ltttcagcagcagcagcagaa 380
20 100
SUGbpll-2Zebrafish DNAsequence from cloneBUSMM01L20inlinkage group2AL591481)
5 cagcagcagcagcagaagnagtagct 30II I I I I I I I I I II I I II I l l l l l I
70639 cagcagcagcagcagcagcagtagct 70664
26 92
SUGbpll-5 143Perna viridismicrosatellite PV1 24sequence.(AYl 90018)
gcctgctgctgctgctgatata 164I l l l l l l l l l l l l l l l l l l l l l
cctgctgctgctgctgatata 3
22 100
SUGbpll-5Zebrafish DNAsequence from cloneDKEY-4P13 in linkagegroup % completesequence. (BX255907)
183686
tcagcagcagcagcagaagcagtagct 34I I I I I I I I I I I I I I I I I I I I I I mitcagcagcagcagcagaagcagcagct 183712
27 96
SUGbpll-5Zebrafish DNA sequencefrom clone BUSM1-101L20in linkage group 2(AL591481)
70639
cagcagcagcagcagaagcagtagct 34I I II I Ifi III II I I I I I III I II Icagcagcagcagcagcagcagtagct 70664
26 96
PERTANIKAJ. TROP. AGRIC. SCL VOL. 28 NO. 2 (2005) 91

SUBHA BHASSU ETAL
Table2Cont'd
Organism Sequences No. of Homologynucleotides (%)
SUGbpll-6Zebrafish DNAsequence from cloneCH211-245M19inlinkage group 5,complete sequence.(CR385090)
144 gtttcgcctgctgctgctgctgaaataa 171Nil 111 I M II111111111111111
124120 gtttggcctgctgctgctgctgaaataa 124147
SUGbpll-6Zebrafish DNA sequencefrom clone CH211-137C4 in linkage group 8,complete sequence.(BX248231)
151 ctgctgctgctgctgaaataacca 1741111111111111J1111 Mill
50048 ctgctgctgctgctgaaacaacca 50071
24 96
SUGbpll-6Zebrafish DNAsequence from cloneDKEY-32D10 in linkagegroup 5, completesequence. (BX469907)
149 gcctgctgctgctgctgaa 167m m M I N I M U M
128335 gcctgctgctgctgctgaa 128353
19 100
SUGbpll-6Perna viridismicrosatellite PV3sequence. (AY190020)
151
24
ctgctgctgctgctgaaataac 172I l l l l l l l l l l l l l l l l l l l l lctgctgctgctgctgaaataac 3
22 100
SUGbpll-6 149Perna viridismicrosatellite PV1 24sequence.
ACCESSION AY190018
gcctgctgctgctgctgaaataac 172MINI II IIMIIIIM Millgcctgctgctgctgctgatataac 1
24 96
SUGbp 11-34Blast analysis is the samewith SUGbpll-6 to thesimilar organism andsame homologoussequences
sequences were identified with Danio rerio orcommonly known as zebra fish and Pernaviridisy or green lipped muscle.
The presence of homologous sequencewith zebra fish is a a very important focal pointin understanding M. rosenbergii genome. Zebrafish is an important model system for cell fatedecisions and pattern formation and forassembling molecular components ofresponsible pathways (Fishman 1999). By
identifying more sequences from M. rosenbergiiand with the understanding on the possiblerelationship that might exist between zebrafish and M. rosenbergii, the role of M. rosenbergiigenes from zebra fish mutations andidentification of zebra fish models forcandidate genes could be established. Thesynthetic relationship between zebra fish andhuman genome is well documented byBarbazuk et al (2000), and shows that similar
92 PERTANIKAJ. TROP. AGRIC. SCI. VOL. 28 NO. 2 (2005)

SINGLE LOCUS MICROSATELLITE DEVELOPMENT FOR M. ROSENBERG/I
work can be carried out with M. rosenbergii.However, there is a possibility that the expectedrelationship might not exist between these twospecies as Zebra fish is a vertebrate and M.rosenbergii is an invertebrate. However, Ohno(1970) proposed that without duplicatedgenes the creation of metazoans, vertebratesand mammals from unicellular organismswould have been impossible. Ohno (1970) wasnot the first to suggest that genome wideredundancy could lead to new evolutionaryopportunities.
Intragenome similarities throughphylogeny and synteny data suggest that thecommon ancestor of zebra fish and puffer fish,a fish that gave rise to 22,000 speciesexperienced a large scale gene duplicationevent and puffer fish has lost many duplicatesthat the zebra fish has retained (Taylor et al2003). Similar possible relationship could existbetween Zebra fish and M. rosenbergii becauseancestor of these two species (zebra fish andpuffer fish) experienced a genome duplicationevent lends support to the idea that genomeduplication and speciation might be casuallylinked (Amores et al 1998). Sequence levelstudies on M. rosenbergii will help to answerpossible evolutionary lineages in thedevelopment of an organism.
The homologous sequences that existedbetween two families, Palaemonidae andMytiloidea, was very interesting due to the factthat similar studies on marine turtlesindicated conservation of flanking sequencesspanning approximately 300 million years ofdivergent evolution (FitzSimmons etal 1995).Few primers that were obtained from Teh etal. (unpublished) showed positiveamplification in M. rosenbergii. Some of theprimers showed polymorphic bandingpatterns (Bhassu et al unpublished data).
CONCLUSIONThis study will be continued to test thehypothesis that microsatellite loci have greatpotential for broader applications such ascomparative gene mapping and assessinggenetic population structures within species.The use of microsatellite loci across species
depends on the conservation of priming siteswithin flanking sequences, which enablesamplification and maintenance of repeatarrays long enough to promote polymorphism(Weber 1990). In this study, the use of RAMSprimer Bpl 1 was designed to amplify (CAG)nrepeats which are trinucleotide regions thatare more conserved than the dinucleotideregions because they are found in the codingregion (Young et al 2000) This could explainthe possible existence of conservation of thesesequences between species such as Zebra fishand Perna viridis and possibly other species.
ACKNOWLEDGEMENTSThe authors would like to thank the Directorof Research, Department of Fisheries,Malaysia, Mr Ismail bin Awang Kechik, for hisvaluable advice and the financial supportunder the BioDiversity program of theDepartment of Fisheries, Malaysia. Ourappreciation also goes to the staff and studentsof the Department of Biology, Universiti PutraMalaysia for their valuable assistance duringthis study.
REFERENCESAMORES, A., A. FORCE, Y L YAN, L. JOLY, C. AMEMIYA,
A. FRITY, R. K. H O , J. LANGELAND, V.PRINCE and
YL. WANG. 1998. Zebrafish hox clusters andvertebrate genome evolution. Science2S2:1711-1714.
BARBAZUK, W. B., I. KORF, C. KADAVI,J. HEYEN, S. TATE,
E. WUN, J. BEDELL, J.D. MGPHERSON and S.L.
JOHNSON. 2000. The synthetic relationship ofthe zebrafish and human genomes.unvw.genome .org/cgi/doi/10.1101/gr. 144700
FISHER, P. J., R. C. GARDNER and T. E. RICHARDSON.
1996. Single locus microsatellite isolated using5'- anchored PCR. Nucleic Acids Research 24:4369-4371.
FISHMAN, M. C. 1999. Zebra fish genetics. Theenigma of arrival. Proc. Natl. Acad. Sci. 96:10554- 10556. USA.
FITZ SIMMON, N. N., C. MORITZ and S. S. MOORE. 1995.
Conservation and dynamics of microsatellite
PERTANIKA J. TROP. AGRIC. SCI. VOL. 28 NO. 2 (2005) 93

SUBHA BHASSU ETAL.
loci over 300 million years of marine turtleevolution. Mol Biol. Evol 12 (3): 432-440.
HANTULAJ., M. DUSABENYAGASANI and R. C. HAMELIN.
1996. Random amplified microsatellites(RAMS) - a novel method for characterizinggenetic variation within fungi. European JournalofForest Pathology 26: 159-166.
GOLDSTEIN, D. B. and C. SCHLOTTERRER. 1999.
Microsatellites: Evolution and Applications. USA:Oxford University Press.
KUMAR, S. V., S. G. TAN, S .C. QUAH and K. YUSOFF.
2002. Isolation of microsatellite markers inmungbean, Vigna radiata. Molecular EcologyNotes 2: 96-98.
MATHER, P. B. and M. DE BRUYN. 2003. Geneticdiversity in wild stocks of the giant freshwaterprawn (Macrobrachiumrosenbergii): Implicationsfor aquaculture and conservation. AquacultureMagazinejune: 1-3.
NEW, M. B. 2000. History and global status offreshwater prawn farming. In Freshwater PrawnCulture the Farming of Macrobrachium rosenbergii,ed. M. B. New and W. C. Valenti, pp. 1-11.Oxford: Blackwell Science.
OHNO, S. 1970. Evolution by Gene Duplication. NewYork: Springer-Verlag.
PRESA, P., M. PEREZ and A. P. DIAZ. 2002.
Polymorphic microsatellite markers for theblue mussels (Mytilusspp.) Conservation Genetics3: 441-443.
SAMBROOK, J., E. F. FRITSCH and T. MANIATIS. 1989.
Molecular Cloning: A Laboratory Mannual 2 edn.New York: Cold Spring Harbor Laboratory,Cold Spring Harbor.
TAUTZ, D., M. TRICK and G. A. DOVER. 1986. Cryptic
simplicity in DNA is a major source of geneticvariation. Nature 322: 652-656.
TAYLOR, J. S., I. BRAASCH, T. FRICKEY, A. MEYER and
Y. Y. DE PEER. 2003. Genome duplication, a traitshared by 22,000 species of ray-finnedfish, xvxvw. genome, org/cgi/doi/10.1101 /gr. 640303.
TEH, C. H., B. H. CHUA, S. Y YONG, S. G. TAN and
K. M. YUSOFF. 2003. Development of singlelocus microsatellite markers for the Green-Lipped Mussel, Perna viridis using a RandomAmplified Microsatellites (RAMS) basedtechnique. In Aquatic Resource andEnvironmental Studies of the Straits of Malacca:Managing the Straits through Science andTechnology, ed. J.S. Bujang, A. Arshaad, M. H.Zakaria and Kawamura, pp. 351-356.
WEBER, J. L. 1990. Informativeness of human 9dC-dA)n -(dG-dT)n polymorphisms. Genomics 7:524-530.
WINNEPENNINCKX, B., T. BACKELJAU and R. D. WACHTER.
1993. Extraction of high molecular weightDNA from mollusks. Trends in Genetics 9: 407.
YUZINE, E., I. PATIMAH, S. G. TAN and P. JOTHY. 1996.
Genetic polymorphism of local population ofudang galah (Macrobrachium rosenbergii de Man)and Udang Putih (Penaeus merguiensis deMan).In Second National Congress on Genetics, ed. M.Osman, M. M. Clyde and Z. Zamrod, pp. 451-457.
YOUNG, E.T., J. S. SLOAN and K. VAN RIPER. 2000.
Trinucleotide repeats are clustered inregulatory genes in Saccharomyces cerevisiae.Genetics 154: 1053-1068.
(Received: 25 February 2006)(Accepted: 7 September 2006)
94 PERTANIKAJ. TROP. AGRIC. SCI. VOL. 28 NO. 2 (2005)